Влияние степени окисления элементов на их биологическую роль
Белорусский
государственный университет
Химический
факультет
Кафедра
неорганической химии
Курсовая
работа
Влияние
степени окисления элементов на их биологическую роль
Студентки 1 курса
группы
химического факультета БГУ
Новожиловой Марии Юрьевны
Руководитель:
Доцент кафедры неорганической химии
Лесникович Юлия Анатольевна
Минск,
2015
Введение
Основное место в живых организмах занимают сложные
органические молекулы, однако их предназначение не может быть реализовано без
определенного содействия ряда низкомолекулярных веществ. Это так называемые
металлы жизни: Na, K, Ca, Mg, Zn, Fe, Mn, Cu, Co, Mo. Видно, что
основная масса биологически активных металлов относится к переходным элементам.
В данной работе будет рассмотрено биологическое
действие пяти металлов жизни: железа, меди, марганца, кобальта и молибдена.
Целью данной работы является изучение биологической роли каждого из
вышеперечисленных металлов, а также выявление общих закономерностей для всех d-металлов, имеющих переменную
валентность.
Для достижения цели работы были поставлены следующие
задачи:
. Провести обзор научной литературы, статей и
рефератов, соответствующих теме курсовой работы.
. Выделить наиболее значимую информацию.
. На основе исследования выявить характерные
особенности биологической роли d-металлов
с переменной валентностью.
. Обобщить собранный материал по данной теме.
Переходные металлы участвуют в жизненно важных
процессах организма, поэтому изучение их свойств и биологической роли является
важной задачей для современной биохимии, фармакологии и медицины.
Общие сведения о свойствах d-элементов. Степени окисления
окисление биологический железо медь
Важным свойством d-элементов является переменная валентность и, как следствие,
разнообразие степеней окисления. Эта связано с незавершенностью предвнешнего
электронного слоя (кроме элементов IB и IIB групп). Возможность существования d-металлов в разных степенях окисления
определяет их широкий диапазон окислительно-восстановительных свойств.
Скорее всего в ходе биологической эволюции отбирались
соединения элементов в промежуточных степенях окисления с мягкими
окислительно-восстановительными свойствами, так как они способствуют плавному
протеканию биохимических реакций. Известно, что ионы Мn2+, Fе2+,
Со2+, Cu2+ при физиологических условиях
практически не проявляют в организме восстановительных свойств, а Mo5+ и Mo6+ проявляют слабые окислительные свойства. Тонкая регулировка
биологических процессов обеспечивает выигрыш по энергии. Из-за этого
функционирование организма становится менее энергоемким и уменьшается
потребление пищевых продуктов [5].
Дополнительное снижение реакционной способности
происходит при взаимодействии этих ионов с биоорганическими лигандами.
Комплексообразование
Комплексообразующая способность у d-металлов достаточно высокая, обычно
значительно выше, чем у s- и p-элементов. Это прежде всего
объясняется возможностями d-элементов
быть как донорами, так и акцепторами электронной пары при образовании
комплексных соединений.
В биохимических реакциях d-элементы часто проявляют себя как
металлы-комплексообразователи. Лигандами при этом выступают биологические
молекулы или анионы неорганических кислот. Ион металла взаимодействует с
электроотрицательными атомами, входящими в различные функциональные группы:
гидроксильные, сульфгидрильные, карбоксильные и аминогруппы.
Важно отметить, что различные окислительные состояния d-металлов стабилизируются
соответствующими лигандами.
Металлопорфирины
Для живых организмов очень важны комплексные
соединения металлов, в которых четыре координационных места занимает частица,
называемая порфином. Она содержит четыре пирролоподобных цикла, соединенных
=CH-группами:
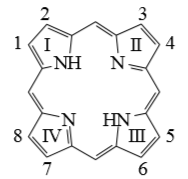
Рисунок
1. Порфин
Ион металла замещает атомы водорода двух пиррольных
колец и одновременно связывается координационными связями с третичными атомами
азота. Благодаря эффекту резонанса связи металла с четырьмя атомами азота
пиррольных колец рассматриваются как одинаковые.
Производными порфина являются порфирины. К ним в
настоящее время относятся представители многочисленного класса циклических
ароматических соединений. Основной каркас молекулы сохраняется во многих
важнейших веществах: гемоглобине, цитохромах, витамине В12.
