Получение шикимовой кислоты
Введение
Впервые шикимовая кислота
(3,4,5-тригидрокси-1-циклогексен-1-карбоновая кислота) была выделена в 1885
году из плодов аниса - растения, произрастающее в Японии - Illicium anisatum. Японское название данного
растения «Шикимино - ки», что и послужило основой для названия данной кислоты.
Дальнейшие исследования данного соединения показали, что оно является
промежуточным продуктом щикиматного пути у микроорганизмов и растений. Данный
метаболитический путь отсутствует у животных.
Шикимовая кислота является промежуточным звеном в синтезе
лигнина, ароматических аминокислот (фенилаланина, тирозина и триптофана),
убихинов, фолиевой кислоты, тетрациклина, фенольных и карболовых соединений и
большинства алкалоидов растений и микроорганизмов. Используется в качестве
реагента в органическом синтезе.
Также используется в качестве исходного вещества в синтезе
осельтамивира (тамифлю) - противовирусного препарата, используемого в лечении
гриппа; зейленона для лечения раковых заболеваний; монопальмитилоксишикимовой
кислоты, обладающей анетикоагуляционной активностью при внутримышечном
введении.
Производные шикимовой кислоты могут использоваться в качестве
гербицидов и антибактериальных средств, так как они могут блокировать
шикиматный путь у растений и бактерий не нанося вреда для животных и человека,
так как у них отсутствует данный метаболитический путь.
Данная кислота может быть использована в качестве исходного
соединения для синтеза ряда ароматических и хиральных соединений.
Как видно из сказанного шикимовая кислота является важным
продуктом для химической промышленности, медицины, фармакологии и сельского
хозяйства. Поэтому встаёт вопрос о получении большого количества данного
соединения. В мировой фармацевтической промышленности существует три подхода к
получению шикимовой кислоты: выделение из растительного сырья,
микробиологический и химический синтез. Химический синтез является
дорогостоящим методом. Синтез из растительного сырья является трудоёмким
методом и не обеспечивает высокого выхода продукта. Поэтому наиболее
перспективным методом является микробиологический синтез.
Цель: изучить литературу по данной проблематике и получение
промежуточной конструкции, содержащей вставку, ограниченную генами tmrB и aroK, на основе плазмиды pMTL21C.
Задачи:
1) Амплифицировать фрагмент ДНК, ограниченный генами tmrB и aroK, с матриц В. Subtilis ВНИИгенетика-15 С10 и В.
Subtilis ВНИИгенетика-15 D4.
2) Продукт амплификации лигировать с плазмидой pMTL21C, для получения
промежуточной конструкции. Плазмиду pMTL21C предварительно подвергают рестрикционному анализу по сайту
рестрикции SmaI.
) Трансформировать полученной лигированной смесью клетки E.coli DH5б.
1. Обзор литературы
.1
Общая характеристик шикимовой кислоты
Шикимовая кислота представляет собой бесцветные игольчатые
кристаллы с температурой плавления 184°С. Это соединение левовращающее и имеет
удельное вращение -157°С. Для данного вещества характерен единственный максимум
в ультрафиолетовом диапазоне, наблюдаемый при л = 235 нм. Для шикимовой кислоты
описано несколько стериоизомеров, но биологической активностью обладает
б-изомер, структура которого изображена на рис. 1.1.
шикимовый плазмида реактив бактериальный
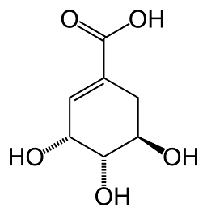
Рисунок 1.1 - Общая структура б-изомера
3,4,5-тригидрокси-1-циклогексен-1-карбоновой кислоты
Молекула шикимовой кислоты в целом не симметрична и имеет 3
стереогенных центра, что обеспечивает возможность существования 23=8
стереоизомеров (4 пары энантиомеров). Природная (−) - шикимовая кислота
имеет (3R, 4S, 5R) - конфигурацию, семь других стереоизомеров ((3S, 4R, 5S)-,
(3R, 4R, 5S)-, (3S, 4S, 5R)-, (3R, 4S, 5S)-, (3S, 4R, 5R)-, (3S, 4S, 5S) - и
(3R, 4R, 5R) - шикимовые кислоты) не имеют биологического и практического
значения. Рацемат из (3R, 4S, 5R) - и (3S, 4R, 5S) - шикимовых кислот имеет
температуру плавления 191-192°C.
Для шикимовой кислоты характерны реакции являющиеся типовыми
для соединений, содержащих карбоксильную группу, три гидроксильные группы и
двойную связь. Стереохимия молекулы позволяет при соответствующих условиях
протекать внутримолекулярным реакциям между карбоксильной группой и одной
гидроксильной группы. Для молекулы также характерны реакции цис-транс переходов
для гидроксильных групп [7].
.2
Шикиматный путь
Шикиматный путь является общим метаболитическим путём для
целого ряда организмов, приводит к образованию разнообразных ароматических
соединений. Ключевым метаболитом этого пути является шикимовая кислота, которую
можно использовать в качестве материала для синтеза ингибиторов нейраминидазы и
препаратов, используемых в противоопухолевой терапии.
Шикиматный путь был обнаружен Бернхардом Дэвисом и Дэвидом
Спринсоном как биосинтетический путь образования ароматических аминокислот
фенилаланина, тирозина и триптофана. Этот путь присутствует у разных групп
микроорганизмов, у растений и паразитов, но отсутствует у животных. Фенилаланин
и триптофан являются важными компонентами рациона питания животных, и животные
синтезируют тирозин из фенилаланина в одну стадию.
