Превращение микроорганизмами соединений углерода
РЕФЕРАТ
По
дисциплине: «Микробиология»
На
тему: «Превращение микроорганизмами соединений углерода»
Выполнила:
студентка 2 курса 2 группы
Попова Алина
Олеговна
Научный
руководитель:
Галиакбарова Алсу
Анваровна
Москва
2015 г
Содержание
Введение
Превращение
микроорганизмами соединений углерода
Гликолиз
Пентозофосфатный
путь
Путь
Энтнера-Дудорова
Анаэробное
дыхание
Брожение
Аэробное
дыхание
Цикл
Кребса
Глиоксилатный
шунт
Строение
дыхательной цепи аэробов
Энергетическая
эффективность процесса полного расщепления глюкозы
Заключение
Список
литературы
Введение
Превращение микроорганизмами соединений углерода
происходит в соответствии с различным механизмами и имеет большое значение в
природе, а также широко используется в различных отраслях промышленности, в
медицине, биотехнологии.
Автотрофные микробы для превращения углекислоты,
не имеющей энергетических свойств, в органические энергетические соединения
нуждаются в тепловых источниках, которыми для них служит солнечная энергия или
химическая энергия окисления минеральных веществ. К фотоавтотрофам относят
цветные бактерии: зеленые содержат в цитоплазме хлорофилл, а пурпурные красный
или коричневый пигмент.
Основной процесс, возвращающий углекислоту в
атмосферу, - разложение органических соединений под влиянием микроорганизмов. В
природе существует множество типов процессов, вызывающихся определенными видами
микробов. Наибольшее значение имеют процессы брожения клетчатки, брожение
пектиновых веществ, спиртовое, молочнокислое брожение, а также уксуснокислое
окисление, маслянокислое брожение.
Известно, что полное окисление углеродного
субстрата до углекислого газа и воды может происходить у микроорганизмов с
помощью трех различных механизмов: в цикле трикарбоновых кислот, в
глиоксилатном цикле и в пентозофосфатном цикле. Поскольку большинство
микроорганизмов использует углеводы в качестве источника энергии, в первую
очередь глюкозу, возможны три пути ее расщепления: гликолиз; окислительный
пентозофосфатный путь; 2-кето-3-дезокси-6-фосфоглюконатный путь (КДФГ-путь).
Перечисленные выше пути катаболизма глюкозы
могут протекать при разных типах энергетического метаболизма: аэробное дыхание,
анаэробное дыхание, брожение. Спектр углеродных соединений, усваиваемых за счет
аэробного дыхания, значительно шире, чем в случае брожения.
Целью данной работы является изучение аэробных
путей превращения микроорганизмами соединений углерода.
В соответствии с целью работы были поставлены
следующие задачи:
) Цели, задачи работы
)Рассмотреть возможные пути расщепления глюкозы.
)Выделить основные типы метаболизма
микроорганизмов.
)Определить составляющие и принцип
функционирования аэробного метаболизма.
)Разобрать механизм дыхательной цепи аэробов и
оценить энергетическую эффективность аэробных процессов.
)Рассмотреть процессы образования органических
кислот и биотрансформации исходных субстратов, отличных от углеводов по своей
химической природе.
микроорганизм аэробный метаболизм
углерод
Превращение микроорганизмами соединений углерода
Полное окисление углеродного субстрата до
углекислого газа и воды может происходить у микроорганизмов с помощью трех
различных механизмов: в цикле трикарбоновых кислот, в глиоксилатном цикле и в
пентозофосфатном цикле. При функционировании каждого из этих циклов в клетке
происходит образование восстановленных пиридиннуклеотидов. Они могут быть
использованы либо для процессов восстановления в ходе биосинтеза, либо для
получения АТФ путем окислительного фосфорилирования. В последнем случае НАДН
становится донором электронов для электронно-транспортной цепи, в которую
входят такие белки-переносчики электронов, как флавопротеиды, убихиноны и
цитохромы, локализованные на внутренней мембране митохондрий.
