Иммуногистохимические изменения в нейронах коры головного мозга при холестазе
Иммуногистохимические
изменения в нейронах коры головного мозга при холестазе
РЕФЕРАТ
Курсовая работа:37 страниц, 23рисунка, 2
таблицы.
Ключевые слова: головной мозг, фронтальная кора,
теменная кора, нейроны, структура, цитотохимия, холестаз.
Цель исследования- оценить иммуногистохимические
показатели нейронов фронтальной и теменной коры больших полушарий головного
мозга в различные сроки подпеченочного холестаза у крыс.
Объект исследования:мозг крыс при холестазе.
Предмет исследования:гистохимические изменения
нейронов мозга крыс при холестазе.
Методы и методики исследования:
Для изучения влияния холестаза на различные
отделы мозга эксперименты проводились самцах крыс Wistar весом 200 ± 25 г. Все
экспериментальные процедуры должны соответствовать Директиве Совета
Европейского сообщества (86/609 / EEC) по уходу и использованию лабораторных
животных.
Экспериментальных крыс анестезируют этиловым
эфиром, а общий желчный проток лигируют на 3-5 мм ниже слияния лобарных
протоков, применяя две лигатуры с последующим пересечением между ними. Животные
контрольной группы подвергаются фиктивной операции. На 2-й, 5-й, 10-й, 20-й,
45-й и 90-й день животных из группы контроля и из группы с холестазом
декапитировали, образцы их мозжечка собирали и фиксировали в
цинк-этанол-формалине при 4 ° С (ночь), а затем залили в парафин.
Блоки парафина (7 мкм) разрезали LeicaRM2125
(микротомом RTS, Германия). Иммуногистохимическое обнаружение проводили с
использованием поликлональных антител кролика, экспозиции в течение 20 часов во
влажной камере. Связывание первичных антител было обнаружено с использованием
Thermo Scientific Super Picture ™ Polymer Detection Kit. Связывание первичных
антител было обнаружено с использованием контейнера Expose Mouse и Rabbit
Specific HRP / DAB IHC Kit ab. 80436 (Abcam, UK).
Для стандартизации исследования во всех образцах
была исследована латеральная зона задней доли мозжечковых полушарий. Кора
головного мозга этой области двусторонне связана с сенсомоторной корой
головного мозга, что позволяет планировать и координировать быстрые движения
тела, следующие один за другим. Гистохимические препараты были исследованы,
сфотографированы и изучены морфометрически с использованием микроскопа Axioscop
2 plus (Zeiss, Германия) с цифровой видеокамерой LeicaDFC 320 (Leica, Германия)
и программой анализа изображений ImageWarp (Bitflow, США).
Средние значения, полученные у животных каждой
экспериментальной группы, анализировались непараметрической статистикой (из-за
небольшого числа животных в группах) в Statistica 10.0 для Windows (StatSoft,
США). Описательная статистика для каждого параметра включала определение
медианного (Me) и межквартильного диапазона (IQR). Различия между значениями в
контрольной и экспериментальной группах считались значимыми при p <0,05 с
использованием U-теста Манна-Уитни.
ВВЕДЕНИЕ
Желчь - секрет, выделяемый печеночными клетками,
содержащий конъюгированный билирубин, соли желчных кислот, холестерин,
фосфолипиды, белки, электролиты, микроэлементы и воду [1].
Онаобладает важными пищеварительными и не
пищеварительными функциями в организме. Нарушение оттока желчи приводит к ее
застою в желчевыводящих путях (холестаз), прекращению ее поступления в
12-перстную кишку и накоплению компонентов желчи в крови.
Холестаз - это нарушение синтеза, секреции и
оттока желчи. Холестаз может протекать с желтухой. Традиционно по
этиологическому принципу холестаз подразделяют на внутрипеченочный и
внепеченочный.
При холестазе, в первую очередь гидрофобные
желчные кислоты и неконъюгированный билирубин, увеличивающиеся в крови,
вызывают значительное токсическое воздействие на мозг. Одной из причин
вызванного холестазом повреждения головного мозга может быть гипоксия из-за
нарушений кровообращения [2]. При холестазе токсичные желчные кислоты нарушают
проницаемость и структуру гематоэнцефалического барьера [3]. Неконъюгированный
билирубин в головном мозге ухудшает энергетический обмен и индуцирует апоптоз,
дефекты клеточной мембраны, что вызывает утечку ферментов и затрудняет перенос
нейротрансмиттеров [3]. Билирубин также препятствует синтезу ДНК и белка; он
непосредственно взаимодействует с фосфолипидами клеточной мембраны и изменяет
внутриклеточный рН. Избыток билирубина индуцирует экзитотоксичность через
рецепторы NMDA глутамата и каспаз-зависимые пути [4]. Кроме того, холестаз
вызывает мальабсорбцию жира, связанную с уменьшением желчных кислот в кишечнике
и абсорбцией липидов и растворимых в липидах витаминов (A, D, E и K). Дефицит
витамина Е связан с прогрессирующим нервно-мышечным синдромом, который может
вызвать атаксию мозжечка.
Мозжечок - это не только центр баланса и
координации движений, но он также участвует в регуляции многих висцеральных
функций и высших психических процессов. В предыдущих исследованиях было
обнаружено, что холестаз повреждает нейроны мозжечка [6] и вызывает, в
частности, потерю клеток Пуркинье и гранулы нейронов [6,7]. Билирубиновая
токсичность клеток мозжечка опосредуется активацией p38 MAP-киназы [8], которая
индуцирует широкий спектр клеточной активности в ответ на внеклеточные стимулы
[9]. В предыдущих исследованиях сообщается о структурных и метаболических
нарушениях в мозжечковых клетках Пуркинье у крыс с холестазом [7]. Но данные о
состоянии синапсов и гамма-аминомасляной кислоты ГАМК-ергической структуры
мозжечка в холестазе все еще отсутствуют.
Морфологи используют термин «холестаз» для
обозначения наличия желчи в гепатоцитах и гипертрофированных клетках Купфера
(так называемый клеточный билирубиностаз), в частности в виде капелек желчи в
расширенных каналикулах (каналикулярный билирубиностаз). При внепеченочном
холестазе желчь находится в расширенных междольковых желчных протоках
(дуктулярный билирубиностаз) и паренхиме печени в виде «желчных озер».
Холестаз, существующий в течение нескольких
дней, вызывает потенциально обратимые ультраструктурные изменения. В
развернутой фазе холестаза происходит ряд гистологических изменений: расширение
желчных капилляров, исчезновение микроворсинок на каналикулярной мембране,
образование желчных тромбов в желчных капиллярах, повреждение клеточных
мембран, ведущее к повышению их проницаемости, нарушение целостности плотных
контактов и парацеллюлярная регургитация желчи в кровь, билирубиностаз,
образование печеночных розеток, пролиферация дуктул в портальных трактах,
перидуктальный отек и склероз, перипортальное и мезенхимальное воспаление,
желчные инфаркты, развитие микроабсцессов, апоптоз, образование ксантомных
клеток. Персистирующий холестаз с сопутствующим воспалением и реакцией
соединительной ткани ведет к необратимому холестазу, а через месяцы/годы к
развитию билиарного фиброза и первичного или вторичного билиарного цирроза[10].
Клиницисты диагностируют холестаз при повышении
уровня желчных кислот, специфических ферментов и холестерина в сыворотке крови.
К маркерам холестаза относят щелочную фосфатазу, лейцинаминопептидазу,
гамма-глютамилтранспептидазу, 5-нуклеотидазу. Основным клиническим признаком
холестаза является кожный зуд.
Внепеченочный холестаз связан с нарушением
оттока желчи, вызванным механическим фактором (холедохолитиаз, стриктура в
дистальной части общего желчного протока). К синдрому внутрипеченочного
холестаза ведет нарушение жизненно важной функции образования желчи под
действием лекарств, инфекционных агентов, а также вследствие аутоиммунных,
метаболических или генетических факторов.
Внутрипеченочный холестаз может быть одним из
синдромов хронических заболеваний печени (гепатиты, циррозы различной
этиологии, метаболические расстройства, опухолевые процессы печени, первичный
билиарный цирроз) или самостоятельным процессом (лекарственный холестаз,
доброкачественный рецидивирующий семейный холестаз, холестаз беременных)
[1,10,11].
