Екологічне обґрунтування кореляційних особливостей зв'язків між організмами та середовищем їх існування
ЗМІСТ
ВСТУП
. ВІДПОВІДНІСТЬ МІЖ ОРГАНІЗМАМИ Й СЕРЕДОВИЩЕМ
ІСНУВАННЯ
.1 Природний добір - адаптація або абаптація?
.2 Пристосування до змінних умов середовища
. ІСТОРИЧНІ ФАКТОРИ ФОРМУВАННЯ ВЗАЄМОЗВ'ЯЗКІВ МІЖ
СЕРЕДОВИЩЕМ ІСНУВАННЯ ТА ОРГАНІЗМАМИ
.1 Переміщення масивів суші
.2 Зміни клімату
.3 Особливості тваринного й рослинного світу островів
.4 Конвергенція й паралелізм
. ПОДІБНІСТЬ МІЖ УГРУПОВАННЯМИ Й ВІДМІННІСТЬ ФОРМ
УСЕРЕДИНІ УГРУПОВАНЬ
.1 Подібність між угрупованнями
.2 Розмаїття всередині угруповань
.3 Екологічна ніша
.4 Спеціалізація всередині видів
.4.1 Екотипи
.4.2 Генетичний поліморфізм
.5 Відповідність між організмами й середовищем, що
змінюється
.6 Пари видів
. РОЗРАХУНКОВА ЧАСТИНА. КІЛЬКІСНА ОЦІНКА ВИДОВОЇ
СТРУКТУРИ ПОПУЛЯЦІЙ
.1 Завдання
.2 Розрахунки
ВИСНОВКИ
ПЕРЕЛІК ПОСИЛАНЬ
ВСТУП
Кореляцією називається неповний зв'язок між досліджуваними
явищами. Це така залежність, коли будь-якому значенню однієї змінної величини
може відповідати декілька різноманітних значень іншої змінної. Під навколишнім
середовищем розуміють сукупність усіх умов, в яких існують організми.
Існують форми відповідності, які спостерігається між
організмами та середовищем, в якому вони живуть. Починати з твердження, що
кожен організм у деякому роді ідеально пристосований до існування там, де він
існує, було б поверхово і не надто оригінально. Можливо, в результаті
всебічного дослідження можливо прийти до такого висновку, але логічним
посиланням це положення служити не може. Тим не менше організми такі, які вони
є, і мешкають там, де вони мешкають, унаслідок своєї еволюційної історії.
Актуальність курсової роботи полягає в тому, що вивчення
зв’язків між організмами та середовищем їх існування дасть змогу робити певні
прогнози про стан організмів при зміні певних факторів середовища, використання
таких даних для збереження конкретних організмів, оптимізувати дію факторів для
культивування організмів в сільському господарстві і т.д.; передбачити вплив
деяких живих організмів на середовище, якщо він буде шкідливим або летальним.
Метою роботи є вивчення особливостей зв’язків між середовищем
існування та живими організмами.
Об’єктами вивчення є організми та їх оточуюче середовище.
1. Відповідність між організмами й
середовищем існування
Екологія вивчає організми і середовище їх існування, тому
дуже важливо зрозуміти зв'язок між ними. Можливо, з усіх тверджень щодо цього
зв'язку основним є наступне: різні організми розподілені по різних середовищах
існування не безладно - між організмами і середовищем є відповідність. Це
цілком укладається в наші уявлення про природу речей. Але яка природа цієї
відповідності?
1.1 Природний добір - адаптація або абаптація?
Поширений штамп, що вживається для опису цієї відповідності,
такий: «Організм X пристосований до...» - і далі йде опис умов, в яких подібні
організми зустрічаються. Так, наприклад, доводиться чути, що «риби пристосовані
до існування у водному середовищі», що «кактуси пристосовані до посушливих умов
пустель» і т. д., але, як не дивно, звичний штамп у таких випадках абсолютно
недоречний. Встановити, чому він недоречний - це, виявляється, найвірніший шлях
до розуміння справжньої природи відповідності між організмом і середовищем.
В основі теорії еволюції шляхом природного відбору,
запропонованої Чарльзом Дарвіном в 1859 р., лежить кілька припущень.
Індивідууми, що утворюють видову популяцію, не тотожні: вони
відрізняються, нехай навіть незначною мірою, за розмірами, за швидкістю
розвитку, по реакціях на температуру та інше.
Відмінності ці, хоча б частково, передаються у спадок. Іншими
словами, властивості індивідуума в якійсь мірі зумовлюються його генетичною
конституцією. Діти успадковують гени батьків і тому, як правило, поділяють
батьківські ознаки.
Потенційно будь-яка популяція в стані «заповнити землю»; так
би воно і сталося, якби кожен індивідуум виживав і до того ж залишав
максимальне число своїх нащадків. Але такого не трапляється: багато особин
гинуть, не встигнувши залишити потомство, а багато (якщо не всі) розмножуються
зі швидкістю, яка далеко не досягає максимальної.
Різні особини залишають різне число нащадків. Це зовсім не
зводиться до того, що у різних індивідуумів число нащадків в одному приплоді
неоднакове. Мається на увазі і ймовірність доживання особин до репродуктивного
віку, і число нащадків в одному приплоді, і виживання і розмноження цих
нащадків, і виживання і розмноження нащадків наступного покоління - і так до
нескінченності.
Нарешті, число нащадків даного організму залежить - не
цілком, не вирішальним чином - від результату взаємодії між цим організмом і
середовищем.
В одних умовах індивідуум виживає й розмножується, в інших -
ні. Тільки в цьому вельми наближеному сенсі і можна вважати, що природа
здійснює відбір. Саме в цьому сенсі певні місцезростання можуть бути названі
сприятливими або несприятливими, і в цьому ж сенсі одні організми можуть
вважатися пристосованими, а інші - ні. Оскільки одні особини залишають більше
нащадків, ніж інші, спадкові властивості популяції можуть в ряді поколінь
змінюватися. У цьому випадку говорять, що має місце еволюція за допомогою
природного відбору.
Мабуть, не позбавлене сенсу твердження, що особини даного
покоління «абаптовані» умовами, в яких жили попередні покоління. Умови минулого
- фільтр, через який деякі сполучення ознак просочилися в даний; але, мабуть,
організми адаптовані до умов нинішніх лише стільки, скільки останні зазвичай
подібні з умовами минулими. Термін «пристосування» («адаптація») залишає
помилкове враження якогось передбачення або принаймні задуму. Організми не
призначені, не адаптовані ні для сьогодення, ні для майбутнього і ні до
теперішнього, ні до майбутнього не пристосовані - вони являють собою живі
наслідки власного минулого. Вони абаптовані своїм минулим.
1.2 Пристосування до змінних умов середовища
Пристосованість - це відносний внесок особин в чисельність
майбутніх поколінь. Теорія природного відбору не пророкує досконалості.
Згідно з визначенням, найбільшою пристосованістю володіють ті
особини популяції, які залишають найбільше число нащадків. На практиці це
поняття нерідко застосовується не до окремої, а до типової особини або до
деякого типу: наприклад, можна сказати, що равлики з жовтими раковинами краще
пристосовані до проживання у піщаних дюнах, ніж равлики з бурими раковинами
(тобто перші з більшою, ніж другі, ймовірністю виживають і виробляють більш
численне потомство).
Пристосованість, все ж, поняття не абсолютне, а відносне.
Число утвореного рослиною насіння або відкладених комахам яєць не є прямою
мірою їх пристосованості; не є нею і саме по собі число залишених ними
нащадків. Пристосованість індивідуума швидше визначається тим відносним
внеском, який він вносить в чисельність майбутніх поколінь: найбільш
пристосовані особини популяції - ті, чисельність потомства яких найбільш велика
в порівнянні з чисельністю потомства, залишеного іншими, менш пристосованими
особинами тієї ж популяції. Особини ж, що вносять найбільший відносний внесок в
чисельність виробленого популяцією потомства, надають і найбільший вплив на її
спадкові ознаки.
Жодна популяція не в змозі вмістити всі можливі генетичні
варіанти, які могли б існувати, впливаючи при цьому на пристосованість. Зі
сказаного випливає, що природний відбір навряд чи призведе до виникнення
досконалих, «максимально пристосованих» індивідуумів. Він сприяє особинам,
найбільш пристосованим з числа існуючих, а такий вибір може виявитися вельми і
вельми обмеженим. Могутність рушійних сил еволюції далеко не безмежна. Теорія
Дарвіна не пророкує виникнення досконалості - навіть у середовищі, що
залишається з покоління в покоління незмінним. З неї випливає лише те, що деякі
особини звичайно залишають більше нащадків, ніж інші, і з цієї причини роблять
найбільший вплив на формування зовнішності майбутніх поколінь.
Вказане вище являє собою основне обмеження, що накладається
на ступінь пристосованості організмів до умов існування. Сама суть природного
відбору така, що організми досягають пристосованості до середовища проживання
через стан «найбільш пристосованих з числа існуючих» або «найбільш
пристосованих з числа вже існуючих»: вони ні в якому разі не «кращі з мислимих».
Ми не плекаємо надій на відшукання в природі досконалості. Причини цього
докладно обговорюються в роботах Джекоба, Гоулда і Левонтін і Харпера.
Крім того, способи пристосування і ступінь пристосованості
організмів до навколишнього середовища підкоряються і іншим обмеженням і
умовам.
2. Історичні фактори формування
ВЗАЄМОЗВ'ЯЗКІВ між середовищем існування та організмами
Винятково важливо усвідомити, що сьогодення нерідко несе за
собою глибокий слід геологічного минулого. Наш світ не був створений творцем,
що перебрав по черзі всі організми «випробував» їх у всіляких умовах і визначив
кожному належне місце. Світ влаштований так, що сучасне поширення живих істот
можна, хоча б частково, віднести на рахунок вигадок історії.
2.1 Переміщення масивів суші
Деякі загадкові картини розселення організмів по материках
представляються незрозумілими з точки зору можливостей переміщення на великі
відстані. Подібні приклади змусили біологів - але насамперед німецького
геофізика Альфреда Вегенера - припустити, що переміщатися повинні були самі
материки. Гіпотеза Вегенера з обуренням відкидалася геологами, але лише до тих
пір, поки результати вивчення палеомагнетизму не змусили їх вдатися до того ж
самого пояснення - на перший погляд фантастичного. Геологів примирила з
біологами концепція рухливих тектонічних плит земної кори, з якої випливає і
рухливість материків (рисунок 2.1). Отже, поки в рослинному і тваринному
царствах відбувалися великі еволюційні події, популяції розділялись на окремі
частини, а масиви суші перетинали цілі кліматичні пояси.