Значение шикиматного пути велико, так как этот путь является
единственным установленным путём биосинтеза ряда важнейших природных
соединений, в том числе значимых в плане их практического использования.
Схема шикиматного пути представлена на рисунке 2.1.
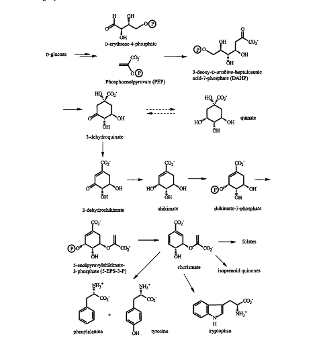
Рисунок 2.1 - Шикиматный путь
В 1935 году немецкий химик-органик Германн Отто Лауренц Фишер
(1888-1960) отметил, что кислоты хинная и шикимовая могут быть метаболитически
тесно связанными с бензоидными ароматическими соединениями. Подтверждения этого
предположения не существовало до 1950-ых годов.
В 1950 г. американский микробиолог Бернард Дэвис (1916-1994),
используя метод отбора с применением пенициллина, получил серию мутантов Escherichia
coli, способных расти лишь в такой питательной среде, в которую добавлялись
ароматические соединения. Многие мутанты нуждались в фенилаланине, тирозине,
триптофане, парааминобензоате и парагидроксибензоате (в следовых количествах).
Неожиданно оказалось, что потребность во всех пяти соединениях можно
удовлетворить путем добавления шикимовой кислоты (шикимата), в то время
считавшимся редким соединением, встречающимся лишь у некоторых растений.
Таким образом, шикимат оказался промежуточным продуктом в
процессах биосинтеза трёх ароматических протеиногенных аминокислот,
парааминобензоата и других существенных ароматических соединений.
При помощи ауксотрофных мутантов Escherichia coli,
Aerobacter aerogenes и Neurospora и с применением изотопно-меченых
соединений был прослежен весь путь от углеводных предшественников до первичных
и вторичных метаболитов ароматической природы. Важнейшая часть работы была
проделана Бернардом Дэвисом в сотрудничестве с Дэвидом Спринсоном (1910-2007)
из Колумбийского университета. Значительный вклад в изучение отдельных стадий
процесса внесли также австралийцы Франк Гибсон (1923-2008), Джеймс Питтард и
многие другие исследователи.
Дэвис и Спринсон установили путь через ДАГФ, долго
считавшийся единственным. В 2004 году американский биохимик Роберт Вайт
опубликовал результаты своих исследований по рассшифровке пути через ДКФФ.
Однако предложенная Вайтом гипотетическая схема биосинтеза ДКФФ в последствии
не подтвердилась. В 2006 году Роберт Вайт и Хуимин Ксу сообщили, что им удалось
выяснить истинный путь биосинтеза ДКФФ у Methanocaldococcus jannaschii.
Шикиматный путь у бактерий используется исключительно для
синтеза ароматических аминокислот. Высшие растения используют эти аминокислоты
не только в качестве строительных элементов белка, но и как предшественников
вторичных метаболитов [9].
Ферменты шикиматного пути у микроорганизмов находятся в
цитоплазме, у растений в пластидах [3].
Шикиматный путь включает в себя 7 каталитических реакций и
сочетает обмен углеводов с синтезом ароматических аминокислот. В результате
первых четырёх реакций происходит образование шикимовой кислоты, а следующие
три реакции обеспечивают ее дальнейшее преобразование в хоризмовую кислоту.
Хоризмовая кислота является предшественником многих ароматических соединений,
фолиевой кислоты, убихинона [3].
Путь биосинтеза ароматических соединений схож в клетках
бактерий, паразитов и растений. Наблюдаемые различия относятся к структуре
ферментов и способа их регулирования.
Первым идентифицированным метаболитом данного пути была
шикимовая кислота, которая и дала ему данное название. Так как шикиматный путь
в дальнейшем приводит к образованию различных ароматических соединений, его
часто называют общим путём биосинтеза ароматических соединений. Однако Бентли
считает такое название не очень корректным, так как не все ароматические
соединения образуются по данному пути [12], [13].
Протекание всех реакций катализируется определёнными
ферментами, ферменты являются продуктами соответствующих генов. Гены
шикиматного пути у бактерий обозначаются aro (от англ. Aromatic - ароматический), так
как ферменты, кодируемые данными генами участвуют в образовании ароматических
соединений [12].
Исходными компонентами шикиматного пути являются
фосфоенолпируват образующийся при гликолитическом окислении глюкозы, и
эритроза-4 - фосфат - промежуточный продукт окисления глюкозы в пентозофосфатном
пути. Шикиматный путь начинается с конденсации фосфоенолпирувата и эритроза -4
- фосфата с образованием 3-дезокси-D-арабиногептулозонат-7-фосфата или ДАГФ и
заканчивается синтезом хоризмата. Промежуточный продуктом является шикимат.
Хоризмат служит общим предшесивенником L - триптофана, L - тирозина, L - фенилаланина и
п-аминобензойной кислоты.
1) фосфоенолпируват + эритроза-4 - фосфат ДАГФ + Фн
Данная реакция регулируется 3-дезокси-D-арабиногептулозонат-7-фосфат-синтазой.
Этот фермент у E. coli представлен тремя изоферментами: ДAГФС - [Phe], ДAГФС - [Tyr] и ДAГФС - [Trp], которые контролируются
соответствующими конечными продуктами. Данные изоферменты кодируются
соответственно генами aroG, aroF, aroH [14].
На второй стадии происходит циклизация ДАГФ, что приводит к
образованию 3-дегидрохинной кислоты или 3-дегидрохината.
