Биологические особенности сибирской ряпушки в связи с её искусственным разведением
ОГЛАВЛЕНИЕ
Введение
Глава 1. Биологическая характеристика сибирской ряпушки в связи со
средой обитания и образом жизни
1.1 Общая
биология вида
.2
Эмбриональное развитие
.3
Постэмбриональное развитие
Глава 2. Влияние различных факторов среды на сибирскую ряпушку
2.1 Влияние
параметров водной среды (температура, гидрохимические показатели)
.2 Влияние
освещенности, уровня и течения воды на сибирскую ряпушку
Глава 3. Управление половыми циклами у сибирской ряпушки различными
методами
Глава 4. Биологические основы кормления сибирской ряпушки
Глава 5. Транспортировка икры, личинок, молоди и взрослых особей
сибирской ряпушки
Глава 6. Биологические основы акклиматизации
Библиографический список
ВВЕДЕНИЕ
Несмотря на произошедшие в последние годы негативные изменения в рыбном
хозяйстве, Российская Федерация продолжает оставаться одним из ведущих
рыбопромышленных государств, определяющих стратегию развития мирового
рыболовства (www.pravo.gov.ru).
Меры охраны естественного воспроизводства сиговых дают ожидаемые результаты
только в районах, где нерестовый фонд находится в удовлетворительном состоянии
и численность нерестовых популяций обеспечивает необходимый уровень пополнения
запаса. Поскольку в большинстве районов мира эти условия нарушены, все большее
развитие получает управляемое воспроизводство, в частности, искусственное
разведение молоди и выпуск ее в естественные водоемы.
Искусственное разведение как способ пополнения запасов проходных сиговых
имеет определенные преимущества - возможность контроля на всех этапах
эмбрионально-личиночного и малькового развития. Это позволяет выращивать
потомство с определенными свойствами, нужного размера и качества, обеспечивает
повышенную сохранность особей, эффективность селективного воздействия с целью
повышения жизнестойкости покатной молоди.
Основные задачи, стоящие перед искусственным рыборазведением:
. Совершенствование биотехники искусственного рыборазведения ценных
промысловых видов рыб;
. Обеспечение населения товарной продукцией таких рыб как карп,
растительноядные, осетровые и др.;
. Сохранение естественного размножения рыб и повышение эффективности
процесса подращивания;
. Обеспечение нормальных условий для нагула ценных рыб и поддержание
генофонда популяций (Пономарева, 2004).
Во внутренних водоемах Сибири сосредоточены основные запасы наиболее
ценных видов рыб Российской Федерации. К ценным видам рыб отнесены
представители четырех семейств: осетровые, лососевые, хариусовые и сиговые.
Статистическая информация собрана по 18 субъектам Федерации, входящим в
Сибирский регион от Республики Саха (Якутия) и Читинской области - на востоке
до Омской и Тюменской областей - на западе. Наиболее крупные бассейны региона -
Обь-Иртышский, бассейны рек Енисей, Пясина, Таймура, оз. Байкал, р. Лена и р.
Колыма (Смешливая, 2015).
В настоящее время запасы промысловых представителей сиговых рыб находятся
в критическом состоянии. Динамика уловов за последние двадцать лет имеет
стабильную тенденцию к спаду.
Сиговые рыбы составляют основу промысловых запасов ценных видов рыб
Сибири и Урала. На 2013 г. их суммарная величина ОДУ определена в 15,7 тыс. т,
что составляет около 80% от общей величины ОДУ сиговых по стране. В 2005 г. ОДУ
этой группы рыб оценивался в 22,7 тыс. т, в 2010 г. - 17,5 тыс. т. За
восемь лет снижение ОДУ составило 7 тыс. т, что составляет 30,8%. Только за
последние три года снижение составило 1,8 тыс. т, 10,1% по отношению к 2010 г.
