Структурная организация молекул нуклеиновых кислот
ФЕДЕРАЛЬНОЕ
АГЕНТСТВО ПО ОБРАЗОВАНИЮ
ГОСУДАРСТВЕННОЕ
ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ
ВЫСШЕГО
ПРОФЕССИОНАЛЬНОГО ОБРАЗОВАНИЯ
«БАШКИРСКИЙ
ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ»
КУРСОВАЯ
РАБОТА
по
дисциплине «Биофизика полимеров»
на тему:
СТРУКТУРНАЯ ОРГАНИЗАЦИЯ МОЛЕКУЛ НУКЛЕИНОВЫХ КИСЛОТ
Выполнил(а):
Шагалина Гульназ Шагибаловна
Уфа 2013
Оглавление
Введение
. Структура и функция нуклеотидов
.1 Азотистые основания
.2 Моносахариды: рибоза и
дезоксирибоза
.3 Остаток фосфорной кислоты
. Физико-химические показатели
нуклеиновых кислот
.1 Выделение нуклеиновых кислот
.2 Хелатирующая способность
.3 Молекулярная масса, содержание и
локализация в клетке ДНК и РНК; виды ДНК и РНК
.4 Физико-химические свойства
нуклеиновых кислот
. Структура нуклеиновых кислот
.1 Правила Чаргаффа
.2 Первичная структура НК
.3 Вторичная структура НК
.4 Оптические характеристики
нуклеиновых кислот
.5 Третичная структура ДНК
Заключение
Литература
Введение
К числу важнейших научных событий нашего века
следует отнести открытие того факта, что генетическая информация кодируется
полимерной молекулой ДНК, образованной лишь четырьмя типами мономерных единиц.
Именно ДНК служит химической основой наследственности. Её молекула содержит в
своей структуре множество генов. ДНК направляет синтез РНК, которая в свою
очередь определяет синтез белка. Значение структуры и функции нуклеиновых
кислот необходимо для понимания сути генетических процессов происходящих в
клетке.
Следует разобрать понятие нуклеопротеид. Сложные
белки состоят из собственного белка и простетической (небелковой части), в роли
которой выступают различные вещества. Если роль небелкового компонента
выполняют нуклеиновые кислоты, то это - нуклеопротеид (нуклеиновая кислота -
белок), то есть сложный белок, а правильнее - надмолекулярный комплекс
(например - хроматин, рибосома). Простетической группой нуклеопротеидов
являются нуклеиновые кислоты, в связи с чем различают дезоксинуклеопротеиды
(ДНП) и рибонуклеопротеиды (РНП). Белковой частью нуклеопротеидов являются
простые белки: гистоны и негистоновые белки, чаще всего кислые белки, если
нуклеопротеиды локализуются в ядрах клеток; протамины - в нуклеопротеидах
сперматозоидов животных и рыб; другие простые белки (альбумины и глобулины),
если нуклеопротеиды локализованы в цитозоле.
В различных нуклеотидах количество нуклеиновых
кислот колеблется от 40 до 65% (например в рибосомах про- и эукариот). В
вирусных нуклеопротеидах количество нуклеиновых кислот не превышает 2- 5 %.
ДНП в клетке локализованы преимущественно в
ядрах, но встречается и в митохондриях, хлоропластах, а РНП - в цитозоле, но
некоторое количество обнаруживаются в ядрах и ядрышках.
В живом организме, в клетке существуют, как
известно два типа нуклеиновых кислот ДНК и РНК. Общее содержание их в клетке
зависит от функционального состояния ее: содержание ДНК в большинстве клеток
1-10%, в мышцах - около 0,2%. Содержание РНК, как правило, в 5-10 раз больше,
так в печени, активно синтезирующей белки соотношение РНК/ДНК составляет 4/10.