Важнейшим свойством порфиринов является наличие в
молекуле координационной полости, ограниченной атомами азота, N4,
имеющей радиус около 2 Å и способной координировать ионы металлов М2+,
М3+, М4+. В результате комплексообразования образуются
так называемые металлопорфирины, обладающие многообразными структурными и химическими
особенностями, высокой биологической и каталитической активностью. При этом
металл либо занимает центр полости N4, образуя плоский
координационный узел из атомов MN4, либо оказывается приподнятым над
плоскостью, в которой лежат атомы азота, и образует координационные узлы
различной геометрической структуры - от тетрагональной пирамиды (L)MN4
и октаэдра (L1)(L2)MN4 до более сложных
геометрических фигур [9].
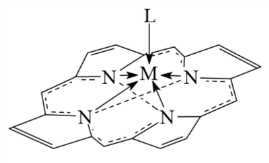
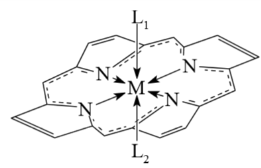
Рисунок
2. (L)MN4 Рисунок 3. (L1)(L2)MN4
Возможности молекул металлопорфиринов выступать в
биологических процессах в качестве ферментов значительно расширяются в связи с
необычайным строением порфиринов и их комплексов.
Металлопорфирины являются макроциклическими комплексами,
и это накладывает отпечаток на их строение и свойства. Однако они отличаются от
бесчисленного множества других групп макроциклических комплексов тем, что
являются ароматическими макроциклами с уникальной сопряженной π-системой. Ароматичность порфиринов
определяет их электронодонорные свойства, то есть способность к снижению
локальных положительных и отрицательных зарядов путем их распределения по
ароматическим орбиталям. Вследствие этого стабилизируются катион- и
анионрадикальные формы, а также различные степени окисления металлов,
возникающие в процессе функционирования биологически активных соединений на
основе металлопорфиринов.
Общие сведения о биологической роли d-элементов
Большинство d-металлов служит в качестве простетических групп ферментов. Металлоферменты
d-элементов - жизненно необходимы. Их
биологическая роль зависит от прочности образуемого комплекса. Причем с
повышением устойчивости металлофермента, повышается специфичность его действия.
К биологическим соединениям с невысокими значениями
констант устойчивости можно отнести соединения, которые стабилизируют сложные
структуры. Например, образование комплексов стабилизирует двойную спираль ДНК.
Комплексы с ДНК образуют ионы Mn2+, Co2+, Fe2+ и они взаимозаменяемы.
В организме присутствуют и менее прочные комплексы,
которые образуются только для выполнения определенных функций, после чего
распадаются: например, образование между ионом металла и ферментом комплексного
соединения в период осуществления катализа. Активаторами часто выступают ионы металла,
в частности Mn2+. Большинство таких ферментов
обладают каталитической активностью, однако без иона металла она ниже.
Специфичность металлов в этих комплексах не выражена. Он может быть заменен на
другой ион металла без потери физиологической активности.
Некоторые соединения настолько прочные, что постоянно
находятся в организме и выполняют определенную функцию. Роль металла в таких
комплексах высокоспецифична: замена его даже на близкий по свойствам элемент
приводит к значительной или полной утрате физиологической активности. Например,
гемоглобин и витамин В12.
Промежуточное положение между этими группами
биокомплексов занимают диссоциирующие металлоферменты. Ионы металлов в этих
комплексах выполняют функции кофактора [5].
Биологическая роль железа
Железо - важнейший биометалл. Среди биоэлементов оно
играет доминирующую роль, так как является активатором многих каталитических
процессов в организме, участвует в транспортировке газов кровью и работе
электрон-транспортных цепей. Степень окисления железа зависит от выполняемой
роли.
Основой миоглобина и гемоглобина, определяющей их
биологические роли и характерный красно-коричневый цвет, служит гем. Он
представляет собой протопорфирин, связанный с атом двухвалентного железа. Атом
железа имеет шесть координационных связей, четыре из которых участвуют в
образовании комплекса железа с плоской молекулой порфирина, а две другие
направлены перпендикулярно кольцу. В миоглобине и гемоглобине одна из этих
связей занята атомом азота, принадлежащим остатку гистидина. Другая связь
свободна и служит для связывания молекулы кислорода.
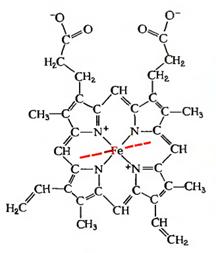
Рисунок 4. Структура гема
В миоглобине и гемоглобине за эту свободную связь
помимо молекулы O2 может конкурировать молекула СО,
которая образует с атомом железа в 200 раз более прочную связь, чем кислород.
При отравлении угарным газом значительная часть гемоглобина переходит в форму
карбоксигемоглобина, препятствуя переносу кислорода из легких в ткани [4].
