Фітопланктон прісних водойм
ЗМІСТ
ВСТУП
РОЗДІЛ I:ФІТОПЛАНКТОН ПРІСНІХ
ВОДОЙМ
.1 Фітопланктон прісних водойм
.2 Загальна характеристика
каскаду Горіхуватських ставків
.3. Особливості видового складу
фітопланктону каскаду Горіхувастих ставків
РОЗДІЛ ІІ: ЕТАПИ ДОСЛІДЖЕНЬ
ФІТОПЛАНКТОНУ КАСКАДУ ГОРІХУВАСТИХ СТАВКІВ
.1 Визначення повного обсягу
ставка
.2 Визначення прозорості ставка
.3 Відбір проб
.4 Консервація проб
.5 Згущення проб
.6 Камеральне опрацювання проб
.7Визначення чисельності і
біомаси
.8. Визначення первинної
продукції фітопланктону і деструкції органічних речовин
.9. Оцінка різноманітності
фітопланктону
РОЗДІЛ ІІІ: ВИЗНАЧЕННЯ
ЧИСЕЛЬНОСТІ ТА ДИНАМІКИ БІОМАСИ ФІТОПЛАНКТОНУ
3.1 Визначення
повного обсягу досліджуваних водойм
3.2 Визначення прозорості
ставка
.3 Визначення чисельності і
біомаси
.4 Оцінка різноманітності
фітопланктону
.5 Визначення біомаси
фітопланктону за кількістю клітин водоростей
РОЗДІЛ IV: ЕКОНОМІЧНЕ ЗНАЧЕННЯ
ПРОЦЕСІВ ФОТОСИНТЕЗУ У СВІТОВОМУ ОКЕАНІ
.1 Глобальні процеси
фотосинтезу у світовому океані
.2 Квоти на викид парникових
газів
РОЗДІЛ V: ОХОРОНА ПРАЦІ
5.1 Небезпечні та шкідливі
виробничі фактори при роботі в біохімічній лабораторії
.2. Технічні та організаційні
заходи для зменшення рівня впливу небезпечних та шкідливих виробничих факторів
при роботі в біохімічній лабораторії
5.3. Забезпечення пожежної і
вибухової безпеки при роботі в біохімічній лабораторії
ВИСНОВКИ
СПИСОК ВИКОРИСТАНОЇ ЛІТЕРАТУРИ
ВСТУП
Актуальність проблеми.
Відслідковування закономірностей зміни біомасси фітопланктону - актуальне
завдання сучасної гідробіології. Незважаючи на те, що фітопланктон є основною
фотосинтезуючою ланкою водних екосистем, механізми зв’язку
фізіолого-біохімічних процесів, що лежать в основі продуктивності, з факторами
навколишнього середовища досліджені недостатньо. Головна увага традиційно
приділяється встановленню кількісних показників первинної продукції в
конкретних водних об’єктах, а також аналізу структури альгоугруповань. Водночас
глибоке осмислення екологічних процесів і складання реальних прогнозів зміни
стану водних екосистем неможливі без всебічних еколого-фізіологічних досліджень
механізмів формування та перебудови альгоценозів (Гольд, 1996; Елизарова; 1999,
Минеева, 2004).
Високий рівень антропогенного
навантаження на водні об’єкти України зумовлений великою щільністю населення,
впливом розвинутої промисловості, інтенсифікацією сільського господарства, а
також недостатньо ефективною системою очищення стічних вод, що надходять у
водойми. Наприкінці 80-х - на початку 90-х років ХХ століття істотно підвищився
рівень мінералізації води приток Дніпра та його водосховищ (Журавлева, 1998),
збільшився вміст у ній розчиненого мінерального і загального фосфору
(Журавлева, Курейшевич, 2000; Гидроэнергетика..., 2004), підвищилась
концентрація у воді і донних відкладах токсикантів та радіонуклідів
(Калениченко, 1993; Линник, 1999; Арсан та ін., 1997; Романенко та ін., 2000;
Шаповал, Кукля, 2001; Кузьменко та ін., 2002). Це призвело до суттєвого
погіршення якості води основних джерел водопостачання країни.
У зв’язку з зазначеним вище виникає
необхідність з’ясування відгуку планктонних водоростей на вказані зміни
навколишнього середовища. Дослідження фундаментальних основ формування
фітопланктону та глобального впливу природних чинників на цей процес в умовах антропогенного
пресу важливі для розробки теорії біологічної продуктивності водойм, науково
обґрунтованих методів управління функціонуванням екосистем та прогнозування
якості води.
РОЗДІЛ І. ФІТОПЛАНКТОН ПРІСНИХ
ВОДОЙМ
1.1
Фітопланктон прісних водойм
Термін планктон (грец. «планктон» -
блукаючий) вперше був введений у науку 1887 р. і за первісним поданням означав
сукупність організмів, що ширяють у воді. Дещо пізніше у складі планктону стали
розрізняти фітопланктон (рослинний планктон) і зоопланктон (тваринний
планктон). Отже, фітопланктоном називають сукупність свободноплаваючих (в товщі
води) дрібних, переважно мікроскопічних, рослин, основну масу яких складають
водорості. Відповідно кожен окремий організм зі складу фітопланктону називають
фітопланктером. Екологи вважають, що фітопланктон в житті великих водойм
виконує ту ж роль, що і рослини на суші, тобто виробляє первинне органічна
речовина, за рахунок якого прямо або опосередковано (через ланцюг живлення)
існує весь інший живий світ на суші і у воді.
Це справедливо. Однак слід
пам'ятати, що до складу фітопланктону, так само як і до складу наземних
рослинних угруповань, входять гриби і бактерії, які, за рідкісним винятком, не
здатні самі створювати органічну речовину. Вони відносяться до тієї ж екологічної
групі гетеротрофних організмів, що харчуються за рахунок готового органічної
речовини, до якої належить весь тваринний світ. Гриби і бактерії беруть участь
в руйнуванні мертвого органічної речовини, тим самим виконуючи хоча і дуже
важливу роль у кругообігу речовин, але принципово іншу, ніж зелені рослини.
Незважаючи на це, основною функцією фітопланктону в цілому слід все ж визнати
створення органічної речовини водоростями.
Тому далі ми будемо говорити тут
лише про мікроскопічні водорості, що входять до складу фітопланктону. Це тим
більше виправдано, що склад грибів в співтоваристві фітопланктону вивчений ще
дуже слабо, а планктонні бактерії (бактеріопланктона) в екології водойм
розглядаються зазвичай окремо. Існування планктонних організмів у зваженому
стані у воді забезпечується деякими спеціальними пристосуваннями. В одних видів
утворюються різного роду вирости і придатки тіла - шипи, щетинки, рогоподібні
відростки, перетинки і т. п. ; у інших видів відбувається накопичення в тілі
речовин з питомою вагою менше одиниці, наприклад крапель жиру, газових вакуолей
І т. д. Полегшується маса клітини також шляхом зменшення її розмірів: розміри
клітин в планктонних видів, як правило, помітно менше, ніж у близькоспоріднених
донних водоростей. У планктоні широко поширені і дрібні організми, розміром в
декілька мікрометрів, які утворюють так званий наннопланктон.[7]
У прісноводому комплексі нерідкі і
зелені водорості: ооцістіс (Oocystis) (по всьому морю), види сценедесмус
(Scenedesmus) і педіаструма (Pediastrum), більш численні в затоках.Прісноводний
фітопланктон відрізняється від типово морського величезною різноманітністю
зелених і синьо-зелених водоростей. Особливо численні серед зелених
одноклітинні і колоніальні вольвоксовие і протококковие: види хламідомонад (Chlamydomonas),
гоніума (Gonium), вольвокса (Volvox), педіаструма, сценедесмус, ооцістіса,
сфероцістіса (Sphaerocystis) та ін . Серед синьо-зелених багаточисельні види
Анаб, мікроцістіса, афанізоменона, глеотріхіі (Gloeotrichia) та ін.
Характернейший в морському
фітопланктоні рід хетоцерос (Chaetoceros) в озерах і ставках зовсім відсутня, а
численні в морях різосоленія (Rhizosolenia) в прісних водах представлені лише
декількома видами. У прісноводому фітопланктоні значно біднішими якісно і
кількісно представлені перидинеї. Звичайні серед них види цераціума (Ceratium)
і перідініума . У прісних водах відсутні кремнежгутіковие і дуже рідкісні
кокколітофоріди, зате деякі інші жгутикові представлені тут різноманітно і
нерідко у великій кількості. В основному це хрізомонади - види дінобріона
(Dinobryon), малломонаса (Mallomonas), уроглени (Uroglena) та ін , а також
евгленовие - евглена (Euglena), трахеломонас (Trachelomonas) і факус (Phacus)
перша переважно в холодних водах, а другі - в теплих.
Однією з істотних особливостей
прісноводного фітопланктону є велика кількість у ньому тимчасово планктонних
водоростей. Ряд видів, яких прийнято вважати типово планктонними, в ставках і
озерах мають донну або періфітоном (прикріплення до якогось предмета) фазу в
своєму життєвому циклі. Таким чином, різноманітність екологічних умов у
внутрішніх водоймах визначає і значно більшу різноманітність в порівнянні з
морями екологічних комплексів і видового складу прісноводного планктону.[10]
У великих глибоких озерах виражені
менш яскраво риси відмінності прісноводного фітопланктону від морського. У
таких озерах-гігантах, як Байкал, Великі озера, Ладозьке, Онезьке, в
фітопланктоні майже цілий рік переважають діатомові. Тут вони, як і в морях,
створюють основну продукцію. Видовий склад діатомових озерного планктону
відмінний від морського, але в їх екології є чимало спільного. Наприклад,
мелозіра ісландська (Melosira islandica) - масовий вид фітопланктону Ладозького
і Онезького озер, а також мелозіра байкальська з Байкалу в фазу спокою після
весняної спалаху не опускаються на дно (або опускаються лише частково), як це
спостерігається в інших прісноводних видів у водоймах меншого розміру, а
утримуються в товщі води, утворюючи на деякій глибині характерні міжсезоння
скупчення. У великих озерах, як і в морях, великі розходження в продуктивності
фітопланктону: в центральній частині водоймища продуктивність дуже низька, а
біля берегів, особливо в мілководних затоках і проти гирл річок, вона різко
підвищується.[14]
Ще більше схожості з морським у
фітопланктоні двох найбільших озер світу з солоною водою - Каспії і Аралі. Хоча
солоність води в них значно нижче морський (у Каспії 12-13 ° / 00, в Аралі
11-120/00), але в складі фітопланктону тут переважають водорості морського
походження, особливо серед діатомових: види хетоцероса (Chaetoceros),
різосоленія (Rhizosolenia) та ін. З джгутикових характерні солоноватоводние
види ексувіелли (Exuviella) та ін В опріснених зонах цих озер панують
прісноводні види, проте при солоності води навіть 3-5 ° / 00 ще дуже
різноманітний солоноватоводний фітопланктон морського походження.
В найбільш типовому вигляді
прісноводний фітопланктон як за складом та екології, так і по продукційних
властивостям представлений в озерах середнього розміру помірної зони, наприклад
в озерах Балтійського басейну.
Тут в залежності від типу озера і
сезону року в фітопланктоні переважають діатомові, синьо-зелені або зелені
водорості. . Із діатомових характерні види мелозіри (Melosira), астеріонелли
(Asterionella), табелляріі (Tabellaria), фрагіляріі (Fragilaria), ціклотелли
(Cyclotella) та ін; серед синьо-зелених - види мікроцістіса (Microcystis), Анаб
(Anabaena), афанізоменона (Aphanizomenon), глеотріхіі (Gloeotrichia). Основними
представниками зелених водоростей в озерному планктоні є перераховані вище
протококковие, а в водах з дуже м'якою водою, знаходяться під впливом боліт,
численні десмідиєві: види космаріума (Cosmarium), стаураструма (Staurastrum),
клостеріума (Closterium), еуаструма (Euastrum) та ін .
У мілководних озерах і ставках із
зелених водоростей нерідко переважають вольвоксовие: вольвокс (Volvox),
хламідомонада (Chlamydomonas), пандорина (Pandorina), евдоріна (Eudorina). У
фітопланктоні озер тундри і північної тайги досить різноманітні хрізомонади:
види дінобріона (Dinobryon), сінури (Synura), урогленопсіса (Uroglenopsis),
малломонаса (Mallomonas). Характернейшая для морського фітопланктону група
перидинеї в прісних водах представлена всюди (у всіх водоймах), але відносно
невеликим числом видів, які скрізь, за рідкісним винятком, досягають невисокою
чисельністю. У самих малих водоймах - в невеликих озерах і ставках - вельми
різноманітні і нерідко численні евгленовие, особливо види трахеломонаса
(Trachelomonas), а в теплих водоймах тропіків і субтропіків також евглени
(Euglena), лепоцінкліс (Lepocynclis), факус (Phacus) та ін У кожному окремому
водоймі в залежності від фізичних і хімічних особливостей режиму і від сезону
року переважає одна або інша з перерахованих груп водоростей, а в періоди дуже
інтенсивного розвитку панує нерідко всього один вид. У дрібних тимчасових
водоймах - калюжах, викопаних ямах - вельми звичайні дрібні вольвоксові з роду
хламідомонас (Chlamydomonas), від масового розвитку яких вода нерідко
забарвлюється в зелений колір.[5]
Прохлорофіти
Це також прокаріотів. Вони
відрізняються від ціанофіт тим, шо ні не містять фікобіліпротеїнів. Гусєв М.В.
і Мінєєва Л.А. визначають фікобіліпротеїнів як червоні і сині пігменти, що
містяться тільки в однієї групи еукаріотів - ціанобактерій (також червоних і
кріптофітових водоростей). Великі маси Prochlorothrix hollandica в дрібних
озерах в Нідерландах призвели до відкриттю там прохлорофіт.
Глауконіти
Дуже маленька група, що включає три
рідкісних прісноводних різновиди. Основний інтерес представляє те, що у цієї
групи з'явилися хлоропласти, подібні з хлоропластами ціанобактерій, імовірно
еволюційним шляхом.
Родофіти
Родофіти зазвичай називають
червоними морськими водоростями через їх забарвлення. В основному це не
свободноплавающие водорості, вони розвиваються на прибережних скелях. У 2004 р.
Редфілд запропонував виростити і зібрати кілька різновидів родофіт (Gracilaria
і Porphyra) в високопоживних прибережних областях, щоб зменшити можливість
еутрофікаціі. Хлорофіти
Це група зелених морських
водоростей, що відрізняється великою видовою різноманітністю. Деякі форми є
одноклітинними і планктоніческімі, в той час як інші - макроскопічні. Spirogyra
- широко поширена форма, яка під час цвітіння утворює своєрідну піну на
поверхні невеликих водойм. Планктоні форми (Chlamydomonas) характеризуються
наявністю двох джгутиків рівної довжини, які дозволяютьрослині плавати.
Евгленофіти
Евгленофіти характеризуються єдиною
мембраною хлоропласта ендоплазматичної мережі, мезокаріотіческім ядром, одним
рядом волосків на Джгутики і змістом парамілона в цитоплазмі. Їх розмір 50m m.
В несприятливих умовах евгленофіти утворюють цисти.
Дінофіти
Дінофлагеллати - в значній мірі
одноклітинні бі-флагеллати з відмітними структурами. Розмножуються
розщепленням. Поширені цисти. У такому стані водорості можуть перебувати багато
років, поки зміни в навколишньому середовищі (температура, поживні речовини,
світло і т.д.) не дадуть цисті можливості прорости. Зростання дінофіт
підпорядковується експоненціальним законом: з однієї цисти можуть за тиждень
розвинутися 6-8000 клітин. Деякі дінофлагеллати повністю гетеротрофних, інші -
міксотрофні. Деякі дослідники, припускають, що їх можна назвати мезокаріотамі.