Особое положение занимают вирусы: у них генетический материал может быть либо
ДНК (ДНК-овые) или РНК (РНК-овые). Приблизительно 1-3% ДНК приходится на
внеядерную (митохондрии, хлоропласты). РНК в клетке распределяется более
равномерно: в клетках высших организмов в ядре -11%, митохондриях -15%, 50% в рибосомах
и 24% в гиалоплазме. нуклеотид химический нуклеиновый
кислота
Нуклеиновые кислоты - это высокомолекулярные
азотсодержащие органические соединения, структурной единицей которых являются
мононуклеотиды, соединенные в полинуклеотидную цепь 3'-5' фосфодиэфирными
связями, отвечающие за вид, форму, состав и функционирование клетки. Из
элементарного состава нуклеиновых кислот обращают внимание содержание фосфора
(10%) и азота (15-18%). Итак, нуклеиновые кислоты - это полимеры,
полинуклеотиды, кирпичиками которых является мононуклеотиды. В свою очередь
мононуклеотид состоит из азотистого основания (производные пурина или
приримидина), пентозы (рибозы или дезоксирибозы) и фосфорной кислоты.
Основная задача молекулярной биофизики состоит в
изучении структурных конформационных изменений НК. До недавнего времени
представлялась весьма жесткой и мало изменчивой макромолекулой. В
действительности, двойная спираль ДНК обладает изменчивой и гибкой структурой,
что очень важно для выполнения биологических функций в клетке. [10]
1. Структура и функция нуклеотидов
Каждый нуклеотид содержит 3 химически различных
компонента: гетероциклическое азотистое основание, моносахарид (рибофуранозу) и
остаток фосфорной кислоты. В зависимости от числа имеющихся в молекуле остатков
фосфорной кислоты различают нуклеозидмонофосфаты (НМФ), нуклеозиддифосфаты
(НДФ), нуклеозидтрифосфаты (НТФ) (рис. 1).
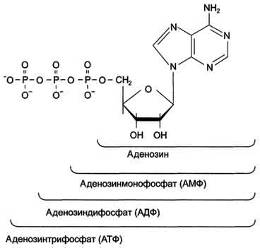
Рис. 1 - Различные нуклеозидфосфаты
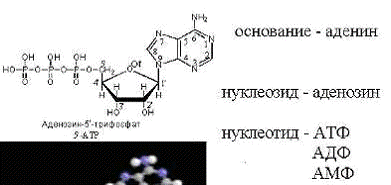
Рис. 2 - Три компонента нуклеотида
.1 Азотистые основания
В состав нуклеиновых кислот входят азотистые
основания двух типов: пуриновые - аденин (А),гуанин (G) и пиримидиновые -
цитозин (С), тимин (Т) и урацил (U) (рис. 3).
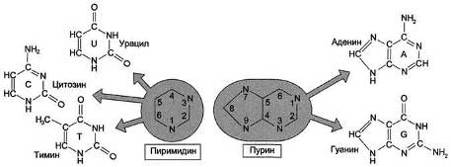
Рис. 3 - Пиримидиновые и пуриновые основания
Более подробно о пиримидине и пурине: пиримидин
С4Н4N2 - шестичленный гетероцикл с двумя
атомами азота.
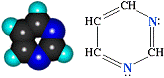
Проявляет свойства очень слабого
основания, т.к. атомы азота в sp2-гибридизованном состоянии довольно
прочно удерживают неподеленную электронную пару. Для пиримидина, подобно
пиридину, характерна 6р-электронная ароматическая система. Поэтому его цикл
обладает повышенной устойчивостью; пурин С5H4N4
- соединение, в молекуле которого сочетаются структуры шести- и пятичленного
гетероциклов (пиримидина и имидазола), содержащих по два атома азота.
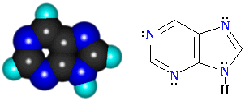
Проявляет амфотерные свойства.
Слабые основные свойства связаны с атомами азота шестичленного (пиримидинового)
цикла. Слабые кислотные свойства обусловлены группой N-H пятичленного цикла
имидазола (по аналогии с пирролом). Эти соединения являются основой
пиримидиновых и пуриновых оснований, входящих в состав природных
высокомолекулярных веществ - нуклеиновых кислот.
.2 Моносахариды: рибоза и
дезоксирибоза
Моносахариды - бесцветные
твердые кристаллические вещества, хорошо растворимые в воде, обычно сладкие на
вкус. К моносахаридам, входящим в состав нуклеиновых кислот, относятся рибоза и
дезоксирибоза.
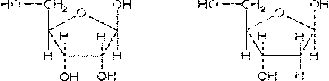
в-рибоза в-дезоксирибоза (C5H10O5)
(C5H10O4)
Рис. 4. Циклические формулы
моносахаридов, входящих в состав нуклеиновых кислот.
