В-лимфоциты. Рецепторы и маркеры. Участие в иммунном ответе
МИНИСТЕРСТВО
ОБРАЗОВАНИЯ И НАУКИ УКРАИНЫ
ТАВРИЧЕСКИЙ
НАЦИОНАЛЬНЫЙ УНИВЕРСИТЕТ
ИМ.
В.И. ВЕРНАДСКОГО
БИОЛОГИЧЕСКИЙ
ФАКУЛЬТЕТ
КАФЕДРА
БИОХИМИИ
Учебная
дисциплина: ИММУНОЛОГИЯ
Тема:
«В-лимфоциты.Рецепторы и маркеры. Участие в иммунном ответе»
Реферат подготовила:
Студент: Левченко
Наталья Николаевна
Проверила:
Залевская Ирина
Николаевна
Симферополь,
2013
Содержание
Введение
Глава 1. Общая характеристика B-лимфоцитов
Глава 2. Рецепторы и маркеры В-лимфоцитов
.1 Антигенраспознающие рецепторы B-клеток: общая характеристика
.2 Маркеры В-лимфоцитов
Глава 3. Субпопуляции В-лимфоцитов
Список использованной литературы
лимфоцит рецептор антиген
Клетки иммунной системы, на которые
возложены ключевые функции по осуществлению приобретённого иммунитета,
относятся к лимфоцитам, которые являются подтипом лейкоцитов.
Лимфоциты - это единственные клетки
организма, способные специфически распознавать собственные и чужеродные
антигены и отвечать активацией на контакт с конкретным антигеном. При весьма сходной
морфологии малые лимфоциты делятся на две популяции, имеющие различные функции
и продуцирующие разные белки.
Одна из популяций получила название
В-лимфоциты, от названия органа «бурса Фабрициуса», где было впервые обнаружено
созревание этих клеток у птиц. У человека В-лимфоциты созревают в красном
костном мозге.
В-лимфоциты распознают антигены
специфическими рецепторами иммуноглобулиновой природы, которые по мере
созревания В-лимфоцитов появляются на их мембранах. Взаимодействие антигена с
такими рецепторами является сигналом активации В-лимфоцитов и их
дифференцировки в плазматические клетки, продуцирующие и секретирующие
специфические для данного антигена антитела - иммуноглобулины.
Главной функцией В-лимфоцитов также
является специфическое распознавание антигена, которое ведет к их активации,
пролиферации и дифференцировке в плазматические клетки - продуценты
специфических антител - иммуноглобулинов, т. е. к гуморальному иммунному
ответу. Чаще всего В-лимфоциты для развития гуморального иммунного ответа
нуждаются в помощи Т-лимфоцитов в виде продукции активирующих цитокинов.
Глава 1. Общая характеристика
B-лимфоцитов
Специфическое иммунологическое
распознавание патогенных организмов - это всецело функция лимфоцитов, поэтому
именно они инициируют реакции приобретенного иммунитета. Все лимфоциты
происходят из стволовых клеток костного мозга, но Т-лимфоциты затем развиваются
в тимусе, тогда как В-лимфоциты продолжают свое развитие в красном костном
мозге (у взрослых особей млекопитающих). Термин B-лимфоциты образован по первой
букве английского названия органов, в которых эти клетки формируются: bursa of
Fabricius (фабрициева сумка у птиц) и bone marrow (костный мозг у
млекопитающих).
Сумка Фабрициуса - один из
центральных органов иммуногенеза птиц, располагающийся в области клоаки и
контролирующий гуморальный иммунный ответ. Удаление этого органа приводит к
отмене синтеза антител. Аналогом фабрициевой сумки у млекопитающих является
красный костный мозг.
Главная функция B-лимфоцитов (а
вернее плазматических клеток, в которые они дифференцируются) - это выработка
антител. Воздействие антигена стимулирует образование клона B-лимфоцитов,
специфического к данному антигену. Затем происходит дифференцировка
новообразованных B-лимфоцитов в плазматические клетки, вырабатывающие антитела.