Кислород транспортируется гемоглобином эритроцитов от
легких к мышцам и удерживается в мышцах миоглобином. Основная масса железа в
организме сконцентрирована в гемоглобине. Одна его молекула состоит из четырех
глобул и содержит четыре гема и, следовательно, четыре атома железа. Так как
кислород фиксируется непосредственно железом, то гемоглобин может присоединить
четыре молекулы O2.
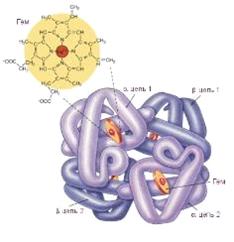
Рисунок 5. Молекула гемоглобина
Молекула О2 связывается с Fe2+
за счет дативной π-связи. Железо поставляет пару электронов, находящуюся
на его d-орбитали, на вакантную p-орбиталь кислорода. Образованию связи
благоприятствует высокая электронодонорная способность порфириновой π-системы и проксимального имидазола.
Атом железа после присоединения кислорода входит в координационную полость
порфирина и располагается центросимметрично [9].
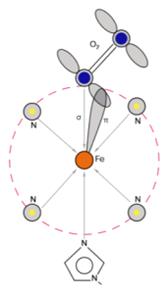
Рисунок 6. Схема координации в оксигенированном
гемоглобине
Кроме переноса кислорода от легких к тканям,
гемоглобин осуществляет перенос конечных продуктов клеточного дыхания, Н+
и СО2, которые потом доставляются из тканей к легким и почкам -
органам, обеспечивавшим выделение этих продуктов.
В клетках тканей питательные вещества окисляются в
митохондриях с использованием кислорода, доставляемого гемоглобином из легких,
при этом в качестве продуктов образуются в основном углекислый газ и вода.
Образование СО2 приводит к повышению в тканях концентрации ионов Н+,
т. е. к понижению рН из-за диссоциации угольной кислоты. На связывание
кислорода гемоглобином очень сильное влияние оказывает pH и концентрация углекислого газа: связывание СO2 и ионов Н+ снижает способность гемоглобина
связывать O2. В тканях с относительно низким значением рН и
высокой концентрацией СО2 сродство гемоглобина к кислороду. И
наоборот, в легочных капиллярах выделение СО2 и сопутствующее ему
повышение рН крови приводит к увеличению сродства гемоглобина к кислороду. Этот
механизм лежит в основе внешнего дыхания [4]. Важным производным гемоглобина
является метгемоглобин, в молекуле которого атом железа находится в степени
окисления 3+. Метгемоглобин не связывает молекулярный кислород. Он образуется
при воздействии на гемоглобин окислителей. Образование метгемоглобина в крови
уменьшает в ней количество функционально важного оксигемоглобина и нарушает доставку
кислорода к тканям.
Миоглобин - кислородрод-связывающий белок,
присутствующий в мышечных клетках. Его функция состоит в том, что он запасает
связанный кислород и способствует его переносу в митохондрии, которые
потребляют кислород в процессе окисления питательных веществ. В молекуле
миоглобина полипептидная цепь координирована железом так же как в гемоглобине.
Но в отличие от молекулы гемоглобина, миоглобин состоит из одной глобулы и
содержит одну гемовую единицу.
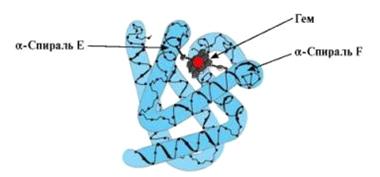
Рисунок 6. Молекула миоглобина
Комплексы железа с порфиринами участвуют не только в
транспорте кислорода, но и выполняют множество других функций. Среди них
процесс переноса электронов.
Важным этапом обмена вещества является отщепление от
питательных веществ водорода. Атомы водорода при этом переходят в ионное
состояние, а освободившиеся электроны поступают в дыхательную цепь. В этой
цепи, переходя от одного соединения к другому, они отдают свою энергию на
образование АТФ, а сами присоединяются к молекуле кислорода. Получившийся ион
кислорода O2- образует с ионами водорода молекулы воды. Перенос
электрона осуществляют железопорфириновые комплексы. Перенос электрона
осуществляется за счет изменения степени окисления железа. Переходы
3+ + e- → Fe2+
Fe2+ − e- → Fe3+
создают возможность передавать электроны от одного
цитохрома к другому.
Цитохромы - железосодержащие белки. Они принадлежат к
классу гемопротеинов, молекулы которых содержат железо, входящее в состав
железопорфириновой группы. Цитохромы обычно делят на три класса: a, b и c.