Рисунок 2.2 ілюструє один повчальний приклад поширення
великої групи великих нелітаючих птахів. Їх поширення піддається осмисленню
лише у світлі гіпотези вказаної вище. Твердження про те, що ему і казуар
займають свої ареали тому, що найкращим чином відповідають умовам Австралії, а
нанду і тинаму - тому, що саме вони найкраще пристосовані до умов Південної
Америки, були б безпідставні. Кожна з цих груп виникла і розвивалася в межах
свого континенту і «абаптована» своїм тамтешнім минулим, але утворення їх
ізольованих і віддалених один від одного ареалів цілком зумовлено доісторичним
переміщенням материків. Ці переміщення спричинили за собою неможливість
потрапляння одних нелітаючих птахів в місця проживання інших.
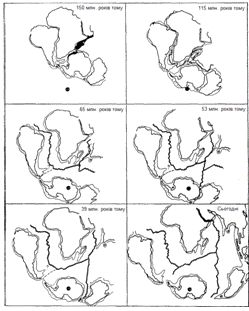
Рисунок 2.1 - Реконструкція послідовних стадій розколу
континенту Гондвана. Показано відділення південних континентів і утворення
Південної Атлантики та Індійського океану.
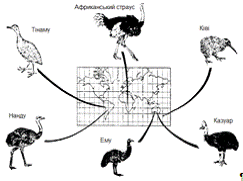
Рисунок 2.2 - Географічне поширення великих нелітаючих птахів
2.2 Зміни клімату
Клімат змінювався набагато швидше, ніж пересувалися масиви
суші, і в сучасній картині географічного поширення організмів багато відображають
певні фази відновлення, що відбуваються після кліматичних зрушень. Для
пояснення того, як виникли ареали тих або інших видів, відомості про історичні
зміни клімату необхідні в тій же мірі, що і уявлення про пристосованості
організмів до сучасних умов існування. У ролі історичного чинника в таких
випадках виступають, зокрема, плейстоценові обледеніння.
Класичні приклади видових ареалів, для тлумачення яких може
знадобитися залучення історичних обставин, - це що складаються з одного або
декількох невеликих відособлених ділянок ареали ряду холодостійких, часто
високоспеціалізованих квіткових рослин аркто-альпійських флор Північної Америки
і півночі Європи. Багато з таких рослин зустрічаються в одному-єдиному місці,
інші населяють два або декілька химерно розкиданих відокремлених «плям» (тобто
мають розірвані ареали - біцентричний або поліцентричний).
Інша точка зору полягає в тому, що сучасні ареали - це
релікти, тобто залишки ареалів популяцій, колись поширених більш широко. Коли з
півночі наступав льодовиковий щит, деякі високоширотні області залишалися
вільними від льоду (хоча і вони сильно промерзали). У цих необледенілих
областях (ескімоською - «нунатаки») збереглися популяції деяких рослин, а коли
льодовик відступив, вони так і залишилися на своїх відокремлених твердинях.
Можна знайти і геологічні свідчення, що підтверджують цю точку зору; вони,
правда, не цілком переконливі. Є також цікаві відомості про особливості
поширення деяких нелітаючих жуків до передбачуваних нунатаків, що є доказом на
користь «реліктової» гіпотези.
Втім, суперечка про нунатаки - це лише невелика частина
набагато більш широкої дискусії між тими, хто вважає, що сучасне поширення
організмів відображає їх відповідність сучасним умовам існування, і тими, хто
вважає, що багато чого в поширенні являє собою спадщина минулого. З
удосконаленням методів пошуку, аналізу і датування біологічних залишків
(особливу роль при цьому відіграє аналіз викопного пилку) розмах кліматичних і
біотичних зрушень, що відбувалися протягом останніх 2 млн. років (тобто в
плейстоцені), тільки починає відкриватися. Ці нові методи дозволяють все
частіше і все з більшою впевненістю встановлювати, що саме в сучасній картині
географічного поширення організмів пов'язано з вузькою спеціалізацією по
відношенню до нинішніх умов їх середовищ існування, а що являє собою відбиток
минулого.
Результати вимірювання вмісту ізотопів кисню в кернах порід
океанічного дна вказують, що в плейстоцені могло бути до 16 циклів обледеніння,
кожен з яких тривав приблизно 125 тис. років. На рисунку 2.3 показано приблизну
динаміку зміни температур за останні 400 тис. років. Оцінка температур отримана
шляхом порівняння співвідношень ізотопів кисню у викопних залишках організмів,
витягнутих з кернів донних відкладень Карибського моря. Штрихова лінія
відповідає температурі, що мала місце 10 тис. років тому - на початку сучасного
потепління. Крива показує, що теплі міжльодяникові, подібні сучасному
траплялися рідко.
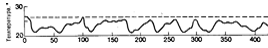
Рисунок 2.3 - Приблизна динаміка температури протягом
послідовних циклів заледеніння, що припали на останні 400 тис. років.
Всупереч поширеній думці, складається враження, що кожне
заледеніння могло затягуватися на цілих 50 і навіть 100 тис. років і що
заледеніння чергувалися нетривалими періодами в 10-20 тис. років, коли
температури піднімалися до сучасних значень або трохи вище. Якщо такий часовий
масштаб відповідає істині, то незвичайний не древній, а сучасний тваринний і
рослинний світ - адже він склався до кінця одного з нечастих катастрофічних
потеплінь!
В додатку 1 показана швидкість зміни післяльодовикового
рослинного покриву околиць оз. Роджерс (шт. Коннектикут) за останні 14 тис.
років за даними пилкового аналізу. Види дерев, пилок яких переважає у
відповідних пилкових профілях, з'являлися почергово: першою була ялина,
останнім - каштан. Кожен наступний поселенець подовжував список видів,
кількість яких неухильно зростала протягом всіх 14 тис. років. Подібна картина
складається і при вивченні пилкових профілів донних відкладень європейських озер.
З ростом числа розшифрованих «пилкових літописів» стало
можливо не тільки скласти уявлення про динаміку рослинного покриву в даному
місці, але і приступити до картографування переміщень різних видів рослин у
міру їх поширення по материках. З'ясувалось, що різні види просувалися з
різними швидкостями, і при цьому не обов'язково в одному і тому ж напрямку.
Рівновага рослинних угруповань тим часом порушувалася, склад їх змінювався, і
зміни ці майже напевно тривають донині.
У Північній Америці першим з «загарбників», котрі рвонули
слідом за відступаючим льодовиком, була ялина; за нею послідкувала смолиста
сосна, вони протягом декількох тисячоліть просувалися на північ зі швидкістю
350-500 м в рік. Веймутова сосна «рушила» приблизно тисячоліттям пізніше-одночасно
з дубом. До числа розторопних переселенців належала і тсуга; вона поширювалася
зі швидкістю 200-300 м в рік і з'явилася у більшості вивчених точок приблизно
через тисячу років після Веймутової сосни. Каштан просувався повільно (100 м в
рік), але, діставшись до нових місць, ставав домінантним видом; на Аллеганах
він з'явився приблизно за 3000 років до того, як досяг Коннектикуту. (Для
рослини з важким насінням і з тривалою фазою ювенільного зростання, що передує
його утворенню, просування навіть на 100 м у рік видається на диво швидким).
Під час відступу льодовика швидкість заселення деякими рослинами виникаючих
можливих місць життя, можливо, лімітувалась швидкістю поширення їх насіння.
По повноті відомостей про зміни клімату тропіки набагато
поступаються помірній зоні. З цієї причини виникає спокуса припустити, що в ті
часи, коли помірні широти знаходилися у владі драматичних змін клімату і
льодовикових вторгнень, тропіки перебували в тому ж стані, що і нині.
Складається картина змін рослинного покриву, паралельних тим, що відбувалися в
помірному поясі: в більш теплі і вологі періоди площа тропічних лісів зростала,
а в періоди більш прохолодні і посушливі, коли переважала савана, -
скорочувалася. У сучасному поширенні як рослин, так і тварин містяться вказівки
на те, яке положення вони займали колись (Рисунок 2.4).
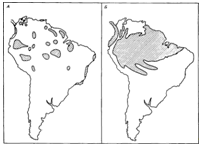
Рисунок 2.4 - А. Приблизне поширення тропічних лісів в
Південній Америці на піку останнього зледеніння. Б. Їх сучасне поширення.
2.3 Особливості тваринного й рослинного світу островів
Фауна і флора островів (будь вони оточені водами океану або
«морем» рослинності, що відрізняється) володіють декількома особливостями, що
відрізняють їх від фауни і флори материків. Стосовно ж до питань, обговорюваних
у цій главі, особливості острівних біот пояснюють три важливих взаємозалежних
положення, а саме:
) відповідно між організмом і середовищем присутня історична
складова;
) варіант «досконалого організму», відповідний даному типу
умов існування, не обов'язково є єдиним;
) природний відбір, впливаючи на абсолютно різні організми,
здатний приводити їх у відповідність з однотипними умовами проживання.
Просто кажучи, фауна і флора островів володіють двома
відмінними рисами: на островах менше видів, ніж на порівнянних материкових
ділянках тих же розмірів; крім того, багато з острівних видів злегка або навіть
суттєво відрізняються від видів, що населяють найближчу порівнювану ділянку
материка. Основних причин тому дві. По-перше, фауна і флора острова обмежені
формами, предки яких зуміли до цього острова дістатися; сила цього обмеження, звичайно,
визначається віддаленістю острова від материка або від інших островів і різна
для різних груп організмів в залежності від властивої цим організмам здатності
до розселення. По-друге, через ту ж саму відособлену швидкість еволюційного
перетворення острівної популяції може виявитися достатньою для того, щоб
подолати наслідки обміну генетичним матеріалом з батьківською популяцією на
материку.
