Уровни организации живого
Реферат
Биология
молекулярный живой природа
биохимические
Биология - это наука о живом. Это целый комплекс
научных дисциплин, изучающих живые организмы, их строение, функционирование,
распространение, происхождение и развитие, а также природные сообщества
организмов, их связи друг с другом, с неживой природой и человеком. Различные
биологические дисциплины изучают живое на разных уровнях его организации.
Стольких уровней организации, как в биологии, нет ни в какой другой
естественной науке.
Существование и развитие неживой природы
определяется сложными физико-химическими процессами, фундаментальными и для
живой природы. Однако, с появлением живых организмов (принципиально
отличающихся по своим свойствам от тел наживой природы) начинают осуществляться
биологические процессы, имеющие специфический характер и подчиняющиеся новым
законам - биологическим. Таким образом, физико-химические процессы в живой
природе являются фундаментальными, первичными, а биологические, возникающие на
их основе, - производными, вторичными.
Признаком живого на молекулярном уровне служат
чрезвычайно многообразные органические соединения. Они являются как
структурными, так и функциональными компонентами организмов, играя важную роль
в процессах обмена веществ и энергии. Основой живого или, другими словами,
субстратом жизни являются белки и нуклеиновые кислоты - биополимеры,
находящиеся в тесном взаимодействии и взаимозависимости. Белки не только
строительный материал живого, они играют важнейшую роль во всех жизненных функциях
(в том числе и в процессе синтеза нуклеиновых кислот), выступая в качестве
биокатализаторов (белки - ферменты). Нуклеиновые кислоты, в свою очередь,
предопределяют структуру всех белков, синтезируемых в организме. Всем живым
организмам на Земле присущ универсальный генетический код - каждой из двадцати
аминокислот, образующих все белки организма, соответствует определенная
последовательность трех нуклеотидов - триплет - в полинуклеотидной цепи.
Таким образом, характерной чертой субстрата
жизни является его структурная организация. Живое вещество, построенное из тех
же химических элементов, что и неживое, характеризуется чрезвычайной сложностью
химических соединений, обусловленной определенной упорядоченностью на
молекулярном уровне. Упорядоченность в пространстве сопровождается
упорядоченностью во времени, обеспечивающей строгую последовательность
процессов, протекающих в живых системах.
Но что же такое жизнь и живое? До сих пор не
сформулировано более-менее приемлемого определения жизни.
Классическим определением жизни стала
формулировка Ф. Энгельса: «Жизнь - это способ существования белковых тел,
существенным моментом которого является постоянный обмен веществ с окружающей
их внешней природой, причем с прекращением этого обмена веществ прекращается и жизнь,
что приводит к разложению белка», И далее: «... обмен веществ состоит в
поглощении веществ, химический состав которых изменяется, которые
ассимилируются организмом и остатки которых выделяются вместе с порожденными в
процессе жизни продуктами разложения самого организма». Этот тезис дополняется
весьма существенным примечанием самого Ф. Энгельса: «И у неорганических тел
может происходить подобный обмен веществ, который и происходит с течением
времени повсюду, так как повсюду происходят, хотя бы и очень медленно,
химические действия. Но разница заключается в том, что в случае неорганических
тел обмен веществ разрушает их, в случае же органических тел он является
необходимым условием их существования». Определение Ф. Энгельса намного
опередило свое время и можно только поражаться тому, как при состоянии науки
того времени ему удалось увидеть главное и указать на самое принципиальное в
характеристике сущности живого.
Выдающийся биохимик нашего времени академик
В.А.Энгельгардт отмечал, что «в способности живого создавать порядок из
хаотического теплового движения молекул состоит наиболее глубокое, коренное
отличие живого от неживого. Тенденция к упорядочению, к созданию порядка из
хаоса есть ни что иное, как противодействие энтропии». Более образно по этому поводу
высказывался выдающийся физик XX века Э. Шредингер: «Живой организм может
избегнуть состояния максимальной энтропии, которое представляет собой смерть,
только путем постоянного извлечения отрицательной энтропии из окружающей его
среды. Отрицательная энтропия вот то, чем организм питается. Или, чтобы
выразить это менее парадоксально, существенно в метаболизме то, что организму
удается освобождать себя от всей той положительной энтропии, которую он
вынужден производить, пока он жив».