Прогрессирование холестаза в ряде случаев
сопровождается проникновением билирубина и других веществ через
гематоэнцефалический барьер и накоплением в нейронах головного мозга
(преимущественно в его подкорковых образованиях, в первую очередь - ядрах
мозга). Проникновение билирубина внутрь клеток блокирует дыхательные ферменты
митохондрий, тормозит реакции с участием аденилатциклазы и К-Na-АТФ-азы,
нарушает функциональное состояние мембраны клетки и приводит к гибели нейронов.
При этом ядра головного мозга приобретают характерный желтый цвет,
обусловленный накоплением в них билирубина, что служит основанием для
использования термина «ядерная желтуха». Наряду с этим термином используется
другой термин - «билирубиновая энцефалопатия»[7,16].
Таким образом выяснение роли в этих процессах
структур головного мозга, является весьма перспективным. Применение
экспериментального холестаза, как модели патологии гепатобилиарной системы,
позволит выяснить участие нервных клеток в адаптационных процессах.
Иммуногистохимических исследований нейронов коры головного мозга при данной
патологии не проводили, что определяет актуальность данной работы.
Цель исследования- оценить иммуногистохимические
показатели нейронов фронтальной и теменной коры больших полушарий головного
мозга в различные сроки подпеченочного холестаза у крыс.
Для достижения поставленной цели необходимо
решить комплекс следующих задач:
1) провести аналитический обзор литературы;
2) изучить уровень экспрессии синаптофизина
при холестазе у крыс;
) изучить гистохимические изменения в
нейронах фронтальной и теменной коры у крыс при холестазе;
) изучить экспресии белка NeuN
в нейронах мозга при холестазе.
Объект исследования:мозг крыс при холестазе.
Предмет исследования:гистохимические изменения
нейронов мозга крыс при холестазе.
Методы и методики исследования:
Для изучения влияния холестаза на различные
отделы мозга эксперименты проводились самцах крыс Wistar весом 200 ± 25 г. Все
экспериментальные процедуры должны соответствовать Директиве Совета
Европейского сообщества (86/609 / EEC) по уходу и использованию лабораторных
животных.
Экспериментальных крыс анестезируют этиловым
эфиром, а общий желчный проток лигируют на 3-5 мм ниже слияния лобарных
протоков, применяя две лигатуры с последующим пересечением между ними. Животные
контрольной группы подвергаются фиктивной операции. На 2-й, 5-й, 10-й, 20-й,
45-й и 90-й день животных из группы контроля и из группы с холестазом
декапитировали, образцы их мозжечка собирали и фиксировали в
цинк-этанол-формалине при 4 ° С (ночь), а затем залили в парафин.
Блоки парафина (7 мкм) разрезали LeicaRM2125
(микротомом RTS, Германия). Иммуногистохимическое обнаружение проводили с
использованием поликлональных антител кролика, экспозиции в течение 20 часов во
влажной камере. Связывание первичных антител было обнаружено с использованием
Thermo Scientific Super Picture ™ Polymer Detection Kit. Связывание первичных
антител было обнаружено с использованием контейнера Expose Mouse и Rabbit
Specific HRP / DAB IHC Kit ab. 80436 (Abcam, UK).
Для стандартизации исследования во всех образцах
была исследована латеральная зона задней доли мозжечковых полушарий. Кора
головного мозга этой области двусторонне связана с сенсомоторной корой
головного мозга, что позволяет планировать и координировать быстрые движения
тела, следующие один за другим. Гистохимические препараты были исследованы,
сфотографированы и изучены морфометрически с использованием микроскопа Axioscop
2 plus (Zeiss, Германия) с цифровой видеокамерой LeicaDFC 320 (Leica, Германия)
и программой анализа изображений ImageWarp (Bitflow, США).
Средние значения, полученные у животных каждой
экспериментальной группы, анализировались непараметрической статистикой (из-за
небольшого числа животных в группах) в Statistica 10.0 для Windows (StatSoft,
США). Описательная статистика для каждого параметра включала определение
медианного (Me) и межквартильного диапазона (IQR). Различия между значениями в
контрольной и экспериментальной группах считались значимыми при p <0,05 с
использованием U-теста Манна-Уитни[16].
нейрон иммуногистохимический полушарие мозг
ГЛАВА 1. АНТИТЕЛА, СТРОЕНИЕ АНТИТЕЛ
Антитела - омолекулы иммуноглобулинов,
синтезируемые и продуцируемые В-лимфоцитами (плазматическими клетками) в
процессе развития гуморального иммунного ответа (рисунок 1.1). Специфически
связываются с комплементарными им эпитопами структурных и не структурных
антигенов. Являются естественными биомолекулами. Их активность генерируется
иммунной системой в процессе иммунного ответа на инфекционный агент или его
токсины. В организме человека циркулирует примерно 108 различных типов антител,
каждое из которых имеет уникальный антигенсвязывающий участок. Благодаря своему
разнообразию они распознают и элиминируют из организма многочисленные
чужеродные для него биологические и комплексные структуры. Секретируемые
антитела обладают разнообразными эффекторными функциями, включая нейтрализацию
антигенов, активацию системы комплемента, опсонизацию и усиление фагоцитарной
реакции. Они также обладают и регуляторными свойствами[12].
Домены - глобулярные образования в структуре
некоторых белков, сформированные дисульфидными связями. Примером являются
вариабельные и константные домены легких и тяжелых цепей молекул
иммуноглобулинов. Домены иммуноглобулинов записывают следующим образом - VH-VL,
CH1-CL1 и т. д.
Домены вариабельные - N-терминальныеV-домены
тяжелых и легких цепей иммуноглобулинов. Формируют комплементарно-детерминирующую
область молекул.
Домены константные - С-терминальные домены
тяжелых и легких цепей молекул иммуноглобулинов. Формируются примерно из
100-110 аминокислотных остатков. Называются С-доменами. Легкие цепи
иммуноглобулинов имеют по одному домену, тяжелые по 3-4 С-домена.
Области вариабельные (V-области) - наиболее
изменчивые области полипептидных цепей молекул иммуноглобулинов (Н и L).
Образуют комплементарно детерминирующие области для эпитопов антигена.
Области гипервариабельные - короткие сегменты
изменчивых по аминокислотному составу доменов легких и тяжелых цепей молекулы
иммуноглобулинов. Обусловливают идиотипическую специфичность молекулы.
Области константные - С-концевые части (домены)
тяжелой или легкой цепи молекулы иммуноглобулинов, отличающиеся постоянством
аминокислотного состава (имеются у всех классов и субклассов иммуноглобулинов).
Участок антигенсвязывающий - функциональное
образование гипервариабельных областей молекулы определенного класса
иммуноглобулинов. Образован вариабельными доменами тяжелой (Н) и легкой (L)
цепей. Комплементарен определенному эпитопу антигена. Специфически
взаимодействует с антигеном, образует с ним комплекс.аb-фрагмент - часть
молекулы иммуноглобулинов (антител). Имеет один антигенсвязывающий участок и
состоит из двух доменов одной легкой и одной тяжелой цепи, связанных
внутренними дисульфидными связями.фрагмент - часть молекулы иммуноглобулинов
(антител). Служит для связи с мембранными рецепторами иммунокомпетентных клеток
и с С1 компонентом комплемента.
Иммуноглобулины (синоним - антитела) -
глобулярные гликопротеиды крови, обладающие защитными свойствами. Молекула
иммуноглобулинов состоит из четырех полипептидных цепей - двух легких (L) и
двух тяжелых (H). Имеется два типа легких (каппа и лямбда) и пять типов тяжелых
(дельта, мю, гамма, альфа, эпсилон) цепей. В структуре молекулы
иммуноглобулинов выделяют функциональные области - Fab и Fc-фрагменты, а также
шарнирную область. Два Fab-фрагмента обусловливают способность молекулы
узнавать и связывать эпитопы антигена, Fc-фрагмент обеспечивает связывание
молекулы с соответствующим рецептором иммунокомпетентных клеток (В- и
Т-лимфоцитов, нейтрофилов, моноцитов, макрофагов, базофилов, тучных и
дендритных клеток, ЕК). Для иммуноглобулинов характерен широкий репертуар антител
разной специфичности[12].
