Мозговое кровообращение
Реферат
Мозговое
кровообращение
Содержание
Введение
.
Морфологические особенности мозгового кровообращения
.
Функциональные особенности мозгового кровообращения
.1
Особенности системы суммарного мозгового кровотока
.2
Особенности системы локального мозгового кровотока
Литература
мозговой кровообращение нервный
ткань
Введение
Нормальная деятельность головного мозга
наитеснейшим образом связана с его постоянно адекватным и хорошо регулируемым
кровоснабжением. Высокая чувствительность нервной ткани к изменениям рО2, рСО2
и содержания глюкозы, с одной стороны, и сравнительно легкая поражаемость ее
сосудистой системы, с другой, позволяют объяснить, почему расстройства функции
головного мозга чаще всего определяются циркуляторными нарушениями.
В 70-80-е годы XX столетия благодаря прежде
всего классическим исследованиям Б.Н. Клосовского, Ю.Е. Москаленко, Г.И.
Мчедлишвили в области физиологии мозгового кровообращения сформировались
основополагающие представления о функциональной организации кровоснабжения
головного мозга. Многочисленные аспекты проблемы мозгового кровообращения были
сведены в две группы. К первой из них были отнесены принципы организации
внутричерепной гемодинамики и суммарного кровоснабжения мозга , тогда как
вторую группу составили сосудистые механизмы, обеспечивающие функцию локального
кровоснабжения нервной ткани.
В настоящее время не вызывает сомнений тот факт,
что процессы суммарного и локального кровоснабжения головного мозга
взаимосвязаны и служат единой цели - адекватному циркуляторно-метаболическому
обеспечению нервной ткани и поддержанию ее гомеостаза.
1. Морфологические особенности мозгового
кровообращения
Поступление артериальной крови в головной мозг
осуществляется двумя парами близко расположенных к выходу из сердца
магистральных сосудов головы: внутренними сонными и позвоночными (сливающимися
в базиллярную) артериями, которые у основания мозга объединяются в Виллизиев
круг.
От Виллизиева круга в каждом из полушарий
отходят по 3 крупные артерии: передняя и средняя, образующие каротидную систему
кровоснабжения мозга, и задняя мозговая артерия, формирующая
вертебробазиллярную систему снабжения кровью затылочных долей, мозжечка,
варолиева моста и продолговатого мозга. Эти крупные мозговые артерии, широко
анастомозируя, направляются кверху и, разветвляясь, переходят в систему
пиальных артерий, расположенных на поверхности мозга. Пиальные сосуды,
соединяясь друг с другом, образуют анастомозы не только в системе одной из
крупных мозговых артерий, но и между ответвлениями разных артерий, что
способствует созданию коллатерального кровотока и надежности кровоснабжения
мозга. От пиальных артерий ответвляются радиальные артерии, которые почти под
прямым углом пронизывают вещество мозга и формируют короткие и длинные
отростки. Первые из них дают кровоснабжение серого вещества, тогда как длинные
отростки обеспечивают кровоснабжение белого вещества мозга. Здесь артериолы,
отходящие от радиальных артерий, образуют капиллярные сети, кровь от которых
поступает в венулы, а те, сливаясь, формируют радиальные вены, которые в
восходящем направлении пронизывают вещество мозга и выходят на его поверхность,
образуя систему широко анастомозирующих пиальных вен. Пиальные вены сливаются в
крупные венозные синусы, снабженные достаточно жесткими стенками. От синусов
венозная кровь по двум основным коллекторам (югулярным венам) направляется к
сердцу. Отток крови от мозга также осуществляется по каналам, впадающим в
позвоночное сплетение, и по анастомозам с орбитальным и крыловидным
сплетениями.
В сосудистой системе головного мозга можно
выделить две взаимосвязанные подсистемы:
а) макроциркуляции, расположенной практически на
поверхности мозга и образующей русло для общего суммарного кровотока;
б) микроциркуляции, обеспечивающей
кровоснабжение вещества мозга и формирующей сосудистое ложе для локального
мозгового кровотока.