При частичном гидролизе нуклеотидов
отщепляется остаток фосфорной кислоты и образуются нуклеозиды, молекулы которых
состоят из остатка пуринового или пиримидинового основания, связанного с
остатком моносахарида - рибозы или дезоксирибозы. (В приложении приведены
структурные формулы основных пуриновых и пиримидиновых нуклеозидов). В случае
образования связи моносахарида с пуринами к названию добавляется суффикс -озин:
аденозин <#"823465.files/image007.gif">
Рис. 5. Адсорбционная хроматография
(схема). Разделение двух разных веществ (А и В), перемещающихся по колонке с
разной скоростью. 1.-нанесение образца на колонку; 2-середина опыта; 3-окончание
опыта.
Рис. 6. Фильтрация через агарозный
гель.
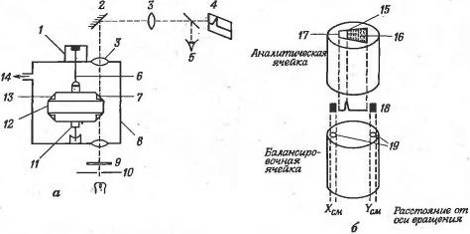
Рис. 7. Схематическое изображение
системы для аналитического ультрацентрифугирования (а) аналитической и
балансировочной ячеек (б). 1 - мотор; 2 - зеркало; 3 - линза; 4 -
фотопластинка; 5 - окуляр; 6 - гибкий вал; 7 - положение аналитической ячейки;
8 - камера ротора; 9 - светофильтр; 10 - источник света; 11- термистор; 12 -
ротор; 13-положение балансировочной ячейки; 14 - к насосу; 15 - граница; 16 -
раствор; 17- растворитель; 18 - шлирен-диаграмма; 19- индексные отверстия.
.2 Хелатирующая способность
В водных растворах нуклеиновые
кислоты проявляют свойства активных полидентатных лигандов. Полидентатность
нуклеиновых кислот обусловлено наличием ионизированных фосфатных групп и
полярных групп азотистых оснований (карбонильных, имино- и др.), способных к
образованию координационных связей с катионами металлов. С помощью
ионизированных фосфатных групп нуклеиновые кислоты хелатируют катионы щелочных
и щелочноземельных металлов, причем с катионами щелочных металлов нуклеиновые
кислоты образуют лабильные, а с катионами щелочноземельных металлов (Mg2+,Ca2+)
- более прочные комплексы. За счет взаимодействия с полярными группами азотистых
оснований нуклеиновые кислоты образуют достаточно стабильные комплексы с
катионами d-металлов.
.3 Молекулярная масса, содержание и
локализация в клетке ДНК и РНК; виды ДНК и РНК
Молекулярную массу ДНК определяют в
основном гидродинамическим и электронно-микроскопическим методами, хотя это
можно делать, измеряя светорассеяние растворов ДНК и некоторыми другими
способами.
В основе гидродинамического метода
лежит линейная зависимость константы седиментации ДНК, определяемой при
ультрацентрифугировании растворов ДНК, от ее молекулярной массы, которую можно
установить по калибровочной кривой или расчитать по формуле:
0,445lgM=1.819+lg(s20͵w-2.7), где
s20͵w -
константа седиментации, приведенная путем экстраполирования к бесконечному
разведению (s), стандартной температуре (20 С) и вязкости воды (w).
Электронно-микроскопический метод
определения молекулярной массы ДНК основан на измерении длины вытянутых молекул
ДНК. Известно, что на 0,1 нм протяженности ее молекулы приходится масса, равная
197 Да. Умножая это значение на экспериментально найденную длину, находят
значение молекулярной массы. Молекулярная масса эукариотических ДНК выше, чем
у ДНК прокариот (например, в одной из хромосом плодовой мушки дрозофилы она
достигает 7,9 х 1010). Кроме того, в состав митохондрий и
хлоропластов входят кольцевые молекулы ДНК с молекулярной массой 106-107.
ДНК этих органелл называют цитоплазматической; она составляет примерно 0,1%
всей клеточной ДНК.
