Зміни деяких морфометричних показників на прикладі мальків коропових риб
Зміст
1. Вступ
2. Сімейство коропові
3. Розвиток коропових риб
3.1 Ембріональний період
3.1.1 І етап. Активація яйця і утворення бластодиска.
3.1.2 ІІІ етап. Бластуляція
3.1.3 ІV етап. Гаструляція.
3.1.4 Презумптивні зачатки
3.1.5 V етап. Органогенез у карпа
3.1.6. VI етап. Відокремлення хвостового відділу від жовткового
мішка
3.1.7 VII етап. Розвиток ембріональної судинної системи
3.1.8 VIII етап. Розвиток зяберно-щелепного апарату
3.2 Личинковий період
3.2.1 І етап. Змішане харчування
3.2.2 ІІ етап. Повне зовнішнє харчування, диференціація непарної
плавцевої складки
3.2.3 ІІІ етап. Розвиток променів в непарних плавцях
3.2.4 ІV етап. Розвиток променів в парних плавцях
3.3 Мальковий період
3.3.1 І етап. Розвиток лускатого покриву
4. Антропогенні фактори, що можуть впливати на розвиток мальків
коропових риб
4.1 Вплив важких металів та їх солей на розвиток мальків коропових
риб
4.2 Вплив рентгенівського випромінювання та радіоактивних речовин
на розвиток мальків
4.3 Вплив пестицидів на розвиток мальків коропових риб.
5. Найбільш поширені аномалії розвитку мальків коропових риб
6. Висновок
7. Перелік посилань
1.
Вступ
У риб, найдавнішої з груп хребетних тварин, існує
сувора система пристосувальницьких взаємодій з абіотичними факторами. Кожен
живий організм, в тому числі і риби, володіє відносно будь-якого діючого на
нього фактора генетично детермінованим, філогенетично набутим, унікальним
діапазоном толерантності, в межах якого цей фактор є для нього стерпним.
Антропогенні впливи, з одного боку, представляють собою нові параметри
середовища існування, з іншого - обумовлюють антропогенну модифікацію вже
наявних природних факторів і тим самим роблять значний вплив на стійкість
організмів до цих чинників. [1]
Період ембріонально-личинкового розвитку є
найбільш чутливим етапом в онтогенезі риб не тільки до дії абіотичних факторів
природного характеру (температура води, вміст кисню, величина pH, швидкість
течії, освітленість тощо), але і до впливу різних токсичних речовин. У цілому
ряді експериментальних робіт виявлено різні порушення у личинок риб під впливом
окремих факторів.
Аналізуючи численні експериментальні роботи,
можна говорити про те, що під впливом різноманітних за походженням
забруднювачів (сира нафта, пестициди, важкі метали тощо) у риб виявляються одні
і ті ж види аномалій розвитку.
Без сумніву можна говорити про те, що наявність
різного виду аномалій у риб є відповідною неспецифічною реакцією на дію
комплексу несприятливих факторів абіотичного походження та різного походження
токсичних речовин. Таким чином, різні морфологічні порушення, які виявляються у
молоді риб, можна ефективно використовувати в якості надійного критерію
екологічного стану водойми.
У своїй роботі я приведу приклади зміни деяких
морфометричних показників на прикладі мальків коропових.
2.
Сімейство коропові
В даний час до цього загону коропоподібних
відносять більше 2000 видів прісноводних (рідше - прохідних) риб, поширених в
Євразії, Північній Америці та Африці. Їх характерною особливістю є відсутність
зубів на щелепах (але так звані глоткові - розташовані на нижньоглоточних
кістках - зуби розвинені у них дуже добре.) Рот у коропоподібні часто висувний,
у багатьох видів з вусиками. Тіло зазвичай вкрита циклоїдною лускою, рідше
голе. Плавальний міхур великий і складається з 2-3 камер.
Загін включає кілька сімейств, в число яких
входить, наприклад, сімейство в*юнові (cob itidae). Але більша частина видів
коропоподібні відноситься до сімейства коропових (Cyprinidae). Це
найбільше сімейство серед риб взагалі, воно включає близько 1800 видів.
Більшість коропових воліє теплі води, але у деяких видів (краснопірок, язей)
ареал простягається до Заполяр'я. Найбільшим представником сімейства є
гігантський вусач (Catlocarpio siamensis), що досягає 3 м у довжину,
мешкає в Південно-Східній Азії. А серед самих дрібних можна назвати дионду (Dionda
diaboli), безбарвного нотрописа (Notropis perpallidus) і
цілий ряд інших американських коропових, чия довжина не перевищує 5 см.
Ікра у більшості коропоподібні клейка - риби
відкладають її на камені, рослинність або інший відповідний субстрат. Але у
деяких видів, наприклад у білого амура (Ctenopharyngodon idella), ікра
розвивається, плаваючи в товщі води.
Для ряду видів коропоподібні характерна турбота
про потомство. Наприклад, живе в Америці чорний толстоголов (Pimephales
promelas) відкладає ікринки в збудоване гніздо, а потім охороняє і
піклується про кладці. Широко відома турбота про потомство у нашого горчака (Rhodeus
sericeus) - самка цього виду з допомогою довгого яйцеклада відкладає ікру в
мантийную порожнину двостулкових молюсків. Серед коропових зустрічаються і
"зозулі", підкидають свою ікру в гнізда інших риб. Однак у більшості
представників загону турбота про потомство не виражена.