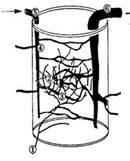
Рис. 1. Схема организации сосудистого модуля
соматосенсорной области коры полушарий мозга (по Woolseyetal., 1996).
- таламо-кортикальные афференты; 2 - дендриты и
тела нейронов IV слоя коры; 3 - капилляры; 4 - радиальные артерии с
ответвляющимися от них артериолами; 5 - пиальная артерия; 6 - пиальная вена.
Макроциркуляция характеризуется многочисленными
анастомозами, что широко используется в нейрохирургической практике путем
наложения сосудистых протезов в условиях тромбоза крупных мозговых сосудов. В
отличие от этого, в микроциркуляторном русле отмечается практически полное
отсутствие анастомозов, поэтому тромбоз или стойкий спазм микрососудов, как
правило, полностью не компенсируется и сопровождается нарушением тех функций
организма, которые регулировались поврежденным нервным центром.
Гистологические особенности мозговых сосудов
связаны, прежде всего, с особенностью строения стенки капилляров, состоящей в
наслоении эндотелиальных клеток друг на друга, что делает места их соединений
практически непроницаемыми (Бредбери, 1983). Пространство между капиллярами и
нейронами заполнено глиальными клетками-астроцитами. Такое пространственное
соотношение между капиллярами, нейронами и астроглией формирует
гематоэнцефалический барьер (см. разд. 3), функцией которого является
избирательная проницаемость из крови в мозговую ткань только необходимых для
мозговой деятельности компонентов. В силу функционирования ГЭБ мозг практически
обладает "своей", отличающейся по химическому составу, внутренней
средой. При такой конструкции через стенку капилляров свободно проникают вода и
растворенные в липидах О2 и СО2, тогда как кислоты - нет. И лишь в некоторых
участках мозга проницаемость капилляров выше: в области гипоталамуса, синего
пятна и ядер шва, т.е. там, где сконцентрированы полимодальные хеморецепторы.
Строение стенки мозговых артериол
характеризуется содержанием в интиме сосудов одного слоя плотно примыкающих и
связанных друг с другом многообразными контактами эндотелиоцитов.
Подэндотелиальная зона наряду с эндотелиоцитами содержит в своем составе
миоэндотелиальные соединения. Учитывая, что эндотелиоциты способны вырабатывать
химические вещества - факторы расслабления и сокращения гладкомышечных клеток
(ГМК), их можно рассматривать как источник местных вазоактивных веществ,
воздействующих на сократительную активность ГМК изнутри сосуда и провоцирующих
сокращение либо расслабление сосудистых стенок. Мышечный слой содержит ГМК,
соединенные друг с другом многочисленными контактами (плазматическими выростами
и нексусами), создавая условия для электротонического проведения возбуждения
между ГМК, что способствует быстрому охвату возбуждением мышечного слоя
сосудов.
Наружная адвентициальная оболочка артериол
содержит в своем составе соединительнотканные элементы со встроенными в них
гранулосодержащими клетками и нервными волокнами, что свидетельствует о наличии
субстрата для местных вазоактивных влияний, провоцирующих сокращение и
расслабление сосудистых стенок. Среди гранулосодержащих клеток, встречающихся в
артериях головного мозга и представляющих местные эндокринные элементы,
идентифицированы хромаффиноциты, тканевые базофилы и меланоциты. Установлено,
что наряду с известными биологически активными веществами - адреналином,
норадреналином, серотонином, гистамином гранулосодержащие клетки могут
синтезировать и депонировать олигопептиды, вызывающие сосудистые эффекты.