Содержание ДНК в клетках организма
определенного вида отличается необыкновенным постоянством, тогда как межвидовые
различия по этому показателю достаточно велики. Количество ДНК в клетке
измеряется пикограммами (1012 г), колеблется от 0,01 пг у кишечной
палочки до нескольких пикограммов в гаплоидных клетках высших организмов.
Кроме внутриклеточной ДНК существует
также ДНК, входящая в состав вирусов и бактериофагов. Количество ее в вирусах
значительно ниже, чем в клетках бактерий (тысячные доли пикограмма).
Молекулярные массы РНК определяют
теми же методами, что и ДНК, но, кроме того, используют электрофорез в
полиакриламидном геле, так как пробег РНК в геле обратно пропорционален их
молекулярным массам. Что касается содержания и локализации РНК в клетках, то
оно не отличается ни однообразием, ни стабильностью: в клетках, где идет
интенсивный биосинтез белков, содержание РНК в несколько раз превышает таковое
ДНК (например, в печени крысы РНК в 4 раза больше, чем ДНК), но там, где синтез
белка мал, соотношение ДНК и РНК может быть обратным (например, в легких крысы
РНК в 2 раза меньше, чем ДНК).
По функциональному значению и
молекулярным массам, равно как и по локализации в клеточном содержимом, РНК
делят на следующие виды.
. Транспортные РНК (тРНК) отличаются
сравнительно невысокими значениями молекулярных масс (25-30 тыс. дальтон). тРНК
составляют 10 % от всех РНК. Эти РНК локализованы в гиалоплазме клетки, ядерном
соке, бесструктурной части хлоропластов и митохондрий и приобретают
специфическую конфигурацию в виде клеверного листа. Они осуществляют
кодирование аминокислот и перенос их в рибосомальный аппарат клетки в процессе
биосинтеза белков.
. Рибосомальные РНК (рРНК)
характеризуются в основном большими молекулярными массами (1-1,5 млн. дальтон),
молекулы крупные, в их состав входит до 5 000 нуклеотидов. Они локализованы в
рибосомах, являясь их структурной основой и выполняя в них разнообразные
функции (формирование активного центра рибосомы; Обеспечение взаимодействия
рРНК и тРНК).
. Информационные, или матричные, РНК
(мРНК) обладают молекулярными массами, варьирующими в широких пределах (от 300
000 до 4*106). мРНК синтезируются в ядре в процессе транскрипции на
определенном участке молекулы ДНК (гене). Функцией мРНК является перенос
генетической информации о структуре белка от ДНК к месту синтеза белка, на
рибосомы.
. Вирусные РНК отличаются
разнообразными и высокими молекулярными массами, лежащими в основном в пределах
нескольких миллионов дальтон. Они являются составными частями вирусных и
фаговых рибонуклеопротеинов, несут всю информацию, необходимую для размножения
вируса в клетках хозяина.
В современной литературе обсуждается
вопрос о целесообразности выделения в отдельные категории еще нескольких видов
РНК: ядерной, хромосомной, митохондриальных, низкомолекулярных регуляторных,
антисмысловых.
2.4 Физико-химические свойства
нуклеиновых кислот
В воде ДНК образует вязкие растворы,
при нагревании таких растворов до 60 °С или при действии щелочей двойная
спираль распадается на две составляющие цепи, которые вновь могут объединиться,
если вернуться к исходным условиям. В слабокислых условиях происходит гидролиз,
в результате частично расщепляются фрагменты -Р-О-СН2- с
образованием фрагментов -Р-ОН и НО-СН2, соответственно результате
образуются мономерные, димерные (сдвоенные) или тримерные (утроенные) кислоты,
представляющие собой звенья, из которых была собрана цепь ДНК.
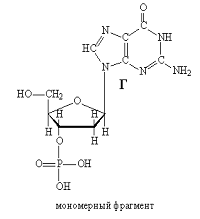
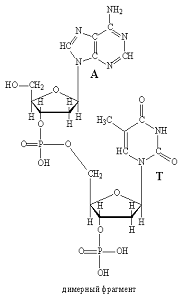
Рис. 8. Мономерный фрагмент. Рис. 9.
Димерный фрагмент.
