Везикулярный транспорт
Везикулярный транспорт
Введение
Везикулярный транспорт, или перемещение макромолекул в составе мембранных пузырьков
(везикул, англ. vesicles) между компартментами клетки, является одним из
базовых клеточных процессов.
Важность везикулярного транспорта
для нормальной жизнедеятельности эукариотической клетки, да и организма в
целом, подчеркивает тот факт, что целый ряд инфекционных агентов (бактерий и
вирусов) поступают в клетки с помощью везикулярного транспорта и обладают
способностью изменять работу отдельных элементов транспортных путей. Выявлен
целый ряд генетически наследуемых заболеваний, связанных с дефектами в белках -
регуляторах везикулярного транспорта, приводящих к дисфункции транспортных
путей, нарушению механизмов сортировки молекул или их правильной доставки.
1. Общие принципы
организации везикулярного транспорта
В общем случае под везикулярным
транспортом подразумевается перенос белков от одного внутриклеточного
компартмента к другому с помощью мембранных везикул.
Транспортные пузырьки
(везикулы) - представляют собой основное
средство передвижения белков и липидов внутри клетки. Это очень экономичный вид
транспорта: переносимые белки и липиды образуют мембрану транспортного
пузырька, а в полости, окруженной этой мембраной, могут находиться грузовые
молекулы, доставляемые к другим органеллам.
Основные принципы
формирования пузырьков:
· Вещества,
синтезируемые в ЭР, предназначены для компартментов, а не для цитозоля
· Вещества проходят в
клетку путём формирования и слияния пузырьков; этот процесс называется
эндоцитозом
· Вещества в полости
пузырьков часто имеют адресную метку, благодаря которой вещества сортируются и
концентрируются в определенных активных участках полости, в которых формируются
пузырьки
· Переносимые
пузырьком белки и липиды изменяются ферментами, которые находятся в полости
каждого компартмента
· Основным местом
гликозилирования белков и липидов является комплекс Гольджи
· Лизосома, основной
гидролитический компартмент клетки, получает свои ферменты из различных
источников
. Молекулярные механизмы
формирования и движения пузырьков
Отпочкование пузырьков
Основным механизмом, с помощью
которого белки и липиды перемещаются в клетке, является почкование
пузырьков. Пузырьки формируются из мембраны одной органеллы, называемой донором,
и затем перемещаются к другой органелле, называемой акцептором. Это
приводит к слиянию донорной и акцепторной мембран и высвобождению содержимого
пузырька в полость акцептирующего компартмента. В результате этого процесса
происходит перестройка клеточных компартментов и поверхности клетки, а также
сохранение или разрушение межклеточных контактов.
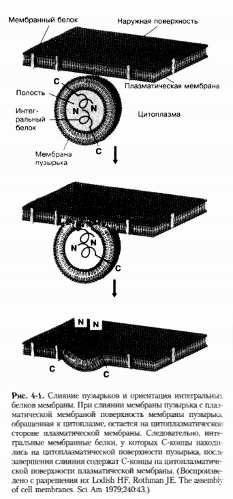
Таким образом, при слиянии
транспортных пузырьков ЭР с акцепторными мембранами CGN определенные рецепторные
белки и липиды возвращаются их КГ обратно в ЭР. Этот процесс называется
ретроградным транспортом. В противоположность ему, при антероградном
транспорте растворимые грузовые белки продолжают перемещаться по секреторному
пути. Раздельный транспорт антероградных грузовых белков и ретроградно
транспортируемых компонентов обеспечивается, по крайней мере, двумя классами
белков со специфической белковой каймой, которые связываются с транспортными
пузырьками. Эти белковые комплексы называются коатомерами или COPs.
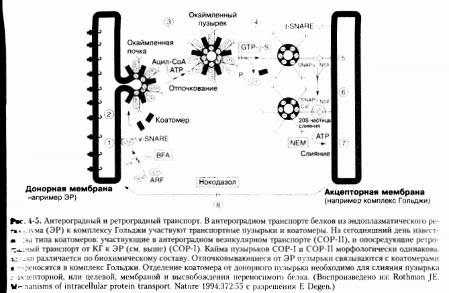
COP-I
Цитозольные белки, которые
закрепляются на мембране КГ и формируют протеиновую кайму, обозначаются как
коатомер или COP-I. В коатомерном комплексе есть небольшой GTP - связывающий белок,
называемый также фактором рибозилирования аденозиндифосфата. Генетическое
исследование дрожжевых клеток подтвердило, что мутации инактивирующие
субъединицы коатомеров или изоформы ARF, блокируют также внутриклеточный везикулярный аппарат и секрецию.