2) ДАГФ 3-дегидрохинат + Фн
Данная реакция регулируется 3-дегидрохинат-синтазой. Для
проявления активности данного фермента необходимо присутствие НАД+ и ионов
цинка и кобальта. Синтез фермента у E. coli регулирует ген aroB [12].
На следующей стадии происходит дегидратация 3-дегидрохината с
образованием 3-дегидрошикимата.
3) 3-дегидрохинат 3-дегидрошикимат + H2O
Данную реакцию катализирует 3-дегидрохинат-дегидротаза или
3-дегидрохинат-дегидрогеназа. У E.coli данный фермент представляет собой димерный белок, кодируемый
геном aroD [6].
На четвёртой стадии происходит превращение 3-дегидрошикимата
в результате НАДФН+ зависимой реакции в одно из вжнейших промежуточных
соединений - шикимовая кислота.
4) 3-дегидрошикимат + НАДФ шикимат
Данная реакция осуществляется шикимат-дегидрогеназой, синтез
которой осуществляется у E. coli геном aroE. Концентрация данного фермента в бактериальной клетке очень мала.
На пятой стадии осуществляется фосфолирование шикимовой
кислоты по третьему атому углерода и образование шикимат-3-фосфата.
5) шикимат + АТФ шикимат-3-фосфат + АДФ
Данная реакция катализируется шикиматкиназой, кодируемой
геном aroL.
На шестой стадии происходит присоединение к фосфолированной
шикимовой кислоте молекулы фосфоенолпирувата с образованием
5-енолпирувилшикимат-3-фосфата.
6) шикимат-3-фосфат + ФЕП 5-енолпирувилшикимат-3-фосфат + Фн
Эта реакция катализируется ферментом
5-енолпирувилшикимат-3-фосфат-синтазой, синтез которого контролируется геном aroA. Активность данного
фермента ингибируется глифосатом, в результате чего происходит накопление
больших количеств шикимата и шикимат-3-фосфата в среде для культивирования.
Этот факт был положен в основу методов получения шикимовой кислоты путём
экстракции из растительного материала контактирующего с глифосатом, а также при
обработке этим реагентом микроорганизмов способных продуцировать шикимовую
кслоту, например E. Coli [13].
На последней стадии происходит дефосфолирование и дегитратация
5-енолпирувилшикимат-3-фосфата и образование конечного продукта хоризмата.
7) 5 - енолпирувилшикимат-3-фосфат хоризмат +Фн
Данная реакция осуществляется хоризмат-синтазой, синтез
которой детерминируется геном aroC. У растений и E.coli даннай фермент активен
при наличии ФМНН2 и в анаэробных условиях [12].
На этой стадии происходит разветвление шикиматного пути. По
одному направлению из хоризмовой кислоты образуется антранилат, который
является предшественником триптофана и алкалоидов, по другому - префенат -
предшественник фенилаланина и тирозина. Также существуют другие пути, которые
приводят к образованию антибиотиков, убихинина, салицилата и других соединений
в зависимости от вида организма [14].
.3
Регуляция шикиматного пути
Большинство ферментов шикиматного пути синтезируются
конститутивно, исключение составляет ДАГФ-синтаза и шикиматкиназа. Их
активность регулируется аллостерически, за счёт взаимодействия с активаторами и
ингибиторами. В качестве ингибиторов часто выступают конечные продукты реакций,
которые они катализируют. Например, шикиматкиназа будет ингибироваться
шикимат-3-фосфатом.
У большинства изученных организмов регуляторный контроль
шикиматного пути основан на первом аллостерическом ферменте - ДАГФ-синтазы. У E.coli как уже говорилось ранее
этот фермент представлен тремя изоферментами: ДAГФС - [Phe], ДAГФС - [Tyr] и ДAГФС - [Trp], которые контролюруется
по типу ретроингибирования конечными продуктами - фенилаланином, тирозином и
триптофаном [14].
Также этот фермент может регулироваться с помощью
транскрипции. Этот механизм регуляции наиболее изучен у E. Coli. Уровень экспрессии генов aroG и aroF регулируется белком
репрессором TyrR в комплексе с тирозином и фенилаланином; а TrpR-белок при взаимодействии
с триптофаном репрессирует транскрипцию гена aroH. TyrR-белок также осуществляет
регуляцию своего собственного гена, а также экспрессию гена шикиматкиназы [15].
.4
Способы получения шикимовой кислоты
Высокая востребованность шикимовой кислоты и наличие широкого
круга областей, где она может быть использована обуславливает постоянный поиск
новых способов её получения. Наиболее популярны следующие способы получения
шикимовой кислоты: химический синтез, выделение из растительного сырья,
микробиологический синтез [16].
Первый успешный синтез шикимовой кислоты в лабораторных
условиях был проведён в 1960 году. Далее были разработаны несколько
модифицированных синтезов данного соединения, а также были применены новые
схемы синтеза шикимата, в которых использовались широкодоступные и недорогие химические
соединения. Но этот способ синтеза имеет ряд недостатков - высокая стоимость,
низкий выход целевого продукта и высокая вероятность образования биологически
неактивных изомеров. Эти недостатки ограничивают использование этого способа
синтеза шикимовой кислоты [17].
Также шикимовую кислоту можно получать из растительного
сырья. В настоящее время шикимовую кислоту выделяют из китайского
звёздообразного аниса. Выход продукта составляет 3-7% от веса сухих семян,
самый высокий из всех исследованных растений. Но данное растение сложно
культивируется и достигает способности к образованию семян на шестой год. И для
получения 1 кг кислоты надо 30 кг семян.
Известно, что многие автотрофы производят шикимовую кислоту,
но её содержание в большинстве автотрофов низкое.