Нуклеиновые кислоты и их растворы
проявляют кислотно-основные свойства; обладают хелатирующей способностью и способностью
к денатурации; оптические, коллоидные, осмотические свойства; высокая вязкость
растворов; в среде живой клетки могут находится в жидкокристаллическом
состоянии.
3. Структура нуклеиновых кислот
.1 Правила Чаргаффа
Для понимания ряда особенностей
структуры ДНК важное значение имели закономерности состава и количественного
содержания азотистых оснований, установленные впервые Э. Чаргаффом. Оказалось,
что азотистые основания ДНК обычно варьируют у разных видов организмов, однако
почти не претерпевают изменений у одного и того же вида в процессе развития или
в зависимости от изменений окружающей среды либо характера питания. Показано
также, что ДНК, выделенная из разных тканей одного и того же вида, имеет
одинаковый состав азотистых оснований. Полученные количественные соотношения
были названы правилами Чаргаффа. При анализе состава очищенной ДНК, выделенной
из разных источников, были сделаны следующие выводы:
1) молярная доля пуринов
<#"823465.files/image012.gif">
) количество аденина
<#"823465.files/image013.gif">
) количество аденина
<#"823465.files/image014.gif">
4) существенным для характеристики
вида (таксономическое значение) оказался так называемый коэффициент
специфичности <#"823465.files/image015.gif">
Это отношение часто выражают в
молярных процентах (Г + Ц), или процентах ГЦ-пар. Для животных и большинства
растений этот коэффициент ниже 1 (от 0,54 до 0,94), у микроорганизмов
<#"823465.files/image016.gif">
Рис. 10. Фосфодиэфирная связь
между нуклеотидами.
Второй, разработанный А. Максамом и В.
Гильбертом, сводится к химической модификации пуриновых и пиримидиновых
оснований в ДНК, избирательном расщеплении фосфодиэфирных связей по месту
модифицированных оснований, разделении продуктов расщепления электрофорезом в
полиакриламидном геле и прямому считыванию структуры фрагмента ДНК с полученных
электрофореграмм (число полос на электрофореграмме продуктов расщепления
фрагмента ДНК с модифицированным азотистым основанием равно числу нуклеотидных
остатков, содержащих этот пурин или пиримидин, а их положение на
электрофореграмме указывает место этого нуклеотидного остатка в анализируемом
фрагменте ДНК).
3.3 Вторичная структура НК
В соответствии с моделью Дж.
Уотсона и Ф. Крика, предложенной в 1953 г. на основании ряда аналитических
данных, а также рентгеноструктурного анализа молекула ДНК состоит из двух
цепей, образуя правовращающую спираль, в которую обе полинуклеотидные цепи закручены
вокруг одной и той же оси. Удерживаются цепи благодаря водородным связям,
образующимся между их азотистыми основаниями. Обе цепи полинуклеотидов в
биспиральной молекуле ДНК имеют строго определенное пространственное
расположение, при котором азотистые основания находятся внутри, а фосфорные и
углеводные компоненты - снаружи.
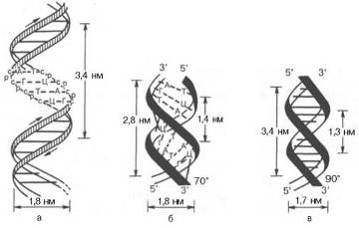
Рис. 11. Схематическое изображение
двойной спирали <#"823465.files/image018.gif">
Рис. 12. Две формы Na-соли ДНК
Позднее была открыта С-форма. В
таблице приведены характеристики А-, В-, С-форм ДНК. Они различаются значениями
шага двойной спирали, углами поворота между соседними парами оснований и наклоном
пар к оси спирали.
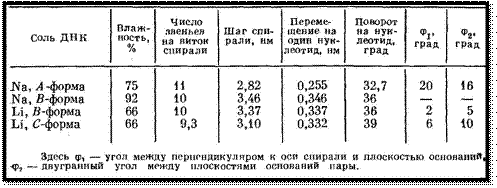
Конформация полинуклеотидной цепи
определяется набором углов поворота, показанных на рисунке:
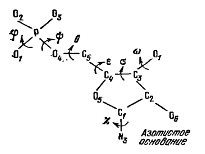
Углы отсчитываются следующим
образом: если смотреть вдоль связи, вокруг которой происходит вращение, то
дальняя связь вращается по часовой стрелке относительно ближней. Отсчет от
цис-положения связей.