Молодь більшості коропових харчується
зоопланктоном, а ось раціони дорослих риб цієї групи досить різноманітні.
Наприклад, головень (Leuciscus cephalus) і жерех (Aspius
aspius) - активні хижаки, що полюють на дрібних риб, білий амур (Ctenopharyngodon
idella) воліє рослинну дієту, підуст (Chondrostoma nasus) і храмуля
(Varicorhinus capoeta) зскрібають обростання з підводних каменів і корчів.
Чимало серед коропових і риб, які харчуються бентосом, збираючи його з дна і
викопуючи з грунту. А ось уклейка (Alburnus alburnus) харчується
в основному комахами - причому не обмежується водними формами і тими, які
падають у воду, але і спритно ловить, вистрибуючи з води, літаючих над
поверхнею комарів і мушок.
Багато коропові риби, навіть відносно невеликих
розмірів, - важливий об'єкт промислу в прісних водах. Це лящ (Abramis brama),
вобла (Rutilus rutilus caspicus), язь (Leuciscus idus),
краснопірка (Scardinius erythrophthalmus) та ін Деякі представники цього
сімейства є об'єктами штучного розведення в ставкових господарствах.
Найбільш відомий серед них, звичайно, короп (Cyprinus
carpio) (рис.1), але дуже давно - вже більше 2 тис. років - у Китаї
розводять і товстолобика (Hypophthalmichthys molitrix)
Досить тривала і історія розведення і селекції
декоративних коропових - численних порід золотої рибки, виведених в Китаї з
срібного карася (Carasius auratus). За деякими даними, початок їх
розведення було покладено майже 2 тис. років тому, в I ст., а до масової
селекції приступили в X-XIII ст. Приблизно таку ж, якщо не більш довгу, історію
має розведення коропів і їх декоративних порід (коі) в Японії. [8]
коропова риба аномалія адаптаційний
3.
Розвиток коропових риб
Для кращого відстеження змін в морфометричних
показниках мальків коропових під впливом антропогенних факторів, розглянемо
розвиток коропових від ембріонального до малькового періоду в нормальних
умовах.
3.1
Ембріональний період
3.1.1
І етап. Активація яйця і утворення бластодиска.
Після з'єднання гамет починається активація яйця,
що помітно по виділенню вмісту кортикальних альвеол (гранул) і утворення
перивителлінового простору. Цитоплазма стягується на анімальний полюс і утворює
горбик бластодиск (рис. 1 А). Протягом етапу завершується друге ділення мейозу,
зближуються чоловічий і жіночий пронуклеуси і незабаром з'являється веретено
першого поділу дроблення. Протягом етапу відбувається затвердіння, тобто
збільшення міцності оболонки.
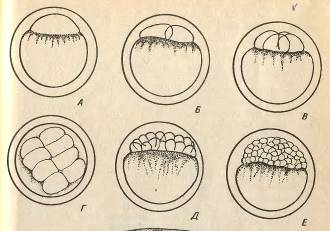
Рис.1. Ранні етапи розвитку карпа: А - І перший
етап - активація яйця та утворення плазмового горбика. ІІ етап - дроблення: Б -
2 бластомери, В - 4 бластомери, Г - 8 бластомерів, Д - 32 бластомери, Е - 128
бластомерів. ІІ етап. Дроблення
Дробленням називається серія міотичних поділів
ядра і цитоплазми зиготи, що не супроводжуються зростанням клітин. Це
призводить до утворення багатоклітинного зародка. Перша борозна дроблення
проходить через центр бластодиска в меридіональному напрямку і поділяє
бластодиск на два, зазвичай, різних за величиною, бластомера (рис.3 Б). Друга
борозна дроблення, також меридіональна, проходить перпендикулярно першій
борозні, і в результаті утворюються чотири бластомера (рис.3 В). Друга борозна
представляє собою площину білатеральної симетрії зародка. Дві борозни третього
поділу дроблення проходить паралельно одна іншій через центри розташовані в два
ряди бластомерів, утворюючи 8 клітин (рис.3 Г). Дві наступні борозни четвертого
поділу призводять до утворення 16 бластомерів. Особливість перших чотирьох
поділок, полягає в тому, що ділення клітин не завершуються і плазмалемма в
нижній частині бластомерів не утворюється. Всі клітини з'єднані в нижній
частині загальним цитоплазматичним шаром, утворюючи синтицій.
Борозна п'ятого ділення дроблення проходить вже
не меридіонально, а паралельно екватору жовтка і це призводить до появи 32
бластомерів і поділу їх на верхні, що не межують з жовтком, і нижні, підстави
яких раніше з'єднані загальним шаром цитоплазми (рис.3 Д). Сполучені нижні
бластомеры утворюють перибласт, який переходить у тонкий цитоплазматичний шар,
що оточує жовток. В результаті шостого ділення дроблення утворюється 64
бластомера, сьомого - 128 (рис.3 Е). Наступні поділи бластомерів призводять до
утворення великої кількості клітин, званих бластодермою.