Эти процессы проходят в лимфоидных органах, регионарных к месту попадания в
организм чужеродного антигена. лимфоциты составляют около 15-18% всех
лимфоцитов, находящихся в периферической крови. После распознавания специфического
антигена эти клетки размножаются и дифференцируются, трансформируясь в
плазматические клетки. Плазматические клетки вырабатывают большое количество
антител (иммуноглобулины Ig), которые являются собственными рецепторами
B-лимфоцитов в растворенном виде.лимфоциты продуцируют и секретируют в кровоток
молекулы антител, являющиеся измененными формами антигенраспознающих рецепторов
этих лимфоцитов. Возникновение антител в крови после появления любого
чужеродного белка - антигена - независимо от того, вреден он или безвреден для
организма, и представляет собой иммунный ответ. Появление антител не просто
защитная реакция организма против инфекционных заболеваний, но явление, имеющее
широкое биологическое значение: это общий механизм распознавания "чужого".
Например, иммунная реакция распознает как чужой и постарается удалить из
организма любой аномальный и, следовательно, потенциально опасный вариант
клетки, в которой в результате мутации в хромосомной ДНК образуется мутантная
белковая молекула. лимфоциты (B-клетки) млекопитающих дифференцируются сначала
в печени плода, а после рождения - красном костном мозге. В цитоплазме
покоящихся B-клеток отсутствуют гранулы, но имеются расеянные рибосомы и
канальцы шероховатого эндоплазматического ретикулума. Каждая B-клетка
генетически запрограммирована на синтез молекул иммуноглобулина, встроенных в
цитоплазматическую мембрану. Иммуноглобулины функционируют как
антигенраспознающие рецепторы, специфичные к определенному антигену. На
поверхности каждого лимфоцита экспрессируется около ста тысяч молекул
рецепторов. Встретив и распознав антиген, соответствующий структуре
антигенраспознающего рецептора B-клетки размножаются и дифференцируются в
плазматические клетки, которые образуют и выделяют в растворимой форме большие
количества таких рецепторных молекул - антител. Антитела представляют собой
крупные гликопротеины и содержатся в крови и тканевой жидкости. Благодаря своей
идентичности исходным рецепторным молекулам они взаимодействуют с тем
антигеном, который первоначально активировал B-клетки, проявляя таким образом
строгую специфичность.
После связывания антигена с
рецепторами B-клетки клетка активируется. Активация B-клеток состоит из двух
фаз: пролиферации и дифференцировки; все процессы индуцируются контактом с
антигеном и T- хелперами. В результате пролиферации увеличивается число клеток,
способных реагировать с введенным в организм антигеном. Значение пролиферации
велико, поскольку в неиммунизированном организме очень мало B-клеток,
специфичных для определенных антигенов. Часть клеток, пролиферирующих под
действием антигена, созревает и дифференцируется последовательно в
антителообразующие клетки нескольких морфологических типов, в том числе и
плазматические клетки. Промежуточные стадии дифференцировки B-клеток отмечены меняющейся
экспрессией разнообразных белков клеточной поверхности, необходимых для
взаимодействия B-клеток с другими клетками.
Каждый лимфоцит, относящийся к
B-лимфоцитам и дифференцирующийся в костном мозге, запрограммирован на
образование антител только одной специфичности.
Молекулы антител не синтезируются
никакими другими клетками организма, и все их многообразие обусловлено
образованием нескольких миллионов клонов B-клеток. Они (молекулы антител)
экспрессируются на поверхностной мембране лимфоцита и функционируют как
рецепторы. При этом на поверхности каждого лимфоцита экспрессируется около ста
тысяч молекул антител. Кроме того, B-лимфоциты секретируют в кровоток
продуцированные ими молекулы антител, являющиеся измененными формами
поверхностных рецепторов этих лимфоцитов.
