Изучение годичной динамики и общей структуры сообщества коллембол в двух биотопах: в черноольшанике мертвопокровном и таволговом
Содержание
Введение
Глава 1. Общая характеристика отряда Collеmbola
. Морфология
.1 Размер и общий вид
1.2 Окраска
.3 Покровы и строение кутикулы
.4 Хетом
.5 Голова
.6 Усики
1.7 Глаза
1.8 Грудь
.9 Ноги
1.10 Брюшко
1.11 Жизненные формы
1.12 Адаптации к основным факторам среды
.13 Структура и динамика популяции
1.14 Население коллембол
.15 Черноольшаник
Глава 2. Материалы и методика
.1 Выбрать район исследования
2.2 Полевые учеты
2.3Техника экстракции
2.4 Камеральная обработка
2.5 Таксономический анализ
2.6 Математическая обработка
данных
2.7 Жизненные формы
.8 Пороги доминирования
2.9 Список использованных характеристик
Глава 3. Результаты
3.1 Температура и влажность
3.2 Видовой состав
3.3 Динамика общей численности(экз. на м2)
3.4 Жизненные формы
3.5 Динамика структуры сообщества
Глава 4. Обсуждение
Выводы
Благодарности
Список литературы
Сайты
Приложение
Введение
collеmbola адаптация популяция доминирование
Изучение почвенных животных в нашей стране началось в конце 19 века,
а к середине 20 века (1939г.) она оформилась в самостоятельную область
– почвенную зоологию, ее возглавил Меркурий Сергеевич Гиляров (1912- 1985). В 1944-1955 старший научный
сотрудник, с 1955 заведующий
лабораторией почвенной зоологии института эволюционной морфологии и экологии АН СССР; одновременно
(1949-1978) профессор Московского
педагогического института им. В. И. Ленина.
Более полувека назад началось активное
использование количественных методов в полевых исследованиях - это
позволило изучать сообщества и
популяции почвенных животных, а также выявить их роль в экосистеме.
К почвенным животным
относятся организмы, живущие
в подстилке или в почве: простейшие, черви, моллюски и членистоногие. Их называют
педобионтами. В данной работе мы будем анализировать мелких почвенных членистоногих- микроартропод, а
если быть точнее коллембол.
По международной классификации микроартроподы- это сборная группа мелких почвенных членистоногих, включающая:
клещей, коллембол, многоножек-симфил,
мелких жуков-ощупников и их личинок.
Микроатроподы обитают в ходах и полостях почвы. Входят в группу мезофауны. В отличии от макрофауны (дождевые черви,
мокрицы, личинки насекомых) они не
способна самостоятельно проделывать ходы.
Коллембол или ногохвостки- это мелкие насекомые, которые можно встретить на поверхности почвы,
в подстилке и на глубине,
возле корней деревьев по всему
земному шару. Они являются важнейшим компонентом
почвенных сообществ и участвуют в круговороте веществ (осуществляют гумификацию растительных остатков) активируют
почвенную микрофлору и являются
индикаторами продуктивности почв, реагируют на рН и гидротермического режима. Поэтому их изучение имеет не только теоретическое, но и практическое
значение. Из-за связи коллембол с почвой на всех этапах жизненного цикла и их
чувствительностью к изменениям
окружающей среды- делает эту группу удобным объектом для изучения почвенных сообществ.
Для моей работы было выбрано два биотопа
черноольшаника. Черноольшаники - своеобразные растительные сообщества,
образующиеся в поймах малых рек,
оврагах и ложбинах стока, характерные для территории
средней полосы России. Особенность этих ассоциаций определяется динамическим режимом увлажнения и средообразующей ролью ольхи черной (Álnus glutinósa), которая вступает в сембиотические
отношения с азотфиксирующими бактериями [17].
Изучение сообществ микроартропод, обычно
рассматривает:
• Общую характеристику (численность, разнообразие, структуру и т.д.)
• Типологию сообщества
• Пространственную структуру
• Динамику (суточную, сезонную, годичную, многолетнюю)- это процессы изменения ее основных
биологических показателей (численность,
биомасса, структуры) во времени в зависимости от экологических факторов.
• Изменения в ходе сукцессии (первичная, вторичная, конвейерная, циклическая).
• Разделение экологических ниш
• Реакцию на антропогенные воздействия
• И многие другие вопросы [14].
Цель моей работы: изучить годичную динамику и общую структуру сообщества коллембол в двух
биотопах: в черноольшанике мертвопокровном
и таволговом.
Для решения этой цели были поставлены следующие задачи:
1. установить видовой состав и структуру населения ногохвосток в черноольшанике;
2. рассмотрим изменение общей численности (тыс. экз. на м^2) по годам
в черноольшанике;
3.динамику доминантных видов (по Энгельману);
. выяснить характер
годичной динамики ногохвосток и их вертикальное распределение (жизненные
формы) коллембол на этих участках
Глава 1.
Общая характеристика отряда Collеmbola
К отряду Collembola
(коллемболы или ногохвостки) относятся
первичнобескрылые насекомые (Apterygota), достигающие 10 мм, но чаще
всего 1-2 мм. Это самая большая группа насекомых среди первичнобескрылых. Известно около 8 тысяч видов. Они распространены повсеместно, в умеренных широтах,
в тропиках так же они встречаются и в
Арктике, и в Антарктике, то есть всюду, где есть мхи и лишайники. Они живут
среди гниющих остатков растений, на поверхности и в глубоких слоях почвы, некоторые обитают и на
поверхностях растений, есть виды, которые перешли
к обитанию на поверхности воды и очень мало видов,
которые живут на трупах.
В отличие от остальных насекомых их брюшко состоит из 6 сегментов, на которых находятся
специфические придатки:
· на I
членике брюшка - вентральная трубка
· на III - зацепка
· на IV -
прыгательная вилка [2] (Рис.1).
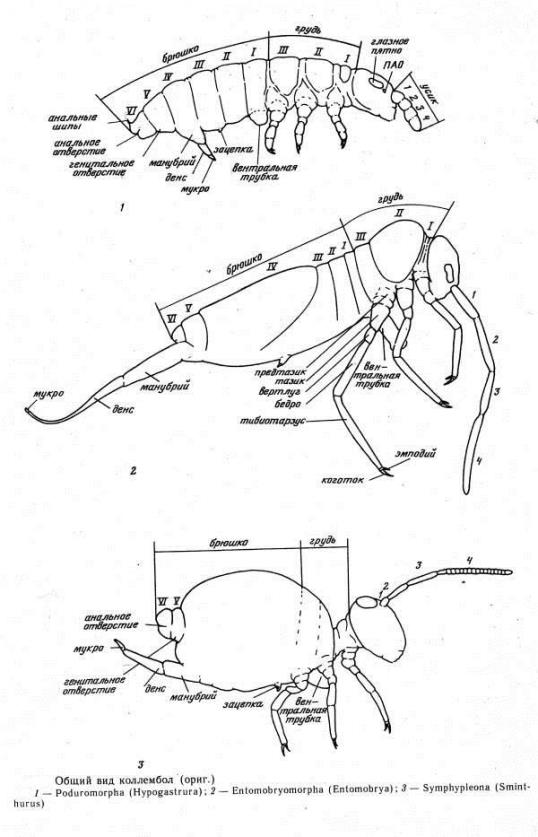
Рис.1
Общий вид коллембол ( из Определитель коллембол фауны СССР, 1988)
В спокойном состоянии прыгательная вилка подогнута под тело и удерживается зацепкой. При прыжке вилка
разгибается и тем самым подбрасывает
тело вверх (Рис.2). Фиксация тела коллембол осуществляется благодаря вентральной трубке, выделяющей слизь.
В природе существуют прыгающие и не прыгающие виды
коллембол. Прыгающие - это те виды,
которые обитают на поверхности, в подстилке и на растениях. Они имеют хорошо
развитую прыгательную вилку. У видов обитающих на поверхности растений и на поверхности почвы тело уплощено
с боков и очень компактное, а также слитные сегменты
брюшка. Живущие в подстилке занимают
промежуточное положение: развитая вилка и членистое брюшко. Не прыгающие или по другому их называют
почвенные, являются
почвообразователями. У таких форм вилка отсутствует и из-за этого они только ползают на коротких грудных
ногах. Их тело сегментировано,
следовательно, они могут изгибать своё тела для перемещения в ходах. Некоторые виды похожи на многоножек.
Ногохвостки по форме тела и окраске разнообразны (Рис.3). Как правило
виды, обитающие в почве белые, а
обитающие на поверхности растений зеленоватые. Обитающие в подстилке- бурые, серые и нередко ярко
окрашенные или с металлическим блеском.

Рис. 2
Стадии прыжка. ()
Голова у них хорошо заметна
с членистыми усиками
(4-6 члеников). Глаза в виде пятнашек
по бокам головы
и представляют собой скопление
простых глазков. Хорошо развиты глаза у поверхностных видов, а у почвенных их нет, но у них бывают особые
пятнышки- орган восприятия влажности. Ротовая часть слабо
развита и находится в головной капсуле. Три
грудных сегмента, снабженные короткими ногами. Грудная часть неясно отделена от брюшной.

Рис.3 Разнообразие коллембол. (из Жизни животных, 1967) Ногохвостки: 1 - Ceratrimeria paucispinosa, 2 - Entomobrya
salta, 3 - E. nivalis, 4 -
Tibiolatra latronigra, 5 - Parachaetoceras pritchardi, 6 - Paronana maculosa, 7 - Pseudachorutes brunneus, 8
- Neanura muscorum, 9 - Katianna purpuravirida,
10 - Bourletiella arvalis, 11 - Parakatianna hexagona, 12 - Isotoma exiguadentata, 13 - Onychiurus
fimetarius, 14 - Tomocerura rubenota, 15 - Proisotomurus novae- zealandiae, 16 - Triacanthella setacea.
У ногохвосток своеобразное размножение. Самцы откладывают сперматофор в виде капелек, в отсутствии самок, а
после самки захватывают его половым
отверстием (Рис.4). Яйца своеобразной формы и чувствительны к влажности (Рис.5). Из яиц выходят
маленькие коллемболы, похожи на
взрослой особи.