Действуя в определенной последовательности, они переносят электроны от
убихинона на молекулярный кислород. Цитохромы дыхательной цепи расположены в
последовательности: b - c1 - c - aa3.
Лучше всех изучен цитохром с. Это небольшой белок с
железопорфириновой группой, присоединенный ковалентной связью к единственной
полипептидной цепи. В этом соединении порфириновое кольцо, содержащее железо
(II) в центре, связано с белком за счет ковалентных связей атомов кольца с
остатком цистеина в молекуле белка. С кислородом цитохром c не реагирует, так
как у него шестое координационное место занято аминокислотным остатком
метионина.
Цитохром аа3 отличается от других
цитохромов. В его состав входят две молекулы гема А, отличающегoся от протогема
гемоглобина наличием у его порфиринового кольца длинной углеводородной боковой
цепи. Кроме того, в нем имеются также два атома меди.
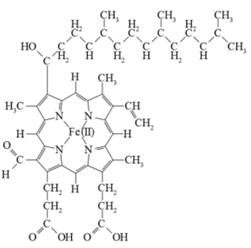
Рисунок 7. Структура гема А
Присоединив электроны, поступившие от цитохрома с, и
перейдя таким образом в восстановленную Fe2+ форму, компонент а цитохрома аа3 передает затем
эти электроны цитохрому а3. Восстановленный цитохром а3 в
свою очередь передает электроны на молекулярный кислород. В этом процессе
вместе с двумя железопорфириновыми группами участвуют два атома меди, что
сопровождается обратимым изменением их валентности [Cu(I) - Сu(II)]. Этот процесс является важным,
поскольку на этом этапе четыре электрона должны быть переданы почти
одновременно на O2 для того, чтобы образовались две
молекулы Н2O
(четыре Н+-иона, которые тоже для этого необходимы, поступают из
водной среды). Из всех переносчиков цепи только цитохром аа3
способен вступать непосредственно в реакцию с кислородом [9].
В общем виде дыхательную цепь можно представить в виде
следующей схемы:
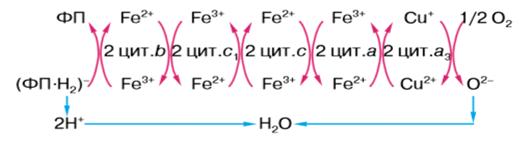
Рисунок 8. Схема процесса клеточного окисления
Для клетки очень важно, чтобы молекула кислорода,
присоединив четыре электрона, полностью восстановилась до двух молекул Н2O. При неполном восстановлении
кислорода образуется Н2O2. Перекись
водорода крайне токсична для клеток. При ее взаимодействии с биоорганическими
соединениями клеток образуются радикалы, инициирующие пероксидное окисление.
Под действием радикалов повреждают клеточные мембраны и ДНК. Аэробные клетки
защищают себя от этого воздействия перекиси с помощью негемового
железосодержащего фермента каталазы, прекращающей перекись водорода в воду и
молекулярный кислород.
Действие каталазы (СаtFе2+) может быть представлено в виде
каталитического цикла из двух последовательных реакций:
СаtFе2+
+ Н2О2 → СаtFе2+•Н2О2
СаtFе2+•Н2О2
+ Н2О2 → СаtFе2+ + 2Н2О + О2
В результате разрушаются две молекулы перекиси
водорода, а молекула каталазы освобождается и может вступать в следующий
каталитический цикл [3].
Биологическая роль меди
Медь - биогенный элемент. В организме она обнаружена в
двух степенях окисления - Cu+ и Cu2+. Ионы меди по сравнению с ионами других металлов активнее
реагируют с аминокислотами и белками, потому что они образуют устойчивые
комплексы, так называемые клешневидные или хелатные.
В ходе эволюции, когда природа создавала систему
переноса кислорода, у нее был выбор между железом и медью. По-видимому,
первоначально у большинства животных пигментом крови служил медьсодержащий
белок гемоцианин, но позднее преимущество получил гемоглобин. Очевидно, что
высшие животные с их возросшей потребностью в кислороде должны были
переключиться на гемоглобин, тогда как моллюски и членистоногие сохранили
гемоцианин, вполне отвечающий их потребностям. У хордовых гемоцианины
обнаруживаются только в плазме крови.
Главная функция меди у высших организмов -
каталитическая. В настоящее время известен целый ряд медьсодержащих ферментов:
церулоплазмин, тирозиназа, супероксиддисмутаза и цитохромоксидаза,
биологическая роль которой была рассмотрена в предыдущей главе. Так же медь
активирует синтез гемоглобина, белков, участвует в образовании костной ткани и
пигмента кожных покровов.