Обобщая достижения современного естествознания в
области теории открытых диссипативных систем, известный биофизик М.В.
Волькенштейн определил живые тела, существующие на Земле, как «открытые
саморегулирующиеся и самовоспроизводящиеся системы, состоящие из биополимеров:
белков и нуклеиновых кислот». Живое никогда не бывает в состоянии
термодинамического равновесия и постоянно совершает работу против достижения
равновесия. Термодинамическое равновесие для живого - это смерть.
Но все высказанные выше характеристики и
определения живого справедливы в той или иной мере и по отношению ко многим
другим неживым неравновесным системам. Кристаллы самовоспроизводятся -
наращивают сами себя, многие природные неживые системы с отрицательными
обратными связями саморегулируются, вихревые потоковые системы создают порядок
из хаоса и производят работу против термодинамического равновесия.
У живых и неживых неравновесных систем нет
принципиальных отличий - свойства всех неравновесных систем подчиняются одним и
тем же законам, но живые системы являются из всех неравновесных систем самыми
сложными:
· Через живые системы постоянно
проходят огромные потоки информации - до 1026 бит.
· Живые системы обладают сложнейшей
саморегуляцией - только на биохимическом уровне они содержат более 50 тыс
отрицательных обратных связей.
· Живые системы представлены огромным
биоразнообразием - более 10 млн видов живых организмов.
· Биологические системы обладают
удивительно развитыми способностями к изменчивости и прогрессивной эволюции,
способностями к самоусложнению ради более эффективного сохранения своего
состояния неравновесия.
· Живые системы обладают способностью
к опережающему отражению - они активно собирают информацию об окружающей среде
и на ее основании предвидят будущие события и выдают опережающие реакции. Они
как бы несут в себе элементы будущего в настоящем.
· Живые системы имеют огромное число
уровней организации - такого числа нет ни у одной неживой неравновесной
системы.
Уровни организации живого
Существование жизни на всех уровнях определяется
структурой низшего уровня. Например, характер клеточного уровня организации
определяется молекулярным и субклеточным уровнями; организменного - клеточным,
тканевым, органным; популяционно-видового - организменным и т.д. Следует
отметить большое сходство дискретных единиц на низших уровнях и все
возрастающее различие на высших уровнях.
По подходу к изучению биологических систем
выделяют следующие уровни организации живой материи на базе разных способов
структурно-функционального объединения составляющих элементов:
Молекулярный уровень. Основу организации на этом
уровне представляют 4 азотистых основания, 20 аминокислот, несколько сотен
тысяч биохимических реакций, почти все из которых сопряжены с синтезом или
разложением АТФ - универсального энергетического компонента живого.
Клеточный уровень. Клетка является минимальной
единицей жизни. Все живое состоит из клеток. Основные механизмы воспроизводства
жизни работают именно на клеточном уровне. Для одноклеточных организмов этот
уровень является также и организменным.
На клеточном уровне функционируют три основных
механизма воспроизведения и реализации генетической информации - репликация
(удвоение молекулы ДНК), транскрипция (синтез комплементарной РНК на матрице
генного участка ДНК) и трансляция (синтез молекулы белка).
Существует так называемая Основная догма
молекулярной биологии - она определяет направление потоков передачи
генетической информации в клетке. Информация передается от ДНК к ДНК во время
репликации; от ДНК к РНК и обратно во время транскрипции и обратной
транскрипции; и от РНК к белку во время трансляции. Передача информации от
белка к РНК или ДНК НИКОГДА не происходит ни у одного живого организма.
Механизмов для такой передачи информации в живой клетке не существует.
Репликация транскрипция трансляция
ДНК ↔ ДНК ↔ РНК → Белок
На клеточном уровне происходит два основных
процесса, необходимых для самовоспроизведения жизни - митоз - деление клетки с
сохранением числа хромосом и генов, и мейоз - редукционное деление, необходимое
для производства половых клеток - гамет. В мейозе также происходит рекомбинация
генетического материала клетки - хромосомная и кроссинговерная, дающая
рекомбинационную изменчивость.
На клеточном уровне функционируют три основные
энергетические цепи, обеспечивающие клетку и организм энергией.