Конформация полинуклеотидной цепи
зависит от конформации сахара - рибозы или дезоксирибозы. Сахар встречается в
четырех конформациях, различающихся тем, какой из атомов пятичленного цикла, С2
или С3 (на рисунке) выведен из плоскости остальных четырех атомов.
Если атом С2 (или С3) находится по ту же сторону от
плоскости, что и С5, то конформация именуется эндо, в
противоположном случае - экзо. Возможны, следовательно, С2-эндо, С3-эндо,
С2-экзо и С3-экзо-формы. Первые две встречаются чаще.
Эксперимент (диффузное рассеяние
рентгеновских лучей, КД и т.д.) и теория показали, что следует говорить не об
отдельных конформациях А, В, С ДНК, а о конформационных семействах А и В.
С-форма, а также Т- и D-формы сходны с В-формой и относятся
к В-семейству. Двойная спираль оказывается структурой, способной к непрерывному
изменению конформации. В пределах семейств А и В конформации меняются
некооперативно, вероятно, путем плавного изменения параметров спирали.
Напротив, переход от одного семейства к другому, АàВ,
происходит резко, кооперативно и сопровождается скачкообразным изменением
конформации сахара от 3-эндо к 2-эндо. Сведения о конформационной структуре
нуклеиновых кислот получаются непосредственно из данных рентгеноструктурного
анализа монокристаллов олигонуклеотидов. На рисунке показаны структуры двух
форм ДНК - В и Z.
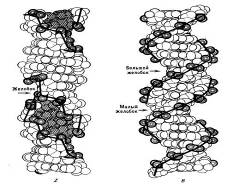
Рис. 13. Z и В-формы
ДНК.
Двойная спираль ДНК в любой
возможной ее конформации характеризуется широкой и узкой спиральными
бороздками, выемками на поверхности макромолекулы.
Двойные спирали ДНК в А-, В-,
С-формах - правые. Позднее была открыта левая двойная спираль ДНК. Так как она
зигзагообразна, эта форма была названа Z-формой. Z-формы
образуются цепями из чередующихся пуринов и пиримидинов. ДНК стабилизируются в Z-форме при
высокой концентрации соли или в результате бромирования или метилирования.
Работа с синтетическими ДНК
показала, что последовательности пурин-пиримидин могут испытывать переходы от
правой В-формы к левой Z-форме, в зависимости от
концентрации соли или химической модификации - от метилирования. Такие переходы
обратимы. Метилирование и демитилирование, по-видимому, существенны для
контроля активности генов.
Возможно и то, что конформационные
превращения ДНК существенны для ее функционирования, для взаимодействия с ферментами
(полимеразы), для регуляции действия генов и т.д.
Очевидно, что имеются две
возможности организации двойной спирали - из параллельных и антипараллельных
цепей. Справедливость второй модели доказана рентгенографическими и
биохимическими данными. Цепи в двойной спирали антипараллельны.
Специфическая двуспиральная
структура ДНК непосредственно объясняет важные факты - репликацию ДНК при
митозе и метаболическую устойчивость ДНК. При репликации двойная спираль
сначала разделяется вследствие разрыва водородных связей и раскручивания цепей.
Каждая из них служит матрицей для сборки новой цепи, комплементарной к матрице.
Мономеры новой цепи соединяются с матрицей, образуя уотсон-криковские пары АТ,
ТА, ГЦ и ЦГ. Одновременно происходит поликонденсация нуклеозидтрифосфатов, и в
результате образуются две двойные спирали, тождественные первоначальной. Такая
полуконсервативная модель (новая спираль содержит одну старую и одну новую
цепь) действительно подтверждается опытом. Ошибки при репликации являются точечными
мутациями. Наибольший гипохромизм, т.е. наибольшая степень двуспиральности,
наблюдается при составе смеси полинуклеотидов 1:1.
.4 Оптические характеристики
нуклеиновых кислот
Интенсивное поглощение нуклеиновых
кислот в УФ-области спектра обусловлено пуриновыми и пиримидиновыми
основаниями. Спектры поглощения отдельных оснований сливаются в одну широкую
полосу и дают характерное поглощение НК с лmax= 260 нм.