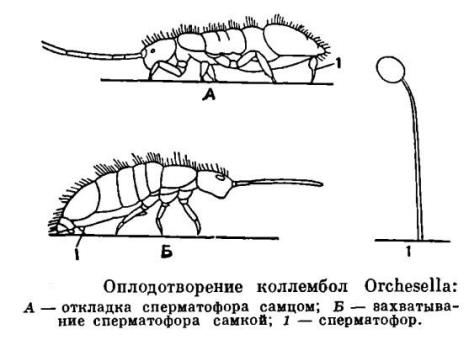
Рис.4 (из Жизни животных, 1967)
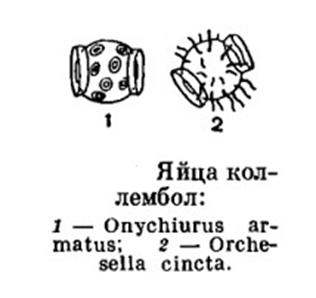
Рис.5 (из Жизни животных,
1967)
Коллемболы играют значительную роль в круговороте веществ в природе, то есть способствуют разложению
опаду, а также гумифицируют
органические вещества. Большинство видов питается пыльцой, спорами и мицелием грибов, зелеными водорослями,
разлагающими тканями растений и
животных в очень редких случаях. Однако некоторые виды могут питаются живыми
тканями растений, тем самым скелетируя листья и могут быть вредителями, разнося споры грибов и способствуя
заболеванию растений.
Морфология
Идентификация коллембол основана на внешней морфологии и на строении ротовых частей, существуют
работы, которые основанные на
строении нервной, трахейной и других систем
органов.
1.1 Размер и общий вид
Как раньше говорилось размер коллембол варьируют
от 1 до 10 мм. Для
взрослых форм наименьший размер примерно составляет 0,2-0,7 мм (Neelidae и некоторые Poduromorpha)
Внешние признаки:
Ранее упоминалось о том, что ногохвостки делятся
на две группы по внешнему виду: членистобрюхие- с удлиненным телом
с хорошо видными члениками и слитобрюхие- с округлым или шаровидным телом со слитыми сегментами (Рис.1).
Также у них есть характерные признаки это: пропорции тела с конечностями, прыгательная вилка, глаза,
пигмент. Все эти и другие
признаки показывают адаптивность к тому или другому месту обитанию в разных ярусах биогеоценоза. Они рассматривают и
жизненные формы ногохвосток [2], [1].
1.2 Окраска
Встречается как пигментная окраска, то есть с красящие
вещества и структурная- это
окраска зависит от расположения чешуй и особенностей
ультраструктуры кутикулы. Кутикула сама по себе бесцветна, но может быть желтоватого цвета или янтарного это
зависит от ее толщены.
Пигмент локализуется в живых геотермальных клетках под кутикулой
и также в специальных пигментных клетках глаза. В природе чаще всего встречаются оттенки сине-фиолетового
пигмента. Окраска при фиксации
меняется- становится светлее. Так же существует красный и желтый пигмент, который локализуется в жировом
теле животного. Эта окраска встречается у многих Poduromorpha. Этот пигмент разрушается в спирте и
из-за этого особи становятся белыми.
Следовательно, яркая окраска
хорошо развита у наземных
обитателей и ослабленная у пещерных и почвенных форм. С возрастом она становится интенсивнее и по большей
части на дорсальной стороне.
Есть равномерная пигментация (Poduromorpha). У тропических видов-
контрастная «Предупреждающая» окраска из белых или цветных пятен, или полос. Например, у Entomobryidae и Symphypleona, а у Entomobryomorpha есть металлический блеск из-за
ребристой чешуи [2].
1.3 Покровы и строение кутикулы
Тело ногохвосток покрыто хетами и чешуйками. Функции покрова включает в себя: защита от
механических повреждений и от иссушения тела.
Защиту от иссушения осуществляют и утолщение кутикулы или сложная орнаментация структуры
эпикутикулы (поверхностный слой). Степень этих
защитных структур зависит от условий обитания.
У коллемболы кожное дыхание, следовательно, кутикула целиком или частично проницаема для газов и водных
испарений. Это еще зависит от степени развития
эпикутикулы [4]. У большинства почвенных форм сплошной
эпикутикулы нет, но присутствуют восковые
колпачки на вершинах
бугорков. Они обеспечивают несмачиваемость покровов [31]. У атмобионтных Symphypleona покровы с многослойной эпикутикулой.
Развитие сплошной водонепроницаемой и
несмачиваемой эпикутикулы характерно для
нейстонных форм (Podura aquatic).
В данном случае эпикутикула защищает
от диффузии воды из вне осмотического
пути.
У коллембол поверхность кутикулы не гладкая, но под микроскопом грануляция видна лишь у Poduromorpha, а также у некоторых Isotomidae и Symphypleona. Реальное строение кутикулы можно
рассмотреть только с помощью
сканирующего электронного микроскопа [36]. Грануляция делится на первичную, вторичную и третичную.
Гранулы на кутикуле могут быть первичными (исходно треугольные по форме и соединяются между собой
связками (комиссурами) [38]. Они присутствуют практически у всех коллембол на
определенных участках тела: на усиках,
вблизи основания хет. Гранулы так же могут быть вторичный
и третичных гранил [36], [27]. Форма первичных гранул может изменятся, то есть:
могут быть с прямыми (Cyphoderus, Entomobrya),
вогнутыми (некоторые Heteromurus, Orchesella) либо выпуклыми сторонами, а также превращается почти в круглые или
эллиптические у некоторых почвенных форм (Tullbergia, Arrhopalites). Модификацией треугольных гранул
являются четырехугольные (слияние 2
треугольных) (Dimorphotoma). Для покровов представителей, обитающих в сухих
условиях, характерные плоские многогранные гранула с большим числом комиссур (до15-17). В световом микроскопе они
видны как сетчатые или мозаичные.
Вторичные гранулы образуются при распухании первичных (Hypogastruridae) либо при выталкивании вверх трех первичных гранул (Neanurinae). Вторичные гранулы могут быть
коническими или полусферическими.
Обычно они разделены участками покровов с первичной
грануляцией. Они видны в световой микроскоп.
При объединении вторичных образуются третичные (Neanurinae).
Третичная и реже вторичная грануляции часто сопровождаются ретикуляцией покровов (образованием
сетчатой структуры на внутренней поверхности
кутикулы в результате локального утолщения.
Ложные глазки (псевдоцелли) у Onychiuridae- кольцевидные
кутикулярные структуры, окруженные валиком и с характерным внутренним строением. Они часто без четких
хитиновых границ. Ложные глазки
располагаются на дорсальной стороне головы, у основания усиков и по заднему краю, на сегментах груди и
брюшка, на базальных участках ног,
вентральной стороне головы.
Диаметр ложных глазков
примерно 5-6 мкм. Глазок отделен от окружающего покрова
вторичными гранулами, глубокой
бороздкой с первичной грануляцией. На внутренней поверхности глазка имеются рёбрышки. Также некоторые
авторыговорят, что через ложные
глазки выделяется полостная жидкость для отпугивания хищников [40], [21].
Возможно, что это вещество продуцируется секреторными клетками, лежащими
пол ложными глазками.
Псевдопоры энтомобриид- небольшие круглые участки покрова на тергитах. Их функция не известна. Но используются как маркеры в связи с расположением специализированных хет [26].
Есть участки с ультрамелкой грануляцией в световом микроскопе кажутся гладкими. Также они имеют характерное
расположение и могут быть участками
прикрепления мышц.
Сенсорное поле встречается на определенных местах
на голове, груди,
и имеют по краям модифицированные хеты
[2]
1.4 Хетом
Хеты относят к структурным образованиям кутикулы и гиподермы. Хеты- это волоски
или щетинки. Они выполняют функцию
механорецепции и защиты
покровов. Где хеты прикрепляются к кутикуле есть щетинковая пора (лунка). Пора может быть округлой,
эллиптической, удлиненной,
расширенной, равной диаметру хеты.
Номенклатуру хет разработали Массу и Эллисон [39]: поверхности может быть
гладкой, с зазубренными, реснитчатые, опушённые односторонне или со всех сторон. Имеют продольную или косую исчерченность, нести ламеллы, расширенные в основании или
на вершине (шпателевидние). По форме
различают притупленные, сплошные, головчатые,
булавовидные. Головчатые хеты могут иметь округлую, треугольную или воронковидную вершину.
Шипы- это сильно утолщенные модифицированные хеты, часто сидящие на папиллах. Если папилл нет, то от обычных хет
они буду отличаются отношение длины
к диаметру. Хеты имеют диаметр, не
превышающий 0,1 собственной длины. Шиповидные хеты- диаметр от 0,1 до 0,25 длины. Шипы -свыше 0,25. Шипы
могут и ветвится.
У многих коллембол подотряда Entomobryomorpha на теле и придатках
имеются чешуйки- сильно модифицированные хеты. Их форма могут быть разнообразной.
Они обычно располагаются в несколько
слоев- черепицеобразно,
предохраняя покровы от иссушения.
На теле ногохвостки также присутствуют разнообразные сенсиллы, выполняющие функцию
специализированных рецепторов. Типичные
сенсиллы имеют удлинённую, палочковидную форму так же различают вздутые, округлые, ланцетовидные и т.д. Ворсинки тоже являются сенсиллами.
Хеты, покрывающие тело, обычно разной длины. Следовательно, их разделяют на макро- (самые длинные),
мезо-, микро- хеты (самые маленькие).
Если различие в длине хет не превышает 1/5 то говорят о гомохетности.
Среди специализированных хет есть трихоботрии- длинные, тонкие, гладкие и покрытые ресничками хеты. У Symphypleona они в виде гладкого
тонкого жгутика на куполовидном основании- ботриотрихиямя. Длинные, часто густо покрытые ресничками
и без куполовидного основания называют
лазиотрихиями встречается у Entomobryomorphs.
Если основания лазиотрихии большое и расположено в
глубине- это истинные лазиотрихии есть у
Tomoceridae; ложные лазиотрихии некоторых Isotomidae имеют небольшое поверхностное основание [39].
Разработана частичная номенклатура хет, которая используется для морфологических исследований отдельных групп.
В современной таксономии коллембол используется
хетотаксия, т.е. число, расположение, форма и
отношение длин отдельных хет на поверхности тела и конечностях. Начало изучению хенотаксии положил Р.