Наиболее существенное падение запасов наблюдается в Обь-Иртышском бассейне, где
величина ОДУ сиговых рыб снизилась за указанный период в два раза. Снижение
запасов обских сиговых видов рыб отчасти связано с влиянием колебаний водности
бассейна, но основной причиной является антропогенный пресс. Наиболее массовые
сиговые рыбы Сибири - ряпушка сибирская (28% от ОДУ), пелядь, омуль (по 18%),
сиг-пыжьян (13%), чир (11%) и муксун (7%). Запасы сиговых рыб распределены
неравномерно. Наиболее значительны они в Обь-Иртышском бассейне - 42% от
величины ОДУ сиговых рыб Сибири. Доля водоёмов Якутии - 30%; Красноярского края
- 18%; Байкала - 10%. По экспертным оценкам фактический вылов находится на
уровне величины ОДУ. В период устойчивого функционирования рыбохозяйственного
комплекса страны в 70-80-е гг. прошлого века в естественных водоёмах Сибири в
среднем вылавливалось 20 тыс. т сиговых в год. На основании расчётов,
основанных на величине промвозврата, определено, что в 2010 г. мероприятия по
искусственному воспроизводству обеспечили 1,55 тыс. т ОДУ сиговых рыб в
регионе. Величина ОДУ, обеспеченная работой рыбоводных предприятий, оценивается
в 430 т по омулю, 440 т по пеляди и 300 т по муксуну. Наиболее масштабно
проводится воспроизводство сиговых рыб. В настоящее время в Сибири и на Урале
действует 14 рыбоводных предприятий, специализирующихся на воспроизводстве этой
группы рыб. К основным, стабильно работающим предприятиям, относятся только
Большеречинский рыбоводный завод (оз. Байкал), Тобольский региональный
рыбопитомник (Обь-Иртышский бассейн). Суммарное производство личинок сиговых в
2010 г. составило 975 млн экз., в 2011 г. - 1455 млн экз.. Необходимо отметить,
что за последние десять лет прослеживается тенденция к снижению количества
ежегодно выпускаемых личинок сиговых рыб. Так, если в 2001-2003 гг. в среднем
выпускалось 1,92 млрд личинок в год, то в 2009-2011 гг. - всего 1,46 млрд
экз. Снижение объёмов искусственного воспроизводства сиговых рыб связано в
основном с организационно-финансовыми причинами (Литвиненко, 2015).
Целью курсовой работы является изучение биологических особенностей
сибирской ряпушки в связи с её искусственным разведением.
Поставленная цель определила следующие задачи :
изучить биологическую характеристику сибирской ряпушки в связи с её
средой обитания и образом жизни;
изучить влияние различных факторов среды на сибирскую ряпушку;
изучить управление половыми циклами сибирской ряпушки различными
методами;
изучить биологические основы кормления сибирской ряпушки;
изучить транспортировку икры, личинок, молоди и взрослых особей сибирской
ряпушки;
изучить биологические основы акклиматизации сибирской ряпушки.
ГЛАВА 1. БИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА
СИБИРСКОЙ РЯПУШКИ В СВЯЗИ СО СРЕДОЙ ОБИТАНИЯ И ОБРАЗОМ ЖИЗНИ
.1 Общая биология вида
Тип: Хордовые - Chordata
Класс: Лучепёрые рыбы - Actinopterygii, Teleostomi
Отряд: Лососеобразные - Salmoniformes
Семейство: Лососевые - Salmonidae
Подсемейство: Сиговые - Coregoninae
Род: Сиги - Coregonus
Вид: Сибирская ряпушка - Coregonus sardinella (Valenciennes, 1848) (рис.
1).
Рот - верхний; нижняя челюсть слегка выдается над передним краем верхней
челюсти. Боковая линия 71-97. Жаберных тычинок 37-52 (меньше, чем у ладожского
рипуса, у которого их насчитывается 43-56).
сибирская ряпушка икра акклиматизация

Рисунок
1 - Сибирская ряпушка, Coregonus sadinella (Valenciennes, 1848
Позвонков (с уростилем) у основной формы (С. sardinella typ. - Обская
губа, оз. Хасейнто, Лена) 55-66, у беломорской ряпушки (С. sardinella maris-
albi-Печора, озера Болыпеземельской тундры) 53-60, у белозерской ряпушки (С.
sardinella vessicus) 54-57. Пилорических придатков (Обь) 53-74. D Ш-IV 8-11(12);
A III-IV 11-14.