Спектр углеродных соединений, усваиваемых за
счет аэробного дыхания, значительно шире, чем в случае брожения. К ним
относятся углеводы, жирные кислоты, н-алканы, одноуглеродные соединения
(метанол), ароматические соединения (фенол, резорцин, салициловая кислота и
т.п.).
Поскольку большинство микроорганизмов использует
углеводы в качестве источника энергии, в первую очередь глюкозу, рассмотрим
основные пути ее катаболизма. К настоящему времени у микроорганизмов хорошо
изучены три основных пути расщепления глюкозы:
) гликолиз, или фруктозодифосфатный путь, или
путь Эмбдена -Мейергофа - Парнаса;
) окислительный пентозофосфатный путь, или
гексозомонофосфатный путь, или путь Варбурга - Диккенса - Хореккера;
) 2-кето-3-дезокси-6-фосфоглюконатный путь
(КДФГ-путь), или путь Энтнера - Дудорова.
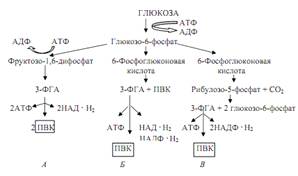
Рис. 1. Схема путей катаболизма глюкозы
(А - путь Эмбдена -Мейергофа - Парнаса, Б - путь
Энтнера - Дудорова, В - путь Варбурга - Диккенса - Хореккера)
Перечисленные выше пути катаболизма глюкозы
могут протекать при разных типах энергетического метаболизма: аэробное дыхание,
анаэробное дыхание, брожение.
Гликолиз
Все пути катаболизма начинаются с того, что
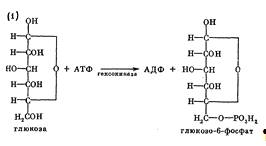
глюкоза, поступившая в клетку, сначала фосфорилируется при участии фермента
гексокиназы и АТФ как донора фосфата. Образуется глюкозо-6-фосфат, который
представляет метаболически активную форму глюкозы в клетке и служит исходным
соединением для любого из трех путей катаболизма углеводов.
Пути расщепления глюкозы состоят из многих
биохимических реакций, каждая из которых катализируется специфическим
ферментом.
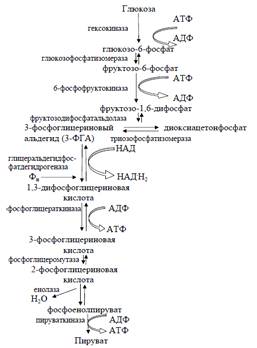
Рис. 2. Гликолитический путь расщепления глюкозы
Наиболее распространенным путем катаболизма
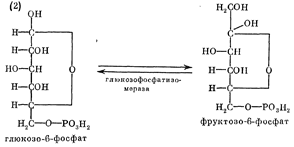
глюкозы у многих микроорганизмов является гликолиз (рис.2). Глюкозо-6-фосфат
под влиянием фермента глюкозо-6-фосфатизомеразы превращается в
фруктозо-6-фосфат (кето-альдольная изомеризация):
На образовавшийся фруктозо-6-фосфат, на первый
атом углерода, ферментом фосфофруктокиназой переносится от АТФ вторая фосфатная
группа (снова происходит фосфорилирование) образуется фруктозо-1,6-дифосфат:
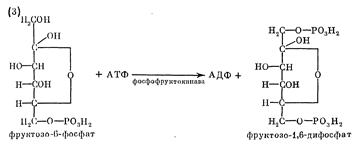
Следующая ступень - разрыв
фруктозо-1,6-дифосфата ферментом альдолазой на 2 фосфотриозы (расщепление связи
С-С): 3-фосфатглицериновый альдегид и диоксиацетонфосфат:
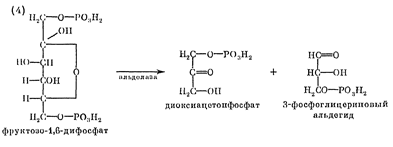
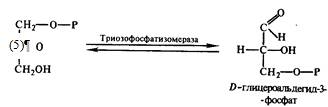
Затем диоксиацетон-3-фосфат подвергается реакции
кето-адольной изомеризации и под действием фермента триозофосфатизомеразы
превращается в 3-фосфатглицериновый альдегид:
Таким образом, из одной молекулы глюкозы
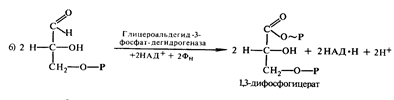
