Функциональная организация предстимульного модально-специфического внимания при произвольной преднастройке и серийном научении
Содержание
Введение
Глава 1. Влияние организация
предстимульного внимания на эффективность обработки значимой информации. Анализ
данных психологических и нейрофизиологических исследований
.1
Экспериментально-психологические исследования предстимульного внимания
.2 Нейрофизиологические
исследования избирательного предстимульного внимания
Эмпирическое исследование
Глава 2. Методические аспекты
исследования
.1 Предварительный
эксперимент. Оптимизация параметров стимулов и уравнивание сложности задач на
различение зрительных и слуховых сигналов
.2 Основной эксперимент
Глава 3. Результаты
.1 Анализ изменений
эффективности решения сенсорных задач в ходе серийного научения
.2 Анализ влияния организации
предстимульного периода на эффективность решения сенсорных задач
.3 Анализ мозговой
организации предстимульного модально-специфического внимания при произвольной
преднастройке и серийном научении
Глава 4. Обсуждение
Выводы:
Список литературы
Введение
Внимание - это одна из наиболее интенсивно изучаемых в современной
психологии и психофизиологии психических функций. Несмотря на это, единого
представления о внимания в психологии не существует. Наиболее распространенным
можно считать общее определение, согласно которому внимание есть концентрация
сознания на определенном реальном или идеальном объекте [James, 1890; Рубинштейн, 1998]. В
когнитивной психологии внимание определяется как процесс избирательной
концентрации на одном аспекте внешней среды при игнорировании других аспектов,
а также как распределение (allocation) энергетических ресурсов, направляемых на обработку информации
[Anderson, 2004]. Внимание исследуется на когнитивном, нейросистемном,
клеточном, синаптическом и генетическом уровнях [Posner, 2004]. Многогранность и комплексный характер
феномена внимания заставляют исследователей использовать различные эпитеты для
его описания: «пассивное», «активное», «произвольное», «непроизвольное»,
«непосредственное», «опосредованное», «избирательное», «бдительное» и др.
Предметом настоящего исследования является предвосхищающее или
антиципационное внимание, внимание к стимулам в будущем [Наатанен, 1998]. Один
из важнейших вопросов, возникающих при изучении внимания - это вопрос о том,
можем ли мы говорить о направленном избирательном внимании еще до того как
сенсорные сигналы доступны для восприятия? Исходя из представлений об активном
характере психических функций [Выготский, 1960] и их зависимости от внутреннего
состояния и целей поведения субъекта, представляется не только оправданным, но
и необходимым рассматривать процессы, происходящие в период, предшествующий
сенсорной обработке внешней информации как важный этап функционирования
механизмов селективного внимания. Согласно концепции П.Я. Гальперина [1958]
внимание есть свернутое действие контроля, а контроль при целенаправленном
поведении предполагает предвосхищение будущих событий и подготовку к ним.
Существуют, по крайней мере, два варианта условий, при которых субъект
может предвидеть появление значимых сигналов: если им предшествует
предупреждающий сигнал - подсказка (избирательное направленное внимание), или
если определенные события закономерно повторяются во времени (имплицитное
предвосхищение в ходе серийного научения). Эти два условия предстимульного
внимания могут возникать в разных жизненных ситуациях и по-разному сказываться
на процессах обработки информации. Представляло интерес проведение
сравнительного экспериментального исследования избирательного направленного
внимания, вызванного инструкцией, и имплицитного предвосхищения для получения новых
данных о влиянии структуры деятельности и организации информационного
пространства на эффективность решения когнитивных задач.
Несмотря на большое количество исследований внимания в психологии и
нейронауках вопрос о возможности избирательной подготовки к анализу значимой
информации остается дискуссионным. В связи с тем, что предстимульный период
является «закрытым» для психологического наблюдения и экспериментального
исследования, об избирательном влиянии антиципационного внимания на обработку
значимой информации можно судить лишь косвенно по изменению скорости и точности
решения когнитивной задачи. Для изучения функциональной организации
предстимульного внимания и оценки его избирательности представляется актуальным
проведение исследования, сочетающего анализ параметров когнитивной деятельности
с психофизиологическими методами неинвазивного изучения мозговой активности в
период предшествующий восприятию целевого сигнала.
Методы: В настоявшей работе для изучения функциональной организации
антиципационного внимания был использован междисциплинарный подход,
предполагающий оценку скорости и точности обработки сенсорной информации и
электроэнцефалографический (ЭЭГ) анализ функционального взаимодействия
различных зон коры головного мозга в период, предшествующий появлению целевых
сенсорных сигналов.
Цель исследования состояла в анализе влияния двух условий антиципационного
(предстимульного) внимания - произвольной преднастройки и серийного научения -
на мозговую организацию процессов подготовки к анализу значимой сенсорной
информации и эффективность распознавания сенсорных сигналов разных
модальностей.
Основная гипотеза исследования: произвольная преднастройка и
серийное научение оказывают различное влияние на функциональную организацию
антиципационного внимания в период подготовки к анализу значимой сенсорной
информации и эффективность решения задач на различение зрительных и слуховых
сигналов.
Экспериментальная проверка основной гипотезы предполагала решение
следующих задач:
) оценка успешности и скорости распознавания зрительных и слуховых
сигналов в условиях направленного произвольного внимания, задаваемого
предварительной инструкцией, и в условиях имплицитного предвосхищения
периодически повторяющейся последовательности сенсорных задач;
) электрофизиологический анализ функционального взаимодействия различных
зон коры головного мозга в период подготовки к решению сенсорных задач в
условиях направленного произвольного внимания, задаваемого предварительной
инструкцией, и в условиях имплицитного предвосхищения периодически повторяющейся
последовательности сенсорных задач
) Сопоставление параметров эффективности когнитивной деятельности с
электрофизиологическими параметрами функциональной организации мозга в разных
условиях предстимульного модально-специфического внимания.
Таким образом, объектом исследования являются параметры
деятельности, отражающие эффективность различения слуховых и зрительных
сигналов и параметры суммарной электрической активности мозга -
электроэнцефалограммы (ЭЭГ), зарегистрированной в период, предшествующий
появлению целевых сенсорных стимулов у взрослых испытуемых.
Практическое применение результатов исследования предстимульного внимания
может быть разнообразным. Поскольку основной функцией внимания с точки зрения
современной психологии и нейронаук является контроль и организации поведения,
можно говорить о его ключевой роли в реализации любой деятельности [Posner, 1995; Coull, 1998; Дормашев, Романов, 2002]. Это определяет
возможность применения результатов исследования функциональной организации
внимания в различных прикладных областях. В области образования такое
исследование может способствовать оптимизации способов подачи учебного
материала для его наиболее эффективного усвоения учащимися, а сравнение
эффективности различных способов организации внимания может быть полезно для
индустрии рекламы.
Глава 1. Влияние организация предстимульного внимания на
эффективность обработки значимой информации. Анализ данных психологических и
нейрофизиологических исследований
Прежде чем перейти к анализу экспериментальных исследований
предстимульного внимания рассмотрим вопрос о правомерности использования самого
термина. Как писал Wittgenstein
[1953], многие концепции не могут дать формального определения вниманию, и
сравнивал этот термин с термином «игра», говоря о том, что мы можем назвать
игрой множество разных вещей, исходя из каких-то общих признаков. Аналогично и
многие феномены в экспериментальной психологии и нейронауке рассматриваются как
проявления внимания. Несмотря на отсутствие единой концепции внимания,
существуют некоторые общие моменты в понимании внимания большинством
современных ученых, которые сходятся во мнение о том, что внимание есть
концентрация сознания на определенном реальном или идеальном объекте [James, 1890; Рубинштейн, 1998]. Благодаря интенсивным
исследованиям механизмов внимания в нейрокогнитивных науках, широкое
распространение получило представление о внимании как самостоятельном
психическом процессе, реализуемом специализированными мозговыми системами,
основными функциями которого являются контроль и организация поведения [Coull, 1998; Мачинская, 2003, обзор;
Posner, Fan, 2008].
Стоит отметить, что одним из переломных моментов в истории изучения
внимания можно назвать переход от исключительно пассивной модели понимания
феномена, внимания в качестве «бутылочного горлышка» к активно-деятельностному
пониманию. Внимание перестало рассматриваться исследователями исключительно как
некоторый ограничитель восприятия, а стало частью деятельности, способностью
мобилизировать внутренние физиологические и психологические ресурсы для решения
конкретных задач. В отечественной психологии активный характер внимания
подчеркивался в концепции П.Я. Гальперина [1958], который рассматривал его как
«свернутое» действие контроля. Такой взгляд на феномен внимания раскрыл его
сложный характер и многостороннюю специфику его проявления, задав, таким
образом, множество векторов для его исследования.
Произвольное внимание рассматривалось П.Я. Гальпериным [1958] как синоним
планомерного внимания, представляющего собой контроль за действием, выполняемый
на основе заранее составленного плана, с помощью заранее установленных
критериев и способов их применения. Наличие такого плана, критериев и способов
действия позволяет вести контроль, а вместе с тем и направлять внимание на то,
на что мы хотим его направить, а не на то, что «само бросается в глаза».
Если непроизвольное внимание может оперировать только в настоящем
времени, так как контроль за тем, что «само бросилось в глаза», может быть
инициирован только наличествующими здесь и сейчас стимулами, произвольное
внимание может быть обращено как к «прошлым», так и «будущим» стимулам и
событиям. В последнем случае в когнитивной психологии используется термин
«ожидание», или «антиципирующее внимание» [Наатанен Р., 1998].
Другим возможным вариантом подготовки субъекта к восприятию будущих
событий и организации своей деятельности в соответствии с ними является
предвосхищение появления значимых сигналов при регулярном повторении порядка их
следования - имплицитное предвосхищение в ходе серийного научения. Рассмотрим
экспериментально-психологические данные, свидетельствующие о влиянии двух типов
предстимульного внимания - произвольного и имплицитного - на процессы обработки
значимой информации.
1.1
Экспериментально-психологические исследования предстимульного внимания
В связи с анализом внимания, обращенного в будущее, т.е. предстимульного
внимания, возникают большие проблемы, связанные с терминологией. За этими
проблемами зачастую стоит, с одной стороны, отсутствие единых принятых
большинством исследователей, терминов и их аналогов в российских исследованиях,
а с другой - наличие принципиально отличающихся форм внимания, которые в силу
тех или иных причин описываются одним и тем же словом или словосочетанием. Так,
говоря об антиципационном внимании, чаще всего имеют в виду экспериментальную
парадигму, и связанную с ней психофизиологическую концепцию внимания,
разработанную М. Познером [Posner,
1978]. В его исследованиях «успеха» (benefits) и «неуспеха», как правило, первый стимул не требовал
реакции испытуемого, а только давал некоторую информацию о целевом стимуле
(например, о возможном локусе его проявления в зрительном поле). Если целевой
стимул действительно появлялся в ожидаемом месте, то время реакции было короче,
чем в случае, когда предупреждающий стимул не содержал информации о месте
целевого сигнала. Такое укороченное время реакции рассматривалось как мера
полезного влияния («успеха») специфического ожидания. Однако если целевой
стимул появлялся в неожиданном месте, то время реакции было больше, чем в
случае, когда предупреждающий стимул не содержал информации о месте его
появления. Такое увеличение времени реакции Познер рассматривал как меру
«потерь» («неуспеха»), связанных с неверным ожиданием. Такая парадигма подразумевает
«выбор» только в том смысле, что субъект избирательно готовит себя к реакции на
некоторые ожидаемые (в будущем) события в большей степени, нежели на другие [Kahnemann, Treisman, 1984]. Наибольшее распространение
при исследовании роли предстимульного периода в эффективности внимания получила
экспериментальная парадигма “flanker test”,
разработанная М. Познером в соавт. [Posner, Fan, 2008]. Целевой стимул в данной
модели представлял собой стрелку, направленную влево или вправо. Целевой стимул
появлялся сверху или снизу экрана. Задача испытуемого состоит в нажатие левой
или правой кнопки в соответствии с направлением стрелки. При постановке этого
эксперимента предполагалось, что эффективность деятельности испытуемого будет
зависеть от привлечения внимания к месту появления целевого сигнала. Для
создание условия привлечения внимания к целевому стимулу, на экране
предъявлялись два вида подсказки, указывающие на место появления целевого
стимула (сверху или снизу экрана): точка в соответствующем месте экрана (периферическая
подсказка), непроизвольно привлекающая внимание к месту появления целевого
стимула, и стрелка в центре экрана указывающая вверх или вниз (центральная
подсказка - инструкция), на основании которой испытуемый произвольно
концентрировал внимание. Помимо варьирования условий внимания в данном
эксперименте вводились периферические стимулы - дистракторы в виде таких же
стрелок как целевой стимул, которые располагались по бокам от целевого стимула.
Периферические стимулы могли быть направлены в ту же сторону, что целевой
стимул или в противоположную сторону. Второй вариант периферических стимулов
повышал нагрузку на процессы внимания и усложнял принятие испытуемым
правильного решения. Исследователей в данном случае интересовали изменения
эффективности деятельности в зависимости от условий внимания и наличия
«конфликтующих» периферических стимулов, игнорирование которых требовало
включения дополнительного контроля внимания (управляющего контроля, cognitive control). В исследовании E. Hahn et al [2011] изменения эффективности деятельности, связанные с
появлением предупреждающего стимула-подсказки, также изучались с использованием
экспериментальной парадигмы, разработанной М. Познером. При этом эксперимент
проводился несколько раз на 55 испытуемых с интервалом в среднем 7.4 месяцев
между сессиями. Было установлено, что повышение эффективности деятельности,
выражающееся в уменьшении времени реакции и увеличении количества правильных
ответов, сохранялось в повторных исследованиях. В исследовании 2004 года M.R. Rueda et al влияние предвосхищения, заданного подсказкой на
эффективность деятельности изучалось на детях разного возраста и
рассматривалось в динамике. В исследовании использовалась экспериментальная
модель flanker test, адаптированная для детей. В этом случае вместо
стрелок испытуемым предъявлялись рисунки, изображающие рыбок. Стимулы были трех
видов: конгруэнтные (рыбки по бокам от центральной рыбки были направлены
головой в одну и ту же сторону), неконгруэнтные (рыбки по бокам были направлены
в противоположную сторону) и нейтральные (предъявлялась только одна рыбка). В
эксперименте принимали участие 4 группы детей по 12 человек (6, 7, 8 и 9 лет).