Квантово-механические расчеты
показали, что интенсивное поглощение света пуриновыми и пиримидиновыми
основаниямипри 260 нм связано с рàр*-
переходами. Определенный вклад в это поглощение вносят nàр-переходы.
Коэффициент молярной экстинции е260 для нуклеиновых кислот почти на
порядок больше, чем е280 для белков. Оптические свойства хромофорных
групп зависят от конформации ДНК. После тепловой денатурации оптическая
плотность D увеличивается примерно на 30%. Явление, связанное с увеличением
оптической плотности НК, называется гиперхромным эффектом. Уменьшенное
поглощение нативных препаратов ДНК есть гипохромный эффект.
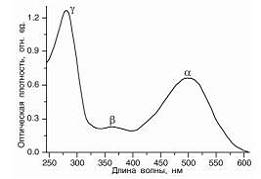
Рис. 14. Спектр поглощения ДНК.
На основании линейной
зависимости температуры плавления ДНК от содержания ГЦ-пар гиперхромный эффект
пытались объяснить разрывом водородных связей. Но исследования одноцепочечных
полинуклеотидов показали, что стабилизацию двойной спирали невозможно объяснить
только горизонтальными взаимодействиями комплементарных пар, стабилизированных
водородными взаимодействиями. Оказалось, что спектры поглощения поли-А и АМФ в
водном растворе, содержащем одинаковое количество нуклеотидов, значительно
отличаются. Интегральное поглощение поли-А существенно меньше, чем поглощение
составляющих его мономеров АМФ, т.е. для поли-А наблюдается гипохромный эффект,
аналогичный и для нативной ДНК. Для поли-А при увеличении температуры
наблюдается гиперхромный эффект.
В том же температурном
интервале для АМФ не наблюдается гиперхромного эффекта. На основании этих
экспериментальных данных сделано заключение, что наряду с горизонтальными
взаимодействиями имеют место вертикальные взаимодействия между основаниями.
Анализ упорядоченной структуры ДНК показывет, что плоскости оснований,
следующие друг за другом вдоль оси спирали, практически параллельны.
Компланарное расположение (стэкинг) оснований обусловливает вертикальные
взаимодействия и вызывает гипохромный эффект в нативной ДНК и в модельном
полинуклеотиде (поли-А) при низкой температуре.
Наибольшая энергия вертикальных
стэкинг-взаимодействий с таким расположением вдоль оси спирали комплементарных
пар: ↓ЦГ и ↑ГЦ. Она составляет - 82,8 кДж*моль-1.
Наименьшая энергия (6,7 кДж*моль-1) отвечает стэкингу ↑АТ и ↓ТА-пар.
Участки ДНК, содержащие АТ-пары, менее стабильны и быстрее денатурируются, чем
участки, содержащие ГЦ-пары. Гиперхромный эффект обусловлен разрушением
структуры ДНК и нарушением стэкинга оснований.
Зависимость оптической
плотности D260 ДНК от температуры называется кривой плавления ДНК. В
области низких температур для нативных молекул ДНК характерен гипохромный
эффект, обусловленный стэкинг-взаимодействиями между основаниями. Тепловая
денатурация ДНК происходит в узком температурном интервале и сопровождается
гиперхромным эффектом. Из кривых плавления ДНК определяется важная
характеристика - температура плавления Тп. Это - температура, при
которой гиперхромный эффект составляет D260/2.
3.5 Третичная структура ДНК
ДНК имеют формы: линейная, кольцевая, 2-х и 1-
цепочечная.
Двуцепочечные ДНК с «липкими» концами могут
образовывать кольцо, которое далее ковалентно сшивается по сахарофосфатной цепи
при помощи ДНК-лигазы.
Третичная структура ДНК у эукариотических клеток
отличается тем, что многократная спирализация ДНК сопровождается образованием
комплексов с белками.
хромосом (хроматид) человека организованы в 23
пары. Средняя длина хромосомы составляет 130 млн. пар оснований (п. о.) и имеет
длину 5 см. Хромосома № 1 - 263 млн. п. о., хромосома № 46 - меньше 50 млн. п.
о. Если проложить все ДНК в В-конформации в линию, то их общая длина превысит 2
метра. Человеческая хромосома 16 имеет 2,5 мкм в длину, а длина самой ДНК - 3,7
см.
