Бактериофаги и их свойства
Министерство
науки и образования Украины
Днепропетровский
национальный университет имени Олеся Гончара
Факультет
заочного и дистанционного образования
Кафедра
микробиологии и вирусологии
Контрольная
работа
По
дисциплине: «Генетика микроорганизмов»
НА ТЕМУ:
«Вирулентные и лизогенные фаги. Способность к интеграции с хромосомой хозяина.
Состояние лизогении»
Выполнила:
ст. гр. ББ-08-2з
Проценко Д.В.
Проверил:
доц. Крысенко О.В.
Днепропетровск
2011
Содержание
Введение
.Вирулентные и лизогенные фаги
.Способность к интеграции с
хромосомой хозяина
.Состояние лизогении
Заключение
Список литературы
Бактериофа́ги
- вирусы, избирательно поражающие бактериальные клетки. Описанные выше
бактериофаги, как правило, лизируют зараженные ими бактерии, и потому их
называют вирулентными. Некоторые фаги, однако, заражают бактерий-хозяев, но не
размножаются в них автономно и не вызывают лизиса. Такие фаги называются
умеренными. Лизогенные бактерии обладают потенциальной способностью
продуцировать фаги, но эту способность нельзя обнаружить, ни морфологическим,
ни серологическим исследованием. Фаг в таком неинфекционном состоянии,
передающийся только дочерним клеткам при делении, называют профагом. В
лизогенных клетках профаг прочно связан с хромосомой клетки-хозяина.
Лизогения - генетически обусловленная
способность бактерий лизироваться с выделением бактериофага через ряд поколений
после непосредственного заражения им. Теория разработана в 1950 французскими
учёными А. Львовым и А. Гутман, показавшими, что лизогенное состояние связано с
присутствием в клетках бактерий потенциально инфекционной структуры - профага.
В каждом поколении лизогенных бактерий подвергается лизису очень небольшая
часть клеток с освобождением от 70 до 150 частиц так называемого умеренного
фага. То есть лизогения - сложная форма вирусной инфекции у бактерий, при
которой от момента заражения бактерий фагом до лизиса клетки проходит большое
число клеточных поколений.
1. Вирулентные и лизогенные фаги
Бактериофа́ги
(фаги) (от др.-греч. φᾰγω
- «пожираю»)
- вирусы, избирательно поражающие бактериальные клетки. Чаще всего бактериофаги
размножаются внутри бактерий и вызывают их лизис. Как правило, бактериофаг
состоит из белковой оболочки и генетического материала одноцепочечной или
двуцепочечной нуклеиновой кислоты (ДНК или, реже, РНК). Размер частиц
приблизительно от 20 до 200 нм [1].
Описанные выше бактериофаги, как правило,
лизируют зараженные ими бактерии, и потому их называют вирулентными. Некоторые
фаги, однако, заражают бактерий-хозяев, но не размножаются в них автономно и не
вызывают лизиса. Такие фаги называются умеренными. Видимо, их размножение
происходит синхронно с размножением бактерии. Лишь очень редко, в одной из
102-105 таких «лизогенных» бактерий, фаг начинает спонтанно размножаться и
клетка подвергается лизису. В этом случае для того, чтобы обнаружить выход
инфекционного фага, в качестве индикатора нужен другой бактериальный штамм, для
которого этот фаг вирулентен. Если смешать лизогенные бактерии с избытком
бактерий-индикаторов и посеять смесь на агаризованную среду, то будут расти
также и колонии лизогенных бактерий [3]. Время от времени некоторые клетки
будут лизироваться и выходящие из них фаговые частицы, будут заражать
находящиеся по соседству чувствительные (индикаторные) бактерии. Это приведет к
появлению бляшек в сплошном бактериальном газоне. Однако в середине каждой
такой бляшки сохранится колония лизогенной бактерии (рис. 4.12).