Гистохимическими методами установлена природа
нервных проводников адвентициального слоя мозговых артерий и артериол,
отражающая источник их иннервации. Она представлена в основном симпатическими
адренергическими волокнами, берущими начало из верхнего шейного узла для
иннервации сосудов, относящихся к системе внутренних сонных артерий, и из
звездчатого ганглия - для артерий и артериол в системе позвоночных сосудов.
Наибольшая плотность иннервации мозговых сосудов присутствует в крупных
артериях мозга, особенно во внутренних сонных. Здесь представлены как
многочисленные адренергические, так и холинергические нервные окончания,
которые располагаются у наружной границы медиального слоя и адвентиции сосуда
на близком (до 26 нм) расстоянии друг от друга, что создает условия для их
функционального взаимодействия. В зонах смежного кровоснабжения главных артерий
мозга холинергические и адренергические нервные волокна образуют сплетения, и
выраженность иннервации сосудов в этих областях высокая. Иннервационный аппарат
пиальных и внутримозговых артерий представлен менее выражено. Эти группы
артерий снабжены адренергическими нервными волокнами. Если эфферентная
холинергическая иннервация присутствует в крупных мозговых артериях, то
адренергические волокна распространяются вплоть до мельчайших артериол. При
этом показано, что по мере уменьшения диаметра мозговых сосудов в процессе их
ветвления интенсивность адренергической иннервации их стенок уменьшается
Известны данные о серотонинергической (исходящей из ядер шва, голубого пятна) и
пептидергической (волокна внутримозговых пептидергических нейронов) иннервации
мозговых сосудов, присущей преимущественно внутримозговым артериям и
артериолам.
С помощью химико-фармакологических приемов
исследования установлено, что нервные влияния на стенку мозговых сосудов
опосредуются через альфа- и бета-адреноредепторы, М-холинорецепторы,
Д-серотониновые рецепторы. При этом стимуляция альфа-адренорецепторов приводит
к сужению артерий мозга, а бета-адренорецепторов и М-холинорецепторов - к их
расширению. Кроме того, через альфа-адренорецепторы реализуется влияние
катехоламинов на углеводный обмен в мозгу.
Благодаря электронно-гистохимическим методам
исследования установлены некоторые особенности иннервации сосудов мозга. Так,
терминальные отделы аксонов, иннервирующих сосудистую стенку артерий мозга,
редко содержат одну терминаль, чаще всего, это комплекс разветвлений с разной
химической специфичностью. При этом взаимодействие терминалей аксонов с ГМК
мозговых артерий может быть двух типов: а) непосредственная взаимосвязь
нервного окончания с ГМК; б) нервное окончание отстоит от миоцита на
расстояние, равное 3-4 диаметрам синаптического пузырька. Если в первом случае
терминали аксонов непосредственно модулируют сократительную активность ГМК
сосуда, то во втором случае объектом для их влияния служат гранулосодержащие
клетки адвентиция. Прямой контакт нервных терминалей с моноаминоцитами
адвентициальной оболочки мозговых сосудов обусловливает их реакцию на нервное
возбуждение, что показано при электрической стимуляции волокон блуждающего
нерва и местном применении ацетилхолина. Наличие в терминалях нервных
проводников, контактирующих с гранулосодержащими клетками адвентиция сосудов
мозга, светлых пузырьков, ферментов холинацетилтрансферазы и
ацетилхолинэстеразы, позволяет их классифицировать как холинергические.
Следовательно, помимо возможности прямого
действия нервных волокон на ГМК мозговых сосудов, имеется основа для
опосредованного их влияния через гранулосодержащие клетки, выделяющие
олигопептиды и биогенные моноамины с разнонаправленным (констрикторным и
дилататорным) действием на мозговые сосуды.
2. Функциональные особенности мозгового
кровообращения
Головной мозг человека даже в условиях
функционального покоя характеризуется непрерывно протекающими высокоемкими
энергетическими процессами аэробного характера, требующими высокого потребления
мозговой тканью кислорода (3-4 мл/100г/мин) и глюкозы (5 мг/100г/мин). В
нервной клетке при рО2 20 мм рт.ст. в каждом мкм3 содержится 2,0*104 молекул
кислорода, а потребляется тем же объемом примерно 1,6*105 молекул в секунду.