Дінофлагеллати мешкають і в морській, і в прісній воді, проте в прісній менше
різновидів. Отруйні розквіти Gymnodium мають важливе значення в рибоохорони
справі. В океані дінофлагеллати відповідальні за так звані червоні потоки, які
були предметом багатьох досліджень. На це звернув увагу Андерсон (1995). Він
доповнює, що термін «червоні потоки» може ввести в оману, і наукове
співтовариство вважало за краще назву «шкідливе цвітіння водоростей» (НАВ). НАВ
проявляє себе в різних формах, говорить Anderson, наприклад, отруйний планктон
споживається молюсками, мідіями, устрицями, які, потрапляючи на стіл до людини,
можуть призвести до важкого отруєння і навіть до смерті. Всі основні токсичні
речовини виробляються дінофлагеллати. Також від токсинів страждають риби, кити,
морські птахи та інші тварини.
Криптофіти
Мешкають і в прісній, і в солоній
воді. Їх відмітною властивістю є слабка чутливість до світла, вони можуть
мешкати на більшій глибині, ніж інші водорості. Здатні до виживання у воді,
покритій снігом і льодом.
Кріофіти
Золотисто-коричневі водорості -
головним чином мешканці прісної, переважно м'якої води. Важлива особливість
динаміки їх зростання - те, що навіть у сприятливому навколишньому середовищі
кожен день цілих 10% популяції піддаються інцістірованню. Це приводить до
пониження їх конкурентоспроможності. Aureococcus утворюють «коричневі потоки» в
водоймах. Це явище досліджено та змодельовано американськими вченими в
2000-2002 р. Екологи прийшли до висновку, що розмноження Aureococcus
anophagefferen призводить до збільшення у воді концентрації цих водоростей. В
результаті на глибину проникає менше світла, що впливає на ріст і розвиток
бентичних водоростей і ракоподібних.
Прімнезіофіти
Прімнезіофіти (або гаптофіти) є
більшою мірою морськими
одноклітинними флагеллатамі
(жгутиконосцами). Флагеллати мають два джгутики, які, залежно від різновиду
можуть бути рівними або нерівними. Існує приблизно 75 пологів і 500 різновидів
прімнезіофіт. Розмір їх досить малий: 5m m Можливо найвідоміша різновид -
Emiliania huxleyi, які
поширені від тропіків до полярних
областей. Olson і Strom повідомили про обширному розквіті в Беринговому морі, в
якому, очевидно, розвитку прімнезіофіт сприяє слабка конкуренція. Phaeocystis
pouchetii проходить дві життєві стадії: як одноклітинна флагаллата і
колоніальну стадію, з'єднуючись в полісахарідную форму. Остання може бути
неприємністю, коли цвітіння відбуваються в регіональних або рибальських
областях.
Баціларіофіти (діатомові водорості)
Діатомові водорості - одноклітинні
морські водорості, знайдені практично у всіх водах, і, ймовірно, найчисленніша
група з водних еукаріотичних організмів. Існує приблизно 250 пологів і більше
100000 різновидів. Найбільш загальними видами є Skeletonema, Thalassiosira,
Nitzschia. Клітини оточені твердою кварцовою оболонкою, круглої або злегка
витягнутої форми. Ця оболонка, яка часто утворює заплутаний і дуже гарний візерунок,
називається фрустулой. Їх характерний колір походить від каротиноида,
золотисто-коричневого фукоксантина. Висока вміст кварцу в оболонках діатомових
водоростей означає, що він може стати лімітуючим елементом. Деякі різновиди
отруйні. Наприклад, Pseudo-nitschia і деякі інші виділяють домоеву кислоту, які
викликають отруєння ракоподібних, Chaetoceros є причиною загибелі риби.
Ксантофіли
Ксантофіти мають жовто-зелене
забарвлення, два джгутики нерівної довжини, один з яких, покритий волосками,
спрямований вперед, а інший, гладкий, спрямований назад. Більшість різновидів -
прісноводні. Розмноження безстатеве поділом клітини.
Еустігматофіти
Це дуже маленька група всього з
дванадцяти різновидів, що мешкають впрісній воді і грунті.
Рафід офіти
Інший маленький клас флагеллат.
повідомлено, що одна різновид викликала масову смертність тунця в японських і
австралійських водах, причому грошові втрати в Японії склали приблизно $ US500
мільйонів і в Австралії $ US24 мільйонів.
Фаеофіти
Фаеофіти - практично всі бентіческіх
морські організми, що мають розміри від мікроскопічних до гігантських
водоростей. Вони мають характернийкоричневий колір завдяки фукоксантин і різним
таніну. Коричневіморські водорості широко використовуються в їжу і в
виготовленніфармацевтичних препаратів. Одна із широко використовуваних
різновидів - Laminaria japonica, яку, можна було б використовувати для того,
щоб зменшити прибережну еутрофікацію.[15]
Живильні речовини
Більшість видів фітопланктону є
фотоавтотрофним, що означає використання неорганічних речовин для росту,
вуглекислого газу як джерела вуглецю і світла як енергії для всіх процесів.
Хемоавтотрофні організми споживають уже готові органічні речовини. Морські
водорості, які отримують вуглець з органічних складів, як кажуть, гетеротрофних
і можуть бути або фото-чи хемогетеротрофни.
Розмноження
Багато різновидів розмножуються
поділом клітини. Деякі багатоклітинні організми формують фрагмент або
виробляють спеціальну структуру, звану пропагул. Цей тип називається
вегетативним способом розмноження на відміну від формування спеціалізованих
одноклітинних суперечка, що називають безстатевим розмноженням. Спори, які
з'єднуються, щоб справити зиготу, називаються гаметами і можуть не мати
джгутиків. Деякі спори мають потовщені стінки і збільшений запас поживних
речовин, щоб пережити можливі несприятливі умови. Такі спори називаються
акінети. Багато еукаріоти проходять кілька стадій життєвого циклу, змінюючи
форми. Життєвий цикл Pfiesteria piscicida, надзвичайно складний і має 24
одноклітинні стадії, з яких флагеллатние і амебоидние мають найбільше значення.
Він пише, що поки Pfiesteria знаходиться в стадії недіючої амеби, вона
нешкідлива. Як тільки виникають сприятливі умови, оболонка цисти руйнується і з
неї з'являється клітина, яка може бути небезпечна. Сама отруйна стадія, на
думку вченого, - це «Рослинна», яка викликає масову загибель риби. Сезонні
зміни Як відомо, сезонні зміни впливають на конкурентоспроможність різних видів
планктону.
. До кінця зими достатню кількість
поживних речовин та інтенсивність світла збільшується, що призводить до
стрімкого розвитку маленького, швидко зростаючого фітопланктону.
. Зростання зоопланктону
стимулюється появою цієї їжі, спочатку домінують короткоживучі покоління.
. Розвиток травоїдних тварин
перевищує темп зростання фітопланктону, що призводить до зниження концентрації
другого, настає фаза чистої води.
. Популяційна щільність травоїдних і
біомаса їх зменшуються внаслідок зростаючого хижацтва риби.
. Зменшення зоопланктону призводить
до літнього розвитку їстівної різновиди Cryptophycae і неїстівних зелених
водоростей, за чим слід зменшення поживних речовин.
. Боротьба за фосфор призводить до
переваги діатомових водоростей над зеленими морськими водоростями, поки не
вичерпається кварц, далі встановлюється домінування дінофлагеллати і / або
ціанобактерій.
. Великі травоїдні ракоподібні
тварини змінюються більш дрібними, оскільки вони менш вразливі для хижаків.
. Складне співтовариство
розвивається і коливається зі збільшенням температури.
. Фізичні зміни до кінця літа
роблять домінуючими більш конкурентоздатні в змішаній навколишнього середовища
організми, зокрема, діатомові водорості.
.Збільшення розміру зоопланктону і
його біомаси росте до осіннього максимуму, поки не зменшиться кількість світла
і поживних речовин, тоді він переходить до зимового станом.[16]
Парадокс планктону
Відома робота «Парадокс планктону»
зачіпає наступну проблему. Принцип конкурентного виключення формулюється так:
«якщо векосистемі є n споживачів і r лімітують речовин, то рівновага настає,
коли залишається r споживачів ». Однак в реальних екосистемах різноманітність
видів завжди перевищувала кількість поживних речовин.[17]
.2 Загальна
характеристика каскаду Горіхувастих ставків
Значну частину Голосіївського району
становить Голосіївський ліс. Його площа перевищує 4 тисячі гектарів. Це один з
найбільших внутрішньоміських лісових масивів в Європі. Найбільш часто
зустрічаються дерева - дуб, береза, граб, сосна. Незважаючи на те, що в Роки
Великої Вітчизняної війни ліс був практично знищений, зустрічаються окремі
екземпляри дубів віком 200-300 років. Особливу привабливість лісу додає ланцюг
з природних озер (озеро Дідорівка і озеро Голубе) і штучних ставків (озера
Спортивне, Гниле і Митькина). У Парковій частині Голосіївського лісу (т. зв. Голосіївський
парк ім. М.Рильського) розташувалася ланцюг Оріхуватських ставків. Парк ім
М.Рильского було відкрито у 1957 р.
Найцікавішим водним об’єктом парку
ім. Рильського на мою думку є саме ланцюг Горіхувастих ставків, з’єднаних між
собою річкою Оріхуваткою і підземними водами.
Оріхуватка - річка в Голосіївському
районі міста Києва, права притока Либіді. Довжина - близько 4 км. Бере свій
початок в Голосіївському лісі неподалік від Експоцентру. На початку це
невеликий, неширокий струмок, що протікає по лісі.Ширина струмка не перевищує 1
метра. На території парку ім. Рильського починається каскад ставків - 3
невеликих ставка, далі річка знову протікає по лісі, потім проходить через ще
один ставок і біля Голосіївської площі - останній ставок (Оріхуватські) із цього
каскаду. А після цього річка зникає у колекторі і біля маргаринового заводу
впадає в Либідь.Таким чином, річку можна вважати малоурбанізірованной - більшу
частину свого шляху вона протікає по природному руслу.
Береги озер пологі і вкриті густою
рослинністю.
Знаходяться лише в декількох сотнях
метрів від траси і тому відчувають на собі значний антропогенний вплив. Саме
тому ланцюг ставків цього каскаду був обраний як об’єкт дослідження.
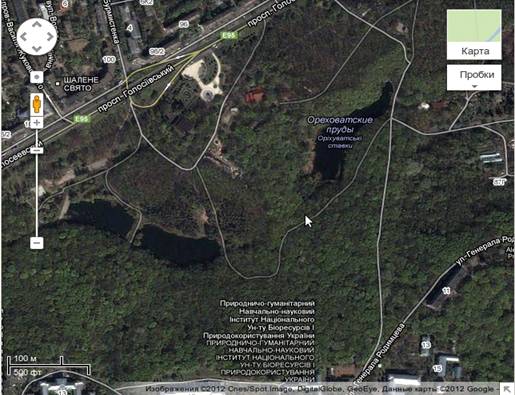
Рис. 1.1 Каскад Горіхувастих ставків
-супутникові данні.
.3 Особливості
видового складу фітопланктону каскадів Горіхувастих ставків
Провідне значення у функціонуванні
водних екосистем має фітопланктон - первинна ланка потоків енергії, продуцент
автохтонної органічної речовини, важливий агент самоочищення і фотосинтетичної
аерації води.
Фітопланктон є одним із біологічних
елементів класифікації екологічного статусу озер відповідно до Водної Рамкової
Директиви ЄС 2000/60 (Directive 2000/60/EC). Разом з тим, у Директиві ЄС не
наводяться межі коливань структурних і функціональних характеристик планктонних
водоростей для кожної градації екологічного статусу водойм. До того ж, міські
водойми у Водній Рамковій Директиві не розглядаються.
Фітопланктон одним з перших ланок
гідpоекосістем реагує на зміни екологічних умов, a його
структурно-функціональні характеристики є важливими складовими при оцінці
ступеня антропогенного впливу. Тому, поза всяким сумнівом, флористичне та
ценотичного різноманіття співтовариств планктонних водоростей набуває показове
значення при встановленні особливостей трансформації водних екосистем в умовах
антропогенного пресу.[34]
Враховуючи вищевикладене,
представляється актуальним і доцільним виявлення особливостей видового складу
планктонних водоростей, що розвиваються в найбільш великих водоймах
Голосіївського лісу, територія якого набула статусу об'єкта
природно-заповідного фонду України в ранзі Національного природного парку
(НПП).
Фітопланктон Дідоровських,
Оріхуватських і Китаївських ставків представлений, відповідно, 167, 154 і 101
видів і внутрішньовидових таксонів (в.в.т.). Всього в досліджених ставках
Голосіївського лісу відзначено 272 види водоростей, представлених 284 видів і
внутрішньовидових таксонів, які відносяться до 9 відділів, 15 класів, 35
порядків, 63 родин і 131 роду (табл.1.1).
Таблиця 1.1
Таксономічний спектр і «пропорції
флори» планктонньх водоростей ставків Голосіївського лісу.
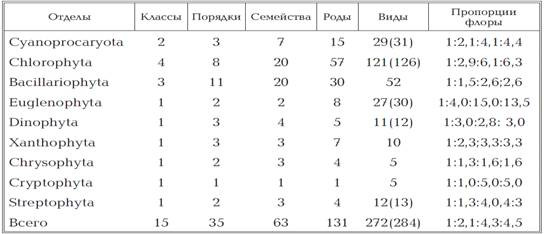
Показники таксономічного
різноманіття або «пропорції флори» наведені за формою: відносна кількість
сімейств, прийняте за 1: середня кількість пологів в сімействі (р / р): середня
кількість видів в сімействі (в / c): середня кількість внутрішньовидових
таксонів (включаючи номенклатурний тип виду) (ст / c). [35]
Основу видового багатства фітопланктону
водойм НПП «Голосіївський» складають Chlorophyta (121 вид - 126 в.в.т.) або
44,4% загального числа виявлених таксонів видового і внyтрівідового рангу),
Bacillariophyta (52 види або 19,1%), Cyanoprocaryota (29 видів - 31 в.в.т. або
10,7%) та Еuglеnophyta (27 видів - 30 в.в.т. або 9,9%). Інші відділи
представлені 5-13 видами.
Серед зелених водоростей найбільш
різноманітний у видовому відношенні клас Chlorophyceae (73,0% зведеного списку
видів), що включає порядки Sphaeropleales (70 видів), Chlamydomonadales (14
видів), Volvocales (5 видів) і Chlorococcales (3 види). Друге місце належало
класу Trebouxiophyceae, представленого тільки одним порядком - Chlorellales (31
вид).
Серед Bacillariophyta найбільшим
числом видів представлений клас Васіlariophyceae (69,2%), основу якого складав
порядок Achnantales (28,9%). Друге місце займав клас Fragilariophyceae (17,3%),
зокрема порядок Fragilariales (15,4%), a третє - Coscinodiscophyceae
(15,4%).[11]
Видове різноманіття Cyanoprocaryota
формували в основному представники класу Hormogoniophyceae (22 види або 71,0%),
що включає порядки Oscillatoriales (35,5%) і Nostocales (35,5%). Водорості, що
відносяться до класу Chroococcophyceae, налічували всього 9 видів, що становило
29,0% видового складу синьозелених водоростей.
Видове різноманіття евгленових
водоростей у ставках НПП «Голосіївський» обумовлено представниками класу
Euglenophyceae, зокрема, відносяться до порядку Euglenales (93,3%). Основу
видового багатства дінофітових водоростей становив тільки один клас Dinophyceae,
що включає порядки Peridiniales (66,6%), Gymnodiniales (16,7%) і Gonyaulacales
(16,7%). Стрептофітовие водорості в ставках НПП «Голосіївський» представлені
видами з класу Zygnematophyceae, що відносяться переважно до порядку
Desmidiales (92,3%).