Супероксиддисмутаза - медьсодержащий фермент, предотвращающий
действие пероксид аниона, который образуется в дыхательной цепи при неполном
восстановлении кислорода:
О2 + е- → О2-
Взаимодействие этого аниона с органическими
соединениями клетки приводит к образованию радикалов и нарушению нормального развития
клетки. Повреждающее действие предотвращается супероксиддисмутазой. Этот
фермент катализирует реакцию:
О2- + О2-
+ 2Н+ → Н2О2 + О2
Образующийся при этом пероксид водорода разлагается
каталазой.
Токсическое действие соединений меди обусловлено тем,
что ионы меди взаимодействуют с тиольными и аминогруппами белков. При этом
образуются биокластеры хелатного типа. Вследствие таких взаимодействий белки
становятся нерастворимыми, теряют ферментативную активность. В результате
нарушается нормальная жизнедеятельность.
Биологическая роль кобальта
Кобальт также является одним из важнейших биогенных
элементов. Ион Со3+ входит в состав витамина В12.
Важнейшей функцией катиона Co2+ является активация ферментных
систем, таких как альдолаза, карбоангидраза. Установлена возможность обратимого
транспорта кислорода с участием ионов двухвалентного кобальта, а также их
участие в протеолизе.
Карбоангидраза - фермент, катализирующий обратимую
реакцию гидратации диоксида углерода:
CO2 + H2O ↔
H2CO3 ↔ H+ + HCO3-.
Фермент обнаружен в эритроцитах, в поджелудочной и
слюнной железе, в слизистой желудка, в почках, в ткани центральной нервной
системы, в сетчатке глаза и т. д. Имеются сведения об участии карбоангидразы в
управлении переносом дыхательных газов, в регуляции pH, в управлении реакциями
биосинтеза с участием бикарбоната (глюконеогенез, синтез аминокислот,
липогенез), в регуляции обновления костной ткани, в регуляции образования мочи,
в реакциях, обеспечивающих образование соляной кислоты в железах желудка,
бикарбонатов в соке поджелудочной железы и в слюне.
Карбоангидраза ускоряет реакцию гидратации двуокиси
углерода в цитоплазме эритроцита в 1500-2000 раз по сравнению с аналогичной
реакцией, протекающей в плазме крови, не содержащей карбоангидразы. Фермент в
зависимости от напряжения двуокиси углерода и от ряда других факторов может
ускорять реакцию гидратации молекул углекислого газа с образованием угольной
кислоты и бикарбонатных ионов. Гемоглобин присоединяет положительно заряженный
ион водорода - по два протона на каждые 4 освободившиеся молекулы кислорода,
что способствует поддержанию буферных свойств крови. В капиллярах
микроциркуляторного русла легких кислород, наоборот, присоединяется к молекуле
дезоксигемоглобина. Водород же присоединяется к бикарбонату, образуя угольную
кислоту, которая диссоциирует на воду и углекислый газ. Это приводит к
высвобождению двуокиси углерода из крови в альвеолярную смесь газов. В тканях
при снижении pH гемоглобин легко отдает кислород. Это обратимое явление лежит в
основе внешнего дыхания, которое было более подробно рассмотрено в главе о
биологической роли железа [4].
Витамин B12
- единственный
витамин в состав которого входит ион металла. Центральная структура
цианокобальмина - порфириноподобная кольцевая система, в которой пара
пиррольных колец связана между собой непосредственно, а не через группу =CH-.
Кобальт (II) находится в положении, которое в геме занимает железо (II).
Считается, что строение кобаламинов обусловливает их своеобразные
каталитические свойства.
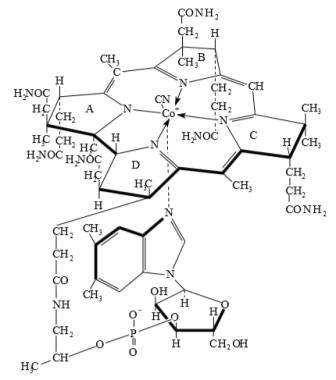
Рисунок 9. Строение витамина В12
Найдено большое число производных цианокобальмина, у
которых CN-группа замещена другим фрагментом. Установлено, что аналоги витамина
В12 являются кофакторами различных ферментов, участвующих в эритропоэзе
[9].
До 90% от общего количества в организме
цианокобальмина находится в виде кофермента, то есть участвует в ферментативных
реакциях.
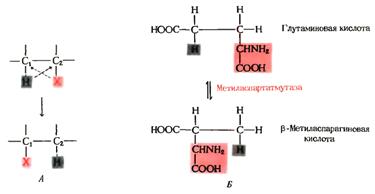
Рисунок 10. Реакции, происходящие с участием
коферментых форм витамина В12.
