Приспособления растений к водному режиму
МИНИСТЕРСТВО
ОБРАЗОВАНИЯ РЕСПУБЛИКИ БЕЛАРУСЬ
Учреждение
образования
«Брестский
государственный университет
имени
А.С. Пушкина»
Биологический
факультет
Кафедра
ботаники и экологии
Курсовая работа
ПРИСПОСОБЛЕНИЯ
РАСТЕНИЙ К ВОДНОМУ РЕЖИМУ
Выполнила:
студентка 3 курса
специальности «Биология и химия»
Кудравец О.Н.
Научный руководитель:
Левковская М. В.
Брест
2015
СОДЕРЖАНИЕ
ВВЕДЕНИЕ
. ВОДА, КАК ЭКОЛОГИЧЕСКИЙ ФАКТОР И ЕЁ ЗНАЧЕНИЕ
ДЛЯ РАСТЕНИЙ
. ЭКОЛОГИЧЕСКИЕ ГРУППЫ НАЗЕМНЫХ РАСТЕНИЙ ПО
ОТНОШЕНИЮ К ВОДЕ
.1 Гигрофиты
.2 Ксерофиты
.3 Мезофиты
.4 Сравнение адаптаций к засухе у ксерофитов и
мезофитов
. ВОДНЫЙ ДЕФИЦИТ И УСТОЙЧИВОСТЬ К ЗАСУХЕ
. АДАПТАЦИОННЫЕ ОСОБЕННОСТИ ВОДНЫХ РАСТЕНИЙ
ЗАКЛЮЧЕНИЕ
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
ВВЕДЕНИЕ
В процессе жизни растения
испытывают различные сезонные колебания климатических факторов (температуры,
влажности и др.). Если дозы этих факторов выходят за пределы, необходимые
организму, они становятся для него экстремальными. Под действием их растение
либо погибает, либо выживает, постепенно адаптируясь к ним с помощью
морфологических и физиолого-биохимических перестроек или путём изменения цикла
онтогенеза [7].
Адаптация
- это приспособление организма к определенным условиям среды, которое
достигается за счет комплекса признаков - морфологических, физиологических. В
результате адаптаций возникают растения, приспособленные к различным условиям
среды. Адаптациями объясняется различный состав экосистем разных экологических
условий [15].
Вода - важнейший экологический
фактор для всего живого на Земле. Для процессов обмена веществ со средой,
составляющих основу жизни, необходимо участие воды в качестве растворителя и
метаболита. Вода, обуславливая необходимое тургорное давление, определённым
образом участвует и в поддержании формы наземных растений как организмов, не
имеющих опорного скелета. Наконец, для большой группы растений, живущих в
водоёмах, морях и океанах, вода является непосредственной средой обитания [5].
Растения «заякорены», т.е.
ведут прикрепленный образ жизни, и потому у них возможны лишь два варианта
адаптаций: снижение интенсивности процессов жизнедеятельности в неблагоприятные
периоды (сбрасывание листьев, перезимовывание в стадии погребенных в почву
органов - луковиц, корневищ, клубней, а также семян и спор, полный криптобиоз у
моховидных) или повышение устойчивости к неблагоприятным факторам за счет
специальных физиологических механизмов [15].
Зная периоды чувствительности
растений к недостатку влаги, можно предотвратить их повреждение и значительную
потерю урожая путём современного полива, смещения сроков сева (с целью
приурочить критические периоды к более влажному времени) или с помощью приёмов,
ускоряющих либо замедляющих наступление тех или иных этапов онтогенеза [7].
Актуальность и
практическая значимость: способность растений
приспосабливаться определённым условиям влажности издавна привлекала внимание
учёных. Интерес к этой проблеме в последние десятилетия особенно возрос в связи
с освоением новых зон возделывания сельскохозяйственных культур в районах с
экстремальными почвенно-климатическими условиями. Для Беларуси характерен
неустойчивый водный режим. Здесь засушливые годы сменяются сильно
переувлажнёнными. В результате растения страдают то от недостатка, то от
избытка влаги. Поэтому для нашей страны проблема повышения устойчивости
растений как к недостатку, так и к избытку влаги чрезвычайна актуальна [7].
Цель работы: изучить
адаптационные особенности растений разных экологических групп по отношению к
различным условиям влажности.
При выполнении курсовой работы
поставлены следующие задачи:
1. Показать
закономерности структурной у функциональной организации растений,
приспособленных к определённым условиям влажности.
. Углубить и
систематизировать знания о многообразии внутренней структуры и вегетативных
органов у растений разных экологических групп по отношению к воде.
3. Выявить
структурные адаптации растений разных экологических групп по отношению к воде к
биотопам на морфологическом и анатомическом уровне организации.
1. ВОДА, КАК
ЭКОЛОГИЧЕСКИЙ ФАКТОР И ЕЁ ЗНАЧЕНИЕ ДЛЯ РАСТЕНИЙ
экологический вода
наземный растение
Тело растения на 50-90% состоит
из воды. Особенно богата водой цитоплазма (85-90%), много её и в органеллах
клетки. Вода имеет первостепенное значение в жизни растений. Процессы водоснабжения
растений тесно связанны с местообитанием. Уже незначительные колебания
водообеспечения вызывают заметные изменения в растительном покрове [6].
Вода препятствует перегреву
растений за счёт её испарения с поверхности надземных органов, главным образом
листьев, - процесса, получившего название транспирации. Благодаря
явлению осмоса и наличию у растительных клеток относительно жёстких клеточных
стенок (КС), вода поддерживает в клетках положительное тургорное давление,
придавая им нужную форму, и отвечает за их растяжение при росте [4].
При недостатке влаги вода из
вакуолей оттягивается, что приводит к утрате тургора и завяданию листьев. Рост
надземных частей растений происходит главным образом ночью, когда уменьшается
отдача воды через транспирацию, и восстанавливается тургесцентность клеток
[10].
Основной источник воды для
наземных растений - атмосферные осадки. Обеспеченность растительного покрова
влагой в первую очередь зависит от общеклиматической характеристики местности.
Большую роль играет соотношение осадков и испаряемости (суммарного годового со
свободной водной поверхности), также весьма неодинаковой в разных районах
земного шара. Области, в которых эта величина превышает годовую сумму осадков,
относят к аридным (сухим, засушливым); здесь растения испытывают
недостаток влаги в течение большей части вегетационного периода. Области, где
растения достаточно обеспечены влагой, называют гумидными (влажными).