Антитела формируются до появления
антигена, и антиген сам отбирает для себя антитела. Как только антиген
проникает в организм человека, он встречается буквально с войском лимфоцитов,
несущих различные антитела, причем у каждого есть свой индивидуальный
распознающий участок. Антиген соединяется только с теми рецепторами, которые в
точности ему соответствуют. Лимфоциты, связавшие антиген, получают пусковой
сигнал и дифференцируются в плазматические клетки, продуцирующие антитела.
Поскольку лимфоцит запрограммирован на синтез антител только одной
специфичности, антитела, секретируемые плазматической клеткой, будут идентичны
своему оригиналу, т.е. поверхностному рецептору лимфоцита и, следовательно,
будут хорошо связываться с антигеном. Так антиген сам отбирает антитела,
распознающие его с высокой эффективностью.
Весь путь развития В-лимфоцитов от
стволовой кроветворной клетки до плазмоцита включает несколько этапов, каждый
из которых характеризуется своим клеточным типом.
Всего выделено 7 таких типов:
) стволовая кроветворная
(гемопоэтическая) клетка - общий предшественник для всех ростков
дифференцировки лимфомиелопоэза;
) общий лимфоидный предшественник
B-клеток и T-клеток для B- и T-клеточного пути развития - наиболее ранняя
лимфоидная клетка, для которой еще не определилось одно из двух направлений
развития;
А) ранняя про-В-клетка - ближайший
потомок предыдущего клеточного типа и предшественник последующих, продвинутых в
дифференцировке клеточных типов (приставка "про" от англ.
progenitor);
Б) поздняя про-В-клетка;
) незрелая В-клетка - завершающая
костномозговое развитие клеточная форма, которая активно экспрессирует поверхностный
иммуноглобулин и находится в стадии отбора на способность взаимодействовать с
собственными антигенами;
) зрелая В-клетка - клеточный тип
периферии, способный взаимодействовать только с чужеродными антигенами;
) плазматическая клетка (плазмоцит)
- эффекторная, антителопродуцирующая клеточная форма, которая образуется из
зрелой В-клетки после ее контакта с антигеном.
Глава 2. Рецепторы и маркеры
В-лимфоцитов
.1 Антигенраспознающие рецепторы
B-клеток: общая характеристика
Антигенраспознающие рецепторы
В-лимфоцитов представляют собой молекулы иммуноглобулинов. Циркулирующие
антитела структурно подобны основной части B-клеточных рецепторов, но лишены их
трансмембранных и цитоплазматических сегментов. Основными классами
мембранно-связанных иммуноглобулинов (mIg), находящихся на поверхности зрелых,
нестимулированных В-лимфоцитов, являются IgM и IgD. На одной В-клетке могут
одновременно присутствовать оба типа молекул, причем они имеют одинаковую
специфичность, и, возможно, что эти антигенные рецепторы могут
взаимодействовать между собой, осуществляя контроль за активацией лимфоцитов и
супрессией лимфоцитов.
Рецептором B-лимфоцитов, узнающим
антиген, является IgM. Мембраносвязанный IgM (mIgМ), как правило, представляет
собой мономерный иммуноглобулин, т.е. отдельную единицу из четырех
полипептидных цепей. Эта молекула имеет гидрофобную последовательность,
расположенную на C-концевом участке тяжелой цепи и предназначенную для фиксации
молекулы на клеточной мембране. Число молекул рецептора достигает 10 - 100тыс.
на клетку.М кодируется тем же набором генов, что и сывороточные аналоги.
Единственным их структурным отличием является дополнительный фрагмент на
С-конце молекулы, играющий роль мембранного якоря.
В онтогенезе первые мембранные формы
IgM появляются на заключительном этапе дифференцировки В-клеток в костном
мозге. Зрелые В- клетки экспрессируют достаточно большое количество этого
белка: 2*105 молекул на одну клетку. Такой рецепторный иммуноглобулин
взаимодействует либо с белковым или корпускулярным антигеном (рис. 3.1), либо с
антигенными детерминантами на поверхности антигенпрезентирующих клеток
(макрофагов, дендритных клеток и др.).