А. Ферменты катализируют перенос атома водорода от
атома С1 к атому С2 и обмен на группу Х.
Б. Реакция, катализируемая метиласпармутазой. Группы H и Х, присоединенные к соседним
атомам углерода, обмениваются местами.
Наиболее важную роль витамина В12 играет в
развитии и формировании эритроцитов.
Биологическая роль марганца
Марганец является биогенным элементом и одним из
десяти металлов жизни. Он принадлежит к числу немногих элементов, способных
существовать в восьми различных состояниях окисления. Однако в биологических
системах реализуются только два из этих состояний: Mn2+, Mn3+.
Особенностью биохимии марганца является способность
заменять ионы некоторых металлов на ион Mn2+. Это объясняется двумя факторами. Во-первых, близкие
значения атомных радиусов. Например, значение атомного радиуса марганца (128
пм) и железа (126 пм) почти одинаковое, и поэтому Mn (II) способен замещать Fe (II) в порфириновом комплексе эритроцита; по той же причине Mn
(II) может заменять и Zn (II). Во-вторых, предпочтение в качестве
лигандов одинаковых групп. Например, марганец и магний тяготеют к карбоксильным
и фосфатным группам, поэтому Mn (II) может заменять Mg (II) в комплексах с ДНК
и соединениях с АТФ [5].
Влияние марганца на жизнедеятельность очень
разнообразно и сказывается главным образом на росте, образовании крови и
функции половых желёз. Так же этот он необходим для активации ряда ферментов:
дегидрогеназ изолимонной и яблочной кислот, аргиназа, холинэстераза,
фосфоглюкомутаза, пируваткарбоксилаза, дезоксирибонуклеаза и т. д.
Малатдегидрогеназа или дегидрогеназа яблочной кислоты
- фермент, катализирующий последний этап цикла Кребса. Цикл Кребса является тем
центром, в котором сходятся практически все метаболические пути. Это общий
конечный путь окисления ацетильных групп органических молекул, играющих роль
«клеточного топлива»: углеводов, жирных кислот и аминокислот.
Гидролиз АТФ до АДФ играет важную роль в организме,
так как обеспечивает его энергией для жизнедеятельности. В организме этот
процесс активируется магнием в результате образования комплексов МgАТФ2- и МgАДФ-. Однако активацию
могут осуществлять и ионы Мn2+. При этом комплексы марганца более
активны, чем магния. Участие марганца в гидролизе можно представить в виде:
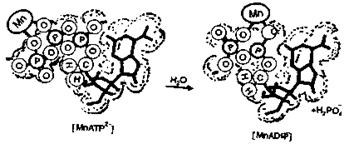
Рисунок 11. Схема гидролиза АТФ при усачтие ионов
марганца
Ионы Мg2+ и Мn2+ осуществляют также активацию нуклеаз. Эти ферменты
катализируют в двенадцатиперстной кишке гидролиз нуклеиновых кислот ДНК и РНК.
В результате они расщепляются на мономерные единицы - нуклеотиды. Наприме,
такой нуклеазой является дезоксирибонуклеаза, которая катализирует гидролиз ДНК
[3].
Связывание аммиака, токсичного продукта превращения
аминокислот в организме млекопитающих, осуществляется через аргинин. Аргиназа -
фермент, катализирующий в печени гидролиз аргинина. В результате аргинин
расщепляется на мочевину и орнитин. Мочевина - нетоксичное для организма
вещество. Оно с потоком крови попадает в почки и выводится с мочой.
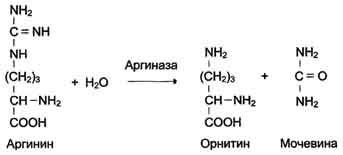
Рисунок 12. Схема гидролиза аргинина
Биологическая роль молибдена
Молибден является одним из важнейших биоэлементов. Как
и марганец, молибден может реализовать восемь различных степеней окисления. В
биохимических процессах он участвует в степенях окисления +5 и +6.
На примере молибдена можно проследить связь и
взаимовлияние метабиологической активности микроэлементов. Избыток молибдена
приводит к уменьшению концентрации меди и кобальта. Непосредственное
взаимодействие между Мо и Сu
может приводить к образованию в желудочно-кишечнем тракте труднорастворимого
соединения CuMoO4 [3].
Молибден образует устойчивые оксокомплексы и, видимо,
поэтому входит в состав ферментов, обеспечивающих перенос оксогрупп. Кроме
кислородных комплексов, молибден образует галогенидные, тиоцианатные и
цианидные комплексы.