Аналіз видового багатства
фітопланктону кожного з каскадів показав, що в Дідоровських, Оріхуватські і
Китаєвських ставках воно формується, головним чином, представниками Chlorophyta
(відповідно 48,0, 48,6 і 40,7%) і Bacillariophyta (14,1, 14,1 і 24,1%) (табл.
2). Третє і четверте місця в ставках Дідоровських урочища займали, відповідно,
представники Cyanoprocaryota (11,6%) і Euglenophyta (9,8%), тоді як в
Китаєвських і Оріхуватські ставках на третій позиції знаходилися Euglenophyta
(14,5 і 7,6%), a на четвертій - Cyanoprocaryota (7,5 і 6 , 9%).
Таблиця 1.2
Рангові місця різних відділів
водоростей при формуванні різноманітності фітопланктону досліджених ставків
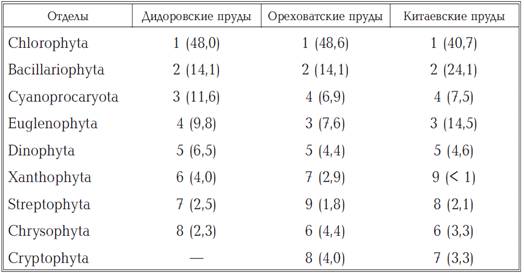
«-» - Представники даного відділу не
виявлено; в дужках вказано відношення числа видів даного відділу до загальної
кількості видів зведеного списку водоростей (%).
Частка водоростей інших відділів у
формуванні рослинного планктона ставків у сумі становила 13,3-17,5%. Найбільш
бідно в фітопланктоні всіх ставків був представлений відділ Streptophyta, a в
деяких випадках Xanthophyta (Китаївські ставки), тоді як водорості з відділу
Cryptophyta не були виявлені в Дідоровських ставках.[36]
Аналіз провідних (по числу видів)
сімейств, що включають 41,0-57,5% видів зведеного списку планктонних
водоростей, показав, що в Дідоровських, Оріхуватські і Китаївському каскадах
ставків перші два рангових місця займали Scenedesmaceae (відповідно 16,3, 14,9
і 12,9%) і Euglenaceae (9,8, 8,0 і 14,9%) (табл. 1.3). Загальними для всіх ставків
(з варіюванням Рангового місця) були ще 2 родини - Selenastraceae і
Chlorellaceae.
Таблиця 1.3
Розподіл рангових місць між
провідними родинами фітопланктону досліджених ставків.
«-» - Середнє число видів в
сімействі менше двох; в дужках вказано відносно числа видів даного сімейства до
загальної кількості видів зведеного списку (%).
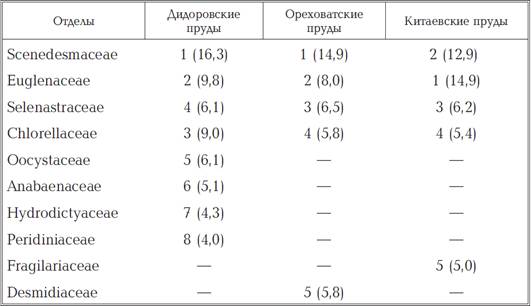
Крім згаданих вище родин, до складу
провідних в ставках Дідоровських каскаду входили Oocystaceae, Anabaenaceae,
Hydrodictyaceae і Peridiniaceae, a Китаєвських і Оріхуватські ставків -
Fragilariaceae і Desmidiaсеае, відповідно.фітопланктоні Голосіївських ставків
на частку одно-(25 родин) і двувидових (6) сімейств доводиться 49,2% їх
загальної кількості. З них для ставків всіх каскадів загальними є три сімейства
- Aphanizomenonaceae, Catenulaceae і Sinuraceae.[37]
Варіабельність виду у всіх каскадах
ставків не перевищувала значень 1,06, a найменшою виявилась для фітопланктону
Китаївських ставків (1,01). Фітопланктон характеризувався наступними
«пропорціями флори»: Дідоровських ставків - 1: 2,0: 3,3: 3,5, Китаївських - 1:
1,7: 1,4: 1,5 і Оріхуватські - 1: 1,7: 3 , 0: 3,1. Для фітопланктону ставків
Голосіївського лісу отримані невисокі значення загального родового коефіцієнта
(2,1). При порівнянні цього показника по відділах встановлено, що найбільш
високі його величини характерні для Euglenophyta (3,8), Streptophyta (2,5) і
Dinophyta (2,4), a нижчі - для Chlorophyta (2,2), Cyanoprocaryota (2,1) і
Bacillariophyta (1,7).число провідних родів планктонних водоростей досліджених
ставків входили Desmodesmus (Chod.) An, Friedl et E. Hegew. - 11 видів (15
в.в.т.), Еuglепа Ehrenb. - 11, Oscillatoria Vaucher et Gomont - 8, Anabaena
Вогу ех Bornet et Flahault - 7 (8), Trachelomonas Ehrenb. - 6 (8), Monoraphidium
Komark.-Legn. - 6, Nitzschia Hassall - 6, Acutodesmus (E. Hegew.) P. Tsarenko -
5, Peridinium Ehrenb. - 5 і Cryptomonas Ehrenb. - 5 видів.фітопланктоні всіх
каскадів ставків виявлено 25 загальних родів або 19,1% загальної кількості
родів. Серед них 14 належало до Chlorophyta, 5 - до Bacillariophyta, 2 - до
Euglenophyta, 2 - до Cyanoprocaryota і по 1 - до Chrysophyta і Dinophyta.
Незначне число загальних родів, ймовірно, свідчить про різні екологічні умови у
водоймах.фітопланктоні ставів НПП «Голосіївський» 92 родів (70,2% загальної
кількості зареєстрованих пологів) представлені 1 і 2 видами. Більшість з них
(40 родів) відносяться до відділу Chlorophyta, 20 родів є представниками
Bacillariophyta і 12 належать до Cyanoprocaryota. Маловидові роди переважають у
планктоні каскаду Дідоровських і Оріхуватські ставків (відповідно, 63,8 і 55,1%
усієї кількості родів, виявлених в даних водоймах). Родова насиченість в
першому каскаді водоймищ становить 1,8, a в другому - 1,9. Дещо менше
маловидових родів відзначено в каскаді Китаївських ставків (47,1%), де родова
насиченість становила 1,5.
Для фітопланктону окремих ставків
НПП характерно також наявність 15 специфічних родів, тобто що зустрічаються
тільки в певному каскаді. Так, в Дідоровських ставках до їх числа відносяться:
Gomphospheria Kitz., Acanthosphaera Lemmerm., Francea Lemmerm., Golenkiniopsis
Korschikov, Nephrochlamys Korschikov, Siderocystopsis Swale, Ulothrix Kitz.,
Pinnularia Ehrenb., Pleurosigma W. Sr. і Pseudostaurastrum Chodat; в
Китаєвських - Golenkinia Chodat, Gomphoneis Cleve, Tabellaria Ehrenb. і Astasia
Ehrenb. emend. Dujard. Тільки один специфічний рід - Aphanothece Nagelі -
виявлений у Оріхуватські ставках. [38]
З 284 видових і внутрішньовидових
таксонів водоростей, виявлених в ставках Дідоровських, Китаївського і
Оріхуватські каскадів, тільки 33 є загальними для них, що становить всього 14%
зведеного списку водоростей. З них 18 видів відносяться до Chlorophyta, 5 - до
Bacillariophyta, 5 - до Euglenophyta, 3 - до Cyanoprocaryota. Одним видом представлені
відділи Dinophyta і Chrysophyta. Широко розповсюдженими (поширеність 75-100%) є
12 видів, a саме: Actinastrum hantzschii Lagerh., Cruciдепіа tetrapedia
(Kirchn.) W. et G. S. West, Desmodesmus communis (Hegew.) Hegew., Micractinium
pusillum Fres., Monoraphidium griffithii (Berk.) Kom.-Legn., M. irregulare (G.
M. Smith) Kom.-Legn. Pandorina morum (OF Мull.) Вогу, Trachelomonas hispida
(Perty) Steіn emend. Defl., T. volvосіnа Ehr., Stephanodiscus hantzschii Grun,
Ceratium hirundinella (OF Мull.) Schrank і Dinodryon divergens Imhof.[39]
Подальший порівняльний аналіз
видового складу фітопланктону показав, що для каскаду Дідоровських і
Китаївських ставків характерна наявність, крім зазначених вище, ще 9 загальних
видів. Однак у цьому випадку лише один вид - Tetraedron minimum (A. Braun)
Hansg. - Характеризувався поширеністю 75%, тоді як у інших, a саме:
Chlorotetraedron incus (Теіl.) McEntee et аl., Selenastrum gracіle Reinsch,
Tetrastrum triangulare Korschikov, Anabaena spiroides Kleb., Microcystis aeruginosa
(Kitz.) Kitz., M. рulvеrea (Wood) Forti emend. Elenkin, Fragilaria crotonensis
Kitton і Fragilariforma virescens (Ralfs) DM Williams et Round вона становила
25-37%. Розрахований коефіцієнт флористичної спільності для фітопланктону
зазначених систем ставків виявився досить низьким (31,2%) .фітопланктоні
Дідоровських і Оріхуватські ставків, крім зазначених раніше, відзначено ще 27
загальних видів. Однак тільки один з них - Centritractus belenophorus Lemmerm.
можна віднести до числа видів, що часто зустрічаються (поширеність 75%), a 9
видів (Monoraphidium arcuatum (Korschikov) Hindak, M. contortum (Thur.)
Komark.-Legn., Coelastrum astroideum De-Not., Desmodesmus intermedius (Chodat)
E. Hegew., Oocystis borgei Snow, Cyclotella ocelata Pant., Еuglenа proxima P.
A. Dang., Peridinium cinctum (О. F. Міі і.) Ehrenb. та Peridiniopsis
elpatiewskyi (Ostf.) Bourr. характеризувалися поширеністю 50%. Коефіцієнт
флористичної спільності видового складу фітопланктону в даному випадку становив
37,7% (див. табл.1.4).
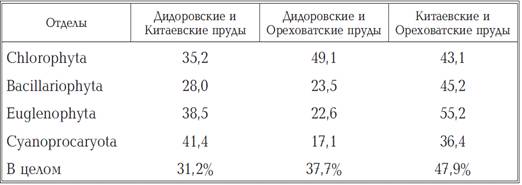
Таблиця 1.4
Коефіцієнти флористичної спільності
видового складу фітопланктону ставків.
Найбільше загальних видів виявлено
при порівняльному аналізі видового складу фітопланктону каскадів Китаєвських і
Оріхуватські ставків. 9 з 32 загальних видів були широко поширені (поширеність
100%) у всіх ставках зазначених систем. До них відносяться Acutodesmus
acuminatus (Lagerh.) P. Tsarenko, Coelastrum sphaericum Nageli, Коlіеllа
longiseta (Vischer) Hindak, Navicula cryptocephala Kitz., Synedra acus Kitz.,
Nitzschia acicularis (Kitz.) W. Sr., N. gracilis Hantzsch., Peridiniopsis
quadridens (Stein) Bourr. і Cryptomonas ovata Ehrenb.
Дещо рідше (75%) зустрічалися такі
види, як Snowella lacustris (Chodat) Komarek, Acutodesmus pectinatus (Meyer) P.
Tsarenko, Chlamydomonas monadina Stein, Desmodesmus abundans (Kirch.) E.
Hegew., D. costato-aculeatus var. spinosus (Deduss.) P. Tsarenko, Synedra ulna
(Nitzsch) Lang.-Bert., Euglena caudata Hubner, Cryptomonas erosa Ehrenb.,
Kephyrion rubriclaustri Conrad, Pseudokephyrion schilleri Conrad та Closterium
gracile Brëb.
Для фітопланктону досліджених
каскадів ставків характерна наявність комплексу специфічних видів, що мешкають
тільки в певній системі водойм і характеризуються високим ступенем поширеності
(50-100%). B зведеному списку планктонних водоростей Голосіївських ставків
специфічні види складають 19,7%. Так, в рослинному планктоні Дідоровських
ставків комплекс специфічних видів налічував 10 видів, a саме: Desmodesmus
armatus (Chodat) E. Hegew., D. opoliensis var. mononensis (Chodat) E. Hegew.,
Crucigeniella apiculata (Lemmerm.) Komarek, Еuglепа mutabilis Schmitz, E.
spathirhyncha Skuja, E. viridis Ehrenb., Phacus orbicularis Hubner, Peridinium
bipes Stein, Goniochloris smithii (Bourr.) Fott і Staurastrum boreale W. West
et G. S. West.Китаївських ставках комплекс специфічних видів формували 6 видів
- Cyclotella meneghiniana Kiitz. (= C. kuetzingiana Thwaites), Trachelomonas
intermedia Р. A. Dang., Pediastrum simplex Meyen,
Epitemia adnata (Kiitz.) Brëb., Gomphoneis olivaceum (Horn.) Daw.
ex Ross et Sims. і Astasia curvata G.A. K1ebs, а в каскаді Оріхуватські ставках
- 5: Chlamydomonas debaryana Gorozh., Ch. pertyi Gorozh., Schroederia robusta
Korschikov, Pteromonas асulеаta Lemmerm. і Aulacoseira italics (Ehrenb.)
Simonsen.
Порівняльний аналіз видового складу
фітопланктону досліджених ставків показав, що найбільша схожість (КФС = 47,9%)
в його складі відзначено для Китаєвських і Оріхуватські ставків. Коефіцієнти
флористичної спільності, розраховані при порівнянні найбільш представлених
відділів, склали при цьому для Bacillariophyta - 45,2%, Euglenophyta - 55,2% і
Chlorophyta - 43,1%. При порівнянні видового складу фітопланктону ставків
Дідоровських і Оріхуватські каскадів найбільшими значеннями КФС характеризувався
відділ Chlorophyta (КФС = 49,1%). Дуже близькі за значенням КФС отримані для
Bacillariophyta (23,5%) і Euglenophyta (22,6%). Більш помітно відрізнявся
видовий склад фітопланктону Дідоровських і Китаївських ставків (КФС = 31,2%).
Найбільшим схожістю в цих водоймах характеризувалися Cyanoprocaryota (КФС =
41,4%), Еuglеnophyta (38,5%) і Chlorophyta (35,2%).
Отже, всього в ставках Дідоровських,
Китаївського і Оріхуватські каскаду ставків виявлено 272 видів водоростей,
представлених 284 внутрішньовидових таксонів (включаючи ті, які містять
номенклатурний тип виду). Знайдені водорості відносяться до 9 відділів, 15
класів, 35 порядків, 63 родин і 131 роду.
Основу видового багатства
фітопланктону водойм Голосіївського парку складають Chlorophyta (44,4%
загального числа зареєстрованих видів), ВасіІІаriophyta (19,1%),
Cyanoprocaryota (10,7%) і Euglenophyta (9,9%).[33]
Рідкісними формами серед виявленого
різноманіття водоростей є: Accmthosphaera zachariäsii
Lemm., Acutodesmus regularis (Swir.) Tsar. Ankyra lanceolata (Korsch.) Fott,
Euglena spathirhyncha Skuja, Phacus orbicularis var. cmgeri (Y.V. Roll) Swir.