3.1.2
ІІІ етап. Бластуляція
На етапі бластуляції ще продовжується дроблення
бластомерів і вперше стає помітною диференціація клітин. Поверхневі клітини
бластодерми ущільнюються і набувають полігональну форму. Їх бічні клітинні
мембрани прилягають один до одного і між ними виникають контакти. В результаті
цього на поверхні бластодерми з'являється епітеліальний пласт - перидерма. Під
перидермою знаходяться глибокі клітини, які нещільно прилягають один до одного,
між ними є щілини і порожнини, що представляють собою порожнину дроблення -
бластоцель (або первинну порожнину тіла).
В ембріології коропових умовно виділяють ранню,
середню та пізню бластули. Для ранньої бластули характерний високий купол клітин
бластодерми, середньої - невелика його сплощення, пізньої - значне сплощення і
вп*ячування в бластодерму в верхній частині жовчного мішка.
Глибоким клітинам бластодерми вже властиві слабкі
рухи. Вони виражаються в періодичній появі в них коротких виступів - лобоподій,
якими клітини торкаються одна одну, не утворюючи між собою контакту і не
здійснюючи спрямованих рухів.
Для процесу бластуляції характерне подовження
клітинного циклу, а саме інтерфази за рахунок збільшення фази S і появи фази G 1, відсутньої на етапі
дроблення. В ядрах виникають ядерця і активізується синтез РНК, в цитоплазмі
зростає число клітинних органел і білкових гранул. Перебудова клітинних циклів
- важлива подія в ранньому ембріогенезі коропових. Ця перебудова створює умови
для морфогенетичної функції ядер самого зародка і означає перехід від дроблення
до бластуляції.
3.1.3
ІV етап. Гаструляція.
Гаструляцією традиційно називають процес
розділення однорідної бластодерми бластули на два шари клітин - зародкові
листки. Розрізняють внутрішній і зовнішній листки, - ентодерму і ектодерму
відповідно. Пізніше між цими листками з'являється третій листок - мезодерма.
Гаструляція здійснюється завдяки інтенсивним морфогенетичним переміщенням
клітин, і результат завжди один - утвориться комплекс осьових зачатків,
розташованих відповідно до основного плану будови тварини.
У коропових риб гаструляція здійснюється двома
типами переміщення клітин бластодерми: обростанням жовтка, тобто епіболією, при
якій клітини мігрують в меридіональному напрямку (рис.2 А), і осьової
конвергенцією, при цьому клітини рухаються в широтному напрямі - до майбутньої
осі зародка (рис.2 Б). При переході до гаструляції відбуваються суттєві зміни в
структурі і властивостях клітин бластодерми. В їх цитоплазмі зростає число клітинних
органел і ускладнюється їх будова. Це призводить до збільшення активності
клітин, встановленню між ними контактів і їх злипанню. Короткі виступи глибоких
клітин - лобоподії - перетворюються на довгі відростки з віялоподібними
мембранами на кінцях - філоподії, завдяки яким і здійснюються контакти і
переміщення клітин.
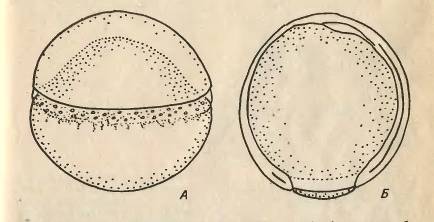
Рис.2. Гаструляція у карпа. А - початок
обростання. Б - завершення обростання.
3.1.4
Презумптивні зачатки
Протягом гаструляції глибокі клітини бластодерми
рухаються по своїх траекторіях до зародкового щитка. Досягнувши його, клітини
розсортовуються і займають певне положення: верхню частину зародка являє
ектодерма, середню - мезодерма і матеріал хорди, нижній шар, що лежить на
перибласті, представлений ентодермою (рис.3).
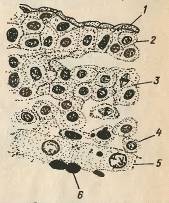
Рис.3. Зріз через зародковий щиток карпа 1 -
перидерма, 2 - ектодерма, 3 - мезодерма, 4 - ентодерма, 5 - перибласт, 6 -
жовток.
Це свідчить про те, що в ранньому ембріогенезі,
на бластулі, відбувається детермінація цих клітин. Вона виникає, очевидно, у
результаті якісних відмінностей клітин, розташованих у різних ділянках
бластодерми.
3.1.5
V етап. Органогенез у карпа
До початку органогенезу тіло зародка у вигляді
валика розташовується на жовтковому мішку (рис.6 А). Висота тіла, особливо
передньої частини, протягом етапу помітно зростає. Диференціація органів
відбувається в краніально-каудальному напрямку. Завершується етап утворенням
зачатка хвостового відділу - хвостової нирки.
Диференціація мезодерми. На самому початку
етапу під головним відділом зародків у вигляді валиків розташовується головна
мезодерма і мезодерма передніх тулубових сомітів (рис.4 А). Головна мезодерма
далі тоншає і розпадається на мезенхімні клітини. Мезодерма передніх сомітів у
вигляді двох тяжів продовжує просуватися в каудальному напрямку.
Тулубова мезодерма, що мігрувала з зародкового
кільця праворуч і ліворуч від осі зародка, утворює два пласта, що зливаються з
мезодермою, що надійшла в зародок спереду. Мезодерма розташовується у вигляді
пластів по обидва боки від хорди, представлена потовщеною частиною верхньої і
звуженої середньою частиною, яка переходить у тонкий вентральний шар -
спланхнотом, або бічну пластину.