Следует заметить, что граница между аридными и гумидными местообитаниями весьма
условна. Иногда выделяют ещё переходные - семиаридные (полуаридные)
местообитания. В случае крайне засушливых районов говорят об экстрааридных
условиях [2].
Основной путь поступления воды
в наземные растения - поглощение её из почвы через корни (и ризоиды). Вода в
почве находится в раздробленном состоянии: она вкраплена в поры разных размеров
и форм и имеет огромную поверхность раздела с почвой, содержит ряд катионов и
анионов. Поэтому почвенная влага весьма неоднородна по физическим и химическим
свойствам. Различают три основные категории почвенной воды, отличающиеся по
механизму удерживания её почвой: гравитационную, капиллярную и связанную (рис.
1) [2].
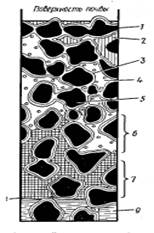
Рисунок 1 - Вода в почве: 1 -
частицы почвы, 2 - гравитационная вода осадков просачивающаяся в почву, 3 и 5 -
гигроскопическая (плёночная и коллоидная) вода, 4 - почвенный воздух с парами
воды, 6 - зона открытой капиллярной воды (часть пор заполнена воздухом, часть
водой), 7- зона замкнутой капиллярной воды (все поры заполнены водой), 8 -
уровень грунтовой воды, 9 - грунтовая вода [2].
Гравитационная вода
- подвижная вода, которая заполняет широкие промежутки между частицами почвы и
просачивается вниз под действием силы тяжести (откуда и её название), пока не
достигнет грунтовых вод [2].
Капиллярная вода
заполняет тончайшие промежутки между частицами почвы и удерживается
капиллярными силами сцепления. Под влиянием испарения с поверхности почвы
капиллярная вода может подтягиваться вверх, создавая восходящий ток влаги [2].
Связанная вода
удерживается на поверхности почвенных частиц адсорбционными силами. Количество
её значительно благодаря огромной поверхности почвенных частиц [2].
Кроме перечисленных форм воды в
почве всегда содержится парообразная влага, занимающая все свободные от
воды поры. Это почти всегда (кроме пустынных почв) насыщенный водяной пар [2].
Разные формы почвенной влаги в
неодинаковой степени доступны корневым системам растений. Наиболее легко
усваивается гравитационная вода, с большим трудом капиллярная. Вся влага, удерживаемая
в почве силами, превышающими осмотическое давление клеточного сока зоны
всасывания корня, не может поступить в растение даже при максимальной величине
его сосущей силы (при завядании). Это так называемая недоступная влага, или
мёртвый запас; он приблизительно соответствует количеству прочно связанной
воды. Количество недоступной влаги в разных почвах неодинаково и зависит от их
физических и химических свойств (размеры частиц и пор, содержание коллоидных
веществ и т. д.) [2].
Наземные организмы постоянно
теряют воду и нуждаются в регулярном пополнении её. В связи с этим у них в
процессе эволюции выработались многочисленные сложные приспособления,
регулирующие водный обмен и обеспечивающие экономное расходование влаги. Эти
приспособления носят анатомо-морфологический и физиологический характер [9].
Влажность среды часто является
фактором, лимитирующим численность и распространение организмов по земному
шару. Например, бук может жить на сравнительно сухой почве, но очень
чувствителен к влажности воздуха. Растения пустынь приспособились к низкой
влажности, а степные и особенно лесные требуют повышенного содержания паров
воды в воздухе [9].
. ЭКОЛОГИЧЕСКИЕ
ГРУППЫ НАЗЕМНЫХ РАСТЕНИЙ ПО ОТНОШЕНИЮ К ВОДЕ
Различные виды растений
по-разному реагируют на изменяющиеся условия водоснабжения. Одни приспособлены
к резким колебаниям содержания воды в среде и при этом сохраняют способность к
поглощению воды из почвы, поддерживая оводнённость клеток и тканей на должном
уровне. Для других губительным оказывается даже незначительное снижение
содержания воды в среде [4].
По отношению к воде Г. Вальтер
предложил разделить растения на две группы: пойкилогидрические (от греч.
poikilos - различный) -
переменно увлажняющиеся и гомойогидрические (от греч. homois
- одинаковый) - постоянно увлажнённые, способные поддерживать относительное
постоянство обводнённости тканей (рис. 2) [1].
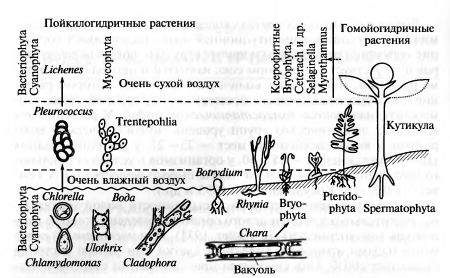
Рисунок 2 - Изменение водного
режима растений в связи с приспособлением их к наземному образу жизни; переход
от пойкилогидричности к гомойогидричности [1].
Пойкилогидрические
растительные организмы не способны сами регулировать водный обмен. К ним
относятся эпифитные и почвенные водоросли, лишайники (рис. 3), некоторые мхи и
папоротникообразные, а также некоторые покрытосеменные растения [4].

Рисунок 3 - Лишайник сосновых
лесов Cetraria islandica - типичный пойкилогидрический организм [13].
У них нет каких-либо
особенностей анатомического строения, способствующих защите от испарения (у
большинства отсутствуют устьица). Они отдают или поглощают воду как физическое
тело, и транспирация у них равна простому испарению [2].
Для подавляющего большинства
этих организмов нужна водная среда или высокая влажность воздуха. В засушливый период
они теряют воду и погибают. Некоторые представители этой группы при
дегидратации сохраняют жизнеспособность, благодаря снижению интенсивности
метаболизма и последующего перехода в состояние анабиоза (криптобиоза). С
окончанием засушливого периода они поглощают воду и возвращаются к активной
жизнедеятельности, восстанавливая рост и другие функции. К этим организмам, в
частности, относятся красные водоросли Bangia
и Porphyra, мох Funaria,
а из высших растений различные виды рода миротамнус, например Myrothamnus
flabellifolia [4].
Гомойогидрические растения
обладают способностью регулировать водный обмен, поддерживая ток воды в системе
почва - растение - атмосфера, а также надлежащую оводнённость клеток, тканей и
органов [4].