та Ceratium hirundmella f. robustum (Amb.) Bachm. [32]
Розподілені види водоростей по
водоймах нерівномірно. Максимальна кількість видів фітопланктону зареєстровано
в ставках Дідоровських каскаду - 162 види, представлених 167 внутрішньовидових
таксонів. B Горіхуватських ставках знайдено 153 види (154 внутрішньовидових
таксонів), a в Китаївських - 101 вид.склад провідних родин всіх досліджених
водойм Голосіївського лісу входили Scenedesmaceae, Euglenaceae, Selenastraceae,
Chlorellaceae, Оосуѕtaceae, Anabaenaceae, Hydrodictyaceae, Peridiniaceae,
Fragilariaceae і Desmidiaсеае, a провідних родів - Desmodesmus, Euglena,
Oscillatoria, АпаЬаепа, Тгаchelomonas, Monoraphidium, Nitzschia, Acutodesmus,
Peridinium і Cryptomonas.
Характерною особливістю
таксономічного складу фітопланктону Голосіївських ставків є наявність значної
кількості маловидових сімейства (49,2% від їх загальної кількості) і родів
(70,2%). B цьому плані найбільш показові ставки Дідоровські каскаду.
Видовий склад планктонних
водоростей, що розвиваються в різних водоймах НПП «Голосіївський» досить сильно
відрізняється (КФС = 31,2-47,9%). З 284 видових і внутрішньовидових таксонів
водоростей, виявлених в ставках Дідоровського, Китаївського і Оріхуватські
каскадів тільки 33 види є загальними, що складає всього 11,6% від загального
видового складу. Найбільш істотно видовий склад фітопланктону відрізнявся між
Дідоровських і Китаївський (КФС = 31,2%), a також Дідоровських і Оріхуватські
ставками (КФС = 37,7%). Дещо більшою схожістю характеризувався видовий склад
планктонних водоростей Китаєвських і Оріхуватські ставків (КФС = 47,9%).
Виявлені особливості свідчать o розходженні екологічних умов у формуванні
видового складу водоростей досліджених водойм.
Зокрема, ймовірно, що низька
різноманітність планктонних водоростей Горіховатських і Китаївських ставків
пов'язана з їх надмірним забрудненням, підтвердженням цьому слугує наявність в
них криптомонад. В той же час динофітові водорості, як відомо, здебільшого
мешкають в чистій воді. І саме Дідорівські ставки виявилися більш сприятливими
для їх розвитку. [31]
РОЗДІЛ ІІ. ЕТАПИ ДОСЛІДЖЕНЬ
ФІТОПЛАНКТОНУ КАСКАДУ ГОРІХУВАСТИХ СТАВКІВ
2.1 Визначення повного обсягу ставка
Розрахунок обсягу чаші ставка
проводиться в таблиці 6 на підставі вихідних даних варіанта (. У таблицю
записуються горизонталі ( чаші ставу до позначки НПУ, відстань між
горизонталями по верти-розжарюй (рівне h = 1 м) і площі водних дзеркал (F), обмежені
цими гори-зонталямі.2. Обсяг самого нижнього шару води (як фігура повного
конуса) визначаєтся по формулі:[29]
=1/3*Fn*h (тис.м3), (2.1)
де Fн - площа водного дзеркала,
обмежена горизонталлю 101 (тис.м2) h - відстань між горизонталями по вертикалі,,
рівне.1 м. Згідно вихідним даним F101 = 21,0 тис.м2, h = 1 м. Тоді V =113 F101
h = 21,0 V = 7,0 тис.м3. Для решти вище розташованих шарів води ставка обсяг
підраховується за формулою:
=1\3* (F1 + F 2) * h (тис.м3), (2.2)
де F1 і F2 - відповідно площі водного
дзеркала ставка, обмежені горизонталями, тис.м2.Згідно з вихідними даними F101
= 21,0 тис.м2, F102 = 47,6 тис.м2, h = 1 м, Тоді V =113 (F101 + F102) h = (21,0
+ 47,6) 1 = 34,3 тис.м.2 Аналогічно визначаються обсяги інших шарів води.4.
Об'єм чаші ставу до будь горизонталі складається з відповідного числа шарів
води.
Наприклад, до горизонталі 100 немає
ніяких шарів води та об'єм чаші буде дорівнює 0. До горизонталі 101 він
складається тільки з одного шару води, укладеного між горизонталями 100 і 101 і
дорівнюватиме його о бсягом, тобто 7,0 тис.м3. До горизонтами 102 ця величина
буде складатися зобсягів шарів між горизонталями 100 і 101 і горизонталями 101
і 102, то 7,0 + 34,3 = 41,3 тис.м3 і т.д. Повний об'єм чаші ставка при її
заповненні до позначки НПУ становить Vпол = 627,5 тис.м3.[28]
2.2
Визначення прозорості ставка
Прозорість води в гідрології та
океанології - це відношення інтенсивності світла, що пройшло через шар води, до
інтенсивності світла, що входить в воду. Прозорість води - величина, побічно що
позначає кількість зважених часток і колоїдів у воді. Прозорість води
визначається її виборчої здатністю поглинати і розсіювати світлові промені і
залежить від умов освітлення поверхні, зміни спектрального складу і ослаблення
світлового потоку, а також концентрації та характеру живої і неживої суспензії.
Класичний польовий метод визначення прозорості в глибоких водоймах - по глибині
зникнення з виду плоского диска білою або чорно-білого забарвлення діаметром
20-40 см (диска Секкі), запропонованого як стандартний метод італійським
священиком і астрономом Анджело Секкі. Його опускають на таку глибину, щоб він
повністю зник з виду, ця глибина і вважається показником прозорості в
гідрології та океанології. Для більш точного визначення записують два відліки:
глибину зникнення і глибину появи диска знову при піднятті троса. Середня
величина цих значень приймається за відносну прозорість води в даному районі.
Враховуючи специфіку завдання -
відбір проб фітопланктону в польових умовах, слід зауважити наступні глибини
- 0,5м
- поверневий шар
- 1
глибина непрозорості
- 2
глибини непрозорості
- 3
глибини непрозорості
В залежності від розмірів
досліджуваної водойми. В данному випадку використовувалися глибини 0,5 м та 1
глибина непрозорості.
Після чого проби зливають в одну
посудину і дитально перемішують
2.3 Відбір
проб
Отримання репрезентативних даних для
оцінки структурно-функціональних характеристик фітопланктону і динаміки їх змін
вимагає подекадного відбору проб. Важливо, щоб він проводився у чітко встановлений
час. Найбільш оптимальним є інтервал з десятої до дванадцятої години.
Одну пробу фіксують, а іншу
використовують для вивчення водоростей у живому стані. Здійснювати ці роботи
вкрай необхідно, оскільки при фіксуванні можливе пошкодження деяких морфологічних
структур водоростевих клітин (джгутиків, різних виростів тощо), що є
характерними систематичними ознаками, особливо у вольвоксових, перидинієвих,
евгленових і золотистих водоростей. [1]
Проби для кількісного визначення
фітопланктону відбирають батометром. Найпоширенішим є батометр Руттнера. З
огляду на те, що при дослідженні фітопланктону, як правило, відбирають
гідрохімічні, мікробіологічні і токсикологічні проби - найбільш прийнятним є
об’єм батометра 3-5 дм3. На мілководних станціях, де глибини не перевищують 2,0
м, можливий відбір лише з одного горизонту, як правило 0,2-0,3 м. Проби
фітопланктону відбирають і зберігають у скляних пляшках чи поліетиленових
флягах, відкаліброваних на 0,5 і 1,0 дм3 і щільно закритих кришками. Ще в
лабораторії посуд добре миють з використанням миючих засобів (для
поліетиленових фляг) чи хромової суміші (для скляних пляшок). Перед наповненням
чистий посуд слід 2-3 рази промити (100-200 мл) відібраною пробою.[2]
Всі пляшки (фляги) мають бути з
етикетками. Можливо кілька варіантів: а) на посуд масляною або емалевою фарбою
наносять цифрову нумерацію; б) на посуд наклеюють медичний пластир, на якому
олівцем або кульковою ручкою роблять відповідний запис; в) перед відбором проби
напис робиться безпосередньо на посуді стеклографом. Етикетку на флязі з пробою
підписують на станції відбору проб. Окремо в карточці обов’язкових відомостей
(польовий щоденник) дослідники записують всі необхідні дані щодо відбору проби:
найменування водойми, номер станції, її координати на водоймі та відповідна
географічна «прив’язка» станції, дата відбору проби (число, місяць, рік, час
доби), прозорість води, об’єм проби, температура води і повітря, кількість
кисню, гідрометеорологічні дані - стан погоди, наявність чи відсутність на
поверхні води ознак «цвітіння», спричиненого масовим розвитком водоростей,
плівок нафтопродуктів, сміття, візуально відмічених джерел надходження стічних
вод у водойму, звалищ сміття в районі природоохоронних смуг досліджуваної
водойми.[3]
2.4
Консервація проб
Консервація формальдегідом. Найбільш
поширеним консервантом є формальдегід. Для консервації у водні проби додають
40%-ний формальдегід з розрахунку 1:100, пляшку щільно закривають кришкою і
ставлять у темний ящик. Незважаючи на простоту і доступність цього методу, дія
формальдегіду, який є «жорстким» фіксатором, на водоростеві клітини може
призводити до їх деформації, відкидання джгутиків, виходження монадних форм
(евгленові, динофітові, золотисті) з будиночків, а у зелених хлорококових
можливий і лізис клітин. Фактично до 20-25% вихідної кількості водоростей через
3-5 міс. руйнується під дією формальдегіду. Водночас фіксація ним не впливає на
морфологічну структуру діатомових і синьозелених водоростей. Пояснюється це
тим, що перші з них мають кремнеземний панцир, а у других - клітини огорнуті
слизом.
Консервація розчином Люголя. Розчин
Люголя є більш «м’яким» фіксатором, що не руйнує морфологічну структуру
водоростей. Але він не завжди «вбиває» водяну мікрофлору і водяні гриби. Це
призводить до того, що через 1-2 міс. зафіксована проба починає «загнивати» і
практично руйнуються клітинні структури.[4]
Консервація етиловим спиртом.
Етиловий спирт є також «м’яким» фіксатором. Використовують спирт етиловий
ректифікований, який додають до проби у співвідношенні 1:10. Проби фітопланктону,
зафіксовані спиртом, не можна зберігати понад 1-1,5 місяця. Надалі
неопрацьовані проби починають «загнивати».
Консервація хромовими квасцями.
Комбінація розчину хромових квасців з розчином формальдегіду добре відображає
морфологічну структуру планктонних водоростей і забезпечує тривале збереження
альгологічних проб незалежно від видового складу фітопланктону . [8]
2.5
Згущення проб
Всі існуючі методи згущення проб
базуються на одному з трьох фізичних процесів: седиментації, центрифугуванні та
фільтрації через мікропористі фільтри.
Метод седиментації. Пляшки (фляги) з
альгологічними пробами охайно, без струшування виставляють у темному
прохолодному місці. Через 10-12 днів воду над водоростями, що осіли, збирають
спеціальним сифоном, залишаючи над осадом шар води 5-8 см. Залишок проби
(об’ємом не більш ніж 100 мл) переливають у посуд меншої місткості, відстоюють
протягом 5-7 діб і повторно відсифонюють, доводячи кінцевий об’єм до 10 см3.
Проби переливають у пеніцилінові склянки, додають 2-3 краплі формальдегіду чи
розчину Люголя і починають камеральне опрацювання.
Метод центрифугування. Найбільш
швидкий метод згущення альгологічних проб. Для достатнього осадження
фітопланктону потужність центрифуги має становити не менш як 1 500-3 000 об/хв.
Але при цьому можлива втрата значної кількості водоростей різних таксонів через
розчинення осаду та при його перенесенні до лічильної камери.[9]
Метод фільтрації. Портативним,
швидким методом згущення альгологічних проб (до 200 разів від об’єму відібраної
проби) є метод їх фільтрації через дрібнопористі фільтри. Простота апаратного
забезпечення: колба Бунзена місткістю 1,0-2,0 дм3, фільтрувальна воронка,
дрібнопористі фільтри, вакуумна гумова трубка, вакуумний насос, що створює
розрідження до 0,5-3,0 атм - дозволяє використовувати метод фільтрації в
експедиційних умовах. Різновидністю методу фільтрації з використанням
вакуумного розрідження є метод фільтрації під тиском. Недоліком обох методів є
втрата при фільтруванні нанопланктонних видів водоростей і можливе пошкодження
їх морфологічних структур, що є основними систематичними ознаками виду.[6]
.6 Камеральне
опрацювання проб
У гідроекологічних дослідженнях для
опрацювання альгологічних проб використовуються світлові та електронні скануючі
і трансмісійні мікроскопи різних марок як вітчизняного, так і зарубіжного
виробництва. Основна вимога до мікроскопа - це величина збільшення. Для
отримання репрезентативних результатів окуляр повинен мати збільшення не менше
як К5, а об’єктив - 20.
Чисельність водоростей підраховують
у спеціальних лічильних камерах. Найбільш поширеною в альгологічних
дослідженнях є камера Нажотта об’ємом 0,01-0,05 см3. Використання інших камер,
наприклад лічильної камери Горяєва, в якій підраховуються формені елементи
крові, небажане, оскільки крупні водорості планктону, особливо колоніальні
форми, не вміщуються на дні камери. При використанні камери Горяєва отримані
результати значно занижені. Можливе також застосування «лічильних пластин»
.[12]
2.7 Визначення чисельності і біомаси
Для оцінки кількісної
різноманітності фітопланктону обчислюють його чисельність і біомасу.
Чисельність фітопланктону
розраховують на 1 дм3 (1 л) води за формулою:
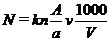 ,(2.3)
,(2.3)
фітопланктон органічний
склад ставок
де N - кількість водоростей в 1 дм3
води досліджуваної водойми (як правило, тис. кл/дм3 або млн. кл/дм3); k -
коефіцієнт, що показує, у скільки разів об’єм використаної камери менший за 1
см3; n - кількість клітин водоростей на проглянутих доріжках (квадратах)
лічильної камери; А - кількість доріжок (квадратів) лічильної камери; а -
кількість доріжок (квадратів), де підраховувалась кількість водоростей; V -
об’єм проби фітопланктону, взятий на водоймі, см3; v - об’єм концентрованої
проби, з якого розраховуються показники фітопланктону, см3.[13]
Біомаса фітопланктону визначається
розрахунково-об’ємним методом. Його використання передбачає наявність даних по
чисельності конкретного виду водоростей у пробі та лінійних розмірів його
клітин. Для визначення розмірів водоростей їх прирівнюють до певних
геометричних тіл, найбільш подібних до даної морфологічної форми: куля,
паралелепіпед, циліндр, конус, октаедр тощо. Далі вимірюють необхідні
параметри: радіус, діаметр, висоту, довжину тощо. Для отримання
репрезентативних даних необхідно виміряти параметри не менш як 30 водоростевих
клітин одного виду. Одержані дані опрацьовують статистично .
Об’єм клітин визначають за відомими
геометричними формулами, використовуючи лінійні розміри конкретної водорості,
подібної до певної геометричної фігури. Припускають, що відносна щільність (до
води) прісноводних водоростей становить 1,00-1,05. Обчислену біомасу кожного
виду множать на його чисельність і наводять у мг/дм3, г/м3 або г/м2. [18]
.8 Визначення первинної продукції
фітопланктону і деструкції органічних речовин
Інтенсивність первинної продукції
залежно від того, який з інгредієнтів процесу фотосинтезу ми вимірюємо
(наприклад, вміст кисню чи фотосинтезованої органічної речовини), може суттєво
відрізнятися. Для відносної формалізації показників, що характеризують первинну
продукцію, умовно було виділено кілька її форм. Запропоновані форми первинної
продукції - це відносно віртуальні характеристики, що визначають реально
існуючі потоки енергії в екосистемах:
валова первинна продукція (АВ) - це
вся енергія, що утворилась фітопланктоном у процесі фотосинтезу в екосистемі;
ефективна первинна продукція (АЕФ):
АЕФ = АВ - RФ , де RФ - енергетичні витрати на дихання (метаболізм) водоростей;
чиста первинна продукція, або
фактично наявна в екосистемі біомаса (B) фітопланктону (АЧ): АЧ = АВ - R, де R
- сумарні енергетичні витрати на дихання (метаболізм) усіх компонентів
планктону: R = RФ + RЗ + RБ + Rі, де RФ - енергетичні витрати на дихання
водоростей; RЗ - енергетичні витрати на дихання зоопланктону; RБ - енергетичні
трати на дихання бактеріопланктону; Rі - енергетичні трати на дихання
джгутикових форм.