Сибирская ряпушка, С. sardinella, обитает в бассейне Северного Ледовитого
океана от pp. Кары и Оби до pp. Колымы и Берингова моря и в р. Анадыре, а также
на Аляске и в р. Мэккензи. Подвид беломорская ряпушка, С. sard, maris-albi,
распространена от Соловецких островов до р. Печоры и о. Колгуева. Подвид
белозерская ряпушка, С. sard, vessicus, водится в Белом озере (Вологодская
обл.), подвид С. sard, baunti - в озерах системы Витима (Решетников, 1980).
Рыба низовьев и придельтовых пространств северных рек; редко живет в
озерах; заходит в солоноватые воды заливов и губ; холодолюбивая рыба,
образующая косяки и мигрирующая для нереста (сравнительно на небольшие
расстояния) вверх по рекам (www.aquacultura.org).
Совершает также суточные вертикальные миграции. Морфологически и
экологически очень пластична; образует местные стада по рекам и внутри системы
одной реки: например, в Оби отмечены: щучьереченское стадо - живет в Обской
губе, нерестует в р. Щучьей; тазовское стадо - зимовка в Обской губе, нерест в
р. Мессо; стадо собственно Обской губы и Ямала - нагул и зимовка в Обской губе,
нерест в самой губе у Нового Порта и в речках, текущих с Ямала.
Восточно-сибирская ряпушка pp. Лены и Колымы (раньше выделялась в отдельный
подвид С. merkii) отличается от других ряпушек своими крупными размерами:
средний вес половозрелых особей в промысловых уловах 300-320 г.
Нерест происходит перед ледоставом (иногда после ледостава), при
температуре воды около 4°С: в конце сентября - начале октября в заполярных
районах и в октябре-начале ноября в более южных районах. В бассейне Оби
нерестилища расположены в среднем и нижнем течении р. Щучьей, частью в Соби,
Войкаре, Сыне и Сосве (щучьереченское стадо), в р. Мессо (тазовское стадо), в
бухте у Нового Порта (новопортовское стадо) и в речках Обской губы (Се-яга,
Тамбей и др.). В Енисее нерестилища расположены между устьями p. Подкаменной
Тунгуски и р. Комса, но основным нерестовым участком является район сел. Бахта
и Мирное, протяжением около 20 км (1200 км от устья). В р. Колыме основные
нерестилища располагаются от Нижнеколымска до с. Крестов (расстояние 237 км),
частью до с. Гороховой и, как исключение, до Родчево и Верхнеколымска
(расстояние от Нижнеколымска 859 км). В Печоре нерестилища расположены ниже
устья р. Усы, в Каре на расстоянии 40 км от устья. В Анадыре ряпушка нерестует
у с. Марково (Шестаков, 1996).
Плодовитость обской ряпушки 7-18,3 тыс., в среднем 11,1 тыс., енисейской
- 2,5-28,6 тыс. икринок.
Зрелая овариальная (ястычная) икра желтого цвета, диаметр икринки 1,5 мм.
Икра вначале слабоклейкая. Инкубация продолжается до конца мая - начала июня.
Личинки и мальки попадаются весною ниже нерестилищ, непосредственно после
вскрытия рек, и частью выходят на пойму, в соры и озера, расположенные вблизи
реки, частью остаются в реках. Молодь кормится в дельтовых участках, губах и
частью в сорах низовьев. Длина сеголетков в Колыме в июле 2,3 см, в августе
4,2, в сентябре 6,6 см.
Сибирская ряпушка живет до десяти лет. Величина промысловых ряпушек в
разных реках очень различна. Половозрелость наступает у обской и енисейской
ряпушки на четвертом-пятом году жизни, у колымской на пятом-шестом, у
хатангской на шестом-седьмом (самцы достигают половой зрелости на год раньше).
Ленская ряпушка созревает на шестом-седьмом году, белозерская и чарандская - на
втором-третьем году. В основном питается планктоном (рачки Bosmina,
Bythotrephes, Diaptomus, циклопы, мизиды), реже придонными организмами (личинки
подёнок, ручейников, комаров) и воздушными насекомыми; как исключение,
заглатывает мальков рыб, растительный детрит, жуков, обрывки травы и т. п. В
период хода почти не питается.