В их клеточных оболочках откладываются
водонепроницаемые вещества, поверхность листьев покрыта кутикулой и т.д.
Благодаря этому у гомойогидрических растений поддерживается на постоянном
уровне
содержание воды в клетках и
давление водяных паров в межклетниках.
Транспирация по величине,
дневной и сезонной динамике значительно отличается от свободного испарения
смоченного физического тела [2].
Они составляют большинство
обитателей суши, однако в эту группу входят и водные растения. К
гомойогидрическим растениям относятся наземные папоротникообразные, все
голосеменные и подавляющая часть покрытосеменных растений. Различия в
водоснабжении и других условиях среды привели в процессе эволюции к
формированию нескольких типов (экологических групп) гомойогидрических растений,
различающихся по характеру и эффективности регуляции их водного обмена [4].
1. Гигрофиты (греч.
hydra - влага) -
растения, обитающие при повышенной влажности, преимущественно атмосферной.
2. Ксерофиты
(греч. xeros - сухой и phyton
- растение) - растения, приспособившиеся к значительному постоянному или
временному недостатку влаги в почве или в воздухе.
3. Мезофиты (греч.
mesos - средний)
- растения, живущие в условиях достаточно умеренного увлажнения [11].
.1 Гигрофиты
Гигрофиты -
растения избыточно увлажнённых местообитаний с высокой влажностью воздуха и
почвы. Для них характерно отсутствие приспособлений, ограничивающих расход
воды, и неспособность переносить даже незначительную её потерю [3].
К гигрофитам в первую очередь
относят тропические растения, которые живут при высокой температуре и влажности
воздуха. Нередко гигрофиты обитают в тени под пологом леса (например,
папоротники) или на открытых пространствах, но обязательно на почвах переувлажненных
или покрытых водой. В умеренном и холодном климате типичными гигрофитами
являются теневые травянистые растения лесов. На открытых местах и влажных
почвах растут световые гигрофиты. Это такие, как калужница (Calta
palustris), плакун-трава (Lythrum
salicaria), росянка (Drosera),
многие злаки и осоки сырых местообитаний, из культурных растений к световым
гигрофитам относят рис, культивируемый на полях, залитых водой [12].
Характерные структурные черты
гигрофитов -тонкие нежные листовые пластинки с небольшим числом устьиц, не
имеющие толстой кутикулы, рыхлое сложение тканей листа с крупными
межклетниками, слабое развитие водопроводящей системы, тонкие
слаборазветвлённые корни [2].
Листья у них обычно крупные
(белокрыльник болотный (Calla
palustris
L.). Исключение
составляет ситник развесистый (Juncus
effuses L.)
и хвощ приречный болотный, у которых листья редуцированы, а функцию фотосинтеза
выполняют зелёные стебли [10].
Способность гигрофитов к
регуляции водного режима ограничена: устьица большей частью широко открыты, так
что транспирация мало отличается от физического испарения. Благодаря
беспрепятственному притоку воды и отсутствию защитных приспособлений
интенсивность транспирации очень высока: у световых гигрофитов в дневное время
листья могут терять за час количество воды, в 4-5 раз превышающее массу листа.
Высокая оводнённость тканей гигрофитов поддерживается в основном за чёт
постоянного притока влаги из окружающей среды. Поэтому гигрофиты можно считать
наиболее близкими к пойкилогидрическим [2].
Другие характерные
физиологические черты гигрофитов, обусловлен-ные лёгкой доступностью влаги,
низкое осмотическое давление клеточного сока, незначительная водоудерживающая
способность, приводящая к быстрой потере запасов воды [2].
В некоторых случаях у растений
сильно увлажнённых местообитаний возникает необходимость удаления избытка
влаги. Обычно это бывает, когда почва хорошо прогрета и корни активно всасывают
воду, а транспирация отсутствует (например, утром или при тумане, когда
влажность воздуха 100%). Избыточная влага удаляется путём гуттации (рис.
4) - выделения воды через специальные выделительные клетки, расположенные по
краю (или на острие) листа [2].

Рисунок 4 - Типы гуттации у
разных растений: 1 - у злаков, 2 - у земляники, 3 - у тюльпана, 4 - у молочая,
5 - у беллевалии сарматской, 6 - у клевера [2].
Избыточное увлажнение почвы
может сказываться на растениях и отрицательно, поскольку затрудняет аэрацию, а
следовательно, дыхание и всасывающую деятельность корней. Борьба растений с
избыточной влагой - это прежде всего борьба за доступ воздуха. У многих
травянистых видов избыточно увлажнённых местообитаний (прибрежных, болотных)
хорошо развита система воздушных полостей в листьях и стеблях, а также
специальная воздухоносная ткань - аэренхима (рис. 5). Благодаря этому
создаётся непрерывный путь от надземных частей к корням, по которому подземные
части растений снабжаются воздухом [2].
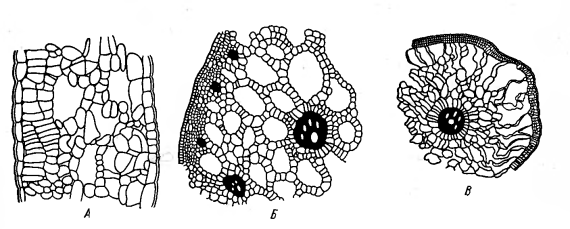
Рисунок 5 - Рыхлые ткани с
системой межклетников на поперечных срезах прибрежного растения Alisma
plantago-aquatica:
А - лист; Б - стебель; В - корень [2].
.2 Ксерофиты
Ксерофиты
- растения сухих местообитаний, способные переносить значительный недостаток
влаги - почвенную и атмосферную засуху. Они распространены, обильны и
разнообразны в областях с жарким и сухим климатом. К этой группе принадлежат
виды пустынь, сухих степей, саванн, колючих редколесий, сухих субтропиков и т.
д [2].
Ксерофиты лучше, чем все другие
растения, способны регулировать водный обмен, поэтому и во время
продолжительной засухи остаются в активном состоянии [13].
В неблагоприятных условиях они
могут сбрасывать листья и приостанавливать рост. Коллоидно-химические свойства
протоплазмы позволяют им выдерживать сильное обезвоживание [10].
Неблагоприятный водный режим
растений в сухих местообитаниях обусловлен, во-первых, ограниченным
поступлением воды при её недостатке в почве и, во-вторых, увеличением расхода
влаги на транспирацию при большой сухости воздуха и высоких температурах.