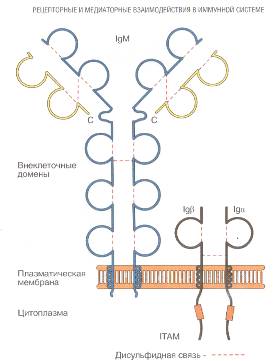
Рис 1. Структура
В-клеточного рецептора
Антигенраспознающие
рецепторы B-клеток были обнаружены достаточно легко, в основном, с помощью
антииммуноглобулиновых антител, меченных либо радиоактивными химическими
элементами, либо флюоресцеином. При связывании антигена с соответствующим
рецептором и под влиянием цитокинов, вырабатываемых моноцитами, макрофагами и
Т-лимфоцитами, происходит активация В-лимфоцитов, которые начинают делиться и
дифференцироваться в плазматические клетки. Часть активированных В-лимфоцитов
превращаются в клетки памяти, которые обеспечивают более быстрый и эффективный
иммунный ответ при повторном контакте с антигеном. С основной частью рецептора
непосредственно связаны дополнителные компоненты (Ig-альфа (СВ79a) и Ig-бета
(CD79b)), соединяющие его с путями внутриклеточной передачи сигнала.
Цитоплазматический
участок у mIg невелик и не пригоден для взаимодействия с С-белками или
тирозинкиназами. Роль CD3 в случае mIgМ, по-видимому, играет ассоциированный с
mIgМ гетеродимер, состоящий из двух соединенных дисульфидной связью
гликопротеинов с молекулярными массами 32-34кДа (IgM-альфа) и 37-39кДа
(IgM-бета, IgM-гамма). Цепи бета и гамма являются продуктами одного гена и
получаются в результате альтернативного сплайсинга. Обе цепи являются
представителями суперсемейства иммуноглобулинов и содержат во внеклеточной
части по одному домену. Цитоплазматические участки этих полипептидов имеют
консервативную последовательность, включающую шесть расположенных определенным
образом друг относительно друга аминокислот. Такая же последовательность
обнаруживается в гамма-, дельта- и дзета-цепях CD3, что предполагает сходные
функции CD3 и IgM-альфа-IgM-бета. Цитоплазматические фрагменты содержат
потенциальные сайты фосфорилирования.
.2 Маркеры В-лимфоцитов
На периферии (вне
костного мозга) В-лимфоциты приобретают характерные для них поверхностно-клеточные
маркёры.
Основные маркёры
В-лимфоцитов - мембранные Ig при этом клетки одного клона (быстро
формирующегося в результате серии последовательных делений потом-ства одной
В-клетки) экспрессируют молекулы Ig, специфически связывающие только один
эпитоп Аг. Такие клетки синтезируют моноклональные AT, способные распознавать и
связывать только один Аг. Аг-связывающий участок мембранного Ig В-лимфоцита
играет роль клеточного Аг-распознающего рецептора. Помимо мембранных Ig,
В-лимфоцит несёт другие маркёры; рецепторы Fc-фрагмента Ig, CD10 {на незрелых
В-клетках), CD19, CD20, CD21, CD22, CD23 (вероятно, участвуют в клеточной
активации), рецепторы к С3b и C3d, молекулы МНС классов I и И.
Мембранный
иммуноглобулин - специфический маркер В-клеток, поскольку он экспрессирован на
всех зрелых В-лимфоцитах и отсутствует на других клетках. Преобладающим классом
мембранных иммуноглобулинов на нативных (не контактировавших с антигеном)
В-клетках является IgM. Он присутствует на поверхности всех нативных В-лимфоцитов,
начиная со стадии незрелых В-клеток (см. раздел 3.3.1.2) (табл. 3.2). На зрелых
нативных
В-клетках наряду с IgM
присутствует IgD. Число молекул иммуноглобулинов на поверхности наивной
В-клетки составляет около 150 000. В процессе иммунного ответа происходит
переключение классов иммуноглобулинов на IgG, IgA и IgE. В-клетки крови и
вторичных лимфоидных органов несут на своей поверхности преимущественно IgG, а
В-клетки слизистых оболочек - IgA.