Первый из них - фотосинтез. Это фотолитическое
восстановление углерода из углекислого газа, сопряженное с переносом электронов
на кислород молекулы воды и высвобождением свободного кислорода. Фотосинтез,
которым владеют растения и синезеленые водоросли - практически единственный
путь получения всей биосферой энергии и органических соединений (за исключением
хемосинтезирующих бактерий, окисляющих соединения серы и др.). При этом
используется световая энергия Солнца, поступающая на Землю. Только благодаря
фотосинтезу Земля имеет в своей атмосфере свободный кислород. Фотосинтез
происходит у растений - в хлоропластах, у синезеленых - в тилакоидах. У
растений около 100 генов, кодирующих ферменты, необходимые для осуществления
фотосинтеза, находятся в хлоропластном геноме. Остальные несколько сотен
необходимых для фотосинтеза генов расположены в ядерном геноме.
Все живые организмы (за исключением анаэробных
бактерий) обладают двумя основными механизмами извлечения энергии, запасенной в
процессе фотосинтеза в сахарах (глюкоза) и подобным им соединениям. Это
гликолиз и окислительное фосфорилирование. В результате этих процессов
химическая энергия глюкозы переводится в энергию химической связи в молекуле
АТФ - аденозинтрифосфата - универсального поставщика энергии внутри клетки.
Гликолиз - это цепь из нескольких десятков
ферментативных реакций, в результате которых глюкоза превращается либо в ацетат
- уксусную кислоту, либо в этиловый спирт, или схожие с ними относительно
простые соединения. Этот процесс идет без доступа свободного кислорода, в его
результате выделяется углекислый газ. Наиболее яркий пример гликолиза - процесс
брожения, обеспечиваемый одноклеточными грибами - дрожжами.
Но гораздо более эффективным в смысле получения
энергии оказывается окислительное фосфорилирование. Этот процесс идет в
биохимическом цикле, называемом циклом Кребса, или циклом трикарбоновых кислот.
В цикле Кребса потребляется свободный кислород атмосферы и глюкоза окисляется
им до углекислого газа и воды. Происходит полное сгорание глюкозы. В цикле
Кребса на одну молекулу глюкозы синтезируется в 14 раз больше молекул АТФ, чем
при гликолизе. Цикл Кребса происходит на мембранах митохондрий - особых
клеточных органелл, также, как и хлоропласты, имеющих свой собственный геном.
Но в геноме митохондрий находится только около десятка генов, кодирующих
ферменты окислительного фосфорилирования. Остальные несколько десятков генов -
до ста, необходимых для этого процесса, давно в процессе эволюции были
перенесены в ядерный геном.
Тканево-органный уровень. Ткань -
совокупность клеток <#"650414.files/image001.jpg">
Репликацию ДНК осуществляет фермент
ДНК-полимераза. Этот фермент способен наращивать ДНК только на 3΄-
конце.
Молекула ДНК антипараллельна, разные ее концы называются 3΄-конец
и 5΄
- конец.
При синтезе новых копий на каждой нити одна новая нить удлиняется в направлении
от 5΄
к
3΄
, а
другая - в направлении от 3΄ к
5-концу. Однако 5΄ конец
ДНК-полимераза наращивать не может. Поэтому синтез одной нити ДНК, той, которая
растет в "удобном" для фермента направлении, идет непрерывно (она
называется лидирующая или ведущая нить), а синтез другой нити осуществляется
короткими фрагментами (они называются фрагментами Оказаки в честь ученого,
который их описал). Потом эти фрагменты сшиваются, и такая нить называется
запаздывающей, в целом репликация этой нити идет медленней. Структура, которая
образуется во время репликации, называется репликативной вилкой.