Следовательно, для преодоления недостатка влаги возможны разные пути:
увеличение её поглощения и сокращение расхода, кроме того, способность
переносить большие потери воды [2].
Большое значение для ксерофитов
имеет разнообразные структурные приспособления к условиям недостатки влаги [2].
Корневые системы обычно сильно
развиты (рис.6), что помогает растениям увеличить поглощение почвенной влаги
[2].
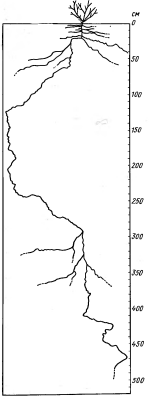
Рисунок 6 - Корневая система
пустынного кустарника Pituranthos
tortuosus [2].
По общей массе корневые системы
ксерофитов нередко превышают надземные части, иногда весьма значительно.
Корневые системы ксерофитов часто бывают экстенсивного типа (рис. 7, А), т.е.
растения имеют длинные корни, распространяющиеся в большом объёме почвы, но
сравнительно мало разветвлённые [2].
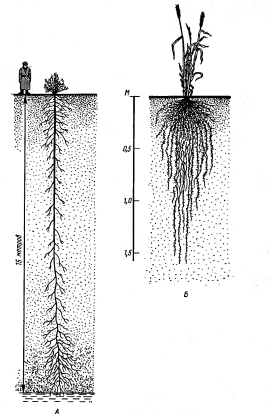
Рисунок 7 - Различные типы
корневых систем: А - экстенсивный (верблюжья колючка Alhagi
camelorum); Б - интенсивный
(пшеница) [2].
Проникновение таких корней на
большую глубину позволяет ксерофитам (например, многим пустынным кустарникам)
использовать влагу глубоких почвенных горизонтов, а в отдельных случаях - и
грунтовых вод. Встречаются и широкораскинутые поверхностные корневые системы,
приспособленные к поглощению скудных атмосферных осадков, смачивающих лишь
верхние горизонты почвы [2].
У других видов (например,
степных злаков) корневые системы интенсивного типа (рис. 7, Б): они охватывают
относительно небольшой объём почвы, но благодаря очень густому ветвлению
максимально используют почвенную влагу [2].
Надземные органы ксерофитов
также отличаются своеобразными (так называемыми ксероморфными) чертами, которые
носят отпечаток трудных условий водоснабжения. У них сильно развита
водопроводящая система, что хорошо заметно по густоте сети жилок в листьях
(рис. 8), подводящих воду к тканям. Эта черта облегчает ксерофитам пополнение
запасов влаги, расходуемой на транспирацию [2].
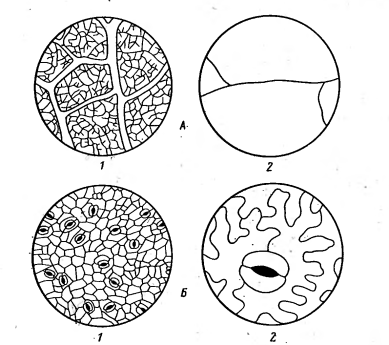
Рисунок 8 - Различие жилкования
(А), размеров и числа устьиц (Б) у ксерофитов и мезофитов: 1 - пустынный
ксерофит - Psoralea
drupacea, 2 - лесной
мезофит - Paris
quadrifolia [2].
Разнообразные структурные
приспособления защитного характера, направленные на уменьшении расхода воды, в
основном сводятся к следующему [2]
Общее сокращение
транспирирующей поверхности. Многие ксерофиты имеют мелкие, узкие, сильно
редуцированные листовые пластинки. Это особенно наглядно при сравнении облика
родственных видов из сухих и влажных местообитаний: например, мелкие
рассечённые листья степной таволги - Filipendula
hexapatela и крупные листовые
пластинки лабазника - F.
ulmaria, обитающего на
сырых лугах и опушках; узкие проволоковидные листья степного типчака - Festuca
sulcata и листья луговой
овсяницы - Festuca
pratensis и т.д. В особо
засушливых пустынных местообитаниях листья древесных и кустарниковых пород
редуцированы до едва заметных чешуек (рис. 9) [2].

Рисунок 9 - Афилльный ксерофит
- Ephedra strobilaceae
[2].
Уменьшение листовой поверхности
в наиболее жаркие и сухие периоды вегетационного сезона (рис. 10) [2].
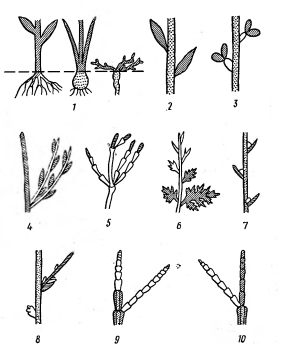
Рисунок 10 - Морфологические
типы растений засушливых областей: 1 - эфемероиды,
2 - Lycium, 3 - Zygophyllum, 4 - Retama, 5 - Ephedra, 6 - Artemisia,
7 - Reaumuria, 8 - Noaea, 9 - Anabasis, 10 -
Haloxylon persicum. Штриховка -ассимилирующие части,
отмирающие при недостатке влаги; белый цвет - части, сохраняющиеся при засухе;
точки - неассимилирующие части [2].
Для многих кустарников
среднеазиатских, североафриканских и других пустынь, а также для некоторых
видов сухих субтропиков Средиземноморья характерен сезонный диморфизм листьев:
ранней весной при ещё благоприятном водном режиме образуются относительно
крупные листья, которые летом, при наступлении жары и сухости, сменяются
мелкими листьями более ксероморфного строения с меньшей интенсивностью
транспирации. У деревьев в тропических областях с ярко выраженным сухим
периодом распространён летний листопад [2].
Защита листьев от больших
потерь влаги на транспирацию. Она достигается благодаря развитию мощных
покровных тканей - толстостенного, иногда многослойного эпидермиса, часто
несущего различные выросты и волоски, которые образуют густое «войлочное»
опушение поверхности листа. Развитие защитных покров на листьях - причина того,
что степной травой имеют тусклые, седоватые оттенки, резко отличающиеся от
яркой зелени лугов [2].