Цепи легкие - пара сравнительно небольших
полипептидных цепей (каппа и лямбда), составляющих мономерную молекулу
иммуноглобулинов. Каждая из них состоит примерно из 220 аминокислот и
разделяется на вариабельный и константный домены. Соотношение количества каппа-
и лямбда-цепей в структуре молекул иммуноглобулинов разных видов животных
варьирует. У человека оно составляет 2:1. Биологический смысл данного
разнообразия до настоящего времени не выяснен. По соотношению каппа- и лямбда-цепей
иммуноглобулинов выявляют нарушение процесса их биосинтеза. Это может указывать
на присутствие в организме опухолевых В-лимфоцитов, продуцирующих данный тип
легкой цепи[14].
.1 Изотипы
иммуноглобулинов
Изотипы иммуноглобулинов - молекулы иммуноглобулинов,
у которых константные (С) домены тяжелых цепей генетически и структурно
различаются. Примером являются пять классов иммуноглобулинов (M, G, A, D и E),
имеющих разные типы тяжелых цепей (мю, гамма, альфа, дельта и эпсилон). Изотипы
иммуноглобулинов отличаются по молекулярной массе и иммунобиологической
активности (рисунок 1.1).
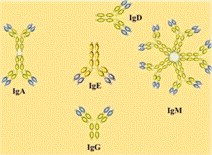
Рисунок 1.1 - Классы иммуноглобулин
Иммуноглобулины G (IgG) - иммуноглобулины с
молекулярной массой 154 кДа. Находятся в мономерной форме. Период полураспада
23-25 дней. Составляют около 85% всех иммуноглобулинов. Способны проникать
через плаценту. Концентрация их в сыворотке взрослого человека колеблется в
пределах от 7 до 18 г/л.
Известны четыре субкласса IgG: IgG1, IgG2, IgG3,
IgG4.высокоэффективны в противоинфекционной защите, являются основным классом
иммуноглобулинов, обусловливающим анамнестический или бустерный ответ.
Антитела IgGвысокоспецифичны к антигенам,
обладают высокой биологической активностью: участвуют в реакциях агглютинации,
связывания комплемента, нейтрализации, стимулируют хемотаксис.
Гамма-цепь иммуноглобулинов (молекулярная масса
51 кДа) состоит из 450 аминокислот. Тяжелая полипептидная цепь включает один
вариабельный домен (VН) и три константных (CН1, CН2 и CН3) домена. Существуют
четыре изотипа гамма-цепи. Различия в их структуре составляют 5%.
Протеолитические ферменты - пепсин и папаин - разрушают полипептидную цепь в
шарнирной области на Fab-, F(ab)2- и Fc-фрагменты.
Иммуноглобулины А (IgA) - иммуноглобулины, молекулярная
масса которых равна 160 кДа. Составляют 5-15% всех иммуноглобулинов организма.
Их концентрация в сыворотке крови 0,5-3,5 г/л, время полужизни 5-6 дней.
Различают мономерные и полимерные (ди-, три), а также сывороточные и
секреторные иммуноглобулины А.
Известны два субкласса иммуноглобулиновА - IgA1
и IgA2.
Сывороточный мономерныйIgAсоставляет 80% от
суммарного количества иммуноглобулинов этого класса, 20% приходится на его
димерную форму.
Секреторный, или экзокринный, иммуноглобулин
(sIgA) является димером или тримером. Он содержит секреторный компонент
(приобретает при прохождении молекулы через секреторную эпителиальную клетку)
и, секретируясь на внешнюю сторону слизистой, защищает ее от адгезии
микроорганизмов, нейтрализует токсины и вирусные частицы (вирионы). Этот
иммуноглобулин имеется в молозиве, кишечном, респираторном, генитальном, ушном
и других секретах, в слюне и слезах.
Классоспецифичность молекулы IgАопределяют
аминокислоты (470), составляющие альфа-цепь, молекулярная масса которой 58 кДа.
Альфа-цепь включает три константных домена (СН1, СН2, СН3) и один вариабельный
(VН). Кроме того, она несет два аллотипа - А2m (1) и A2m (2). Шарнирная область
локализуется между СН1 и СН2 доменами. Дополнительный сегмент, состоящий из 18
аминокислот, содержит цистеиновые последовательности, к которым дисульфидными
связями присоединяется J-цепь. Последовательности аминокислот, определяющие
специфичность субклассов, локализуются в первом и втором константных доменах и
в шарнирной области[14].
Иммуноглобулины М (IgM) - наиболее
высокомолекулярные иммуноглобулины (молекулярная масса 900 кДа). Составляют
5-10% всех классов иммуноглобулинов. Они пентамеры. Их концентрация в сыворотке
крови 0,4-2,2 г/л, период полураспада 5 дней. Теоретически они имеют десять активных
центров. Синтез их стимулируется полисахаридами.
Антитела IgM класса образуются преимущественно
при первичном иммунном ответе и служат свидетельством развития острого
инфекционного процесса. Они обладают высокой биологической активностью:
принимают участие в реакциях агглютинации, связывания комплемента, опсонизации,
бактериолиза, усиливают фагоцитоз. Специфичность этих антител невысокая.
Антитела изотипа IgM образуют с антигеном крупные иммунные комплексы.
Иммуноглобулины Е (IgE)- иммуноглобулины с
молекулярной массой до190 кДа и высокой цитофильностью. Находятся в основном в
мономерной форме. Составляют менее 1% суммарной концентрации иммуноглобулинов.
В норме их содержание в сыворотке крови невысокое (примерно 0,25 мг/л.). Период
полураспада IgE 2,5 дня. Они термолабильны, не образуют преципитатов с
антигеном in vitro[12].
Специфические к аллергенам IgE-антитела
фиксируются на Fc-рецепторах тучных клеток, базофилов и, таким образом,
участвуют в аллергических реакциях немедленного типа. Они также играют
существенную роль в противопаразитарной защите организма. Эпсилон-цепь
иммуноглобулинов (молекулярная масса 72 кДа) полипептидная, состоит из 550
аминокислот, одного вариабельного (VН) и четырех константных (CН1, CН2, CН3,
CН4) доменов. Константная область молекулы состоит из 428 аминокислот.
Иммуноглобулины D (IgD) - иммуноглобулины с
молекулярной массой 185 кДа. На долю IgD приходится около 1% всех
иммуноглобулинов. Период их полураспада 3 дня. Они мономеры. Экспрессируются в
основном на мембране В-лимфоцитов. Молекула IgD состоит из двух тяжелых
дельта-цепей и двух легких (лямбда или каппа).
.2 Субклассы
иммуноглобулинов
Субклассы иммуноглобулинов -
подразделениеиммуноглобулинов классов G и A на субклассы IgG1-IgG4, IgA1 и
IgA2, основанное на различиях в них аминокислотной последовательности Н-цепей.
Субклассы иммуноглобулинов G на 90% гомологичны
друг другу (за исключением гипервариабельных областей). Классы иммуноглобулинов
гомологичны друг другу только на 60%. Существенные различия в их структуре
позволяют получать моноспецифические сыворотки к отдельным классам и субклассам[14].
Субклассы IgG различаются по иммунобиологической
активности и обусловливают многие протективные свойства. По сродству молекул с
FcRI и FcRII рецепторами они распределяются в следующем порядке -
IgG1-IgG3-IgG4-IgG2. С рецепторами FcRIII типа взаимодействуют субклассы IgG1 и
IgG3. IgG4 не связывают комплемент, не взаимодействуют с белком А стафилококка.
IgG2 синтезируются в ответ на воздействие антигенов углеводной природы, с
трудом проникают через плаценту, не сенсибилизируют кожу. IgG3 наиболее эффективно
активируют систему комплемента. IgG1 наиболее активно связываются с мембраной
моноцитов.
Период полураспада разных субклассов IgG
различное:
- IgG3 - 7 дней;
- IgG1, IgG2, IgG4 - 23 дня.
Содержание их в крови также разное: IgG1 - 70%,
IgG2 - 20, IgG3 - 7, IgG4 - 3.
Что же касается субклассов иммуноглобулинов А,
то в крови IgA1 - 93%, IgA2 - 7%. Имеется определенная связь между субклассами
и Gmаллотипами. Концентрация определенных субклассов IgG ассоциируется с
Gmаллотипами. УровеньIgG1 в 1,2 раза выше у носителей G1m аллотипа, уровеньIgG2
субкласса в 1,5 раза выше для носителей G2m аллотипа, соответственно уровень
IgG3 субкласса в 2,6 раза выше у носителей G3m аллотипа. При развитии ответа
организма на инфекцию и иммунизацию это соотношение сохраняется. Кроме того,
чувствительность индивидуума к определенной инфекции ассоциируется с дефицитом
определенных субклассов иммуноглобулинов. Индивидуальные особенностиHLAсистемы
и Gmаллотипов синергически вовлечены в модуляцию иммунного ответа. 75% иммунодефицитных
состояний, связанных с субклассами иммуноглобулинов обусловлены дисфункцией Т -
иВ-лимфоцитов. Гены субклассов иммуноглобулинов располагаются на длинном плече
хромосомы 14 в следующем порядке: G3-G1, A1-G2-G4-E-A2[12].