Розтлумачимо другий пункт більш докладно. Біологи визнають
організми належать до одного й того ж виду, якщо ці організми здатні
схрещуватися, вільно обмінюючись при цьому генетичним матеріалом і виробляти
плідне потомство. Такий обмін генами (як, і перетасування генів у результаті
рекомбінації) сприяє згладжуванню генетичної неоднорідності популяції. Навпаки,
репродуктивна ізоляція (наприклад, подібна до тієї, що існує між острівною і
материковою популяціями) зазвичай створює передумови еволюційного становлення
більш локалізованих, суто місцевих відповідностей між організмами і
середовищем. Дійсно, репродуктивна ізоляція - це, мабуть, істотний крок на
шляху до розщеплення єдиного предкового виду тварин на два. Ця обставина, поза
всяким сумнівом, дуже багато дає для розуміння того, чому на островах багато
ендемічних видів, а також особливих «рас» або «підвидів», що відрізняються від
материкових форм, але не настільки, щоб іменуватися окремими видами.
Один із найяскравіших прикладів острівного видоутворення
являють плодові мухи роду Drosophila, що населяють Гавайські острови. Цей же
приклад, безсумнівно, найкращим чином вивчений в генетичному відношенні.
Гавайський острівний ланцюг - вулканічного походження,він формувалася поступово
протягом останніх 40 млн. років у міру того, як центральна частина
Тихоокеанської літосферної плити, потроху зрушуючи в північно-західному напрямку,
проходила над магматичним вогнищем (таким чином, о. Нііхау - найдавніший з
Гавайських островів, а сам о. Гавайї - наймолодший). На рисунку 2.5 зображено
Гавайський острівний ланцюг, простежено родинні відносини між видами шляхом
вивчення поперечної смугастості хромосом. Найдавніші види - D.primaeva (вид 1)
і A attigua (вид 2), вони зустрічаються лише на найдавнішому з заселених
дрозофілами островів (Нііхау). Ці види, як і всі інші з числа нині існуючих,
позначені чорними кружечками; види гіпотетичні, існування яких в минулому
доводиться допустити для встановлення зв'язків між видами нині існуючими,
позначені білими кружечками. Позначення кожного виду поміщено над тим островом
(або островами), де він зустрічається (при цьому острова Молокаї, Ланаї і Мауї
об'єднані в одну загальну групу). На островах Нііхау і Кахоолаве дрозофіл
немає. Перший поселенець з роду Drosophila, мабуть, досяг Гавайського
архіпелагу 40 млн. років тому - тоді, коли жодного з теперішніх островів ще не
існувало.
Різноманітність гавайських дрозофіл разюча: у всьому світі,
мабуть, налічується близько 1500 видів роду Drosophila, але з них щонайменше
500 зустрічаються виключно на Гавайських островах. Таке різноманіття, мабуть,
хоча б частково пояснюється тим, що Гавайї рясніють «островами на островах»: у
багатьох місцях потоки лави відгородили ділянки рослинності (такі ізоляти
називаються «кіпука»).
Особливий інтерес представляють приблизно 100 видів
«узорчатокрилих» дрозофіл. Генеалогічні зв'язки цих мух можна простежити,
вивчаючи поперечну смугастість гігантських хромосом у клітинах слинних залоз їх
личинок.
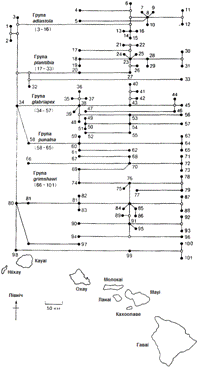
Рисунок 2.5 - Еволюційне генеалогічне древо Drosophila
Гавайських островів.
Відповідне «еволюційне древо» зображено на рисунку 2.5, кожен
з видів вказаний над зображенням того острова, на якому він зустрічається
(видів, що зустрічаються більш ніж на одному острові, тільки два). Історична
складова в тому, «хто де живе», в наявності: більш древні види населяють і
більш давні острова, а в міру утворення нових островів до них добиралися
небагато переселенців, які давали початок новим видам. Принаймні деякі з цих
видів, судячи з усього, відповідають тим же самим умовам існування, яким на
інших островах відповідають інші види. Прикладом можуть бути два
близькоспоріднених види -D. adiastola (вид 8) і D. setosimentum (вид І): перший
зустрічається тільки на о. Мауї, а другий - тільки на о. Гавайї, але умови, в
яких вони мешкають, явно ідентичні. Найцікавіше, звичайно, те, яку важливу роль
ізоляція, супроводжувана природним відбором, грає в утворенні нових видів (і,
таким чином, нових відповідностей між організмами і середовищем).
Найвідоміший приклад острівної еволюції і острівного
видоутворення - знамениті дарвінові в'юрки Галапагоських островів. 13 видів
в'юрків складають приблизно 40% загального числа видів птахів, що населяють
острови. В'юрки займають найрізноманітніші середовища існування і споживають
найрізноманітнішу їжу,що зображено на рисунку 2.6; всі вони, проте, полягають у
близькій спорідненості і швидше за все походять від якогось загального предка,
що нагадував сучасного в'юрка, що мешкає на о. Кокос.
На відокремлених Галапагоських островах природний відбір
впливав на той матеріал, якому судилося туди потрапити, і приводив до
виникнення організмів, які відповідають таким умовам, яким в інших місцях
відповідають організми абсолютно інших видів.
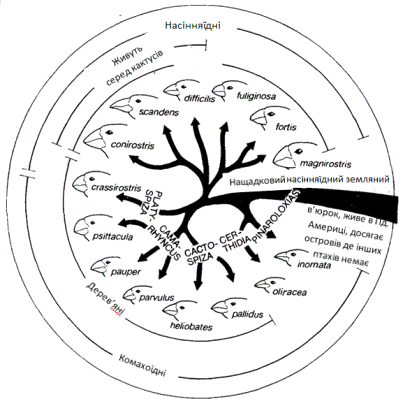
Рисунок 2.6 - Дарвінові в'юрки поїдають найрізноманітнішу їжу
і населяють найрізноманітніші місцеперебування; настільки ж різноманітні за
своєю формою, незважаючи на близьку їх спорідненість і схожість дзьобів.
2.4 Конвергенція й паралелізм
Відповідність між організмами і середовищем нерідко
проявляється у подібності будови та способу життя організмів, що мешкають в
подібних умовах, але належать до різних філетичних ліній (тобто до різних гілок
еволюційного дерева). Явища подібної схожості до того ж сприяють спростуванню
уявлення про те, ніби кожному типу середовища існування відповідає один і
тільки один варіант «досконалого організму».
Свідоцтва виникнення подібності особливо переконливі, коли
відповідні філетичні лінії видалені, а подібними функціями наділені структури
зовсім різного еволюційного походження, тобто структури аналогічні, а не
гомологічні. Коли спостерігається таке явище, ми говоримо про конвергентну
еволюцію.
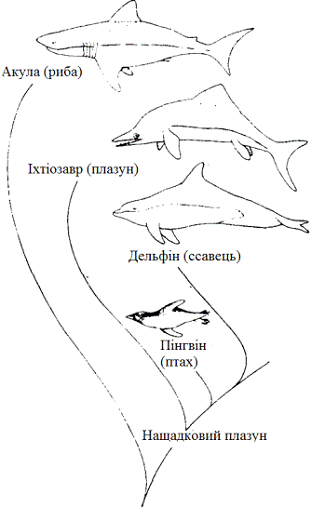
Рисунок 2.7 - Приклад конвергентної еволюції: схожість
зовнішньої будови тіла великих морських хижаків, що представляють різні
генеалогічні лінії тваринного царства.
Великі водні хижаки виникли в чотирьох абсолютно різних
групах: серед риб, плазунів, птахів і ссавців. Подібність зовнішньої будови,яка
зображена на рисунку 2.7, тим більш чудово, що вона приховує глибокі
відмінності внутрішньої будови та обміну речовин, що свідчать про настільки ж
глибокі відмінності еволюційної історії зображених організмів.
Насіння багатьох вищих рослин поширюється птахами. Птахів
приваблює наявність м'ясистої цукристої тканини, яку вони поїдають і
перетравлюють. Під цією тканиною насіння, захищене товстими оболонками; воно
проходять через травний тракт неушкодженим, а іноді перебування в ньому навіть
стимулює його проростання. У різних представників рослинного царства м'якоть
плодів бере своє походження від абсолютно різних структур. Ряд прикладів
наведено на додатку 2. Багато з зображених рослин належать до далеко віддалених
один від одного філогенетичних ліній, наприклад тисс - голонасінна рослина, а
всі інші - покритонасінні. Втім, навіть у межах одного лише сімейства розоцвітих
для утворення привабливої м'ясистої оболонки насіння «задіяні» вельми
різноманітні структури. Суниця, яблуня, персик - всі вони утворюють привабливу
для птахів м'якоть і (або) захищають своє насіння абсолютно різними способами.
Класичний приклад такого роду паралельної еволюції -
адаптивна радіація плацентарних і сумчастих ссавців. Сумчасті потрапили в
Австралію в крейдяний період (близько 90 млн. років тому), коли єдиними
ссавцями, що мешкали там були представники своєрідної групи однопрохідних, або
яйцекладущих (в даний час представленої лише єхидною і качкодзьобом). Потім в
еволюції їх послідкувало виникнення безлічі форм (радіація), у багатьох
відношеннях в точності повторило процеси еволюційної радіації ссавців на інших
континентах. На рисунку 2.8 наведені приклади екологічних відповідностей між
сумчастими і плацентарними ссавцями. І зовнішня будова, і спосіб життя тварин
повторюються з такою вражаючою детальністю, що мимоволі виникає відчуття, ніби
середовище проживання плацентарних і сумчастих ссавців складалася з
«екологічних ніш», в які еволюція акуратно «вставила» відповідні екологічні
еквіваленти. (Важливо пам'ятати, що в протилежних групах, які витерпіли
конвергентну еволюцію, у сумчастих і плацентарних радіація почалася з одним і
тим же набором обмежень і можливостей, оскільки обидві групи походять від
спільних предків.)
Можна припустити, що настільки разючу подібність зовнішньої
будови ряду рослин (найчастіше аж ніяк не споріднених) відображає настільки ж
разюча подібність умов, в яких вони виростають. Насправді ж ці рослини
«вмонтовані» в абсолютно непорівнювані рослинні співтовариства показані в
таблиці 2.1. Треба думати, високогір'я, а часто і острови, де ростуть мегафіти
з числа дводольних, мають мало спільного як з підлісками, до складу яких
входять пальми і деревоподібні папороті, так і з посушливим місцем проживання
багатьох алое і агав.