Результаты исследования продемонстрировали, что при наличии стимула-подсказки у
детей всех возрастов сокращается время выполнения задания и увеличивает процент
правильных ответов.
Таким образом, когнитивные исследования демонстрируют возможность
избирательного влияния предстимульного внимания, задаваемого инструкцией, на
обработку целевых сигналов.
Все вышеупомянутые примеры и экспериментальные парадигмы, связанные с
антиципирующем вниманием, представляют собой подготовку к анализу релевантного
стимула, обусловленную предварительной инструкцией-подсказкой. Исследование же
второго типа организации предстимульного внимания, который мы назвали
имплицитным предвосхищением, основанным на периодически повторяющемся порядке
появления релевантных сигналов разной модальности, не так популярно. В
контексте данного исследования мы предполагали, что такое повторение последовательности
сигналов с определенным порядком усваивается, и на основании этого
«неосознаваемого знание» формируется способность к предвосхищению. Чаще всего в
рамках изучения усвоения повторяющихся последовательностей речь идет об
имплицитном научении.
Говоря об имплицитном научении, A. Cleeremans et al [1998] описал в своей работе «Implicit learning: news from
the front» основные экспериментальные парадигмы изучения данного феномена.
Экспериментальные ситуации, связанные с имплицитным научением обычно включают
в себя три компонента:
) Наличие определенной сложной экспериментальной среды заданной
правилами в условиях имплицитного научения.
2) Мера оценки, насколько хорошо испытуемый усвоил знание о среде,
выполняя одинаковые или разные задания
) Мера, отражающая степень осознанности усвоенных знаний
испытуемым
При исследовании имплицитного научения используются 3 основные
экспериментальные парадигмы, имеющие схожий дизайн: усвоение искусственной
грамматики (artificial grammar learning (AGL)) [Reber, 1967, 1989, 1993], усвоение последовательностей -
серийное научение (sequence learning (SL)) [Nissen, Bullemer, 1987; Reber, 1993; Lewicki et al, 1988] и контроль динамичной системы (dynamic system control
(DSC)) [Berry and Broadbent, 1984].
Поскольку в данной работе в качестве одной из моделей для изучения
влияния предстимульного внимания на эффективность решения когнитивной задачи
использовалась парадигма усвоения повторяющейся последовательности сенсорных
стимулов, представляется важным более подробно остановиться на исследованиях,
посвященных изучению серийного научения.
При изучении усвоения последовательностей испытуемых просят реагировать
(обычно используется моторная реакция) на каждый элемент последовательности
событий, следующих в определенном и периодически повторяющемся порядке, при
этом регистрируется время реакции. Испытуемым не сообщается, что за
последовательностью повторяющихся стимулов стоит определенная закономерность.
Чаще всего в подобных исследованиях используются последовательности зрительных стимулов.
[Nissen, Bullemer, 1987; Reber, 1993; Lewicki et al, 1988]. При использовании
повторяющегося порядка стимулов испытуемые демонстрируют более высокую скорость
реакции, чем при случайном предъявлении стимулов.
Так, например, M.J. Nissen и P. Bullemer [1987] исследовали имплицитное научение в условиях
предъявления периодически повторяющихся последовательностей зрительных
стимулов. Испытуемым предъявлялись зрительные стимулы (звездочки), которые
появлялись в четырех возможных местах. Кнопка ответного устройства
располагалась напротив каждого потенциального места появления стимула. После
ответа следующий стимул появлялся через 500 мс. Для одной группы испытуемых
появление стимулов было задано определенной 10-ти элементной
последовательностью, в то время как для другой было случайным.
Испытуемые, которым предъявлялась повторяющаяся последовательность,
демонстрировали более высокую скорость реакции и делали меньше ошибок по
сравнению со второй группой. В исследовании Kushner et al [1991] также изучалось изменение успешности деятельности в
ходе усвоения определенной заданной последовательности сигналов. В этом случае
испытуемым на мониторе компьютера последовательно в разных местах экрана
предъявлялось 5 стимулов. Задача состояла в том, чтобы предугадать в каком
месте должен появиться шестой стимул. Для релевантного стимула было
предусмотрено 3 позиции. Первые 5 стимулов предъявлялись в хаотичном порядке,
однако появление 6-ого стимула зависело от пространственного расположения
второго и четвертого стимулов относительно друг друга. Эксперимент был разделен
на три этапа. На первом этапе испытуемым было необходимо решить большое
количество такого рода заданий (2430). На втором этапе правило лежавшее в
основе появления шестого стимула менялось (972 проб), и теперь аналогичной
последовательности предъявления первых пяти стимулов из первого этапа
соответствовало другое место появление шестого стимула в эксперименте второго
этапа. На третьем этапе правило опять менялось (972 проб): предъявление шестого
стимула было случайным. При этом испытуемым не разглашались особенности и
детали эксперимента. Результаты исследования показали, что в ходе первого этапа
точность предугадывания места появления шестого стимула постепенно
увеличивалась, достигая показателя, значительно превышающего уровень случайного
угадывания. В начале второго этапа точность ответов падала до уровня случайного
угадывания, однако к концу этапа опять же значимо возрастала. В течение всего
третьего этапа испытуемые давали случайные ответы. Стоит отметить, что в
постэкспериментальных опросах испытуемые не могли ответить, какой принцип лежал
в основе появления последовательности стимулов. Это значит, что у испытуемых
формировалось имплицитное предвосхищение, возникающее на основании определенной
последовательности появления в пространстве определенных стимулов.
Рассмотренные выше данные дают основание предполагать, что закономерность
повторяющейся последовательности сигналов усваивается, и испытуемый готовится к
появлению очередного стимула заранее, хотя часто не может дать вербальный отчет
о порядке стимулов.
Существуют разные точки зрения на механизмы усвоения последовательности
сенсорных сигналов при серийном научении. Рассмотрим некоторые модели серийной
организации движений.
Под серийной организацией движения обычно понимают способность ЦНС
осуществить заранее упорядоченную последовательность движений в точном
соответствии с запланированным временным расписанием (например, игра на
музыкальном инструменте) [Курганский, 2014].
Одна из самых ранних и широко известных моделей - это модель цепочек.
Согласно этой модели серийный порядок обеспечивается образованием устойчивых
однонаправленных связей между последовательными элементами серии. Двигательное
воспроизведение серии основано на том, что для каждого элемента
последовательности выполняется ассоциированное с ним движение, причем такое
выполнение служит пусковым сигналом (например, в качестве пускового может быть
использован афферентный сигнал выполненного движения) для выполнения следующего
элемента серии. В рамках данной модели всю последовательность воспроизводимых
движений можно представить как цепочку рефлекторных ответов, в которой текущее
движение является реакцией на предыдущее, что связывает ее с концепцией
рефлекторной дуги.
K.S. Lashley [1951] возражал против
такой модели указывая на трудности, с которыми сталкивается такого рода схема.
Одной из таких проблем является наличие повторных элементов в серии, как
например в слове EVERY, где элемент
«E» занимает первое и третье место.
Таким образом, элемент E указывает как на V (второй элемент), так и на R
(четвертый элемент). В результате образуется развилка, и не ясно, каким образом
принимается решение, что первый раз процесс припоминания должен пройти по ветке
E-V, а второй - по E-R. Другой недостаток модели цепочек состоит в том, что
одна единственная ошибка, допущенная при выполнении серии, приведет к тому, что
и все последующие элементы серии будут либо не соответствовать заданному
порядку, либо вообще не будут выполнены [Курганский, 2014].
Одним из наиболее популярных семейств моделей серийной организации
движений являются CQ-модели (competitive queuing) [Grossberg, 1978; Rhodes et al., 2004; Bullock, 2004]. В рамках этого класса моделей
предполагается, что все элементы серии находятся в общей очереди и "соревнуются"
за право быть выполненными.
Все модели при этом имеют следующие уровни: уровень планирование
последовательности, уровень выбора и уровень реализации элементов серии. Первый
уровень определяет степень активации определенных нейронов, выраженной в
частоте их разряда. Распределение уровней активации задает желаемый порядок
выполнения элементов, и величина активации выражает степень срочности
выполнения элемента. На уровне выбора определяется порядок выполнения
элементов. Нейроны этого уровня, мембранный потенциал которых находится ниже
порогового уровня и количество которых однозначно соответствует количеству
нейронов вышележащего уровня, получают на вход сигналы, пропорциональные
активации нейронов уровня планирования и оказывают тормозящее воздействие на
своих соседей с помощью латерального торможения. Латеральное торможение тем
сильнее, чем выше уровень входного сигнала. Если нейрон уровня выбора достигнет
порога разряда, его разрядом будет инициирована одна из двигательных программ.
Одновременно "нейрон-победитель" окажет тормозное воздействие на
соответствующий нейрон уровня планирования по аксонной коллатерали, снизив его
активацию до нуля. В итоге будет побеждать тот элемент, которому соответствует
наибольшая активация.
В рамках анализа параметров деятельности невозможно понять, насколько
процессы, происходящие в предстимульный период как при серийном научении, так и
при произвольном внимании, могут быть избирательными, возможна ли действительно
избирательная настройка и подготовка к обработке информации или эффект
настройки состоит в общей активации. Для экспериментального исследования этих
проблем в нейрокогнитивной науке используются нейрофизиологические методы
анализа, позволяющие оценить работу различных корковых зон и глубинных структур
мозга в предстимульный период.
.2
Нейрофизиологические исследования избирательного предстимульного внимания
Избирательное ожидания сигнала с определенными свойствами на нейронные
процессы, обеспечивающие восприятие значимого сигнала, изучалось с помощью анализа
вызванных потенциалов мозга (ВП) уже давно в экспериментах B. Porjesz и H. Begleiter [1975]. Испытуемые заранее обучались
различать слабые и сильные вспышки, после чего сами предъявляли себе сигналы
определенной интенсивности. Таким образом, создавалась ситуация ожидания
сигнала с определенными свойствами. В процессе предъявления вспышек
экспериментатор заменял слабые или сильные стимулы на сигналы средней
интенсивности, не сообщая об этом испытуемому. При анализе ВП оказалось, что их
параметры определяются не реальной, а ожидаемой интенсивностью. Результаты
электрофизиологических исследований позволили предположить существование
опережающих активных избирательных влияний, регулирующих функциональное
состояние мозговых структур, участвующих в обработке релевантного стимула.
Вопрос о возможности селективной настройки мозговых систем в предстимульном
периоде произвольного внимания долгое время оставался дискуссионным.
Ch. Brunia
[1999] на основе анализа механизмов антиципирующего внимания пришел к выводу о
ключевой роли префронтальной коры и таламических ядер в обеспечении нисходящих
избирательных влияний на проекционные и ассоциативные области коры при
подготовке к анализу релевантного стимула. Префронтальная кора, по его мнению,
"организует" антиципирующее поведение, активируя корково-корковые и
таламо-корковые нейронные цепи (loops), таламические ядра также активируются
префронтальной корой, в особенности медиодорзальное ядро и подушка. В свою
очередь ретикулярное ядро осуществляет тормозные влияния на передачу информации
в релейных ядрах. Исследования с помощью позитронно-эмиссионной томографии
подтверждают участие фронто-таламической регуляторной системы в обеспечении
селективной преднастройки мозговых систем [McAlonan
et al., 2000; P.W. Burgess,
2001; Zikopoulos, Barbas, 2006, 2007; Marzinzik et al., 2008].
Существует много экспериментальных данных [Kastner and Ungerleider, 2000; Corbetta and Shulman, 2002; Serences and Yantis, 2006], которые свидетельствуют о
том, что предвосхищающее внимание к зрительным стимулам характеризуется
активацией различных регионов фронтальной, теменной и затылочной областей при
этом есть основания полагать, что облегчение идентификации целевого стимула при
наличии о нем предварительной вспомогательной информации может быть опосредовано
нисходящими управляющими сигналами из фронтальной и теменной коры,
модулирующими активность сенсорно-специфических зон [Desimone and Duncan, 1995; Kastner and Ungerleider, 2000; Corbetta and Shulman, 2002; Moore et al., 2003; Serences and Yantis,2006; S.L. Bressler
et al, 2008; Knudsen,
2007]. В работе S.L. Bressler и соавторов [2008] с помощью анализа
гемодинамической реакции в ходе фМРТ исследования анализировали направленные
статистические связи между активностью префронтальных (фронтальное глазодвигательные
поле - FEF) и теменных (внутренняя теменная
борозда - IPS) ассоциативных корковых зон и
активностью затылочных сенсорно-специфических областей в предстимульный период
зрительно - пространственного направленного внимания. В этом эксперименте
зрительный сигнал появлялся слева или справа от центра экрана и испытуемые
должны были определять его местоположение, нажимая на соответствующие ключи.
Предстимульное внимание моделировалось слуховым стимулом - подсказкой (слова
«слева» или «справа»). Было показано, что предстимульное
зрительно-пространственное внимание сопровождается усилением влияний от
фронтальных и теменных зон на зрительные специфические области (нисходящих
влияний (top-down influences)) и фронтальных зон на теменные
области. При этом степень нисходящих влияний от теменных зон правого полушария
на затылочные области обоих полушарий положительно коррелировала с
эффективностью решения когнитивной задачи. Активизация нисходящих управляющих
влияний префронтальной коры в процессе предстимульного направленного
зрительно-пространственного внимания показана в работе G.V. Simpson и соавт. при исследовании
магнито-энцефалограммы (МЭГ), зарегистрированной в период между предупреждающим
стимулом-подсказкой и целевым сигналом. Связанная с направленным вниманием
активность мозга была выявлена через 300 мс после появления предупреждающего
стимула во фронтальной глазодвигательной (FEF) области и латеральной париетальной зоне (LPF). Вслед за активацией этих зон
наблюдалась согласованная активность областей коры, входящих во
фронто-париетальную систему внимания: верхней лобной извилины (SFG), средней лобной извилины (MFG), латеральной (LIP) и передней (AIP) частей внутренней теменной борозды.