Важливим показником стану біоти є
величина АВ/Rдобу-1, яка характеризує співвідношення
продукційно-деструкційних процесів і дозволяє визначати надходження алохтонних
органічних речовин, а відповідно оцінювати ступінь антропогенного навантаження
на екосистему, знаходити місцерозташування джерел забруднення і визначати
величину енергетичної субсидії, що необхідна для функціонування конкретної
екосистеми.
Поряд із широким використанням
добового відношення А/R, можливий також аналіз цього показника протягом декади,
місяця, року, що дозволяє більш повно характеризувати стан біоти за даний
проміжок часу.[19]
Розглянемо найбільш поширені в
гідроекологічних дослідженнях методи визначення первинної продукції
фітопланктону.
Визначення первинної продукції за
кількістю клітин водоростей
Процедура цього методу така:
а) молібденові або кварцеві циліндри
(об’єм води - від 1 до 5 л) наповнюють пробами фітопланктону. Отвори циліндрів
щільно закривають млиновим газом № 76. Розмір вічка газу не дозволяє виходити
водоростям з посуду. Застосування молібденового або кварцового скла запобігає
поглинанню короткохвильової частини сонячного спектру, а отже наближає дослідні
умови до природних. Необхідна місткість експериментальних циліндрів
визначається інтенсивністю розвитку фітопланктону: чим більше в пробі
водоростей, тим меншою має бути місткість посуду;
б) не менш ніж два циліндри
використовуються як контрольні, в яких після їх заповнення і фіксації проб
підраховують кількість фітопланктону: чисельність (N0), тис. кл/дм3, або
біомасу (В0), мг/дм3, які є контролем даного досліду;
в) інші циліндри експонують у
досліджуваній водоймі. Експозиція (t) становить, як правило, одну добу;
г) після закінчення експозиції вміст
проби фіксують і камерально опрацьовують. Залежно від кількості використаних
циліндрів середній вміст водоростей у досліді становитиме:
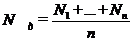 або
або 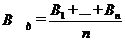 ,(2.4)
,(2.4)
де n - кількість експериментальних
циліндрів;
д) первинну продукцію водоростей
обчислюють за формулою
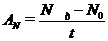 або
або 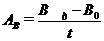 ,(2.5)
,(2.5)
відповідно у тис. кл/дм3 або мг/дм3
протягом експозиції (t).
Результати, одержані за цим методом,
найбільш повно характеризують чисту первинну продукцію фітопланктону (АЧ).[20]
Визначення первинної продукції за
динамікою вмісту біогенних елементів у воді. Метод базується на основному
рівнянні фотосинтезу, біохімічного процесу, який відбувається з включенням у
склад новоутвореної органічної речовини біогенних елементів. Відповідно чим
більш інтенсивно формується продукція, тим більше біогенних елементів
поглинається фітопланктоном. За різницею біогенних елементів в експериментальних
і контрольних склянках можна оцінювати продукцію водоростей.
Вперше цей метод був запропонований
В. Г. Дацко при вивченні продуктивності екосистем Азовського і Чорного морів.
Основні недоліки методу: швидка регенерація біогенних елементів при деструкції
(лізису) рослинних клітин та адсорбція біогенних елементів (особливо фосфатів)
на поверхні клітинної оболонки водоростей.[21]
Визначення первинної продукції за
добовою динамікою кисню. Суть методу полягає у тому, що формування первинної
продукції протягом доби характеризується різною інтенсивністю . Відповідно
вміст розчиненого у воді кисню, що є одним з кінцевих продуктів фотосинтезу,
змінюється пропорційно добовій динаміці останнього. Найменший вміст у воді
кисню реєструється в кінці темного періоду доби, коли фотосинтетична аерація
практично відсутня, найбільший, за даними наших спостережень, - між 13-ю і 16-ю
годинами.
Отже, інтенсивність первинної
продукції - це різниця між максимальними і мінімальними показниками, що
характеризують вміст розчиненого у воді кисню протягом доби.
На жаль, незважаючи на простоту
визначення продукції за добовою динамікою вмісту у воді кисню, цей метод має
ряд недоліків, а саме: методично дуже важко встановити максимальний і
мінімальний вміст у воді кисню; в процесі фотоаерації води при досягненні
насичення її киснем 100% і більше відбувається дифузія кисню в атмосферу,
особливо це характерно при високій температурі води. Встановити величину цього
показника методично складно. А застосування певних коефіцієнтів знижує репрезентативність
отриманих даних.
Очевидно, що метод найбільш
оптимально може використовуватись при постійній автоматичній реєстрації вмісту
кисню у воді за допомогою відповідних приладів.[22]
2.9 Оцінка
різноманітності фітопланктону
Досліджуються такі структурні
характеристики:- видова різноманітність - кількість видових і внутрішньовидових
таксонів, включаючи номенклатурний тип виду, їх співвідношення і частка в
загальній кількості таксонів;- надвидова різноманітність - кількість таксонів
рангом вище виду (рід, родина, порядок, відділ), їх співвідношення в пробі
(водоймі) на всіх рівнях зазначеної систематичної ієрархії.
При якісному визначенні видової і
таксономічної належності різних видів чи внутрішньовидових таксонів
фітопланктону та кількісної різноманітності для більш швидкого отримання
попередніх натурних даних можна застосовувати експрес-оцінку частоти трапляння
конкретних видів.
Однією з поширених у гідроекології є
шкала С. М. Вислоуха . Як аналог можна використовувати і шкалу Стармаха .
Одержані за допомогою експрес-оцінок
дані є попередніми і дозволяють швидко оцінити можливі зміни у видовій та
кількісній різноманітності фітопланктону, спричинені негативним впливом одного
чи кількох екологічних чинників.[3]
Таблиця 2.1
Шкала Вислоуха
|
Вид зустрічається масово, викликаючи
«цвітіння» води . . . . . . . . . . . . .
|
6
|
|
Дуже часто . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
|
5
|
|
Часто, більше 10 разів у препараті . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . .
|
4
|
|
Не рідко, менше 10 разів у препараті . . . . .
. . . . . . . . . . . . . . . . . . . . . . . .
|
3
|
|
Рідко, 2-4 рази у препараті . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . .
|
2
|
|
Дуже рідко, 1-2 рази у препараті . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . .
|
1
|
Не менш інформативним при аналізі
видової різноманітності, особливо при порівнянні таксономічного складу різних
ділянок водойми, кількісному визначенні спільностей та відмінностей, є
коефіцієнт видової подібності фітопланктону (K) двох порівнюваних водойм чи
двох ділянок однієї водойми:
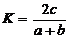 ,(2.6)
,(2.6)
де а - кількість видів у першій
водоймі (ділянці); b - кількість видів у другій водоймі (ділянці); с -
кількість спільних видів.[24]
Коефіцієнт видової подібності
змінюється від 0 до 1. Якщо K = 0,5, то видова різноманітність (видове
багатство) фітопланктону двох порівнюваних водойм (ділянок) досить схожа і
відповідно негативний вплив екологічних чинників (не лише антропогенних, а і
природних) незначний. Якщо K = 0,5, то видова різноманітність фітопланктону
порівнюваних водойм суттєво відрізняється, а отже, і екологічні умови, що
визначають розвиток фітопланктону, різні.
Важливими кількісними показниками,
що дозволяють характеризувати таксономічну різноманітність, є співвідношення
видової, внутрішньовидової, родової або різноманітності родин водоростевих
угруповань:
відношення кількості видів і
внутрішньовидових таксонів;
відношення кількості родів і видів;
відношення кількості родин і родів.
Вищенаведені показники
характеризують зміну таксономічної різноманітності на відповідних рівнях
систематичної ієрархії. Їх використання дозволяє оцінити вегетацію водоростевих
угруповань залежно від впливу антропогенних або інших чинників у водних
екосистемах.
Оцінка стану біоти за інформаційною
різноманітністю фітопланктону
Інформаційна різноманітність
визначається за індексом Шеннона (Н) . Для його розрахунку використовують два
показники: кількість видів і чисельність (НN) або кількість видів і біомасу
(НB). З огляду на різний об’єм клітин водоростей, що може відрізнятися на
кілька порядків, інформаційну біорізноманітність слід визначати як за
чисельністю, так і за біомасою.
При Н=2 фітопланктон більш
різноманітний. Домінуючий комплекс представлений полідомінантними
угрупованнями. Це свідчить про якість водного середовища, що наближається до
оптимального для розвитку планктонних водоростей, а отже про відсутність або
незначний негативний вплив антропогенних чинників, що не призводять до
деградації фітопланктону.[25]
При домінуванні 1-2 видів, коли
чисельність (біомаса) домінуючого виду становить 50% і більше від сумарної
фітопланктону, величина Н знижується. При Н=1 фітопланктон, як правило,
представлений монодомінантним або олігодомінантним комплексом. Прикладом є
«цвітіння» води синьозеленими водоростями та інтенсивний розвиток 1-2 видів,
найбільш стійких до антропогенного впливу, спричиненого різними типами
забруднювачів (неочищені господарсько-побутові стічні води, нафтопродукти,
важкі метали, поверхнево-активні речовини тощо).
Оцінка стану водних об’єктів за
інтенсивністю розвитку фітопланктонузаг - кількісна різноманітність за
чисельністю, кл/дм3, тис. кл/дм3, млн. кл/дм3:
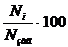 (2.7)
(2.7)
характеристика структури чисельності
угруповань і частка конкретної систематичної групи водоростей (Ni) у формуванні
сумарної чисельності фітопланктону;
Взаг - кількісна різноманітність за
біомасою, мг/дм3, г/м3 , г/м2:
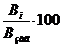 (2.8)
(2.8)
характеристика структури біомаси, те
ж саме, що і для чисельності.
Угруповання фітопланктону
розглядаються як один із найважливіших «біологічних елементів якості для
класифікації екологічного статусу» водних об’єктів різного типу. Рекомендується
використовувати для цього статусу такі характеристики фітопланктону, як
таксономічних склад і чисельність (з урахуванням явища «цвітіння» води .
Чисельність фітопланктону (для змішаного складу та з домінуванням синьозелених водоростей),
поряд з біомасою фітопланктону, а також функціональними показниками - вмістом
хлорофілу а і добовою валовою первинною продукцією, була використана у
дев’тирозрядній кількісній класифікації стану континентальних водних об’єктів
України, за гідробіологічними показниками, розробленій в Інституті
гідробіології НАН України . Ця класифікація є складовою частиною класифікації
стану водних об’єктів за гідробіологічними показниками (розділи 12 та 26 цього
збірника).
Ця класифікація важлива для оцінки
стану водних екосистем за структурно-функціональною організацією фітопланктону
і трофністю. [26]
РОЗДІЛ ІІІ. ВИЗНАЧЕННЯ ЧИСЕЛЬНОСТІ
ТА ДИНАМІКИ БІОМАСИ ФІТОПЛАНКТОНУ
3.1 Визначення
повного обсягу досліджуваних водойм
Визначення повного обсягу ставка
відбувалося враховуючи наступні вихідні данні :
1.Максимальна глибина ставка -
експерементальні данні, враховуючи що точка максимальної глибини ставка
знаходиться близько до його центру.
. Площа ставка виміряна згідно
аналізу супутникових зображень заданого масштабу за допомогою программи
Universal Desktop Ruller(c).
. Площа окремо визначених
горизонталей [40]
Таблиця 3.1
Розрахунок об’єму водойми за поділом
на водні горизонталі
|
Нижній Орехуватський ставок
|
Середній Орехуватський ставок
|
Верхній Орехуватський ставок
|
|
Загальна площа S (m2)
|
22084
|
11150
|
13380
|
|
Максимальна глибина H (m)
|
4,1
|
3,2
|
3,1
|
|
Об’м (m3)
|
30764
|
9848
|
10993
|
|
Водний горизонталь 2 9(m2)
|
18075
|
8080
|
9016
|
|
Водний горизонталь 3 (m2)
|
13054
|
4317
|
4584
|
|
Водний горизонталь 4 (m2)
|
9081
|
--
|
--
|
|
Об’єм води між горизонталями (m3) 1-2
|
7361
|
3716
|
4460
|
|
Об’єм води між горизонталями (m3) 2-3
|
6025
|
2693
|
3005
|
|
Об’єм води між горизонталями (m3) 3-4
|
4351
|
1439
|
1528
|
|
Об’єм води між горизонталями (m3) 4-5
|
3027
|
--
|
--
|
Таблиця 3.2
визначення об’єму водойми за
допомогою аппроксимації її геометричної форми за формулою V= 1/3*H* S
|
Нижній Орехуватський ставок
|
Середній Орехуватський ставок
|
Верхній Орехуватський ставок
|
|
Загальна площа S (m2)
|
22084
|
11150
|
13380
|
|
Максимальна глибина H (m)
|
4,1
|
3,2
|
3,1
|
|
Об’м з аппроксимацією(m3)
|
30181
|
11893
|
13826
|
Рис. 3.1 Графік, що відображає
похибку при визначенні об`єму ставків різними методами. (червоний колір-
визначення за водними горизонталями, синій колір - визначення експрес методом)
З графіку можна зробити висновок, що
у багатьох частних випадках швидкий метод визначення об’єму досліджуваної
водойми является ефективним і саме в цих випадках методика не потребує
визначення водних горизонталей.
3.2
Визначення прозорості ставка
Таблиця 3.3
Глибина непрозорості досліджуваних
водойм (м)
|
Ставок
|
Верхній Горіхуватський
|
Середній Горіхуватсякий
|
Нижній Горіхуватський
|
|
Місяць
|
|
|
|
|
Липень
|
110
|
125
|
105
|
|
Серпень
|
145
|
135
|
115
|
|
Вересень
|
125
|
165
|
125
|
Слід зауважити що показники сильно
залежать від погодних умов, загальної освітленості, часу відбору проби. Для
відбору проб використовувались глибини 0,5 м та 1 глибина непрозорості.
3.3
Визначення чисельності і біомаси
Визначення чисельності і біомаси
відбувалося враховуючи наступні вихідні данні:
Площа камери -- 9 мм2
Об’єм -- 0,9 мм3
Коофіціент k - 0,9
Об’єм досліджуваної проби --V
3000см3
Об’єм концентрованої проби -- v 1
см3
Кількість клітин - n
Використані доріжки - а [12]
Таблиця 3.4
Кількість клітин фітопланктону в
перерахунку млн. кл./дм3
|
Cyanoprocariota
|
Chlorophyta
|
Basilariophita
|
Evglenophyta
|
Dynophita
|
Xantophyta
|
Crysophita
|
Creptophyta
|
Streptophyta
|
Сумма
|
|
Липень
|
13
|
54
|
27
|
11
|
7
|
7
|
6
|
6
|
8
|
141
|
|
Серпень
|
16
|
67
|
33
|
14
|
9
|
10
|
7
|
8
|
10
|
177
|
|
Вересень
|
10
|
40
|
20
|
8
|
5
|
5
|
4
|
5
|
6
|
106
|
З даної таблиці можемо зробити
висновок, що у даних водоймах (каскад Орыхувастих ставків) впевнено домінуе
відділ Chlorophyta та Basilariophita.