Молибден входит в состав ряда ферментов. В организме
человека к ним относятся альдегидогидроксидазы, ксантиндегидрогеназы,
ксантиноксидазы и сульфитоксидаза.
Ксантиноксидаза - важнейший фермент в обмене азота. Он
катализирует оксиление пуринов. Конечным продуктом реакции является мочевая
кислота, которая затем выводиться из организма с мочой. В ходе ферментативной
реакции Mo6+ переходит в Mo5+, а потом в Mo4+.

Рисунок 13. Схема оксиления пурина при участии
ксантиноксидазы
Предполагается, что в ходе каталитического процесса
молибден образует связь с азотом и кислородом ксантина [3].
Так же ксантиониксидаза может катализировать окисление
альдегидов.
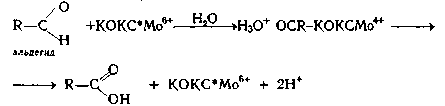
Рисунок
14. Схема оксиления альдегида при участии ксантиноксидазы
Сульфитоксидаза представляет собой димер, образованный
двумя идентичными субъединицами, каждая из которых содержит по одному атому
молибдена и одной молекуле цитохрома. Она превращает сульфит в сульфат и строго
специфична к своему субстрату.
SO32- + H2O → SO42- + 2H+ +2e
Эта реакция является конечной стадией окислительной
деградации цистеина, метионина и сульфатидов. Нарушение функции сульфитоксидазы
приводит к большим неврологическим отклонениям и ранней смерти. Фермент
присутствует в печени, где он локализуется в межмембранном пространстве
митохондрий. Его физиологический акцептор электронов - цитохром с [1].
Практическая часть. Колебательные
реакции
Перекись водорода. Для безопасной работы следует
использовать защитную одежду, защитные очки и резиновые перчатки; при попадании
на кожу или слизистые тщательно промыть водой; не ставить горючих материалов
вблизи места хранения перекиси водорода.
Концентрированная серная кислота. Работать с
концентрированной серной кислотой следует под тягой в защитной одежде, в
защитных очках и в резиновых перчатках. При попадании на кожу тщательно промыть
пораженное место водой и нейтрализовать кислоту содой.
Иодат и бромат калия. Соли являются сильными
окислителями, поэтому работать с ними следует в защитной одежде, в защитных
очках и резиновых перчатках. При попадании на кожу или слизистые тщательно
промыть водой.
Используемые посуда и реактивы
Для реакции Бриггса-Раушера: электронные весы, чашка
Петри, стеклянный шпатель, мерный стакан, мерный цилиндр, электрическая плитка,
кристаллизатор, мерная пипетка, магнитная мешалка, H2O2(30%), KIO3(тв), H2SO4(конц), крахмал, малоновая кислота, MnSO4·5H2O
Для реакции Белоусова-Жаботинского: электронные весы,
чашка Петри, стеклянный шпатель, мерный стакан, мерный цилиндр, мерная пипетка,
магнитная мешалка, KBrO3(тв), H2SO4(конц), Ce(SO4)2(тв), лимонная кислота.
Методика реакции Бриггса-Раушера
Готовится 3 раствора. Назовем их «раствор 1», «раствор
2» и «раствор 3».
Раствор 1. 40 мл 30% H2O2 доводят дистиллированной водой до
100 мл.
Раствор 2. Нагревают 60-70 мл дистиллированной воды, а
затем в ней растворяют 4,3 грамма иодата калия (из-за плохой растворимости
иодата). Потом этот раствор ставят на ледяную баню и добавляют 0,4 мл
концентрированной серной кислоты. После охлаждения раствора его объем доводят
до 100 мл.
Раствор 3. 30 мл дистиллированной воды доводят до
кипения. В другом стакане смешивают 0,3 г крахмала и 5 мл воды и тщательно
перемешивают. Затем эту суспензию вливают в закипевшую воду и снова
перемешивают. Еще в одном стакане готовиться раствор сульфата марганца(II) и малоновой кислоты. На 200 мл воды
добавляют 1,4 г пентагидрата сульфата марганца(II) и 5,2 г малоновой кислоты. Затем к нему приливают
крахмальный раствор и доводят объем конечного раствора до 300 мл.
Приготовленные растворы нужно слить в определенной
последовательности. Так же важно, чтобы конечный раствор тщательно
перемешивался, поэтому лучше проводить опыт на магнитной мешалке. Сначала
отбирают 100 мл раствора 3 и добавляют к нему раствор 2. После того, как
раствор немного пожелтеет, к нему добавляют раствор 1 и через секунд 30-60
наблюдаются колебания.