Роль предвосхищающих нисходящих модулирующих влияний ассоциативных
структур коры в избирательной модуляции активности корковых зон,
непосредственно участвующих в обработке сенсорной информации, демонстрируется
также в работах с искусственным «выключением» или наоборот «активированием»
префронтальных и теменных зон с помощью методов направленной магнитной
стимуляции отдельных мозговых структур (Transcranial Magnetic Stimulation (TMS)). Исследования, в которых метод TMS применялся для изменений активности
фронтального глазодвигательного поля человека [Ruff et al., 2006] и корковых
зон в области внутритеменной борозды [Ruff et al., 2008], а также исследования
с применением метода электрической микростимуляции фронтального
глазодвигательного поля обезьян [Moore and Armstrong, 2003], демонстрируют, что
активность зрительной коры зависит от модулирующих влияний, исходящих от
фронтального глазодвигательного поля и корковых зон, расположенных в области
внутритеменной борозды. Стимуляция этих зон, может привести к поведенческим
эффектам схожим с эффектами произвольного вниманием.
Роль избирательной предстимульной настройки функциональных систем мозга в
решении когнитивных задач продемонстрирована в исследовании Д.А. Фарбер в
соавт. [2014]. Исследование проводилось на взрослых испытуемых, при этом
анализировалось функциональное взаимодействие префронтальных областей коры с
другими корковыми зонами в период подготовки к опознанию неполных изображений
различного уровня фрагментации при их последовательном приближении к полному
изображению. Сопоставлялись электрофизиологические показатели функциональных
связей корковых областей в период, предшествующий появлению предупреждающего
сигнала (неспецифическое мобилизационное внимание) и в период, предшествующий
появлению целевого изображения (направленное предстимульное внимание). Было
обнаружено, что у испытуемых, успешно справившихся с заданием, предстимульное
внимание, направленное на анализ целевого сигнала, сопровождалось усилением
связей префронтальной коры и зрительных ассоциативных зон в правом полушарии в
тех случаях, когда целевой стимул еще не был опознан. В тех случаях, когда
целевое изображение опознавалось, аналогичные изменения преобладали в левом
полушарии. У неуспешных испытуемых, напротив, степень функционального
взаимодействия корковых зон при направленном предстимульном внимании была ниже,
чем при неспецифическом внимании, как в правом, так и в левом полушарии.
В электрофизиологических исследованиях показано, что в период ожидания
целевых стимулов происходит избирательная сонастройка нейронной активности не
только тех областей мозга, которые осуществляют регуляторные модулирующие
влияния, но и тех корковых зон, которые непосредственно участвуют в обработке
значимого сигнала. Так, B. Verhoef и коллеги [Verhoef et al., 2011] регистрировали нейронную активность и локальные
корковые потенциалы у макак при ожидании целевого зрительного стимула в задачах
на различение объемных выпуклых и вогнутых изображений. Они наблюдали рост
степени функционального взаимодействия между отдельными нейронами и локальными
нейронными сетями в зонах коры дорзальной и вентральной зрительных систем,
специфическим образом участвующих в обработке 3D информации - нижней височной (inferior temporal - IT) и передней интерпариетальной (anterior interparietal - AIP). При этом изменения нейронной активности в зоне AIP положительно коррелировали с
успешностью различения объемных изображений. В исследованиях на нейронах
приматов [цит. по Knudsen, 2007]
было показано, что нисходящий контроль фронто-париетальных систем внимания
избирательно увеличивает сенсорную чувствительность нейронов зрительных полей V4 и MT, рецептивные поля которых совпадают с характеристиками
целевых зрительных стимулов.
Избирательная настройка сенсорно-специфических корковых зон в процессе
подготовки к различению релевантных сигналов была обнаружена и в исследованиях
электрической активности мозга человека. Анализ ЭЭГ взрослых и детей младшего
школьного возраста [Мачинский и соавт., 1990; Мачинская, 2001] позволил
обнаружить, что ожидание сенсорных сигналов разной модальности (слуховых и
тактильных), определяемое предварительной инструкцией, сопровождается
формированием избирательных функциональных связей сенсорно-специфических и
ассоциативных корковых зон. Так при подготовке к различению слуховых сигналов
растет степень синхронизации по альфа-ритму височных (слуховых) зон с лобными и
заднеассоциативными зонами, а при подготовке к различению тактильных сигналов
аналогичные изменения происходят в соматосенсорной коре. В этих же зонах коры
усиливается вызванная электрическая активность мозга (ССП) уже при обработке
значимых сигналов по сравнению с ситуацией простого наблюдения.
Как показано в работе J.Bollinger
и коллег [2010] подготовка к анализу внешних сигналов с определенными
свойствами влияет не только на корковые зоны, участвующие в анализе простых
сенсорных сигналов, но и на те области коры, которые обеспечивают восприятие и
сохранение в рабочей памяти лицевых паттернов и сложных сцен. В этом
исследовании с помощью фМТР была обнаружена активация «лицевой области»
нижневисочной коры, а также рост функционального взаимодействия этой корковой
зоны с корковыми областями, расположенными в нижнелобном соединении (inferior frontal junction - IFJ), среднелобной (middle frontal gyrus - MFG) и
нижнелобной извилинах (inferior frontal
gyruss - IFG), и внутренней теменной борозде (intraparietal sulcus) в период после предъявления подсказки,
сигнализирующей о типе целевого сигнала (лицевого паттерна). Рост степени
функционального взаимодействия между зрительной лицевой областью и IFJ оказывал положительное влияние на
сохранение информации о целевом лицевом паттерне в рабочей памяти, а
аналогичные изменения совместной активности зрительной лицевой области и MFG - на сохранение значимой информации
в долговременной памяти.
Представленные выше нейрофизиологические исследования направленного
предстимульного внимания свидетельствуют о возможности избирательной настройки
функциональных систем мозга на обработку значимой информации и роль фронтальных
и фронто-париентальных модулирующих систем внимания в предвосхищающей подготовке
сенсорно-специфических областей коры к анализу значимой информации.
Что касается имплицитных установок, В 1997 году J. Doyon et al было проведено исследование, в котором анализировалась роль
полосатого тела, мозжечка и лобных долей при имплицитном усвоении
последовательности. В этой работе принимали участие испытуемые с болезнью
Паркинсона, с поражениями мозжечка или с ограниченным поражением лобных долей,
а также испытуемые без органических поражений мозга. В качестве
экспериментальной модели использовался тест повторяемых последовательностей
(Repeated Sequence Test), при выполнении которого испытуемые должны были
воспроизводить определенную последовательность движений пальцев в соответствии
с последовательностью зрительных сигналов. Испытуемым предъявлялось 4 блока
проб (40 предъявлений 10-ти элементной последовательности) в течение 6 дней. В
рамках последней экспериментальной сессии испытуемые так же проходили 2 теста
для оценки их эксплицитного (declarative) знания последовательности. Только пациенты
с синдромом Паркинсона с билатеральной дисфункцией полосатого тела или пациенты
с поражением мозжечка не смогли улучшить свои результаты в последних трех
экспериментальных сессиях, что говорит о снижении эффективности (impairment) процесса имплицитного научения в
этих случаях. Дальнейший анализ свидетельствовал о том, что возникающие при
патологии стриопалидарной системы или мозжечка нарушения имплицитного усвоения
последовательности зрительно-моторных реакций не могут быть объяснены общим
дефицитом когнитивных функций, волнением или тяжестью двигательных симптомов. В
свою очередь, уровень эксплицитного (declarative) знания последовательности не
различался среди трех клинических групп и соответствующих им групп испытуемых
без нарушений. Эти сведения говорят в пользу того, что в отличие от
эксплицитной памяти (declarative memory) поэтапное усвоение нового
зрительно-моторного навыка зависит от «сохранности» полосатого тела и мозжечка,
но не лобных долей. Роль полосатого тела во время серийного научения подтверждается
так же рядом других работ [J. Doyon et al, 1997; S. L. Rauch et al, 1997] с использованием функциональной МРТ. Вместе с тем,
нельзя исключить роль коры головного мозга в реализации процессов имплицитного
предвосхищения. В исследовании S.M. Landau и M. D’Esposito [2006] усвоение и моторное
воспроизведение последовательностей изучалось по средством фМРТ на испытуемых
играющих и не играющих на пианино. При этом было установлено, что усвоение
последовательностей в обеих группах сопровождается увеличением билатеральной
активации в соматосенсорной и теменной коре. Наблюдалась более интенсивная
активация в сети префронтальных, сенсомоторных и теменных областей правого
полушария у пианистов по сравнению с не пианистами.
Значительный интерес для выявления возможных различий в функциональной
организации мозга при произвольном направленном внимании и имплицитном
предвосхищении представляет исследование David Soto и др. [2012]. Внимание, как направляемое инструкцией,
так и формирующееся при имплицитном научении, тесно связано с актуализацией
следов предшествующего опыта. Исследование D. Soto в соавт.
исходит из представлений о различном влиянии процессов рабочей памяти, которые
активируются при произвольном внимании, и прайминга, основанного на имплицитном
запоминании, на организацию внимания человека. Было выдвинуто предположение,
что РП и прайминг могут обеспечиваться различными нейронными механизмами. Роль
процессов памяти в изменениях активности зрительной коры при смещениях внимания
(attention biases), остается неясной, так как в большинстве
исследований память и избирательные процессы не разграничиваются. В работе D. Soto и соавт применялся метод TMS для изучения роли рабочей памяти и прайминга в
модуляции активности зрительной коры при сдвигах внимания. В эксперименте
испытуемые должны были искать кольца Ландольта, о появление которых
сигнализировали цветные стимулы-подсказки. Информация о стимулах должна была
удерживаться в рабочей памяти эксплицитно или же имплицитно на основании их
повторяющейся последовательности и в дальнейшем проверяться тестом на
узнавание. С помощью магнитной стимуляции затылочной доли больших полушарий
было смоделировано влияние актуализации следов памяти на успешность поиска.
Характер такого влияния зависел от «вида» памяти. В условиях рабочей памяти
(эксплицитной), использование TMS
одновременно с подсказками о появлении целевого стимула увеличивало успешность
поиска по сравнению с условием, когда стимуляция сопровождала подсказку о
появлении нерелевантного стимула. Обратный эффект наблюдался в условиях
прайминга (имплицитной памяти). Эти данные рассматриваются авторами как
свидетельства различных механизмов влияния эксплицитного и иплицитного
запоминания на зрительное внимание при решении зрительных когнитивных задач.
Таким образом, анализ результатов экспериментально-психологических и
нейрофизиологических исследований подтверждает правомерность основной гипотезы
исследования о различном влиянии произвольного избирательного внимания и
имплицитного предвосхищения на эффективность когнитивной деятельности, что
может быть обусловлено различной функциональной организацией мозга в
предстимульный период. В настоящей работе для экспериментального исследования
этого предположения использовался комплексный подход, сочетающий анализ параметров
деятельности при различении сенсорных сигналов и электрофизиологический анализ
функционального взаимодействия корковых зон в период, предшествующий их
появлению, в двух экспериментальных условиях - при произвольном внимании,
направляемом инструкцией и имплицитном серийном научении.
Эмпирическое
исследование
Задача исследования:
) Проанализировать функциональную организацию коры больших полушарий при
решении когнитивных задач в двух условиях, отличающихся типом организации
предстимульного внимания на основании количественного анализа ритмических
составляющих суммарной электрической активности мозга (ЭЭГ), зарегистрированной
в период ожидания целевого стимула.
Глава 2.
Методические аспекты исследования
Влияние организации предстимульного внимания оценивалось при решении
следующей когнитивной задачи: испытуемый должен был определить, какой из
двух последовательно предъявляемых с небольшим интервалом зрительных или
слуховых стимулов с определенными сенсорными характеристиками появился первым,
и нажать на один из трех возможных ответных ключей (третий ключ испытуемый
должен был нажать, если он не был готов дать ответ в отведенное время).
Стимульный материал:
Зрительные стимулы: вытянутые прямоугольники двух оттенков серого
(светлого и темного) с угловыми размерами 2.5 ´ 2.5 градуса, появляющиеся с
небольшим временным интервалом на черном экране монитора под углом 45 градусов
друг к другу в центральном поле зрения (рис. 1).




Рис. 1. Все возможные варианты следования пар зрительных стимулов (в
наложении: первым появляется стимул, изображенный на заднем плане)
Слуховые стимулы: звуки двух разных частот (300 Гц и 3000 Гц) также
следующие с небольшим интервалом друг за другом, предъявляющиеся биноурально.
Решению основной задачи исследования предшествовал предварительный выбор
оптимальных параметров сенсорных характеристик стимулов в специально
организованном предварительном эксперименте.
Техническое и программное обеспечение эксперимента:
Эксперимент проводился на компьютере с помощью специального программы EEGExProc, разработанной в лаборатории
нейрофизиологии Института возрастной физиологии РАО. Программа позволяет
заранее создавать шаблон проведения эксперимента, управлять предъявлением
стимулов, регистрировать ответные реакции испытуемого и синхронизировать
события эксперимента с записью электроэнцефалограммы. В ходе проведения
эксперимента автоматически велся протокол, куда в формате TXT заносилась вся последовательность
событий эксперимента. Протокол затем обрабатывался в среде MATLAB для вычисления средних по условиям
эксперимента показателей деятельности для каждого испытуемого. Количественная
оценка ЭЭГ для исследования динамики функционального взаимодействия корковых
зон производилась offline в среде MATLAB с помощью специально разработанных
методик (см. раздел «Регистрация и анализ ЭЭГ»)
Для регистрации ЭЭГ использовался 128-канальный электроэнцефалограф Geodesic EEG system (Electrical Geodesics, Inc., USA).
.1
Предварительный эксперимент. Оптимизация параметров стимулов и уравнивание
сложности задач на различение зрительных и слуховых сигналов
В предварительном эксперименте приняли участие 9 взрослых здоровых
испытуемых в возрасте от 20 до 50 лет, правшей с нормальным или
скорректированным до нормального зрением. Все испытуемые дали письменное
информированное согласие на участие в исследовании. Цель предварительного
эксперимента состояла в определении оптимальных параметров зрительных и
слуховых стимулов. Поскольку сложность задания изменялась в зависимости от
интервалов между двумя сенсорными сигналами в паре стимулов, то на данном этапе
исследования использовались разные длительности интервалов и затем отбирались
те, при которых как зрительные, так и слуховые задачи решались на надпороговом
уровне с вероятностью 60-80%
Предварительный эксперимент состоит из двух этапов, каждый из этапов в
свою очередь состоит из двух сессий: «слуховой» и «зрительной». На первом этапе
предварительного эксперимента испытуемому предъявлялись пары зрительных и
слуховых стимулов, варьирующих в зависимости от межстимульного интервала.