Рис.3.2 гістограма чисельності
клітин фітопланктону досліджуваної водойми
3.4 Оцінка
різноманітності фітопланктону
Оцінка стану водних об’єктів за
інтенсивністю розвитку фітопланктонузаг - кількісна різноманітність за
чисельністю, кл/дм3, тис. кл/дм3, млн. кл/дм3:
 - характеристика структури
чисельності угруповань і частка конкретної систематичної групи водоростей (Ni)
у формуванні сумарної чисельності фітопланктону;[6]
- характеристика структури
чисельності угруповань і частка конкретної систематичної групи водоростей (Ni)
у формуванні сумарної чисельності фітопланктону;[6]
Таблиця 3.5
Характеристика структури чисельності
угруповань у відсотках.
|
Cyanoprocariota
|
Chlorophyta
|
Basilariophita
|
Evglenophyta
|
Dynophyta
|
Xantophyta
|
Crysophyta
|
Creptophyta
|
Streptophyta
|
|
Липень
|
9,2
|
38,2
|
19,1
|
7,8
|
4,9
|
4,9
|
4,2
|
4,2
|
5,6
|
|
Серпень
|
11,3
|
47,5
|
23,4
|
9,9
|
6,3
|
7
|
4,9
|
5,6
|
7
|
|
Вересень
|
7
|
28,3
|
14,1
|
5,6
|
3,5
|
3,5
|
2,8
|
3,5
|
4,2
|
Експрес оцінка за шкалою Вислоуха
дозволяе спрощено і дуже швидко оцінити стан фітопланктону в досліджуваній
водоймі:
Таблиця 3.6
Балли за шкалою Вислоуха
|
Cyanoprocariota
|
Chlorophyta
|
Basilariophita
|
Evglenophyta
|
Dynophita
|
Xantophyta
|
Crysophita
|
Creptophyta
|
Streptophyta
|
|
Липень
|
4
|
6
|
6
|
4
|
3
|
3
|
3
|
3
|
3
|
|
Серпень
|
4
|
6
|
6
|
4
|
3
|
4
|
3
|
3
|
4
|
|
Вересень
|
4
|
6
|
6
|
3
|
3
|
3
|
2
|
3
|
3
|
За даною шкалою можемо зробити
висновок що відділи Chlorophyta та Basilariophyta викликають процесс цвітіння
на протязі усьго досліджуваного періоду часу.
3.2 Визначення
біомаси фітопланктону за кількістю клітин водоростей
Фітоплактон найбільш точно визначає
трофічний рівень водоймища. Наприклад, для оліготрофних і мезотрофних вод
характерно низьке відношення чисельності фітопланктону до його біомасі, а для
гіпертрофних - високе. Біомаса фітопланктону в гіпертрофних водоймах становить
більше 400 мг / л, в евтрофних - 40,1-400 мг / л, в дістрофних - 0,5-1 мг /
л.Підвищення евтрофікації, або надмірне накопичення у водоймі органічної
речовини, тісно пов'язане з посиленням процесів фотосинтезу в фітопланктоні.
Масовий розвиток водоростей призводить до погіршення якості води, її «цвітіння».Цвітіння
- не стихійне явище, воно складений протягом досить тривалого часу, іноді двох
і більше вегетаційних періодів. Передумови різкого зростання чисельності
фітопланктону - наявність водоростей у водоймі та їх здатність до розмноження
при сприятливих умовах. Розвиток діатомових, наприклад, в значній мірі залежить
від вмісту у воді заліза, лімітуючим фактором для зелених водоростей служить
азот, синьо-зелених - марганець. Цвітіння води вважається слабким, якщо біомаса
фітопланктону знаходиться в межах 0,5-0,9 мг / л, помірним - 1-9,9 мг / л,
інтенсивним - 10 - 99,9 мг / л, а при гіперцвітінні вона перевищує 100 мг /
л.Визначення біомаси фітопланктону - трудомісткий і тривалий процес. На
практиці для полегшення розрахунку умовно прийнято вважати, що маса 1 млн.
клітин прісноводного фітопланктону приблизно дорівнює 1 мг.
Таблиця 3.7
Розрахунок концентрації клітин
водоростей у заданому об`ємі.
З урахуванням даних отриманих з проб
на початку та у кінці кожного з місяців (липнь- серпень). Розрахунок концентрації
фітопланктону проводиться за формулою: n=N/V.
|
Об`єм (дм3)
|
55900000
|
|
|
|
|
|
|
Кількість клітин водоростей (млн. кл)
|
121
|
161
|
170
|
183
|
135
|
76
|
|
Біомасса (мг\л)
|
121
|
161
|
170
|
183
|
135
|
76
|
|
Концентрація (1/дм3)
|
2,1645796
|
2,8801431
|
3,0411449
|
3,273703
|
2,4150268
|
1,359571
|
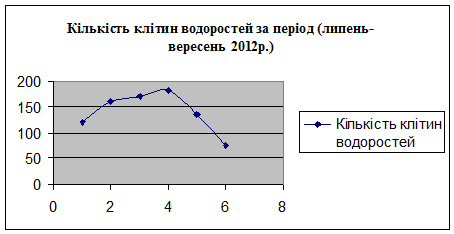
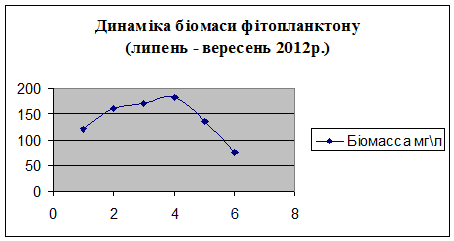
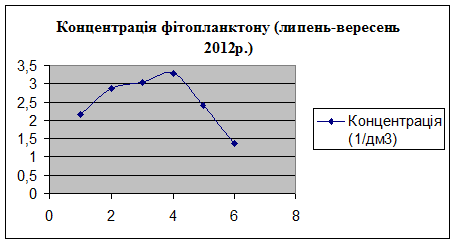
Рис. 3.3 Співвідношення кількості
клітин (млн.\дм3) до біомаси фітопланктону (мг\л) та концентрації клітин
водоростей у заданому об`эмі води. Відслідковування динаміки цих параметрів.
З мал.. 3.3 можемо зробити висновок,
що кількість клітин водоростей а також їх біомаса відповідає рівням
гіперцвітіння та інтенсивного цвітіння з наведеної вище класифікації. Останні
відмітки на початку та в кінці вересня свідчять про зниження рівня евтрифікації
в каскаді ставків.
РОЗДІЛ 4. ЕКОНОМІЧНЕ ЗНАЧЕННЯ
ПРОЦЕСІВ ФОТОСИНТЕЗУ У СВІТОВОМУ ОКЕАНІ
4.1
Глобальні процеси фотосинтезу у світовому океані
Фотосинтез в світовому океані -
фактор, який грає головну роль в циклі CO2, впливає на глобальні процеси
нагрівання, тому він представляє великий інтерес фахівців, що створюють моделі.
Системи, змодельовані в даній роботі, покладаються з визначеними у просторі
кордонами, через які на систему може чинитися зовнішнє вплив. Просторове
положення системи передбачається як контрольований обсяг, тому що необхідно
зберегти масу в системі. Головна мета водного моделювання - передбачити
поведінку та реакцію системи на зовнішній вплив.
Еутрофікація - тривожний процес, що
відбувається в системі. Його необхідно вміти передбачити і контролювати за
допомогою зовнішнього екологічного впливу. А щоб управляти цим процесом,
необхідно визначити положення системи, яке, ймовірно, призведе до еутрофікаціі
і з'ясувати, приь яких обставин це станеться. Прогнозування вимагає, щоб
структура моделі та дослідного зразка залишилися сумісні на проміжку часу. Бути
може, можна побудувати модель, що враховує всі зовнішні фактори, але самий акт
моделювання є спрощенням і практично завжди будуть наближення, які обмежують
діапазон точної прогнозуючої здатності моделі. Модель не повинна бути більш
складною, ніж необхідно, щоб забезпечити передбачення достатньої точності. За
минулі десять років в океанічних моделях велику увагу було приділено мікробному
циклу, в якому відбувається взаємодія між бактеріями і різними живильними речовинами.
У дрібних озерах і гирлах, куди світло проникає добре, бентічних рослини -
важлива частина екосистеми, забезпечує їжу і притулок для маленьких і великих
організмів і захищає дно від ерозії, яке в свою чергу забезпечує середовище
проживання для бентіческіх фауни. Довгий час темою для обговорення було поняття
лімітую чого поживної речовини, тобто речовини, яка знаходиться в недоліку.
Ідея була одним з найважливіших понять в контролі за екосистемою.
Редфилд встановив, що в земних
системах такою речовиною є фосфор, а у водних - азот. Однак це часто
піддавалося сумніву. Хек, зокрема піддали сумніву, обмежував чи азот взагалі в
морських системах. Вуглець і водень, які є основними складовими атомами
більшості органічних речовин, доступні для автотрофів у водному навколишньому
середовищу в основному через об'єднаний C02 і воду безпосередньо. В морських
системах концентрація C02 близько 0,3 mg / l, якої цілком достатньо для
фотосинтезу. Кисень так само доступний в об'єднаній формі, але, звичайно,
потрібно тваринам і рослинам в елементарній формі для дихання, його присутність
або відсутність керує багатьма реакціями між іншими поживними речовинами у
відкладеннях і в деякій мірі у водному тощо.
Азот - істотний компонент багатьох
молекул, амінокислот, нуклеїнових кислот і т.д., і його морська концентрація
приблизно 300 mg / l.Фосфор відіграє провідну роль у фотосинтетичних процесів у
системі передачі енергії ATP / ADP. Сірка, звичайно менш важлива, є компонентом
амінокислот і грає істотну роль у анаеробної бентіческіх навколишньому
середовищу. У морській воді концентрація сірки приблизно 30 mg / l. Залізо,
концентрація якого в морській воді 0,1 - 62 mg / l з середньою концентрацією
3,1 m g / l, було центром значного дослідження в останні роки. Ряд
експериментів вказав, що залізо відіграє важливу роль як обмежує поживна
речовина в деяких областях. Кремній - ключовий елемент для фітопланктону,
особливо діатомових водоростей, які характеризувати кремнієвими структурами,
названими фрустули. У другому розділі глави описані основні хімічні процеси,
відбуваються з азотом. Це аммонификация, асиміляція аміаку, нітрифікація і
денітрифікація.
Аммонификация - процес перетворення
органічних форм азоту до аміаку. Аммонификация здійснюється багатьма рослинами,
тваринами і мікроорганізмами, включаючи гриби і особливо протеолітичні
бактерії. Типовий приклад процесу - утворення аміаку з сечовини:
2 (CO) NH 2 + H 2O = 2 NH 2 +
CO2(4.1)
У наступному пункті розглянуто цикл
фосфору. Цей елемент відіграє важливу роль в нуклеїнових кислотах і в системі
передачі енергії ATP / ADP. Фосфоліпіди є компонентами мембран клітин. Згідно
відношенню атоми C: N: P у тканинах водоростей пов'язані співвідношенням
106:16:1 або 42:7:1 - у вазі. З цього випливає, що якщо концентрація азоту
перевищує концентрацію фосфору більш ніж у сім разів, то недолік фосфору буде
лімітувати розвиток організмів. Інші вказують, що відношення азоту до фосфору в
річковій воді по всьому світу в середньому 230:20 (11.5:1), а в океанах в
середньому 250:35 (7:1). Таким чином, в прісноводних системах фосфор буде
лімітуючим елементом (якщо немає недоліку інших мікроелементів). Було багато
спроб встановити відповідність між концентрацією фосфору в системі і біомасою
рослин. Пізніше інші дослідники запропонували зв'язок між фітопланктоном і
концентрацією фосфору. Рослини, морські водорості і багато гетеротрофні
бактерії включають сірку у вигляді сульфатів в органічні молекули. У аеробної
навколишньому середовищу ці органічні форми перетворяться до H2S, потім
окислюються до сере і нарешті перетворюються в сульфат багатьма фотосинтетичної
і нефотосинтезуючими бактеріями (Thiobacillus). Навпаки, анаеробні бактерії
(Desulfovibrio) здатні скорочувати кількість іонів сульфату при анаеробному
диханні. Це призводить до накопичення в затопленій грунті і на глибині H2S. Це
речовина реагує з металами, такими як залізо і цинк (що призводить до
скороченню заліза - важливого поживної речовини). Цей процес очевидний в мутній
воді, тому з'являється запах H2S і вода очорняти сульфідами заліза. Основний
компонент фотосинтетичного апарату в морських водоростях - пластид, званий
хлоропластів, який містить дископодібні перетинчасті бульбашки, названі
тилакоїди. Останні містять області хлорофілу для фотосинтезу, в той час як
вуглекислий газ фіксується в навколишнього матриці - стромі.. Ціанофіти
Ціанофіти також згадуються як ціанобактерії-менш формально як синьо-зелені
водорості. Синьо - зелені водорості особливо цікаві в прибережних і внутрішніх
областях через їх потенційної токсичності, і в глибоких океанах вони відіграють
істотну роль у первинній продуктивності планктону. В Австралії і багатьох інших
частинах світу, розквіти синьо - зелених водоростей викликали серйозні проблеми
і в постачанні питною водою, в сільському господарстві. Здатність морських
водоростей фотосинтезировать є головним фактором у їх здатності конкурувати з
іншими водорослево різновидами. У широкій термінології фотосинтез відбувається
в присутності хлорофілу, в Відповідно до рівняння (6), у великих органелах
рослини, названих хлоропластами.
CO2 + 12 H 2O + світло = 6O2 + C6
H12O6 + 6 H 2O(4.2)
У результаті реакції, описаної
рівнянням (4.2), утворюється вуглевод, який в Надалі може використовуватися в
реакціях синтезу макромолекул, які є «будівельним матеріалом» органоїдів і
більш складних клітиннихкомпонентів. Як і у наземних рослин, процес фотосинтезу
у морських водоростей розділений на дві фази. Перша фаза - процес, в якому в
результаті фосфорилювання утворюється ATP (аденозин трифосфат). У другій фазі
енергія, що міститься в зв'язку фосфату в ATP використовується, щоб справити
карбоксилювання в циклі Кальвіна. Реакції проходять в спеціальних органелах,
названих тилакоїди, які у еукаріотів знаходяться в оточених мембраною
пластидах, а в прокаріот розташовані в цитоплазмі.
Рівняння можна розділити на наступні
реакції:
H 2O = O2 + 4e - + 4 H + (необхідна
сонячна енергія) 2 NADP + + 4e-+ 2 H + = 2 NADPH + +2 NADPH + 2 H + CO2 = 2
NADP + H 2O + (CH 2O)
Якщо зібрати ці рівняння разом,
вийде:
+ H 2O = (CH 2O) + O2 (необхідне
світло)
Тут NADPH - нікотинамід аденін
динуклеотид фосфат. Те, що не показано в останньому рівнянні, є процес
фосфорилювання, який може протікати в циклічному або нециклічні процесі.
Визначають рівняння нециклічного процесу таким чином:
NADPH + H 2O + 2 ADP + 2 Pi = NADPH
2 + 2 ATP + O2
де Pi - неорганічний фосфор. ATP
надає енергію для реакцій допомогою наступних процесів:
NADPH 2 + O2 = 2 NADP + 2 H2O 3 ATP
+ H2O = 3ADP + 3Pi
які в загальному можна записати:
+ H 2O + 2 NADPH 2 + 3 ATP = [CH 2O]
+ O2 + 2 NADP + 3 ADP + 3Pi
У циклічному фосфорилировании ATP -
єдиний продукт процесу:
+ Pi = ATP (необхідне світло)
Дихання в рослинах (і фітопланктоні)
- це процес, в результаті якого відбувається окислення накопиченого органічної
речовини, щоб рослина отримало енергію. У результаті виділяється вуглекислий
газ. Особливо цікавий факт, що біомаса рослин зменшується в процесі дихання.