Поэтому продолжается поиски альтернативных растений для
синтеза данного соединения. Недавно выделена чистая шикимовая кислота (выходы
2,4-3,7%) из семян Liqutdambar styracifina, которое растёт в изобилии в восточной части
Северной Америки и Мексики. Её высокое содержание обнаружено и в листьях гингко
(Gingo biloba) [1].
Было установлено, сто глифосат блокирует действие
шикиматкиназы. Поэтому обработка данным гербицидом растений увеличивает выход
шикимовой кислоты[13].
Но несмотря на то что методы выделения шикимовой кислоты
постоянно совершенствуются, а также расширяется круг используемых растений,
этот метод имеет ряд недостатков: является трудоёмким, поступление
растительного сырья для выделения ограничивается сезоном года, когда содержание
шикимовой кислоты максимально, и данного сырья недостаточно для удовлетворения
существующих потребностей в шикимовой кислоте.
Поэтому наиболее перспективными методами в синтезе данного
соединения является микробиологический синтез. На сегодняшний день одна треть
от всего количества шикимовой кислоты получается путеё синтеза с использование
сверхпродуцентов E. Coli.
Для создания микроорганизмов-сверхпродуцентов шикимовой
кислоты применяется различные подходы. Например Джоном Фростом с коллегами был
сконструирован штамм E.coli SP1.1/pKD12.112, дефектный по гену
шикиматдегидрогиназы, продуктивность которого составила более 50 г. шикимата на
литр культуральной жидкости. Но данные бактерии также секретировали в среду
большое количество дегидрошикимата и хинной кислоты, поэтому для получения
чистой шикимовой кислоты необходимо было производить трудоёмкую процедуру
очистки[18].
Луиза Ёхансен с коллегами создали сверхпродуцент E.coli W3110.shik1 за счёт модификации гена
aroL, который детерминирует
образование фермента шикиматкиназы[19].
Сверхпродуценты шикимовой кислоты были созданы на основе Bacillus subtilis. За счёт увеличения
числа копий генов, кодирующих шикиматдегидрогеназу и ингибирование генов,
отвечающих за синтез шикиматкиназы. Были получены штаммы с выходом шикимата 14
г./л[21].
1.5
Применение шикимовой кислоты
Шикимовая кислота используется в синтезе противовирусного
препарата осельтамивира, который является эффективным средством против вируса
гриппа H5N1 особенно на начальных этапах лечения. Также препарат применяется
для лечения гриппа А и В.
Осельтамивир - пролекарство, при пероральном приеме
подвергается гидролизу и превращается в активную форму - осельтамивира
карбоксилат. Механизм действия осельтамивира карбоксилата связан с
ингибированием нейраминидазы вирусов гриппа типа А и В. Нейраминидаза -
поверхностный гликопротеин вируса гриппа, является одним из ключевых ферментов,
участвующих в репликации вирусов гриппа А и В.
При ингибировании нейраминидазы нарушается способность
вирусных частиц проникать внутрь клетки, а также выход вирионов из
инфицированной клетки, что приводит к ограничению распространения инфекции в
организме. При ингибировании нейраминидазы нарушается способность вирусных
частиц проникать внутрь клетки, а также выход вирионов из инфицированной
клетки, что приводит к ограничению распространения инфекции в организме.
Уменьшает выраженность симптомов и сокращает продолжительность заболевания,
ускоряет элиминацию вируса из организма, снижает частоту развития осложнений
гриппа, требующих проведения антибиотикотерапии. Осельтамивир входит в состав
препарата тамифлю[10], [11].
Производные шикимовой кислоты обладают многими
фармакологическими эффектами: антикоагулирующими, противовоспалительными,
антиоксидантными[4].
Шикимовая кислота широко применяется в качестве реагента в
органическом синтезе как в органической химии, так и в фармакологии. Описан
способ получения фенола из шикимовой кислоты. Китайскими учёными был
синтезирован зейленон, препарат который широко используется при химиотерапии
раковых заболеваний. А также известно о синтезе монопальмитилоксишикимовой
кислоты, обладающей анетикоагуляционной активностью при внутримышечном введении.
Группой китайских учёных синтезировано производное шикимовой кислоты -
триацилшикимовая кислота, также обладающая интикоагуляционным и
антитромбическом действием[2].
Производные шикимовой кислоты могут использоваться в качестве
гербицидов и антибактериальных средств, так как они могут блокировать
шикиматный путь у растений и бактерий не нанося вреда для животных и человека,
так как у них отсутствует данный метаболитический путь.
Данная кислота может быть использована в качестве исходного
соединения для синтеза ряда ароматических и хиральных соединений. При кислотной
дегидротации шикимовой кислоты образуется п-гидросибензойная кислота. Эта
кислота является предшественником парабенов. 20
Таким образом, шикимовая кислота является важным реагентом в
синтезе различных соединений в органической химии, фармокологии и сельском
хозяйстве.
2. Материалы и методы
.1
Бактериальные штаммы и плазмиды
В работе использовали бактериальный штамм E.coli DH5б
и плазмиду pMTL21C, матрицы для полимеразной цепной реакции В. Subtilis ВНИИгенетика-15 С10 и В.
Subtilis ВНИИгенетика-15 D4, характеристика которых
представлена в таблице 2.1.