Таким образом, была установлена роль пузырьков COP-I в процессе внутриклеточного
транспорта.
COP-II
Второй белок каймы, состоящий из
пяти субъединиц, был биохимически и генетически охарактеризован в дрожжевых
клетках. Этот белковый комплекс, называемый COP-II, участвует в
антероградном транспорте и способствует переносу пузырьков от ЭР к КГ.
SNAREs и направление пузырьков
Результаты экспериментов казали, что
окаймленные коатомерами пузырьки не могут слиться с мембраной акцептора, до тех
пор пока белковая кайма не будет удалена. Неокаймленные транспортные пузырьки
могут сливаться с мишенями, но для этого в цитозоле должен присутствовать
N-этилмалеимиду, вызывающему его инактивацию. В дрожжевых клетках
существует эквивалент NSF: продукт гена sec 18. При мутации в белке sec18 секреция нарушается, так как транспортные пузырьки,
отпочкованные от ЭР, накапливаются в высоких концентрациях, но не могут слиться
с акцепторной мембраной Гольджи.
Межвидовая
консервативность: NSF
NSF - это тример, содержащий три идентичные субъединицы массой 76 кДА
и присоединяющий растворимые белки SNAPs (soluble NSF attachment proteins), которые способствуют связыванию с мембранами комплекса Гольджи.
Комплекс NSF-SNAPs связывается с мембранами с помощью мембранного рецептора,
называемого ЛОВУШКОЙ (SNARE).
Пузырьки, покрытые
клатрином
Поглощение лиганд-рецепторных
комплексов с поверхности плазматической мембраны происходит путём образования
окаймленных ямок, которые затем превращаются в окаймленные пузырьки. Эти
специфические белковые комплексы строятся из белка клатрина. Клатрин формирует
корзиноподобную клетку, в которую втягивается мембранный бислой.
Пузырьки, покрытые клатрином,
участвуют в движении веществ от плазматической мембраны к эндосомам и от
комплекса Гольджи к плазматической мембране.
Направление движения
пузырьков
Вновь сформированные пузырьки могут
слиться с различными мембранами. Для обеспечения правильного направления каждый
пузырёк снабжен специальной рецепторной молекулой, называемой ЛОВУШКА-п (для
пузырька). Акцепторная, или целевая, мембрана также обладает рецептором,
связывающимся с ЛОВУШКОЙ-п, и он называется ЛОВУШКА-ц (для целевой мембраны).
Ловушки обеспечивают правильную доставку пузырьков, отпочкованных от донорной
мембраны.
Формирование комплекса
слияния
Для слияния двух мембран (то есть мембраны
пузырька и целевой мембраны) требуется специальный белок слияния (фузионный
белок). Этот белок дестабилизирует гидрофильные силы в месте взаимодействия
двух мембран. Когда две мембранные поверхности приближаются друг к другу,
гидрофобный домен белка слияния направляет молекулы воды в разные стороны, и
наружные липидные слои сливаются друг с другом. То же происходит при слиянии
внутренних слоёв.
GTPазы: стыковка и слияние
В клетках существует два типа GTPаз, и оба типа участвуют
в контролировании процессов молекулярной сигнализации. К первому типу относятся
тримеры, белки имеющие три субъединицы (α, β и γ). Мономеры, состоящие
из одного полипептида, составляют второй тип GTPаз.
G-белки
Тримерные GTPазы - это крупное
семейство белков, называемых обычно
G-белками. Они участвуют в
переносе сигналов с наружной стороны клетки внутрь. Каждая тримерная
субъединица участвует в переносе различных сигналов к целевым молекулам в
клетке.
Мономерные GTPазы
Вторым крупным классом GTPаз являются мономеры. На
сегодняшний день выявлено 5 подсемейств:
|
GTPазы
|
Функция
|
|
Ras
|
Участвует в росте и дифференцировке
|
|
Rho
|
Связан с активностью интегрина и формированием актинового
цитоскелета
|
|
Rab
|
Участвует в транспорте пузырьков в клетке
|
|
ARF
|
Связан с формированием пузырьков
|
|
Ran
|
Участвует в транспорте белков ядра
|
Rab - белки
Rab-белки участвуют в везикулярном транспорте следующим образом.