образуются две молекулы 3-фосфатглицеринового альдегида. Далее происходит
окисление каждой молекулы 3-фосфатглицеринового альдегида до
1,3-дифосфоглицериновой кислоты (1,3-ФГК) под действием фермента
глицеральдегид-3-фосфатдегидрогеназой.В этой реакции участвуют кофермент этого
фермента НАД+ и неорганический фосфат:
Фосфатная группа на первом углеродном атоме в
цепи присоединена макроэргической связью и может реагировать под действием
фосфоглицераткиназы с АДФ с образованием АТФ:
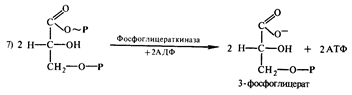
Затем 3-фосфоглицериновая кислота подвергается
перестройке под влиянием фосфоглицеромутазы и изомеризуется в
2-фосфоглицериновую кислоту:

После этого при отщеплении молекулы воды
(дегидратации) с участием фермента енолазы из 2-фосфоглицериновой кислоты
образуется фосфоенолпировиноградная кислота, обладающая макроэргической связью:
Фосфоенолпировиноградная кислота под влиянием
пируваткиназы отдает свою фосфатную группу и запас энергии молекуле АДФ с
образованием АТФ и пировиноградной кислоты:
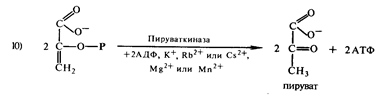
Таким образом, при распаде одной молекулы
глюкозы образуется четыре молекулы АТФ, в которых аккумулируется освободившаяся
энергия. Поскольку в начале процесса на активирование глюкозы были затрачены
две молекулы АТФ, чистый выход АТФ на одну молекулу глюкозы составляет две
молекулы. Суммарное уравнение гликолиза можно записать следующим образом:
С6Н12О6 + 2АДФ + 2H3PO4 + 2 НАД+→ 2С3Н4О3
+ 2АТФ + 2НАД · Н2
) Пентозофосфатный путь расщепления углеводов
характерен для некоторых представителей семейства Enterobacteriaceae, а также
для гетероферментативных молочнокислых бактерий, и некоторых маслянокислых
бактерий. В этом цикле глюкозо-6-фосфат, образующийся путем активирования
глюкозы молекулой АТФ, превращается через ряд промежуточных реакций в
6-фосфоглюконовую кислоту, которая подвергается окислению и декарбоксилированию
с образованием рибулозо-5-фосфата, СО2 и НАДФ · Н2 . Рибулозо-5-фосфат
включается в сложный цикл, приводящий к образованию из трех его молекул двух
молекул глюкозо-6-фосфата и одной молекулы 3-фосфоглицеринового альдегида. Глюкозо-6-фосфат
может снова включаться в цикл, а 3-фосфоглицериновый альдегид может быть
превращен в пировиноградную кислоту.
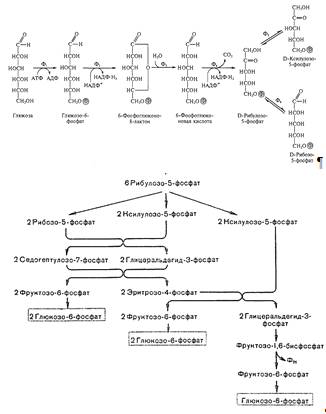
Рис. 3. Пентозофосфатный путь окисления
С энергетической точки зрения этот путь
катаболизма углеводов в 2 раза менее эффективен, чем гликолитический, так как
при окислении одной молекулы глюкозы образуется только одна молекула АТФ.
Однако большое значение этого пути в том, что он обеспечивает клетки бактерий
пентозами (рибулозо-5-фосфатом), которые являются предшественниками нуклеотидов
и нуклеиновых кислот. Кроме того, в этом цикле образуются две молекулы НАДФ ·
Н2, которые необходимы клетке для восстановительных реакций биосинтеза.