Сегментації, тобто розчленування на окремі блоки
- соміти, піддається лише потовщена дорсальна частина мезодерми. При цьому
відбувається об'єднання мезодермальних клітин в групи, розділені вузькими
щілинами. Число сомітів поступово зростає, завдяки відокремленню їх із задньої
несегментованої частини мезодерми (рис.4 Б-Г).
Протягом етапу відбувається диференціація соміта.
Насамперед від нього відділяється спланхнотом, і в області його відділення від
"ніжок соміта" відособлюється проміжна мезодерма. Частина її,
називається нефротомом, утворює видільну систему зародка і личинки. Передній
короткий відрізок тяжа формує переднирку, а решта, довгий відрізок - вивідний
проток. Інша частина проміжної мезодерми у вигляді мезенхімних клітин
зміщується під хорду і зливається в поздовжній тяж клітин, що утворюють пізніше
магістральні судини - спинну аорту і задні кардинальні вени. Третя частина
мезенхімних клітин являє собою склеротомну мезенхіму. Вона мігрує до хорди і
охрящуває згодом на її оболонці в чотирьох місцях на кожен майбутній хребець:
праворуч і ліворуч у верхній і нижній частинах. Два дорсальний хрящових зачатка
утворюють пізніше, вже у личинок, невральну дугу хребця, а два вентральних -
нижні відростки хребців. тіла хребців утворюються в личинковому періоді життя.
Ще деяка кількість відокремленої від соміта мезенхіми, званої дерматомною,
переміщується під епітелій і утворює сполучнотканинний шар шкіри. Після
відокремлення від сомітів нефро-, склеро- і дерматома, а також мезенхіми
кровоносних судин залишаються їх осі, які утворюють тулубову мускулатуру.
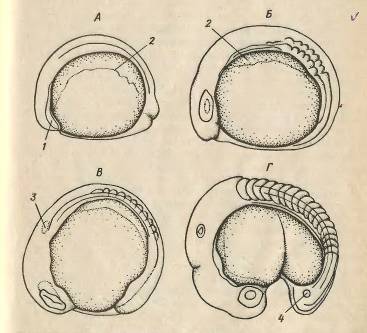
Рис. 4. Органогенез у карпа. А - закладка
зародка, Б - поява зачатків очей, В - поява слухових плактод, Г - поява в очах
хрусталиків, порожнин в слухових зачатках купферового міхурця, 1 - головна
мезодерма, 2 - бічна пластинка, 3 - слуховаплакода, 4 - купферов міхурець.
Диференціація ентодерми. Під
мезодермальним шаром зародка розташовується тонкий шар ентодерми, що лежить на
перибласті. задня частина ентодермального зачатка розширена і в ній приблизно в
середині етапу з'являється характерний для коропових риб провізорний орган -
купферів пухирець (рис.6 Г). Цей пухирець являє собою маленьке розширення в
ентодермі. З подовженням ентодермального зачатка він переміщається в
каудальному напрямку, потім зменшується і зникає.
Дифференціація ектодерми. Верхній
ектодермальный шар зародка має потовщену посередині невральну пластинку.
Клітини її, конвергуючи до центру, утворюють щільний нервовий тяж, потовщений у
головному відділі. По мірі росту зародка відбувається подовження неврального тяжа.
Слухові зачатки у вигляді плакод з'являються у
зародків на стадії 8-12 тулубових сегментів в області заднього мозкового міхура
(рис.4 В). За слуховими плакодами у риб також закладаються зачатки органів
бічної лінії. Вони розростаються під епідермісом вперед - над і під очима, і
назад, у тулуб.
3.1.6.
VI етап. Відокремлення хвостового відділу від жовткового мішка
На цьому етапі хвостова нирка, подовжуючись,
утворює хвостовий відділ зародка. У ньому диференціюється хорда, спинний мозок,
соміти. Суттєвим моментом цього етапу є закладка серця. Воно розвивається з
мезодермальних клітин, відділяються попереду від правого і лівого бічних
пластинок, в місці їх з'єднання. На ранній стадії серце представлено трубкою,
яка незабаром починає пульсувати. розташовується серце з лівої сторони зародка
між головою і жовтковим мішком. Продовжується подальша диференціація нервової
системи, що приводить до утворення п'яти відділів головного мозку. (рис.5).
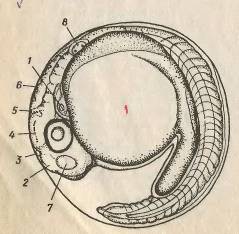
Рис. 7. відокремлення хвостового відділу від
жовточного мішка: 1 - серце, 2 - передній мозок, 3 - епіфіз, 4 - середній
мозок, 5 - зачаточний мозочок, 6 - довгастий мозок, 7 - нюхальна капсула, 8 -
слуховий міхурець.
Кишечник представлений щільним зачатком,
розташованим під хордою в поглибленні жовткового мішка. задню частину тіла і
хвіст зародка оздоблює вузька непарна плавникова складка, що розвивається за
рахунок ектодерми і містить мезенхімальні клітини. у передній частині зародка
нижче тулубових міотомів закладаються грудні плавці. На цьому етапі починається
нервово-м'язова моторика. Зародки стають рухливими і перевертаються в оболонці.