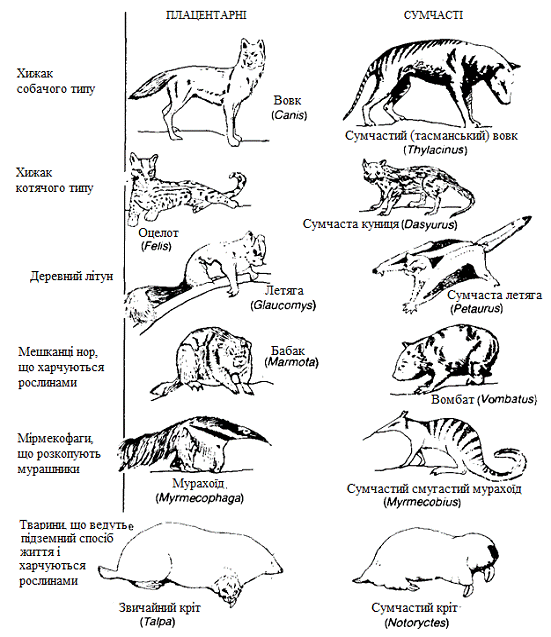
Рисунок 2.8 - Паралельна еволюція сумчастих і плацентарних
ссавців. Тварини зіставлених видів нагадують один одного як за зовнішнім
виглядом, так і за особливостями способу життя.
Ймовірно, схожість всіх цих рослин можна частково пояснити
наступним чином. Кожне з них має набір узгоджених ознак, що виявляються
одночасно; відбір ж у кожному окремому випадку впливав лише на одну з цих
ознак, причому не обов'язково на одну і ту ж. Можливо, чим менше дана рослина
схильна до розгалуження, тим крупніші органи, що розвиваються на його гілках.
Можна допустити, що відбір на укрупнення суцвіть з неминучістю призводить і до
укрупнення листя, що сприяє зміцненню стебла. Може бути, в інших середовищах
існування відбір на крупнолистість настільки ж неминуче тягне за собою
збільшення розмірів суцвіть.
Таблиця 2.1 Мегафіти («деревця з верхівковими листовими
розетками») і їх місце проживання
|
Деревоподібні папороті
|
Тропічний або субтропічний ліс - зазвичай підлісок
|
|
Саговники
|
Тропічний або субтропічний ліс - зазвичай підлісок
|
|
Покритонасінні
|
|
|
Однодольні
|
|
|
Пальми
|
Тропічний або субтропічний ліс - зазвичай підлісок
|
|
Liliaceae і Amaryllidaceae (лілійні і амарилісові),
наприклад Yucca, Agave, Aloe, Cordyline, Dracaena
|
Тропіки або субтропіки - багато проростає в
посушливих областях і пустелях, а інші утворюють підлісок тропічних або
субтропічних лісів
|
|
Xanthorrhoeaceae (трав'яні дерева)
|
Посушливі області - на відкритих місцях або в
підлісках
|
|
Дводольні
|
|
|
Boraginaceae (бурачникові), наприклад Echium
|
Канарські острови
|
|
Campanulaceae (дзвоникові), наприклад деревоподібне
Lobelias
|
Гавайські острови і гори Африки
|
|
Compositae (складноцвіті), наприклад Espeletia,
Senecio
|
Високогір’я Анд і гори Африки
|
Практика рослинництва, зрозуміло, має в своєму розпорядженні
приклади, що в якійсь мірі підтверджують цю точку зору. Культурний соняшник
виведений з диких предкових форм, для яких характерні часте галуження,
дрібнолистість, наявність численних дрібних суцвіть і невеликі розміри насіння.
Культурна форма соняшнику виведена в результаті відбору на крупнонасінність
(великі насіння легко збирати). У цього соняшнику одне величезне суцвіття,
нерозгалужене стебло і дуже великі листи; у багатьох відношеннях він теж
перетворився на типовий мегафіт. Різноманітних і глибоких формоутворюючих
зрушень добилися селекціонери і в межах роду Brassica. З розгалужених, відносно
мілколистих диких форм виведені кольорова і спаржева капуста - справжнісінькі
мегафіти з масивним і міцним нерозгалуженим стеблом, що має дуже великі листи і
величезне суцвіття. У іншого представника роду Brassica висота стебел досягає
2-2,5 м; на їх вершинах розташовуються мутовки великих листів. Ці нерозгалужені
стебла розвинені настільки, що використовуються в якості сировини для
промислового виробництва палиць! У всякому разі, яка б не була причина
подібності мегафітов, природу відповідних селективних сил зрозуміти важче, ніж
при зіставленні плодів або великих водних хижаків.
Ще одне ускладнення виникає через те, що явища конвергенції і
паралелізму найчастіше орієнтуються по вражаючому зовнішній схожості. І хоча
сумчастий кролячий бандікут і плацентарний кролик дуже схожі один на одного (разючу
подібність їм надають довгі вуха) і обидва влаштовують нори, бандікут споживає
головним чином тваринну їжу (личинок комах), а плацентарний кролик - рослинну.
Навпаки, кенгуру своєю зовнішністю навряд чи нагадує вівцю, але при цьому як
кенгуру, так і вівця - великі травоїдні тварини, при спільному проживанні
харчуються майже однієї і тієї ж їжею.
Для позначення групи видових популяцій, схожим чином
використовують однотипні ресурси середовища проживання, Рутом введено термін
«гільдія». Популяції, що мало нагадують один одного - кенгуру та вівці,
очевидно, входять до складу або одних і тих же, або аналогічних гільдій. Кролик
і бандікут придбали зовнішню схожість, але їх популяції належать до зовсім
різних гільдій. Іншими словами, є вагомі підстави стверджувати, що в певному
сенсі відповідність між організмами та середовищем іноді, можливо, проявляється
не стільки в зовнішній схожості, скільки у подібності трофічній (у тому, чим
тварини харчуються і хто харчується ними).
3. Подібність між
угрупованнями й відмінність форм усередині угруповань
Явища конвергентної і паралельної еволюції переконують у
тому, що простий список видів, родів і навіть родин, представлених в двох даних
областях, може практично нічого не повідомити про екологічну подібність цих
областей. Крім того, хоча наявність певних видів часто свідчить про певну
екологічну ситуацію, ці види майже завжди входять до складу різноманітних
спільнот, утворених формами,що дуже сильно розрізняються. Лише у виняткових
випадках в природі зустрічаються співтовариства, які з одної єдиної популяції
якогось унікального виду. У переважній більшості випадків природні угрупування
включають цілу гаму життєвих форм з різноманітними біологічними властивостями.
З цих причин прийнятне тлумачення відповідностей між організмами і середовищем
не повинно зводитися лише до «висмикування» з різних угрупувань організмів, що
сильно нагадують один одного зовнішнім виглядом і способом життя. Таке
тлумачення має бути також в змозі співвіднести з особливостями середовища
особливості цілих спільнот і пояснити не тільки схожість, але і відмінності між
організмами, що живуть в одних і тих же умовах.
3.1 Подібність між угрупованнями
Класичний тип рослинності, подібний «середземноморському»
маквису, можна виявити як по берегах Середземного моря, так і в Каліфорнії,
Чилі, Південній Африці і Південній Австралії. Всі ці області характеризуються
майже однаковими кліматичними умовами. Однотипність рослинного покриву стає
очевидною як при вивченні аерофотознімків, так і при поверхневому огляді
ділянок місцевості з швидко проїжджаючого автомобіля; проте таксономічні списки
відзначаються в цих областях видів (і навіть представлених ними сімейств)
ніяких вказівок на схожість не містять. Описати і виміряти цю подібність
виявилося надзвичайно важко. Часто-густо вона проявляється в «архітектурі»
різних рослин, а її не так-то просто охарактеризувати кількісно. У цьому
полягає причина частого вживання розпливчастих, якісних термінів.
Найпростішою, а у багатьох відношеннях і найбільш задовільною
класифікацією життєвих форм рослин, не враховуючи їх систематичного положення,
досі залишається класифікація, запропонована датським ботаніком Раункиер. Ріст
пагонів вищих рослин визначається закладкою меристем в точках росту, і Раункиер
підрозділив рослини відповідно до того, де розташовані і як захищені їхні
верхівкові меристеми, що показано в таблиці 3.1. В додатку 3 наведено кілька
прикладів «спектрів Раункиера» для флор різних областей; вони зіставлені зі
спектром, отриманим Раункиером і зараз (тропіки були відносно мало вивчені).
Впадає в очі разюча схожість між типами рослинності різних областей; воно
показує, що середовищі проживання можуть відповідати не тільки організми, а й
цілі денотичні комплекси.
Таблиця 3.1 - Класифікація життєвих форм рослин по Раункиеру
|
Фанерофіти
|
Бруньки поновлення або верхівкові точки росту
розташовані на повітряних пагонах а) Вічнозелені рослини без ниркових луски
б) Вічнозелені рослини з нирковими чешуями в) Рослини з обпадаючим листям і з
нирковими чешуями г) Рослини, висота яких не перевищує 2 м
|
|
Хамефіти
|
Бруньки поновлення або верхівкові точки росту
розташовані на приземних пагонах або на приземних частинах пагонів а)
напівчагарникові хамефіти, тобто рослини, що викидають прямостоячі пагони,
щорічно відмирають від вершини до тієї їх частини, на якій знаходяться
бруньки відновлення б) пасивні хамефіти зі слаборозвиненими невідмираючими
пагонами, що стеляться над поверхнею ґрунту або лежать на ній в) активні
хамефіти, пагони яких стеляться над поверхнею ґрунту або лежать на ній, так
як вони не відмирають і ростуть в горизонтальному напрямку г) подушкоподібні
рослини
|
|
Гемікриптофіти
|
Бруньки поновлення або верхівкові точки росту
розташовані безпосередньо під поверхнею ґрунту а) потогемікриптофіти з
нормально облистяними повітряними пагонами; верхні листки кілька
недорозвинені б) напіврозеткові рослини, велика частина листя яких
розташована на приземних частинах пагонів з укороченими міжвузлями; ці листи
бувають найбільшими в) розеткові рослини, все листя яких зосереджене в
прикореневій розетці
|
|
Криптофіти
|
Бруньки поновлення або верхівкові точки зростання
знаходяться в ґрунті (або у воді) а) Геокриптофіти або геофіти, що включають
форми, що володіють 1) кореневищами, 2) цибулинами, 3) стебловими бульбами,
4) кореневими бульбами б) Болотні рослини (геліофіти) в) Водні рослини
(гідрофіти)
|
|
Терофіти
|
.2 Розмаїття всередині угруповань
Існує безліч пояснень різноманітності, що має місце всередині
спільнот.