В состав BCR помимо
иммуноглобулина входит еще несколько молекул. Две из них - СD79a и СD79b -
составляют интегральную чаcть BCR, еще три - CD19, CD21 и CD81 - функционально
ассоциированы с ним и формируют физическую связь с BCR только при активации
клетки. Варианты молекул CD79 - a и b - называют еще Igα
и Igβ. При помощи нековалентных связей они формируют гетеродимеры,
связанные с мембранным иммуноглобулином. Эти молекулы имеют сходные размеры и
молекулярную массу (около 40 кДа). Участие Igα и
Igβ в передаче сигнала основано на связи их цитоплазматической части с
внутриклеточными тирозинкиназами.
Молекулу CD19 относят к
суперсемейству иммуноглобулинов. CD19 играет важную сигнальную функцию,
поскольку эта молекула связана с киназой PI3K. CD21 - рецептор для компонентов
комплемента (CR2), участвующий в усилении антигенного сигнала, а также в
регуляции активности В-лимфоцитов. СD81 относят к тетраспанинам (4 раза
пронизывают мембрану); функция этой молекулы точно не определена.
С цитоплазматическими
участками мембранного иммуноглобулина связана тирозинкиназа Fyn, а с молекулами
CD79, CD19 и CD81 - тирозин-киназы Blk, Lyn, Lck, а также Syk, участвующие в
передаче активационного сигнала. Кроме того, около цитоплазматической части
молекулы CD19 располагается липидная киназа PI3K. Такое обилие сигнальных
ферментов, связанных с компонентами BCR, обеспечивает запуск и передачу
активационных сигналов при связывании антигена.
Все В-лимфоциты обладают
рядом общих свойств: они продуцируют антитела и иммуноглобулин (Ig),
экспрессируют антиген-распознающий Ig-рецептор (B-Cell Receptor - BCR) и
поверхностные маркеры CD 19 и CD45 (В220). Вместе с тем можно выделить
несколько субпопуляций В-клеток, различающихся по происхождению,
дифференцировке, фенотипу и функциональным свойствам.
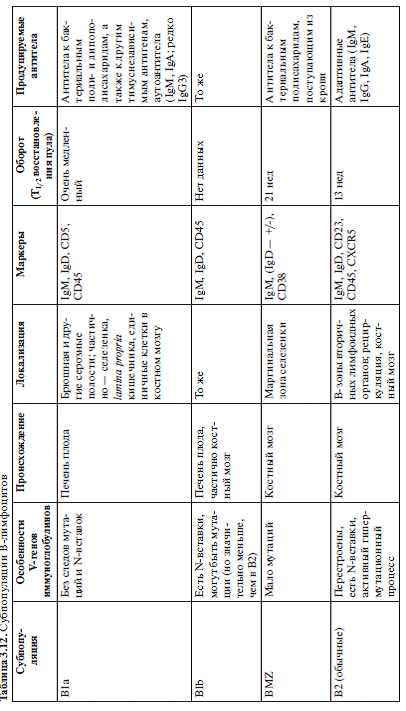
Выделяют 3 основные
субпопуляции В-клеток (табл.1). Одна из них рассмотрена выше - В2-клетки
(иногда их называют обычными В-клетками), локализующиеся преимущественно в
селезенке, костном мозгу, лимфоузлах, пейеровых бляшках и отдельных фолликулах
лимфоидной ткани кишечника. Гистологическая единица, являющаяся местом
сосредоточения В2-клеток - лимфоидный фолликул. Эти клетки составляют
подавляющеебольшинство циркулирующих В-лимфоцитов и играют основную роль в
гуморальном иммунном ответе. Две другие субпопуляции - В1- и В-клетки
маргинальной зоны (MZВ-клетки). Большинство данных о различных субпопуляциях
В-лимфоцитов получено на мышах. Сведения о субпопуляцияхВ-клеток человека
крайне скудны.