Пузырёк сформированный на мембране донора, содержит ЛОВУШКУ-п и молекулу Rab в GTP-связанной конфигурации.
Когда пузырёк состыковывается с соответствующей ЛОВУШКОЙ-ц, Rab-GTP гидролизуется. Этот
процесс инициирует слияние пузырька с мембраной акцептора. Rab в GDP - связанной конфигурации отщепляется от мембраны и возвращается к
мембране донора, где GTP-активирующий белок восстанавливает Rab, катализируя связывание
с GTP.
Это приводит к тому, что Rab связывается с липидной частью мембраны и участвует в новом
направляющем процессе. Для каждой внутриклеточной мембраны существуют
специфические Rab-белки, которые обеспечивают определенное направление пузырьков.
3. Эндоцитоз
Эндоцитоз - везикулярный захват
жидкостей, макромолекул или небольших частиц в клетку. Существует по крайней
мере три механизма эндоцитоза:
1. Пиноцитоз, который дословно обозначает «клеточное питьё». Он также
называется клатрин-независимым эндоцитозом.
2. Рецепторно-опосредованный
эндоцитоз или клатрин-зависимый эндоцитоз.
Этому процессу уделяется большое внимание, поскольу он вызывает определенные
заболевания у человека.
3. Фагоцитоз, который дословно означает «клеточная еда».
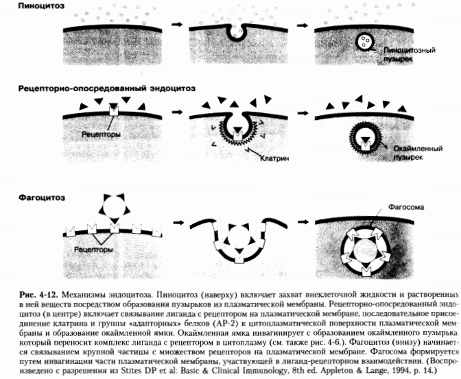
Пиноцитоз
Пиноцитоз - это конститутивный
процесс, то есть он принимает участие в постоянном динамическом образовании
небольших пузырьков на поверхности клетки. Точнее, мелкие инвагинации и
пузырьки образуются на поверхности клетки, затем они поглощаются и сливаются с
другими пузырьками, находящимися близко к поверхности и формируют первичные
эндосомы. На периферии клетки эти пузырьки встраиваются в поверхностную
мембрану.
Таким образом, на клеточной
поверхности образуются и постоянно работают пузырьки, которые доставляют
вещества в клетку и восстанавливают плазматическую мембрану. Эти пузырьки
переносят небольшие молекулы, воду и растворимые белки, то есть вещества,
относящиеся к жидкой фазе внеклеточной среды. Несмотря на маленькие размеры
пиноцитозных пузырьков, их многочисленность позволяет им доставлять в клетку
большое количество веществ.
Рецепторно-опосредованный
эндоцитоз
Рецепторно-опосредованный эндоцитоз
использует для переноса молекул специфические поверхностные рецепторы.
Рецепторно-опосредованный эндоцитоз обладает определенными перимуществами:
· Специфичность. Только клетки определенного типа экспрессируют поверхностные
рецепторы, что обеспечивает избирательное связывание молекул во внеклеточном
растворе. Экспрессия определенных растворов у клеток одного типа являются общим
механизмом развития и формирования ткани.
· Способность
к концентрированию лиганда на поверхности клетки. По законам диффузии молекулы перемещаются из среды с высокой
концентрацией. Таким образом, если клетки могут избирательно удалять лиганды из
окружающего раствора, они действуют как водосток для этих молекул. В конце
концов, лиганд будет удалён из окружающей жидкости.
Основные характеристики
рецепторно-опосредованного эндоцитоза
После того как раньше эндосомы
сливаются с другими эндосомами, содержащими ATP-азную протонную помпу,
и pH
в полости понижается, лиганд-рецепторный комплекс может пойти по одному из
следующих путей.