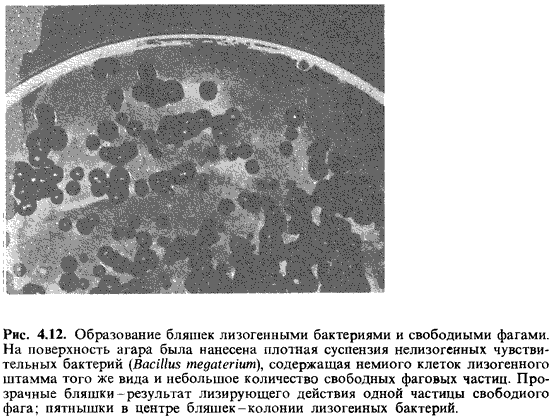
Лизогенные бактерии обладают потенциальной способностью
продуцировать фаги, но эту способность нельзя обнаружить, ни морфологическим,
ни серологическим исследованием. Фаг в таком неинфекционном состоянии,
передающийся только дочерним клеткам при делении, называют профагом. Подобно
другим признакам бактериальной клетки, наличие в ней профага наследуется.
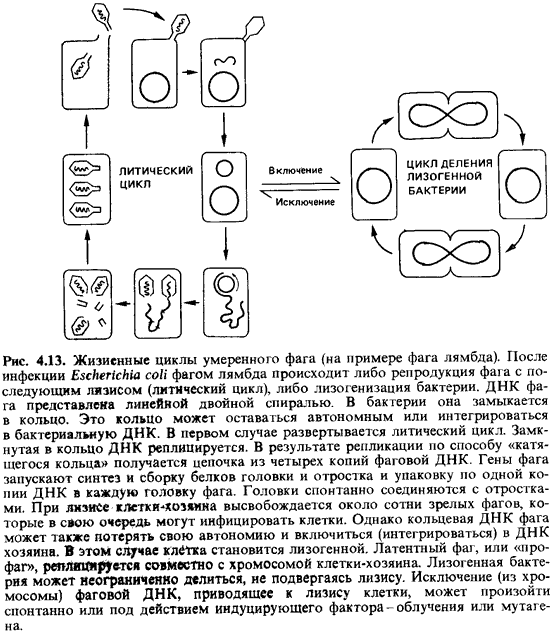
Поскольку все потомство лизогенной клетки тоже лизогенно, профаг, очевидно,
должен реплицироваться синхронно и регулярно вместе с хромосомой клетки-хозяина
(рис. 4.13) [4].
Лизогенные бактерии иммунны к заражению теми
фагами, которые присутствуют в них в виде профага. Обеспечиваемый профагами
иммунитет обусловлен не невозможностью адсорбции (как при устойчивости к
вирулентным фагам), а образованием особого цитоплазматического
белка-репрессора, препятствующего размножению вегетативных фагов. Этот же
репрессор препятствует обратному переходу профага в вегетативное состояние и
подавляет синтез фаговых белков. Возникновение лизогенного состояния связано,
таким образом, с образованием репрессорного белка[6].
Спонтанно, без воздействия извне лизогенные
бактерии лизируются редко. Однако целый ряд факторов (ультрафиолетовые лучи,
митомицин С или алкилирующие агенты) может индуцировать в каждой клетке
развитие профага, ведущее к образованию и высвобождению инфекционного фага.
Успех такой индукции зависит от генетической конституции профага,
физиологического состояния хозяина и условий культивирования. Индукция связана,
очевидно, с устранением или инактивацией имеющихся молекул репрессора.
Некоторые мутанты умеренных фагов образуют термолабильный репрессор, и тогда
достаточно уже повышения температуры до 44°С, чтобы вызвать лизис бактерий [5].
. Способность к интеграции с хромосомой хозяина
лизогенная
бактерия профаг хромосома конверсия
Интеграция и индукция фага λ
(лямбда).