В1-клетки локализуются
преимущественно в серозных полостях -
брюшной и плевральной.
Небольшое количество В1-лимфоцитов, преимущественно клетки, секретирующие
антитела, выявляют в селезенке, где на их долю приходится 1-5% от числа
В-клеток. Некоторые В1-клетки мигрируют (через сальник) в слизистую оболочку
кишечника и брыжеечные лимфоузлы (до 50% IgA-продуцентов в лимфоидной ткани
кишечника - В1-клетки). В лимфатических узлах у мыши они отсутствуют. Выделяют2
субпопуляции В1-клеток. Основной дифференциальный признак при этом - экспрессия
мембранной молекулы СD5 (известной как один из маркеров Т-клеток). В1а-клетки
одновременно несут на поверхности молекулы IgM и CD5. CD5 отсутствует на всех
остальных В-лимфоцитах, в том числе на В1b-клетках, в остальном очень схожих с
В1а-клетками. Для В1-клеток характерен ≪активированный
фенотип≫, что проявляется в экспрессии на их поверхности костимулирующих
молекул СD80 и СD86. Это свойство обеспечивает способность В1-лимфоцитов
выполнять функции АПК.а- и B1b-клетки экспрессируют BCR, содержащий мембранную
форму IgM. Известны исключения: описано переключение IgM на IgA в lamina
propria кишечника. Перестроенные V-гены мембранного IgM В1а-клеток не содержат
N-вставок (т.е. в их перестройке не участвует фермент TdT). Разнообразие
V-генов В1-клеток существенно ниже, чем у В2-клеток. Это связано с различиями в
условиях развития: В1а-клетки в онтогенезе появляются раньше других
субпопуляций - еще до рождения. Они развиваются в печени плода при участии IL-5
и IL-10 из клеток-предшественников, отличных от таковых у обычных В-клеток. Еще
в эмбриональном периоде В1-клетки мигрируют в серозные полости, где они
существуют в течение всей жизни организма.
В1-клетки способны к
самоподдержанию путем очень медленной пролиферации, восполняющей убыль клеток,
погибающих по механизму апоптоза.
В1b-клетки также
развиваются в печени эмбрионов, а после рождения - в костном мозгу из других
клеток-предшественников. B1b-лимфоциты расселяются на периферии несколько позже
В1а-клеток - непосредственно перед рождением и сразу после него. При
перестройке V-генов в B1b-клетках формируется некоторое количество N-вставок.
B1b-клетки также мигрируют в серозные полости и сохраняются там путем
самоподдержания.
Обе разновидности
В1-клеток могут дифференцироваться в антителообразующие клетки без стимуляции
антигеном. При этом они секретируют преимущественно IgM-антитела (в кишечнике -
также IgA). Большинство этих антител специфично к собственным белкам организма
(ДНК, гистонам, коллагену, компонентам цитоскелета, антигенам групп крови и
т.д.); многие из них полиспецифичны, т.е. способны взаимодействовать с
несколькими антигенами, в том числе аутологичными. Эти антитела имеют низкое
сродство (аффинность) к антигенам, включая аутоантигены, и не способны вызвать
повреждение тканей. Примерно половина сывороточного IgM секретируется
В1-клетками. Естественные антитела, продуцируемые В1а-лимфоцитами, часто
специфичны к микробным антигенам и опсонизируют патогены, играя важную роль в
реакциях врожденного иммунитета.