Йосии [43]. В дальнейшем многие авторы
использовали описание хетома (набор хет) для диагностики видов и построения филогенетических схем высших
таксонов. Также созданы разные
номенклатурные системы для отдельных групп- [24], [25], [33], [30], [37].
Традиционным является описание хетомов половозрелый стадий. Р. Йосии [44] выделил у коллембол
гомохетность- размер и форма хет примерно
одинаковые, гетерохетность- дифференцированные хеты. По количеству дорсальных хет выделял поли- или плюрихетность
(большое число хет), олигохетность, ахетность.
У некоторых таксонов возможно образование новых хет в дополнение к примитивному хетому- неохетоз или
наоборот уменьшение числа хет по
сравнению с примитивным вариантом- паурохетоз
[2].
1.5 Голова
Голова более или менее овальной, или пятиугольной формы. У большинства Entomobryomorpha и Poduromorpha голова прогнатическая, то есть ротовое отверстие на переднем
конце и ее длинная оси параллельна оси
тела. У остальных подотрядов голова в основном гипогнатическая, ротовое
отверстие обращено вниз, а длинная ось головы перпендикулярна оси тела.
У Symphypleona усики прикрепляются к «макушке»-
верхней части головы, а у остальных к
средней или передней части.
На голове можно различить: наличники, лоб, верхнюю губу, область
усиков, глаз, щек и темени.
Если посмотреть область
рта в профиль, то она будет прямоугольной, но у некоторых подотрядов с редуцированным ротовым
аппаратом она имеет вид
конуса.
Ротовой аппарат энтогнатного типа: мандибулы и максиллы расположены в ротовой полости, наружу выступает только
апикальные части. Сверху прикрыты
верхней губой с рядом хет (прямые, изогнутые
или раздвоенные). Расположение хет на верхней губе обозначается
формулой. В таксономии используют
структуру дистального края, строение первичных
и вторичных верхнегубных папилл.
Нижняя губа ограничивает ротовую полость
снизу. Она состоит из двух симметричных частей, которые разделяются вентральной линией и образуют так называемый
нижнегубной треугольник. Каждая часть
состоит из ментума, субментума (границы не заметны) и лабиальной пальпы (тактильная функция), которая несет хеты и
папиллы. У форм с сосущем ротовым
аппаратом папиллы редуцируются. Форма и структура хет на основании нижней губы
и вдоль латеральных краев
нижнегубного треугольника важна в таксономии многих групп.
У коллембол известно три типа строения ротового аппарата: грызущий, режуще- сосущий и колюще-
сосущий. У видов с грызущим ротовым
аппаратом мандибулы имеют жевательную пластинку с многочисленными рядами конических зубов,
которые перетирают пищи,
а также есть апикальные
зубцы. Структура у разных таксонов относительно постоянна. У форм с сосущим ротовым аппаратом малярная
пластинка на мандибулах утрачена, а
апикальная часть модифицирована или редуцирована. У Brachystomellinae
мандибулы отсутствуют.
Максилы устроены сложнее: кардо лежит в основании, прикрепляется к головной капсуле при помощи
фулькрум. Служит шарниром,
то есть помогает в движении остальной части максиллы. Далее
идет стипес- центральная и удлиненная часть. К нему дистально
причленяются наружная и внутренняя
лопасть (галеа и лациния). Наружная лопасть состоит из максиллярного щупика и сублобальной пластинки с несколькими
волосками. У форм с жующим ротовым
аппаратом головка максиллы устроена сложнее.
Подвиды, питающиеся размягченными тканями или жидким содержимым, максиллы упрощаются и используются для
прокалывания стенок клеток или оболочек
гиф [2], [1].
1.6 Усики
Усики так же называют антенны. У большинства таксонов они состоят из четырех первичных члеников,
каждый с собственной мускулатурой.
Нумерация члеников ведется от основания. У некоторых один или два последних членика поделены на
несколько или много вторичных (без
собственной мускулатуры)- Tomoceridae, Entomobryomorpha, Symphypleon. Кольцевые вторичные членики томоцерид хорошо заметны по рядам круговых хет. У взрослых есть вариация,
например, у Orchesellinae первый и вророй членик поделен на два, то есть усики 6- члениковые, а у Poduromorpha и Neelidae 3-й и 4-й членики более или менее
слиты. Так же дистальные части могут обламываться и регенерация может быть не полная- Entomobryomorpha. У большинства коллембол 4-й членик длиннее 3-го.
На антеннах располагаются различные рецепторы. У Sminthurididae 2-й и 3-й членики самцов коленчато
изогнутые и приспособлены для удержания
самки в период размножения. Между основанием усиков и глазным пятном располагается постантеннальный
орган. Его функции до конца еще не
изучены, но возможно это хемо- и гигрорецепция [1], [2].
.7 Глаза
Глаза у коллембол состоят максимум из восьми простых глазков. Последние расположены на темном
овальном или четырехугольном пятне.
Размеры глазков могут
быть одинаковыми или неодинаковыми. У почвенных,
подстилочных и почвенных форм встречаются все стадии редукции и окраски пятна, а в некоторых семействах
вовсе отсутствуют.
.8 Грудь
Грудь коллембол состоит
из трех сегментов. Сегмент делится на тергит-
спинная часть, стернит- брюшная часть и они соединяются плейральной областью. Грудь от брюшка не отделяются. Каждый
сегмент несет пару ног.
.9 Ноги
Ноги состоят из следующих частей: из тазика, вертлуга, бедра и голенелапки. К дистальной части
голенелапки примыкает коготковый
сегмент, по-другому он называется- претарзус, с одной или двумя хетами. Претарзус несет крупный непарный
коготок- эмподий. У некоторых внешняя
сторона коготка прикрыта туникой- прозрачная мембрана, либо имеется латеральная базальная зубчатая
ламелла. При ходьбе в состоянии покоя
коллембол опираются на кончиках коготка, касаясь тибиотарзальными волосками субстракта. Так же используют как опору и эмподии.
1.10 Брюшко
Брюшко разделено на сегменты, принято подразделяют на шесть. Если все
сегменты приблизительно одного размера, их называют гомономные. Если IV
сегмент, который несет прыгательную вилку длиннее остальных, то в этом случае называют гетеромными. У членистобрюхих границы
сегментов хорошо видны из-за
того, что там более тонкая грануляция или идет полоса без хет. У некоторых видов первые четыре сегмента и
последние сегменты брюшка слиты. В первом случаи
их называют- “большим
брюшком”, а во втором-
“малым брюшком”. У слитобрюхих все сегменты сливаются.
I, III, IV сегменты несут специфические придатки- характерны только для отряда Collembola.
На I сегменте располагается брюшная
трубка или по другому ее называют
вентральной. Это округлый или цилиндрический
придаток, состоящий из основания и двух боковых лопастей- мешковидные коксальные пузырьки. Брюшная
трубка может быть короткой как у Poduromorpha или крупной- Entomobryomorpha, но выпячивавшиеся пузырьки обычно не заметны. Вентральная трубка
несет хеты, их количество различно
у разных таксонов. Брюшная
трубка выполняет несколько функций: во-первых,
это газообмен и осморегуляция, адсорбция воды и ионов. У некоторых виды из
вентральной трубки выделяется специальная жидкость для ухода за хетами, а также это жидкость удерживает влажность
[34]. Во-вторых, используется для
фиксации тела при движении на гладкой вертикальной поверхности и использует для перевёртывания при не
правильном приземлении после прыжка
[34]. В этом случаи пузырьки (или тяжи) выворачиваются.
У видов имеющих прыгательную вилку на III сегменте располагается
зацепка, которая обычно несет хеты. У нее непарное основание и дистальные парные концы с зубцами
(максимум 4-5 шт.), которые и закрепляют вилку.
Форма зубцов модифицируется.
Сама прыгательная вилка
располагается на IV сегменте и состоит
из непарного основания- манубрий, от которого отходят парные образования- денс и закачивается небольшим
когтевидным придатком- мукро. Иногда эти
части могут сливаться частично или полностью. Если вилка в расправленном состоянии, то можно отличить
дорсальную и вентральную поверхность
(задняя и передняя). Длина вилки варьирует у разных таксонов. Например, у
Campylothorax Iongicornis в
подогнутом состоянии она может достигать рта,
либо прыгательная вилка может полностью быть редуцирована вместе с ней редуцируется и зацепка, это
происходит у почвенных форм.
Денс обычно имеет хитиновые утолщения
или ребра. Они служащие для сцепления с зубцами на зацепке. У некоторых таксонах
на денс возникают шипы или шиповидные хеты (Isotomidae, Neelidae, Poduridae). На дорсальной поверхности денс часто несет поперечные складки (Entomobryidae).
Мукро может быть очень разнообразным по своей форме, может нести зубцы и ламеллы, реже шипы и хеты.
На V сегменте брюшка располагаются
генитальное отверстие. У коллембол
есть половой диморфизм, который проявляется в
положении половый щели: у самцов- продольная, а у самок- поперечное. Так
же половой диморфизм может
проявится и в пигментации. У самцов шипики
расположены на разных участках тела, голове или усиках. Надо отметить
что самки больше самцов. Наиболее
четко половой диморфизм проявляется у слитобрюхих чем у членистобрюхих видов.
На VI сегменте- анальное отверстие,
открывается терминально или
вентрально, трехлучевое. Оно ограниченно непарной дорсальной и двумя парными вентролатеральными
лопастями, на которых расположены обычные
хеты. У самок большинства слитобрюхих форм по бокам
анального отверстия
расположены парные анальные придатки. Их формы разнообразны- от щетинковидных до лопаточковидных.
На VI, реже на V сегменте могут находится анальные шипы [2].
1.11 Жизненные формы
У представителей различных таксонов коллембол хорошо выделяются черты конвергентного сходства,
связанные с обитанием определенных ярусов
биоценоза.
По систематики Г. Гизина [32] и ряда других авторов коллемболы делятся на три больших ярусных
группы, в соответствии с внешними
особенностями строения, адаптирующими животных к месту обитанию:
· Атмобиос (поверхностные)- животные с напочвенным местом обитания.
· Гемиэдафон (подстилочные)-обитают в полупочве или в подтилке.