Использовалось 8 интервалов: 5мс, 10мс, 20мс, 40мс, 80мс, 150мс, 300мс, 500мс
(+15 мс, с учетом времени развертки экрана компьютера). На этом этапе
вычислялся процент правильно выполненных заданий по всей выборке испытуемых. На
основании этих данных было выбрано 3 интервала наиболее соответствующих
вышеуказанным критериям. На втором этапе предъявлялись пары стимулов с тремя
выбранными по результатам первого этапа межстимульными интервалами. По
результатам второго этапа был выбран наиболее оптимальный межстимульный
интревал, который затем использовался в основном эксперименте.
Процедура эксперимента:
Зрительная сессия
Перед началом эксперимента испытуемым давалась подробная инструкция о
характере задания и требованиях к ее решению. После того, как у испытуемого не
оставалось вопросов по поводу прохождения эксперимента, запускалась
экспериментальная модель. С самого начала на черном фоне в центре экрана
появлялся серый фиксационный крест. После того как испытуемый был готов к
прохождению эксперимента, он нажимал левую клавишу компьютерной мыши. После
этого через 500 мс в центре экрана на 15мс появлялся зрительный стимул
темно-серого или светло-серого цвета. Затем через фиксированный интервал (5мс,
10мс, 20мс, 40мс, 80мс, 150мс, 300мс или 500мс) в центре экрана на 15мс
появлялся второй стимул, отличающийся цветом и наклоном от первого. Для каждого
варианта межстимульного интервала использовались 4 пары зрительных стимулов,
отличающиеся порядком следования светлых и темных полос и их наклоном (рис. 1)
(всего 32 пробы). Различные пары стимулов следовали в псевдослучайном порядке.
После предъявления пары зрительных стимулов в центре экрана снова появляется фиксационный
крест. Задача испытуемого состояла в определении порядка следования светлых и
темных полосок и нажатии одной из трех возможных кнопок клавиатуры. Варианты
ответа: «первым появился прямоугольник темно-серого цвета», «первым появился
прямоугольник светло-серого цвета», «не знаю». Ответ испытуемый мог давать в
течение 10 с. Ответы испытуемого автоматически заносились в протокол в формате
(.txt). В протоколе фиксировались последовательности предъявления пар стимулов,
их характеристики, ответ и время реакции испытуемого.
Слуховая сессия
В данной сессии целевые стимулы предъявлялись через наушники биноурально.
Порядок следования событий в сессии был таким же, как в сессии со зрительной
стимуляцией. В отличие от зрительной сессии в данном эксперименте использовалось
два варианта заданий, отличающихся только порядком следования высоких и низких
звуков (всего 16 проб).
Ответы испытуемых группировались в соответствии с межстимульными
интервалами - 5мс, 10мс, 20мс, 40мс, 80мс, 150мс, 300мс и 500мс. Таким образом,
для «зрительной сессии» было проанализировано 36 типов реакций испытуемых (4´9), а для «слуховой сессии» - 18 (2´9). Для каждого типа реакций был
рассчитан процент правильных ответов по группе испытуемых. На основании
процента верно решенных задач обеих модальностей из 8-ми интервалов было
отобрано 3 интервала - 20мс, 40мс и 80мс, при которых процент верно решенных
задач в группе испытуемых попадал в интервал от 60 до 80%.
Процедура второго этапа предварительного эксперимента, как «зрительной»,
так и «слуховой» сессии была аналогична процедуре первого этапа. Количество
интервалов между предъявляемыми стимулами было уменьшено до трех - 20мс, 40мс,
80мс. Каждому испытуемому предъявляется по 20 задач с каждым из интервалов.
Таким образом, в каждую сессию второго этапа эксперимента входило 60
экспериментальных проб.
По каждому испытуемому был подсчитан индивидуальный процент правильных
ответов по каждому интервалу - 20мс, 40мс, 80мс (всего по 20 ответов). На
основании среднего по группе процента верно решенных задач из трех интервалов
нами был выбран интервал 40 мс как для слуховой, так и для зрительной задачи
при котором, в случае со «зрительной сессией», процент верно решенных задач
составил - 66,1%, а в случае со «слуховой сессией» - 61,5%.
.2
Основной эксперимент
В основном эксперименте использовались сенсорные зрительные и слуховые
сигналы, протестированные и отобранные в предварительном эксперименте. Во всех
пробах использовался интервал между сигналами равный 40 мс, что согласно данным
предварительного эксперимента удовлетворяло требованию надпороговой трудности
сенсорной задачи, как для зрительной, так и для слуховой модальности.
Аналогично предварительному эксперименту задача испытуемого состояла в
определении того, какой из двух зрительных или слуховых сигналов с определенным
характеристиками следовал первым в паре стимулов, предъявляемых с коротким
интервалом.
Основной эксперимент состоял из двух сессий - направленное
внимание и серийное научение, различающихся порядком следования пар зрительных
и слуховых стимулов и наличием или отсутствием стимула-подсказки. Порядок сессий чередовался от
испытуемого к испытуемому для исключения систематического влияния
продолжительности исследования.
В сессии направленное внимание появлению целевых стимулов предшествовало
предъявление стимулов-подсказок со схематическим изображением уха или глаза,
сигнализирующих о модальности (рис. 2). Модальность целевых стимулов
чередовалась в псевдослучайном порядке.

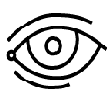
Рис. 2. Стимул-подсказка «ухо» и «глаз», указывающие на слуховую и
зрительную модальность соответственно.
В сессии серийное научение пары стимулов предъявлялись без подсказки, а
последовательность их предъявления повторялась и подчинялась определенному
правилу.
Испытуемые: 20 взрослых здоровых людей (10 мужчины и 10 женщин) в
возрасте 23±5.7 лет, правшей с нормальным или скорректированным до нормального
зрением.
Процедура эксперимента:
Сессия направленное внимание
Перед началом эксперимента испытуемым давалась инструкция (см. ниже).
Каждая проба эксперимента начиналась с появления фиксационного крестика в
центре экрана монитора. После этого, через промежуток времени в 1000-1500мс в
центре экрана на 80мс предъявлялся предупреждающий стимул, информирующий
испытуемого о модальности целевого сигнала. За предупреждающим стимулом через
3500-4000мс следовала пара либо слуховых (если стимул-подсказка был изображение
уха), либо зрительных стимулов (если стимул-подсказка был изображение глаза). В
этой сессии эксперимента последовательность появления зрительных и слуховых
задач была псевдослучайной и испытуемый мог давать двигательный ответ в течение
2 с. В качестве ответного устройства использовался геймпад, три кнопки которого
испытуемый использовал для ответов. Количество когнитивных задач в зрительной и
слуховой модальности было одинаковым - по 40 предъявлений. Таким образом,
эксперимент состоял из 80 проб. Последовательность событий в одной пробе
схематично представлена на рис. 3.
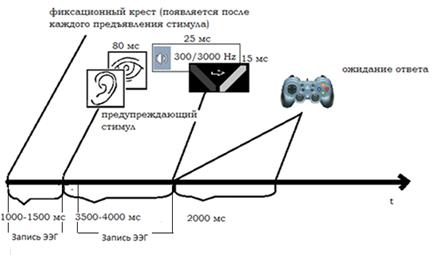
Рис. 3. Последовательность событий в одной экспериментальной пробе в
сессии направленное внимания
Сессия серийное научение
В этой сессии предъявлялись сенсорные задачи с теми же параметрами
целевых сигналов и интервалами между ними. Особенностью этой сессии было
периодическое повторение экспериментальных проб с одинаковым порядком
следования зрительных и слуховых задач и отсутствие стимулов-подсказок. Каждая
проба состояла из 8 предъявлений пар целевых сигналов (рис. 4): [темно-серый
прямоугольник (наклон вправо)/светло-серый прямоугольник (наклон влево)] -
[светло-серый прямоугольник (наклон вправо)/темно-серый прямоугольник
(наклон влево)] - [высокий тон/низкий тон] - [низкий тон/высокий
тон] - [высокий тон/низкий тон] - [светло-серый прямоугольник
(наклон вправо)/темно-серый прямоугольник (наклон влево)] - [темно-серый
прямоугольник (наклон вправо)/светло-серый прямоугольник (наклон влево)] -
[низкий тон/высокий тон]. Используемое количество задач в одной
повторяющейся последовательности было подобрано таким образом, чтобы
избежать осознанного усвоения последовательности целевых сигналов и вместе с
тем обеспечить возможность ее имплицитного усвоения. Сессия состояла из
30 проб, которые при обработке данных были разделены на три последовательных
этапа по 10 проб в каждом. Это было необходимо для того, чтобы можно было
отследить динамику имплицитного усвоения последовательности и при анализе
параметров деятельности выбрать этап, на котором усвоение уже произошло.
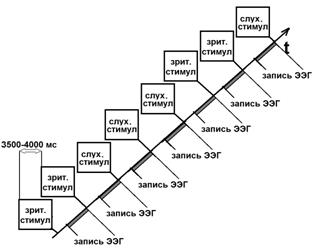
Рис. 4. Схематичное изображение последовательности событий в одной
экспериментальной пробе в сессии серийное научение
Инструкция испытуемому:
Вам будут предъявляться пары зрительных и слуховых стимулов. В паре
зрительных стимулов будут использоваться полоски двух оттенков серого
(светло-серый и темно-серый), которые с небольшим интервалом будут следовать
друг за другом. Порядок появления светло-серых и темно-серых полосок может
меняться. Ваша задача - определить, какая из двух полосок, светлая или темная
появлялась первой и нажать на одну из трех кнопок ответного устройства: кнопку
2, если первой была темная полоска, 3 - если первой была светлая полоска, 1 -
если Вы не знаете ответа. В паре слуховых стимулов будут использоваться два
тона, высокий и низкий, которые также с небольшим интервалом будут следовать
друг за другом, порядок следования высокого и низкого тона в паре также может
меняться. Ваша задача - определить, какой из двух тонов был первым, высокий или
низкий, и нажать на одну из трех кнопок: 2 - если первым был низкий тон, 3 -
если первым был высокий тон, 1 - если Вы не знаете ответа. Отвечать надо как
можно точнее и быстрее.
Исследование состоит из двух сессий. В одной сессии появлению пар
стимулов будет предшествовать подсказка, сигнализирующая о типе сигналов:
изображение глаза перед зрительными стимулами, изображение уха - перед
слуховыми стимулами. Ориентируясь на эту подсказку, Вы можете настроиться на
восприятие зрительных или слуховых сигналов.
В другой сессии пары зрительных и слуховых сигналов будут появляться без
предупреждения. Время предъявления каждого из стимулов и интервал между ними
очень короткие, и может показаться, что это один стимул, однако их там два.
Ваша задача определить, какой из стимулов был первым.
Регистрация и анализ ЭЭГ
В области экспериментального исследования функциональных корково-корковых
взаимодействий широко применяется метод электроэнцефалографии (ЭЭГ). ЭЭГ -
неинвазивный метод исследования функционального состояния головного мозга путем
регистрации суммарной электрической активности множества нейронов головного
мозга. В основе колебаний потенциалов, регистрируемых с поверхности головы в
виде ЭЭГ, лежат изменения внутриклеточных мембранных потенциалов (МП) корковых
пирамидных нейронов [Гусельников, 1976]. Характерной особенностью ЭЭГ является
ее ритмический характер. Общность ритмических сигналов в паре корковых зон
оценивается с помощью функции когерентности. Рост значений функции
когерентности ЭЭГ для колебаний определенной частоты в паре корковых зон рассматривается
как показатель функционального взаимодействия между ними.
В настоящем исследовании в процессе решения когнитивных задач
регистрировалась многоканальная ЭЭГ с помощью 128-канального
электроэнцефалографа Geodesic EEG system (Electrical Geodesics, Inc., USA).
ЭЭГ регистрировалась в полосе частот от 0.1 до 70 Гц и оцифровывалось с
частотой 250 Гц. Подготовка записанной электроэнцефалограммы для анализа
когерентности проводилось в 2 этапа. На первом этапе при визуальном просмотре
кривых ЭЭГ на сегментах равной длительности, характеризующих определенное
экспериментальное условие, отмечались каналы и участки сигнала, содержащие
артефакты - помехи различного происхождения, связанные с движениями глаз
испытуемого, нарушением контакта электрода с поверхностью головы, мышечным
напряжением и др. (рис. 5).

Рис. 5. Пример сегмента ЭЭГ, с отмеченными артефактными каналами и
участками сигнала
На втором этапе свободные от артефактов эпохи ЭЭГ разбивались на короткие
неперекрывающиеся сегменты длительностью 200 мс. Это было связано с тем, что
наличие артефактных фрагментов в изначально одинаковых по длительности
сегментах ЭЭГ не позволяло использовать весь сегмент для дальнейшего анализа, и
уменьшало длительность его «полезных» для обработки участков. В связи с тем,
что артефактные участки могли различаться по длительности и моменту появления,
это приводило к различиям в длительности отрезков ЭЭГ свободных от артефактов.
Функции КОГ, вычисленные по эпохам разной длительности сравнивать между собой
не корректно. Именно поэтому для дальнейшего анализа использовались «составные»
эпохи, образованные из одинакового числа коротких сегментов длительностью 200
мс. При этом сегмент длительностью 200 мс является достаточно длинным для его
дальнейшей обработки и достаточно коротким для того, чтобы полезная ЭЭГ могла
быть разбита на максимальное количество таких сегментов при минимальной потери
информации. Количество таких сегментов в связи с разной длиной свободной от
артефактов записи, варьировалось от испытуемого к испытуемому. Для того чтобы
избежать влияния этого обстоятельства на анализируемые параметры когерентности
ЭЭГ, при расчетах индивидуальных параметров когерентности использовалась
последовательность, составленная из десяти 200-миллисекундных сегментов записи.
Выбор количества суммируемых отрезков был обусловлен размером самой короткой
“чистой” записи в исследуемой выборке.
Функциональные корково-корковые взаимодействия оценивались с помощью
12-канальной векторной авторегрессионной модели (ВАР-модель) [Курганский,
2010]. Использование ВАР-моделей позволяет оценить классическую функцию
когерентности J(f) =|Ckm(f)| (КОГ), пример функции КОГ
представлен на рис. 6.