Миграции сибирской ряпушки в бассейне Оби сравнительно невелики по
расстояниям, что объясняется близостью мест размножения к местам нагула.
В Оби ряпушка новопортовского стада из Обской губы подходит для нереста в
бухту и к берегам в сентябре - начале октября. Из Тазовской губы в р. Мессо ход
начинается со второй половины августа, усиливается во второй половине сентября
и заканчивается в первой половине октября. К р. Щучьей из Обской губы ряпушка
подходит с конца июня - первых чисел июля и держится около устья реки до конца
августа, затем поднимается вверх и в октябре нерестует. Из Енисейского залива
ход ряпушки начинается со второй половины июня и продолжается в течение
июля-августа; к нерестилищам, расположенным в Туруханском и Ярцевском районах
(села Мирное, Бахта, в 1200 км от устья), она подходит в конце августа -
сентябре-октябре. В низовьях Лены и Колымы нерестовый ход наблюдается во второй
половине августа - сентябре.
Производители после нереста отходят от берегов и скатываются в дельту и
губы. Ход ряпушки вверх по р. Анадырю начинается в августе, в конце сентября
она достигает с. Марково (Берг, 1948).
.2 Эмбриональное развитие
Эмбриональный период развития сиговых рыб различными авторами разделяется
на 7, 8, 9, 10 либо 13-14 этапов. Несоответствие в числе этапов развития имеет
место на стадиях органогенеза и позднее. Исходя из того, что "этап
развития - это отрезок развития, на протяжении которого происходит становление
новой функции у группы одновозрастных особей" (Сытина, 1973).
Эмбриональный период развития протекает с момента оплодотворения икринки
до выхода личинки. В это время зародыш растет и развивается только за счет
запаса питательных веществ, содержащихся в икринке.
Эмбриональный период сибирской ряпушки продолжается 220-240 суток
(Макеева, 2011).
Эмбрионы ряпушки эвритермны, эмбриональное развитие по данным О. А.
Лебедевой (1983), в норме может происходить при температуре воды от +0 до 10 оС,
при содержании кислорода от 5,0 до 14,0 мг/л. Д. Прайс, проводя инкубацию икры
сиговых рыб при постоянной температуре от 0 до 12 оС, обнаружил, что
успешное развитие эмбрионов происходит в узком диапазоне температур (от 0,5 до
6 оС), а наиболее благоприятная температура +0,5 оС.
Скорость эмбрионального развития сиговых рыб зависит от температуры
среды. Установлено, что повышение температуры воды до известных пределов
вызывает ускорение развития. Оплодотворение и прохождение начальных стадий
эмбриогенеза при более низкой температуре воды приводит к сокращению
продолжительности развития в зимний период. Уменьшение длительности
эмбриогенеза сближает даты вылупления. Динамика развития эмбрионов сибирской
ряпушки при разных температурах воды показана в таблице 1. В процессе
эмбриогенеза происходит закономерное изменение количества каротиноидных
пигментов в цитоплазме и в желтке (Микулин, 1978). Интенсивность пигментации
икры, в основном, обусловлена интенсивностью цвета жировых капель, собственно
желток и плазма окрашены слабо (Валюшок, 1988).