Причем, нервная ткань практически не обладает ни субстратом для анаэробных
окислительных процессов, ни запасами кислорода, а, значит, для нормального функционирования
мозга необходимым является высокая интенсивность его кровоснабжения. В связи с
этим, головной мозг, весящий в среднем 1400-1500 г (примерно 2% массы тела), в
состоянии функционального покоя получает около 750 мл/мин крови, что составляет
примерно 15% общего сердечного выброса. Объемная скорость кровотока при этих
условиях соответствует 50-60 мл/100г/мин, однако следует отметить, что серое
вещество обеспечивается кровью интенсивнее, чем белое вещество, что связано с
его более высокой клеточной активностью. У детей 1 года жизни величина общего
мозгового кровотока на 50-55% больше, а в старческом возрасте примерно на 20%
меньше, чем у человека в зрелом возрасте.
Функциональное назначение системы мозгового
кровообращения, как и в любом другом регионе, направлено на то, чтобы
своевременно и в адекватных количествах доставить кислород и питательные
вещества к нервным клеткам, удалить продукты клеточного метаболизма и тем самым
обеспечить гомеостаз нервной ткани. Между тем хорошо известно, что не все функциональные
единицы мозга, даже в условиях физиологического покоя, работают одинаково
активно в одно и то же время, а значит имеют разные метаболические потребности.
Последнее обстоятельство позволяет предположить, что специфичное для мозга
мозаичное распределение активности нейронов требует в свою очередь и
избирательной доставки энергетического материала нервным клеткам, которая может
быть обеспечена только за счет гетерогенного распределения кровотока в мозгу
между относительно покоящимися и более активными участками нервной ткани.
Экспериментальным подтверждением вышеизложенному
служат результаты исследования распределения кровотока по коре полушарий мозга
у человека. Так, у пациентов в состоянии лежа на спине с закрытыми глазами, при
полном расслаблении скелетных мышц интенсивность кровотока в лобных долях была
выше на 20-30%, а в затылочной и височной долях ниже на 20-30% по сравнению с
другими отделами коры головного мозга. Во время функциональной нагрузки, когда
пациенты производили ритмическое сжатие кисти руки, в контралатеральном
полушарии интенсивность кровотока в средней части центральной борозды
увеличивалась на 50%, а в лобной и премоторных областях коры головного мозга
снижалась на 30% по сравнению со средним общеполушарным уровнем кровотока.
Следовательно, в головном мозге наряду с
сосудистыми механизмами, обеспечивающими необходимый уровень суммарного
мозгового кровотока, должны функционировать механизмы, обеспечивающие локальное
кровоснабжение отдельных участков мозга при изменении их функциональной
активности. Каждая в отдельности организация системы суммарного и локального
кровоснабжения в головном мозге имеет свои отличительные черты.
.1 Особенности системы суммарного мозгового
кровотока
Суммарное кровенаполнение мозга осуществляется в
сложных биофизических условиях. Мозг расположен в ригидном костном образовании
черепе (исключение составляют дети грудного возраста, имеющие роднички,
придающие некоторую подвижность черепной коробке), где помимо вещества мозга
содержатся кровь и цереброспинальная жидкость, являющиеся практически
несжимающимися жидкостями. Поэтому здесь имеет место прямой гидравлический
контакт несжимающихся жидкостей в черепной коробке. Для сохранения неизменным
объема внутричерепного содержимого возможен отток некоторого количества
цереброспинальной жидкости через ограниченное в размерах отверстие в более
растяжимый спинномозговой канал. Такие сложные биофизические условия для
суммарного мозгового кровотока требуют наличия надежных механизмов
предохранения вещества мозга от избыточности кровенаполнения.