Устьица у ксерофитов обычно
защищены от чрезмерной потери влаги, например, расположены в специальных
углублениях в ткани листа («погружённые устьица»), иногда снабжённых волосками
и прочими дополнительными защитными устройствами. У ковылей и других степных
злаков существует интересный механизм защиты устьиц в самые жаркие и сухие часы
дня: при больших потерях воды крупные тонкостенный водоносные клетки эпидермиса
теряют тургор, и лист свёртывается в трубку (рис. 11); так устьица оказываются
изолированными от окружающего сухого воздуха внутри замкнутой полости, где
благодаря транспирации создаётся повышенная влажность [2].
Во влажную погоду клетки
эпидермиса восстанавливают тургор, и листовая пластинка вновь развёртывается
[2].
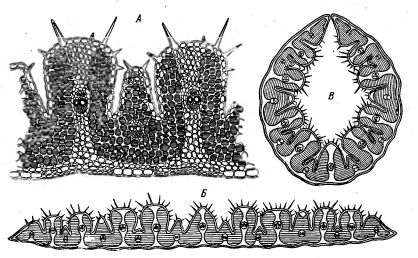
Рисунок 11 - Поперечный срез
листа ковыля-волосатика - Stipa
capillata: А - участок среза
при большом увеличении; Б - во влажную погоду (пластинка листа развёрнута); В -
при засухе (лист свёрнут) [2].
Усиленное развитие механической
ткани. Клетки тканей листьев у ксерофитов отличаются мелкими размерами и весьма
плотной упаковкой, т.е. малым развитием межклетников, благодаря чему сильно
сокращается внутренняя испаряющая поверхность листа [2].
Формирование некоторых черт
ксероморфной структуры (в частности, мелких размеров клеток) - прямое следствие
недостатка воды, тормозящего растяжение клеток в растущем листе [2].
Разнообразны физиологические
адаптации ксерофитов, помогающие им успешно выдерживать недостаток влаги [2].
У ксерофитов обычно повышено
осмотическое давление клеточного сока, позволяющее всасывать воду даже при
больших водоотнимающих силах почвы, т.е. использовать не только легкодоступную,
но и труднодоступную почвенную влагу. При достаточном водоснабжении большинство
ксерофитов имеет довольно высокую транспирацию, но при наступлении засушливых
условий (в жаркие и сухие полуденные часы, в периоды длительных летних засух)
они сильно сокращают её. При этом играет роль и закрывание устьиц, и сильное
обезвоживание листа при начинающемся подвядании [2].
Основную роль в засухоустойчивости
ксерофитов в настоящее время отводят физиологическим механизмам [2].
) К числу этих
механизмов принадлежит высокая водоудерживающая способность тканей и клеток,
обусловленная рядом физиологических и биохимических особенностей (содержание «защитных
веществ», высокая вязкость и эластичность цитоплазмы, большая доля связанной
воды в общем водном запасе и т.д.) [2].
) Большое значение для
выживания ксерофитов при резком недостатке воды имеет их способность переносить
глубокое обезвоживание тканей без потери жизнеспособности и возможности
восстановления нормального содержания воды в растении при возобновлении
благоприятных условий [2].
) Ещё одна система
адаптаций, обеспечивающая выживание ксерофитов в аридных условиях, - выработка
сезонных ритмов, дающих возможность растениям использовать для вегетации
наиболее благоприятные периоды года и резко сократить жизнедеятельность (вплоть
до впадение в состояние покоя или анабиоза) во время засухи [2].
В процессе эволюции возникли
разные группы ксерофитов, отличающимися механизмами выживания в условиях
жёсткого водного дефицита [5].
)суккуленты (ложные
ксерофиты). Это многолетние растения, запасающие воду в стеблях, листьях
или корнях. Отсюда произошло их название: succulents
по-латыни - сочный. Эти органы становятся толстыми из-за сильно развитой
водозапасающей паренхимы. После дождей в них запасается вода, которая долгое
время покрывает потребности растения [5].
Другим общим признаком
растений-суккулентов является экономное расходование воды, которое достигается
за счёт низкой интенсивности транспирации [5].
У кактусов небольшая
интенсивность транспирации связана, во-первых, с сокращением испаряющей
поверхности благодаря превращению листьев в колючки. При этом фотосинтетическую
функцию листьев выполняют стебли. Во-вторых, у них не только мало устьиц, но
днём они закрыты, а открываются лишь ночью, когда температура воздуха ниже, а
его влажность выше. В результате в отличие от других растений транспирация у
суккулентов днём идёт медленнее, чем ночью [5].
Уменьшению интенсивности
транспирации способствует толстая кутикула и опушение стеблей и листьев [5].
Корневые системы слабые,
поверхностные (рис. 12) и, по-видимому, используют не только влагу редко
выпадающих дождей, но и росу. Часто корни весьма динамичны во времени: они
отмирают в засуху, но после дождей быстро (в течение 2-4 дней) отрастают новые,
а в очередной засушливый период опять засыхают [2].
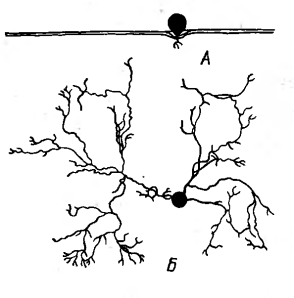
Рисунок 12 - Корневая система
кактуса - Ferocactus
wislizenii: А - вид сбоку
(корни на глубине около 2 см); Б - вид сверху [2].
Интенсивность фотосинтеза
невелика, рост и накопление массы идут очень медленно, вследствие чего они не
отличаются высокой биологической продуктивностью и не образуют сомкнутых
растительных сообществ [2].
)тонколистные ксерофиты
(полуксерофиты), представляющие собой полную противоположность первому типу.
Представителем является верблюжья колючка. Её тонкие листья с большим
количеством устьиц и проводящих пучков, не имеющие хорошо развитой кутикулы и
склеренхимы, интенсивно транспортируют: в течение час вода в тканях несколько
раз сменяется. Длинные корни (10 м и более) во много раз больше надземных
органов, достигают грунтовых вод и непрерывно снабжают побеги водой. Устьица
широко открыты даже в очень жаркие и сухие дни, поэтому фотосинтез идёт быстро.
Листья часто покрыты волосками, защищающими хлорофилл от интенсивного света
[5].
)Жестколистные ксерофиты
(настоящие ксерофиты). Эти растения способны резко сокращать транспирацию при
недостатке воды. У ряда представителей этого типа сильно развита кутикула или
опушение, отполированная поверхность листьев способна к сильному отражению
солнечных лучей. Корневая система неглубокая, но разветвлённая [5].