Методика реакции
Белоусова-Жаботинского
Как и в реакции Бриггса-Раушера, в данном процессе
важно постоянное перемешивание, поэтому опыт ведется на магнитной мешалке и
готовиться раствор объемом 30 мл и более (при меньших объемах не получиться
перемешивать раствор с помощью мешалки).
Отмеряют 40 мл дистиллированной воды. В ней растворяют
8 г лимонной кислоты. К раствору добавляют 8 мл разбавленной серной кислоты (1
часть концентрированной кислоты на 3 части воды). Затем в нем растворяют 0,64 г
сульфата церия(IV). Растворение
сульфата удобно проводить на магнитной мешалке. Так же важно добавлять данную
соль после серной кислоты, так как в ее присутствии она растворяется лучше, чем
в чистой воде. К конечному раствору добавляют 0,8 г бромата калия. В качестве
индикатора добавляют несколько капель ферроина.
Механизм колебательных реакций
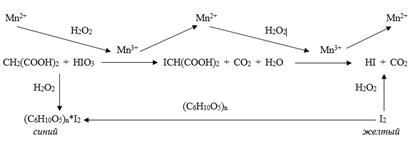
Для реакции Бриггса-Раушера механизм реакции можно
представить в виде схемы:
Механизм реакции Белоусова-Жаботинского может быть
описан следующим набором уравнений:
1) HOOC-CH2-C(OH)(COOH)-CH2-COOH
+ Ce4+ → HOOC-CH2-CO-CH2-COOH + Ce3+
+ CO2 + H2O
) Ce3+ + BrО3- → Ce4+ + Вr-
) Вr - + BrО3- → BrO- + BrО2-
3.1) Вr - + BrO- → Br2
3.2) 3Вr- + BrО2- → 2Br2
4) HOOC-CH2-CO-CH2-COOH
+ 5Br2 → Br3C-CO-CHBr2 + 5Вr- + 2CO2 + 5H+
Результаты опыта
Реакция Бриггса-Раушера. Цвет раствора колеблется от
желтого к синему в течении нескольких минут. Период колебаний около 30 секунд.
Раствор после окончания реакции имеет синюю окраску.
Реакция Белоусова-Жаботинского. Цвет раствора без
добавления ферроина колеблется от бесцветного к желтому (Ce3+ - бесцветный, Ce4+ - желтый). При добавлении ферроина
окраска меняется от красной к синей. Реакция продолжается около 15 минут с
периодом колебаний 1-1,5 минуты. В процессе реакции образуется белая взвесь.
Заключение
В данной работе было рассмотрено значение для
организма только пяти d-элементов,
но несмотря на это, можно проследить взаимосвязь биологической роли со степенью
окисления элементов.
Подводя итог, можно выделить следующие роли d-металлов в живых организмах:
· транспорт ионов металлов и других
неорганических компонентов;
· катализ реакций гидролиза;
· катализ
окислительно-восстановительных реакций;
· перенос электронов;
· перенос групп;
· активация и ингибирование ферментов;
· металлолигандный гомеостаз.
Осуществление вышеперечисленных реакций возможно
только из-за того, что d-металлы
способны находиться в разных степенях окисления, а также имеют высокую
склонность к комплексообразованию. Эти свойства переходных металлов объясняются
строением электронных оболочек этих элементов.
Из-за разнообразия биологического действия, соединения
d-металлов могут применяться в
диагностике и лечении множества заболеваний.
Список использованных источников
1. Бабенко О., Утупов Т., Аликулов З. А. Роль молибдоферментов в
токсичности этанола и нитратов.
. Белоусов Б. П. Периодически действующая реакция и ее механизм
//Химия жизнь - 1982. - №7. - С. 145-147.
. Ершов Ю. А., Попков В. А., Берлянд А. С. Общая химия.
Биофизическая химия. Химия биогенных элементов. - М. : Высш. шк., 2000.
. Ленинджер, А. (1985). Основы биохимии: в 3-х т. Т. 1-3.
. Логинова Н. В. Основные направления исследований в
бионеорганической химии. - 2008.
. Миронов А. Ф. Цитохромоксидаза. Строение и механизм действия.
. Муштакова С. П. Колебательные реакции в химии //Соросовский
образовательный журнал. - 1997. - Т. 7. - С. 31-36.
. Першин Р. В. Колебательные реакции, или химические ритмы //Химия
в школе. - 2002. - №7. - С. 62-70
. Улахович Н. А. Комплексы металлов в живых организмах
//Соросовский образовательный журнал. - 1997. - №. 8. - С. 27-32.