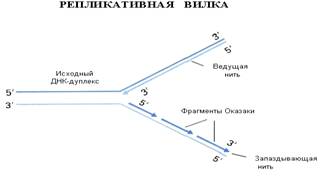
Если мы посмотрим на реплицирующуюся ДНК
бактерии, а это можно наблюдать в электронном микроскопе, мы увидим, что у нее
вначале образуется "глазок", затем он расширяется, в конце концов,
вся кольцевая молекула ДНК оказывается реплицированной. Процесс репликации
происходит с большой точностью, но не абсолютной. Бактериальная ДНК-полимераза
делает ошибки, то есть вставляет не тот нуклеотид, который был в матричной
молекуле ДНК, примерно с частотой 10-6. У эукариот ферменты работают точнее,
так как они более сложно устроены, уровень ошибок при репликации ДНК у человека
оценивается как 10-7 - 10 -8 . Точность репликации может быть разной на разных
участках геном, есть участки с повышенной частотой мутаций и есть участки более
консервативные, где мутации происходят редко. И в этом следует различать два
разных процесса: процесс появления мутации ДНК и процесс фиксации мутации. Ведь
если мутации ведут к летальному исходу, они не проявятся в следующих
поколениях, а если ошибка не смертельна, она закрепится в следующих поколениях,
и мы сможем ее проявление наблюдать и изучить. Еще одной особенностью
репликации ДНК является то, что ДНК-полимераза не может начать процесс синтеза
сама, ей нужна «затравка». Обычно в качестве такой затравки используется
фрагмент РНК. Если речь идет о геноме бактерии, то там есть специальная точка
называемая origin (исток, начало) репликации, в этой точке находится
последовательность, которая распознается ферментом, синтезирующим РНК. Он
относится к классу РНК-полимераз, и в данном случае называется праймазой.
РНК-полимеразы не нуждаются в затравках, и этот фермент синтезирует короткий
фрагмент РНК - ту самую «затравку», с которой начинается синтез ДНК.
Транскрипция. Транскрипция - синтез РНК на ДНК,
то есть синтез комплементарной нити РНК на молекуле ДНК осуществляется
ферментом РНК-полимеразой. У бактерий, например, кишечной палочки - одна
РНК-полимераза, и все бактериальные ферменты очень похожи друг на друга; у
высших организмов (эукариотов) - несколько ферментов, они называются
РНК-полимераза I, РНК-полимераза II, РНК-полимераза III, они также имеют
сходство с бактериальными ферментами, но устроены сложнее, в их состав входит
больше белков. Каждый вид эукариотической РНК-полимеразы обладает своими специальными
функциями, то есть транскрибирует определенный набор генов. Нить ДНК, которая
служит матрицей для синтеза РНК при транскрипции называется смысловой или
матричной. Вторая нить ДНК называется некодирующей (комплементарная ей РНК не
кодирует белки, она "бессмысленная").
В процессе транскрипции можно выделить три
этапа. Первый этап - инициация транскрипции - начало синтеза нити РНК,
образуется первая связь между нуклеотидами. Затем идет наращивание нити, ее
удлинение - элонгация, и, когда синтез завершен, происходит терминация,
освобождение синтезированной РНК. РНК-полимераза при этом «слезает» с ДНК и
готова к новому циклу транскрипции.
Элонгационный комплекс довольно стабилен, т.к.
он должен выполнить большую работу. То есть, сам по себе он с ДНК не
«свалится». Он способен перемещаться по ДНК со скоростью до 50 нуклеотидов в
секунду. Этот процесс называется перемещение (или, транслокация).
В результате траскрипции образуется нить
одноцепочечной РНК, которая отходит от ДНК-РНК-полимеразного комплекса и по
пути к рибосомам цитоплазмы (где происходит трансляция - синтез белка)
подвергается дальнейшему процессингу (разрезание на части, обрезание концов и
вырезание некодирующих частей).
Трансляция. Перейдем к трансляции - синтезу
белков. Трансляция происходит на рибосомах и катализируется ими. Рибосома
состоит из двух субчастиц: большой и малой.
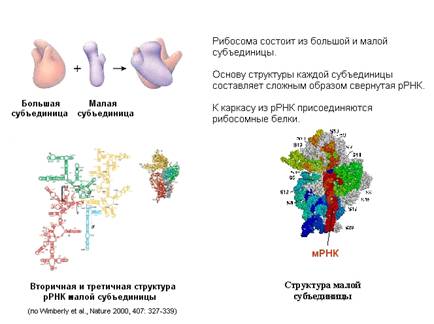
Каждая субчастица состоит из нескольких десятков
белков. Белки в рибосоме держатся на каркасе, состоящем из рибосомной РНК.
Формирование рибосомы начинается с того, что рибосомная РНК сворачивается и на
нее в определенном порядке начинают налипать белки. На рисунке представлена
рибосомная РНК. В ней самокомплементарные участки нити РНК спариваются, образуя
шпильки (вторичная структура), и затем РНК сворачивается (третичная структура
РНК), образуя каркас субчастиц.