3.1.7
VII етап. Розвиток ембріональної судинної системи
На цьому етапі закладаються і починають
функціонувати багато дефінітивних судин і ряд провізорних, які виконують роль
органів дихання, а також йде формування клітин крові. На ранніх стадіях
еритроцити ще не пофарбовані, оскільки не містить гемоглобіну. Серце має великий
вигин і представлено двома змінно пульсуючими відділами: передсердям і
шлуночком. В зябровій області закладаються дуги. Судин в цій області ще немає,
тому кров із серця надходить в голову і спинну аорту за переднім парних
мандібулярих дуг аорти. Аорта розташовується під хордою і кров тече в ній в
каудальному напрямку. Досягнувши початку хвоста, аорта переходить у хвостову
артерію, яка згинається вниз, повертається вперед і стає хвостовою веною. У
тулубовому відділі хвостова вена продовжується в задню, поки ще непарну
кардинальну вену, розташовану під спинною аортою. Попереду грудних плавців
кардинальна вена роздвоюється і її гілки, зливаючись з передніми парними
кардинальними венами, що несуть кров з голови, утворюють кюв*єрові протоки
(рис.6 - А). Ці протоки впадають у передсердя.
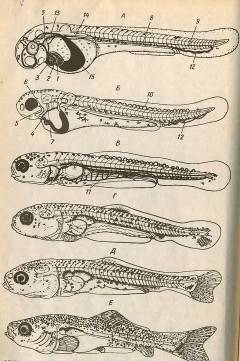
Рис.6. Завершення ембріонального періода (А, Б) і
личинковий період (В-Е) розвитку карпа. А - розвиток ембріональної судинної
системи; Б - Розвиток зяберно-челюсного апарата; В - змішане харчування; Г -
повне зовнішнє харчування; Д - розвиток променів на непарних плавцях; Е -
розвиток променів на парних плацях; 1 - венозний синус; 2 - передсердя; 3 -
шлуночок; 4 - черевна аорта; 5 - мандібулярна і 6 - гіоїдна дуги аорти; 7 -
зяберна кришка; 8 - спинна аорта; 9 - хвостова артерія; 10 - сегментарні
судини; 11 - жовточно-підкишкова вена; 12 - нижня хвостова вена; 13 - передня і
14 - задня кардинальні вени; 15 - проток Кюв є.
Протягом етапу відбувається поступове випрямлення
голови, що супроводжується появою вигину головного мозку. Тіло зародка оточує
непарна плавникова складка. Вона починається на спині, оздоблює хвіст і триває
вперед під хвостовим відділом до області анального отвору.
Зародки стали дуже рухливими і постійно рухаються
в яйцевих оболонках.
3.1.8
VIII етап. Розвиток зяберно-щелепного апарату
На цьому етапі відбувається вихід карпа з
ікринок, чому сприяють активні рухи зародків і зниження міцності яйцевих
оболонок. Риб, що вилупилися, називають передличинками (або вільними зародками)
У них відбувається швидка резорбція жовткового мішка, а довжина його не
змінюється.
У будові кровоносної системи відбулися значні
зміни. Зменшення передньої частини жовткового мішка призводить до поступового
вкорочення кюв*єрових протоків і зниження їх дихальної функції. В якості заміни
їм на жовточному мішку утворюється інша система судин, представлена
жовточно-підкишковими венами. На більш ранніх етапах розвитку кров із серця
надходила в спинну аорту лише по мандібулярній дузі. З появою зябрових артерій
мандібулярна дуга не редукувала. Вона стала постачати кров'ю розвиваючу на
цьому етапі хибнозябру, розташовану за очима. Протягом етапу відбувається
швидкий розвиток щелепного і зябрового апаратів, а також інших систем органів,
готуючих організм до активного способу життя.
3.2
Личинковий період
3.2.1
І етап. Змішане харчування
Характерна особливість цього етапу полягає в
переході личинок до активного способу життя. У зв'язку з цим у них заповнюється
повітрям плавальний міхур, що дозволяє личинок тримаються в товщі води (рис.8 -
В). Тіло личинки випрямлене, і його як і раніше оточує непарна плавцева
складка, розширена в хвості. Грудні плавці помітно збільшилися, займають
вертикальне положення і стали рухливими. В основі їх з'явився кістковий зачаток
плечового пояса - клейтрум. Зяброва кришка майже повністю закриває дуги. У
зябрових пелюстках починає циркулювати кров, що свідчить про посилення
зябрового дихання. Однак провізорна дихальна система ще продовжує
функціонувати. З боків голови і тіла розташовуються горбки органів бічної лінії
- невромасти. Вони представлені чутливими і опорними клітинами. Жовтковий мішок
став невеликим і вузьким. Кишечник на всьому протязі має порожнину, почалася
його слабка перистальтика. Під переднім відділом кишки розташовується печінка.
Рот став кінцевим, рухливим, але ще не цілком закривається. На тілі є
багаточисленні меланофори, голова і спина вкриті жовтим дифузним пігментом.
Личинки активно плавають і харчуються дрібними малорухомими водними
організмами, але основне їх живлення відбувається за рахунок жовткового мішка.