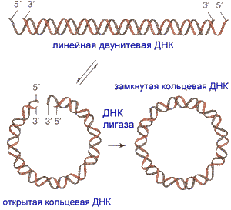
Рис. 15. Линейная, кольцевая третичные структуры
ДНК.
Понятно, что уместить такой длины ДНК в ядре
возможно только путем ее определенной упаковки. При образовании третичной
структуры ДНК человека происходит в среднем уменьшение ее размеров в 100 тысяч
раз.
Заключение
Нуклеиновым кислотам принадлежит очень важная
роль в обеспечении специфического синтеза биополимеров в организме человека,
животных, растений и микробов. В ядре клетки расположены хромосомы. В свою
очередь, хромосомы состоят из двух частей, соединенных перемычкой. В каждой из
этих частей находится хроматин. Он склонен образовывать спиралеобразные нити -
хроматиды, перед делением клетки. В хроматидах содержится ДНК. Какой компактной
должна быть структура ДНК, чтобы вместиться в хромосому! Именно в ней и
содержится наследственная информация или ген. ДНК имеет структуру закрученных
ступенек и определяется, как двойная закрученная спираль. Ее составляют две
очень тонкие нити, соединенные перемычками. Единицей этой структуры является
нуклеотид. Нуклеотид включает в себя 3 компонента: сахар, остаток фосфорной
кислоты и азотистого основания (тимин, аденин, цитозин а также гуанин).
Ступеньки спиральной лестницы как раз образованы последним, т.е. соединением
аденин с тимином и цитозин с гуанином, причем их связь водородная. Перилами же
лестницы являются остатки фосфорной кислоты, а сахар - это связующая в общей
цепи молекулы ДНК. Гены или отдельные фрагменты составляют молекулу ДНК. Репликация
ДНК Прямо перед делением клетки посредством митоза, ее ДНК реплицируется, т.е.
удваивается.
Структура двойной спирали характерна для
подавляющего большинства ДНК. Но в настоящее время известны разные конформации
молекулы ДНК.
Спектры поглощения растворов ДНК позволяют
выяснить оптическую плотность ДНК, Т плав.
Измерение спектра поглощения раствора ДНК
проводят на спектрофотометре. В свою очередь измерения используются для
последующего расчета концентрации растворов нуклеиновых кислот. Метод определения
концентрации нуклеиновых кислот в растворе основан на существовании у ДНК и РНК
максимума поглощения при длине волны 260 нм. Это означает, что в растворах
нуклеиновых кислот максимальная фотометрическая абсорбция наблюдается при 260
нм и прямо коррелирует с концентрацией ДНК или РНК.
Литература
1. А.С. Спирин Молекулярная
биология: Структура и биосинтез нуклеиновых кислот, 1986. - 496с.
. А.Ю. Ершов Общая химия.
Биофизическая химия. Химия биогенных элементов. Учебник для вузов, 2003. -
423с.
. А.Я. Николаев Биологическая химия,
2004 - 254с.
. Аппель Б., Бенеке И., Бенсон Я.,
под ред. С. Мюллер Нуклеиновые кислоты от А до Я, 2012. - 352с.
. В.Ф. Антонов и др. Биофизика:
Учебник, 2000. - 288с.
. И.Ф. Шаяхметов. Основы
биотехнологии растений: Учебное пособие, 2007. - 136с.
. Л.Н. Стрельникова ред. Химия и
жизнь, научно-популярный журнал, 2010. - 68с.
. М.В. Волькенштейн. Биофизика,
1988. - 588с.
. М.В. Волькенштейн. Молекулярная
биофизика, 1990. - 650с.
. П.Г. Костюк, Д.М. Гроздинский,
В.Л. Зима и др. Биофизика, 1988. - 505с.
. Ревин В.В., Максимов Г.В.
Биофизика. Учебник, 2000. - 206с.
. Рубин А.Б. - Биофизика (том 1,2),
2003. - 642с.
. Э. Чаргафф, Дж. Дэвидсон.
Нуклеиновые кислоты, 1962. - 456с.
. Ю.А. Владимиров, Д.И. Рощупкин
Биофизика, 1999. - 480с.
. Ю.Б. Филиппович Основы биохимии,
1993. - 495с.