Изучение фага лямбда (λ), лизогенного
для Escherichia coli K12, позволило выяснить, каким образом профаг связан с
бактериальной хромосомой. Лизогенизация бактерий этим фагом может служить
примером жизненного цикла умеренного бактериофага. Длина хромосомы фага лямбда
оставляет всего 2% длины бактериальной хромосомы [1].
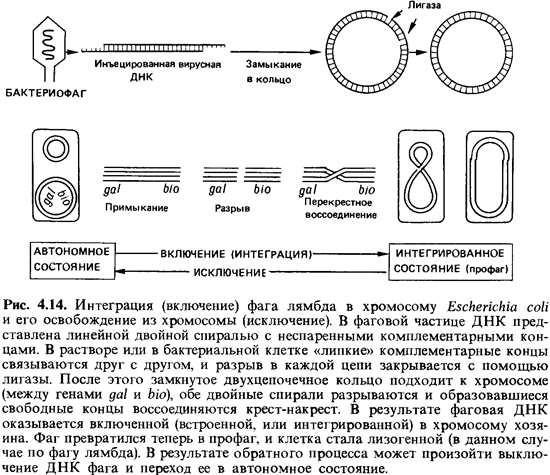
В свободных фаговых частицах ДНК присутствует в
виде линейной (не кольцевой) двойной спирали (рис. 4.14). Каждая из цепей на
одном конце выступает за пределы дуплекса на 12 нуклеотидов. Эти два
одноцепочечных конца комплементарны друг другу; путем спаривания оснований они
могут соединяться друг с другом, поэтому их называют «липкими» концами. Если
поместить такие молекулы ДНК in vitro в раствор, то благодаря взаимодействию
между комплементарными основаниями одноцепочечных концов наступает равновесие
между линейными и кольцевыми ДНК. Такое же замыкание в кольцо происходит после
того, как фаг лямбда инфицирует клетку. При этом оба разрыва между концами
цепей закрываются полинуклеотидлигазой. Функция этого бактериального фермента
состоит в том, чтобы устранять разрывы в отдельных цепях двойных спиралей ДНК
путем связывания нуклеотидов. Таким образом, для замыкания линейной ДНК фага в
кольцо никакие фаговые ферменты не нужны [7].
В лизогенных клетках профаг прочно связан с
хромосомой клетки-хозяина. При конъюгации клеток профаг вместе с хромосомой
хозяина переносится из клетки-донора в клетку-реципиент. Генетические
эксперименты показывают, что фаг лямбда присоединен к хромосоме хозяина в
совершенно определенном месте (между галактозным опероном и биотиновым
локусом). Вначале предполагали, что ДНК бактериофага только прикрепляется к
хромосоме бактерии в этом участке. Однако в результате составления генетических
карт фага, а также из опытов по рекомбинации стало ясно, что фаговая ДНК при
лизогенизации не просто прикрепляется к бактериальной ДНК, а включается в нее
[3].
Включение (интеграция) ДНК профага в хромосому
клетки-хозяина происходит, очевидно, в результате разрыва и перекрестного
воссоединения. За эту реакцию ответствен фермент, названный лямбда-интегразой.
Он узнает две разные, негомологичные последовательности нуклеотидов - одну в
хромосомной ДНК и одну в ДНК фага - и тесно сближает обе двойные спирали друг с
другом; затем последние разрываются и снова соединяются крест-накрест.
Отдельные этапы этой специфической рекомбинации показаны на рис. 4.14.
В интегрированном состоянии фаговая ДНК
реплицируется вместе с бактериальной и подвержена тем же регуляторным
воздействиям, что и удвоение бактериальных хромосом. Информация, содержащаяся в
фаговой ДНК, в это время не проявляется [8]. Только в результате перехода
профага в вегетативное состояние восстанавливается автономия фаговой ДНК и
начинается размножение фага. Этот обратный процесс может произойти спонтанно
или в результате индукции (например, под действием ультрафиолетового
облучения). Исключение фаговой ДНК из бактериальной хромосомы происходит,
вероятно, путем обращения процессов, приведших к ее включению, и осуществляется
очень точно: более 99% фаговых частиц, освобождающихся из лизогенных клеток,
идентичны с исходным (инфицирующим) фагом. Это означает, что фаговая ДНК при ее
выключении выщепляется точно в том же месте, где происходила интеграция. Только
в редких случаях (одном из 100 000) выключение ДНК фага происходит аномально
[9].