При избытке кровоснабжения мозга в нервной ткани
может произойти излишняя гидратация, а отсюда высокая вероятность развития
отека мозга с последующими (несовместимыми с жизнью) повреждениями жизненно
важных центров.
Ведущей причиной избыточности кровоснабжения
головного мозга может служить увеличение системного артериального давления.
Однако в норме при участии ауторегуляторных сосудистых реакций мозг предохранен
от избыточности кровенаполнения при повышении давления вплоть до 160-170 мм
рт.ст. Помимо феномена ауторегуляции кровотока, предохранение головного мозга
(близко расположенного к сердцу органа) от высокого кровяного давления и
избыточности пульсации происходит и за счет особенностей конструкции сосудистой
системы мозга. В частности, эту функцию достаточно эффективно выполняют
многочисленные изгибы (сифоны) по ходу сосудистого русла, которые способствуют
значительному перепаду давления и сглаживанию пульсации кровотока.
В активно работающем мозге формируется
потребность в увеличении интенсивности кровоснабжения. Благодаря феномену
функциональной гиперемии, такая возросшая потребность полностью
удовлетворяется, никак не вступая в противоречие с необходимостью
предотвращения головного мозга от избыточности кровенаполнения. Объясняется это
специфическими особенностями мозгового кровообращения.
При повышенной активности всего организма
(усиленная физическая работа, эмоциональное возбуждение и т.п.) суммарный
мозговой кровоток в мозге может увеличиваться примерно на 20-25%, что не
оказывает повреждающего действия, поскольку вещество головного мозга снабжается
кровью сосудами, находящимися на его поверхности. Поверхностно расположенные
пиальные сосуды проходят в каналах мягкой мозговой оболочки, за счет расстояния
до твердой мозговой оболочки располагают резервом для некоторого
кровенаполнения. Многочисленные анастомозы в системе пиальных сосудов
обеспечивают высокую надежность в равномерном поступлении крови к различным
отделам головного мозга.
На уровне сосудов, образующих систему для общего
мозгового кровотока, нет возможности перераспределять кровь между различными
областями мозга, но имеет место преимущественное снабжение кровью коры мозга
через сонные артерии, продолговатого мозга - через позвоночные артерии, а
структуры промежуточного и среднего мозга - через те и другие.
2.2 Особенности системы локального мозгового
кровотока
Текущие физиологически активные состояния
человека (включая умственную, интеллектуальную деятельность) характеризуются
развитием процесса активации в строго соответствующих нервных центрах, где и
формируются доминантные очаги. В этом случае нет необходимости в увеличении
суммарного мозгового кровотока, зато есть необходимость внутримозгового
перераспределения крови в пользу активно работающих зон (областей, участков)
мозга. Эта функциональная потребность реализуется путем активных сосудистых
реакций, развивающихся в пределах соответствующих областей мозга.
Особенностями локального кровоснабжения в
головном мозге являются высокая гетерогенность и изменчивость распределения
локального кровотока в микроучастках нервной ткани. Наиболее высокий уровень
кровотока отмечен в корковых структурах и в ядрах гипоталамуса. При
одновременном измерении кровотока в соседних участках одной и той же структуры
мозга обнаруживается существенное различие его абсолютных величин. Так, если в
двух точках коры мозга, отстоящих друг от друга на расстоянии до 200-250 мкм,
значения кровотока близки по величине, то на больших расстояниях разница между
ними достигает 1,5-2-кратной величины. Размеры зон, в которых наблюдается
синхронность спонтанных колебаний кровотока, составляют в среднем 250-300 мкм.
Существенные колебания кровотока наблюдаются и
по слоям коры мозга. Наибольшая его величина отмечается в IV-V слоях коры, а
наименьшая - на границе с белым веществом. Такая закономерность в распределении
кровотока в коре головного мозга объясняется неравномерностью
цитоархитектоники, плотности васкуляризации мозга, гетерогенностью активации
нейронных популяций.