3.2.2
ІІ етап. Повне зовнішнє харчування, диференціація непарної плавцевої складки
Жовток резорбувався і личинки живляться тільки
зовнішньою їжею. У них добре сформувалися щелепи. Зябра закриті зябрової кришкою,
а в міжзяберній перетинці утворилися промені. Личинки повністю перейшли на
жаберное дихання. Задній кінець хорди злегка загинається вгору, розвиваються
нижні дуги хребців. Почалася диференціація непарної плавниковой складки.
кишечник розширився але як і раніше представлений прямою трубкою. посилилася
пігментація тіла, і в цілому личинки в зеленуватий колір, що робить їх менш
помітними серед рослин. Активному пошуку харчових організмів сприяють зір і
органи почуттів бічної лінії.
3.2.3
ІІІ етап. Розвиток променів в непарних плавцях
На цьому етапі завершується формування хрящового
черепа личинок і центр ваги зміщується вперед. Кінець хорди ще більше
загинається вгору. Починають швидко розвиватися промені в непарних плавцях,
спочатку в хвостовому, потім у спинному і анальному. Мускулатура хвостового
плавця розвивається за рахунок останніх хвостових міотомів. Виникають промені -
лепідотрихії утворюються за рахунок шкірного окостеніння. З появою перших
променів у спинному плавці закладаються черевні плавці у вигляді мезенхімних
скупчень. Прианальна складка зберігається ще широкою і служить стабілізатором
при русі (рис.8-Д). В кишечнику розвивається перша пара петель. Личинки стали
ще більш рухливими, тримаються поодинці на невеликій глибині біля берега. Харчуються
більш великими, ніж раніше ракоподібними і дрібними формами хірономід.
3.2.4
ІV етап. Розвиток променів в парних плавцях
На цьому етапі завершується розвиток лепідотрихій
в спинному, грудному і черевних плавцях. Відбувається повне відокремлення спинного
і анального плавців. Промені у хвостовому плавці стають членистими і більш
гнучкими. Міотоми набувають більш складної форми: їх верхні та нижні кінці
вигинаються вперед. Це дозволяє личинкам здійснювати складні й швидкі рухи. У
нюховому отворі розвивається перемичка, що ділить його на дві ніздрі: передню і
задню. В нюховій ямці відбувається диференціація рецепторних клітин. Триває
розвиток черепа, в кришці якого з'явилися окостеніння. Розвинені всі
окостеніння зябрової кришки. Ускладнилося будова ротового апарату, який стає
рухомим і здатним до всмоктування їжі. Личинки утворюють зграйки і живляться
крупними ракоподібними і хірономідами.
3.3
Мальковий період
.3.1
І етап. Розвиток лускатого покриву
На цьому етапі пропорції тіла коропа стають
схожими з дефінітивні. Проте очі в них більші порівняно з очима дорослих
особин. На тілі починається розвиватися лускатий покрив. Луска розвивається під
епідермісом, де серед колагенових волокон з'являються клітини-остеобласти, що
продукують мінералізований шар луски. Луска розташовується в кишенях, утворених
сполучнотканинними клітинами. Череп, підвісок і зяброві дуги мальків ще значною
мірою хрящові. Рот став нижнім, всмоктувальним, з'явилася здатність рити грунт.
Розвиваються вусики. Зростає довжина петель кишки. Нюховий отвір повністю
розділено на дві ніздрі. Преанальна плавцева складка редукувала. Мальки
тримаються в придонних шарах води, серед рослинності. У їхньому раціоні
переважає бентос. [4]
4.
Антропогенні фактори, що можуть впливати на розвиток мальків коропових риб
Техногенне забруднення водного середовища - одна
з глобальних екологічних проблем тимчасовості. Сила впливу цього фактора на
гідробіонтів, які виступають в якості маркерів техногенного пресингу на
водойми, визначається його якісними і кількісними ознаками.
До найпоширеніших забруднювачів техногенної
природи відносяться важкі метали і нафтопродукти. Нафта може потрапляти у воду
в результаті людської діяльності: використання нафти як палива і промислової
сировини. [2]
Інші забруднювачі - метали (ртуть, свинець, цинк,
мідь, хром, олово, марганець), радіоактивні елементи, отрутохімікати, що
надходять з сільськогосподарських полів, гербіциди, які, як відомо,
застосовують не тільки для знищення бур'янів на посівах сільськогосподарських
культур, але й для боротьби із заростанням водосховищ, ставків, зрошувальних
каналів.
Поряд з отрутохімікатами сільськогосподарські
стоки містять значну кількість залишків добрив (азоту, фосфору, калію), що
вносяться на поля. Крім того, великі кількості органічних сполук азоту і
фосфору потрапляють із стоками від тваринницьких ферм, а також з
каналізаційними стоками. Підвищення концентрації поживних речовин у ґрунті
призводить до порушення біологічного порушення рівноваги у водоймі.
Всі ці речовини, надходячи у водойми із стоками з
промислових та сільськогосподарських угідь, і частково з опадами, викликають
розвиток аномалій у мешканців цих водойм. [12]
Значний вплив також здійснюють Рентгенівські
промені, електромагнітне випромінювання, радіоактивні речовини, міський або промисловий
шум, інфразвук, ультразвук. [6]
4.1
Вплив важких металів та їх солей на розвиток мальків коропових риб
Важкі метали та їх солі (Сu, Zn, Hg, Cd, Pb,
Sn, Fe, Mn, Ag, Cr, Co, Ni, As, Al) - широко поширені промислові забруднювачі.