Как только профаг в результате выключения
перешел в вегетативное состояние, он опять становится автономным и может
размножаться в бактериальной клетке как вирулентный фаг. Выключение, таким
образом, приводит к лизису бактерии и высвобождению фага лямбда.
Двухцепочечная ДНК вируса может встроиться в
хромосому клетки-хозяина с помощью фермента интегразы. Такой процесс
встраивания в хромосомную ДНК называют интеграцией. Вирусный геном в форме
интегрированной ДНК, синтезированной по проникшей в клетку вирусной РНК с
помощью обратной транскриптазы, называется провирусом . Провирус становится
частью генетического материала клетки, реплицируется вместе с клеточной ДНК и
при делении передается дочерним клеткам. В скрытой (латентной) форме провирус
может пребывать бесконечно долгое время, переходя от родителей к потомкам через
сперматозоид или яйцеклетку. Способность вирусных геномов к интеграции с
геномом клетки была предсказана замечательным исследователем-вирусологом Львом
Александровичем Зильбером (1894-1966) [6].
Лизогения (от греч. lýsis
- разложение,
распад и ...geneia - происхождение, создание) генетически обусловленная
способность бактерий лизироваться с выделением бактериофага через ряд поколений
после непосредственного заражения им. Теория разработана в 1950 французскими
учёными А. Львовым и А. Гутман, показавшими, что лизогенное состояние связано с
присутствием в клетках бактерий потенциально инфекционной структуры - профага.
Изменение свойств бактериальной клетки, связанное с присутствие профага,
получило название феномена лизогенной конверсии или фаговой конверсии. Впервые
феномен фаговой конверсии описал Фримен в 1951 г. при наблюдении токсигенности
у дифтерийной палочки. Он показал, что продукция экзотоксина у дифтерийных
бактерий связана с обязательным присутствием профага в клетке. В настоящее
время для многих микроорганизмов доказано, что способность выделять экзотоксин
детерминирована фагами, находящимися в клетке. Присутствие профага в бактерии,
с одной стороны, губительно для нее, а с другой - делает ее иммунной к
заражению гомологичным или близкородственным фагом [10]. В каждом поколении
лизогенных бактерий подвергается лизису очень небольшая часть клеток (Лизогения
1 клетка на миллион) с освобождением от 70 до 150 частиц так называемого
умеренного фага. Частота перехода профага в инфекционное состояние (индукция
профага) может быть увеличена рядом агентов (например, ультрафиолетовыми
лучами). После заражения бактериальной клетки умеренным фагом процесс инфекции
может развиваться по одному из двух направлений (см. рис.1): по пути
литического цикла, который так же, как и при заражении бактерий вирулентными
фагами, заканчивается лизисом клеток и выходом потомства фага в окружающую
среду; по пути лизогенизации, когда в результате биосинтетических процессов в
клетке вырабатывается иммунитет к инфицирующему фагу, фаговая ДНК включается в
ДНК бактерии и в дальнейшем реплицируется вместе с ней как её составная часть
(профаг), а бактерия выживает и становится лизогенной. Судьба клетки решается
на первых этапах инфекции и зависит главным образом от времени формирования
иммунитета. Если состояние иммунитета наступает раньше, чем развитие инфекции
достигнет стадии, необратимо ведущей к лизису, то может осуществиться
лизогенизация. В геноме бактерий могут содержаться одновременно профаги
нескольких разных фагов. В этом случае клетка обладает иммунитетом в отношении
всех этих фагов. В результате лизогенизации может произойти изменение некоторых
свойств бактериальной клетки (Лизогенная конверсия), обусловленное
приобретением бактерией новой генетической информации [5].