Эти клетки могут
принимать участие в адаптивном иммунном ответе, что в большей степени свойственно
В1b-клеткам. Ответ В1-клеток преимущественно тимуснезависимый. В1-клетки
постоянно циркулируют между селезенкой и брюшной полостью, но не поступают в
фолликулы, поскольку не экспрессируют CXCR5 - рецептор хемокина BLC (CXCL13). С
этим связано то обстоятельство, что процессы ≪усовершенствования≫
гуморального иммунного ответа в виде переключения изотипов и повышения сродства
к антигенам, не затрагивают или минимально затрагивают В1-клетки.
Еще одна разновидность
В-лимфоцитов -B-клетки маргинальной зоны (MZB). Они локализуются почти
исключительно в маргинальной зоне селезенки, отделяющей белую пульпу от
красной. Фенотипически эти клетки более сходны с В2-, чем с В1-клетками. Они
происходят от тех же костно-мозговых клеток-предшественников. Основной мембранный
иммуноглобулин MZB-клеток -IgM, экспрессируемый сильнее, чем на В2-клетках. В
то же время IgD присутствует на мембране в очень малом количестве. Эти клетки
сходны по своему фенотипу с активированными В-лимфоцитами. На них присутcтвуют
молекулы CD69, CD25, CD38, в малом количестве CD23. Обращает на себя внимание
наличие молекулы CD1d, участвующей в презентации липидных антигенов.
Отделение линии
MZB-клеток от общей линии В2-клеток происходит на переходной стадии
транзиторных клеток (Т3), когда будущие MZB-клетки ослабляют экспрессию не IgM
(как В2-клетки), а IgD и утрачивают молекулу CD23. На MZB-лимфоцитах не
экспрессируется хемокиновый рецептор CXCR5, позволяющий клеткам мигрировать в
фолликулы. Ключевой фактор дифференцировки MZB-клеток - Notсh-2. Под влиянием
сфингозин-1-фосфата и при участии молекул адгезии LFA-1 и VLA-4 они мигрируют в
маргинальные зоны селезенки. MZB-клетки не участвуют в рециркуляции, но
осуществляют ≪челночные≫ миграции до лимфоидных фолликулов и обратно, получая информацию
об антигенах, поступающих в селезенку с кровью. Срок жизни MZB-лимфоцитов
сопоставим со сроком жизни организма. Снижение численности MZB-клеток,
вызываемое повреждающими факторами, достаточно быстро устраняется.клетки
участвуют в гуморальном иммунном ответе на возбудители, поступающие в кровоток.
Они осуществляют тимуснезависимый иммунный ответ на инкапсулированные патогены.
Благодаря сильной экспрессии молекул MHC-II и костимулирующих молекул
MZB-клетки обладают выраженной способностью к взаимодействию с Т-хелперами,
однако их участие в тимусзависимом иммунном ответе изучено плохо. При ответе на
антигены MZB-клетки дифференцируются в короткоживущие антителообразующие
клетки. V-гены MZB-клеток редко затрагиваются мутациями, что характерно для
развития плазматических клеток вне зародышевых центров. В этих клетках не
происходит переключения классов иммуноглобулинов и даже MZB-клетки памяти несут
на своей поверхности IgM, а не IgG. IgM+ клетки памяти преобладают в
маргинальной зоне селезенки человека.
Список использованной
литературы
1. А. Ройт, Дж. Брюсстофф, Д. Мейл. Иммунология- М.: Мир,
2000
. Лебедев К.А. - Иммунология в клинической практике, 1996
. Иммунология(в 3 т.) / Под. ред. У. Пола.- М.:Мир, 1988
. Ярилин А.А - Иммунология, 2010
. Хаитов P.M., Игнатьева Г.А., Сидорович И.Г., Иммунология:
Учебник. - М.: Медицина, 2000. 432 с: ил. (Учеб. лит. Для студ. медвузов).
. http://medbiol.ru/medbiol/immunology/imm-gal/00005492.htm
. http://vmede.org
. http://immuninfo.ru/immunologiya