Переключение синтеза классов Ig - процесс
перестройки генов иммуноглобулинов плазматических клеток, обеспечивающий
переключение синтезирования изотипа тяжелых цепей (например, с IgM на IgG или
IgE). Переключение биосинтеза классов антител (этап адаптивного иммунитета)
зависит от Т-системы лимфоцитов и находится под ее контролем. Этот процесс
связан с межклеточными В-Т-контактами посредством взаимодействия мембранных
молекул CD40 В-клеток и CD40LТ-лимфоцитов. На переключение классов
иммуноглобулинов ключевое влияние оказывают цитокины. Под регуляторным влиянием
ИЛ-4 осуществляется переключение биосинтеза иммуноглобулинов с IgМна
продуцирование IgG1 и IgE, а под влиянием гамма-интерферона биосинтез
субклассов IgG2a и IgG3. Трансформирующий бета-фактор обусловливает
переключение антител на биосинтез IgA и IgG2b[13,14].
ГЛАВА 2. СИНАПТОФИЗИН
Для изучения экспрессии синаптофизтина
эксперименты проводились самцах крыс Wistar весом 200 ± 25 г. Все
экспериментальные процедуры должны соответствовать Директиве Совета
Европейского сообщества (86/609 EEC) по уходу и использованию лабораторных
животных. Это исследование было одобрено Комитетом по биомедицинской этике
Гродненского государственного медицинского университета. Экспериментальных крыс
анестезируют этиловым эфиром, а общий желчный проток лигируют на 3-5 мм ниже
слияния лобарных протоков, применяя две лигатуры с последующим пересечением
между ними. Животные контрольной группы подвергаются фиктивной операции. На
2-й, 5-й, 10-й, 20-й, 45-й и 90-й день животных из группы контроля и из группы
с холестазом декапитировали, образцы их мозжечка собирали и фиксировали в
цинк-этанол-формалине при 4 ° С (ночь), а затем залили в парафин.
Блоки парафина (7 мкм) разрезали LeicaRM2125
(микротомом RTS, Германия). Иммуногистохимическое обнаружение Syn проводили с
использованием поликлонального первичного антитела кролика, Synaptophysin
Antibody (PA5-27286, Thermo Scientific, США), разбавленного 1: 400 при 4 ° C,
экспозиции в течение 20 часов во влажной камере. Связывание первичных антител
было обнаружено с использованием Thermo Scientific Super Picture ™ Polymer
Detection Kit. Глутаматдекарбоксилазу (GAD) детектировали с мышиными
моноклональными первичными антителами против GAD 67, ab. 26116 (Abcam, UK),
разбавленный 1: 2000 при 4 ° С, воздействие в течение 20 ч во влажной камере.
Связывание первичных антител было обнаружено с использованием контейнера Expose
Mouse и Rabbit Specific HRP / DAB IHC Kit ab. 80436 (Abcam, UK).
Для стандартизации исследования во всех образцах
была исследована латеральная зона задней доли мозжечковых полушарий. Кора
головного мозга этой области двусторонне связана с сенсомоторной корой
головного мозга, что позволяет планировать и координировать быстрые движения
тела, следующие один за другим. Определены уровни экспрессии Syn и GAD в структурах
коры головного мозга. Гистохимические препараты были исследованы,
сфотографированы и изучены морфометрически с использованием микроскопа Axioscop
2 plus (Zeiss, Германия) с цифровой видеокамерой LeicaDFC 320 (Leica, Германия)
и программой анализа изображений ImageWarp (Bitflow, США)[15].
Средние значения, полученные у животных каждой
экспериментальной группы, анализировались непараметрической статистикой (из-за
небольшого числа животных в группах) в Statistica 10.0 для Windows (StatSoft,
США). Описательная статистика для каждого параметра включала определение
медианного (Me) и межквартильного диапазона (IQR). Различия между значениями в
контрольной и экспериментальной группах считались значимыми при p <0,05 с
использованием U-теста Манна-Уитни[16].
У контрольных животных экспрессия синаптофизина
была обнаружена в многочисленных пресинаптических мембранах на дендритах клеток
Пуркинье в молекулярном слое мозжечка, а также в аксональных синапсах и вокруг
тел клеток Пуркинье. Экспрессия Syn была особенно высокой в гранулярных слоях
клубочков мозжечка (рисунок2.1 B, C). Интенсивность экспрессии синаптофизина у
контрольных животных существенно не изменялась в разные периоды времени после
ложной операции (таблица 2.1).
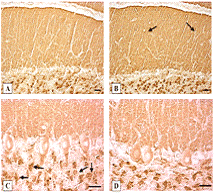
Рисунок 2.1 - Экспрессия синаптофизина в коре
мозжечка крыс[15]:
рисунок 2.1 A, C-контроль (через 10 дней после
фиктивных операций);
рисунок 2.1 B, D - 10 дней холестаза.
в молекулярном слое (показанном стрелками)
увеличение экспрессии синаптофизина; С-стрелки показывают гранулированные
слоистые клубочки. Цифровая микрофотография. Масштабные бруски - 20 мкм.
Увеличение: А, B-x200; C, D-x400.
Таблица 2.1 - Экспрессия синаптофизина в
молекулярном слое коры мозжечка крыс (единица оптической плотности ×
103[15])
|
День
|
Контрольная группа
|
Экспериментальная группа
|
|
2
|
187.03 ± 7.5
|
182.27 ± 12.7
|
|
5
|
182.77 ± 19.0
|
211.47 ± 5.62 ***
|
|
10
|
189.43 ± 15.4
|
261.18 ± 19.9 ***
|
|
20
|
187.54 ± 19.1
|
188.74 ± 17.6
|
|
45
|
186.77 ± 27.2
|
184.46 ± 5.6
|
|
90
|
180.0 ± 12.3
|
182.07 ± 14.0
|
На 2-й день после лигирования общего желчного
протока в молекулярном слое коры мозжечка экспрессия синаптофизина не
различалась, на 5-й день он увеличивался на 16%, а на 10-й день - на 38%, по
сравнению с контрольной группой. На последующих временных интервалах
исследования он не отличалсяот контрольной группы (рисунок
2.2
B, C, таблица
2.1)
Экспрессия глутаматдекарбоксилазы (GAD67) в
цитоплазме клеток Пуркинье была обнаружена (как в телах, так и дендритах), а
также вокруг тел клеток Пуркинье. В молекулярном слое также были видны
глутаматдекарбоксилаза-иммуноположительные звездчатые и корзиночные клетки,
образующие многочисленные аксональные и аксодендритные синапсы на клетках
Пуркинье (рисунок2.2 A, C). В гранулированном слое были обнаружены
глутаматдекарбоксилаза-иммуноположительные нейроны, а также терминалы на
периферии мозжечковых клубочков (рисунок2.2А, В).
На второй день после лигирования общего желчного
протока в цитоплазме клеток Пуркинье экспрессия глутаматдекарбоксилазы
уменьшилась на 10% (таблица2.2). На 5-й день холестаза экспрессия
глутаматдекарбоксилазы в цитоплазме остальных клеток Пуркинье увеличилась на
28%. Корзины вокруг перикариона стали более заметными, вытянутыми, удлиненными
в направлении гранулированного слоя и проявляли более темное окрашивание из-за
увеличения глутаматдекарбоксилазы-иммунореактивности на 11%. На 10-й день
холестаза экспрессия глутаматдекарбоксилазы в цитоплазме выживших клеток
Пуркинье увеличилась на 41% и в их корзинах на 39%, соответственно, по
сравнению с контролем (рисунок 2.2 В, таблица 2.2). Холестаз в течение 20 дней
приводил к увеличению экспрессии глутаматдекарбоксилазы в цитоплазме выживших
клеток Пуркинье на 61% и увеличению их корзин на 33% по сравнению с контролем
(рисунок2.2 D, таблица 2.2). Однако на 45-й и 90-й день после лигирования
общего желчного протока иммуногистохимические различия неопределялись по
сравнению с контрольными (таблица 2.2)[15].