)Путь Энтнера - Дудорова встречается у прокариот
реже других. Он характерен в основном для псевдомонад и уксуснокислых бактерий.
У анаэробов он встречается довольно редко. От пентозофосфатного пути он
отличается тем, что 6-фосфоглюконовая кислота превращается в пировиноградную
кислоту и 3-фосфоглицериновый альдегид. Последний может превращаться в
пировиноградную кислоту. Специфичны для пути Энтнера-Дудорова 2 следующие
реакции:1)дегидратирование 6-фосфоглюконовой кислоты, приводящее к образованию
2-кето-3-дезокси-6-фосфоглюконовой кислоты; 2)расщепление продукта первой реакции
на 2 С3-фрагмента. Из одной молекулы глюкозы при функционировании этого пути
синтезируется одна молекула АТФ, по одной молекуле НАДФ · Н2 и НАД · Н2.
Следует подчеркнуть, что путь Энтнера - Дудорова является самым кратчайшим
механизмом расщепления углеводов до пировиноградной кислоты.
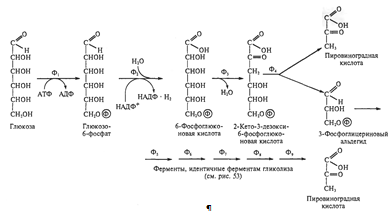
Рис. 4. Путь Энтнера-Дудорова
Таким образом, можно сделать вывод, что
важнейшим продуктом, образующимся при указанных путях метаболизма, является
пировиноградная кислота (ПВК), которая подвергается дальнейшим превращениям.
Пируват занимает центральное положение в метаболизме клеток и может служить
предшественником многих продуктов. Затем ПВК используется в таких процессах как
анаэробное дыхание, брожение, аэробное дыхание.
Анаэробное дыхание
При анаэробном дыхании конечным акцептором
электронов могут являться углеводы, в числе других органических веществ, но не
молекулярный кислород. Бактерии, способные к анаэробному дыханию, имеют
укороченную дыхательную цепь, так как не содержат всех тех переносчиков,
которые свойственны дыхательным цепям, функционирующим в аэробных условиях.
Цикл Кребса у строгих анаэробов не функционирует и не выполняет энергетических
функций, поэтому основное количество молекул АТФ синтиезируется в процессе мембранного
фосфорилирования.
Брожение
Брожением называется анаэробный процесс
превращения безазотистых органических веществ (главным образом углеводов)
микроорганизмами, при котором происходит накопление продуктов неполного
окисления (спиртов, органических кислот, углеводов и др.) и который
сопровождается выделением энергии. Биологическое значение брожения заключается
в образовании энергии для осуществления жизнедеятельности микроорганизмов
подобно дыханию животных и растений.
Различают следующие виды брожения по характеру
накапливающихся при брожении главных продуктов: молочнокислое,
пропионовокислое, масляно-кислое, ацетоноэтиловое и ацетонобутиловое,
анаэробное разложение клетчатки, которые вызываются различными бактериями, и
спиртовое брожение, вызываемое главным образом дрожжами.
Аэробное дыхание
При аэробном дыхании донором водорода или
электронов являются органические (реже неорганические) вещества, а конечным
акцептором - молекулярный кислород.
При аэробном дыхании пируват, образованный в
ходе гликолиза и пути Энтнера-Дудорова, полностью окисляется (то есть
превращается в СО2) в циклическом процессе, известном как цикл трикарбоновых
кислот (ЦТК) или цикл Кребса (рис. 5). До включения в цикл пируват с помощью
ферментов превращается в ацетил-КоА. ЦТК служит для полного окисления
двухуглеродного соединения -ацетата до С02 с отщеплением водорода.