· Эуэдафон (почвенные) - почвенное формы, а если быть точнее в минеральных горизонтах почвы [2].
После С.К. Стебаева эту классификацию дополнила и расширила, введя в нее
ряд количественных критериев для более дробных подразделений. Что предлагает выделят четыре группы жизненных форм:
· Поверхностные или верхнеподстилочные формы. У них полный набор глаз (8+8), сильно развита
пигментация, а также конечности и
прыгательная вилка. В себя включает четыре жизненные формы:
1. Антобионты- обитатели макрохитов, поверхность рыхлой подстилки. Животные, относящиеся к
этой форме, совершают суточные
миграции в травостой, на деревья и кустарники. Характеризуются яркой окраской, длинными усиками, ногами
и прыгательной вилкой.
так же у них крупные размеры
(1,8-6,0 мм), простое строение антеннального
или отсутствием
постантеннального органа, усложнение коготков ног, покровы с эпикутикулой или защитным слоем из
макрохет и чешуек. (Orchesella, Entomobrya,
sminthurus и др.)
2. Верхнеподстилочные формы- обитатели верхних слоев подстилки. Имеют средние размеры тела,
умеренно длинные усики и ноги, полный набор
глаз. (Hypogastrura,
Pseudachorutes,
Brachystomella и
др.) Сюда следует
отнести и очень крупных Onychiuridae с прыгательной вилкой, хотя у них нет глаз;
обитателей моховых субстратов с максимальным для рода количеством глазков (5+5) и без развитой вилки Morulina
3. Нейстонные формы- обитатель водных поверхностей или переувлажненных грунтов. Из-за своего
места обитания они приобрели
специфические черты: мукро сильно расширен и снабжен широкими веслообразными ламеллами или многочисленными зубцами,
коготки ног удлинённые, с расширенным
эмподием и часто длинной
субапикальнойнитью- филаментом. (Podura aquatica,
некоторые виды рода Isotomurus, Sminthurides и
др.)
4. «Кортицикольные» формы- обитатели лишайников и мхов на скалах и стволах деревьев, сухих
подстилок. У них интенсивная пигментация,
уплотненная кутикула с ячеистым орнаментом. У большинства имеются многочисленные головные волоски на
тибиотарзусе, эмподий часто
редуцирован (ряд видов родов Pseudachorutes, Vertagopus,
Xenylla и др.) Часть имеет меньше глазков, то есть меньше
8+8 и могут не иметь прыгательную
вилку, но это следствие филогенетических ограничений.
· Гемиэдафические (полупочвенные) формы. Характеризуются частичная редукцией глаз, пигмента, прыгательной
вилки. Среди них встречаются:
1. Нижнеподстилочные формы- адаптированы к обитанию в толще мхов, в ферментативном слое
подстилки. У этой формы наблюдается первая
стадия редукции глаз (от 6+6 до 4+4), ослабленная пигментация, Вилка укорочена или редуцирована. (Tomocerus, часть Xenylla,
Microgatrura и др.)
2. Подстилочно- почвенные формы- со второй
стадией редукции глаз (от 3+3 до 1+1), слабая пигментация
покровов, вилка короткая или
отсутствует. (Arrhopalites,
часть Folsomia, Neanura, Anurida и др.)
· Эуэдафические (почвенные) формы-
у них есть черты сходства,
то есть полная редукция
глаз, пигмента и части прыгательной вилки. Характерно
дальнейшее уменьшение относительного размера придатков, то есть усиков и ног, с шириной тела. Они разделяется на:
1. Верхнепочвенные формы- населяют
верхние слои почвы,
а в глубь проникают по крупным ходам и трещинам. Относительно крупные эуэдафические коллемболы (более 1
мм). Суда относят большинство Onychiurinae,
некоторые Tullbergiinae,
а также крупные
безглазые Cryptopygus, Folsomia и
Anurida.
2. Глубокопочвенные формы- проникают в глубь по тонким почвенным капиллярам. Объединяют
мелких представителей тех же и
некоторых других родов (менее 1мм).
· Специализированные обитатели пещер (троглобионты), муравейников и термитников (синэкоморфы)- полная
редукция глаз и пигмента, но
сохраняют ряд черт поверхностных форм, то есть
развитые конечности и прыгательная вилка. (Многие Plutimurus, Sinella и др.)
[14], [2] (Рис.6, 7).

Рис.6 ( по Стебаевой, 1970)
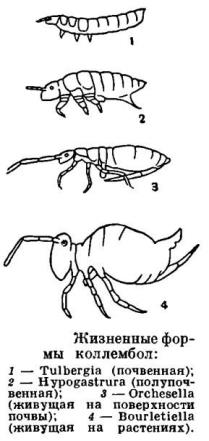
Рис. 7 (из Жизни животных, 1967)
1.12 Адаптации к основным факторам среды
1) Влажность- кожное дыхание обуславливает, высокую чувствительность к влажности окружающего воздуха [34].
Но особенности строения покровов.
Эти физиологические и поведенческие адаптации
позволяют приспособится к местообитанию с разными гидротермическими режимами. Влажность среды определяет успех
линьки и яйцекладки. В период линьки
особи очень чувствительны к обезвоживанию, а яйца и вовсе могут погибнуть от обезвоживания.
Адаптации:
· Кутикулярное сопротивление испарению- дефицит насыщения воздуха, атмосферной влагой
умноженное на скорость потери воды)
· Чешуйчатый покров- частичное его нарушение приводит к снижению выживаемости при малой влажности.
· Ангидробиоз- коллемболы могут находятся в высушенном состоянии несколько месяцев, а
при благоприятных условиях вновь
восстанавливают свою жизнедеятельность.
· Сезонное изменение гидротермического режима местообитания, то есть взывает экоморфоз-
морфологические изменения в кутикуле и
кишечнике. Сильное увеличение жирового тела до 70%, возрастание в клетках
концентрации уратов и протеинов. В это время животные уходят
в глубь почвы у там голодают.
· У некоторых видов описано голодание во время засухи из-за того, что в пустые кишечники
испарение воды из тела меньше,
после наступает тепловое
оцепенение. Некоторые
таксоны (Hypogasrturidae) способны
находится в таком состоянии год.
· Поведенчиская реакция на возрастание сухости воздуха. В такой ситуации виды начинается усиленная
активность особей, происходит
перераспределение особей в пространстве и усиливается их агрегированность.
(Распределение одного и того же вида в подстилке сильно меняется в зависимости от ее влажности)
· Вертикальная миграция, так же определяется условиями влажности. Особи уходят в глубь из пересыхающих сверху
субстратов. Это зависит от размера самих коллембол и от размера
скважин. (У почвенных видов- вертикальная миграция, а у напочвенных- горизонтальная.)
· Могут выходить на стволы деревьев и травостой при высокой влажности (например, после дождя)
при этом повышается ночная активность.
2) Температура-как у всех пойкилотермных организмов, скорость развития сильно зависит от поступления тепла в организм
из вне. Это сопровождается повышение
температуры укорачивает межлиночный период
при этом увеличивается смертность.
Адаптации:
· Основные физиологический механизм переохлаждения - это накопление криопротекторов (глицерин,
маннит, глюкоза, ксилоза, трегалоза,
фруктоза) [34]. Так как образование льда в тканях
смертельно для любого организма.
· Поведенческие адаптации для избежание замерзания- быстрое опорожнение кишечника вслед за
резким снижением температуры. У
кормящихся особей устойчивость к холоду ниже. Ювенильны особи более устойчивы к понижению температуры
чем взрослые.
· Холодоустойчивость особей меняется в течении года. Резистентность возрастает под
влиянием сокращающейся длины дня.
· Особи, которые активны под снегом или выходящие на него имеют темную окраску, следовательно,
появляются только днем, и используют
тепло солнечной радиации.
3) Свет- отношение коллембол к свету определяется принадлежностью к той или иной жизненной форме. У атмобионтных (поверхностных) ногохвосток пигментация и
развитие глаз свидетельствуют о значительной
роли светового фактора в их жизни. У почвенных форм преобладает отрицательный фототаксис. Тогда как пещерные виды не проявляют фотопреферендума.
4) Газовый состав воздуха и химические свойства среды
обитания- в местах
обитания коллембол бывает
возможно временное создание
анаэробных условий, к чему многие виды хорошо адаптированными.
Доказано что виды обитающие в гниющей растительности и навозе устойчивы к некоторым токсичным
газам. Так же рад видов может обитать в
условиях литорали или на сильно засоленных почвах, следовательно, испытывают постоянное влияние
солевых растворов. При этом осморегуляция поддерживается разными
способами, например, концентрация гемолимфы
меняется вслед за изменением солености воды- Archisotoma pulchilla [42].
5) Кислотность (pH) среды- считается что кислотность слабо
влияет на распределение коллембол и
их жизнеспособность, так как они не
подвергаются прямому воздействию щелочных и кислотных растворов, которые находятся в почве. Из-за того, что они
обладают воздушным дыханием и
гидрофобной кутикулой. Но есть данные которые говорят о том, что при низкой кислотности снижается плодовитость у самок [35].
По мнению Хэгвара,
связь с pH почвы у коллембол косвенная. Автор считает более вероятной
гипотезой- это изменение
конкурентной способности видов
при сдвигах реакции среды. Коллемболы- слабые
индикаторы непосредственных значений pH, но население их быстро реагирует на интегральные изменения микробиологической обстановки в лесной подстилке, вызываемые кислыми
дождями [2].
1.13 Структура и динамика популяции
Возрастной состав коллембол определяется по размеру тела или по отдельным его частям, так как рост не
прекращается на протяжении всей жизни,
но на поздних этапах он замедляется. Так же возраст определяется по морфологии генитального поля.
Размножение коллембол, связанно с растянутыми сроками откладки яиц, это является основой
разновозрастной популяции. Синхронизация редкое явление и определяется внешними условиями среды.
У большинства ногохвосток жизненный цикл не фиксирован в годовом периоде, то есть лон изменчивый.
Лишь у некоторый видов, есть механизмы,
которые регулируют время появление отдельных генераций. Например, у Sphaeridia зимуют лишь яйца, весной появляется
молодь, которая быстро растет. Летом
в популяция по большой части состоит из яиц.