У водойми вони надходять з природних джерел (гірських порід, поверхневих шарів
ґрунту і підземних вод), зі стічними водами багатьох промислових підприємств і
атмосферними опадами, які забруднюються викидами з димовими. Важкі метали як
мікроелементи постійно зустрічаються в природних водоймах і органах
гідробіонтів.
Важкі метали досить стійкі. Потрапляючи у
водойми, вони включаються в кругообіг речовин і піддаються різним
перетворенням. Неорганічні сполуки швидко зв'язуються буферною системою води і
переходять в слаборозчинні гідроокису, карбонати, сульфіди і фосфати, а також
утворюють металлорганічні комплекси, адсорбуються донними осадами. Під впливом
живих організмів (мікробів та ін) ртуть, свинець, миш'як піддаються
метилованню, перетворюючись в більш токсичні алкільні з'єднання.
Велика частина неорганічних сполук металів
надходить в організм риб з їжею. Через зябра і шкіру проникають розчинні
диссоціюючі солі і металоорганічні з'єднання. Антропогенні джерела багаторазово
(у 2 - 13 разів) підвищують концентрацію важких металів у воді. З цим чітко
корелює вміст металів в органах мальків.
Токсична дія більшості важких металів на риб
обумовлено їх іонами. Концентровані розчини їх солей, володіючи
в'язко-припікаючою дією, порушують функції органів дихання. У слабких розведеннях,
проникаючи в організм, вони порушують проникність біологічних мембран, знижують
вміст розчинних протеїнів, які зв'язуються із сульфгідрильними та аміногрупами
білків і викликають тим самим пригнічення активності ферментів. Гідроокис
заліза і марганцю, осаджуючись на зябрах, порушують газообмін, що призводить до
асфіксії. З підвищеним забрудненням води сполуками титану, заліза, кадмію,
хрому та інших металів пов'язують ураження мальків пухлинами (епідермальна
папілома, псевдопухлина зябер, карцинома печінки) і виразковою хворобою, а
також деформацію скелета і запалення плавників. [11]
Одним з найбільш токсичних металів є кадмій, який
акумулюється в організмі переважно в нирках і печінці. Він накопичується у
вигляді комплексів з білками, які беруть участь у ферментативних процесах.
Потрапляючи в організм малька ззовні, кадмій чинить інгібуючу дію на цілий ряд
ферментів, руйнуючи їх. Його дія заснована на
зв'язуванні групи-SH цистеїнових залишків у білках і інгібуванні SH-ферментів.
Він може також інгібувати активність цинковмісних ферментів, заміщаючи цинк.
До важких металів відноситься також марганець,
який є есенціальним елементом, однак при надмірному надходженні в організм
надає токсичну дію. При хронічному отруєнні сполуки марганцю виступають як
цитоплазматичні отрути, здатні викликати важкі порушення в нервовій системі,
нирках і органах кровообігу. [14]
4.2
Вплив рентгенівського випромінювання та радіоактивних речовин на розвиток
мальків
Промені Рентгена надають різке негативне дію на
мальків. Як показав дослід Г.В. Самохвалової (1935), доза в 4000r є летальною для
рибок. Менші дози викликають зменшення посліду і дегенерацію статевих залоз.
Опромінення статевонезрілих самців викликає у них недорозвинення вторинних
статевих ознак.
Радіоактивне випромінювання впливає сильніше на
ікру і ембріонів ніж на дорослих організмів.
Розвиток атомної промисловості, а також
випробування атомних і водневих бомб призвели до значного зростання
радіоактивності повітря і води та акумуляції радіоактивних елементів у водних
організмах. Основним радіоактивним елементом, що має значення в житті риб, є
стронцій 90
(Sr90). Стронцій проникає в
організм малька головним чином через шлунково-кишковий тракт, а також через
зябра і шкіру. Основна маса стронцію (50-65%) концентрується в кістках, значно
менше в нутрощах (10-25%) і зябрах (8-25%) і зовсім небагато - у м'язах (2-8%).
Але накопичений головним чином в кістках стронцій, викликає появу в м'язах
радіоактивного ітрія - I 90.
Мальки акумулюють радіоактивність як з води, так
і з інших організмів, які служать їм їжею. Акумуляція радіоактивних речовин у
молодих риб йде більш швидко, ніж у дорослих, що пов'язано з більшою
інтенсивністю обміну речовин у перших.
Накопичення стронцію в кістках риб викликає
розвиток так званої Уровської хвороби, пов'язаного з порушенням кальцієвого
обміну. [5]
4.3
Вплив пестицидів на розвиток мальків коропових риб.
В даний час в сільському господарстві та інших
галузях використовують велику кількість пестицидів, асортимент і об'єм
виробництва яких постійно збільшується. До складу пестицидів найчастіше входять
такі хімічні сполуки як: сірчана кислота, нітрат натрію, ціанамід кальцію,
ціанамід натрію, ціанамід калію, хлорат натрію, арсеніт натрію, борати та
багато ін. Ці речовини, потрапляючи у воду, а потім в організм гідробіонтів,
включаються в метаболізм, можуть викликати порушення процесів життєдіяльності
та пластичного обміну риб. [16]
Під дією пестицидного навантаження у мальків та
молодих коропових риб можуть спостерігатися наступні порушення: геморагії,
виразки на зовнішніх покривах тіла, набряки тканин, точкові крововиливи,
жовтувате забарвлення поверхні в області анального отвору, сітчасті крововиливи
на грудних і черевних плавцях, зниження тургору м язів. [10]
5.