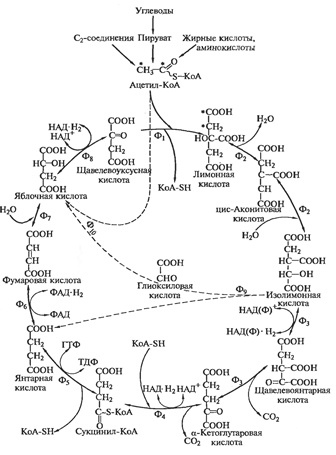
Рис. 5. Цикл Кребса. Цикл трикарбоновых кислот и
глиоксилатный шунт.
А)Рассмотрим процесс, в результате которого
пируват, образовавшийся при расщеплении глюкозы, окисляется до ацетил-КоА и СО2
при участии набора ферментов, объединенных структурно в так называемый
пируватдегидрогеназный комплекс. Эта мультиферментная система, находящаяся у
эукариотических клеток в митохондриях, а у прокариотических - в цитоплазме.
Этот процесс включает 5 стадий:
)Пируват теряет свою карбоксильную группу в
результате взаимодействия с тиаминпирофосфатом, присоединенным к
пируватдегидрогеназе (Е1). В результате образуется гидроэтильное производное
тиаминпирофосфата с гидроксиэтильной группой:

)Оксиэтильная группа окисляется с образованием
ацетильной, которая одновременно переносится на амид липоевой кислоты,
связанной с ферментом дегидролипоилацетилтрансферазой (Е2).
)Происходит перенос ацетильной группы на коэнзим
А с образованием конечного продукта ацетил-КоА:
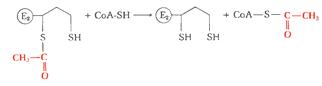
)При участии фермента дигидролипоилдегидрогеназы
(Е3) осуществляется перенос атомов водорода от восстановленных сульфгидрильных
групп дигидролипоамида на FAD:
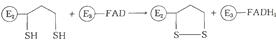
5)Восстановленная FAD-группа
дигидролипоилдегидрогеназы передает водород на NAD+ с образованием NADH:
Суммарную реакцию можно представить следующим
образом:
Б)Рассмотрим последовательность реакций,
протекающих в цикле Кребса. Образовавшийся в результате окисления
пировиноградной кислоты ацетил-КоА является исходным субстратом цикла
трикарбоновых кислот.
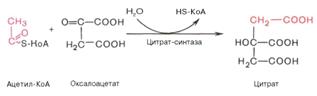
)Одна молекула ацетил-КоА вступает в реакцию
конденсации с оксалацетатом, катализируемую ферментом цитратсинтазой, в
результате чего образуется лимонная кислота и свободного коэнзима А:
)Лимонная кислота с помощью фермента аконитазы
подвергается изомеризации, в процессе которой происходит перенос гидроксигруппы
к другому атому углерода. Реакция идет через образование промежуточного
продукта цис-аконитата:
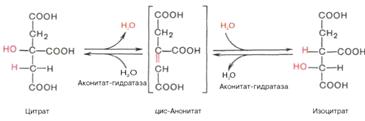
)Окислительное декарбоксилирование изоцитрата:
гидроксигруппа изоцитрата окисляется до карбонильной с помощью NAD+ и
одновременно отщепляется карбоксильная группа в β-положении.
Промежуточный продукт реакции - оксалосукцинат. Это первая реакция цикла, в
которой восстанавливается NAD+- кофермент фермента изотрат дегидрогеназы:
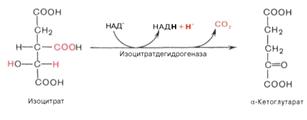
4)α-кетоглутаровая
кислота подвергается окислительному декарбоксилированию с образованием
высокоэнергетического соединения сукцинил-КоА, содержащем тиоэфирную связь. α-кетоглутаратдегидрогеназный
комплекс напоминает по своей структуре пируватдегидрогеназный комплекс:
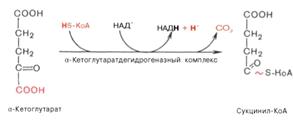
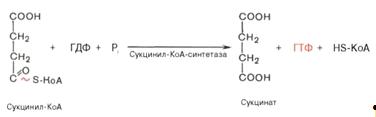
) Сукцинил-КоА при участии ГДФ и неорганического
фосфата превращается в сукцинат. Одновременно происходит образование
высокоэргической фосфатной связи ГТФ за счет высокоэргической тиоэфирной связи
сукцинил-КоА. Это единственная в цикле реакция субстратного фосфорилирования,
катализируемая ферментом сукцинил-КоА-синтетазой:
)Сукцинат дегидрируется в фумаровую кислоту.