)Растения, избегающие засуху
благодаря короткому онтогенезу (эфемеры), не превышающему несколько недель.
Они успевают прорасти, зацвести и дать зрелые плоды в тот короткий срок
времени, когда идут весенние или осенние дожди, влажность почвы высокая и нет
экстремальных температур. К этому типу ксерофитов относятся растения степей,
например, тюльпаны, маки, вероники и др [5].
)Близки к эфемерам. Все
они сбрасывают листья, когда становится жарко и почва пересыхает, и переходят в
состояние покоя. К этому типу ксерофитов относятся колючий астрагал,
белая полынь [5].
)Растения, способные
выносить сильное обезвоживание клеток. Эти растения произрастают в
условиях, характеризующихся периодами сильной засухи [5].
.3 Мезофиты
Эта группа включает растения,
произрастающие в средних (т.е. достаточных, но не избыточных) условиях
увлажнения. Сюда относятся растения лугов, травяного покрова лесов, лиственные
древесные и кустарниковые породы из областей умеренно влажного климата [2].
Мезофиты имеют корневые системы
как экстенсивного, так и интенсивного типа, со всеми переходами между ними [2].
Для листа характерна
дифференцировка тканей на более или менее плотную палисадную паренхиму (за
исключением листьев ярко выраженного теневого характера) и рыхлую губчатую
паренхиму с системой межклетников. Сеть жилок сравнительно негустая. Покровные
ткани могут иметь отдельные ксероморфные черты (чаще всего редкое опушение), но
не столь ярко выраженные, как у ксерофитов [2].
Для них характерны умеренные
величины осмотического давления, содержания воды в листьях, предельного водного
дефицита. Величина транспирации в большей степени зависит от условий
освещённости и других элементов микроклимата. Один и тот же мезофильный вид,
попадая в разные по водоснабжению условия, обнаруживает известную пластичность,
приобретая в сухих условиях более ксероморфные, а во влажных - более
гигроморфные черты. Это
хорошо
иллюстрирует рисунок 13 и
14,
показывающие, что с усилением сухости и освещённости у мезофитов наблюдается не
только определённая ксерофилизация листа, но изменяется и интенсивность
физиологических процессов [2].
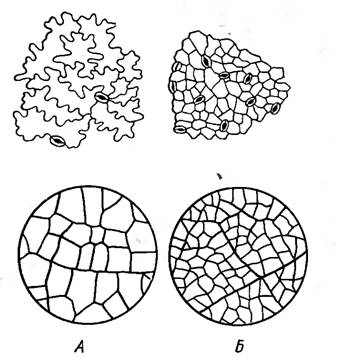
Рисунок 13 - Нижний эпидермис и
густота жилкования в листьях ясеня: А - в лесу; Б - на полезащитной полосе [2].
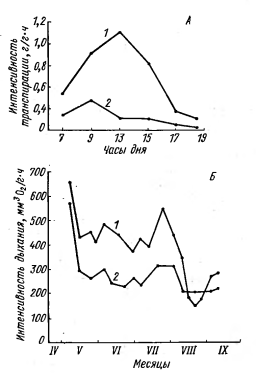
Рисунок 14 - Изменение
интенсивности физиологических процессов у травянистых растения-мезофитов в
местообитаниях с различным микроклиматом. А - дневной ход транспирации у сныти
- Aegopodium
podagraria; Б - сезонный ход
дыхания листьев у ежи - Dactylis
glomerata: 1 - на открытой
лесной поляне, 2 - под пологом дубового леса [2].
Мезофиты - группа весьма
разнообразная не только по видовому составу, но и по различным экологическим
«оттенкам», обусловленным разным сочетанием факторов в природных
местообитаниях. Они связаны переходами с другими экологическими типами растений
по отношению к воде, так что чёткую границу между ними провести очень трудно
[2].
.4 Сравнение
адаптаций к засухе у ксерофитов и мезофитов
У ксерофитов выработались
эволюционные адаптации к перенесению засухи, которые шли по пути формирования
следующих механизмов:
· Поддерживание
оводнённости тканей, необходимой для нормальной жизнедеятельности (запасание
воды в стеблях, листьях, корнях; экономное расходование воды за счёт низкой
транспирации);
· Развитие
приспособлений для нормального течения метаболизма в условиях дефицита воды в
организме (С4 - и САМ-пути фотосинтеза);
· Эффективное восстановление
(репарация) клеточных структур и функций после сильного обезвоживания;
· Быстрое завершение
онтогенеза в период достаточного обеспечения водой, до наступления засушливого
периода.
У ксерофитов вырабатываются
анатомические приспособления к недостатку воды, которые проявляются в редукции
листьев, их опушённости, наличии толстой кутикулы, заглублении устьиц в ткани
листа.
У мезофитов возникают
онтогенетические адаптации, которые генетически детерминированы, отсутствуют в
условиях нормального увлажнения, образуются в ответ на действие стрессора. Они
проявляются в сокращении потерь воды за счёт торможения увеличения листовой
поверхности; уменьшении площади листовой поверхности (сбрасывание листьев);
стимуляции роста корневой системы; закрытии устьиц; аккумуляции
низкомолекулярных осмотически активных соединений.
Когда мезофиты
приспосабливаются к временной засухе, у них выявляются признаки ксерофитов.
Возникновение ксероморфной
структуры, как и редукция листьев, опушённость, толстая кутикула служит анатомическим
приспособлением к недостатку воды [15].
3. ВОДНЫЙ ДЕФИЦИТ И
УСТОЙЧИВОСТЬ К ЗАСУХЕ
Недостаток воды в растительных
тканях возникает тогда, когда расход начинает превышать поступление. Засухи
бывают почвенные и атмосферные. Атмосферная засуха характеризуется
низкой относительной влажностью воздуха, почвенная засуха, как правило,
следует за атмосферной и возникает при длительном отсутствии дождей. Водный
дефицит вызывает серьёзные нарушения в первую очередь процессов роста и
фотосинтеза. Во время засухи наряду с обезвоживанием происходит перегрев
растений, поэтому засуха является наиболее жёстким стрессовым воздействием.
Более устойчивы к засухе такие виды растений, которые способны расходовать воду
более экономно и запасать её [7].