Таблица 2.1 - Характеристика бактериального штамма и
плазмиды, использованных в работе
|
Название
|
Характеристика
|
Источник
|
|
E.coli DH5б
|
F - endA1
glnV44 thi-1 recA1 relA1 gyrA96 deoR nupG Ц80dlacZДM15 Д (lacZYA-argF) U169,
hsdR17 (rK - mK+), л-
|
23
|
|
pMTL21C
|
AmpR,
CmR, ColE-репликон, tac-промотор, полилинкер (3,648kb)
|
22
|
|
В. Subtilis ВНИИгенетика-15 С10
|
Получен путём
ступенчатого мутагенеза штамма В. Subtilis
ВНИИгенетика-15 и последующего отбора мутантов, резистентных к структурным
аналогам триптофана
|
Коллекция
гафедры
|
|
В. Subtilis ВНИИгенетика-15 D4
|
Получен путём
ступенчатого мутагенеза штамма В. Subtilis
ВНИИгенетика-15 и последующего отбора мутантов, резистентных к структурным
аналогам триптофана
|
Коллекция
гафедры
|
Краткая
характеристика штамма E.coli DH5б
Бактерии рода Escherichia грамотрицательные,
палочковидные бактерии. Эти бактерии являются факультативными анаэробами. Штамм
DH5б один из наиболее часто
используемых штаммов для технологии рекомбинантной ДНК. Штамм DH5б был разработан Ханаханом
в лаборатории для экспериментов по клонированию. Этот штамм был разработан с
содержанием нескольких мутаций, которые позволяют отбирать высокоэффективных
трансформантов. Эти мутации обладают различными характеристиками.
Мутация recA1снижает уровень гомологичной
рекомбинации, что обеспечивает большую стабильность вставки. И является
желательным, так как любой вектор, имеющий вставку может дестабилизироваться
общей рекомбинации в клетке реципиента.
Мутация deoR позволяет бактериям расти
на минимальной среде, которая содержит один источник углерода (инозин), а также
обеспечивает эффективную вставку больших фрагментов ДНК, что может быть полезно
при построении генных библиотек для больших фрагментов.
Мутация gyrA96 необходима для штаммов, используемых для
клонирования, так как продукт гена GyrА, ДНК-гиразы могут привести к делеции
между прямыми повторами, эти повторы могут произойти и в клонированной вставке
в плазмиде.
Мутация lacZДM15 деактивирует активность LacZ в
бактериях, которые продуцируют неактивную форму в - галактозидазы. Штаммы с
этой мутации не усваивают X-gal и остаются бесцветными на среде содержащей этот реагент. Но если
плазмида имеет LacZ альфа-субъединицу (например, PUC - плазмиды) и эту плазмиду
трансформируют в клетку, она дополняет ген LacZ и в - галактозидаза образуется.
Поскольку альфа - субъединицы находятся рядом с сайтом множественного
клонирования (MCS) векторов pUC, вставка в MCS будет нарушать альфа-субъединицу
и колонии, которые будут содержать данную вставку будут белые, а колонии без
вставки синие.
Этот штамм кишечной палочки был использован в недавних
исследованиях, для хранения цифровых данных[5], [8].
2.2
Среды и реактивы
Бактериальную культуру выращивали в на плотной питательной
среде (полноценный агар). Агаризованная среда содержала 1,5% агара, источником
углерода была глюкоза в конечной концентрации 0,2%. Аминокислоты, основания и
витамины добавляли в концентрации 20, 10 и 1 мкг/мл соответственно.
Питательный бульон
|
Питательный
бульон для культивирования микроорганизмов (панкреатический гидролизат рыбной
муки)
|
20 г.
|
|
Дистиллированная
вода
|
1 л
|
|
Доводили pH 90 7,0-7,2 Стерилизовали путём
автоклавирования при 0,5 атм 45 минут
|
Агаризованная полноценная среда (на основе рыбного
гидролизата)
|
Питательный
бульон для культивирования микроорганизмов
|
15 г.
|
|
Смесь
аминокислот
|
5 г
|
|
Агар-агар
|
15 г.
|
|
Дистиллированная
вода
|
1 л
|
|
pH до 7,0-7,2 Стерилизовали путём
автоклавирования при 0,5 атм 45 мин
|
ТЕ-буфер (pH 7.4) 50 мл
|
Состав раствора
|
Стоковый
раствор
|
Количество
|
|
10 мМ Tris pH 8.0
|
1М
|
0,5 мл
|
|
EDTA1
мМ pH 8.0
|
0,5М
|
0,1 мл
|
|
Дистиллированная
вода
|
|
до 50 мл
|
Отмывающий раствор
|
Состав раствора
|
Стоковый
раствор
|
Количество
|
|
Tris
25мМ pH 8.0
|
1М
|
12,5 мл
|
0,5М
|
10,0 мл
|
|
NaCl
150мМ
|
5М
|
15,0 мл
|
|
Дистиллированная
вода
|
|
До 500 мл
|
Раствор 1 для выделения плазмидной ДНК. (200 мл)
|
Состав раствора
|
Стоковый
раствор
|
Количество
|
|
Глюкоза 50 мМ
|
2 М
|
5 мл
|
|
EDTA
10мМ pH 8,0
|
0,5 М
|
4 мл
|
|
Tris
25 мМ pH 8,0
|
1,0 М
|
5 мл
|
|
Лизоцим E. coli 3 мг/мл
|
|
0,6 г
(добавлять перед использованием)
|
|
Дистиллированная
вода
|
|
до 200 мл (186
мл)
|
Раствор 2 для выделения плазмидной ДНК. (30 мл)
|
Состав раствора
|
Стоковый
раствор
|
Количество
|
|
NaOH
0.2 н
|
2н NaOH
|
3 мл
|
|
SDS
1%
|
SDS
10%
|
3 мл
|
|
Дистиллированная
вода
|
|
До 30 мл (24
мл)
|
Раствор 3 для выделения плазмидной ДНК. (100 мл) pH 4,8
|
Состав раствора
|
Количество
|
|
Ацетат калия
(натрия) 5М
|
60 мл
|
|
Ледяная
уксусная кислота
|
11,5 мл
|
|
Дистиллированная
вода
|
28,5 мл
|
X-gal 20 мг/мл
|
Состав раствора
|
Количество
|
|
Х-gal
|
65.2 мг
|
|
раствор DMFA
|
До 3,26 мл
|
IpTg100 мМ (23,83 мг/мл)
|
Состав раствора
|
Количество
|
|
IpTg
|
238.4
мг
|
|
Деионизированная
вода
|
до 10.4 мл
|
Приготовление 0,7% агарозы
|
Агароза
|
0,7г
|
|
TАЕ - буфер
|
100 мл
|
|
Нагреть до
полного расплавления агарозы, охладить до 50-60оС.