Найбільш поширені аномалії розвитку мальків коропових риб
Пухлина в одному очному яблуці відноситься до
групи порушень морфології очей. Одне з очних яблук збільшено в розмірі і має
неправильну форму з-за того, що в ньому присутня пухлиноподібне утворення. Як
правило, пухлина складається з пухкої тканини позбавленої пігменту або в деяких
випадках слабо пігментованою. Розміри таких новоутворень варіюються від 0,1 до
1,1 мм.
Відсутність одного черевного плавця. Даний тип
аномалії відноситься до групи порушень морфології плавників і виражається в
повній відсутності одного черевного плавця при наявності нормального другого
Порушення пігментації очного яблука віднесено
до групи порушень пігментації тіла. Дана морфологічна аномалія проявляється в
тому, що очне яблуко частково або повністю позбавлене пігменту; іноді воно
зовсім прозоре. По всім іншим параметрам очне яблуко, як правило, не має
відхилень від норми.
Непігментовані пухлини на голові відносяться до
групи непігментованих новоутворень на тілі. Даний тип порушення виражається в
присутності безбарвного, або слабопігментованого, новоутворення або пухлини,
локалізовані зазвичай за очними яблуками в тім'яній області голови, рідше - між
очима. Розміри таких новоутворень можуть варіювати від 1,2 до 1,7 мм. [15]
6.
Висновок
Найбільшу частку серед виявлених морфологічних
порушень у мальків коропових риб під дією антропогенних факторів складають
аномалії, що входять у групу порушень пігментації тіла. До даних потворностей
відносяться поодинокі або множинні пігментовані новоутворення в області очних яблук,
пігментовані новоутворення в міотомах тулуба і в основі хвостового плавця,
поверхневі пігментоми тулуба. Ці аномалії є не тільки найбільш масовими серед
порушень даної групи, а й становлять найбільшу частину серед усіх виявлених
аномалій у молоді риб. В групу порушень пігментації тіла входять також
відсутність пігментних клітин в стандартних місцях локалізації, що призводить
до порушення видоспецифічного пігментного малюнка, але ці порушення
зустрічаються у молоді риб відносно рідко.
Перераховані вище морфологічні аномалії віднесені
до малолетальних або нелетальних потворностей, так як вони виявляються навіть
на пізніх стадіях личиночного розвитку і у мальків. Переважну частину аномалій,
виявлених у риб такого віку, становлять саме порушення пігментації тіла. Втім,
і на більш ранніх стадіях розвитку переважають аномалії цієї групи.
Непігментровані новоутворення, об'єднані в окрему
групу, складають приблизно 5% від загальної кількості морфологічних порушень.
До аномалій даної групи відносяться непігментовані або слабопігментовані
пухлини в області очних яблук (всередині і зовні ока), а також непігментовані
або слабопігментовані новоутворення в міотомах тулуба. Ці відхилення в
морфології можна віднести до вад внутрішньої будови, ступінь їх летальності для
риб набагато вище, ніж ступінь летальності пігментованих новоутворень. Всі
каліцтва даної групи віднесені до розряду напівлетальних аномалій. Як правило,
молодь риб, що є носієм даних порушень, перестає зустрічатися серед риб вже на
стадіях розвитку пізніх мальків. [13]
Порушення фізіологічних процесів і, як наслідок,
поява новоутворень у риб є прямим доказом канцерогенного впливу окремих
компонентів забруднення водного середовища. Крім дегенеративних змін в bulbus
arteriosus, аневризм, мінералізації тканин і глануломатозного запалення,
відзначені також різні неоплазми: лепідоцитоми, нейробластоми нюхових органів,
карциноми стравоходу.
Зустрічальність великої кількості молоді
коропових з перерахованими вище аномаліями є прямим наслідком і доказом
присутності у воді цілого комплексу забруднюючих речовин, які потрапили туди в
результаті діяльності людини.
7.
Перелік посилань
1. Тертишний О.С., Товстик В.Ф., "Рибництво з основами
гідрології". - 2009. - 288 с.
. Хімко Р.В. Мережко В.І. Бабко Р.В. "Малі річки. Дослідження,
охорона, відновлення". Київ, 2003-238с.
. С.Н. Александров. "Садковое рыбоводство" М.: ACT;
Донецк: Сталкер, 2005. - 270 с.
. Макєєва А.П. "Ембріологія риб", видавництво
Московського університету, Москва, 1992 р.
. Никольский Г.В. "Экология рыб", Высшая школа, Москва 1963г.
. Жукинский В.Н. "Влияние абиотических факторов среды на
разнокачественность и жизнеспособность рыб в раннем онтогенезе" М.:
Агропромиздат, 1986.248 с.
7.
<http://www.coolreferat.com>
.
http://uk. wikipedia.org
.
<http://uarybka.com>
.
http://vestnik. udsu.ru
.
http://www.ssc. smr.ru <http://www.ssc.smr.ru>
.
<http://www.cnshb.ru>
.
http://archive. nbuv.gov.ua
.
http://www.unn.ru