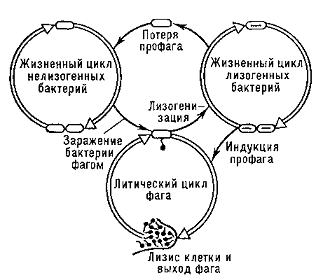
Рис .1. Лизогения
Искусственно полученные лизогенные бактерии по
своим свойствам не отличаются от лизогенных бактерий, найденных в естественных
условиях. У небольшой части потомства лизогенной клетки происходит «исцеление»
- потеря профага. Утратившие профаг клетки дают начало нелизогенным линиям.
Частота этого процесса может быть увеличена, например, действием
ультрафиолетовых лучей. То есть лизогения - сложная форма вирусной инфекции у
бактерий, при которой от момента заражения бактерий фагом до лизиса клетки проходит
большое число клеточных поколений.
Заключение
Итак, фаги могут быть вирулентными и умеренными.
Вирулентные фаги проникают в микробную клетку, размножаются в ней и вызывают ее
лизис. Умеренные фаги вступают в своеобразные симбиотические взаимоотношения с
микробной клеткой: проникнув в клетку, они включают свой геномом в хромосому
бактерий и реплицируются вместе с ней. Бактерии, несущие умеренный фаг,
получили название лизогенных; фаг, присутствующий в них, называют профагом, а
симбиоз бактериальной клетки с профагом - феноменом лизогении. Лизогения широко
распространена и практически выявлена почти у всех видов бактерий. В связи с
этим лизогению следует считать не исключительным, а нормальным состоянием
микроорганизмов. Фаг (в состоянии профага) может в течение многих лет находится
в бактериях, не теряя способности при определенных условиях превратиться в
полноценную фаговую частицу. В эволюционном аспекте процесс лизогенизации можно
расценивать как выгодный и фагу, и бактериальной клетке. Лизогенная культура
приобретает ряд новых полезных для нее свойств: невосприимчивость к повторному
заражению гомологичным вирулентным фагом, способность передавать фаг по
наследству и продуцировать ряд веществ, синтез которых детерминируется
профагом.
Изменение свойств бактериальной клетки,
связанное с присутствие профага, получило название феномена лизогенной
конверсии или фаговой конверсии. Впервые феномен фаговой конверсии описал
Фримен в 1951 г. Из того, что мы узнали, очевидно, что присутствие профага в
бактерии, с одной стороны, губительно для нее, а с другой - делает ее иммунной
к заражению гомологичным или близкородственным фагом.
Список литературы
1. Кочемасова З.Н. Микробиология:
Учеб. Пособие/З.Н. Кочемасова. - М.: Знание, 1988. - 356с.
. Покровский В.Н. Бактериофаг -
вирус бактерии/ В.Н. Покровский. - М.: Знание,1986. - 64с. - (Медицина).
. Смородинцев А.А. Беседы о вирусах/
А.А. Смородинцев. - М.: Просвещение, 1975. - 120с.
. Шаров Г. Антибиотики, бактерии и
фаги/ Г. Шаров// Наука и жизнь. - 2001. - №9. - С. 98-100.
.Захаров И.А. Курс генетики
микроорганизмов / И.А. Захаров.- Минск: Высшая школа, 1978.- 192 с.
. Хейс У. Генетика бактерий и
бактериофагов / У. Хейс. - М.: Наука, 1965.- 254 с.
. Адаме М. Бактериофаги. / Адаме М.
Москва: изд-во иностранной литературы, 1961.- 320 с.
. Крюгер Д., Роитер М., Шредер К.
Молекулярная биология взаимодействия вирус-клетка хозяина бактериофагов.
. Баев А.А. Бактериофаги. - Пущино.:
НЦБИ, 1982. - С. 3.
. Габрилович И.М. Лизогения. Минск,
1970. - С. 120-147.