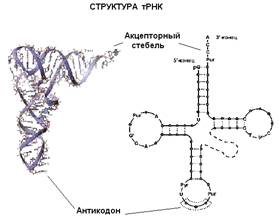
Еще один вид РНК, участвующей в синтезе белка,
это транспортная РНК (т-РНК). Молекулы т-РНК относительно небольшие (по
сравнению с рибосомной или матричной РНК). Для каждой из 20 аминокислот
существует своя транспортная РНК. Все т-РНК имеют общую вторичную структуру. За
счет спаривания комплементарных участков молекулы т-РНК образуется три
"стебля" с петлями на концах и один "стебель", образованный
5'- и 3'-концами молекулы т-РНК (иногда образуется еще дополнительная пятая
петля). Изображение этой структуры похоже на крест или клеверный лист.
"Голова" на этом листе представлена антикодонной петлей, здесь
находится антикодон - те три нуклеотида, которые комплементарно взаимодействуют
с кодоном в м-РНК. Противоположный антикодонной петле стебель, образованный
концами молекулы, называется акцепторным стеблем - сюда присоединяется
соответствующая аминокислота. Распознают подходящие друг другу т-РНК и
аминокислоты специальные ферменты, называемые аминоацил-т-РНК синтетазами. Для
каждой аминокислоты есть своя аминоацил-т-РНК-синтетаза.
На рибосоме находится матричная РНК (м-РНК). С
кодоном (тремя нуклеотидами) м-РНК комплементарно связан антикодон транспортной
РНК, на которой висит остаток аминокислоты. На рисунке видна такая структура
(т-РНК вместе с аминокислотой, которая называется аминоцил-т-РНК).
Процесс трансляции, также как и процесс
транскрипции, связан с перемещением вдоль молекулы нуклеиновой кислоты, разница
в том, что рибосома шагает на три нуклеотида, в то время как РНК-полимераза -
на один.
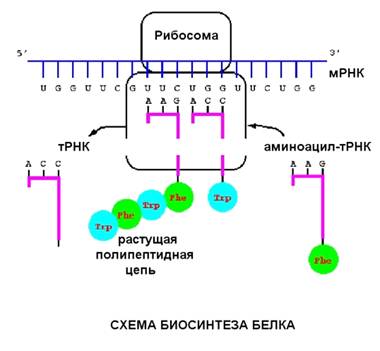
Аминоцил-т-РНК входит в рибосому, комплементарно
связываясь с кодоном м-РНК, затем происходит реакция при которой аминокислотные
остатки связываются друг с другом, а т-РНК удаляется.
"Словарь" для перевода с языка
нуклеотидов на язык аминокислот называется генетическим кодом. Аминокислот -
20, нуклеотидов - 4, число комбинаций из 4 по 2 = 16, а аминокислот 20, поэтому
кодировка не двух, а трехбуквенная, каждая тройка называется кодоном. Каждая
аминокислота кодируется тремя нуклеотидами в м-РНК (которая, в свою очередь,
кодируется ДНК).
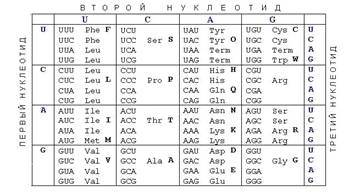
В таблице на рисунке боковые столбцы кодируют
левую и правую букву кодона, верхняя строка - среднюю. Например, кодон AUG
кодирует аминокислоту метионин. Число комбинаций из 4 по 3 = 64, то есть
некоторые аминокислоты кодируются несколькими кодонами. Три кодона не кодируют
никакую аминокислоту, они называются терминирующими. Когда они попадаются в
м-РНК, рибосома прекращает свою работу и готовая полипептидная цепь
выбрасывается наружу.
Генетический код универсален. Он один и тот же у
всех живых организмов. Есть небольшие отличия в генетическом коде митохондрий и
коде археобактерий.
Митоз и Мейоз. При подготовке к делению клетки
происходит репликация ДНК, каждая хромосома, состоящая из одной молекулы ДНК,
реплицируется, и возникают две новые хромосомы - молекулы ДНК, идентичные
родительской. Удвоенные хромосомы остаются связанными в районе центромеры - там
наблюдается недорепликация ДНК. После репликации ДНК конденсируется, хромосомы
приобретают более компактную укладку, и в таком состоянии их можно увидеть в
световом микроскопе. Между делениями эти хромосомы не столь конденсированы и в
большей степени расплетены. Две сестринские хроматиды, соединенные в
центромерном районе недореплицированным участком, приобретают вид буквы Х.