|
10Х ТАЕ - буфер pH = 7.6
|
Tris
основной
|
24,2 г
|
|
Ледяная
уксусная кислота
|
5,71 мл
|
10 мл
|
|
Дистиллированная
вода
|
До 500 мл
|
В работе использовался коммерческий препарат антибиотика
ампицилин в концентрации 10000 мкг/мл. Аммпицилин растворяли в стерильной воде.
Изопропилтио в - галактозид (IPTG) и в-галактозиаду (X-Gal) производства «MBI Fermentas» (Литва) готовили в
соответствии с рекомендациями изготовителя и использовали в концентрации 100
ммоль/л и 20 мг/мл соответственно.
.3
Методы исследования
Манипуляции
с ДНК
Тотальную ДНК использовали в качестве матрицы в полимеразной
цепной реакции. Рестрикцию ДНК и последующее лигирование фрагментов
осуществляли в условиях, рекомендуемых фирмой изготовителем ферментов «MBI Fermentas» (Литва). В качестве
реперной ДНК при определении размеров рестрикционных фрагментов использовали
синтетический репер 1 kb ladder (Fermentas, Литва))
Выделение
плазмидной ДНК
Плазмидную ДНК из бактерий E.coli выделяли методом щелочного
лизиса. Выделение производили из ночной культуры E.coli, которую выращивали при
37 ОС.
Использовали растворы следующего состава: лизирующие (буфер I: 50 мМ глюкоза, 10мМ EDTA, 25 мМ Tris, лизоцим в концентрации
3 мг/мл; буфер II: 0,2н NaOH, 1% SDS) и нейтрализующий (3М ацета K, pH4,8). ДНК осаждали
изопропанолом и ресуспендировали в 200 мкл ТЕ буфера (10 мМ Tris pH 8.0, 1 мМ EDTA pH 8.0), дальнейшую очистку
препарата ДНК вели с использованием12 моль/л LiCl.
Трансформация
бактерий
Трансформацию бактерий E.coli осуществляли кальциевым
методом. К клеткам предварительно переведённых в состояние компетентности
добавляли ДНК в концентрации до 1 мкг.
1) Бактериальную культуру выращивали в течении ночи на
питательном бульоне.
2) Ночную культуру разводили в 25 раз.
) Далее доращивали до log-фазы на качалке 200-250
оборотов 37 ОС до 2 часов.
) Бактериальную культуру переносим в стерильные
эппендерфы по 1,5.
5) Центрифугировали 5000 об/ мин 5 мин 4оС
6) Супернатант удаляли, осадок мягко ресуспендировали в
100 мкл 0,1М CaCl2. Эппендорфы оставляли на лядяной бане на 1 час для достижения
компетентности.
) К компетентным клеткам добавляли в концентрации ДНК
до 1 мкг и оставить в тех же условиях на 30-45 минут.
) Осуществляли температурный шок: 40оС - 30
сек + 2 мин на ледяной бане.
) В эппендорфы добавляли 1 мл питательного бульона, и
помещали на качалку на 30 мин, 200-250 оборотов 37оС для экспрессии.
) Центрифугировали 5000 об/мин 5 мин.
) Производили высев на чашки с селективной средой
содержащей ампициллин в концентрации 50 мкг/мл, IpTg в концентрации 0,5
ммоль/л, Х-gal в концентрации 0,1 мг/мл.
) Учёт результатов производили через 24 часа.
Электофоретический
анализ
Электофоретический анализ ДНК осуществляли общепринятыми
методами. Для продуктов использовали 0,7% агарозный гель и ТАЕ-буфер. Для
визуализации фрагментов ДНК использовали бромистый этидий (добавляли в буфер) в
конечной концентрации 0,5 мкг/мл. Для нанесения проб использовали буфер
следующего состава: 40% сахароза, 0,25% бромфенеловый синий. Гель фотографировали,
используя
Размер фрагментов ДНК устанавливали на основании их
электрофоретической подвижности в агарозном геле, в качестве реперной ДНК
использовали 1 kb DNA Ladder (Fermentas, Литва)
Полимеразная
цепная реакция
При постановке полимеразной цепной реакции (ПЦР) использовали
набор FermentasTM (Литва) и амплификатор Thermo scientific Pico 24 Thermal Cycler.
Реакционную смесь для ПЦР готовили в объёме 20 мкл, состав
реакционноё смеси представлен в таблице 2.