1. Рецептор возвращается после
высвобождения груза
2. Рецептор и переносимый
лиганд возвращаются, как в случае с трансферрином
. Рецептор и лиганд
разрушаются в лизосоме
. Рецептор и лиганд
транспортируются через клетку и доставляются к противоположной стороне
мембране; это происходит главным образом в полярных клетках

Фагоцитоз
Фагоцитоз - это захват клетками
относительно крупных частиц с помощью клатрин-независимого, актин-зависимого
механизма. Фагоцитоз представляет собой ключевой механизм защиты
организма-хозяина от микроорганизмов; фагоцитоз поврежденных или постаревших
клеток необходим для обновления тканей и заживления ран. Фагоцитоз особенно
важен для многоклеточных организмов. К тому же фагоцитоз является общим
механизмом, используемым микроорганизмами для защиты от прямого разрушающего
действия антител и белков комплемента и цитотоксических клеток.
Фагоцитоз у млекопитающих
осуществляют в основном клетки трех типов: нейтрофилы, моноциты и макрофаги. На
поверхности этих клеток расположены специальные рецепторы, предназначенные для
распознавания и проведения процесса фагоцитоза. Эти рецепторы распознают
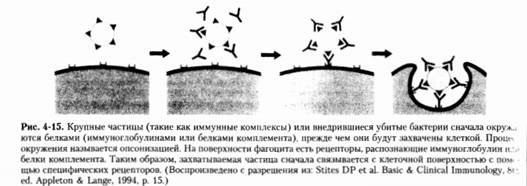
неантиген-связывающий участок иммуноглобулинов или других молекул, которые
входят в состав иммунной системы организма-хозяина. Антитела и белки
комплемента в плазме окружают поверхность клетки микроорганизма, после чего
микроб связывается с рецептором фагоцита и начинается фагоцитарный процесс.
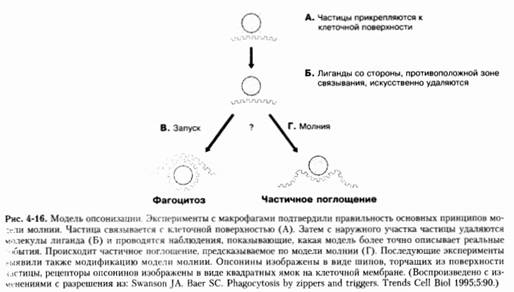
Этот процесс называется опсонизацией.
Захват опсонизированных частиц
происходит по механизму молнии, в котором частица сначала связывается с Fc-рецептором на
поверхности фагоцита. Затем следует последовательное соединение с другими поверхностными
рецепторами клетки. Это соединение приводит к образованию фагосомы, которая
принимает форму захваченной частицы. Таким образом, плазматическая мембрана
плотно прилегает к частице, словно на липучке.
везикулярный пузырек транспорт
клеточный
. Трансцитоз
Вещества, проникающие в с помощью
специфических рецепторов, почти всегда формируют под плазматической мембраной
пузырьки, называемые ранними эндосомами. Эти пузырьки служат местами сортировки
поглощенных лигандов и рецепторов. В полости ранней эндосомы значение рН
приблизительно равно 6,0, что часто приводит к разрыву связи между лигандом и
рецептором. Если это происходит, то и лиганд, и рецептор обычно сортируются в
другой пузырёк, который отпочковывается от ранней эндосомы, сливается с
лизосомой и переваривается. Однако некоторые комплексы лиганда с рецептором не
распадаются при пониженном рН. Вместо этого они сортируются в другие участки
ранней эндосомы и образуют почкующиеся пузырьки, которые затем сливаются с
другими внутриклеточными мембранами или с различными участками плазматической
мембраны. Этот процесс называется трансцитозом. Трансцитоз - это
механизм, посредством которого молекулы, пришедшие в клетку извне, могут
доставляться к различным местам внутри клетки или даже перемещаться от одного
слоя клеток к другому.
5. Экзоцитоз
У эукариот
клеточный процесс, при котором внутриклеточные везикулы (мембранные пузырьки)
сливаются с внешней клеточной мембраной. При экзоцитозе содержимое секреторных
везикул (экзоцитозных пузырьков) выделяется наружу, а их мембрана сливается с
клеточной мембраной. Практически все макромолекулярные соединения (белки,
пептидные гормоны и др.) выделяются из клетки этим способом. У прокариот везикулярный
механизм экзоцитоза не встречается, у них экзоцитозом называют встраивание
белков в клеточную мембрану (или в наружную мембрану у грамотрицательных
бактерий), выделение белков из клетки во внешнюю среду или в периплазматическое
пространство.