Рисунок 2.2 - Экспрессия GAD67
в
коре мозжечка крыс[15]:
рисунок 2.2 А, С - контроль (10 дней и 20 дней
после фиктивных операций, соответственно);
рисунок 2.2 B - 10 дней;
рисунок 2.2 D - 20 дней холестаза., D-стрелки
показывают увеличенные корзины с повышенной иммунореактивностью вокруг тел
клеток Пуркинье. Цифровая микрофотография. Шкала шкалы -20 мкм Увеличение x400.
Таблица 2.2 - Экспрессия глутаматдекарбоксилазы
в коре головного мозга крыс (единица оптической плотности ×
103[15])
|
День
|
Тела клеток Пуркинье
|
Область вокруг клеток Пуркинье
|
|
Контроль
|
Экспериментальная группа
|
Контроль
|
Экспериментальная группа
|
|
1
|
2
|
3
|
4
|
5
|
|
2
|
182.12 ± 2.48
|
163.81 ± 4.85 **
|
197.27 ± 11.0
|
188.02 ± 14.42
|
|
5
|
205.11 ± 5.37 ***
|
189.73 ± 16.09
|
211.18 ± 8.75 ***
|
|
|
10
|
166.37 ± 3.92
|
235.04 ± 4.27 ***
|
189.72 ± 12.88
|
264.26 ± 18,64 ***
|
|
|
20
|
155.51 ± 3.86
|
250.26 ± 3.88 ***
|
189.27 ± 10.12
|
251.16 ± 17.40 ***
|
|
|
45
|
166.37 ± 3.79
|
168.13 ± 4.44
|
188.22 ± 8.70
|
190.02 ± 14.29
|
|
|
90
|
166.36 ± 3.05
|
165.43 ± 4.13
|
189.73 ± 11.27
|
184.44 ± 16.81
|
|
|
|
|
|
|
|
|
|
Временное увеличение синаптофизина в
молекулярном слое мозжечка может отражать усиленный синаптогенез, увеличение
количества синаптических пузырьков в синапсах, что является компенсаторной
реакцией.
Увеличение экспрессии глутаматдекарбоксилазы
вокруг их тел указывает на активацию клеток ГАМК-эргической области, которые могут
компенсировать гиперрезистентные клетки Пуркинье, возможно, для поддержки
функций мозжечка[8].
ГЛАВА 3. ГИСТОХИМИЧЕСКИЕ ИЗМЕНЕНИЯ В НЕЙРОНАХ
ФРОНТАЛЬНОЙ, ТЕМЕННОЙ КОРЫ
3.1 Гистохимические
изменения в нейронах фронтальной коры крыс
Холестаз в течение 2 суток вызывает только в
пятом слое нейронов статистически значимое снижение активности НАДН-ДГ на 15,7
% (Z=3,13; p=0,002); снижение содержания РНК на 8,2 % (Z=3,13;p=0,002), и
усиление активности КФ на 17,1 % (Z=-3,13; p=0,002) иЛДГ на 23,7 % (Z=-2,23;
p=0,025).
Пятисуточный подпечёночный холестаз у
экспериментальныхкрыс вызывает статистически достоверное снижение активностиСДГ
в третьем - на 7,0 % (Z=2,71; p=0,007) и пятом - на 5,2 % (Z=2,36;p=0,018)
слоях коры. Активность НАДН-ДГ в третьем слое нейронов уменьшается на 3,9 %
(Z=2,71; p=0,007), а активность ЛДГ - повышается на 13,9 % (Z=-2,14; p=0,032).
Десятисуточный подпечёночный холестаз у
экспериментальных животных вызывает существенное снижение активности одних
ферментов и усиление других. Так, активность СДГ вовтором слое фронтальной доли
коры больших полушарий снижается на 42,6 % (Z=3,13; p=0,002) (рисунок 3.1)в
третьем - на45,8 % (Z=3,13; p=0,002) и в пятом - на 45,5 % (Z=3,13; p=0,002).
Активность НАДН-ДГ у опытных животных во втором слое нейронов снижается на 43,3
% (Z=3,13; p=0,002) (рисунок 3.2), в третьемслое - на 35,4 % (Z=3,13; p=0,002),
а в пятом - на 39,1 % (Z=3,13;p=0,002) (рисунок 3.3). Активность Г-6-ФДГ
снижается на 17,1 %(Z=3,0; p=0,003), а в третьем и пятом слоях снижение более выраженноеи
составляет соответственно - 48,0 % (Z=3,13; p=0,002) и42,3 % (Z=3,13; p=0,002)
(рисунок 3.4). Активность кислой фосфатазы возрастает: во втором слое на 39,7 %
(Z=-3,13; p=0,002), втретьем - на 79,9 % (Z=-3,13; p=0,002), в пятом - 34,0 %
(Z=-3,13;p=0,002) (рисунок 3.5). Активность ЛДГ также усиливается вовсех
изучаемых слоях коры. Если во втором слое это усилениесоставляет 27,0 %
(Z=-2,11; p=0,035), в третьем - 29,8 % (Z=-2,62;p=0,009) (рисунок 3.6), то в
пятом - 29,2 % (Z=-3,13; p=0,002). Содержание РНК уменьшается в цитоплазме
нейронов всех изучаемых слоев коры: на 17,3 % (Z=2,11; p=0,035), 15,3 %
(Z=3,13;p=0,002) и 21,9 % (Z=3,13; p=0,002) соответственно во 2, 3 и 5
слояхкоры фронтальной доли (рисунки 3.7).
Через 45 суток выявлено статистически достоверное
повышение активности ферментов в нейронах второго слоя для СДГ - на6,3 %
(Z=-2,75; p=0,006), для КФ - на 11,8 % (Z=-2,36; p=0,018) и снижение активности
Г-6-ФДГ на 6,8 % (Z=2,11; p=0,035). В третьем слоеотмечено повышение активности
КФ на 17,5 % (Z=-3,13; p=0,002) ив пятом слое повышение активности для Г-6-ФДГ
на 8,6 % (Z=-2,87;p=0,004)[28].
Через 90 суток после перевязки общего жёлчного
протока невыявлено статистически достоверных гистохимических изменений[16].
Рисунок 3.1 - Активность СДГ в нейронах 2 слоя
фронтальной коры[16]: активность СДГ в нейронах 2 слоя фронтальной коры мозга
крыс в контроле - А (10 суток после ложной операции) и снижение её в опыте - Б
(10 суток холестаза). Окраска по Нахласу и др. Ув. 200. Цифровая
микрофотография

Рисунок 3.2 - активность НАДН-ДГ в нейронах 2
слоя фронтальной коры[16]: активность НАДН-ДГ в нейронах 2 слоя фронтальной
коры мозга крыс в контроле - А (10 суток после ложной операции) и снижение её в
опыте - Б (10 суток холестаза). Окраска по Нахласу, Уокеру, Зелигману. Ув. 800.
Цифровая микрофотография

Рисунок 3.3 - Активность НАДН-ДГ в нейронах 5
слоя фронтальной коры[16]: Активность НАДН-ДГ в нейронах 5 слоя фронтальной
коры мозга крыс в контроле - А (10 суток после ложной операции) и снижение её в
опыте - Б (10 суток холестаза). Окраска по Нахласу, Уокеру, Зелигману

Рисунок 3.4 - активность Г-6-ФДГ в нейронах 5
слоя фронтальной коры[16]: активность Г-6-ФДГ в нейронах 5 слоя фронтальной
коры мозга крыс в контроле - А (10 суток после ложной операции) и снижение её в
опыте - Б (10 суток холестаза). Окраска по Гесс, Скарпели, Пирсу. Ув. 400.
Цифровая микрофотография.