Окисление сукцината катализируется сукцинатдегидрогеназой, в молекуле которой с
белком прочно (ковалентно) связан кофермент FAD:

)Образовавшаяся фумаровая кислота гидратируется,
под влиянием фермента фумаратгидратазы (фумаразы).Продуктом реакции является
яблочная кислота. Так как фумаратгидратаза обладает стереоспецифичностью - в
ходе реакции образуется L-яблочная кислота:

)Под действием NAD+-зависимой малатдегидрогеназы
L-малат дегидрируется и превращается в оксалоацетат:

Затем происходит конденсация образовавшейся
молекулы щавелевоуксусной кислоты с другой молекулой ацетил-КоА и запускается
следующий оборот цикла.
Суммарное уравнение цикла трикарбоновых кислот
можно представить:
ацетил-КоА + 3 NAD++ FAD+ГДФ +Н3РО4 →2СО2+
3NADН∙Н+ +ФАДН2+ГТФ
Цикл Кребса - это ключевой этап дыхания всех
клеток, использующих кислород, центр пересечения множества метаболических путей
в организме. Кроме значительной энергетической роли циклу отводится также и
существенная пластическая функция, то есть это важный источник
молекул-предшественников, из которых в ходе других биохимических превращений
синтезируются такие важные для жизнедеятельности клетки соединения как
аминокислоты,углеводы,жирныекислоты.
Но так как из цикла постоянно происходит
постоянный отток для биосинтезов промежуточных метаболитов, приводящий к
понижению уровня щавелево-уксусной кислоты, возникает необходимость в ее
дополнительном синтезе. У некоторых микроорганизмов это обеспечивается с
помощью последовательности из двух реакций, получивших название
11)глиоксилатного шунта (рис.4).В первой из них изолимонная кислота под
действием изоцитратлиазы расщепляется на янтарную и глиоксиловую кислоты. Во
второй реакции, катализируемой малатсинтетазой, глиоксиловая кислота
конденсируется с ацетил-КоА с образованием яблочной кислоты, превращающейся
далее в щавелево-уксусную кислоту. В результате двух новых реакций происходит
синтез С4-кислоты из двух С2-остатков. Глиоксилантный шунт не работает при
выращивании на субстратах, катаболизирование которых приводит к образованию
пировиноградной кислоты. Он включается при выращивании микроорганизмов на
С2-соединениях.
Передача (водорода) электронов на кислород
происходит не прямым путем, а в результате сложного многоступенчатого процесса
дыхательной цепи. Далее рассмотрим состав и функционирование цепи транспорта
электронов у аэробных микроорганизмов.
Строение дыхательной цепи аэробных
микроорганизмов
В переносе электронов от субстратов к
молекулярному кислороду принимают участие:
)Пиридинзависимые дегидрогеназы, для которых
коферментами служат либо NAD, либо NADF;
)Флавинзависимые дегидрогеназы (флавиновые
ферменты), у которых роль простетической группы играют FAD или FMN;
)Цитохромы, содержащие в качестве простетической
группы железопорфириновую кольцевую систему.
Среди компонентов дыхательной цепи обнаружены
также убихинон (кофермент Q) и железосерные белки.
Общую схему переноса водорода (электронов) к
молекулярному кислороду можно представить следующим способом:
Субстрат→ NAD∙H2→ Флавопротеин→
Кофермент Q→Цитохром b→
→ Цитохром с1 → Цитохром с →
Цитохром аа3→ О2
Дыхательная цепь у прокариот находится в
цитоплазматической мембране, а у эукариот - в мембране митохондрий. Дыхательная
цепь построена таким образом, что одни ее компоненты переносят только атомы
водорода, а а другие - только электроны. Причем переносчики атомов водорода и
переносчики электронов последовательно чередуются в дыхательной цепи.