а) У природі не буває однорідних місць існування. Навіть
однорідний на перший погляд внутрішній простір лабораторної
пробірки неоднорідний у силу неминучої наявності кордону - скляної стінки. З
цією обставиною часто стикаються мікробіологи, коли вирощувані ними організми
підрозділяються на дві форми: пристінну і ту, що залишається в товщі
культурального середовища. Ступінь неоднорідності середовища залежить від
розмірів організму, що сприймає її. Поруч з гірчичним насінням грудочка ґрунту
- гора, а для гусениці один-єдиний листок може стати довічним прожитком.
Проростання насіння,що лежить в тіні листка, може затримуватися, тоді як
насіння, що лежить за межами тієї ж самої тіні, може без перешкод прорости. Те,
що з боку представляється екологу однорідним середовищем, для організму, що
живе в ньому, може виявитися мозаїкою з абсолютно непридатних і цілком
прийнятних ділянок.
б) У зв'язку зі сказаним слід зауважити, що всередині
більшості (можливо, всіх) місцеперебувань маються градієнти умов або доступних
ресурсів. Ці градієнти бувають просторовими або тимчасовими; останні в свою
чергу можуть бути циклічними (зв'язаними, наприклад, з добової або сезонної
ритмікою), спрямованими (накопичення забруднюючої речовини в озері) або
безладними (зв'язаними, наприклад, з пожежами, градом і тайфунами).
в) Поява в деякій частині середовища одних організмів негайно
ж підвищує її різноманітність для інших. Сама присутність організму означає, що
труп його врешті-решт можна буде з'їсти, а якщо він живий, то може збагачувати
навколишнє середовище послідом, сечею або опалим листям. Його власне живе тіло
може служити місцем проживання організмів інших видів або ресурсом для хижаків,
паразитів і хвороботворних організмів, яким до появи його в співтоваристві
нічого було там робити.
Своїм походженням різноманітність, існуюча всередині
природних угрупувань, може бути зобов'язана неоднорідності всіх трьох
перерахованих типів. «Тлумачення» цього розмаїття-вправа в якійсь мірі
тривіальна. Тим, що в рамках одного і того ж угрупування співіснують рослина,
яка використовує сонячне світло, що живе на цій рослині гриб, що поїдають цю
рослину травоїдна тварина і оселився в організмі тварини гельмінт, нікого не
здивуєш. Разом з тим для більшої частини співтовариств характерна наявність
цілого ряду популяцій різних організмів, що володіють вельми схожою будовою і
дуже сильно (принаймні на перший погляд) нагадують один одного способом життя.
Тлумачення й осягнення різноманітності такого роду - вправа вже зовсім не
тривіальна.
Деякі відмінності організмів різних видів,що спільно мешкають
в межах одного і того ж угрупування, демонструють антарктичні тюлені. Вважають,
що нащадкові форми тюленів виникли в Північній півкулі, де їх викопні рештки
відомі з міоцену; одна з груп тюленів, однак, перемістилася на південь, у більш
теплі води, і, мабуть, в кінці міоцену або початку пліоцену (близько 5 млн.
років тому) заселила Антарктику. Коли туди проникли тюлені, Південний океан,
мабуть, ряснів їжею і був вільний від великих хижаків (що, до речі, знову має
місце в наші дні). Судячи з усього, саме в таких умовах група зазнала
дивергентної еволюції як на рисунку 3.1. Тюлень Уедделла, наприклад, годується
головним чином рибою і володіє неспеціалізованою зубною системою;
тюлень-крабоїд споживає майже виключно криль, і його зуби пристосовані до
відцідження криля з морської води; у тюленя Росса маленькі, гострі зуби, а
поїдає він в основному пелагічних кальмарів; у морського ж леопарда зуби
великі, загострені, «хапальні», і харчується він найрізноманітнішою їжею, в
тому числі іншими тюленями, а в певну пору року і пінгвінами.
Насправді, можливість і ступінь співіснування тюленів різних
видів в одних і тих же спільнотах зумовлюється, ймовірно, відмінностями між
ними. Якщо тюлені двох або більше видів споживають приблизно одну і ту ж їжу
приблизно на одній і тій же глибині (як, наприклад, субантарктичний морський
слон і тюлень Росса), то вони ізольовані географічно (їх ареали перекриваються
лише в незначній мірі). Якщо ж існують багатовидові угруповання, то вони
складаються з видів тюленів, що використовують у їжу різні організми (з
тюленя-крильоїда, тюленя Росса і морського леопарда; із морського леопарда і морського
слона; із тюленя Уедделла і морського леопарда - див. рисунок 3.1).
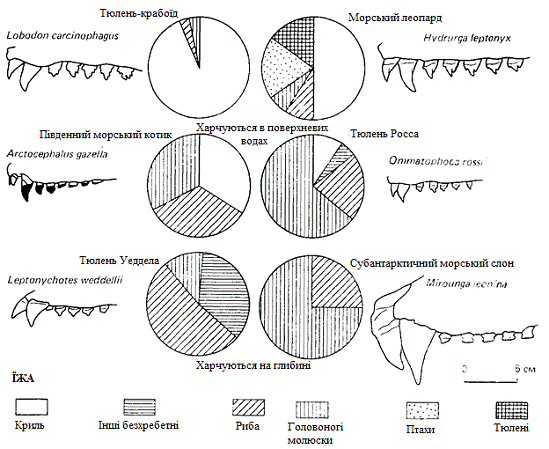
Рисунок 3.1 - Склад їжі деяких представницький групи
антарктичних ластоногих, глибина, на якій вони добувають їжу, і будова їх
зубів.
Можна відзначити ряд істотних обставин.
) Співіснуючі види, як правило, в якихось деталях
відрізняються один від іншого, але організми кожного з цих видів відповідають
середовищу свого проживання і існують тому, що саме середовище неоднорідне.
) Таку диференціацію найлегше виявити при вивченні
близькоспоріднених видів (таких, як антарктичні тюлені), оскільки на тлі
безлічі загальних рис відмінності виступають найбільш виразно. Але, можливо, не
менш важливо зрозуміти природу співіснування членів однієї і тієї ж гільдії, що
зовсім не обов'язково складаються в близькому спорідненні. Наприклад,
антарктичні води рясніють крилем, і на споживанні цього корму спеціалізуються
тюлень-крабоїд і в дещо меншій мірі морський леопард; але криль поїдають ще
кальмари, риби, птиці і вусаті кити, тому в тій запеклій головоломці, яку являє
собою подібне товариство, повний аналіз відповідностей і адаптацій не може
зводитися до вивчення окремих таксономічних груп, а повинен включати розгляд
цілих гільдій.
) Розібраний приклад з антарктичними тюленями ставить
набагато більше питань, ніж дає відповідей. Чи є неминучими відмінності між
співіснуючими видами? Якщо неминучі, то наскільки вони повинні відрізнятися
(інакше кажучи, чи існує яка-небудь межа їх схожості)? Чи взаємодіють між собою
популяції різних видів, подібних антарктичним тюленям, в даний час або
дивергентна еволюція в минулому призвела до відсутності таких взаємодій в
сучасних гільдіях? Ні на одне з цих питань не можна дати простої і однозначної
відповіді, але кожному з них у наступних розділах буде приділено увагу.
.3 Екологічна ніша
Екологічна ніша - фізичний простір з властивими йому
екологічними умовами, що визначають існування будь-якого організму, місце виду
в природі, що включає не лише становище його в просторі, а й функціональну роль
у біоценозі та ставлення до абіотичних факторів середовища існування.
Екологічна ніша характеризує ступінь біологічної спеціалізації даного виду.
Місцезростання, - говорить Ю. Одум, - це адреса мешкання
виду, а екологічна ніша - це система занять в тій системі видів, до якої він
належить. Іншими словами, знання екологічної ніші дає можливість відповісти на
питання, як, де і чим живиться вид, чиєю здобиччю є сам, яким чином і де він
відпочиває і розмножується, які для нього необхідні умови середовища
(температура, вологість і т.д.)
За станом розміщення екологічних ніш види поділяють на
алопатричні і симпатричні. Перший тип об'єднує види, які мають різні ареали і
їх екологічні ніші розміщені порізно або частково заходять одна на одну.
Конкуренція в цьому випадку виключається географічною ізоляцією.
Другий тип розміщення видів відрізняється від першого
сумісним їх проживанням на значній території і частковим або повним включенням
одна в одну.
Виділяють ще й третій тип розміщення екологічних ніш, який
називають суміжною алопатрією: обидва ареали і екологічні ніші стикаються. На
рис. 3.35 зображено декілька різних випадків розташування ніш. Вирізняють
потенційну (фундаментальну) і реальну (реалізовану) екологічні ніші. Потенційна
ніша являє собою всю сукупність необхідних для виду умов місцезростання без
будь-якого тиску з боку другого виду і відповідає максимально можливій
експансії з боку першого. Реальна екологічна ніша - це частина існуючої
потенційної ніші, яка фактично зайнята видом в біотопі. На рис. 3.36 зображене
взаємне розташування фундаментальної (потенційної) і реалізованої (реальної)
ніші.