У растительных организмов
имеется несколько адаптивных стратегий, с помощью которых удаётся переживать
засушливые периоды. Одни виды растений способны накапливать и удерживать воду,
поддерживая таким образом необходимую степень гидратации своих клеток и
органов. Другие приспособились функционировать в условиях дефицита воды в
организме. К третьей группе относят растения, которые «избегают» засухи за счёт
организации своего жизненного цикла таким образом, чтобы вегетировать в период
достаточного обеспечения водой [7].
Многие виды растений резко
ускоряют свой рост весной, когда в почве накапливается большое количество
влаги, до наступления засушливого лета. У таких растений формируется большая
листовая поверхность и быстро развивается мощная корневая система, которая выбирает
всю влаги из почвы, накопившуюся в зимний период. При этом растения или быстро
завершают свой жизненный цикл, или запасают воду для завершения репродуктивного
этапа развития. Такой тип роста и развития растений называют детерминированным,
поскольку он определяется периодами дождей или засухи. При резком наступлении
засушливого периода растения, как правило, сбрасывают часть листьев за счёт
формирования у черешка отделительного слоя под влиянием фитогормона этилена.
Если водный дефицит развивается постепенно, то растительному организму лучше
удаётся приспособиться к недостатку воды, заранее замедлив темпы ростовых
процессов [7].
Совсем другая стратегия
развития у растений, произрастающих в условиях нормальной обеспеченности водой.
В этом случае развиваются большие листья, а вегетация и цветение идут весь
летний период. Такой тип развития растений называют недетерминированным.
У растений с этим типом роста дефицит воды лимитирует не только размеры
отдельных листьев, но и их количество [7].
Одной из самых ранних реакций
растительных организмов на водный дефицит является синтез фитогормона
абсцизовой кислоты, которая вызывает закрывание устьиц и таким образом снижает
потерю воды при транспирации. Недостаток воды инициирует рост корней в более
влажные участки почвы при условии, что в растущие кончики корней поступает
необходимое количество
фотоассимилятов.
Особое
значение
в
осморегу-ляции
имеет пролин, содержание которого резко возрастает при засухе [7].
Зная периоды особой
чувствительности растений к недостатку влаги, можно предотвратить их
повреждение и значительную потерю урожая путём своевременного полива, смещения
сроков сева (с целью приурочить критические периоды к более влажному времени)
или с помощью приёмов, ускоряющих либо замедляющих наступление тех или иных
этапов онтогенеза [14].
4. АДАПТАЦИОННЫЕ
ОСОБЕННОСТИ ВОДНЫХ РАСТЕНИЙ
Гидрофиты: эта
группа включает в себя растения, нормально растущие в воде, а в случае
укоренения на суше корни их распространяются в переувлажнённой почве, что
неблагоприятно для других растений. Низкая растворимость и небольшая скорость
диффузии кислорода в воде или в пересыщенной почве создаёт такие условия, при
которых могут расти только специально-адаптированные виды [6].
В связи со спецификой условий
существования основной структурной особенностью гидрофитов являются губчатость,
«рыхлость» их тканей, наличие крупных межклетников, полостей, заполненных
воздухом, т. е. преобладание аэренхимы. Система воздухоносных
межклетников встречается и у мезофитов, но у гидрофитов она развита особенно
сильно и является их типичной чертой строения. Полости погружённых в воду
частей растений обычно сообщаются с устьицами, расположенными на непогружённых
листьях, что способствует процессу газообмена и проникновению кислорода к
органам, расположенным в среде с малыми количествами кислорода. Имеются
воздушные полости и в плодах некоторых гидрофитов, а это обеспечивает их
плавучесть и, следовательно, способствует распространению. Губчатость тканей и
воздушные полости в листе и черешке также поддерживают их плавучесть. У
полностью погружённых растений (Elodea)
воздушные полости могут аккумулировать кислород в течение дня, а затем этот
кислород используется на дыхание ночью, когда аккумулируется СО₂
[6].
Помимо губчатости органы
подводных гидрофитов отличаются от надводных отсутствием кутикулы и перидермы,
а также не функционирующими устьицами, наличие которых говорит об эволюционном
происхождении водных растений от наземных. Отсутствие кутина и суберина у
погружённых органов способствует поглощению воды и питательных веществ
непосредственно поверхностью органа - дополнительно к тому, что растение может
получить через корни [6].
Корни гидрофитов обычно короче
и менее разветвлены, чем у ксерофитов и мезофитов, а их окончания нередко
лишены корневых волосков. Наконец, для гидрофитов характерна атрофированность
поддерживающих, механических и водопроводящих тканей; недостаток механической
ткани компенсируется плавучестью отдельных органов [6].
Листья водных растений,
погружённые в воду, сильно рассечены на узкие нитевидные доли (роголистник,
уруть, пузырчатка) (рис. 15) или имеют тонкую просвечивающую пластинку
(подводные листья кубышек и кувшинок, листья погружённых рдестов) [2].
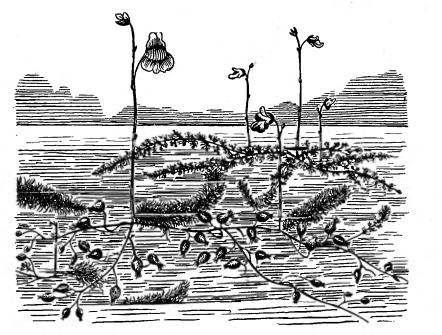
Рисунок 15 - Пузырчатки: Utricularia
grafiana (на переднем
плане) и U. minor
[2].
Подобные черты характерны и для
водорослей (например, нитчатые водоросли, рассечённые талломы харовых, тонкие
прозрачные талломы многих глубоководных видов) (рис. 16) [2].
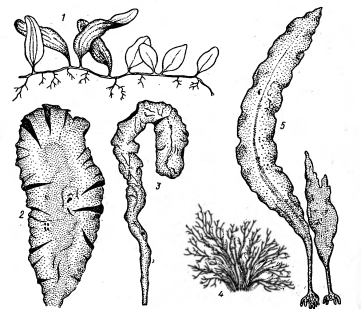
Рисунок 16 - Различные морские
донные водоросли: 1 - Caulepra, 2 - Ulva, 3 -
Enteromorpha, 4 - Ahnfeltia, 5 - Laminaria [2].
Всё это даёт возможность
гидрофитам увеличить отношение площади тела к объёму, т.е. развивать большую
поверхность при сравнительно небольших затратах органической массы [2].