Так, в исследованиях, проведенных на крысах,
показано, что нейроанатомическая структура коры головного мозга состоит из
плотно упакованных кольцеобразных комплексов нервных клеток, так называемых
"бочонков" или "колонок" (Сухов, 1992). Диаметр таких
бочонков составляет 250-300 мкм. Плотность капилляров внутри бочонков нервных
клеток максимальна. Таким образом, гетерогенность кровотока в коре мозга может
быть связана с разной плотностью васкуляризации нервной ткани, которая в свою
очередь зависит от неравномерности распределения активности нервных клеток.
Измерение локального мозгового кровотока у
испытуемых в 254 точках по всей латеральной поверхности полушария с помощью
детекторов клиренса радиоактивного ксенона позволило обнаружить очаги
функциональной гиперемии в коре, которые перемещались по полушарию в
зависимости от вида функциональной деятельности мозга или модальности
сенсорного стимула. Такие очаговые изменения кровотока и его перераспределение
по полушарию были тесно связаны с уровнем активации соответствующих корковых
областей.
Исследование локального кровоснабжения в
сенсомоторной зоне коры головного мозга у человека с помощью позитронной
эмиссионной томографии в покое и при движении пальцев выявило при выполнении
функциональной нагрузки увеличение кровотока в соответствующих двигательных
зонах коры. Таким образом, яркой отличительной чертой локального кровоснабжения
головного мозга является гетерогенность распределения мозгового кровотока.
Проведение сравнительно-физиологического
исследования гетерогенности распределения кровотока в ряду позвоночных от
амфибий до высших млекопитающих показало, что с развитием мозга мозаичность
распределения кровотока увеличивается. Это дало основание предположить, что в
процессе эволюции происходило совершенствование системы оперативного и надежного
обеспечения адекватного кровоснабжения мозга при изменении функциональной
активности нейронов от отдельных нейронных ансамблей до его обширных зон.
Интенсивность кровоснабжения локальных участков
мозга зависит от следующих параметров:
а) плотности функционирующих капилляров;
б) анатомической и функциональной геометрии
микроциркуляторного русла;
в) характера кровотока.
Согласно большому количеству морфологических
данных стенка мозговых капилляров не имеет специализированных сократительных
элементов и лишена вазомоторной иннервации. В связи с этим, изменение числа
функционирующих капилляров не может считаться активной реакцией со стороны
самих капилляров. К пассивному "расправлению" нефункционирующих
капилляров могут привести уменьшение тонуса прекапиллярных сфинктеров,
расширение артериол в комбинации с сужением венул или без участия такого
сужения.
Со времени открытия Крогом (1919) такой
закономерности ее значение традиционно преувеличивается. Исследования
последнего времени показали, что в мозге такой реакции не наблюдается:
капилляры мозга в отличие от других органов постоянно открыты, чему
способствует постоянство оттока по периваскулярным пространствам
интерстициальной жидкости, окружающей микрососуды мозга. Изменение числа
открытых капилляров может регулироваться лишь благодаря сокращению
эндотелиальных клеток. Однако сократительная активность эндотелиальных клеток
не является их специализированной функцией и направлена не столько на изменение
числа открытых капилляров, сколько на изменение их проницаемости.
Микрососудистые сети мозга обладают
пластичностью, под которой условились понимать способность или свойство
микрососудистых сетей изменять свою конфигурацию при изменении параметров
гемодинамики. При этом сосуд обладает двумя видами подвижности:
) может изменять величину диаметра своего
просвета;
) может изменять свою длину.
Среди основных факторов, которые способствуют
изменению архитектоники микроциркуляторного русла мозга, выделяют:
внутрисосудистое давление, напряжение сдвига на сосудистой стенке, потоковые
характеристики крови, вязкость крови и гематокрит, растяжение сосудов внешними
силами и др.