Флавопротеины и хиноны осуществляют перенос атомов водорода, а FeS-белки и
цитохромы - электронов.
Функционирование дыхательной цепи осуществляется
следующим образом:
В присутствии восстановленного субстрата (S-H2)
- донора электронов и соответствуюзщей дегидрогеназы пиридиновое кольцо
восстанавливается путем связывания в четвертом положении одного протона и двух
электронов, а второй протон остается в среде:
Субстрат-Н2 +NAD+→Субстрат +NADH +H+
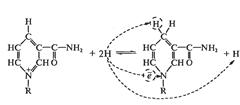
Следующим акцептором атомов водорода является
группа флавиновых ферментов:
NADH+ H+ +FAD → FADH2+ NAD+
В некоторых случаях, например окисления
сукцината, флавиновые ферменты могут играть роль первичных дегидрогеназ, т.е.
прямо, без участия NAD и NADF-зависимых дегидрогеназ, принимать электроны и
протоны при окислении субстратов.Такой фермент, в частности,
сукцинатдегидрогеназа.
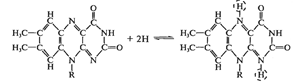
Восстановленная форма флавиновых ферментов
передает атомы водорода (протоны и электроны) убихинону (коферменту Q):
Кофермент Q + FADH2→Кофермент Q∙Н2+
FAD
Кофермент Q - жирорастворимое соединение,
которое служит «сборщиком» водорода, поставляемого различными коферментами и
простетическими группами. Здесь атом водорода расщепляется на протон и
электрон. Протон выделяется в среду, электрон передается на систему цитохромов,
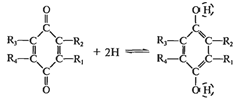
которые включаются в дыхательную цепь в определенной последовательности:
Цитохром b→ Цитохром с1 → Цитохром с
→ Цитохром аа3
Все цитохромы представляют собой гемопротеины.
При этом в обратимой окислительно-восстановительной реакции участвует атом
железа гемма, таким образом они переносят только электроны:

Каждый из этих цитохромов находясь в окисной
Fe3+ форме, присоединяет один электрон и переходит в закисную Fe2+ форму.
Последним в ряду переносчиков электронов стоит
цитохром аа3 (цитохромоксидаза), который является терминальным дыхательным
ферментом, непосредственно взаимодействующим с кислородом. Восстановленная
форма цитохрома Fe2+ вновь окисляется в Fe3+ форму молекулярным кислородом.
Образовавшийся «активный» кислород присоединяет два протона из окружающей
среды, в результате чего образуется молекула воды:
H++О2-→Н2О
Во время прохождения электронов по цепи энергия
освобождается отдельными небольшими порциями. Это осуществляется за счет
разницы окислительно-восстановительных потенциалов. Для синтеза 1 АТФ
достаточно 150-300 мВ. Такая разница достигается в электронно-транспортной цепи
в трех определенных местах, называемых пунктами сопряжения, так как именно там
находится АТФ-фаза.