Рисунок 3.5 - Активность КФ в нейронах 5 слоя
фронтальной коры[16]: активность КФ в нейронах 5 слоя фронтальной коры мозга
крыс в контроле - А (10 суток после ложной операции) и снижение её в опыте - Б
(10 суток холестаза). Окраска по Гомори. Ув. 1200. Цифровая микрофотография

Рисунок 3.6 - Активность КФ в нейронах 5 слоя
фронтальной коры[16]: активность ЛДГ в нейронах 3 слоя фронтальной коры мозга
крыс в контроле - А (10 суток после ложной операции) и снижение её в опыте - Б
(10 суток холестаза). Окраска по Гесс, Скарпели, Пирсу. Ув. 1200. Цифровая
микрофотография

Рисунок 3.7 - Содержание РНК в нейронах 3 слоя
фронтальной коры[16]: содержание РНК в нейронах 3 слоя фронтальной коры мозга
крыс в контроле - А (10 суток после ложной операции) и снижение её в опыте - Б
(10 суток холестаза). Окраска по Эйнарсону. Ув. 1200. Цифровая микрофотография
Таким образом все вышесказаноесвидетельствует о
тяжелых морфо-функциональных нарушенияхво фронтальной коре мозга при холестазе
у крыс, что приводитк гибели части её нейронов. С другой стороны, в
сохранившихсянейронах развиваются значительные адаптационные изменения,ведущие
к восстановлению структуры и метаболизма нейронов вотдалённые сроки после
устранения холестаза[28].
3.3 Гистохимические
изменения в нейронах теменной коры крыс
Холестаз в течение 2 суток приводит к изменению
активностиферментов: во втором слое нейронов активность ЛДГ увеличивается на
15,8 % (Z=-2,36; p=0,018); в 3 слое снижается: СДГ - на 3,2 %(Z=1,98; p=0,048),
НАДН-ДГ - на 6,8 % (Z=2,87; p=0,025), Г-6-ФДГ - на5,9 % (Z=2,36; p=0,018) и
содержание РНК - на 4,5 % (Z=2,24; p=0,025);в 5 слое активность КФ повышается
на 3,0 % (Z=-2,62; p=0,009).
После 5-суточного холестаза происходит
статистически значимое снижение активности оксидоредуктаз: СДГ снижается во
2слое на 8,4 % (Z=2,43; p=0,015); активность НАДН-ДГ во 2 слое снижается на
19,1 % (Z=3,0; p=0,003) и в 3 - на 9,7 % (Z=2,57; p=0,01);активность Г-6-ФДГ в
клетках 3 слоя снижается на 9,7 % (Z=2,86;p=0,004). Активность КФ во 2 и 5
слоях повышается на 11,9 %(Z=-2,29; p=0,022) и 16,1 % (Z=-2,57; p=0,01)
соответственно. ЛДГ также повышается во 2 слое на 15,9 % (Z=-3,0; p=0,003) и в
5 - на 13,6 % (Z=-3,0; p=0,003).
Холестаз в течение 10 суток приводит к снижению
активностиоксидоредуктаз в теменной доле коры больших полушарий головногомозга.
В частности, активность СДГ снижается во втором слоена 37,8 % (Z=2,36;
p=0,018), в третьем - на 48,7 % (Z=3,13; p=0,002) и впятом - на 45,7 % (Z=3,13;
p=0,002) (рисунки 6.18, 6.19). АктивностьНАДН-ДГ у опытных крыс во втором слое
клеток уменьшается на30,9 % (Z=3,13; p=0,002) от исходных контрольных значений.
Нейроны третьего слоя имеют ту же закономерность: здесь в опыте активность
снижена на 8,6 % (Z=2,36; p=0,018) от контрольных данных.
В пятом слое клеток снижение активности не такое
значительное:в опыте активность уменьшается на 5,8 % (Z=2,75; p=0,006) от
контрольных значений (рисунки 6.20, 6.21, 6.22). Г-6-ФДГ во всех изученных
слоях также имеет тенденцию к снижению в опыте: на 33,9 %(Z=3,13; p=0,002),
14,8 % (Z=3,13; p=0,002) и 36,1 % (Z=3,13; p=0,002)соответственно во 2, 3 и 5
слоях коры больших полушарий (рисунок 3.8). Кислая фосфатаза после
десятисуточного холестаза активизируется во всех изученных структурах: во
втором слое - на 7,1 %(Z=-2,87; p=0,004), в третьем слое - на 51,0 % (Z=-3,13;
p=0,002), в пятом слое - на 29,3 % (Z=-3,13; p=0,002) по сравнению с
контрольными данными (рисунок 3.9). Активность ЛДГ усиливается во второмслое -
на 49,7 % (Z=-3,13; p=0,002), в третьем слое - на 35,9 % (Z=-3,13;p=0,002) и в
пятом слое - на 25,6 % (Z=-3,13; p=0,002) по сравнениюс контролем (рисунки
3.10, 3.11). Содержание РНК имеет тенденцию к снижению. После эксперимента ее
количество у опытныхживотных уменьшается во втором слое - на 33,6 % (Z=3,13;
p=0,002),в третьем слое - 23,3 % (Z=3,13; p=0,002) и в пятом слое - 22,9
%(Z=3,13; p=0,002) по сравнению с контрольными данными (рисунки 3.12-3.19).
Через 45 суток обнаружено достоверное изменение
активности ферментов только в третьем слое коры: для Г-6-ФДГ выявленоеё
снижение на 14,5 % (Z=2,24; p=0,025), для КФ и ЛДГ повышениесоответственно на
11,3 % (Z=-2,36; p=0,018) и 6,6 % (Z=-2,24; p=0,025),и снижение содержания РНК
- на 6,5 % (Z=-1,98; p=0,048)[29].
У животных через 90 суток после начала
эксперимента не обнаружено статистически значимых изменений активности
ферментов и содержания РНК в перикарионах нейронов[16].

Рисунок 3.8 - Активность СДГ в нейронах 2 слоя
теменной коры[16]: активность СДГ в нейронах 2 слоя теменной коры мозга крыс в
контроле - А (10 суток после ложной операции) и снижение ее в опыте - Б (10
суток холестаза). Окраска по Нахласу и др. Ув. 400. Цифровая микрофотография
Рисунок 3.9 - Активность СДГ в нейронах 5 слоя
фронтальной коры[16]: активность СДГ в нейронах 5 слоя фронтальной коры мозга
крыс в контроле - А (10 суток после ложной операции)и снижение ее в опыте - Б
(10 суток холестаза). Окраска по Нахласу и др. Ув. 800. Цифровая
микрофотография

Рисунок 3.10 - Активность НАДН-ДГ в нейронах 2
слоя теменной коры[16]: активность НАДН-ДГ в нейронах 2 слоя теменной коры
мозга крыс в контроле - А (10 суток после ложной операции) и снижение ее в
опыте - Б (10 суток холестаза). Окраска по Нахласу, Уокеру, Зелигману. Ув. 400.
Цифровая микрофотография
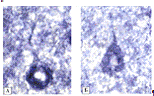
Рисунок 3.11 - Активность НАДН-ДГ в нейронах 3
слоя теменной коры[16]: активность НАДН-ДГ в нейронах 3 слоя теменной коры крыс
в контроле - А (10 суток после ложной операции) и снижение ее в опыте - Б (10
суток холестаза). Окраска по Нахласу, Уокеру, Зелигману. Ув. 800. Цифровая
микрофотография

Рисунок 3.12 - Активность НАДН-ДГ в нейронах 5
слоя теменной коры мозга[16]: активность НАДН-ДГ в нейронах 5 слоя теменной
коры крыс в контроле - А (10 суток после ложной операции) и снижение её в опыте
- Б (10 суток холестаза). Окраска по Нахласу, Уокеру, Зелигману. Ув.
400.Цифровая микрофотография

Рисунок 3.13 - Активность Г-6-ФДГ в нейронах 5
слоя теменной коры[16]: активность Г-6-ФДГ в нейронах 5 слоя теменной коры
мозга крыс в контроле - А (10 суток после ложной операции) и снижение её в
опыте - Б (10 суток холестаза). Окраска по Гесс, Скарпели, Пирсу. Ув. 300.