Таблица 1 - Динамика развития эмбрионов сибирской ряпушки при разных
температурах воды (сут) (Черняев, 1987)
|
Контролируемые этапы и
стадии развития
|
Температура воды, оС
|
|
0,2
|
2,0
|
4
|
6
|
|
Начало дробления
|
1-2
|
1-2
|
1
|
1
|
|
Морула крупных клеток
|
2-11
|
2-7
|
1-5
|
1-3
|
|
Бластула
|
7-14
|
6-10
|
4-7
|
3-5
|
|
Обрастание желтка
бластодермой, образование нервной трубки, хорды
|
18-37
|
11-23
|
9-19
|
7-15
|
|
Обособление хвостового
отдела от желточного мешка
|
34-43
|
21-28
|
19-25
|
15-20
|
|
Начало пигментации глаз и
пульсации сердца
|
50-62
|
34-48
|
28-37
|
24-28
|
|
Начало вылупления эмбрионов
|
135
|
99
|
90
|
80
|
|
Массовое вылупление
эмбрионов
|
163-166
|
120-131
|
104-114
|
92-107
|
|
Завершение вылупления
эмбрионов
|
170
|
150
|
133
|
121
|
Подготовка эмбрионов к выходу из оболочки один из критических периодов эмбрионального
развития (Мешков, 1965). Известно, что выделение ферментов из желез вылупления
происходит под влиянием изменений светового и температурного режимов, химизма
воды, а также вследствие действия на икру толчков и встряхиваний (Богданов,
2006).
.3 Постэмбриональное развитие
Вылупление личинок ряпушки происходит дружно в короткий срок за 6-8 дней
при температуре воды 3,5-7,0 °С, но при более низких температурах (1,5-5,0 °С)
этот процесс растягивается до 10-15 дней. Предличинки мелкие: длина тела -
7,50-7,53 мм, масса - 2,10-2,18 мг (табл. 2).
Таблица 2 - Морфометрические
показатели предличинок сибирской ряпушки массового вылупления
|
Показатель
|
Колебания средних величин
|
|
Длина, мм общая тела
|
7,91-7,95 7,50-7,53
|
|
Высота, мм тела желтка
|
0,57-0,60 0,56-0,57
|
|
Масса, мг тела желтка
|
2,10-2,18 0,07-0,08
|
|
Масса желтка от массы тела,
%
|
3,20-3,80
|
|
Диаметр жировой капли, мм
|
0,67-0,69
|
Личиночные этапы охватывают развитие от начала активного питания внешней
пищей до малькового периода. Время наступления этапов определяется условиями
обитания, температурным фактором, обеспеченностью кормом. Ряпушка сибирская
переходит на экзогенное питания, как и другие сиговые, когда количество желтка
определяется как остаточное. В этот момент особи внутри вида, независимо от
сроков и стадии вылупления, длительности и температуры выдерживания, имеют
одинаковую, степень развития жаберно-челюстного аппарата, дифференцировку
плавниковой каймы, интенсивность пигментации и близкие линейно-весовые
показатели. У личинок, не получающих экзогенный корм после резорбции желтка,
вначале происходит увеличение длины тела и количества жаберных лепестков на
жаберных дугах, дифференциация плавников, а в дальнейшем развитие прекращается:
исчезает скопление мезенхимы в спинном плавнике, уменьшается высота тела,
исчезает жировая капля или сохраняются ее небольшие остатки, снижаются масса и
упитанность тела. При сравнении степени развития сибирской ряпушки с другими
видами сиговых при переходе на личиночный этап установлено, что она более
развита, чем пелядь, но менее, чем муксун, занимая между ними промежуточное
положение (Сергиенко, 2015).
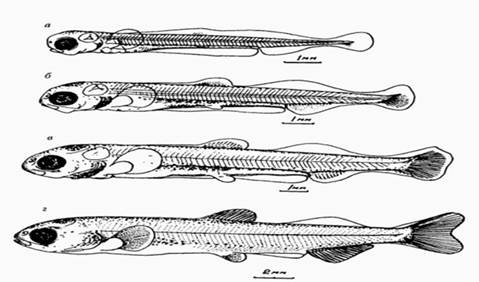
Рисунок 2 - Личиночное развитие сибирской ряпушки (Богданов, 1998).
В таблице 3 приведена морфологическая характеристика этапов личиночного
периода сибирской ряпушки.