Разновозрастность популяции в конце весны
и осенью [23]. У некоторых видов без диапаузы в отдельные временные отрезки могут быть представлены одной возрастной группой.
Соотношение потомства и взрослого населения есть экологическая характеристика вида, которая отражает масштабы
вымирания до взрослого состояния. Следовательно, у них возрастная стабильности популяции выглядит в виде пирамиды с
широким основанием из-за высокой смертности
в природе. Коллемболы являются «r-стратегами»- постоянное воспроизводство.
В динамики большинства видов есть определенная цикличность. По
приуроченности подъёма численности популяций к определенным сезонам года авторы выделяют фенологические
группы видов. Например, в ельниках
подмосковской области выделяют: весенне-летние, летние, летне-осенняя, осенне-зимние [9]. Сезонная приуроченность численности
может сдвигается от влияния погодных
условий [2]
1.14 Население коллембол
Группировки формируются по принципу закона Тинемана: чем экстремальнее условия биотопа тем
беднее на них видовой набор и тем выше
может быть численность отдельного вида.
Как правило видовой состав беден в условиях критических температур, переувлажнение, аэробность
среды и т.д. [20], а также на скалах с
экогенетическими сукцессиями, на днищах высохших озер [15] и т.д. В Мезофитных условиях с толстым слоем
подстилки или моховой дернины
появляется десятки видов ногохвосток [2]
1.15 Черноольшаник
Черноольшаник -это своеобразные растительные сообщества, образующиеся в поймах малых рек, оврагах и ложбинах
стока, характерные для территории
средней полосы России. Особенность этих ассоциаций
определяется динамическим режимом увлажнения и средообразующей ролью ольхи черной (Álnus glutinósa),
которая вступает в сембиотические отношения с азотфиксирующими бактериями [17].
Ольха черная- быстрорастущее листопадное дерево семейства березовых, высотой до 30 м, с темно-бурой корой.
Распространено в Европе, Западной
Сибири, на Кавказе, в Малой Азии и Северной Африке. Растут обычно во влажных местообитаниях и на низинных болотах,
оврагов, поймах рек и т.д. Ольха
черная живет до 80 - 100 лет.
Наиболее распространенными типами черноольшаников являются черноольшаник таволговый, крапивный, снытевый и
кисличный. При увеличении
обводненности почв и усилении застойности вод
развиваются черноольшаники осоковые, болотно-папоротниковые и ивняковые.
Черноольшаник таволговый располагается на плоских или с незначительным уклоном понижения, поймы
рек и ручьев. Почвы обычно торфянисто- и торфяно-глеевые,
торфяные маломощные и среднемощные, среднеобводненные,
слабопроточные. Мертвопокровный черноольшаник
располагается наоборот на склонах пойм рек.
Глава 2.
Материалы и методика
Методика состоит из нескольких стандартных этапов:
2.1 Выбрать район исследования (район выбирался научными руководителями.)
В моем случаи выбор пал на черноольшаники (Рис. 8) Материалом послужили результаты
исследований в ассоциации черноольшаника
мертвопокровного и таволгового на территории Новой
Москвы на Малинском биогеоценологическом стационаре
ИПЭЭ РАН с 2000 по 2009 г.

Рис.8 Пойменный черноольшаник
2.2
Полевые учеты
(Материал был представлен Боковой А.И.)
Заключается в том, чтобы пробы содержали в себе примерно одинаковый процент субстрата
(подстилка, почва, мох и т.д.) Почвенные
пробы брали при помощи бура (Рис. 9) на всей толщине подстилки, в среднем она равняется 5-10 см. Бур удобен для изучения вертикального распределения микроатропод.
Пробы обычно берутся до глубины«необитаемого слоя».
В каждой ассоциации за один срок учета отбирали по 10 проб. Всего было
взято 10 серий (с 2000 до 2009 г.). Одну большую или одну маленькую пробу нельзя брать, так как любая оценка численности строится на статистическом
методе. Но в этих случаях можно получить
некоторые представления о фауну сообществ.
Схема взятия проб:
1. Биотопы должны быть более или менее выравненный микрорельеф и растительный
рельеф, но есть и случайный выбор проб
2. Пробы на биотопах с выраженной гетерогенностью среды должны браться микростациями, то есть
в определенной последовательности и
определенном месте. Например, в лесу сначала
проба берется у подножья дерева,
после в середине
проекции кроны древа, следом в
окне межу деревьями и т.д.
Так же надо отметить чем больше проб в выборке, тем точнее будут результаты анализа.
Сроки учетов приходились на летние месяцы (июль - август). Всего было взято 200 проб из которых извлечено 3541 экземпляр коллембол.
Существуют и другие
методы: ловушки Барбера,
кошение сачком, флокация
песка, эксгаустерный сбор [14].

Рис. 9 Пробоотборники почвы.
2.3
Техника
экстракции (Экстрагирование проводилось
научными руководителями.)
Коллембол экстрагировали по стандартной методике при помощи эклекторов Тулльгрена [3] (Рис
10). Воронка Тулльгрена состоит из
картонного эклектора, сита с диаметром ячеик около 1,5-2 мм, а к узкой части воронки крепится пробирки с
70% этиловом спирте.
Продолжительность выгонки определялась временем высыхания субстрата, но во всех случаях
это составляла не менее 3 суток. Так же
можно использовать лампу для более быстрого высыхания почвы.
Принцип работы:
1. На сито в воронке кладется проба и ставится сушится.
2. По мере высыхания почвы все животные стремятся к влажному месту обитания, следовательно,
они спускаются глубже и в итоге
проваливаются в воронки. Где скатываются по краю и попадают в пробирку со спиртом.
3. В пробирку поместить листик с номером пробы, названием места где собрали и числом сбора [13]
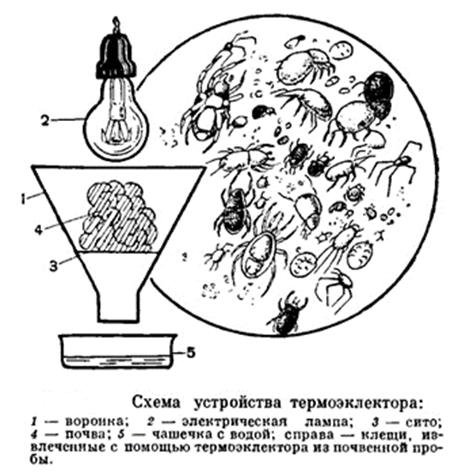
Рис. 10 Воронка Тульгрена ()
2.4 Камеральная обработка
После экстрагированные пробы фильтровались на разлинованной ручкой фильтре с помощью дистиллированной водой.
Сама разборка экстрагированного
материала проводили на этих фильтрах в чашке Петри под бинокулярным микроскопом
при 8-12-кратное увеличение.
Коллембол вылавливали при помощи препаровальной иглы вслед за этим животных помещали на стекло с
каплей жидкости Фора, покрывались покровным стеклом. На одном препаратном стекле помещалось три пробы. После готовый
препарат надо подписать: время сбора,
место сбора, так же над каждой пробой написать ее номер. После уже полностью готовые препараты помещались на планшет и ставились в термостат с температурой
примерно 45 °C (Рис. 11, 12).

Рис. 11 Разбор материала
Рис. 12 Коллембола под бинокуляром.
2.5 Таксономический анализ (материал определен Боковой А.И. и
Кузнецовой Н.А.)
Весь собранный материал определяли под микроскопом при разных увеличениях. Для таксономического анализа
использовали ряд определителей: Palissa, 1964; Fiellberg, 1980; Определитель фауны коллембол России и сопредельных
стран, 1994 [41], [29], [1]; и другие
ключи. Все определенные виды вносилась в таблицу по типу вид-проба с использованием
методом «конвертов». Всего в материалах представлено 63 вида ногохвосток.
2.6 Математическая обработка данных
Всю статистическую обработку данных проводили при помощи разных формул в программе Exel.
· Таблица вид- проба включает в себя данные по одной серии проб. В ней представлено количество экземпляров вида.
Следовательно, в заголовке должно отмечаться: географическое положение,
дата сбора, название биотопа. В этой таблице также
проводились расчеты:
1. Общего количества по видам
(x).
2. Общего количества всех видов
(y).
3. Число проб в одной серии (n)= 10
4. Доля вида (w):
5. Процент вида (q):
6. Среднее арифметическое (M) -это количество экземпляров в пробе (или численность на кв. м):
7. Среднее квадратное отклонение (SD)- основной показатель
того, насколько данные отклоняются от среднего
значения:
8. Ошибка средней (m)-показатель того, насколько полученная
средняя может быть далека от реальной средней в природе:
9. Среднее квадратичное отклонение:
10 .Площадь пробы в м2(S):
𝑆 = 8
∗𝑛
11 .Общая численность экз. на
м2:
12 .Стандартный коэффициент вариации (V)-мера относительного
разброса случайной величины; показывает, какую долю среднего
значения этой величины составляет её
средний разброс. Надо отметить что SD-среднее квадратичное отклонение, а M-средняя арифметическая [14, стр. 44, 47-49]:
Иногда предлагается условная классификация вариабельности
выборки на основе коэффициента
вариации: при V≤10% выборка вариабельна слабо, при 10≤V≤20% - средне, при V≥20%- сильно.
13 .Коэффициент Жаккара (𝐼𝑗 )- показывает долю общих видов
в объединённом списке. A-
число видов в первом биотопе, B-число
видов во втором биотопе, С- число общих видов,
отмеченные на первом и втором биотопе:
.7
Жизненные
формы
При определении жизненных форм принята система С.К. Стебаевой [16]. Выделяли три основных класса: поверхностно- обитающие, полупочвенные и
почвенные. (см. литобзор жизненные
формы)
2.8 Пороги доминирования
За основу выделения классов доминирования принята шкала Энгельмана [28]: 0-1,3%-редкий; 1,31-3,9% малочисленный, 3,91-12,4%
субдоминант, 12,4-39,3% - доминант, 39,4-100% - эудоминант. Обилие вида выражали в % от общей
численности [14].
2.9 Список использованных характеристик
- общая численность коллембол для удобства сравнения экстраполировали на 1 м2
- общее число видов в биотопе (по данным нескольких серий учетов)
- видовое разнообразие - индекс Шеннона
- относительное обилие (в % от общей численности):
доминирующих и субдоминирующих видов, жизненных форм.