.4 Спеціалізація всередині видів
До цих пір в якості одиниць, відповідних середовищу, ми
розглядали види (або таксони більш високого рангу), але деякі з випадків
найбільш глибокої, спеціалізованої відповідності умовам існування можна виявити
саме на підвидовому рівні. Обговорюючи особливості тваринного і рослинного
світу островів, ми підкреслювали, що якщо генетичний обмін і рекомбінація
володіють нівелюючою дію, виникнення більшої частини найбільш помітних
міжпопуляційних відмінностей пов'язано швидше за все з географічною ізоляцією
різних частин вихідних популяцій, що тягне за собою припинення схрещування. І
все-таки в тих випадках, коли «місцеві» сили природного відбору дуже великі,
вони можуть подолати нівелюючий вплив статевого розмноження і рекомбінації. У
таких випадках навіть за відсутності повної репродуктивної ізоляції
«сприйтливі» генотипи можуть в місцевих умовах володіти такими перевагами, що
«несприятливі» генетичні комбінації будуть неухильно усуватися (елімінуватися).
Потік генів при цьому не переривається, і окремі популяції залишаються
частинами одного і того ж виду, але всередині нього виникають спеціалізовані
локальні раси.
Швидкість обміну генетичним матеріалом усередині популяції
залежить від рухливості самих організмів або, як це буває в популяціях
прикріплених організмів, від рухливості їх гамет, пилку чи насіння. Якщо
особини популяції вільно переміщаються в пошуках статевих партнерів (або якщо
безперешкодно поширюються їх гамети, спори або насіння), то ймовірність того,
що вплив локального спеціалізованого природного відбору приведе до виникнення
локалізованих, спеціалізованих відповідностей між організмами і середовищем,
невелика.
Утворення спеціалізованих локальних популяцій найбільш
характерно для організмів, що залишаються протягом більшої частини життєвого
циклу прикріпленими. Причина в тому, що рухливі організми значною мірою
контролюють умови свого існування: вони можуть уникнути згубних або
несприятливих місцеперебувань або віддалитися з них і приступити до активного
пошуку нових; нерухомі ж організми - вищі рослини, багато морських водоростей,
корали - подібною свободою не володіють ; по завершенні разселюючої стадії життєвого
циклу їм доводиться або жити в умовах, що існують там, де вони осіли, або
гинути. Як зауважив Бредшоу, «... рослина не в змозі перебігти на нове місце
або сховатися в затишному куточку». Все, що може вища рослина - це,
розростаючись і «переростаючи» з місця на місце, вишукувати ресурси або
вибиратися за межі несприятливого ділянки; видерти себе з корінням і пересадити
себе за власним вибором в інше місце вона ніяк не може. Її потомство (насіння,
пилок або гамети) схильне всім примхам пасивного поширення вітром, водою або
тваринами (на поверхні або всередині тіла). З цих причин популяції нерухомих
організмів піддаються особливо сильній дії природного відбору.
У найбільш різкій формі відмінності між способами «досягнення
відповідності» із середовищем проживання, характерні для рухливих і
прикріплених організмів, можна спостерігати на березі моря, де літораль
поперемінно перетворюється то в наземне, то в водне місце проживання.
Прикріплені водорості, гідроїдні поліпи, губки, моховинки, молюски та вусоногі
раки - всі ці організми стикаються з двома протилежними крайнощами свого
існування і стійко їх переносять. При цьому рухливі члени того ж водного
співтовариства (креветки, краби, риби) переміщуються слідом за своїм водним
місцем життя, а птиці, що годуються біля урізку води, пересуваючись то вперед,
до моря, то назад, до берега, слідують за поперемінними скороченнями і
розширеннями свого наземного місце проживання. Прикріпленим організмам
доводиться виносити повний добовий цикл змін у їхньому навколишньому середовищі,
а рухливим організмам така витривалість ні до чого: вони переміщаються слідом
за приливами і відливами. Відповідність таких рухливих організмів середовищу
свого існування дозволяє їм уникати впливу багатьох чинників природного
відбору. Рухливість наділяє організм здатністю підбирати місце проживання за
власною міркою. Нерухомий організм повинен бути скроєний за міркою свого
місцеперебування.
.4.1 Екотипи
Термін «екотипи» був вперше використаний стосовно до деяких
рослин - для опису внутрішньовидових генетично зумовлених локальних
відповідностей між організмами і середовищем. Вирощуючи рослини з різноманітних
природних місцеперебувань в одному загальному протягом одного або більше
вегетаційних сезонів, дослідники виявили значні внутрішньовидові відмінності
між цими рослинами. При постановці подібних досліджень важливо, щоб рослини, що
походять з різних місць, вирощувалися і порівнювалися в одних і тих же умовах,
тому що деякі з відмінностей між природними популяціями можуть виявитися
фенотиповими реакціями на умови існування, не пов'язаними з генотиповими
відмінностями. До прикладу, на інтенсивно використовуваному пасовищі повзучі
столони конюшини розгалужені дуже сильно, а на сінокісних лугах - набагато
менше, але відмінності ці можуть виникнути між частинами однієї і тієї ж
рослини - варто тільки помістити їх у різні умови,як наприклад на рисунку 3.2.
Коли рослина конюшини Trifolium repens, розростаючись, проростає з ділянки, де
злаків немає, на ділянку, де домінує злак Lolium регеппе, характер росту
конюшини змінюється. Наведений на малюнку план показує, як розташовані на
поверхні ґрунту столони повзучої конюшини після проростання його в злакову
дерновину. Зверніть увагу, що, опинившись у дерновину, мережа столонів конюшини
порідшала, міжвузля їх подовжилися, а самі столони стали набагато рідше
гілкуватися.
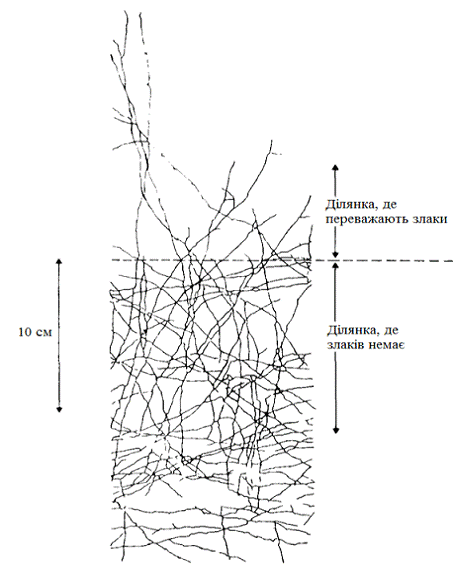
Рисунок 3.2 - Зміна характеру росту конюшини
У роботах по вивченню стійкості до забруднення токсичними
важкими металами (свинцем, цинком, міддю і т. д.) з граничною ясністю показано,
що в рослин просторовий масштаб локальної спеціалізації може бути надзвичайно
дрібним. На краях ділянок, які опинилися в результаті розробки рудних родовищ
забрудненими, інтенсивність відбору проти «сприйнятливих» генотипів різко
змінюється, і популяції, що займають забруднені ділянки, можуть дуже сильно
відрізнятися за ступенем стійкості до важких металів, будучи при цьому
розділеними відстанню менш ніж в 100 м (у запашного колоска - менш ніж 1,5 м).
І все це - незважаючи на те, що вітер і комахи безперешкодно переносять пилок
через ці кордони.
В окремих випадках вдалося встановити час, з якого
розпочалося вплив відбору на стійкість до підвищених концентрацій важких
металів. У Суонсі (Південний Уельс) така стійкість, очевидно, складалася з тих
самих пір, як 300 років тому почався видобуток руд; відомий випадок, коли
рослини, що росли під огорожею з оцинкованого заліза, придбали стійкість до
цинку не більш ніж за 25 років; створюючи дуже густий експериментальний засів
сильно забрудненої важкими металами ділянки ґрунту, вдавалося отримати стійку
популяцію рослин в результаті відбору, що діяв протягом одного покоління! У
порівнянні з найпотужнішим тиском відбору потік генів з його нівелюючим впливом
досить слабкий; на користь цього аргументу свідчить як швидкість відбору, так і
чіткі просторові межі між екотипами.
.4.2 Генетичний поліморфізм
Переходячи від екотипів до ще більш детального аналізу
популяційних структур, біологи набувають все більших навиків виявлення
пов'язаних з відбором мінливості усередині невеликих локальних популяцій. Така
мінливість відома як поліморфізм. Точніше кажучи, генетичний поліморфізм - це
«співіснування в межах одного і того ж місцеперебування двох або більше чітко
помітних внутрішньовидових форм, причому в таких співвідношеннях, що постійна
присутність рідкісної з цих форм не може бути віднесена тільки на рахунок
безперервного мутагенезу та імміграції». Далеко не всі прояви такого роду
мінливості відображають відповідності між організмами і середовищем; навпаки,
буває так, що за деякими з них стоять явні невідповідності. Невідповідності ці
можуть виникати через те, що розселяючи стадії однієї спеціалізованої форми
проникають в місце проживання іншої. Вони можуть виникати і тоді, коли при
зміні умов одна форма витісняється іншою, краще пристосованою до умов, що
змінилися. Такий поліморфізм називається минущим. Спільноти завжди змінюються,
дуже багато спостережуваних в природі явищ поліморфізму, мабуть, носять саме
такий тимчасовий характер -у тій мірі, в якій ні одна популяція ніколи не
встигне за змінами умов існування і ніколи не буде в змозі ці зміни
передбачити.
І все ж у багатьох випадках внутрішньопопуляційний
поліморфізм активно підтримується природним відбором, і при цьому
найрізноманітнішими способами.
а) У деяких випадках гетерозиготи мають підвищену
пристосованість, але через менделевське розщеплення вони постійно поповнюють
популяцію породжуваними ними менш життєздатними гомозиготами. Приклад такого
«гетерозису» - існування серповидної клітинної анемії в популяціях людей, що
знаходяться в осередках малярії. Індивідууми, гетерозиготні за відповідним
локусу, страждають легким недокрів'ям, але рідко хворіють малярією; при цьому
вони безперервно породжують гомозиготних індивідуумів - або уражених важким недокрів'ям,
або сприйнятливих до малярії.
б) Інтенсивність відбору може змінюватися в межах деякого
діапазону, причому на одній з його меж відбір може сприяти одній формі (морфе),
а на іншій границі - іншій. При проміжній інтенсивності відбору можуть виникати
поліморфні популяції.
в) Іноді відбір буває частотно-залежним: будь-яка з
внутрішньовидових форм найбільш життєздатна тоді, коли вона зустрічається рідше
всіх інших. Вважають, що саме так йде справа з незвично пофарбованими формами
жертв: вони життєздатні тому, що хижаки їх не впізнають і не чіпають.