Рис. 6. Пример графика функции когерентности ЭЭГ для пары отведений F7 левого полушария (нижнелобное) и O1 (затылочное), вычисленной для
одного из испытуемых
Математически ВАР-модель представляет собой выражение, описывающее
зависимость векторного сигнала от самого себя в прошедшие моменты дискретного
времени n:
x(n) = 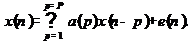
Здесь
a(p) обозначает квадратную матрицу, элементы которой akm(p) показывают, какой
вклад в текущее значение сигнала xk(n) в канале k вносит сигнал xm(n - p),
зарегистрированный p отсчетов назад в канале m. Порядок P выбирается с таким
расчетом, чтобы объяснить всю линейную корреляционную структуру исходного
многоканального сигнала x(n). Последнее означает, что многоканальная
последовательность остатков авторегрессии e(n) не обладает никакой линейной
корреляционной структурой, т.е. является белым шумом и характеризуются
ковариационной матрицей V.
В
частотной области ВАР-модель описывается матричным соотношением:
(f)
= H(f)E(f), (2)
где
X(f) и E(f) обозначают векторы-столбцы Фурье-образов, соответственно, сигнала
x(n) и остатков авторегрессии e(n), а H(f) для каждой частоты f является
квадратной M ×
M матрицей, где М - число каналов [цит.
по Курганский и Мачинская, 2012].
Для
нейрофизиологии непосредственный интерес представляют взаимодействия между
областями коры. Между тем, в силу влияния объемного проведения биотоков мозга
оценка силы функциональных связей на основе непосредственно регистрируемой ЭЭГ
(в пространстве сенсоров) является лишь косвенным методом оценки истинных корковых
взаимодействий. Регистрация ЭЭГ высокой плотности (64 канала и более) позволяет
оценить распределение биоэлектрической активности (аналог фокального
потенциала) непосредственно в кортикальной ткани. Тем самым появляется
возможность оценить силу истинных корковых функциональных связей
непосредственно в пространстве источников.
Вычисление
биоэлектрической активности в пространстве источников проводится в несколько
этапов. Эти этапы включают разбиение всего объема корковой ткани на множество
субсантиметровых элементарных ячеек вокселов (voxel - Volume Element).
Поскольку апикальные дендриты пирамидных клеток, являющихся основными
источником ЭЭГ, расположены параллельно друг другу и при этом перпендикулярно
поверхности коры, каждому вокселу соответствует вполне определенное направление
электрического диполя. В каждый момент времени n наблюдаемая в отведениях ЭЭГ
представляет собой взвешенное среднее алгебраическое влияний диполей всех
вокселов в этот же момент времени. В сокращенном матричном виде это записывается
как
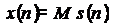
Матрица
M вычисляется с помощью методов решения прямой задачи электроэнцефалографии
[Hallez et al., 2007].
Зная
матрицу M, с помощью методов решения обратной задачи электроэнцефалографии
[Grech et al., 2008], можно найти матрицу W, которая осуществляет обратную
операцию - преобразует наблюдаемую ЭЭГ в сигналы, порождаемые в каждом вокселе.
Матричная запись этого преобразования выглядит как
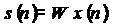
Большое
число вокселов (несколько тысяч) исключает возможность построения единой
векторной авторегрессионной модели для всех вокселов. Поэтому оценка силы
функциональных связей в пространстве источников проводилась лишь для немногих
сигналов, характеризующих активность в специально выбранных участках коры -
областях интереса (regions of interest
- ROI).
Исходя
из литературных данных (Глава 1), в качестве ROI были выбраны
следующие билатерально-симметричные области коры, координаты которых были
определенны в соответствии с атласом Монреальского неврологического института (MNI) c
помощью SPM anatomy toolbox
(V1.8) [Eickhoff et al., 2005]
Таблица 1
|
Области коры
|
Сокращенное обозначение
областей коры
|
MNI - координаты
|
V1 (L)
|
X = -9.8407, Y = -88.1789 Z =
-8.0946
|
|
V1(R)
|
X = 9.8407, Y
= -88.1789 Z = -8.0946
|
|
Слуховая проекционная зона
(Auditory 1)
|
A1 (L)
|
X = -39.9763, Y
= -32.8018 Z = 14.5984
|
|
A1 (R)
|
X = 39.9763, Y =
-32.8018 Z = 14.5984
|
|
Латеральная префронтальная
кора (Lateral prefrontal cortex)
|
LPFC (L)
|
X = -44, Y = 40
Z = 20
|
|
LPFC (R)
|
X = 44, Y = 40 Z
= 20
|
|
Фронтальная
глазодвигательная область (frontal eye
field)
|
FEF (L)
|
X = -30, Y = -4
Z = 60
|
|
FEF (R)
|
X = 30, Y = -4 Z
= 60
|
|
Латеральная
интрапариетальная зона (lateral intraparietal cortex)
|
LIP (L)
|
X = -31.5696, Y
= -55.1989 Z = 49.6191
|
|
LIP (R)
|
X = 31.5696, Y =
-55.1989 Z = 49.6191
|
|
Интрапариетальная кора (intraparietal cortex)
|
IPC (L)
|
X = -51.6499, Y
= -55.941 Z = 44.9097
|
|
IPC (R)
|
X = 51.6499, Y =
-55.941 Z = 44.9097
|
В таблице 1 показаны MNI-координаты центров выбранных ROI, каждая из которых содержит
множество вокселов (от десятков до сотен). Для того, чтобы охарактеризовать
типичную для каждой ROI зависимость
фокального потенциала от времени применялся следующий прием. Для каждого ROI вокруг его центра (см. таблицу 1)
строилась сфера радиусом 10 мм. Сигналы всех вокселов, оказавшихся внутри этой
сферы, рассматривались как многоканальный сигнал (фокальный потенциал),
характеризующий данную область коры. К этой многоканальной записи фокального
потенциала применялся метод принципиальных компонентов, и в качестве
представителя ROI выбирался первый компонент, который
характеризует наиболее выраженную общую активность, присутствующую в
многоканальной записи.
Затем на основе полученной 12-канальной записи фокального потенциала
вычислялись коэффициенты авторегрессионной модели. Дальнейшие вычисления
функции когерентности на основе этих коэффициентов проводились в точности так
же, как и в пространстве сенсоров.
Вычисления матриц M и W осуществлялось с помощью программной системы
spm8, разработанной в Wellcome Trust Centre for Neuroimaging
(#"781081.files/image014.jpg">
Рис. 7. Среднее по выборке испытуемых значение ВР (в мс) при решении
зрительной и слуховой задачи в процессе серийное научение. 1, 2, 3 - этапы
серийного научения. Линии погрешности на графике отражают величину ошибки
среднего
Таким образом, по мере повторения последовательностей чередования
слуховых и зрительных сигналов происходило сокращение ВР для обеих
модельностей, что свидетельствует о наличии эффекта серийного научения по этому
параметру деятельности. Однако этот эффект был больше выражен при решении
слуховой задачи. Как при различении зрительных, так и при различении слуховых
сигналов время решения на первом этапе существенно отличалось от времени
решения, как на втором, так и на третьем этапе, второй и третий этапы не
различались. Это дает возможность считать, что эффект научения проявляется уже
на втором этапе предъявления сенсорных сигналов.
Точность
Зависимость успешности решения когнитивной задачи (процента правильных
ответов) от модальности целевых сигналов и этапа предъявления повторяющихся
последовательностей слуховых и зрительных стимулов представлена на рис. 8.
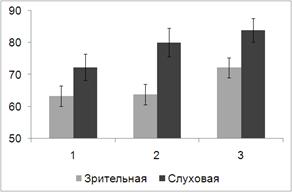
Рис. 8. Средний по выборке испытуемых процент правильных ответов при
решении зрительной и слуховой задачи в процессе серийное научение. 1, 2, 3 -
этапы серийного научения. Линии погрешности на графике отражают величину ошибки
среднего
Дисперсионный анализ выявил основной эффект фактора МОДАЛЬНОСТЬ (F(1,19) = 7.930, p = 0.011), при этом слуховая задача
на всех этапах предъявления стимулов решалась испытуемыми более успешно, чем
зрительная. С другой стороны, для обеих модальностей выявлен основной эффект
фактора ЭТАП (F(2, 18) = 8.610, p = 0.002), успешность решения
когнитивной задачи возрастает по мере предъявления повторяющихся последовательностей
сенсорных сигналов. Парные сравнения выявили значимые различия между первым и
третьим (p = 0.001) и вторым и третьим (p = 0.020) этапами.
Таким образом и для параметра «точность» обнаружен эффект серийного
научения при решении сенсорных задач обеих модальностей. Эффект научения в этом
случае проявляется несколько позже, т.е. от второго к третьему этапу
предъявления последовательностей зрительных и слуховых сигналов.
По результатам анализа сессии с предъявлением повторяющихся
последовательностей слуховых и зрительных сигналов можно сделать два вывода:
в ходе проведенного эксперимента у испытуемых значимо возрастает
эффективность деятельности как по показателю ВР, так и по показателю
успешности, что дает возможность предполагать формирование имплицитного
предвосхищения появления сигналов разных модальностей в предстимульный период;
значимые изменения эффективности деятельности происходят при переходе от
первого ко второму (для ВР) или от второго к третьему (для успешности) этапам
предъявления сенсорных сигналов. Такой характер изменений параметров решения
когнитивных задач позволяет считать, что при предъявлении первых 10 проб
реакция испытуемого еще не обусловлена имплицитным научением и может
рассматриваться как реакция выбора, а при предъявлении последних 10 проб уже
происходит формирование имплицитного предвосхищения появления целевых сигналов.
.2 Анализ
влияния организации предстимульного периода на эффективность решения сенсорных
задач
Учитывая результаты предыдущего этапа анализа данных, для оценки влияния
организации предстимульного периода на эффективность решения сенсорных задач
показатели деятельности - ВР и успешность - сопоставлялись в трех
экспериментальных условиях: (1) реакция выбора без преднастройки (показатели
первого этапа сессии «серийное научение»), (2) направленное внимание,
обусловленное инструкцией-подсказкой (показатели сессии «направленное
внимание»), (3) имплицитное предвосхищение (показатели третьего этапа сессии
«серийное научение»). В каждом из условий для каждого испытуемого
подсчитывалось среднее время реакции и процент правильных ответов при решении
слуховой и зрительной задачи. Эти показатели служили переменной для
дисперсионного анализа GLM по
схеме MANOVA. В качестве внутрииндивидуальных
факторов использовались МОДАЛЬНОСТЬ (2 уровня: зрительная и слуховая) и УСЛОВИЕ
(3 уровня: реакция выбора, направленное внимание, имплицитное научение).
Время реакции
Зависимость времени реакции (ВР) от модальности целевых сигналов и
условий преднастройки представлен на рис. 9.
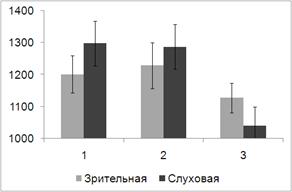
Рис. 9. Среднее по выборке испытуемых значение ВР (в мс) при решении
зрительной и слуховой задачи в разных экспериментальных условиях. 1 - реакция
выбора без преднастройки, 2 - направленное внимание, обусловленное инструкцией-подсказкой,
3 - имплицитное предвосхищение. Линии погрешности на графике отражают величину
ошибки среднего.
Дисперсионный анализ выявил основной эффект фактора УСЛОВИЕ (F(2, 18) = 6.549, p = 0.007). При этом как показали
парные сравнении влияние этого фактора преимущественно связано с различием ВР в
ситуации реакции выбора (1) и имплицитного научения (3). Различия между этими
двумя условиями описаны в предыдущем параграфе. Средние значения ВР в ситуации
направленного внимания меньше, чем в условиях реакции выбора (1287 мс и 1297 мс
соответственно), однако эти различия не значимы. Было также обнаружено значимое
взаимодействие факторов МОДАЛЬНОСТЬ и УСЛОВИЕ (F(2, 18) = 4.098, p = 0.034). Взаимодействие обусловлено наличием некоторого снижения ВР при
направленном слуховом внимании и отсутствием позитивного влияния на скорость
реакции условий направленного внимания при решении зрительной задачи.
Точность
При оценке точности выполнения сенсорной задачи из анализа были исключены
данные испытуемых, у которых невозможно было установить процент правильных
ответов в сессии «направленное внимание», поскольку в значительном числе
случаев испытуемые не давали ответ в отведенное время. На данном этапе были
проанализированы показатели точности 16 испытуемых.
Зависимость успешности решения когнитивной задачи (процента правильных
ответов) от условий преднастройки и модальности целевых стимулов представлена
на рис. 10.
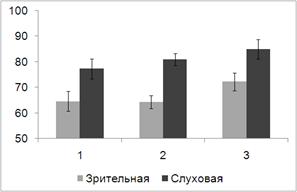
Рис. 10. Средний по выборке процент правильных ответов при решении
зрительной и слуховой задачи в разных экспериментальных условиях. 1 - реакция
выбора без преднастройки, 2 - направленное внимание, обусловленное
инструкцией-подсказкой, 3 - имплицитное предвосхищение. Линии погрешности на
графике отражают величину ошибки среднего
Дисперсионный анализ выявил основной эффект фактора МОДАЛЬНОСТЬ (F(1,15) = 12.663, p = 0.003), при этом слуховая задача
при всех экспериментальных условиях решалась испытуемыми более успешно, чем
зрительная. С другой стороны, выявлен основной эффект фактора УСЛОВИЕ (F(2, 14) = 5.861, p = 0.014), успешность решения
когнитивной задачи выше при направленном внимании (72.65%), чем при реакции
выбора (71%), и еще выше при имплицитном научении (78.67%). Однако значимые
различия выявлены только между реакцией выбора и имплицитным научением (p = 0.009). Позитивный эффект
направленного внимания был обнаружен только для слуховой задачи: в этом случае
средний процент решенных задач при реакции выбора был равен 77.3%, а при
направленном внимании 81%. При решении зрительной задачи эти два
экспериментальных условиях сопровождались практически одинаковым уровнем
точности выполнения сенсорной задачи (64.6% и 64.3%).
Таким образом, в условиях данной экспериментальной модели позитивный
эффект направленного внимания хотя и наблюдался, но не был значимым. Кроме
того, он наблюдался только при решении слуховой задачи. В этом случае он
касался как времени реакции, так и точности выполнения задания.