Цифровая микрофотография

Рисунок 3.14 - Активность КФ в нейронах 2, 3, 5
слоёв фронтальной коры[16]: активность КФ в нейронах 2, 3, 5 слоёв фронтальной
коры мозга крыс в контроле - А (10 суток после ложной операции) и повышение её
в опыте - Б (10 суток холестаза). Окраска по Гомори. Ув. 100. Цифровая
микрофотография

Рисунок 3.15 - Активность ЛДГ в нейронах 2 слоя
теменной коры[16]: активность ЛДГ в нейронах 2 слоя теменной коры мозга крыс в
контроле - А (10 суток после ложной операции) и снижение её в опыте - Б (10
суток холестаза). Окраска по Гесс, Скарпели, Пирсу. Ув. 400. Цифровая
микрофотография
Рисунок 3.16 - Активность ЛДГ в нейронах 3 слоя
теменной коры[16]: активность ЛДГ в нейронах 3 слоя теменной коры мозга крыс в
контроле - А (10 суток после ложной операции) и снижение её в опыте - Б (10
суток холестаза). Окраска по Гесс, Скарпели, Пирсу. Ув. 1000
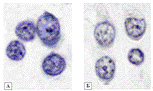
Рисунок 3.17 - Содержание РНК в нейронах 2 слоя
теменной коры[16]: содержание РНК в нейронах 2 слоя теменной коры мозга крыс в
контроле - А (10 суток после ложной операции) и снижение её в опыте - Б (10
суток холестаза). Окраска по Эйнарсону. Ув. 1000. Цифровая микрофотография
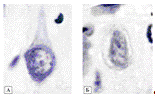
Рисунок 3.18 - Содержание РНК в нейронах 3 слоя
теменной коры[16]: содержание РНК в нейронах 3 слоя теменной коры мозга крыс в
контроле - А (10 суток после ложной операции) и снижение её в опыте - Б (10
суток холестаза). Окраска по Эйнарсону. Ув. 1000. Цифровая микрофотография

Рисунок 3.19- Содержание РНК в нейронах 5 слоя
теменной коры[16]: содержание РНК в нейронах 5 слоя теменной коры мозга крыс в
контроле - А (10 суток после ложной операции) и снижение её в опыте - Б (10
суток холестаза). Окраска по Эйнарсону. Ув. 1000. Цифровая микрофотография
Таким образом после перевязки общего жёлчного
протока у крыс в теменной коре мозга развиваются несколько более глубокие
структурные и гистохимические изменения по сравнению с фронтальной корой[29].
ГЛАВА 4. ЯДЕРНЫЙ БЕЛОК НЕРВНЫХ КЛЕТОК NEUN
Белок NeuN локализуется в ядрах и перинуклеарной
цитоплазме большинства зрелых нейронов ЦНС млекопитающих. NeuN был открыт в
1992 году группой исследователей, получившей моноклональные антитела (клон
А60), выявлявшие этот известный ранее ядерный белок [17]. Антитела к NeuN
позволяют выявлять нейроны, но не связываются с глиальными клетками. Синтез
белка NeuN начинается в постмитотических нейробластах на достаточно поздних
стадиях дифференцировки [18].Следует отметить, что этот маркер не вполне
применим при изучении развития нервных клеток мозжечка, поскольку он не
синтезируется в клетках Пуркинье и некоторых нетипичных нейронах коры мозжечка
[19]. Для выявления последних в качестве селективных нейрональных маркеров
могут быть использованы кальбиндин и кальретинин.
Из оригинальной работы Mullen
и соавторов[21] следовало, что экспресия этого белка связана с нейрональной
дифференцировкой и созраняется в течение всей жизни клеток, что в свою очередь
указывало на роль этого белка как постоянного регулятора общих проявлений
нейронального фенотипа. Однако, несмотря на продолжительное использование антител
к белку NeuN в практике
научных исследований и клинико-диагностических исследований[22-25] конкретные
функции данного белка в нервных клетках долгое время оставались непонятными.
Ранее было описано в исследованиях профессора
Зиматкина, что у потомства крыс с холестазом, вызванным на 17-сутки
беременности нарушается морфогенез палеоцеребеллюма. В раннем периоде
постнатального развития это выражается в торможении роста коры, нарушении
динамики развития наружного зернистого слоя, размеров перикарионов и плотности
расположения клеток Пуркинье. В более позднем периоде онтогенеза это
проявляется в уменьшении количества нормохромных клеток Пуркинье и увеличении
гиперхромных, а также содержания в их цитоплазме рибонуклеопротеинов.
На 2-15 сутки в нейронах наружного зернистого
слоя у потомства крыс с холестазом обнаружено отставание убыли экспрессии
даблкортина, который является маркером незрелых нейронов, а во внутреннем
зернистом слое - уменьшение плотности расположения иммунопозитивных по NeuN
нейронов, замедление нарастания в них экспресии NeuN.
Это свидетельствует о замедлении миграции нейронов из наружного зернистого слоя
и формирования внутреннего зернистого слоя в постнатальном онтогенезе[20].
При электронно-микроскопическом исследовании
клеток Пуркинье палеоцеребеллюма 7- и 15-суточных крысят группы «холестаз» ГрЭС
развита хуже, чем в контроле, обнаруживалась ее дезорганизация, на цистернах
меньшее количество рибосом, наблюдались участки цитоплазмы, заполненные
преимущественно многочисленными свободными рибосомами и полисомами, что
свидетельствует о замедлении дифференцировки клеток Пуркинье. Митохондрии
характеризовались полиморфизмом, наблюдалась их гипертрофия, в некоторых
просветление матрикса, очаговый лизис крипт, увеличение числа лизосом, которые
контактируют и измененными митохондриями. Наблюдались тесные контакты
митохондрий с органеллами. В некоторых клетках Пуркинье наблюдалось расширение
цистерн гладкой эндоплазматической сети. В ядре наблюдается усиленный выход
субъединиц рибосом в цитоплазму. На 45 сутки цитоплазма гиперхромных клеток
Пуркинье более осмиофильна, имеется большое количество свободных рибосом и
уменьшение связанных, митохондрии имели просветленный матрикс с нечеткими
кристами, гипертрофия эндоплазматической сети и комплекса Гольджи[20].
ЗАКЛЮЧЕНИЕ
В исследовании лигирование общего желчного
протока у крыс индуцирует временное увеличение (на 5-й и 10-й день) экспрессии
синаптофизина в молекулярном слое коры мозжечка. Большинство синапсов
образованы терминалами аксонами гранулярной клетки (параллельные волокна) на
дендритах клеток Пуркинье. Syn, интегральный мембранный белок синаптических
везикул, встречается во всех нервных окончаниях и участвует в синаптической
передаче [11,12], играя важную регуляторную роль в образовании синапсов [13].
Временное увеличение синаптофизина в молекулярном слое мозжечка может отражать
усиленный синаптогенез, увеличение количества синаптических пузырьков в
синапсах, что является компенсаторной реакцией. Холестаз, который приводил к
гибели гранулярных клеток и клеток Пуркинье [8], структурные и метаболические
нарушения в мозжечковых клетках Пуркинье, появились на 5-й день после перевязки
желчных протоков. Он достиг максимума в 10-20 дней и уменьшился в 45-90-е дни.
В исследовании, проведенном в 20-90-е дни после лигирования желчного протока,
экспрессия синаптофизина в молекулярном слое коры мозжечка не отличалась от
экспрессии в контрольной группе. Восстановление может быть объяснено ростом
новых добавочных желчных протоков.
Увеличение экспрессии глутаматдекарбоксилазы в
выживших ГАМК-эргических клетках Пуркинье на 5-20-й день холестаза указывает на
их активацию. Увеличение экспрессии глутаматдекарбоксилазы вокруг их тел
указывает на активацию клеток ГАМК-эргической области, которые могут
компенсировать гиперрезистентные клетки Пуркинье, возможно, для поддержки
функций мозжечка.
В заключение, холестаз у крыс индуцирует
увеличение экспрессии синаптофизина и глутаматдекарбоксилазы в коре мозжечка,
необходимых для поддержки функций мозжечка.
В раннем периоде постнатального развития это
выражается в торможении роста коры, нарушении динамики развития наружного
зернистого слоя, размеров перикарионов и плотности расположения клеток
Пуркинье. В более позднем периоде онтогенеза это проявляется в уменьшении
количества нормохромных клеток Пуркинье и увеличении гиперхромных, а также
содержания в их цитоплазме рибонуклеопротеинов.
На 2-15 сутки в нейронах наружного зернистого
слоя у потомства крыс с холестазом обнаружено отставание убыли экспрессии
даблкортина, который является маркером незрелых нейронов, а во внутреннем
зернистом слое - уменьшение плотности расположения иммунопозитивных по NeuN
нейронов, замедление нарастания в них экспресии NeuN.
Это свидетельствует о замедлении играции нейронов из наружного зернистого слоя
и формирования внутреннего зернистого слоя в постнатальном онтогенезе.