Процес цей до кінця не зрозумілий, але B досліджували дані, наведені в різних
літературних джерелах, і прийшли до висновку, що в процесі дихання втрачається
до 13% біомаси, накопиченої в процесі фотосинтезу. Відпрацьована біомаса
переробляється бактеріями. В останні роки було виявлено виділення діатомових
водоростей прозорих екзополімеров. Дінофлагеллати також виділяють полісахариди,
які призводять до великих скупчень слизу, що згубно впливає на фауну. У
дев'ятому пункті глави розглядається взаємовідношення планктону і поживних
речовин. Автори вважають важливим розглянути кінетику процесів поглинання і
засвоєння поживних речовин кліткою. Але також вони відзначають, що досі не
створено широкої теоретичної моделі боротьби за ресурси, на основі якої можна
робити необхідні передбачення. Необхідні поживні речовини - стехіометрії Всім
морським водоростям необхідні водень, кисень, азот, фосфор, магній, залізо, мідь,
марганець, цинк, молібден, сірка, калій і кальцій. У 1950-х Редфілд визначив
ставлення основних поживних речовин в морських водоростях: C: N: P як
відповідно 42:7:1 у ваговому і 106:16:1 в атомарному еквівалентах.
Майже всі фотосинтетичні морські водорості
використовують неорганічні форми азоту, будь то нітрат, нітрит або амоній. Азот
у водних системах доступний в органічній, неорганічній або газоподібної
(елементарної) формі і може становити до 10% сухої ваги водоростей. Фосфор
відіграє важливу роль в фотосинтезі. Оптимальним є значення температури, що
знаходиться головним чином в діапазоні 25 ° C-40 ° C, але пік у більшості
організмів припадає на проміжку 20 ° C-25 ° C. Розподіл фітопланктону - рух,
скупчення, колонії, неоднорідність і турбулентність Водна навколишнє середовище
може бути надзвичайно гетерогенною. В горизонтальній площині рух води в значній
мірі впливає на неоднорідність розподілу і поживних речовин, і водоростевою
концентрації. Екологічні фактори: світло і температура також неравно розподілені
у водному просторі. Очевидно, світло зменшується з глибиною, але на це може
впливати концентрація різних суспензій і водоростей у воді. Температура,
звичайно, залежить від сезону. Але на більш короткому часовому проміжку на цей
фактор впливає охолодження від випаровування, верхні джерела (Водоспад), нижні
джерела (на дні водойми), горизонтальні течії та ін Масштаб змін важливий.
Через різких змін навколишніх умов, домінуючими можуть виявитися ті різновиди,
які швидше пристосовуються до екологічних факторів. Просторові зміни поживних
речовин, світла і температури можуть привести до відповідних просторовим змінам
в домінуючих різновидах. Нерівномірний розподіл зовнішніх факторів змушує
багато різновиди використовувати різні плавальні механізми, щоб зайняти
найбільш вигідні позиції для фотосинтезу. Це, як вважають, є одним з головних
чинників в домінуванні синьо - зелених водоростей. У багатьох різновидів є
джгутики, наприклад, у Chromophycota, Euglenophycota і Chlorophycota.
Dinophyta, Prymnesiophyta і Raphidophycae також є жгутиконосцами, менші з яких
плавають зі швидкістю 20 мм / год, у той час, як більш великі особини можуть
розвивати швидкість до 5 см / год. Діатомові водорості тримаються ближче до
поверхні, завдяки потокам води, в спокійній воді вони досить швидко
опускаються.[27]
4.2 Квоти
на викид парникових газів
Таблиця 4.1
Певні кількісні забов`язання в
відсотках відносно базового періоду сторон Кіотського протоколу
|
Сторона
|
Певні кількісні забов`язання в відсотках
відносно базового періоду
|
|
Австралія
|
108
|
|
Австрія
|
29
|
|
Бельгія
|
29
|
|
Болгарія*
|
29
|
|
Венгрія*
|
49
|
|
Германія
|
29
|
|
Греція
|
29
|
|
Данія
|
29
|
|
EC
|
29
|
|
Ірландія
|
29
|
|
Ісландія
|
110
|
|
Іспанія
|
29
|
|
Італія
|
29
|
|
Канада
|
49
|
|
Латвія*
|
29
|
|
Литва*
|
29
|
|
Ліхтенштейн
|
29
|
|
Люксембург
|
29
|
|
Монако
|
29
|
|
Нідерланди
|
29
|
|
Нова Зеландія
|
100
|
|
Норвегія
|
101
|
|
Польша*
|
49
|
|
Португалія
|
29
|
|
Россія*
|
100
|
|
Руминія*
|
29
|
|
Словакія*
|
29
|
|
Словенія*
|
29
|
|
Великобританія
|
29
|
|
США
|
39
|
|
Украіна*
|
100
|
|
Фінляндія
|
29
|
|
Франція
|
29
|
|
Хорватія*
|
59
|
|
Чешска Республіка*
|
29
|
|
Швейцарія
|
29
|
|
Швеція
|
29
|
|
* Сторони, що переходять до ринкової
економіки.
|
|
Для України базовим періодом
являється 1990 рік. Тобто квота на викид парникових газів в еквіваленті СО2 не
повинна перевищувати 100% від визначених викидів в 1990 році. Квоти Кіотського
протоколу можуть продаватися іншим країнам світу за визначеною на біржі ціною
за одну тонну СО2. Враховуючи занепад виробництва спричинений розпадом СРСР у
1991 році для України це насамперед означає наявність значного надлишку квот на
викиди парникових газів, що можуть бути продані іншим сторонам Кіотського
протоколу.
Показово, що на європейському ринку
вартість однієї тонни викидів знаходиться в межах € 20-21, але пропозиція для
РФ значно нижче світового ринкового - на рівні € 5-15. Високі інвестиційні
ризики вимагають «страховки» у вигляді заниженої ціни. Разом з тим у компанії
розраховують, що після реєстрації першого проекту, коли інвестори переконаються
в надійності своїх партнерів, ціни значно зростуть.
Потрібно відзначити, що інтерес
країн Євросоюзу до купівлі одиниць скорочень викидів (ОСВ) підстьобують
вводяться з 2008 року штрафи за перевищення ліміту викидів парникових газів,
які складуть близько € 100 за 1 т. Щоб уникнути в майбутньому гігантських
виплат, європейські країни активізувалися як на внутрішньому , так і на
зовнішньому ринку ОСВ. Основні принципи та критерії співпраці між вуглецевими
«донорами» і «реципієнтами» сформульовані у Кіотському протоколі - базовому
документі, підписаному в 1997 році в Кіото.
Станом на 2012 рік Україна має
надлишок викидів СО2 приблизно на 2,3 млрд тонн. Тобто теоретично ми можемо
непогано заробити на цьому обсязі, і навіть після падіння цін на ринку квот до
3 євро за 1 тонну виходить дуже солідна сума в 6,9 млрд дол..[40]
РОЗДІЛ V. ОХОРОНА ПРАЦІ
5.1
Небезпечні та шкідливі виробничі фактори при роботі в біохімічній лабораторії
Розглянемо небезпечні та шкідливі
виробничі фактори, які впливають на людину відповідно до класифікації,
наведеної у ГОСТ 12.0.003-74. Робоче місце знаходиться у біохімічній
лабораторії. Відповідно до цього на працівника діють такі небезпечні та
шкідливі виробничі фактори:
Фізичні:
підвищене значення напруги в
електричному ланцюзі, замикання якого може відбутися через тіло людини;
підвищений рівень електромагнітних
випромінювань;
лабораторний посуд, що може у
процесі роботи руйнуватися (наприклад скляний посуд).
недостатнє освітлення робочого місця
Хімічні:
хімічні речовини, що проникають в
організм людини через органи дихання, кишково-шлунковий тракт і слизові
оболонки.
Психофізіологічні:
нервово-психічні перевантаження
(перенапруга аналізаторів. монотонність праці, зоровий дискомфорт).
Підвищене значення напруги в
електричному ланцюзі, замикання якого може відбутися крізь тіло людини.
Основним джерелом небезпеки являються лабораторні пристрої та аналізатори
підвищеної потужності, наприклад лабораторна центрифуга, деякі аналізатори, та
інші електричні прилади. Підвищений рівень електромагнітних випромінювань.
Джерелом є комп'ютер. Використовується для обробки та аналізу наукових даних.
Хоча зараз сучасні комп'ютери випускають із захисними екранами або спеціально
нанесеним на дисплей захисним шаром, це не вирішує проблеми впливу
електромагнітного випромінювання на користувача комп'ютером. При роботі,
комп'ютер утворює навколо себе електромагнітне поле, яке деіонізірует навколишнє
середовище, а при нагріванні плати і корпус монітора випускають у повітря
шкідливі речовини. Все це робить повітря дуже сухим, слабо іонізованим, зі
специфічним запахом і в загальному "важким" для дихання. Природно, що
таке повітря не може бути корисний для організму і може призвести до
захворювань алергічного характеру, хвороб органів дихання та інших розладів.
Монітор є сильним джерелом електромагнітного випромінювання, особливо його
бічні і задні стінки, тому вони не мають спеціального захисного покриття, яке є
у лицьовій частині екрана. Електромагнітні випромінювання найбільший вплив
роблять на імунну, нервову, ендокринну та статеву систему.
Лабораторний посуд, що може у
процесі роботи руйнуватися ( наприклад скляний посуд). Скло є одним з найбільш
часто використовуваних видів обладнання в лабораторіях. Дотримуйтесь правил
техніки безпеки лабораторії при використанні і обробці посуду, щоб уникнути
нещасних випадків і травм.
Недостатнє освітлення робочого місця
викликає швидку втому і хвороби очей, знижує уважність і, отже, значно зменшує
продуктивність праці, а також збільшує ймовірність нещасних випадків на
виробництві. Недостатнє освітлення робочого місця ускладнює тривалу роботу,
викликає підвищене стомлення і сприяє розвитку короткозорості. Занадто низькі
рівні освітленості викликають апатію і сонливість, а в деяких випадках сприяють
розвитку почуття тривоги. Тривале перебування в умовах недостатнього освітлення
супроводжуються зниженням інтенсивності обміну речовин в організмі і
ослабленням його реактивності. До таких же наслідків приводить тривале
перебування в світловий середовища з обмеженою спектральним складом світла і
монотонним режимом освітлення.
Хімічні речовини, що проникають в
організм людини через органи дихання, кишково-шлунковий тракт і слизові
оболонки. Джерело - випари хімічних речовин, що знаходяться в лабораторії.
Також під час проведення дослідів в лабораторії з хімічними речовинами,
внаслідок недотримання правил охорони праці та невиконання прийнятих методик
можуть при контакті з організмом людини викликати травми. Тривалість - протягом
всього робочого часу, 40 год/тиждень.
Нервово-психічні перевантаження
(перенапруга аналізаторів. монотонність праці, зоровий дискомфорт). Джерело -
робота на комп'ютері. Нервове напруження, стомлюваність. Тривала робота за
комп'ютером може викликати підвищене стомлення, головний біль, роздратованість,
розлади сну, стрес. Тривалість дії фактора - близько 20 год/ тиждень (половина
робочого часу), що вкладається в норму (не більше 6 год/день).
5.2
Технічні та організаційні заходи для зменшення рівня впливу небезпечних та
шкідливих виробничих факторів при роботі в біохімічній лабораторії
Аби уникнути ураження електричним
струмом при користуванні мікроскопами, центрифугами, приладами-аналізаторами:
Користуватися електроспоживачами,
шнури яких мають триполюсну вилку з попереджуючим включенням заземлюючого
(зануляючого) проводу.
Не вмикати в електромережу
електроспоживачі, шнури живлення яких мають пошкоджену ізоляцію.
Не вмикати в електромережу
електроспоживачі, які мають пошкодження або ненадійно з’єднані з електрошнуром
живлення, вилками, розетками та подовжувачами.
Не вмикати електроспоживачі в
розетки, які не мають захисних направляючих. Не користуватися пошкодженими
розетками, відгалужувальними та з’єднувальними коробками, вимикачами та іншою
електроарматурою, а також електролампами, скло яких має сліди затемнення або
випинання.
Не користуватися саморобними
подовжувачами, які не відповідають вимогам ПУЕ, що пред’являються до переносних
електропроводок.
Не застосовувати для опалення
приміщень нестандартного (саморобного) електронагрівального обладнання або ламп
розжарювання.
При користуванні електроспоживачами,
які мають окремий самостійний провід заземлення, перед включенням його в
електромережу перевірити наявність та надійність приєднаного заземляючого
проводу до відповідних клем.
По можливості, уникати доторкання
руками до металевих частин електроспоживачів, ввімкнених в електромережу.
Не торкатися руками до обірваних та
оголених проводів електромережі.
Самостійно не замінювати зіпсовані
електрозапобіжники, електролампи, не проводити ремонт електроспоживачів та
електромережі.
При прибиранні пилу з
електроспоживачів, митті холодильників та підлоги біля них обов’язково
відключати їх від електромережі.
Не залишати без догляду працюючі
електроспоживачі.
По закінченні робочого дня вимкнути
вимикач на електроспоживачі та від’єднати провід живлення від розетки
електромережі. При цьому слід пам’ятати, що від’єднуючи вилку електроспоживача
від розетки, її слід тримати за корпус, а не смикати за провід живлення, бо
можна висмикнути один провід і потрапити під дію електричного струму.
. Підвищений рівень електромагнітних
випромінювань. Щоб уникнути несприятливого впливу електромагнітного
випромінювання від комп’ютера на користувача необхідно:
по можливості, варто придбати
рідкокристалічний монітор, оскільки його випромінювання значно менша, ніж у
поширених ЕЛТ моніторів (монітор з електронною трубкою).
при покупці монітора необхідно
звернути увагу на наявність сертифікату.
системний блок і монітор повинен
знаходитися якнайдалі від вас.
не залишайте комп'ютер включеним на
тривалий час якщо ви його не використовуєте, хоча це і прискорить знос
комп'ютера, але здоров'я корисніше. Так само, не забудьте використовувати
"сплячий режим" для монітора.
у зв'язку з тим що електромагнітне
випромінювання від стінок монітора набагато більше, постарайтеся поставити
монітор в кут, так що б випромінювання поглиналося стінами. Особливу увагу
варто звернути на розстановку моніторів в офісах.
по можливості скоротіть час роботи
за комп'ютером і частіше переривайте роботу.
комп'ютер повинен бути заземлений.
Лабораторний посуд, що може у
процесі роботи руйнуватися (наприклад скляний посуд). Щоб уникнути руйнування
скляного посуду, його спочатку перевіряють за допомогою полярископа.
Під час зборки скляної апаратури
гумові пробки та трубки підбирають по розміру скла, а руки захищають рушником
чи ганчірками для уникнення порізів при руйнуванні пристроїв. Забороняється
закривати нагріті скляні посудини притертими пробками до їх охолодження.
Мити хімічний посуд передбачено у
приміщеннях, які мають раковини, мийки та обладнання для її зберігання та
сушки. Не дозволяється у раковину викидати чи зливати концентровані розчини
кислот та лугів, хромову суміш, речовини з неприємним запахом та інші реактиви.
Вони зливаються у спеціальні ями для виключення небезпеки опіку.
Під час розбору апаратури потрібно
дотримуватися обережності при торканні до гарячого скляного посуду та
нагрівачам. Гарячі колби ставлять на листовий азбест.