- общая численность экз.\пробу
Глава
3. Результаты
.1
Температура
и влажность
Максимальная температура была зафиксирована в 2003 г.-28 °C, а
минимальная в 2007 г.-17 °C.
Средняя температура по всем годам
равняется 22 °C. Давление в летние время колебалось
в пределах от 742 до 755 мм. рт. ст. Погода
в эти года была малооблачна, кроме 2001-2002, 2004
и 2009 г.- пасмурная. Дождь был в 2001 г. и 2007 г. Максимальная влажность в 2002 г.- 87,75
%, а минимальная в 2008 г.-39,75. Средняя влажность по всем годам составляла
79,84%. Но влажность и температура
может немного варьировать так как черноольшаника
произрастает на влажных территориях, следовательно, тут % влажности будет выше, а температура ниже
(Таблица 1).
Таблица 1 Метеоданные по московской области
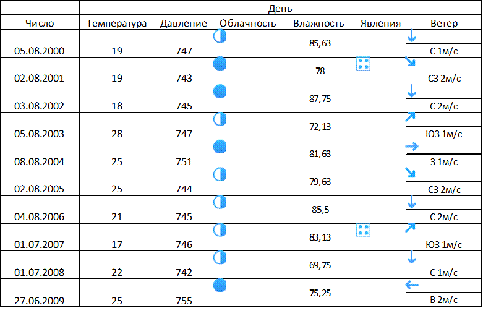
 -дождь,
-дождь,  -ясно,
-ясно,  -малооблачно,
-малооблачно,  -облачно,
-облачно,  -пасмурно
-пасмурно
3.2 Видовой состав
Общий видовой состав
в двух биотопах Московской области
равен 58 видам (Таблица 2).
Таблица 2 Видовой состав сообществ.
|
Семейство, вид
|
Мертвопокровный
биотоп
|
Таволговый
биотоп
|
|
Hypogastruridae
|
|
|
|
Willemia anophthalma
|
|
+
|
|
Willemia denisi
|
+
|
|
|
Sсhoetella ununguiculata
|
|
+
|
|
Neanuridae
|
|
|
|
Micranurida pygmea
|
+
|
+
|
|
Pseudachorutes sp.
|
+
|
+
|
|
Friesea truncata
|
+
|
+
|
|
Neanura muscorum
|
+
|
+
|
|
Onychiuridae
|
|
|
|
Mesaphorura sp.
|
+
|
+
|
|
Protaphorura armata
|
+
|
|
|
Protaphorura sp.
|
+
|
+
|
|
Micraphorura absoloni
|
+
|
+
|
|
Stenaphorura sp.
|
+
|
|
|
Stenaphorura
quadrispina
|
|
+
|
|
Neotullbergia
crassicuspis
|
+
|
|
|
Hymenaphorura polonica
|
+
|
|
|
Isotomidae
|
|
|
|
Vertogopus cinereus
|
|
+
|
|
Isotoma viridis
|
+
|
+
|
|
Desoria olivacea
|
|
+
|
|
Desoria hiemalis
|
+
|
|
|
Desoria sp.
|
+
|
+
|
|
Desoria blufusata
|
+
|
+
|
|
Isotomiella minor
|
+
|
+
|
|
Paraisotoma notabilis
|
+
|
+
|
|
Folsomia candida
|
+
|
|
|
Folsomia fimetaria
|
+
|
|
|
Folsomia quadrioculata
|
+
|
+
|
|
Isotomurus fucicolus
|
+
|
+
|
|
Entomobryidae
|
|
|
|
Willowsia nigmaculata
|
|
+
|
|
Heteromurus nitidus
|
+
|
+
|
+
|
|
Pseudosinella sp.
|
+
|
|
|
Entomobrya corticalis
|
+
|
|
|
Entomobrya longinosa
|
|
+
|
|
Entomobrya marginata
|
+
|
|
|
Entomobrya nivalis
|
+
|
+
|
|
Entomobrya sp.
|
|
+
|
|
Lepidocyrtus lignorum
|
+
|
+
|
|
Lepidocyrtus violaceus
|
+
|
|
|
Orchesella bifasciata
|
|
+
|
|
Orchesella flavescens
|
+
|
+
|
|
Wiellowsia buski
|
+
|
|
|
Tomoceridae
|
|
|
|
Tomocerus vulgaris
|
+
|
+
|
|
Pogonognathellus flavescens
|
+
|
+
|
|
Katiannidae
|
|
|
|
Sminthurides schoetti
|
+
|
+
|
|
Sminthurides sp.
|
+
|
+
|
|
Sminthurididae
|
|
|
|
Sphaeridia pumilis
|
+
|
+
|
|
Caprainea marginata
|
+
|
+
|
|
Allacma fusca
|
+
|
|
|
Sminthurinus niger
|
+
|
+
|
|
Sminthurinus
sp. 1
|
+
|
+
|
|
Sminthurinus aureus
|
|
+
|
|
Sminthurinus
sp. 2
|
|
+
|
|
Arrhopalitidae
|
|
|
|
Arrhopalites gr.caecus
|
+
|
|
|
Dicyrtomidae
|
|
|
|
Ptenothrix atra
|
|
+
|
|
Dicyrtoma fusca
|
+
|
+
|
|
Dicyrtoma minuta
|
+
|
|
|
Neellidae
|
|
|
|
Neelides murinus
|
+
|
|
|
Megalothorax minimus
|
+
|
+
|
|
Прочие
|
|
+
|
|
Итого
|
46
|
35
|
В мертвопокровном черноольшанике- 46 видов во все сроки учета, а в таволговом- 35 вида. Это может быть связанно с тем, что в
черноольшанике мертвопокровном больше «питательных веществ» или были оптимальные условия для проживания
сообществ. «Питательные вещества»-
это поступающие в почву опавшая листва, которая состоит из водорастворимых органических компонентов (сахара, фенолы, углеводороды, глицериды
и пр.), целлюлозы, лигнина и ряда других компонентов. Доля этих соединений в
растительном субстрате может сильно
варьировать, в зависимости от органа и вида растения [22]. Растительные полисахариды являются основным источником
энергии в остатках растений, их
содержание в опаде составляет примерно 20-30%
[5]. Отмирающие части древесных растений можно разделить на листовой, корневой и древесный опад. Поступление в почву листового
и корневого опада неравномерно во времени и климатических условиях, то есть большая часть опада
приходится на осень, что особенно четко
выражено для лиственных деревьев.
При сравнении видового состава по всем годам в отдельности выявляется некая закономерность в чередовании повешения количества видов в определённый год на
одном биотопе и понижение в тот же год на другом
биотопе. (Например, мертвопокровный в
2001 было обнаружено 22 вида, а
таволговом 15 видов) (Таблица 3, Рис.13).
Таблица 3 Число видов коллембол.
|
Год
|
число видов в
ч. Мертвопок ровном
|
число видов в
ч. Таволгов ом
|
|
2000
|
15
|
13
|
|
2001
|
22
|
15
|
|
2002
|
18
|
16
|
|
2003
|
13
|
21
|
|
2004
|
20
|
16
|
|
2005
|
22
|
10
|
|
2006
|
10
|
10
|
|
2007
|
12
|
16
|
|
2008
|
19
|
9
|
|
2009
|
20
|
11
|
|
Общее кол-в
|
о 171
|
137
|
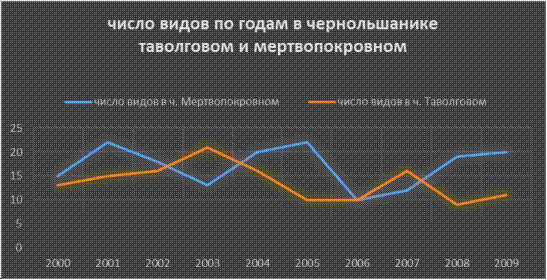
Рис.13
В мертвопокровном черноольшанике в 2000г. и 2005г. было зафиксировано максимальное количество видов- 22 вида, в 2007г. минимальное 12 видов. В таволговом,
максимальное было в 2003г.- 21 вид, а минимальное 2008г.-
9 видов. В 2006 было одинаковое количество видов- 10, это может быть
связанно с тем что в этом году не быто
серьезных отличай в температурном режиме,
световом, влажности и т.д.
(то есть температура с 1 по 4 августа повышалась на 1 градус, а влажность повышалась на 10%). В этот год
так же идет совпадение по жизненным
формам у поверхностных (20%) и подстилочных (40%)
видов.
Поле анализа данные по влажность, температуре и числу видов, я заметила, что изменения количество видов
меняется вместе с климатическими
условиями. Например, при максимальной влажности
87,75% на мертвопокровном участке количество видов уменьшилось с 22 до 18, а в таволговом осталось
примерно таким же 15 и 16. При максимальной температуре 28 в
мертвопокровном произошла понижение с
18 до 13, а на таволговом наоборот повышение с 16 до 21. Следовательно виды на мертвопокровном участке
больше зависят от влажности так как мертвопокровный
черноольшаник располагается на склонах
(избыток воды уходит к подножью), а таволговые виды от температуры, так как занимает пониженные и западинные участки
пойм рек (вода тут остается и следовательно постоянная влажная почва, но при высоких температурах кочки (на
которых брались пробы) подсыхают
следовательно более благоприятные условия)
Но не смотря на то что на одном биотопе 47 видов, а на втором 44 нельзя говорить об их различии в
видовом составе. Это подтверждает и
коэффициент Жаккара который равен 0,44. Исходя из этого два биотопа похожи друг на друг примерно на
половину. Коэффициент изменяется от 0
(общих видов нет вообще) до 1 (списки видов одинаковы, но это не означает что сообщества одинаковы,
так как обилие видов может
отличатся).
3.3 Динамика общей численности(экз. на м𝟐)
Теперь разберемся с общей численностью (экз. на м2). В
черноольшанике мертвопокровном обнаружилась закономерность:спады сопровождаются
повышением плотности сообществ, но с 2003-
2004 были спады, а с 2006 по 2008г.г. повышение. Максимальный пик зафиксирован
в 2002г. примерно 47 экз. на м2, а в 2006г. было минимальное обилие 17экз. на м2 (Таблица 4, Рис. 14).