Таблица 2.2 - Состав реакционной смеси для полимеразной
цепной реакции
|
Компонент
|
Объём (мкл)
|
Исходная
концентрация
|
Конечная
концентрация
|
|
ДНК
|
1-4
|
-
|
0,1-5 нг/мкл
|
|
Праймер 1
|
1
|
10 мкмоль/л
|
0,5 мкмоль/мл
|
|
Праймер 2
|
1
|
10 мкмоль/л
|
0,5 мкмоль/мл
|
|
Буфер
|
2
|
10Х
|
1Х
|
|
Смесь dNTP
|
2
|
2 ммоль/л
|
0,2 ммоль/л
|
|
ДНК-полимераза
|
0,06
|
5 ед/мкл
|
0,0025 ед/мкл
|
|
mQ вода
|
до 20 мкл
|
-
|
-
|
Фрагмент бактериальной хромосомы B. subtilis размером 885 п.н.,
содержащий гены tmrB и aroK - амплифицировали с помощью праймеров ShikF-NotI (5’ - GCGGCCGCCATTAGTGTAAAGTGGTGAAC-3’) и ShikR-BamH1 (5’ - CCGGATCCGCCCAGAGATACGATTTTG-3’) при
режиме амплификации: 94 ОС - 5 минут (один цикл); 94 ОС -
30 секунд, 54 ОС - 5 секунд, 52 ОС-25 секунд, 94ОС-30
секунд, 54 ОС-5 секунд, 52 ОС-25 секунд, 68 ОС-
1 минута 30 секунд (10 циклов); 94 ОС-30 секунд, 54 ОС-30
секунд, 68 ОС- 1 минута 30 секунд, (20 циклов); 72 ОС - 5
минут. Праймеры содержали на 5’ - концах сайты для рестриктаз NotI и BamHI соответственно.
3.
Результаты и обсуждение
Наиболее перспективным методом получения шикимовой кислоты
является микробиологический синтез.
В общем пути биосинтеза ароматических соединений образующийся
шикимат подвергается фосфолированию под действием шикиматкиназы с образованием
шикимат-3-фосфата. Для накопления шикимата надо инактивировать данный фермент.
Данный фермент кодируется геном aroI, который расположен достаточно далеко от
остальных генов данного метаболитического пути. Одним из таких вариантов
является постановка гена aroI под регулируемый промотор в составе
бактериальной хромосомы. Данный метод позволяет регулировать уровень экспрессии
гена в зависимости от стадии культивирования штамма-продуцента: индуцировать на
ранних этапах роста культуры, позволяя ей проявлять свои прототрофные свойства
и блокировать на поздних, вызывая накопление шикимовой кислоты.
Для амплификации целевого фрагмента, ограниченного генами tmrB и aroK, также содержащего
регуляторные элементы указанных генов, кодирующие последовательности которых
расположены на «-» и «+» цепи ДНК соответственно, использовали праймеры
содержащие на своих 5’ концах навески с сайтами распознавания для рестриктаз BamHI и NotI.
Данный фрагмент амплифицировали с матриц штаммов В. Subtilis ВНИИгенетика-15 С10 и
ВНИИгенетика-15 D4, размер данного фрагмента должен составлять 885 п.н.
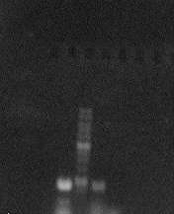
Рисунок 3.1 - Электрофореграмма продуктов амплификации.
- продукт амплификации по матрице ВНИИгенетика-15 D4
- маркер1 kb DNA Ladder
- продукт амплификации по матрице ВНИИ генетика-15 С10
Так как концентрация продукта амплификации с матрицы ВНИИгенетика-15
D4 была больше, этот
продукт использовался в дальнейшем работе.
Полученный продукт лигировали с ДНК плазмиды pMTL21C. C ДНК плазмиды
предварительно осуществляли рестрикцию по сайту рестрикции SmaI. Лигированной смесью
трансформировали клетки E. coli DH5a. Селекцию вели на полноценной питательной агаризованной
среде, содержащий ампициллин в концентрации 50 мкг/мл, X-Gal, IPTG. Из клонов
трансформантов, имеющих белую окраску выделяли плазмидную ДНК и проверяли путём
рестрикционного анализа по сайту рестрикции BamHI.
С данным продуктом рестрикции осуществляли полимеразную
цепную реакцию.
|
Наименование
реагента
|
Исходная
концентрация
|
Желаемое
количество / концентрация в конечном растворе
|
Объем реактива
(добавляемый в пробирку)
|
|
ДНК вставки
|
50 ng/µl
|
91,7 ng
|
1,3 µl
|
|
ДНК вектора
|
50 ng/µl
|
108,3 ng
|
2,2 µl
|
|
10x Ligation Buffer
|
10X
|
1X
|
2.0
µl
|
|
T4 ligase
|
10
u/µl
|
5 u
|
0.5 µl
|
|
H2O
|
до 20 µl
|
|
14,02 µl
|
|
Условия
реакции: реакционная смесь оставляется на ночь при 4-8оС) Для инактивации
фермента выдерживаем смесь 10 минут при 65°С.
|
Суммарное количество ДНК в лигазной смеси - 200 ng
Отношение вектор / вставка = 1/3
Пусть x - масса вектора в ng, у - масса вставки в ng.
x+y=200 (y/1000)/(x/3543)=3=200-y
y/1000=3*(200-y)/3543=91,7 нг x=108,3 ng
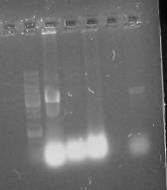
Рисунок 3.2 - Электофореграмма продуктов рестрикции по BamHI
1- маркер1 kb DNA Ladder
2- плазмида pMTL21C
- продукт рестрикции BamHI
- продукт рестрикции BamHI
- продукт рестрикции BamHI
- продукт рестрикции BamHI

Рисунок 3.3 - Электофореграмма продуктов амплификации по
матрице BamHI
-плазмида pMTL21C
2 - продукт амплификации по матрице BamHI
Заключение
В ходе данной работы были получены ампликоны с матриц с
матриц В. Subtilis ВНИИгенетика-15 С10 и В. Subtilis ВНИИгенетика-15 D4.
Осуществлено лигирование с ДНК плазмидой pMTL21C.