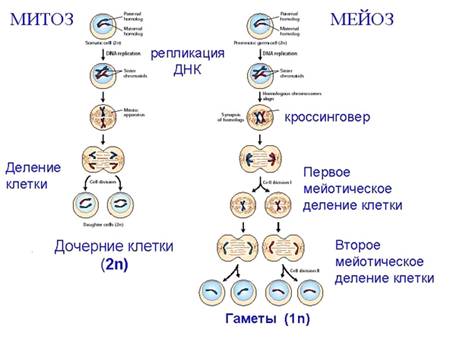
Процесс деления, при котором исходно диплоидная
клетка дает две дочерние, также диплоидные, клетки, называется митозом.
Имеющиеся в клетке хромосомы удваиваются, выстраиваются в клетке, образуя
митотическую пластинку, к ним прикреплены нити веретена деления, которые
растягиваются к полюсам клетки и клетка делится на две дочерние, каждая из
которых содержит копию исходного набора хромосом.
При образовании гамет, т.е. половых клеток -
сперматозоидов и яйцеклеток - происходит деление клетки, называемое мейозом.
Исходная клетка имеет диплоидный набор хромосом - 2n. Происходит репликация ДНК
каждой хромосомы, как и при митозе. Но, если при митозе в каждой хромосоме
хроматиды просто расходятся, то при мейозе перед первой стадией деления -
редукционной стадией - хромосома, состоящая из двух хроматид, коньюгирует, то
есть приходит в тесный контакт с другой, гомологичной ей хромосомой, также
состоящей из двух хроматид, и происходит кроссинговер - обмен гомологичными
участками хромосом. Затем гомологичные хромосомы - каждая из них реплицирована,
то есть состоит из двух хроматид - с обмененными между собой участками
расходятся и образуются клетки с гаплоидным набором хромосом, но генный
(алллельный) состав этих хромосом уже отличается от материнского. В этих
клетках произошла рекомбинация генов. Второе деление мейоза происходит без
нового синтеза ДНК, и является чисто митотическим.
В результате мейоза возникает четыре гаплоидных
клетки - гаметы, каждая из которых уже несет перетасованный между гомологами,
обновленный набор хромосом. Это так называемая рекомбинационная изменчивость -
во-первых, происходит случайное расхождение материнских и отцовских гомологов,
во-вторых, за счет кроссинговера хромосомы обмениваются гомологичными
участками. В результате этих двух процессов в гаметах возникают новые
комбинации аллелей генов. Третий этап рекомбинационной изменчивости происходит
во время оплодотворения. Случайная отцовская гамета сливается со случайной
материнской, в результате возникает зигота, прошедшая три стадии
рекомбинационной изменчивости.
1.Бабушкин
А.Н. Современные концепции естествознания: Курс лекций. 4-е изд., стер. - СПб:
Издательство «Лань», М.: ООО Издательство «Омега-Л», 2008. - 224с.
.Горбачев
В.В. Концепции современного естествознания: Учеб.пособие для студентов вузов. -
М.: ООО «Издательский дом «ОНИКС 21 век»: ООО «Издательство «Мир и
образование», 2007. - 592с.
.Грушевицкая
Т.Г., Садохин А.П. Концепции современного естествознания: Учеб. пособие для
вузов. - М.: ЮНИТИ-ДАНА, 2007. - 670с.
.Дубнищева
Т.Я. Концепции современного естествознания. - Основной курс в вопросах и
ответах: Учебное пособие. 2-е изд. испр. и доп. - Новосибирск: Сиб-е ун-е
изд-во, 2008. - 592с.
.Дубнищева
Т.Я. Концепции современного естествознания: Учеб. пособие. 7-е изд, испр. и
доп. - М.: Издательский Центр «Академия», 2009. - 608с.
.Концепции
современного естествознания: Учебник для вузов /Под ред В.Н. Лавриненко, В.П
Ратникова. - 3-е изд., перераб. и доп.- М.: ЮНИТИ-ДАНА, 2009. - 317с.
.Рузавин
Г.И. Концепции современного естествознания: Учебник для вузов. - М.: ЮНИТИ,
2007. - 287с.
.Свиридов
В.В. Концепции современного естествознания: Учебное пособие.- 2-изд. - СПб.:
Питер, 2009. - 349с.