.3 Анализ
мозговой организации предстимульного модально-специфического внимания при
произвольной преднастройке и серийном научении
Влияние условий преднастройки на функциональные связи лобных
и каудальных областей коры
Учитывая литературные данные (глава 1) о мозговой организации
направленного внимания представлялось целесообразным проанализировать влияние
избирательной преднастройки на функциональное взаимодействие фронтальных и
каудальных зон коры. Для этого был проведен дисперсионный анализ в парах
областей, объединяющих фронтальную глазодвигательную область (FEF-V1, FEF-IPC,
FEF-LIP), а также латеральную префронтальную кору с затылочными зонами и
теменными областями (LPFC-V1, LPFC-IPC, LPFC-LIP).
Для выявления особенностей функциональной организации избирательного
внимания, направляемого инструкцией-подсказкой проводилось сопоставление
величины когерентности сигналов ЭЭГ в диапазоне альфа-ритма в предстимульный
период, предшествующий появлению целевого сигнала, и в период мобилизационной
готовности после появление фиксационной точки и до появления предупреждающего
стимула. Статистическое сравнение проводилось по схеме MANOVA: УСЛОВИЕ (1 - мобилизационная готовность, 2 -
зрительное направленное внимание, 3 - слуховой направленное внимание) ´ ПОЛУШАРИЕ (1 - левое, 2 - правое) ´ ЛОКАЛИЗАЦИЯ (этот фактор зависит от
конкретных анализируемых пар ЭЭГ сигналов в разных областях коры).
При анализе изменений функционального взаимодействия в парах областей с
участием FEF был обнаружен основной эффект
фактора УСЛОВИЕ - F(2,8) = 24.644, p < 0.0001. Сравнение средних
значений КОГ (рис. 11) свидетельствует о росте функционального взаимодействия в
анализируемых парах отведений при избирательном направленном внимании по
сравнению с мобилизационной готовностью. Сравнение каждого типа
модально-специфического направленного внимания с мобилизационной готовностью
выявило значимые различия для обеих модальностей (F(1,9) = 28.073, p <0.0001 - для зрительной модальности и F(1,9) = 36.697, p <0.0001).
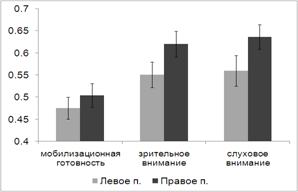
Рис. 11. Средние значения альфа КОГ предстимульной ЭЭГ в парах областей с
«фокусом» в FEF при направленном зрительном и
слуховом внимании и мобилизационной готовности
Для обеих модальностей отмечаются особенности роста функционального
взаимодействия FEF c теменными ассоциативными зонами в
правом и левом полушариях (рис. 12). В левом полушарии максимальный рост
отмечается для связи FEF с IPC, а в правом полушарии для связи с LIP. Межполушарные различия динамики
функционального взаимодействия при направленном внимании были больше выражены
при подготовке к решению слуховой задачи, что отразилось в значимом
взаимодействии УСЛОВИЕ´ПОЛУШАРИЕ ´ЛОКАЛИЗАЦИЯ (F(2,8) = 5.979, p = 0.026).
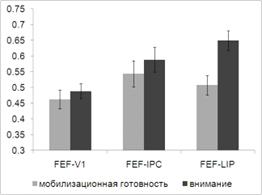
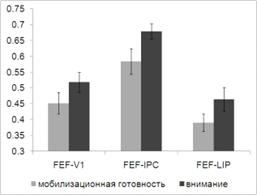
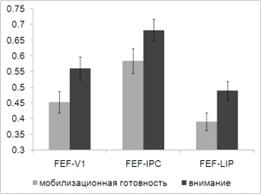
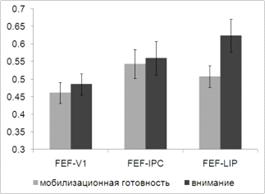
Рис. 12. Средние значения альфа КОГ предстимульной ЭЭГ в парах областей с
«фокусом» в FEF при направленном внимании. Верхний
ряд - слуховая задача, нижний ряд - зрительная задача, слева - левое полушарие,
справа - правое полушарие.
Для связей с фокусом в LPFC
также выявлено значимое влияние фактора УСЛОВИЕ (F(2,8) = 18.779, p = 0.001). Функциональные связи латеральной префронтальной коры при
избирательном внимании также оказались выше, чем при мобилизационной готовности
(рис. 13), что отразилось в значимом влияние основного эффекта фактора УСЛОВИЕ F(2,8) = 18.779, p = 0.001). Рост КОГ отмечался для
обеих модальностей (F(1,9) = 31.894, p <0.0001 для зрительной
модальности, F(1,9) = 20.741, p = 0.001 - для слуховой модальности).
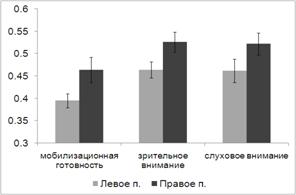
Рис.13. Средние значения альфа КОГ предстимульной ЭЭГ в парах областей с
«фокусом» в LPFC при направленном зрительном и
слуховом внимании и мобилизационной готовности
При сопоставлении избирательного внимания с мобилизационной готовностью
как для зрительной, так и для слуховой модальности был выявлен основной эффект
фактора ПОЛУШАРИЯ (F(1,9) = 12.868, p = 0.006 - для зрительной задачи, F(1,9) = 13.039, p = 0.006 - для слуховой задачи), при
этом КОГ в правом полушарии была выше, чем в левом полушарии.
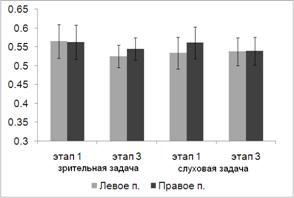
Рис. 14. Средние значения альфа КОГ предстимульной ЭЭГ в парах областей с
«фокусом» в FEF на разных этапах серийного научения
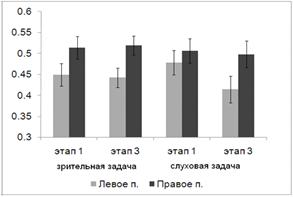
Рис. 15. Средние значения альфа КОГ предстимульной ЭЭГ в парах областей с
«фокусом» в FEF LPFC на разных этапах серийного научения
Предстимульное функциональное взаимодействие лобных и каудальных отделов
коры на первом и третьем этапах сессии серийное научение сопоставлялось также с
функциональным взаимодействием этих отделов коры при мобилизационной готовности
отдельно для каждого этапа и каждой сенсорной задачи.
Для зрительной задачи при сопоставление КОГ в парах отведений с
фокусом в FEF в условиях ЭУ_СН1 и ЭУ_МГ было
выявлено значимое влияние фактора условие (F(1,9) = 5.888, p =
0.038. При сопоставлении КОГ в условиях ЭУ_СН3 и ЭУ_МГ основной эффект фактора
УСЛОВИЕ не был обнаружен. Однако было выявлено значимое взаимодействие
УСЛОВИЕ*ПОЛУШАРИЕ*ЛОКАЛИЗАЦИЯ (F(2,8) = 6.932,p = 0.018)
Сравнения КОГ в отдельных парах отведений правого и левого полушария
показало, что на третьем этапе сессии серийное научение при решении зрительной
задачи степень функционального взаимодействия в паре областей FEF-LIP правого полушария с учетом поправки на множественные
сравнения (3 пары областей в каждом полушарии) значимый (p < 0.05/3=0.017) была выше, чем
при мобилизационной готовности
(F(1,9) = 10.404, p = 0.010)
Для пар отведений с фокусом в LPFC при сравнении предстимульной КОГ как на первом, так и на третьем этапе
серийного научения с КОГ в ситуации мобилизационной готовности было обнаружено
значимое влияние фактора УСЛОВИЕ: для первого этапа научения
F(1,9)
= 13.036, p = 0.006; для третьего - F(1,9) = 5.729, p = 0.040
При этом в обоих случаях КОГ была значимо выше в период, предшествующий
появлению целевого сигнала в сессии серийного научения, чем при мобилизационной
готовности. Сравнение КОГ в отдельных парах областей показало значимые различия
в паре LPFC-V1 правого полушария (F(1, 9) = 8.099, p = 0.016).
Для слуховой задачи при сопоставление КОГ в парах отведений с фокусом в FEF в условиях ЭУ_СН1 и ЭУ_МГ не было
обнаружено влияния фактора УСЛОВИЕ. При сопоставлении КОГ в условиях ЭУ_СН3 и
ЭУ_МГ было выявлено взаимодействие
УСЛОВИЕ*ПОЛУШАРИЕ*ЛОКАЛИЗАЦИЯ на уровне тенденции
(F(2,8) = 4.193, p = 0.06)
Сравнения КОГ в отдельных парах отведений правого и левого полушария
показало, что на третьем этапе сессии серийного научения при решении слуховой
задачи степень функционального взаимодействия в паре областей FEF-LIP правого полушария так же как при решении зрительной
задачи значимо выше, чем при мобилизационной готовности (F(1,9) =10.449, p = 0.010).
Для пар отведений с фокусом в LPFC при сравнении предстимульной КОГ в сессии серийного научения с КОГ в
ситуации мобилизационной значимое влияние фактора УСЛОВИЕ было выявлено только
для первого этапа сессии F(1,9)
= 8.229, p = 0.019. При этом КОГ была значимо
выше в период, предшествующий появлению целевого сигнала в сессии серийное
научение, чем при мобилизационной готовности.
В соответствии с задачей исследования представляло интерес провести
сопоставление анализируемых параметров ЭЭГ при избирательном внимании и
имплицитном предвосхищении (третий этап сессии серийного научения). В этом
случае в схеме MANOVA (см. выше)
учитывались два варианта экспериментальных условий - условие ЭУ_НВ из первой
сессии и условие ЭУ_СН3 из второй сессии. В целом уровень функционального
взаимодействия фронтальных корковых зон с затылочными и теменными
ассоциативными зонами оказался выше при направленном предстимульном внимании,
чем при имплицитном предвосхищении (рис. 16, 17), однако эти различия не
достигали уровня значимости. Было обнаружено взаимодействие факторов УСЛОВИЕ,
МОДАЛЬНОСТЬ и ПОЛУШАРИЕ для пар областей с фокусом в FEF на уровне тенденции (F(1,9) = 4.317, p =
0.068). При этом различия между КОГ в условиях внимания и имплицитного
предвосхищения для зрительной модальности были больше выражены в левом
полушарии, а для слуховой - в правом (рис. 16, 17).
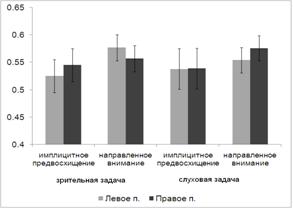
Рис. 16. Средние значения альфа КОГ предстимульной ЭЭГ в парах
областей с «фокусом» в FEF в
условиях имплицитного предвосхищения и направленного внимания
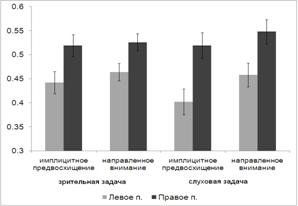
Рис. 17. Средние значения альфа КОГ предстимульной ЭЭГ в парах областей с
«фокусом» в LPFC в условиях имплицитного
предвосхищения и направленного внимания
Влияние условий преднастройки на функциональные связи
сенсорно-специфических и ассоциативных зон
Для выявления модально-специфических перестроек функциональных корковых
связей при направленном внимании параметры КОГ ЭЭГ в диапазоне альфа-ритма
сопоставлялись в экспериментальных условиях ЭУ_МГ и ЭУ_НВ для пар областей,
объединяющих сенсорно-специфические зоны коры (V1 и А1) с фронтальными (FEF и LPFC)
и теменными (LIP и IPC) зонами, а также для пар областей, связывающие
сенсорно-специфические зоны между собой для каждой модальности в левом и правом
полушарии отдельно. С учетом поправки на множественные сравнения (8 пар
областей в каждом полушарии) значимый (p<0.05/8=0.0062), рост альфа КОГ был выявлен для зрительной
модальности в парах V1-A1 и V1-FEF левого
полушария (t(9) = 5.255, p = 0.001 и t(9) =
4.163, p =0.002 соответственно). При этом
направленное внимание сопровождалось ростом КОГ по сравнению с ситуацией мобилизационной
готовности (mean 0.496 ± 0.15, mean = 0.391 ± 0.06 для мобилизационной
готовности и mean 0.65 ± 0.15, mean 0.471 ± 0.006 для направленного
зрительного внимания). Для слуховой модальности значимые различия альфа-КОГ
также наблюдались в парах областей левого полушария V1-A1 и V1-LPFC (t(9) =
3.888, p = 0.004 и t(9) = 3.862, p =
0.004 соответственно). Также как для зрительной задачи направленное внимание
сопровождалось ростом функционального взаимодействия по альфа-ритму в этих
корковых зонах (mean 0.496 ± 0.15,
mean = 0.391 ± 0.06 для мобилизационной
готовности и mean 0.604 ± 0.13, mean 0.472 ± 0.009 для направленного
слухового внимания). Значимых различий КОГ с учетом поправки на множественные
сравнения в правом полушарии в парах областей, объединяющих
сенсорно-специфические зоны с ассоциативными выявлено не было.
Аналогичный анализ был проведен для сессии серийное научение. В этом
случае параметры альфа-КОГ сопоставлялись в экспериментальных условиях ЭУ_СН1 и
ЭУ_СН3. При этом не было установлено значимого роста альфа-КОГ в условии ЭУ_СН1
по сравнению с ЭУ_СН3 для всех вышеупомянутых пар отведений. В тех же парах
областей сопоставлялись параметры альфа КОГ предстимульной ЭЭГ на первом и
третьем этапе серийного научения с параметрами КОГ в ситуации мобилизационной
готовности. При этом сопоставление параметров альфа-КОГ в ЭУ_СН1 (mean = 0.625 ± 0.14) и ЭУ_МГ (mean = 0.496 ± 0.16) выявило значимые
различия для слуховой модальности в паре областей левого полушария V1-A1 (t(9) =
-5.149, p = 0.001).