Энергетическая эффективность процесса полного
окисления глюкозы
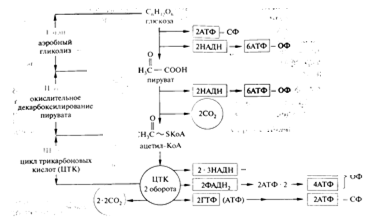
Рассмотрим 3 этапа энергетического выхода
аэробного дыхания(рис.6):
)Гликолитическое расщепление одной молекулы
глюкозы в аэробных условиях дает две молекулы пирувата, две молекулы NADH и две
молекулы АТФ:
Глюкоза +2Н3РО4+2АДФ +2 NAD+→2Пируват
+2АТФ+2 NADН+2H++2Н2О
Затем две молекулы NADН поступают в дыхательную
цепь:
NADН+2H++6Н3РО4+6АДФ+ О2→2 NAD++6АТФ+8 Н2О
)Окисление 2 молекул пирувата дает две молекулы
ацетил-КоА и две молекулы СО2:
Пируват+2 КоА+6Н3РО4+6АДФ+ О2→
2ацетил-КоА+ 2СО2+6АТФ+8Н2О
Затем две молекулы NADН, образованные в
результате этого окисления, поступают в дыхательную цепь:
NADН+2H++6Н3РО4+6АДФ+ О2→2 NAD++6АТФ+8 Н2О
) Окисление одной молекул ацетил-КоА до СО2 и
Н2О в цикле трикарбоновых кислот дает следующие продукты:
ацетил-КоА + 3 NAD++ FAD+ГДФ +Н3РО4+2О2 →
→2СО2+ 3NADН+Н+ +FАDН2+ГТФ
Образовавшийся ГТФ отдает отдает свою концевую
фосфатную группу на АДФ, вследствие чего образуется АТФ:
ГТФ + АДФ→ АТФ+ ГДФ;
Образовавшиеся три молекулы NADН и FАDН2
отправляются в дыхательную цепь.
Так как две молекулы пирувата окисляются с
образованием двух молекул ацетил-КоА, то цикл Кребса совершает два оборота для
полного расщепления глюкозы.
В итоге получаем суммарное уравнение для гликолиза
и дыхания:
Глюкоза+38Н3РО4+ 38АДФ+ 6О2→6 СО2 +38АТФ
+44 Н2О
Заключение
Микробы получают энергию в основном при
освобождении ее из безазотистых органических веществ. Различают две формы
катаболизма (энергетического обмена): брожение - без доступа свободного
кислорода и дыхание, или окисление - с кислородом. При дыхании энергия
освобождается полностью и конечными продуктами явялются СО2 и Н2О.
В данной работе проведено изучение основных, в
первую очередь аэробных, путей превращения микроорганизмами соединений
углерода. Выявлено, что процессы распада углеводов в микробной клетке
преследуют три основные цели: накопление энергии в форме макроэргических
связей; образование предшественников, необходимых для синтеза компонентов
клетки и метаболитов; создание окислительно-восстановительных механизмов,
необходимых для превращения этих предшественников в промежуточные или конечные
продукты. Все углеводы, ассимилируемые микроорганизмами, вступают в процессы
ферментативного распада, характерные для метаболизма глюкозы. При рассмотрении
этого вопроса представляют интерес три процесса: гликолиз; пентозофосфатный
(гексозомонофосфатный; апотомический) путь распада; путь Энтнера-Дудорова.
Также был подробно изучены состав и принцип функционирования аэробного метаболизма.
Важным вопросом стало изучение механизма
дыхательной цепи аэробов и сравнение энергетической эффективности аэробных
процессов расщепления углеводов и жиров.
Также в работе детально рассмотрены пути
образования уксусной, лимонной, щавелевой кислот, глюконовой кислоты,
разложение клечатки, окисление углеводородов, жиров и высокомолекулярных кислот
жирного ряда.
Список литературы
Борисов
Л.Б. Медицинская микробиология, вирусология, иммунология. М.: Медицинское
информационное агентство. - 2001. - 736 с.
Воробьева
Л.И. Промышленная микробиология. - М.: Изд-во МГУ, 1989. - 294с.
Голубев
В.Н., Жиганов И.Н. Пищевая биотехнология. - М.: Делипринт, 2001.- 123 с.
Гусев
М.В., Микробиология: учебник/ Гусев М.В., Минеева Л.А. - 3-е издание. -М:
Академия. - 2007. - 464 с.
Елинов
Н.П. Основы биотехнологии: Для студентов институтов. - СПб: Наука, 1995. - 600
с.
Медицинская
микробиология, вирусология и иммунология. Под ред. Академика РАМН А.А.
Воробьева М.: Медицинское информационное агентство. - 2004.- 691 с.
Мудрецова-Висс
К.А., Кудряшова А.А., Дедюхина В.П. Микробиология, гигиена и санитария. - М.:
Деловая литература, 2001. - 388 с.
Донецкая
Э.Г.-А. - Клиническая микробиология - ГЭОТАР Медиа, 2011.