В микрососудах мозга линейная скорость кровотока
является крайне вариабельной величиной. Так, в одном и том же капилляре мозга
линейная скорость кровотока изменяется в пределах от 0,05 до 2,5 мм/с в течение
всего нескольких секунд. Обращает на себя внимание факт наличия в мозге высокой
изменчивости скорости движения эритроцитов в венулах. Эта закономерность была
выявлена для венул коры головного мозга диаметром 8-12 мкм. Вариабельность
линейной скорости кровотока в микрососудах мозга тесно связана с локальными
особенностями структуры кровотока. Для микрогемодинамики в головном мозге
характерными чертами являются: попеременное запустевание микрососудов,
изменения потока в противоположных направлениях (колеблющийся поток),
переменное движение эритроцитов и пристеночного слоя плазмы, высокий уровень
псевдотурбулизации потока крови. В капиллярах коры головного мозга, ввиду их
узкого просвета, эритроциты при движении сильно деформируются. При этом
эритроциты значительно вытягиваются в длину, и соотношение их длины к
поперечнику составляет 2:1 или даже 2,5:1. На такую деформацию мембран
эритроцитов затрачивается значительная энергия. Целью такой деформации
эритроцитов, по-видимому, является сохранение пристеночного слоя плазмы в
капиллярах. Даже в узких капиллярах коры мозга этот пристеночный слой плазмы
имеет место, и его толщина составляет около 0,5 мкм.
При уменьшении объемной скорости кровотока в
локусе нервной ткани местный гематокрит по сравнению с другими органами резко
уменьшается. Положительная роль такой перестройки структуры кровотока
заключается в увеличении текучести крови, а ее отрицательное значение
определяется уменьшением количества эритроцитов в потоке.
Изменчивость скорости кровотока в микрососудах
мозга определяется также особенностями движения лейкоцитов. При переходе
лейкоцита из узкого капилляра мозга в более широкую венулу скорость движения
эритроцитов на короткое время резко возрастает. При ишемии мозга ввиду резкого
угнетения выделения NO из эндотелиоцитов мозговых сосудов увеличивается адгезия
лейкоцитов к стенкам капилляров, что закупоривает их просвет и становится
причиной невозможности восстановления кровотока в микрососудах после ишемии.
Для локального кровоснабжения мозга характерными
являются медленные колебания кровотока с частотой 6-7 циклов в минуту и
амплитудой до 10% от исходного уровня. Возможными причинами флуктуации
локального мозгового кровотока считаются перистальтическая ритмическая
активность ГМК артериол и ритмические колебания тонуса прекапиллярных
сфинктеров.
Изменения мозгового кровообращения, связанные с
характером нейродинамических процессов в головном мозге, сводятся к увеличению
интенсивности кровоснабжения функционально активных участков мозга с
одновременным уменьшением кровотока в других зонах, активность которых в данный
момент снижена. При этом местные сосудистые реакции характеризуются коротким
латентным периодом - в пределах 1-2 с. Так, применение радиоизотопных методов
исследования мозгового кровотока позволило выявить, что у человека при
произвольных движениях кисти руки интенсивность кровотока в контралатеральном полушарии
увеличивается в прецентральной и постцентральной извилинах соответственно на 22
и 21,8%, а в затылочной и височной областях снижается. При реализации
зрительной функции также характерно специфическое распределение кровотока в
коре полушарий. Когда пациенты открывают глаза и рассматривают предметы, то
повышение кровотока в среднем на 20% имеет место в зрительной коре, включая ее
ассоциативные зоны, а также локально во фронтальной области, ответственной за
движения глазных яблок, и добавочной моторной коре. Местные изменения мозгового
кровотока при повышении биоэлектрической активности различных областей мозга
вследствие выполнения человеком соответствующих функциональных проб в среднем
составляют ±20-30%, а в отдельных случаях могут достигать ±60%.