Таким образом, электрофизиологический анализ позволил обнаружить
существенные различия мозговой организации предстимульного
модально-специфического внимания при произвольной преднастройке и серийном
научении. Предстимульный период произвольного избирательного внимания,
задаваемого стимулом-подсказкой сопровождался ростом функционального
взаимодействия по альфа-ритму между дорзальными и латеральными зонами
префронтальной коры и теменными и затылочными областями по сравнению с
неспецифической мобилизационной готовностью, предшествующей появлению
стимула-подсказки. При периодическом повторении последовательности чередования
слуховых и зрительных задач на стадии уже сформированного имплицитного научения
вовлечение фронто-париетальных модулирующих систем коры в процесс подготовки к
анализу целевых стимулов выражено существенно меньше. В особенности это
касается слуховой задачи, которая характеризуется наиболее существенным
сокращением времени реакции и повышением точности решения сенсорных задач в
ходе серийного научения.
Глава 4.
Обсуждение
На основании анализа поведенческих результатов было установлено, что к
третьему этапу выполнения заданий в сессии серийное научение по мере повторения
последовательностей слуховых и зрительных сигналов происходит сокращение ВР при
решении задач обоих модальностей, что свидетельствует о наличии эффекта
серийного научения по этому параметру деятельности. Этот эффект был больше
выражен при решении слуховой задачи. Для параметра «точность» также был
обнаружен эффект серийного научения при решении сенсорных задач обеих
модальностей, при этом слуховые задачи опять решались более успешно. В целом
результаты анализа параметров деятельности в этой сессии позволяют считать, что
у испытуемых действительно формировалось имплицитная преднастройка к анализу
сенсорных сигналов, что и способствовало более эффективному решению когнитивной
задачи.
Сравнение времени реакции в трех экспериментальных условиях (направленное
внимание, имплицитная преднастройка и реакция выбора) показало, что предъявление
предупреждающего стимула-подсказки оказывает положительный эффект (по сравнению
с реакцией выбора) как на точность, так и на скорость решения когнитивной
задачи. Однако, это эффект оказался статистически незначимым и зависящим от
модальности сенсорного сигнала. Он наблюдался только для слуховых стимулов.
Значимых различий точности и скорости решения когнитивной задачи при сравнении
двух условий предстимульного внимания между собой выявлено не было.
В сессии с повторяющейся последовательностью чередования слуховых и
зрительных стимулов, было обнаружено существенное повышение эффективности
деятельности в процессе серийного научения. Точность и скорость ответов
испытуемых в этом случае может возрастать в связи со снижением уровня
неопреденности при принятии решения в процессе повторения событий и переходом к
автономной, по классификации M. Posner и P. Fitts [1976],
стадии решения когнитивных задач. Согласно концепции этих авторов выполнение
повторяющихся заданий, включающих двигательный компонент, проходит несколько
стадий. На первой стадии (когнитивной) происходит ориентировка в задании,
произвольное следование инструкции, коррекция ошибок на основе обратной связи.
Выполнение задания на этой стадии характеризуется большим количеством ошибок,
нестабильностью и вариабельностью. На второй стадии (ассоциативной) происходит
«оттачивание» выполнения на основе установления связей между внешними событиями
и соответствующей двигательной реакцией. На третьей стадии (автономной) роль
внимания существенно падает, деятельность становится менее произвольной,
двигательные реакции более точными, вариабельность ответов снижается. Скорость
реакции в сессии серийное научение может также возрастать по мере формирования
моторного навыка, обусловленного большим количеством выполняемых в ходе этой
сессии двигательных ответов (240 (80×3)).
По результатам анализа поведенческих данных были выявлены особенности
выполнения заданий разной модальности: как в сессии направленное внимание, так
и в сессии серийное научение слуховые задачи решались более успешно, чем
зрительные. Для сессии направленное внимание эффект модальности может быть
связан с особенностями экспериментальной модели. Особенность в данном случае
заключается в том, что модальность стимулов-подсказок при зрительной задаче совпадала
с модальностью целевых стимулов, а при слуховой не совпадала. Возможно, при
совпадении модальности предупреждающего и целевого стимулов опознание
предупреждающего стимула как самостоятельная когнитивная задача в большей
степени интерферирует со следующей вслед за ней основной задачей различения
целевых стимулов, чем в тех случаях, когда модальности предупреждающего и
целевого сигналов не совпадают. Это вероятно и объясняет менее выраженный
позитивный эффект предстимульного внимания в зрительной задаче. В сессии
серийное научение меньшая выраженность позитивных изменений скорости и точности
решения зрительной задачи могла быть обусловлена зрительным утомлением, что
отмечали и сами испытуемые.
В целом наличие статистически значимых различий показателей эффективности
деятельности при имплицитном предвосхищении и реакции выбора, а также наличие
позитивного эффекта направленного внимания по отношению к реакции выбора
говорят в пользу обоснованности исследования влияния условий преднастройки на
мозговую организацию предстимульного модально-специфического внимания.
Анализ ЭЭГ позволил обнаружить разную мозговую организацию
предстимульного модально-специфического внимания в условиях произвольной
преднастройки, задаваемой инструкцией, и в условиях серийного научения.
Согласно полученным в настоящем исследовании результатам, при произвольном
предстимульном внимании как зрительном, так и слуховом возрастают
функциональные связи между префронтальными (FEF и LPFC)
и каудальными (V1, IPC, LIP) зонами коры (рис. 18).
Эти изменения можно трактовать как активацию лобных управляющих систем
мозга, осуществляющих нисходящие (top-down) модулирующие влияния на
сенсорно-специфические зоны. Нисходящие модулирующие влияния подготавливают
сенсорные системы к анализу целевых стимулов. Данные о росте степени
функционального взаимодействия между дорзальными и латеральными зонами
префронтальной коры и теменными ассоциативными областями согласуются с
результатами исследований других авторов, большинство из которых указывают на
роль фронто-париетальных систем в механизмах произвольного внимания и удержания
значимой информации в рабочей памяти в процессе решения когнитивных задач [Desimone and Duncan, 1995; Kastner and Ungerleider, 2000; Corbetta and Shulman, 2002; Moore et al., 2003; Serences and Yantis,2006; Bressler et al, 2008; Knudsen, 2007, см. главу 1].
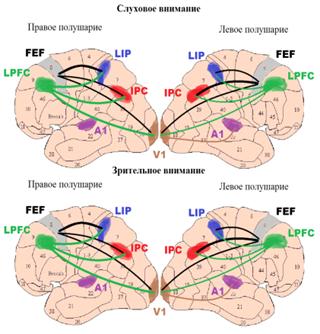
Рис. 18. Функциональная организация предстимульного
модально-специфического внимания при произвольной преднастройке
Линиями соединены зоны коры, для которых выявлен значимый рост
функционального взаимодействия при ожидании целевого сигнала по сравнению с
мобилизационной готовностью
Участие нейронов префронтальных (FEF и LPFC)
и теменных (LIP) зон коры в избирательной настройке
мозга при направленном предстимульном внимании продемонстрировано в
исследования на приматах [Bushman,
Miller, 2007, 2009]. Согласно результатам
проведенного исследования вовлечение фронто-париетальных корковых связей в
процессе подготовки к анализу значимой информации было более выраженным в
правом полушарии. Это согласуется с литературными данными о преимущественном
участии дорзолатеральной префронтальной коры правого полушария в механизмах
поддержания внимания и избирательного когнитивного контроля [Posner, Petersen, 1990; Gazzaley et al., 2004; Мачинская и соавт., 2012].
Анализ динамики функциональных связей с участием сенсорно-специфических
зон коры позволил обнаружить рост функционального взаимодействия между
зрительной и слуховой корой в предстимульный период направленного внимания как
при решении зрительных, так и при решении слуховых задач. Следует отметить, что
изменения связей между сенсорно-специфическими зонами коры отмечались только в
левом полушарии. Эти наблюдения согласуются с данными исследования Р.И. Мачинской
и соавт. [1992] об особенностях функциональной организации левого и правого
полушария при избирательном модально-специфическом внимании. В этом
исследовании было показано, что в предстимульный период направленного
модально-специфического внимания в левом полушарии растет степень
функционального взаимодействия корковых зон с фокусом в сенсорно-специфических
областях, тогда как в правом полушарии изменения касаются преимущественно
дистантных связей между ассоциативными зонами коры.
Активация фронто-париетальных систем внимания, выявленная при
произвольной преднастройке, была существенно меньше выражена в предстимульный
период, предшествующий решению сенсорных задач в условиях сформированного серийного
научения (рис. 19).
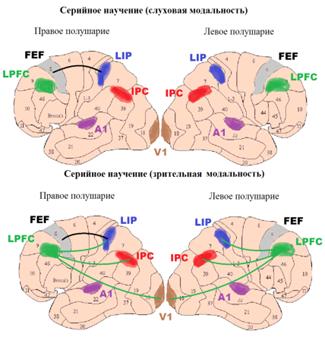
Рис. 19. Функциональная организация предстимульного внимания при серийном
научении
Линиями соединены зоны коры, для которых выявлен значимый рост
функционального взаимодействия в предстимульный период на стадии имплицитного
научения по сравнению с мобилизационной готовностью.
В этой экспериментальной ситуации отсутствовал рост функционального
взаимодействия корковых зон в дорзальной фронто-париетальной модулирующей
системе. В латеральной фронто-париетальной системе увеличение степени
функционального взаимодействия корковых зон было отмечено только для зрительной
задачи. Эти данные могут рассматриваться как нейрофизиологическое подтверждение
высказанного выше предположения о переходе когнитивной деятельности в
автономную фазу в результате серийного научения. Менее выраженное вовлечение
лобных зон коры в подготовку к решению зрительной задачи и отсутствие значимых
перестроек электрической активности этих зон при решении слуховой задачи
говорят о формировании в ходе серийного научения более экономных, специфичных и
автономных функциональных систем обработки информации и принятия решений, не
требующих дополнительного участия управляющих структур мозга.
В ходе данного исследования не удалось выявить функциональные системы,
формирование которых обеспечивает подготовку к решению когнитивной задачи на
этапе сформированного серийного научения. По-видимому, для решения этого
вопроса необходимо включить в электофизиологический анализ помимо уже
исследованных префронтальных, теменных и сенсорно-специфических областей зон
коры те области мозга, которые связаны с моторными компонентами серийного
научения. Согласно исследованию O.
Hikosaka [2002] мозговые системы, которые обеспечивают моторные компоненты
серийного научения, включают в себя дополнительную моторную кору (supplementary motor area - SMA) и
кору, расположенную ростральнее этой области (presupplementary motor area - preSMA).
Вопросы, требующие дополнительных исследований и возможные
подходы к их решению:
Какое влияние оказывает количество решаемых в ходе эксперимента задач на
функциональную организацию предстимульного внимания? Этот вопрос возникает в
связи с тем, что в данном исследовании количество задач было разным в сессиях
направленное внимание и серийное научение, и это могло повлиять на результаты
выполнения заданий. Для исследования этого вопроса необходимо уравнять
количество предъявлений целевых стимулов в этих экспериментальных условиях
Каково влияние модальности предупреждающего стимула? Для решения этого
вопроса необходимо провести аналогичное исследование направленного внимания с
использованием слухового стимула в качестве стимула-подсказки
Какова роль моторных и премоторных зон коры в подготовке мозга к решению
когнитивной задаче при серийном научении? Для исследования этого вопроса
следует включить эти области коры в электрофизиологический анализ
внутрикоркового взаимодействия.
Выводы:
1. Обнаружено позитивное влияние направленного внимания и имплицитного
предвосхищения в ходе серийного научения на скорость и точность решения
сенсорных задач. Позитивный эффект серийного научения в целом был более
выражен, чем эффект направленного внимания. При этом слуховая задача решалась
испытуемыми успешнее, чем зрительная, как в условиях направленного внимания,
так и при серийном научении.
2. Функциональная организация коры головного мозга в период
предстимульного модально-специфического внимания при произвольной преднастройке
и серийном научении существенно различается:
- В предстимульный период направленного избирательного внимания
возрастает степень функционального взаимодействия во фронто-париетальных
дорзальной и латеральной модулирующих системах при решении сенсорных задач
обеих модальностей. Рост функционального взаимодействия в латеральной
фронтально-париетальной системе в условии направленного произвольного внимания
более выражен в правом полушарии. Функциональные связи между
сенсорно-специфическими областями (затылочной и височной) наблюдаются только в
левом полушарии.
На этапе сформированного имплицитного научения в предстимульный
период не выявлен рост функционального взаимодействия корковых зон в дорзальной
фронто-париетальной модулирующей системе. В латеральной фронто-париетальной
системе увеличение степени функционального взаимодействия корковых зон отмечено
только для зрительной задачи.
. Основная особенность функциональной организации предстимульного периода
при серийном научении состоит в меньшем вовлечении фронтальных систем
нисходящего контроля. Можно предположить, что серийное научение способствует
формированию более экономных специализированных и автономных функциональных
систем, настраивающих мозг на обработку информации и принятие решения, которые
не требуют дополнительного контроля со стороны лобных управляющих структур.
Список
литературы
1. Выготский,
Л.С. Проблема развития и распада высших психических функций / Л.С. Выготский //
Развитие высших психических функций. М.: АПН РСФСР. - 1960. -364-383 c. Докл. на конф. Ин-та эксперимент.
медицины. 28 апреля 1934 г.
2. Гальперин,
П.Я. К проблеме внимания / П.Я. Гальперин // Доклады АПН РСФСР. - 1958. - N3
. Гусельников,
В.И. Электрофизиология головного мозга / В.И. Гусельников // М.: Высшая школа.
- 1976. - 211 с.
. Дормашев,
Ю.Б. Психология внимания / Ю.Б. Дормашев, В.Я. Романов// М.: Тривола. 1995. -
352 с.
. Курганский,
А.В. Механизмы формирования координационной структуры серийных движений у
взрослых и детей. Диссертация. Наук / А.В. Курганский // Диссертация докт.
биол. наук. М. - 2014. - 391 с.
. Курганский,
А.В. Некоторые вопросы исследования кортико-кортикальных функциональных связей
с помощью векторной авторегрессионной модели многоканальной ЭЭГ / А.В.
Курганский // ЖВНД. - 2010. - Т. 60. - N5.
. Курганский,
А.В. Фронтальные билатерально-синхронные теат-волны на ЭЭГ детей 7-8 лет с
трудностями обучения: качественный и количественный анализ/А.В. Курганский,
Р.И. Мачинская // Физиология человека. - 2012. - Т. 38. - N3. - 37-47 с.