г) дрібномасштабна просторова структура популяції та її
місцеперебування буває дуже складним, і в різних частинах цієї «клаптикової
ковдри» відбір може діяти в різних напрямках. Підтримка відповідності між
організмами та середовищем в такій ситуації неминуче залежить від розсіювання
численних розселяючих стадій: якщо чисельність їх досить висока, то досить
висока і вірогідність того, що частина їх вкорениться саме в тому «клапті», де
відповідна форма найбільш життєздатна. Можливий і інший варіант: організм може
бути довговічний і здатний переміщуватися, використовуючи при цьому умови та
ресурси найбільш підходящих «клаптів».
Вражаючий приклад поліморфізму природної популяції виявлено в
результаті низки робіт, присвячених вивченню конюшини повзучої, зростаючої на
довголітньому (не молодше 80 років) пасовищі в Північному Уельсі. В ході одного
з цих досліджень з ґрунту були викопані 50 рослин конюшини. Рослини ці потім
містилися в теплиці, і кожну з них було піддано генетичному аналізу з цілого
ряду локусів, пов'язаних з ознаками, майже всі з яких свідомо володіли
селективною значимістю. За комбінацією властивостей, які, мабуть, впливають на
пристосованість рослин в природних умовах, кожен з 50 клонів відрізнявся від
усіх інших.
Ясно, що мінливість конюшини на цьому маленькому полі хоча б
частково обумовлена генетично і відображає відповідності між організмами і
середовищем.
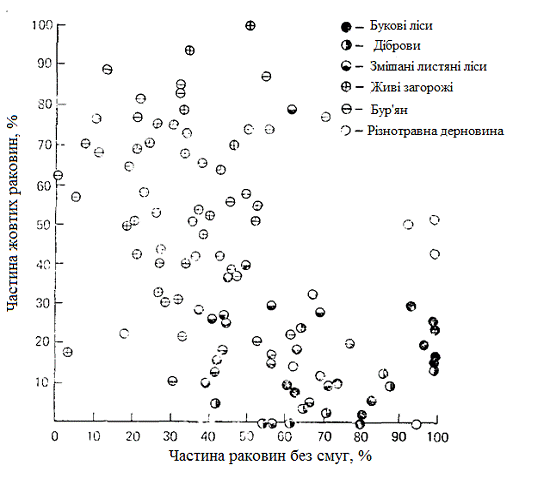
Рисунок 3.3 - Частота зустрічання раковин
Прикладом поліморфізму у тварин може служити поліморфізм в
популяціях равликів Сераеа nemoralis. Равлики малорухливі, а популяції їх
займають найрізноманітніші середовища існування, що різко розрізняються по
фізичних умовах і ступенях тиску хижаків. Ця обставина привела до локальної
внутрішньопопуляційної диференціації за зовнішнім виглядом раковини. На рис.
3.3 зображена частота зустрічальності равликів Cepaea nemoralis з жовтими
раковинами, позбавленими смуг, в різних середовищах існування. (Жовті раковини
протиставляються буро-рожевим, а раковини без смуг - смугастим.) У наявності
непогана відповідність між найбільш поширеним забарвленням і колірним фоном:
наприклад, в букових лісах найчастіше зустрічаються равлики з буро-рожевими
раковинами, а в живих огорожах - зі смугастими жовтими раковинами, що
сприймаються як зелені.
Будь-яка з популяцій равликів поліморфна хоча б по одній з
двох ознак: кольору раковини і наявності на ній малюнка у вигляді смуг.
Чисельні співвідношення різних форм в різних місцях зосередження равликів
неоднакові, але в кожному місці існування для найбільш звичайної форми
характерна візуальна відповідність своєму оточенню. Очевидно, на тлі
відповідної рослинності та відповідного ґрунтового покриву равлики цих форм не
помітні для дроздів, що харчуються ними.
.5 Відповідність між організмами й середовищем, що змінюється
популяція середовище розселення організм
З плином часу будь-які умови існування змінюються, але в
одних випадках вони схильні до більш сильних змін, а в інших - до менш сильних.
Ні будова організму, ні його поведінка не можуть відповідати мінливим умовам,
не змінюючись адекватним чином. Можна виділити три основні типи змін середовища
проживання.
а) Циклічні зміни, тобто періодично повторювані, як при зміні
пір року, при припливах і відливах і при почерговому настанні світлого і
темного часу доби.
б) Направлені зміни, при яких напрям зміни залишається
стабільним протягом періоду, тривалість якого може бути дуже високою, у
порівнянні з тривалістю життєвого циклу організмів,що переживають цю зміну.
Прикладами спрямованих змін можуть служити прогресуюча ерозія берегів,
накопичення донних відкладів в естуаріях рік і зміни, що відбуваються на одній
з фаз циклу обледеніння.
в) Хаотичні зміни; для всіх змін цього типу характерна
аритмія і відсутність певного напряму. Приклади хаотичних змін - коливання
термінів настання мусонних дощів, непередбачувані зміни часу виникнення і
траєкторій циклонів і ураганів, шквали, викликані ударом блискавки, пожежі.
Найсприятливіша відповідність між організмами та умовами, що
змінюються неминуче передбачає певний компроміс між пристосуванням до змін і
здатністю до їх переживання. В умовах багаторазового впливу циклічних змін на
послідовні покоління організмів природний відбір привів до виникнення ряду особливостей
способу життя, які і самі по собі є циклічними. До числа таких особливостей
відносяться діапауза комах, щорічне скидання листя листопадними деревами,
добові рухи листя, приливно-відливних ритм переміщень у літоральних крабів,
річний цикл функціонування репродуктивних систем і сезонний цикл зміни густоти
хутра у ссавців.
Існують два основних способи, якими організми пристосовують
свої реакції до змін у навколишньому середовищі: а) зміна у відповідь на зміну
зовнішніх умов і б) реагування на сигнальний фактор, що передбачає зміну
зовнішніх умов. У тому випадку, якщо циклічні зміни умов середовища виражені
слабо і до того ж вельми варіабельні, найкращим способом адаптації організмів
до таких умов може бути пряме реагування на них. Такого роду непередбачуваність
характерна для випадання опадів; із рисунку 3.4 видно, що вона властива навіть
тим місцевостям, де відзначається чітко виражений сезонний ритм. Тут показано
діаграми, що відповідають трьом різним місцевостям. Кожна з діаграм відображає
багаторічну динаміку кількості опадів, що випадають протягом того місяця, який
у відповідній місцевості є самим дощовим (в середньому).
У найбільш різкій формі вона простежується на нижній
діаграмі, що відбиває багаторічні зміни кількості опадів, що випадали в Каїрі протягом
січня. У таких умовах будь-якій рослині, запрограмованій на проростання у
відповідь на дію якогось фактора, що свідчить про настання сезону дощів, навряд
чи судилося б вижити. Дійсно, проростання пустельних рослин найчастіше
викликається не якоюсь прикметою про наближення дощів, а дощем як таким.
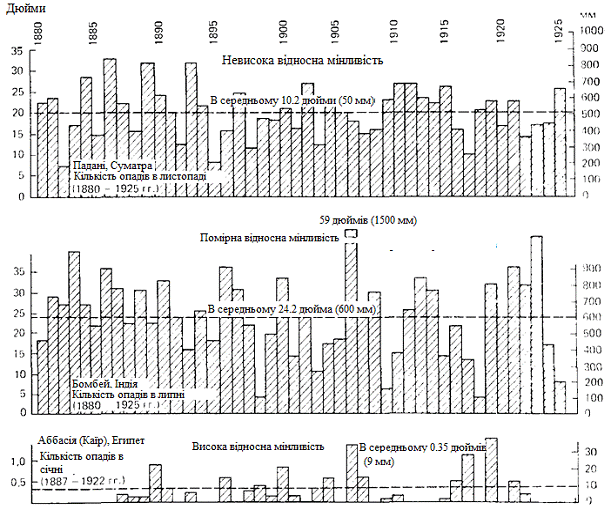
Рисунок 3.4 - Мінливість кількості опадів і
непередбачуваність їх випадання.
Втім, організмам, що реагують на зміну умов безпосередньо,
дечим доводиться розплачуватися: деяких переваг вони позбавлені. Ссавцю, у
якого густота хутра змінюється у відповідь на похолодання, доведеться тремтіти
від холоду до тих пір, поки зміна волосяного покриву не завершиться; але якщо
ссавець реагує не на саме похолодання, а на пов'язаний з ним сигнал, що
свідчить про нього (такий, наприклад, як зменшення довготи дня), то збільшення
густоти волосяного покриву може початися завчасно. Реагування на подібні
сигнальні стимули вельми характерне для тих тварин і рослин, які існують в
умовах, що змінюються з явною і відносно стабільною циклічністю.
Існує спокуса писати про живих істот так, ніби вони здатні до
передбачення (або про умови середовища так, ніби вони передбачувані); хочеться
написати і про ознаку, що дозволяє організмам будувати плани на майбутнє. На
перший погляд циклічні особливості способу життя як нібито узгоджені з якимись
прогнозами; насправді ж це зовсім не так. Циклічні особливості являють собою
ознаки, що виникли в результаті природного відбору в умовах багаторазово
пережитих в минулому циклічних змін середовища проживання. Будова та спосіб
життя живої істоти зумовлюються не його власною здатністю до передбачення
майбутнього, а життям і смертю попередніх поколінь.
Явища, подібні нічному мороку і зимовим холодам, пов'язані з
плином часу, і тривалість їх можна виміряти, а наступ - вгадати, використовуючи
в якості годин Сонце (висота Сонця над горизонтом може служити мірою часу доби,
а тривалість світлового дня - показником пори року). До цих зовнішніх годин
деякі організми додали свій власний внутрішній фізіологічний годинник
(ендогенний ритм). Такий годинник потрібно час від часу звіряти з сонячним і
встановлювати по ньому, але і без звірки фізіологічний годинник може
функціонувати досить довго.