Экзоцитоз может
выполнять три основные задачи:
· доставка на клеточную мембрану липидов, необходимого для роста
клетки;
· высвобождение различных соединений из клетки, например, токсичных
продуктов метаболизма или сигнальных молекул (гормонов или нейромедиаторов);
· доставка на клеточную мембрану функциональных мембранных белков
таких как рецепторы или белки-транспортёры. При этом часть белка, которая была
направлена внутрь секреторной везикулы, оказывается выступающей на наружной
поверхности клетки.
Типы
экзоцитоза
1. Кальций-независимый конститутивный экзоцитоз встречается практически во всех
эукариотических клетках. Это необходимый процесс для построения внеклеточного
матрикса и доставки белков на внешнюю клеточную мембрану. При этом секреторные
везикулы доставляются к поверхности клетки и сливаются с наружной мембраной по
мере их образования.
2. Кальций-зависимый
неконститутивный экзоцитоз встречается,
например, в химических синапсах или клетках, вырабатывающих макромолекулярные
гормоны. Этот экзоцитоз служит, например, для выделения нейромедиаторов. При
этом типе экзоцитоза секреторные пузырьки накапливаются в клетке, а процесс их
высвобождения запускается по определённому сигналу, опосредованному быстрым
повышением концентрации ионов кальция в цитозоле клетки. В пресинаптических
мембранах процесс осуществляется специальным кальций-зависимым белковым
комплексом SNARE.
Этапы
экзоцитоза
· Транспортировка
везикулы от места синтеза и формирования (аппарат Гольджи) до места доставки
осуществляется моторными белками вдоль актиновых филаментов либо микротрубочек
цитоскелета. Этот этап может потребовать перемещения секретируемого материала
на значительное расстояние, как, например, в нейроне. Когда везикула достигает
места секреции, она входит в контакт со специфическими удерживающими факторами
клеточной мембраны.
· Удержание доставленной
везикулы обеспечивается относительно слабыми связями на расстоянии более 25 нм
и может служить, например, для концентрирования синаптических везикул около
пресинаптической мембраны.
· Стыковка везикулы с мембраной
является непосредственным продолжением первой фазы доставки, когда мембрана
везикулы входит в близкий контакт с мембраной клетки (5-10 нм). Это включает
прочное соединение белковых компонентов обеих мембран, вызванным внутримолекулярными
перестановками, и предваряет формирования SNARE комплекса.
· Стимуляция (прайминг)
везикулы фактически соответствует образованию особого SNARE комплекса между
двумя мембранами и осуществляется только в случае нейронального экзоцитоза.
Этот этап включает процессы молекулярных перестановок и АТФ-зависимые
модификации белков и липидов, происходящие непосредственно до слияния мембран в
ответ на подъём уровня свободного кальция. Этот кальций-зависимый процесс
необходим для контролируемого быстрого выброса нейромедиатора и отсутствует в
случае конститутивного экзоцитоза.
· Слияние мембраны везикулы с
мембраной клетки приводит к высвобождению, или выбросу, содержания
секретируемой везикулы во внеклеточное пространство и объединению липидного
бислоя везикулы с внешней мембраной. В случае синаптического выброса процесс
слияния, так же как и стимуляция, осуществляется SNARE комплексом.
Заключение
Везикулярный транспорт подчиняется
общим принципам организации во всех клетках, начиная от культуры клеток дрожжей
и заканчивая клетками организма человека, и играет важную роль в целом спектре
физиологических процессов, когда имеет место контроль за слиянием везикул с
клеточной мембраной (синтез и секреция гормонов и цитокинов). Нарушения в
везикулярном транспорте происходят при многих заболеваниях, включая ряд
неврологических и иммунологических нарушений, сахарный диабет. Когда нарушается
эта удивительная упорядоченность транспорта молекул, клетка переходит в
состояние полного хаоса.
Важно, чтобы студенты смогли понять
основные молекулярные механизмы везикулярного транспорта и оценить огромное
число молекул, участвующих в этом процессе.
Список использованной
литературы
1. Фаллер Д.М., Шилдс Д. Молекулярная биология клетки. Руководство
для врачей. Пер с англ. М.: БИНОМ - Пресс, 2003
2. Мушкамбаров Н.Н. Кузнецов С.Н. Молекулярная биология.
Учебное пособие для студентов медицинских вузов, Москва: Наука, 2003