Таблица 4 Динамика общей
численности коллембол в мертвопокровной
ассоциации.
|
Мертвопокровный
|
|
Год
|
общая численность (экз. на м2)
|
Ошибка средней (m)
|
|
2000
|
44
|
18,55
|
|
2001
|
31
|
31,62
|
|
2002
|
47
|
31,45
|
|
2003
|
33
|
8,44
|
|
2004
|
27
|
25,47
|
|
2005
|
45
|
45,54
|
|
2006
|
17
|
0
|
|
2007
|
22
|
3,71
|
|
2008
|
26
|
19,35
|
|
2009
|
18
|
15,39
|
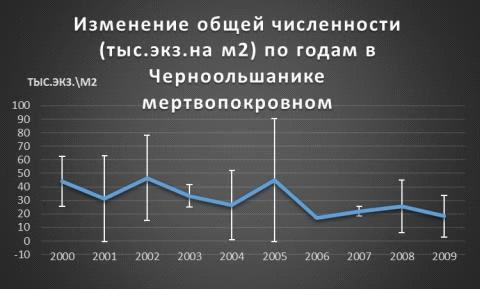
Рис. 14
В таволговом черноольшанике такой резкой закономерности не нашлось: в отличии от мертвопокровного
где все идет скачкообразно, тут идут
плавные подъёмы и спады плотности. Максимальное численность была обнаружена в
2002г.- 37 экз. на м2 точно
также как и в мертвопокровном, и
минимальное в 2005г.- 4 экз. на м2. Ошибка
средней показывает нам насколько полученные результаты могут быть далеки от реальной средней в природе
(Таблица 5, Рис. 15).
Таблица
5 Динамика общей численности коллембол в
таволговой ассоциации.
|
Таволговый
|
|
Год
|
общая численность (экз. на м2)
|
Ошибка средней (m)
|
|
2000
|
14
|
10,95
|
|
2001
|
11
|
4,53
|
|
2002
|
37
|
18,85
|
|
2003
|
36
|
32,93
|
|
2004
|
8
|
3,6
|
|
2005
|
4
|
0,32
|
|
2006
|
5
|
0
|
|
2007
|
17
|
8,73
|
|
2008
|
10
|
0,82
|
|
2009
|
10
|
0,78
|
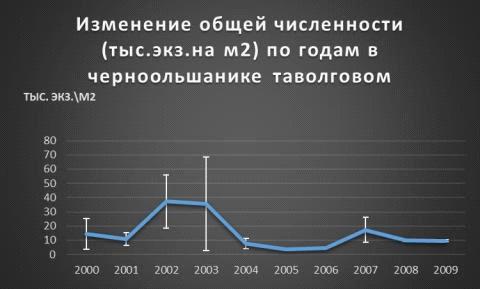
Рис. 15
3.3 Жизненные формы
Теперь рассмотрим динамику жизненных форм. Просматривая графики можно заметить, пространственную организацию населения коллембол, которая
характеризуется выраженной ярусностью данных
групп, то есть приуроченность к разным слоям почвы (поверхностные, подстилочные, почвенные) хотя соотношение и
менялось по годам.
Максимальный процент поверхностных форм в ч. мертвопокровном замечен в 2005г.-54,5%, а минимальный в 2002г.-
27,8%. А вот с подстилочными формами
все с точность наоборот, максимум в 2002г.-
44,4%, минимум в 2005г.- 22,7%. Почвенные достигли высоты сразу в двух годах
2004 и 2009г.г.- 35%, а минимум в 2005г.- 22,7%.
В 2006г. процентное
соотношение жизненных форм было одинаково- 33,3%
(Таблица 6, Рис. 16).
Таблица
6 Жизненные формы в мертвопокровной ассоциации.
|
спектр жизненных форм в Ч.
Мертвопокровном (%)
|
|
|
2000
|
2001
|
2002
|
2003
|
2004
|
2005
|
2006
|
2007
|
2008
|
2009
|
Общий процент
|
|
поверхно стные
|
40,0
|
45,5
|
27,8
|
38,5
|
35,0
|
54,5
|
20,0
|
33,3
|
31,6
|
30,0
|
356,2
|
|
подстило чные
|
33,3
|
27,3
|
44,4
|
30,8
|
30,0
|
22,7
|
40,0
|
33,3
|
36,8
|
30,0
|
328,7
|
|
почвенные
|
26,7
|
27,3
|
27,8
|
30,8
|
35,0
|
22,7
|
30,0
|
33,3
|
31,6
|
35,0
|
300,1
|
|
ПРОЧИЕ
|
|
|
|
|
|
|
10,0
|
|
|
5,0
|
15
|
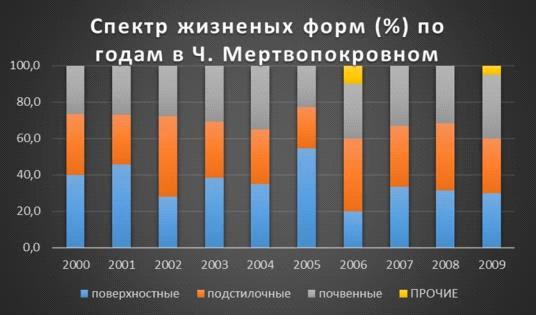
Рис. 16
Теперь так же разберем и таволговый
черноольшаник: поверхностные достигли своей высоты в 2000г.- 61,5%, а
нижняя точка была зафиксирована в 2008г.- 11,1%.
Подстилочные: максимум в 2008г.-
44,4% и минимум в 2009г.-
27,3%. Почвенные в отличии от ч.
мертвопокровного два раза вышли на нижней отметке
в 2002 и 2007 г.г.- 6,3%, а пик зафиксирован в 2008г.- 44,4% (Таблица 7, Рис. 17).
Таблица 7 Жизненные формы в таволговой
ассоциации.
|
спектр жизненных форм в Ч.
Таволговом (%)
|
|
|
2000
|
2001
|
2002
|
2003
|
2004
|
2005
|
2006
|
2007
|
2008
|
2009
|
Общий процент
|
|
поверхност ные
|
61,5
|
46,7
|
50,0
|
42,9
|
37,5
|
30,0
|
20,0
|
56,3
|
11,1
|
36,4
|
392,3
|
|
подстилочн ые
|
30,8
|
33,3
|
43,8
|
33,3
|
31,3
|
40,0
|
40,0
|
37,5
|
44,4
|
27,3
|
361,7
|
|
почвенные
|
7,7
|
20,0
|
6,3
|
23,8
|
12,5
|
30,0
|
20,0
|
6,3
|
44,4
|
36,4
|
207,3
|
|
ПРОЧИЕ
|
|
|
|
|
18,8
|
|
20,0
|
|
|
|
38,8
|
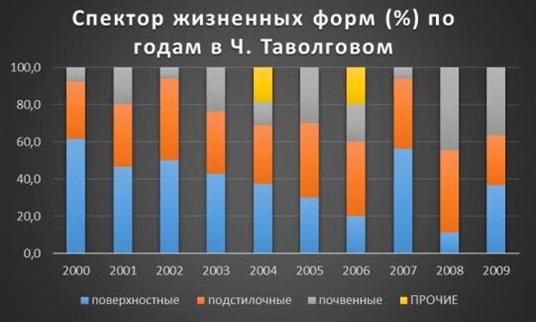
Рис. 17
Если их сравнивать вместе, то отчетливо видно, что поверхностных и подстилочных видов больше в ч. таволговом
чем в ч. мертвопокровном. А вот с
почвенными все наоборот в ч.
мертвопокровном их больше чем в таволговом.
К прочим отнесли виды, у которых не смогли определить жизненную форму или ювенильные особи.
3.4 Динамика структуры сообщества
Видовая структура представлена классами
доминирования выделены по Энгельману: редкие (субрецеденты) <1,3%; малочисленные (рецеденты) 1,3-
3,8%; субдоминанты 3,9-12,3%;
доминанты 12,4-39,3%; эудоминанты >39,4%. Я буду рассматривать доминантные виды. На разных участках были
обнаружены одинаковые доминантные
виды Lepidocyrtus lignorum, Folsomia quadrioculata,
Paraisotoma notabilis. Также
можно обратить внимание на то что в
таволговом черноольшайнике больше доминантных
видов.
Как правило, динамику численности коллембол определяют изменения в плотности доминирующих видов [29]. По доминантным таблицам и графикам видно в черноольшанике таволговом такой вид как
Isotomurus fucicolus появлялся толь в 2001 году. Теперь рассмотрим сменяемость доминантных видов по годам в тавоговом черноольшанике: 2000г. главным домининтом был Isotomiella minor-33,3%, в 2001 его сменил Folsomia quadrioculata-37,4%, в 2002-2003 он стал эудоминантом (41,6% и 40,8%). В 2004 он
вернулся к доминантам, но главнее оказался
вид Lepidocyrtus lignorum 29,6%. В 2005г. Paraisotoma notabilis- 30,8%.
В 2006 сразу из субдоминантов поднялся до эудоминанта вид Pogonognathellus flavescens -57,9%. В2007г. Lepidocyrtus lignorum 30,4%. В 2008 году он стал эудоминантом
42,3% и в 2009г. оказалось два
равноценных доминанта Folsomia quadrioculata и Paraisotoma notabilis- 20% (Таблица 8, Рис. 18). Следовательно,
на этом биотопе происходят переходы
от одного доминанта к другому скачкообразно.