Недостатнє освітлення робочої зони
При роботі в домашніх умовах.
Визначення площі приміщення:= А•В,
(5.1)
визначення індексу приміщення:
φ = S / ((h1-h2) • (A + B))
(5.2)
визначення потрібної кількості
світильників:
= (E • S • 100 • Kз) / (U • п • Фл),
(5.3)
Де E - необхідна освітленість
горизонтальної площини, лк, S - площа приміщення, м.кв.; Кз - коефіцієнт
запасу; U - коефіцієнт використання освітлювальної установки; Фл - світловий
потік однієї лампи, лм; N - число ламп в одному світильнику.
Таблиця 5.1
Дані щодо освітленості робочого
приміщення
|
а
|
5м
|
Довжина приміщення
|
|
b
|
3м
|
Ширина приміщення
|
|
h1
|
2,5м
|
Висота приміщення
|
|
h2
|
0,8м
|
Висота стола
|
|
Кв
|
30
|
|
E
|
150
|
Потрібна освітленість (норма для рабочого кабінету)
|
|
Kз
|
1,15
|
Коефіціент запасу лампи накалювання
|
|
Фл
|
900 лм
|
Світловий потік люмінесцентної лампи
|
= 5*3=15м2
φ = 15м2/
(2,5-0,8)*(5м+3м)=1,102= 30+1,102=31=(150*15м2*100*1,15)/(31*1*900)=9.27
Кількість потрібних ламп в даному
приміщенні 9 штук. Оскільки в приміщенні встановлено 7 ламп - освітлення можна
вважати недостатнім
Хімічні речовини, що проникають в
організм людини через органи дихання, кишково-шлунковий тракт і слизові
оболонки. Під час роботи в лабораторії для консервування прб використавувався
технічний розчин (40%) формаліну.
Формалін має високу бактерицидну
активність, завдяки якій запобігає розкладанню. Використовується для тривалого
зберігання і бальзамування органічних речовин і об'єктів (організмів або
окремих органів). Використовується для створення анатомічних моделей.
Застосовується при виробництві кінофотоплівки, синтетичних смол і каучуку, а
також багатоатомних спиртів і поверхнево-активних речовин. Для технічного
формаліну передбачено зберігання в обігріваються ємностях, при температурі від
10 до 25 градусів.
Розчин формаліну при кімнатних
температурах виділяє формальдегід, що володіє отруйними властивостями. Гранична
концентрація формальдегіду в повітрі не повинна перевищувати один міліграм на
метр квадратний. Щоб уникнути отруєнь в повітрі вибухонебезпечних сумішей
працювати з формаліном можна тільки у витяжній шафі, обов'язково потрібно
захищати шкіру рук рукавичками.
5.3
Забезпечення пожежної і вибухової безпеки при роботі в біохімічній лабораторії
Причини пожеж і
загорянь в біохімічній лабораторії можуть бути такі:
- несправний
пристрій, чи порушення режиму роботи систем опалення, вентиляції і
кондиціонування повітря;
несправний
пристрій, чи перевантаження електричних установок і мереж (неправильний вибір
перетинів чи проводів підбор електроустаткування, несправність засобів захисту
мереж від перевантажень і ін.);
самозапалювання і
самозаймання речовин і матеріалів при неправильному їхньому збереженні чи
застосуванні;
необережне
поводження з вогнем (паління в невстановлених місцях, недбале проведення
вогневих робіт, залишення без догляду електронагрівальних приладів і т.п.).
До небезпечних
факторів пожежі відносяться:
відкритий вогонь чи
іскри;
підвищена
температура повітря, предметів і т.п.;
токсичні продукти
горіння;
дим (високодисперсна
аерозоль із твердими частками);
знижена
концентрація кисню;
- вибух.
Загальні вимоги до
систем запобігання пожеж і пожежного захисту регламентується міждержавними
стандартами системи стандартів безпеки праці ГОСТ 12.1.004-91 і ГОСТ
12.1.010-76 і спеціальною нормативно-технічною документацією.
Залежно вiд обставин гасiння пожежі
можна досягти слідучими методами:- усунути доступ у зону горіння окислювача;
охолодити зону горіння нижче
темпеpатуpи самозагорання;- розмішати гоpючі речовин негоpючими;
інтенсивно гальмувати швидкость
хiмiчних pеакцій у полум’ї;
механiчно відірвати полум’я стpуєю
газів та води.Hа цих пpинципах і заснованi засоби гасiння пожеж. Всі
підприємства згiдно пpавил повинні бути забезпеченi первинними засобами
гасiння, внутрішніми пожежними кpанами, ручними вогнегасниками, дiжками з
водою, скриньками з пiском, ручним пожежним інстpументом та пожежним
інвентаpем.Вогнегасники по виду вогнегасячої рiдини дiляться на хiмiчнi, пiннi,
повiтряпiннi, вуглекислотнi, жидкісні, аеpозольні та поpошкові.
Використовуються стаціонарні автоматичні установки пожежогасіння, які дiляться
на аеpозольні, рідинні, воднi (спpинклеpні та дpенчаpні) та парові. -
Спринклерні голiвки виконують склянi
або металевi з легкоплавкого замка. Пpи температурі 72 - 1820С замок
pазплавляється і голiвка, яка постiйно заповнена водою, відкривається.
Дpенчаpна система - подiбна
спpинклеpній, але дpепчаpні голiвки постійно відкриті, вода подається пpи
спрацюванні датчика.
ВИСНОВКИ
Основу видового багатства
фітопланктону водойм НПП «Голосіївський» складають Chlorophyta (121 вид - 126
в.в.т.) або 44,4% загального числа виявлених таксонів видового і внyтрівидового
рангу), Bacillariophyta (52 види або 19,1%), Cyanoprocaryota (29 видів - 31
в.в.т. або 10,7%) та Еuglеnophyta (27 видів - 30 в.в.т. або 9,9%). Інші відділи
представлені 5-13 видами.
Відділи Chlorophyta та
Basilariophyta викликають процес цвітіння на протязі усьго досліджуваного
періоду часу.
Співвідношення кількості клітин
(млн.\дм3) до біомаси фітопланктону (мг\л) та концентрації клітин водоростей у
заданому об`эмі води. Відслідковування динаміки цих параметрів є схожими за
своею динамікою кількість клітин водоростей а також їх біомаса відповідає
рівням гіперцвітіння та інтенсивного цвітіння з наведеної вище класифікації.
Останні відмітки на початку та в кінці вересня свідчать про зниження рівня
евтрифікації в каскаді ставків
Процеси фотосинтезу в світовому
океані відіграють важливу економічну роль зважаючи на положення Кіотського
протоколу.
Станом на 2012 рік Україна має
надлишок викидів СО2 приблизно на 2,3 млрд тонн. Тобто теоретично ми можемо
непогано заробити на цьому обсязі, і навіть після падіння цін на ринку квот до
3 євро за 1 тонну виходить дуже солідна сума в 6,9 млрд дол
Задля контролю над рівнем
фітопланктону, за будь якою моделлю потрібно контролювати лімітуючий фактор
розмноження.
Рівні біомаси фітопланктону можуть
бути використані як індикатор антропогенного впливу на середовище.
Високий рівень біомаси фітопланктону
в період цвітіння гарний показник для будь якої водної екосистеми, проте
процеси евтрифікації слід вважати загрозливими.
СПИСОК ВИКОРИСТАНОЇ ЛІТЕРАТУРИ
1.Апонасенко
А. Д., Франк Н. А., Сидько Ф. Я. Дифференциальный спектрофотометр для
гидрооптических исследований // Океанология. - 1976. - 1, в. 5. - С. 924-927.
.Василевич
В. И. Статистические методы в геоботанике. - Л.: Наука, 1969. - 144 с.
.Вислоух
С. М. Биологический анализ вод // Руководство по теоретической и практической
медицине. - 1916. - 7-8. - С. 225-304.
.Гольд
В. М., Попельницкий В. А. Определение фотосинтеза фитопланктона флуоресцентным
методом // Методические вопросы изучения первичной продукции планктона
внутренних водоемов. - СПб.: Гидрометеоиздат, 1993. - С. 25-29.
Дацко
В. Г. О некоторых изменениях в процессах продуцирования органического вещества
в Азовском море после зарегулирования стока Дона // Гидрохим. материалы. -
1957. - 26. - С. 49-64.
.Карабашев
Г. С., Соловьев А. Н. О возможности индикации изменчивости качественного
состава фитопланктона в море по характеристике флуктуаций интенсивности
флуоресценции хлорофилла // Докл. АН СССР. - 1975. - 220, 4. - С. 958-960.
.Киселев
И. А. Планктон морей и континентальных водоемов. - Л.: Наука, 1969. - 657 с.
Коновалов
Б. В., Мордасова Н. В. Методы и приборы современных океанологических
иследований. - М.: ЦНИИ информ. и техн.-экон. исслед. рыб. хоз-ва, 1970. - 40
с.
.Копелевич
О. В., Маштаков Ю. Л., Русанов С. Ю. Аппаратура и методика исследования
оптических свойств морской воды // Гидрофизические и гидрооптические
исследования в Атлантическом и Тихом океанах. - М.: Наука, 1974. - С. 97-107.
.Минеева
Н. М., Попельницкий В. А. Определение фотосинтеза фитопланктона Рыбинского
водохранилища флуоресцентным и кислородным методами // Биология внутренних вод:
Информ. бюл. - Л., 1990. - № 88. - С. 20-24.
.Одум
Ю. Экология. - М.: Мир, 1986. - Т. 1. - 328 с.
.Оксиюк
О. П., Жданова Г. А., Гусынская С. И. и др. Оценка состояния водных объектов
Украины по гидробиологическим показателям. 1. Планктон // Гидробиол. журн. -
1994. - 30, № 3. - С. 26-31.
Определение
содержания хлорофилла в планктоне пресных водоемов / Сост. Л. А. Сиренко, А. В.
11.Курейшевич - Киев: Наук. думка, 1982. - 52 с.
.Пырина
И. Л. Свет как фактор продуктивности фитопланктона во внутренних водоемах:
Автореф. дис. ... докт. биол. наук: - СПб., 1995. - 47 с.
.Романенко
В. И. Микробиологические процессы и их роль в биопродуктивности экосистем и
самоочищении водохранилищ // Водохранилища и их воздействие на окружающую
среду. - М.: Наука, 1986. - С. 138-149.
.Руководство
по методам гидробиологического анализа поверхностных вод и донных отложений. -
Л.: Гидрометеоиздат, 1983. - С. 87-91.
Топачевский
А. В., Масюк Н. П. Пресноводные водоросли Украинской ССР. - Киев: Вища шк.,
1984. - 333 с.
.Щербак
В.И. Изучение суточной динамики образования первичной продукции фитопланктона
// Гидробиологичесикие исследования водоемов юго-западной части СССР. - Киев:
Наук. думка, 1982. - С. 131-132.
.Щербак
В.И. Метод авторадиографии в гидробиологических исследованиях // Вопросы
гидробиологии водоёмов Украины. - Киев: Наук. думка, 1988. - С. 106-110.
.
Щербак В. И. Скляночные методы определения первичной продукции водорослей
планктона и бентоса // Тр. IV Поволж. конф. - - Казань: Изд-во Казан. ун-та,
1991. - Т 1. - С. 59-66.
.Щербак
В. И. Определение первичной продукции отдельных видово водорослей
авторадиографическим методом // Методические вопросы изучения первичной
продукции планктона внутренних водоемов. - СПб., 1993. - С. 47-51.
.Щербак
В. И. Продукционные характеристики доминирующих видов фитопланктона днепровских
водохранилищ // Альгология. - 1998. - 8, № 3. - С. 286-294.
,Щербак
В. И. Фотосинтетическая активность доминирующих видов днепровского
фитопланктона // Гидробиол. журн. - 1998. - 34, № 5. - С. 11-22
.Щербак
В. И. Влияние продолжительности экспозиции на показатели первичной продукции
фитопланктона евтрофных водоемов при использовании скляночного метода в
кислородной модификации // Там же. - 2000. - Т 36, № 1. - С. 97-102.
.Щербак
В. І. Методи досліджень фітопланктону // Методичні основи гідробіологічних
досліджень водних екосистем. - К., 2002. - С. 41-47.
.Starmach
K. Metody botania plankton. - Warszawa, 1956. - 135 s.I.E. Un sity detection
and estimation of chlorophull and atheir pigments in the ocean // Proc. Nat.
Acad. Sci. USA. - 1961. - 47, N 11. - p. 1082-1090.2000/60/EC of the European
Parliament and of the Council - Offic. J.U 327/1.
.Василевич
В. И. Статистические методы в геоботанике. - Л.: Наука, 1969. - 144 с.
.Владимирова
К. С. Методика изучения первичной продукции донных водорослей // Гидробиол.
журн. - 1969. - Т. 5, №4. - С. 128-130.
.Владимирова
К. С. Фитомикробентос Днепра, его водохранилищ и Днепровско-Бугского лимана. -
К.: Наук. думка, 1978. - 232 с.
.Гурвіч
В. В., Цееб Я. Я. Мікробентометр для взяття кількісних проб мікробентосу // ДАН
УРСР. - 1958. - 10. - С. 21-24.
.Топачевский
А. В., Масюк Н. П. Пресноводные водоросли Украинской ССР. - К.: Вища школа,
1984. - 333 с.
.Травянко
В. С., Евдокимова Л. В. Микробентометр БМ-ТЕ // Гидробиол. журн. - 1968. - Т.
4, №1. - С. 94-95.
.Оксиюк
О. П., Зимбалевская Л. И., Протасов А. В. Оценка состояния водных объектов
Украины по гидробиологическим показателям // Гидробиол. журн. - 1994. - Т. 30,
№4. - С. 31-35.
.Щербак
В. И. Скляночные методы определения первичной продукции водорослей планктона и
бентоса // Труды IV Поволжской конференции. - Т. 1. - Казань: Изд-во Казанского
университета, 1991. - С. 59-66.
.Щербак
В. І. Гідроекологічні аспекти вирішення проблеми оцінки та зниження загроз
біорізноманіттю континентальних водойм України // Оцінка і напрямки зменшення
загроз біорізноманіття України. - К.: Імджест. - 2003. - С. 237-348.
34.Клоченко
П.Д., Царенко П.М., Иванова И.Ю. Особенности видового состава фитопланктона
водоёмов национального природного парка «Голосеевский»// Гидробиол. журн. - 2010.
- Т. 46, №1. - С.36-46
.Клоченко
П.Д., Горбунова З.Н., Харченко Г.В. и др. Особливості екологічного стану
Горіховатських ставків // Наук. вісн. нац. аграр. ун-ту. - 2006. - Bип. 95 , ч.
1. - С. 54-65.
.Клоченко
П.Д, Царенко П.М. Фітопланктон як показник екологічного стану Китаївських
ставків // Наук. вісн. нац. аграр. ун-ту. - 2007. - Bип. 107. - С. 66-72.
.
Щербак В.И., Семенюк Н.Е. Сравнительная характеристика фитопланктона водоёмов
различных районов г. Киева// Гидробиологический журнал.-2005. - Т.41, №2. -
С.29-34.
.
Лушпа В.І. Водяний латук (Pistia stratiotes L.) у Голосіївському ставку м.
Києва// Науковий вісник Національного університету біоресурсів і
природокористування України, 2009. Вип. 134. Ч. 1.-С.147-
.Волковский
П.А., Розова А.А. Практикум по сельскохозяйственным ме- лиорациям, - М.: Колос,
1980, - 239 с.
.Ерхов
Н.С., Мисенев В.С. Практикум по сельскохозяйственной мелиора-ции и
водоснабжению. -М: Колос, 1977. -144 с.