Життєві цикли більшості птахів і ссавців довше календарного
року, і багато дрібних ссавці, в умовах клімату з чітко вираженою сезонністю,
зимують, впадаючи в сплячку, тобто знижуючи інтенсивність обміну і ховаючись в
затишних місцях. Багато інших ссавці чинять інакше: на зміну умов існування
вони відповідають зміною ступеня теплоізоляції тіла. У найбільш різкій формі
сезонні зміни густоти хутра відзначаються у великих ссавців. Теплоізолююча
здатність зимового хутра чорного ведмедя на 92% вище, ніж річного. Подібні
циклічні зміни товщини, густоти і фарбування зовнішніх покривів характерні для
сезонних реакцій тих тварин, які не можуть уникнути несприятливої пори року;
але з усіх сезонних ритмів поведінки та способу життя рухомих тварин
найчудовіші - ті, що припускають переміщення. Це можуть бути переміщення в
притулку, а можуть бути і масові переселення в інші кліматичні області (на
зразок щорічних міграцій північних оленів, бізонів та багатьох птахів).
Вкоріненим вищим рослинам і прикріпленим тваринам цього, звичайно, не дано,
зате саме у цих організмів можна спостерігати приклади найбільш яскраво
виражених сезонних ритмів, що зачіпають особливості їх будови.
Для багатьох водних рослин річний цикл зміни умов зовнішнього
середовища постає передусім у вигляді коливань рівня води; на ці сезонні ритми
можуть накладатися хаотичні зміни, викликані сильними засухами або раптовими
повенями. У прибережній частині водойми виростають рослини найрізноманітніших
форм. Рослини, що ростуть біля урізку води, зазвичай забезпечені повітряним
листям, яке при зануренні у воду різко знижує фотосинтетичну активність і
навіть пошкоджується. Листя рослин, що ростуть на більш глибоких місцях,
зазвичай плавають на поверхні води. На ще більш глибоководних ділянках у рослин
повністю занурене, довге, гнучке стрічкоподібне листя, що не ушкоджується
швидкою течією. А в найбільш глибоководній зоні ростуть рослини із зануреними і
тонко розсіченим листям. Для багатьох рослин характерне листя лише одного з
перерахованих спеціалізованих типів. Крім того, у багатьох водних рослин одна і
та ж особина може нести на собі листя більш ніж одного типу. Такий фенотиповий
поліморфізм розширює межі коливань рівня води, при якому можлива ефективна
життєдіяльність цих рослин.
Для організму, який не може уникнути несприятливих умов,
найбільш ефективним вирішенням проблеми виживання в змінному середовищі можуть
виявитися сезонні зміни особливостей будови. У посушливих місце проживання
фенотиповий поліморфізм буває ще більш різким, ніж у водних рослин. Деякі
рослини протягом одного року утворюють три покоління листя, причому листя
кожного з поколінь володіють своєю характерною будовою. У Teucrium pollium під
час вологого сезону утворюються порівняно велике листя. Під час більш
посушливого сезону вони опадають, а на зміну їм приходять дрібні листочки, або
лусочки. Іноді зникає і це «друге» листя, і рослина проводить найбільш
посушливий період лише з зеленими стеблами та колючками.
.6 Пари видів
Серед найбільш яскравих прикладів відповідності між
організмами і середовищем ми знаходимо такі, у яких очевидне виникнення
залежності організмів одного виду від організмів іншого виду. Саме так йде
справа в багатьох випадках взаємин між споживачами та їх їжею, наприклад, у
випадку залежності коали від листя евкаліпта (Eucalyptus) або гігантської панди
від паростків бамбука. Цілий комплекс особливостей будови, обміну речовин і
поведінки утримує тварину в рамках його вузької харчової ніші і не дозволяє
йому скористатися тим, що в інших обставинах могло б здатися цілком придатним
додатковим джерелом їжі. Такі ж жорсткі відповідності характерні для взаємин
між деякими паразитами і їх господарями. Наприклад, видоспецифічність іржавого
гриба, пристосованого до існування в абсолютно певному і чітко обмеженому
середовищі, а саме в організмі господарів суворо визначених видів.
Якщо в ході еволюції між двома різними видами виникла взаємна
залежність, то відповідність може бути ще жорсткішою. Два переконливих приклади
- мутуалістичний зв'язок між азотфіксуючими бактеріями і корінням бобових
рослин і часто дуже тонка взаємна відповідність між комахами-запилювачами і
квітками запилюваних ними рослин. Найбільш тісні відповідності між організмами
і середовищем виникли в тих випадках, коли вирішальним фактором життєдіяльності
організмів одного виду є присутність організмів іншого виду: в таких випадках
все середовище проживання одного організму може вичерпуватися іншим організмом.
Якщо популяція піддається впливу змінних чинників фізичного
середовища (якщо, наприклад, вона існує в умовах нетривалого вегетаційного
періоду, підвищеного ризику заморозків або засухи, багаторазового застосування
гербіцидів), то організми можуть врешті-решт придбати постійну різнобічну
стійкість. Самі фізичні фактори в результаті еволюції організмів ні
перетворюватися, ні еволюціонувати не можуть. При взаємодії ж організмів різних
видів зміни одних організмів приводять до змін у житті інших, і будь-який з
взаємодіючих видів може створювати умови відбору, направляючі еволюцію іншого
виду. В ході такого процесу коеволюції міжвидова взаємодія може постійно посилюватися
і поглиблюватися, а результат цього процесу ми можемо спостерігати в природі як
пару видів загнали один одного у колію все більш і більш поглибленої
спеціалізації.
Поглядаючи на різноманіття живої природи, важко встояти перед
почуттям подиву, захоплення і зачарування тим, що так легко прийняти за
досконалість. Не треба забувати, що здатність дивуватися і захоплюватися - це
специфічна риса біології нашого власного виду! Будучи натуралістом, еколог
докопується до причин і наслідків; йому не годиться задовольнятися
«поясненнями», призначеними лише для того, щоб показати, яким чином в цю ж саму
мить все «досягло досконалості в цьому найкращому зі світів».
4.
РОЗРАХУНКОВА ЧАСТИНА. Кількісна оцінка
видової структури популяцій
4.1 Завдання
1) Розрахувати кількість взаємодій між видами у
біоценозі.
2) Розрахувати індекс
домінування.
4) Визначити три показники
видової різноманітності популяцій.
5) Розрахувати показник загальної різноманітності
організмів - індекс Шеннона.
6) Оцінити показник
вирівнюванності Піелу.
7) Визначити індекс видового
багатства Маргалефа.
4.2
Розрахунки
1) Кількість взаємодій між видами знаходимо за формулою:
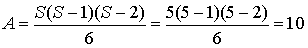 (4.1)
(4.1)
де S - загальна кількість видів.
2) Індекс домінування:
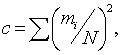
деmi - кількість особин і-го виду;
N -
загальна кількість живих організмів.
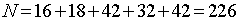
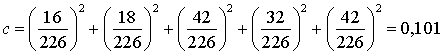 (4.2)
(4.2)
3) Розраховуємо індекс різноманітності Сімпсона:
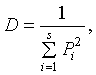
де Pi - частка і-го виду 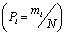 .
.
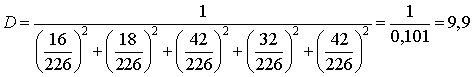 (4.3)
(4.3)
4) Визначаємо три показники
видової різноманітності популяцій:
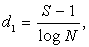
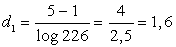 (4.4)
(4.4)
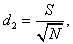
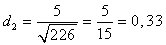 (4.5)
(4.5)
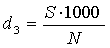
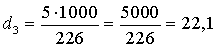 (4.6)
(4.6)
5) Обчислюємо показник загальної різноманітності
організмів - індекс Шеннона:
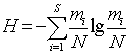
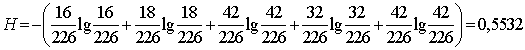 (4.7)
(4.7)
6) Оцінюємо індекс Піелу:
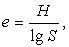
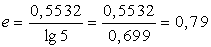 (4.8)
(4.8)
7) Визначаємо індекс видового
багатства Маргалефа:
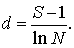
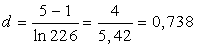 (4.9)
(4.9)
ВИСНОВКИ
В курсовій роботі виконано наступне:
) Розглянуто відповідність між організмами і
середовищем існування, через природний добір та пристосування
) Встановлено деякі історичні взаємозв’язки між
організмами та середовищем існування на прикладі: змін внаслідок переміщень
масивів суші, змін клімату, острівного відчуженого життя, а також явищ
конвергенції та паралелізму
) Показано подібність між угрупуваннями та
взаємозв’язки в них самих за рахунок таких явищ як подібність між
угрупуваннями, розмаїття в них самих, спеціалізація всередині видів, пари видів
і поняття екологічна ніша.
) В розрахунковій роботі дано кількісну оцінку видовій
структурі популяцій і отримано такі результати:
¾ кількість взаємодій між видами А =
10,
¾ індекс домінування с = 0,101;
¾ індекс різноманітності Сімпсона D =
9,9;
¾ показники видової різноманітності популяції1
= 1,62 = 0,333 = 22,1;
¾ індекс Шеннона H = 0,5532,;
¾ індекс Піелу е = 0,79;
¾ індекс видового багатства Маргалефа d = 0,738.
ПЕРЕЛІК
ПОСИЛАНЬ
1. Бігон
М., Харпер Дж., Таудсен К. Екологія, особи, популяції та угрупування. Т - 1.
1989. - 667 с.
2. Кучерявий
В. П. Екологія. 2001 - 500 с.
. Воронков
Н. А. Екологія загальна, соціальна, прикладна. 1999. - 346 с.
. Бігон
М., Харпер Дж., Таудсен К. Екологія, особи, популяції та угрупування. Т - 2.
1989. - 477 с.
5. Казначеев
В. П. Учение В. И. Вернадского о биосфере и ноосфере. 1989. - 248 с.
6. Одум
Ю. Екологія. Т - 1. 1986. - 328 с.
. Білявський
Г.О. Основи екології: теорія і практикум / Г. О.Білявський, Л. І. Бутченко, В.
М. Навроцький - К.: Лібра, 2002. - 352 с.
. Уіттекер
Р. Угрупування та екосистеми. 1989. - 487 с.
. Вернадський
В. І. Вибрані праці. 1969. - 567 с.
10. Реймерс
Н. Ф. Екологія (теорія, закони, правила, принципи та гіпотези). 1994. - 534 с.
11. Федоров
В. Д., Гільманов Т. Г. Екологія. 1980. - 347 с.