. Мачинская,
Р.И. Нейрофизиологические механизмы произвольного внимания (Аналитический
обзор) / Р.И. Мачинская // Журнал высшей нервной деятельности. - 2003. - Т.53.
- N2. - 133-150 c.
. Мачинская,
Р.И. Сравнительное электрофизиологическое исследование регуляторных компонентов
рабочей памяти у взрослых и детей 7-8 лет. Анализ когерентности ритмов ЭЭГ /
Р.И. Мачинская, А.В. Курганский// Физиология человека. - 2012. - Т. 38. - N1. - 5-19 c.
10. Мачинская, Р.И. Формирование нейрофизиологических механизмов
произвольного избирательного внимания у детей младшего школьного возраста.
Диссертация / Р.И. Мачинская // Диссертация докт. биол. наук. М. - 2001.
11. Мачинский, Н.О. Электрофизиологическое исследование функциональной
организации мозга при направленном внимании. Сообщение 1. Взрослые в норме /
Н.О. Мачинский, Р.И Мачинская, В.Д. Труш // Физиология человека. - 1990. - 5-16
c.
. Мачинский, Н.О. Электрофизиологическое исследование
функциональной организации мозга при направленном внимании. Сообщение 2. Дети
11-13 лет в норме / Н.О. Мачинский, Р.И Мачинская, В.Д. Труш // Физиология
человека. - 1990. - 5-13 c.
13. Наатен,
Р. Внимание и функция мозга: Учеб. Пособие / Пер. с англ. Под ред. Е.Н.
Соколова / Р. Наатен // М.: Изд-во МГУ. - 1998. - 560 с.
14. Рубинштейн,
С.Л. Основы общей психологии / С.Л. Рубинштейн // СПб. - 1998.
. Фарбер,
Д.А. Функциональная организация мозга в период подготовки к опознанию
фрагментарных изображений / Д.А. Фарбер, Р.И. Мачинская, А.В. Курганский, Н.Е.
Петренко // ФГНУ «Институт возрастной физиологии РАО». - 2014. - 25 с.
16. Anderson,
J.R. Cognitive psychology and its implications (6th ed.)/J.R. Anderson // Worth
Publishers. - 2004. - P.519
. Bollinger,
J. Expectation-driven changes in cortical functional connectivity influence
working memory and long-term memory/J. Bollinger, M.T. Rubens, T.P. Zanto, A.
Gazzaley // J. Neurosci. - 2010. - P. 14399-14410.
. Bressler,
S.L. Top-down control of human visual cortex by frontal and parietal cortex in
anticipatory visual spatial attention / S.L. Bressler, W. Tang, C.M. Sylvester,
G.L. Shulman, M. Corbetta // J. Neurosci. - 2008. - P. 10056-10061.
20. Brunia,
C.H. Neural aspects of anticipatory behavior / C.H. Brunia//ActaPsychol.
(Amst). - 1999. - V. 101. - P. 213-242.
21. Bullock,
D. Adaptive neural models of queuing and timing in fluent action/ D.
Bullock//Trends Cogn Sci. - 2004. - P. 426-433.
22. Burgess,
P.W., Brain regions involved in prospective memory as determined by positron
emission tomography/ P.W. Burgess, A. Quayle, C.D. Frith // Neuropsychologia. -
2001. - V. 39. - P. 545-555.
23. Buschman,
T.J. Top-down versus bottom-up control of attention in the prefrontal and
posterior parietal cortices/T.J. Buschman, E.K. Miller//Science. - 2007. - P:
1860-1862.
. Buschman,
T.J. Serial, сovert, shifts of attention
during visual search are reflected by the frontal eye fields and correlated
with population oscillations / T.J. Buschman, E.K. Miller//Neuron. - 2009. - P.
386-396.
. Cleeremans,
A. Implicit learning: News from the front / A. Cleeremans, A. Destrebecqz, M.
Boyer//Trends in cognitive psychology. - 1998. - P. 406-416.
. Corbetta
M. Control of goal-directed and stimulus-driven attention in the brain//M.
Corbetta, G.L. Shulman/Nat Rev Neurosci. - 2002. - P. 201-215.
27. Coull,
J.T. Neural correlates of attention and arousal: insights from
electrophysiology, functional neuroimaging and psychoparmacology / J.T.Coull //
Progress in Neurobiology. - 1998. - V. 55. - P.343-361.
28. Cui,
J. BSMART: A Matlab / C toolbox for analysis of multichannel neural time series
/ J. Cui, L. Xu, S.L. Bressler, M. Ding, H. Liang // Neural Networks. - 2008 -
P. 1094-1104.
. Desimone,
R. Neural mechanisms of selective visual attention / R. Desimone, J. Duncan //
Annu Rev Neurosci. - 1995. - P. 193-222.
. Doyon,
J. Role of the striatum, cerebellum, and frontal lobes in the learning of a
visuomotor sequence / J. Doyon, D. Gaudreau, R. Laforce, M. Castonguay, P.J.
Bedard, F. Bedard, J.-P. Bouchard//Brain and Cognition. - 1997. - P. 218-245.
31. Eickhoff,
S.B. A new SPM toolbox for combining probabilistic cytoarchitectonic maps and
functional imaging data / S.B. Eickhoff, K. E. Stephan, H. Mohlberg, C.
Grefkes, G. R. Fink, K. Amunts, K. Zilles // Neuroimage. - 2004. - P.
1325-1335.
. Fitts,
P.M. Human performance / P.M. Fitts, M.I. Posner//Belmont. CA: Brook-Cole. -
1967. - P.162.
33. Gazzaley,
A. Functional connectivity during working memory maintenance/ A. Gazzaley, J.
Rissman, M. D’esposito // Cognitive, Affective, & Behavioral Neuroscience.
- 2004. - P.580-599.
. Grech,
R. Review on solving the inverse problem in EEG source analysis/ R. Grech, T.
Cassar, J. Muscat, K.P. Camilleri, S.G. Fabri, M. Zervakis, P. Xanthopoulos, V.
Sakkalis, B. Vanrumste//J Neuroeng Rehabil. - 2008. - P. 25.
. Grossberg,
S. Behavioral contrast in short term memory: serial binary memory models or
parallel continuous memory models? / S. Grossberg//J Math Psychol. - 1978. -
P.199-219.
. Hahn,
E. Test-retested reliability of attention network test measures in schizophrenia
/ E. Hahn, T.M. Ta, C. Hahn, L.K. Kuehl, C. Ruehl, A.H. Neuhaus, M. Dettling //
Schizophrenia Research. - 2011. - V. 133. - P. 218-222.
. Hallez,
H. Review on solving the forward problem in EEG source analysis/H. Hallez, B.
Vanrumste, R. Grech, J. Muscat, W. De Clercq, A. Vergult, Y. D'Asseler, K.P.
Camilleri, S.G. Fabri, S. Van Huffel, I. Lemahieu // J. Neuroeng Rehabil. -
2007. - P. 46.
. Hikosaka,
O. Central mechanisms of motor skill learning/O. Hikosaka, K. Nakamura, K.
Sakai, H. Nakahara // Curr Opin Neurobiol. - 2002. - P. 217-222.
. James,
W. Principles of psychology/W. James // New York: Holt
. Kahnemann,
D. Changing views of attention and automatic/ D. Kahnemann, A. Traismann //
Parasuraman R., Davies D.R. (Eds). Varieties of attention. New York: Academic
Press. - 1984. - P. 29-61.
. Kastner,
S. Mechanisms of visual attention in the human cortex/S. Kastner, L.G. Ugleiter
// Annu Rev Neurosci. - 2000. - P. 315-341.
. Knudsen,
E.I. Fundamental components of attention/E.I. Knudsen // Annu Rev Neurosci. -
2007. - P. 57-78.
. Kushner,
M. Implicit detection of event interdependencies and a PDP model of the process
/ M. Kushner, A. Cleeremans, A.S. Reber // Paper presented at the Proceedings
of the Thirteenth Annual Conference of the Cognitive Science Society. - 1991.
. Landau,
S.M. Sequence learning in pianists and non-pianists: An fMRI study of motor
expertise/S.M. Landau, M. D’Esposito // Cognitive, Affective, and Behavioral
Neuroscience. - 2006. - P. 246-259.
45. Lashley,
K.S. The problem of serial order in behavior / K.S. Lashley // Jeffress L.A.
(Ed.) New York: Wiley. Cerebral mechanisms in behavior. - 1951. - P. 112-131.
. Lewicki,
P. Acquisition of procedural knowledge about a pattern of stimuli that cannot
be articulated/ P. Lewicki, T. Hill, E. Bizot // Cognit. Psychol. - 1988. -
N20. - P. 24-37
47. Marzinzik,
F. The human thalamus is crucially involved in executive control operations/F.
Marzinzik, M. Wahl, G.H. Schneider, A. Kupsch, G. Curio // J. Cogn. Neurosci. -
2008. - P. 1903-1914.
. McAlonan,
K. Thalamic reticular nucleus activation reflects attentional gating during
classical conditioning / K. McAlonan, V.J. Brown, E.M. Bowman//J. Neurosci. -
2000. - P. 8897-8901.
. Moore,
T. Selective gating of visual signals by microstimulation of frontal cortex /
T. Moore, K.M. Armstrong // Nature. - 2003. - P. 370-373.
. Moore,
T. Visuomotor origins of covert spatial attention/T. Moore, K.M. Armstrong, M.
Fallah // Neuron. - 2003. - P. 671-683.
. Nissen,
M.J. Attentional requirement of learning: evidence from performance measures /
M.J. Nissen, P. Bullemer // Cognit. Psychol. - 1987. - N19. - P. 1-32.
. Pascual-Marqui,
R.D. Low resolution electromagnetic tomography: a new method for localizing
electrical activity in the brain / R.D. Pascual-Marqui, C.M Michel, D. Lehmann
// International Journal of Psychophysiology. - 1994. - P. 49-65.
. Porjesz,
В. The effects of stimulus
expectancy on brain evoked potentials/ В. Porjesz, H. Begleiter // Psychophysiology. - 1975. -V. 12. - P.
152-157.
. Posner,
M.I. Attention as an organ system/M.I. Posner, J. Fan // Topics in integrative
neuroscience: from cells to cognition Pomerantz J.R. - Cambridge: Cambridge
University Press. - 2008. - P. 31-62.
55. Posner,
M.I. Attention in cognitive neuroscience: An overview in M.S. Gazzaniga (Ed.),
Cognitive neurosciences / M.I. Posner // Cambridge, MA: MIT Press. - 1995. - P.
615-624.
56. Posner,
M.I. Chronometric exploration of mind // M.I. Posner // Hillsdale, NJ: Lawrence
Erlbaum Associates
. Posner,
M.I. Cognitive neuroscience of attention / M.I. Posner // The Guilford Press. -
2004. - P. 466.
58. Posner,
M.I. The attention system of the human brain / M. I. Posner, S.E.
Petersen//Annu Rev Neurosci. - 1990. - P. 25-42.
. Rauch,
S.L. Striatal recruitment during an implicit sequence learning task as measured
by functional magnetic resonance imaging/S.L. Rauch, P.J. Whalen, C.R. Savage,
T. Curran, A. Kendrick, H.D. Brown, G. Bush, H.C. Breiter, B.R. Rosen // Hum
Brain Map. - 1997. P. 124-132.
. Reber,
A.S. Implicit learning and tacit knowledge: An Essay on the Cognitive
Unconscious / A.S. Reber // Oxford University Press. - 1993.
61. Reber,
A.S. Implicit learning and tacit knowledge / A.S. Reber // J. Exp. Psychol.
Gen. - 1989. - N118. - P. 219-235.
. Reber,
A.S. Implicit learning of artificial grammars / A.S. Reber // J. Verbal Learn.
Verbal Behav. - 1967. - N6. - P. 855-863.
. Rhodes,
B.J. Learning and production of movement sequences: behavioral,
neurophysiological, and modeling perspectives / B.J. Rhodes, D. Bullock, W.B.
Verwey, B.B. Averbeck, M.P. Page // Hum Mov Sci. - 2004. - P. 699-746.
. Rueda,
M.R. Attentional control and self-regulation/M.R. Rueda, M.I. Posner, M.K.
Rothbart // R.F. Baumeister, K.D. Vohs (Eds). Handbook of self-regulation:
research, theory, and applications. New York: Guilford Press. - 2004. - P.
283-300.
. Ruff,
C.C. Concurrent TMS-fMRI and psychophysics reveal frontal influences on human
retinotopic visual cortex / C.C. Ruff, F. Blankenburg, O. Bjoertomt, S.
Bestmann, E. Freeman, J.D. Haynes, G. Rees, O. Josephs, R. Deichmann, J. Driver
// Curr Biol. - 2006. - P. 1479-1488.
. Ruff,
C.C. Distinct causal influences of parietal versus frontal areas on human
visual cortex: evidence from concurrent TMS-fMRI / C.C. Ruff, S. Bestmann, F.
Blankenburg, O. Bjoertomt, O. Josephs, N. Weiskopf, R. Deichmann, J.
Driver//Cereb Cortex. - 2008. - P. 817-827.
. Serences,
J.T. Selective visual attention and perceptual coherence/J.T. Serences, S.
Yantis//Trends Cogn Sci. - 2006. - P. 38-45.
. Soto,
D. Distinct causal mechanisms of attentional guidance by working memory and
repetition priming in early visual cortex/D. Soto, D. Llewelyn, J. Silvanto //
J. Neuroscience. - 2012. - P. 3447-3452.
. Verhoef,
B.E. Synchronization between the end stages of the dorsal and the ventral
visual stream/B.E. Verhoef, R. Vogels, P. Janssen // J. Neurophysiol. - 2011. -
P. 2030-2042.
70. Wittgenstein,
L. Philosophical investigations / L. Wittgenstein // Oxford, UK: Blackwell.
71. Zikopoulos,
B. Circuits formultisensory integration and attentional modulation through the
prefrontal cortex and the thalamic reticular nucleus in primates / B.
Zikopoulos, H. Barbas // Reviews in the neurosciences. - 2007. - P.417-438
. Zikopoulos,
B. Prefrontal projections to the thalamic reticular nucleus form a unique
circuit for attentional mechanisms // J. Neurosci. - 2006. - P. 7348-7361.