Осуществлена трансформация клеток E.coli DH5б.
Список использованной литературы
1. Бочков, Д.В. Исследование содержания
шикимовой кислоты в некоторых ратениях Алтайского края/ Д.В Бочков, С.В.
Сысолятин, А.И. Калашников, И.А. Сурмачёва // Химия растительного сырья 2011
№1. - с. 119-122
2. Бочков, Д.В. Поиск сырья для выделения
шикимовой кислоты/ Д.В Бочков, С.В. Сысолятин, А.И. Калашников, И.А. Сурмачёва,
А.А. Ламберова, А.С. Буянова, М.Э. Ламберова // Химия растительного сырья 2011
№3. - с. 81-87
3. Iwona Gientka. SHIKIMATE PATHWAY
IN YEAST CELLS: ENzYMES, FUNCTIONING, REGULATION - A REVIEW // Iwona Gientka,
Wanda Duszkiewicz-Reinhard // Рolish journal of food and nutrition sciences 2009, Vol. 59,
No. 2, pp. 113-118
. Fengli Chen. Extraction and
Chromatographic Determination of Shikimic Acid in Chinese Conifer Needles with
1-Benzyl-3-methylimidazolium Bromide Ionic Liquid Aqueous Solutions/ Fengli
Chen, Kexin Hou, Shuangyang Li, Yuangang Zu, and Lei Yang // Journal of
Analytical Methods in Chemistry Volume 2014 (2014), Article ID 256473, 12
pages.
. Malachi Griffith. Genetic
Modification of the Escherichia coli Strain DH5б to Allow the Selection of
Plasmids Carrying Complementary Yeast Genes // Projects in Biology 05.4111/6
Supervised by Dr. R.D. Gietz April 2, 2001
. Dewick, M. THE SHIKIMATE
PATHWAY:AROMATIC AMINO ACIDS AND PHENYLPROPANOIDS/ M. Dewick // Medicinal
Natural Products, 2002.
. Denis, V. Shikimic acid: review
of its analytical, isolation, and purification techniques from plant and
microbial sources/ Denis V. Bochkov, Sergey V. Sysolyatin, Alexander I.
Kalashnikov, and Irina A. Surmacheva // J Chem Biol. Jan 2012; 5 (1): c. 5-17.
8. Интернет-портал DH5-Alpha E.coli [Электронный ресурс] -
Режим доступа http://microbewiki.kenyon.edu/index.php/DH5-Alpha_E.coli.
. Интернет-портал Шикиматный путь
[Электронный ресурс] - Режим доступа
http://www.pandia.ru/text/77/454/37664.php.
. Интернет-портал Осельтамивир (Тамифлю)
[Электронный ресурс] - Режим доступа http://it-apharm.ru/oseltamivir.html.
. Интернет-портал Осельтамивир
(Oseltamyvir): инструкция, применение и формула [Электронный ресурс] - Режим
доступа http://www.rlsnet.ru/mnn_index_id_2869.htm
12. Hermann K.M. The
shirimatepathway:early steps in the biosynthesis of aromatic compounds. // The
Plant cell. -1995. Vol.7. - c. 907-919.
. Rangashari S., Fiedmay T.C.,
Hartvan G., Hemmiinghaus J.V., Kretzmer K.A. Processes for producing and recovering
shicimic acid.: пат. WO 2009/052303 А2 СI2P 7/42 (2006.01).-опублю 23.04.2009.
. Brouns K.D. Regulation of
aromatic amino acid biosyntesis in E.coli K12 // Genetics. - 1960. Vol.60. - c.
31-48
15. Panina E.M., Vitreschak A.G., A.A. Mironov and gelfand. Regulation
of Aromatic Amino acid Biosyntesis in Gamma - Proteobacteria // J. Mol.
Vicrobiol/ Biotechnol. - 2001.-Vol.33-/529-543
. Adachi O., Ano Y., Toyama H.,
Matsushita K. //High shikimate production from quinate with two enzymatic
systems of acetic acid bacteria. Biosci. Biotechnol. Biochem.2006. Vol.70.
17. Ambhaikar N. Shicimic acid //
Material of Group Meeting, 1.12.2005. - 2005.-7c.
. Frost J.W. Methods and materials
for production of shicimic acid.: patent US 2007/0087424 A1, C12P 7/42, C12N
1/21, C07H21/04? C12n 15/74.-опубликовано 19.04.2007
. Johansson L., Lindskog A.,
Silfversparre G., Cimander C., // Shicimic acid production by modified strain
of E.coli under phosphate-limited and carbon limited condition. //Biotechnology
and Bioengineering.2005. Vol.92. No.5., p541-552.
. Escalante A., Calderyn R.,
Valdivia A., Anda R., Hernandez G., Ramfirez O.T., Gosset G and Bolivar F.
Metabolic engineering for the production of shicimic acid in an evolved E.coli
strain lacking the phosphoenolpyruvate:carbohydrate phosphotransferaze system.
// Microbial Cell Factories. -2010.-Vol. 9. - c. 21.
. Iomantas Yurgis A.V., Abalkina
E.G., Polanuer B.M., Yampolskay T.A., Bachina T.A., Kozlov Y.B. //Method for
production shicimic acid // US6436664.2002-08-20.
. Chambers S.P., prior S.E.,
Barstov D.A., Minton N.P., The pMTL nic-cloning vectors. Improved pUC
polylinker regions to facilitate the the use of nucleotide sequencing //Gene. -
1988.-Vol.68, №1.-с. 139-149
23. Интернет-портал E. coli genotypes [Электронный ресурс] -
Режим доступа http://openwetware.org/wiki/E._coli_genotypes#DH5.CE.B1.