У растений, лишь частично
погружённых в воду, хорошо выражена гетерофиллия - различие строения надводных
и подводных листьев на одной и той же особи. Первые имеют черты, обычные для
листьев наземных растений (дорзовентральное строение, хорошо развитые покровные
ткани и устьичный аппарат), вторые - рассеченные или очень тонкие листовые
пластинки. Гетерофиллия отмечена у водного лютика - Ranunculus
diversifolius (рис. 17), кувшинок
и кубышек (различие плавающих и подводных листьев), стрелолиста и других видов
[2].
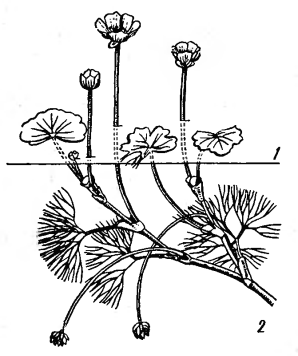
Рисунок 17 - Гетерофиллия у
водного лютика разнолистного - Ranunculus
diversifolius. Листья: 1 -
надводные, 2 - подводные [2].
ЗАКЛЮЧЕНИЕ
Растение в ходе эволюции должно
приспособиться к тем условиям водоснабжения, которые характерны для данного
местообитания. Приспособления у растений тем сложнее, чем больше дефицит влаги
[6].
Можно указать ряд
морфологических и физиологических признаков, которые отличают растения,
растущие при затруднённом водоснабжении от растений оптимальных условий [6].
Онтогенез гигрофитов
проходит при сравнительно благоприятных условиях водоснабжения и достаточной
влажности воздуха, и поэтому они не имеют каких-либо особых
анатомо-морфологических или физиологических приспособлений к засухе [6].
Признаками
ксероморфизма являются следующее:
Морфолого-анатомическое
признаки:
· уменьшение размеров
стебля при одновременном увеличении объёма корневой системы;
· относительно более
мелкие размеры клеток листьев и межклетников. Пластинка листа толще, но меньше
по размерам. Устьица более мелкие, но их больше на единицу площади. Сеть жилок
гуще, т. е. промежутки между ними меньше;
· кутикула и
клеточные стенки толще; обычно отмечается отложение кутина или восковых веществ
и развитие опушения;
· палисадная ткань
развита сильнее, губчатый мезофилл развит слабее. Клетки ксилемы по размерам
меньше, ксилема содержит больше механических элементов [6].
Физиологические
признаки:
· более высокая
интенсивность транспирации на единицу поверхности и фотосинтеза в периоды
благоприятного водоснабжения;
· более высокая
вязкость цитоплазмы и её проницаемость;
· большая
устойчивость к завяданию и долговечность растений, более раннее цветение и
плодоношение [6].
Поскольку мезофиты по
своему водному режиму занимают промежуточное положение между ксерофитами и
гигрофитами, то в строении их листьев сочетаются промежуточные черты
гигроморфной и ксероморфной организации [6].
В связи со спецификой условий
существования гидрофитов основной структурной особенностью их является
губчатость, «рыхлость» тканей, наличие крупных межклетников, полостей,
заполненных воздухом, т.е. преобладание аэренхимы. Система воздухоносных
межклетников встречается и у мезофитов, но у гидрофитов она развита особенно
сильно и является их типичной чертой строения [6].
Сравнение адаптаций разных
видов к недостатку воды возможно лишь в пределах близкородственных групп -
видов или родов, а так же отдельных органов у одного растения [6].
Нельзя утверждать, что все виды
реагируют на снижение водоснабжения одинаково. Если сравнить степень реакции
разных видов, то можно сказать, что более и менее сходно реагируют только
эволюционно близкородственные виды. Но мы не можем утверждать, что и виды
одного семейства реагируют одинаково, так как в состав семейства входят весьма
различные жизненные формы. Этим объясняется многообразие форм приспособления к
недостатку воды, разнообразие экологических групп по отношению к влажности [6].
СПИСОК
ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
1. Березина, Н. А. Экология растений:
учеб. пособие для студ. высш. учеб. заведений / Н. А. Березина, Н. Б.
Афанасьева. - М.: «Академия», 2009. - 400 с.
. Горышина, Т. К. Экология растений:
учеб. пособие / Т. К. Горышина. - М.: Высш. школа, 1979. - 368 с., ил.
. Дроздов, В. В. Общая экология: учеб.
пособие / В. В. Дроздов. - СПб.: РГГМУ, 2011. - 412 с.
. Физиология растений: учеб. для студ.
вузов / И. П. Ермакова [и др.]; под общ. ред. И. П. Ермакова. - 2-е изд., испр.
- М.:
«Академия», 2007. - 640 с.
. Кузнецов, В. В. Физиология растений:
учебник / В. В. Кузнецов,
Г. А. Дмитриева. - 2-е изд., перераб. и доп. -
М.: Высш. школа, 2006. - 742 с.
. Культиасов, И. М. Экология растений /
И. М. Культиасов. - М.: Изд-во Моск. ун-та, 1982. - 384 с.
. Медведев, С. С. Физиология растений:
учебник / C. С. Медведев. -
СПб.: Изд-во С.-Петерб. ун-та, 2004. - 336 с.
. Миркин, Б. М. Основы общей экологии:
учебник / Б. М. Миркин,
Л. Г. Наумова. - М.: «Логос», 2003. - 142 с.
В. А. Радкевич. - Минск: Высш. школа, 1983. -
320 с.
. Сергейчик, С. А. Экология: учеб.
пособие / С. А. Сергейчик. - Минск: Современная школа, 2010. - 400 с.
. Ботаника с основами фитоценологии:
Анатомия и морфология растений: учеб. для вузов / Т. И. Серебрякова, Н. С.
Воронин,
А. Г. Еленевский и др. - М.: ИКЦ «Академкнига»,
2006. - 543 с.
. Степановских, А. С. Экологи / А. С.
Степановских.-М.: ЮНИТИ-ДАНА, 2001 - 703 с.
. Чернова, Н. М. Общая экология: учеб. для
студ. пед. вузов /
Н. М. Чернова, А. М. Былова. - М.: Изд-во
«Дрофа», 2004. - 416 с.
. Шабельская, Э. Ф. Физиология растений: учеб.
пособие для биол. спец. пед. ин-тов / Э. Ф. Шабельская. - Минск: Высш. школа,
1987. - 320 с.
. Яковец, О. Г. Физиология стресса: курс
лекций / О. Г. Яковец. - Минск: БГУ, 2009. - 101 с.