Одним из принципиально важных вопросов проблемы
регуляции регионарного кровообращения остается исследование физиологических
закономерностей перераспределения локального кровотока между микроучастками
коры головного мозга, имеющими разный уровень функциональной активности.
Наиболее удачным объектом для разработки этого вопроса представляется
соматосенсорная область коры головного мозга, где эволюционно сформировались и
по сравнению с другими органами получили наивысшее развитие по автономности
своего функционирования гетерогенные клеточные популяции. Очевидно, популяции
нервных клеток, отличающиеся друг от друга активностью функционирования,
нуждаются в адекватном гемоциркуляторном обеспечении и, следовательно, наличии
оперативных механизмов местной регуляции кровотока. В связи с этим для
выявления закономерностей функциональной организации кровоснабжения
микроучастков коры головного мозга представляется необходимым комплексное
исследование характера сосудистых реакций, нейронной активности и кислородного
обеспечения в пределах отдельных функционально специализированных популяций
нейронов.
Литература
1.
Физиология человека: В 3-х томах. Т. 1. Пер. с англ. / Под ред. Р. Шмидта и Г.
Тевса. - 3-е изд. - М.: Мир, 2005. - 323с.
.
Николс Джон, Мартин Роберт, Валлас Брюс, Фукс Пол От нейрона к мозгу / Пер. с
англ. П.М. Балабана и др. - М.: Эдиториал УРСС, 2003, - 672 с.
.
Основы нейрофармакологии: Учеб. пособие для студентов вузов / Е.И. Белова. -
М.: Аспект Пресс, 2006. - 176 с.
.
Шульговский В.В. Физиология центральной нервной системы. Учебник - М.: МГУ,
1997, - 397 с.
5.
Физиология центральной нервной системы: учеб. пособие / Т.В. Алейникова, В.Н.
Думбай, Г.А. Кураев, Г.Л. Фельдман. Ростов н/Д: Феникс, 2006. - 376 с.
6.
Общая физиология нервной системы. В серии: Руководство по физиологии. Л.,
"Наука", 1979. 717 с.
.
Фундаментальная и клиническая физиология: Учебник для студ. высш. учебн.
заведений / Под ред. А.Г. Камкина и А.А. Каменского. - М.: Издательский центр
"Академия", 2004. - 1072 с.
8.
Алейникова Т.В. Принципы переработки информации в зрительной системе лягушки. -
Ростов-на-Дону: "ИРУ", 1985. - 128 с.
.
Линдсей П., Норман Д. Переработка информации у человека. - М.: "Мир",
1974. - 452 с.
.
Прибрам К. Языки мозга. - М.: "Прогресс", 1975. - 464 с.
.
Сомьен Дж. Кодирование сенсорной информации. - М.: "Мир", 1975. - 416
с.
.
Чораян О.Г. Информационные процессы в нервной системе. - Ростов -на-Дону:
"ИРУ",1976. - 163 с.
.
Ещенко Н.Д. Энергетический обмен в головном мозге // Биохимия мозга / Под.ред.
И.П. Ашмарина и др. - СПб. -1999. - С. 124-168.
15.
Неговский В.А. Патофизиология и терапия агонии и клинической смерти. - М.:
Медгиз, 1954.
.
Николаев А.Я. Биологическая химия. - М.: Медицинкое информационное агенство,
2004.
.
Фердинанд Хухо. Нейрохимия. Основы и принципы. - М.: Мир, 1990.
.
Эккерт Р., Рэнделл Д., Огастин Дж. Физиология животных: механизмы и адаптация.
Том 1. Пер. с англ. - М.: Мир, 1991.
.
Хананашвили Я.А. Основы организации кровоснабжения органов. - Ростов-на-Дону,
2001., 160 с.
.
Хананашвили Я.А. Лекции по физиологии регионарного кровообращения. -
Ростов-на-Дону, 2009, 86 с.