После перевязки общего жёлчного протока укрыс во
фронтальной коре мозга развиваются глубокие структурные и гистохимические
изменения. Незначительные изменения отдельных параметров появляются на 2-5
сутки опыта, а достигаютмаксимального уровня они через 10-20 суток. Затем у
выжившихживотных, у которых образуются обходные жёлчевыводящие путии холестаз
исчезает, постепенно нормализуется. К отдалённымпоследствиям холестаза (90
сутки) можно отнести очаги отсутствия нейронов во всех слоях фронтальной коры,
уменьшение в нихобщего числа нейронов и увеличение числа глиальных клеток.
Этосвидетельствует о тяжелых морфо-функциональных нарушенияхво фронтальной коре
мозга при холестазе у крыс, что приводитк гибели части её нейронов. С другой
стороны, в сохранившихсянейронах развиваются значительные адаптационные
изменения,ведущие к восстановлению структуры и метаболизма нейронов вотдалённые
сроки после устранения холестаза[28].
После перевязки общего жёлчного протока у крыс в
теменной коре мозга развиваются несколько более глубокие структурные и
гистохимические изменения по сравнению с фронтальной корой. Они также
развиваются постепенно и максимального уровня достигают через 10-20 суток.
После 45 суток (у выживших животных) они частично нормализуются, а через 90
суток в сохранившихся нейронах - полностью нормализуются. В отдаленные сроки в
коре определены очаги отсутствия нейронов во всех слоях, уменьшение в них
общего числа нейронов, но увеличение числа глиальных клеток[29].
БИБЛИОГРАФИЧЕСКИЙ СПИСОК
1. Болезни
печени и желчевыводящих путей. Под ред. В.Т.Ивашкина. М.: Медицина, 2005.
2. Green J, Beyar R, Bomzon L,
Finberg JP, Better OS (1984) Jaundice, the circulation, and the kidney. Nephron
37:
145-152. [Crossref] <http://www.ncbi.nlm.nih.gov/pubmed/6738765>
3. Greenwood J, Adu J, Davey
AJ, Abbott NJ, Bradbury MW (1991) The effect of bile salts on the permeability
and ultrastructure of the perfused, energy-depleted, rat blood-brain barrier. J
Cereb Blood Flow Metab 11: 644-654. [Crossref]
<https://www.ncbi.nlm.nih.gov/pubmed/2050752>
4. Ostrow JD, Pascolo L, Shapiro SM,
Tiribelli C (2003) New concepts in bilirubin encephalopathy. Eur
J Clin Invest 33: 988-997. [Crossref] <http://www.ncbi.nlm.nih.gov/pubmed/14636303>
. Grojean S, Koziel V, Vert
P, Daval JL (2000) Bilirubin induces apoptosis via activation of NMDA receptors
in developing rat brain neurons. Exp Neurol 166:
334-341.
6. Francavilla R, Miniello VL,
Brunetti L, Lionetti ME, Armenio L (2003) Hepatitis and cholestasis in infancy:
clinical and nutritional aspects. Acta Paediatr Suppl 91:
101-104. [Crossref] <http://www.ncbi.nlm.nih.gov/pubmed/14599052>
. Emelyanchik SV, Zimatkin SM
(2014) Structural and histochemical changes in Purkinje cells in the rat
cerebellum in cholestasis. Neurosci Behav Physiol 44: 467-471.
. Lin SZ, Yan C, Wei X, Paul
SM, Du YS (2003) p38 MAP kinase mediates bilirubin-induced neuronal death of
cultured rat cerebellar granule neurons. Neurosci Lett 353:
209-212.
. Zarubin T, Han J (2005)
Activation and signaling of the p38 MAP kinase pathway. Cell Res 15: 11-18.
[Crossref] <http://www.ncbi.nlm.nih.gov/pubmed/15686620>
10. Надинская
М.Ю. Заболевания печени, протекающие с синдромом внутрипеченочного холестаза.
Consilium medicum 2001; 4 (6): 286-92.
11. Gores
GJ. Mechanisms of cell injuri and death in cholestasis and hepatoprotection by
ursodeoxycholic acid. J Hepatol 2002; 32 (Suppl. 2): 11-3.
. Хаитов
Р.М., Игнатьева Г.А., Сидорович И.Г. Иммунология. Норма и патология. Учебник. -
3-е изд., М., Медицина, 2010. - 752 с.
. Хаитов
P.M., Игнатьева Г.Л., Сидорович И.Г. Иммунология. Учебник // Москва. Медицина.
- 2000. - 432 с.: илл.
. Хаитов
Р.М., Ярилин А.А., Пинегин Б.В. Иммунология. Атлас. М. 2011 г. - 624с.
15. Karnyushko,
O.A., Zimatkin, S.M. Synaptophysin and glutamate decarboxylase expression in
the rat cerebellum structures in cholestasis // Brain Nerves. - 2017. - Vol. 1,
№ 2. - Р.1-3.
16. Емельянчик,
С.В., Зиматкин, С.М. Мозг при холестазе: монография. - Гродно : Гродненский
государственный университет , 2011. - 265 с.
17. Mullen
R. J., Buck C. R., Smith A. M. NeuN, a neuronal specific nuclear protein in
vertebrates // Development. 1992. Vol. 116. № 1. P. 201-211.
18. Коржевский
Д. Э., Петрова Е. С., Кирик О. В., Отеллин В. А. Оценка дифференцировки
нейронов в эмбриогенезе крысы с использованием иммуноцитохимического выявления
даблкортина // Морфология. 2008. Т. 133. № 4. С. 7-10.
19. Wolf
H. K., Buslei R., Schmidt-Kastner R. et al. NeuN:
a useful neuronal marker for diagnostic histopathology // J. Histochem.
Cytochem. 1996. Vol. 44. № 10. P.
1167-1171.
20. Карнюшко,
О.А. Нарушения развития нейронов мозжечка у потомства крыс с холестазом и их
коррекция УДХК / О.А. Карнюшко, С.М. Зиматкин // Новости мед.-биол. Наук. 2016.
Т. 12, №4. С. 185-190.
. Mullen
R. J., Buck C. R., Smith A. M. NeuN, a neuronal specific nuclear protein in
vertebrates / Development. 1992. V. 116. P. 201-211.
. Hess
D. C., Hill W. D., Martin-Studdard A., Carroll J., Brailer J., Carothers J.
Bone marrow as a source of endothelial cells and NeuN-expressing cells after
stroke / Stroke. 2002. V. 33. P. 1362-1368.
. Tanvig
M., Blaabjerg M., Andersen R. K., Villa A., Rosager A. M., Poulsen F. R.,
Martinez-Serrano A., Zimmer J., Meyer M. A brain slice culture model for
studies of endogenous and exogenous precursor cell migration in the rostral
migratory stream / Brain Res. 2009. V. 1295. P. 1-12.
. Weyer
A., Schilling K. Development and cell type-specific expression of the neuronal
marker NeuN in the murine cerebellum / J. Neurosci. Res. 2003. V. 73. P.
400-409.
. McPhail
L. T., McBride C. B., McGraw J., Steeves J. D., Tetzlaff W. Axotomy abolishes
NeuN expression in facial but not rubrospinal neurons / Exp. Neurol. 2004. V.
185.P. 182-190.
. Williamson,
C. Intrahepatic Cholestasis of Pregnaney / С.
Williamson, V.L. Geenes// Obstet. Gynecol. - 2014. - Vol. 124, N. 1. - P.
120-133.
27. Дудук,
Н. И. Холестаз беременных и его последствий для матери и потомства / Н. И.
Дудук, С. М. Зиматкин// Журн. ГрГМУ. - 2011. - №1. - С. 3-6.
. Емельянчик,
С.В., Зиматкин, С.М. Структурные и гистохимические изменения в нейронах
теменной коры мозга крыс при подпеченочном холестазе // Новости
медико-биологических наук. №1 (Т.7), 2013. - С. 38-44.
. Емельянчик,
С.В. Гистохимические изменения в нейронах 2 и 3 слоев фронтальной коры
головного мозга при 10-суточном экспериментальном холестазе у крыс / С.В.
Емельянчик, С.М. Зиматкин, П.И. Суходольский // Актуальные проблемы экологии -
2007: тезисы докладов III Международной научно-практической конференции, г.
Гродно, 21 - 23 ноября 2007 г./Учреждение образования «Гродненский гос. ун-т
им. Я.Купалы»; отв. ред. Н. Канунникова. - Гродно : ГрГУ, 2007. - С.92-93.