Таблица
8 Структура сообщества коллембол в таволговой
ассоциации.
|
Классы доминирования
(Выделенны по Энгельману) по годам - черноольшайник таволговай
|
|
Год.
|
F.quadriocul ata
|
I.min or
|
S.pumilis
|
I.fucic olus
|
L.lignorum
|
P.notabili s
|
P.flavesc ens
|
Desoria
blufusata
|
Entomob ryidae
jvv.
|
|
2000
|
15,9
|
33,3
|
20,3
|
0
|
0
|
5,8
|
1,4
|
1,4
|
|
2001
|
37,4
|
20,9
|
4,7
|
16,3
|
5,8
|
5,8
|
3,5
|
2,3
|
0
|
|
2002
|
41,6
|
8,7
|
0
|
0
|
15,8
|
11,1
|
2
|
11,7
|
0,3
|
|
2003
|
40,8
|
9,9
|
0,7
|
0
|
4,6
|
17,3
|
1,1
|
2,5
|
0,4
|
|
2004
|
13
|
3,7
|
1,9
|
0
|
29,6
|
0
|
5,6
|
0
|
13
|
|
2005
|
19,2
|
3,8
|
0
|
0
|
7,7
|
30,8
|
11,5
|
7,7
|
0
|
|
2006
|
10,4
|
2,6
|
0
|
0
|
13,2
|
2,6
|
57,9
|
0
|
0
|
|
2007
|
3,6
|
4,3
|
2,2
|
0
|
30,4
|
8
|
2,9
|
13,8
|
1,4
|
|
2008
|
16,7
|
5,1
|
0
|
0
|
42,3
|
14,1
|
0
|
0
|
12,8
|
|
2009
|
20
|
10
|
17,1
|
0
|
0
|
20
|
0
|
11,4
|
0
|
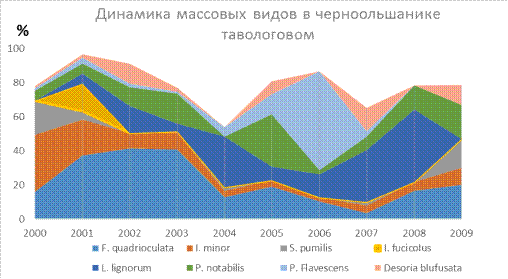
Рис. 18
В черноольшанике мертвопокровном на протяжении всей выборки
был один главный доминант Folsomia quadrioculata,
который в 2006 стал
эудоминантом 71,6% (Таблица 9, Рис. 19). Из это можно сделать вывод что в черноольшанике
мертвопокровном ядро доминантов (из 5
доминатов выделяется 2), следовательно, он стабильнее чем черноольшаник таволгового [9]. Стабильность также обуславливается синхронностью в изменениях
численности доминантных видов и в
регуляции колебаний плотности населения.
Таблица
9 Структура сообщества коллембол в
мертвопокровной ассоциации.
|
Классы доминирования
(Выделены по Энгельману) по годам - черноольшаник
мертвопокровный
|
|
Год.
|
F. quadrioculata
|
P. notabilis
|
L. lignorum
|
Protaphorura sp.
|
Mesaphorura sp.
|
|
2000
|
51,1
|
16,5
|
2,6
|
7,1
|
1,4
|
|
2001
|
26,4
|
17,6
|
6,8
|
13,2
|
2
|
|
2002
|
27,1
|
13,7
|
25,7
|
12,9
|
5,1
|
|
2003
|
33,3
|
10,5
|
10,9
|
19,1
|
9,4
|
|
2004
|
34,6
|
9,4
|
6,3
|
6,8
|
5,8
|
|
2005
|
35,3
|
28,1
|
5,6
|
6,7
|
2,5
|
|
2006
|
71,6
|
7,4
|
2,5
|
3,7
|
0
|
|
2007
|
33
|
6,8
|
19,3
|
0
|
5,1
|
|
2008
|
30,4
|
13,2
|
8,8
|
6,4
|
19,6
|
|
2009
|
32,9
|
11,6
|
8,2
|
4,8
|
8,9
|
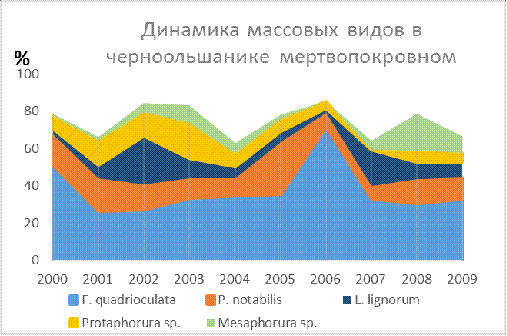
Рис. 19
Теперь рассмотрим максимальные и минимальные точки вида по всем годам.
Глава 4. Обсуждение
Как я уже говорила, динамика населения меняется с изменениями влажности и температурным режимом. Это подтверждает
работа [5]: показано, что динамика
населения ногохвосток в мезофитных лесах
обычно тесно связана с температурным режимом
почв.
Таким образом, в пойменном берёзово-осиновом лесу, формируются умеренно флуктуирующие сообщества коллембол, отличающиеся от стабильных по
амплитуде варьирования, прежде
всего, спектров доминантов структуру этого населения, во-первых, отличает расширенным набор потенциальны доминантов, а во-вторых более высокая доля редких
и малочисленных видов [18], тоже самое
наблюдается и таволговом черноольшанике, где редких и малочисленных видов по всем годам рана 19, а в мертвопокровном
6. В таволговом много потенциальных
доминантов которые в определенные
года выходят на этом уровне.
Вертикальное распределение ногохвосток связанно с погодными условиями. Похожая работа была написана в 1989 [11] - сезонная динамика и вертикальное
распределение ногохвосток в луговых почвах
тесно связанно с ходом погодных
условий, то есть ранней весной
и осенью были обнаружены пики численности и закономерные миграции коллембол во всех слоях почвы.
Также это подтвердилось в работе [11]-
гидротермический режим почвы влияет на вертикальное распределение коллембол, и на видовой состав ногохвосток.
Например, в 2007 г., когда был
дождь, влажность равнялась 83, а температура была 17, то вертикальное распределение отличалось от других 56% поверхностных видов, а почвенных 6%.
В это работе было показано что поверхностных и полупочвенных видов больше чем почвенных (поверхностные виды
составляют 31,6 %, полупочвенные
виды составляют 42,1 %, почвенные виды составляют
26,3 %.) Это закономерность замечена на моих биотопах (в мертвопокровном средний процент по всем годам приблизительно равен: поверхностные 35,6
%, полупочвенные 32,9 %, почвенные 30 %,
а таволговом: поверхностные 39,2 %, полупочвенные 36,17 %, почвенные 20,7 %.)
Высокие летние температуры на фоне недостатка влаги, как правило, ведут к снижению численности
ногохвосток в таёжных экосистемах [7]. В моей работе в 2008 г. при температуре равной 22 и влажность равнялась 70 заметное снижение
численности видов в черноольшанике таволговом (найдено 9 видов),
а также замечено
самый низкий процент поверхностных видов (11%), а подстилочных и почвенных форм где влажность больше,
одинаковый процент (44%).
Выводы
В ходе исследований на двух биотопах,
черноольшаник мертвопокровный и таволговой, было взято 200 проб, из которых выделено 3541 экземпляр коллембол. Всего в период
исследования было обнаружено
47 видов в мертвопокровном черноольшанике и 44
в таволговом, а общих
видов на этих двух участках
обнаружено- 28 видов. Из них большую часть составляют
поверхностные виды. На динамику
коллембол в большей степени влияет погодные условия т.е. количество осадков (влажность), температура воздуха, солнечный
свет, а также они могут реагировать
на pH среды) на конкретном биотопе. Для избежание не благоприятных условий
коллемболы могли мигрировать в
вертикальном или горизонтальном направлении.
Выяснилось, что в мертвопокровном черноольшанике динамика по годам происходит скачкообразно, в отличии от таволгового где обилие коллембол переходит более плавно.
По классам доминирования выяснилось, что доминантов больше в таволговом черноольшанике, но
устойчивый вид обнаружен в мертвопокровном- Folsomia quadrioculata.
В жизненных формах обнаружены все ярусы,
конечно соотношение их меняется
это как говорилось ранее связанно в погодные
условия.
Если сложить проценты по всем годам на отдельные жизненные формы (общий процент), то помучится что поверхностные и подстилочные виды в таволговом биотопе превышают мертвопокровный, а почвенные
наоборот. (см. таблицу 4-5). Но если
сравнивать по общему количеству всех видов (общее число видов), которые относятся к данной жизненной форме по годам то преимущество имеет мертвопокровный.
(см. таблицы 10-11) Но нельзя
говорить о их разницы в видовом составе так как он различаются не так сильно друг от друга (3 вида)
Благодарности:
Я хотел бы поблагодарить своего научного руководителя Бокову Анну Ивановну за ее советы и поддержку на протяжении
всей работа, а также Кузнецову Наталью
Александровну за его помощь в
экспериментальной работе и за ценные советы.
Список литературы
1. Бабенко А. Б. и др., Отв. ред. Н. М. Чернова. 1994. Определитель коллембол фауны России
и сопредельных стран: Семейство Hypogastruridae / Рос. АН, Ин-т эволюц. морфологии и экологии животных А. Н. Северцова, С.-5-335.
2. Бабенко А.Б., Кузнецова Н.А., Потапов М.Б., Стебаева С.К., Ханисламова Г.М., Чернова Н.М. 1988.
Определитель
коллембол фауны СССР. С.5-46.
3. Гиляров М. С. 1975. Методы почвенно- зоологических исследований. Наука. С. 5-281.
4. Гиляров М. С., Семенова Л. М. 1963. Эволюция кутикулы членистоногих\\ Успехи соврем. Биологии. Т. 56, вып. 5.
С. 208-227.
5. Кременица А. М. 2005. Сезонная динамика населения ногохвосток в пахотных
горизонтах под ячменем в структуре
десятипольного севооборота.
6. Кременица A.M. 2005. Структура комплекса ногохвосток (Collembola) чернозема обыкновенного нижнего
дона: Автореф. дис. канд. биол. наук.
-Ростов-на-Дону. - 24с.
7. Кузнецова Н. А. Многолетняя динамика популяций коллембол в лесной и производной
экосистемах // Зоол. журн. 2007. Т. 86, № 1.
8. Кузнецова, С.А., Александрова Н.Б., Б.Н. Кузнецов. 2001. Состав и превращения основных
компонентов автогидролизованной
древесины сосны, ели и осины.
9. Кузнецова H. А., Бабенко А. Б. 1984.Многолетняя динамика
численности коллембол в ельнике-зеленомошнике // Фауна и экология ногохвосток
10. Кутырева Л. Т., Бабушкин Н. М. 1989. Сезонноя динамика коллембол в почве
разнотравного-вейникового луга
11. Кузнецова Н.А. Организация сообществ почвообитающих коллембол. М.: ГНО «Прометей» МПГУ,
2005. - 244 с
12. Научная библиотека диссертаций и авторефератов disserCat #"896874.files/image026.jpg">