Таблица 3 - Морфологическая
характеристика этапов личиночного периода сибирской ряпушки
|
Показа
|
|
|
|
Этап
|
|
|
|
|
тель
|
I
|
II
|
III
|
IV
|
V
|
VI
|
VII
|
|
Места
|
Начало
|
Область
|
Область
|
Лучей хво
|
Начало пиг
|
Боковой
|
Спинного
|
|
пигмента
|
пигмента
|
переднего
|
слуховых
|
стового
|
ментации
|
части туло
|
плавника,
|
|
ции
|
ции обла
|
мозга, обо
|
пузырей,
|
плавника у
|
боковой ли
|
вища, лу
|
слабая пиг
|
|
|
сти перед
|
нятельных
|
начало пиг
|
основания,
|
нии, хвосто
|
чей хвосто
|
ментация
|
|
|
него мозга
|
долей,
|
ментации
|
начало пиг
|
вой плавник
|
вого плав
|
туловища,
|
|
|
|
верхней
|
лучей хво
|
ментации
|
пигменти
|
ника за
|
хвостовые
|
|
|
|
челюсти,
|
стового
|
нижней че
|
рован у ос
|
середину,
|
лучи пиг
|
|
|
|
жаберных
|
плавника у
|
люсти
|
нования
|
слабая
|
ментирова
|
|
|
|
крышек,
|
основания
|
|
|
пигмента
|
ны почти до
|
|
|
|
жаберных
|
|
|
|
ция спин
|
конца
|
|
|
|
дуг
|
|
|
|
ного плав
|
|
|
|
|
|
|
|
|
ника
|
|
|
Количе
|
6 лепест
|
8-10 ле
|
12-16 ле
|
Жаберные
|
-
|
-
|
-
|
|
ство ле
|
ков,
|
пестков,
|
пестков, в
|
лепестки за
|
|
|
|
|
пестков +
|
2-4 бугор
|
2-4 бугорка
|
конце ста
|
кладывают
|
|
|
|
|
бугорков
|
ка
|
|
дии закла
|
ся на всем
|
|
|
|
|
на 2й и
|
|
|
дываются 6-
|
протяжении
|
|
|
|
|
3й жа
|
|
|
8 бугорков
|
верхних дуг
|
|
|
|
|
берных
|
|
|
на верхней
|
до конца
|
|
|
|
|
дугах
|
|
|
стороне дуг
|
|
|
|
|
|
Хвостовой
|
Начало за
|
4-5 гипу
|
Конец хор
|
20 ребри
|
20 лучей не
|
20 лучей
|
По 4 боко
|
|
плавник
|
кладки ги
|
ралий, 12-
|
ды загиба
|
стых лучей,
|
доходят до
|
доходят до
|
вых луча с
|
|
|
пуралий и
|
14 мезен
|
ется кверху;
|
одна линия
|
конца плав
|
конца
|
каждой сто
|
|
|
мезенхим
|
химных
|
18 лучей,
|
членисто
|
никовой
|
плавнико
|
роны,
|
|
|
ных лучей
|
лучей
|
начинается
|
сти, в конце
|
складки, в
|
вой склад
|
3 линии чле
|
|
|
|
|
их окосте
|
этапа 2 по
|
конце этапа
|
ки, две ли
|
нистости.
|
|
|
|
|
нение (реб
|
логие выем
|
закладыва
|
нии члени
|
Лучи выхо
|
|
|
|
|
ристость)
|
ки на конце
|
ется вторая
|
стости,
|
дят за край
|
|
|
|
|
|
хвостовой
|
линия чле
|
появляются
|
плавниковой
|
|
|
|
|
|
плавнико
|
нистости
|
боковые
|
складки
|
|
|
|
|
|
вой складки
|
|
лучи по бо
|
|
|
|
|
|
|
|
|
кам плавника
|
|
|
Спинной
|
Слабое
|
Скопление
|
Мускульные
|
Начало за
|
8 лучей, вы
|
12 лучей не
|
12 ребри
|
|
плавник
|
скопление
|
мезенхимы
|
почки
|
кладки хря
|
сотой до се
|
до конца
|
стых лучей
|
|
|
мезенхимы
|
и закладка
|
|
щевых лу
|
редины
|
плавнико
|
до конца
|
|
|
|
мускуль
|
|
чей
|
плавниковой
|
вой склад
|
плавниковой
|
|
|
|
ных почек
|
|
|
складки
|
ки
|
складки
|
|
Анальный
|
Слабое
|
Скопление
|
Мускуль
|
Мускуль
|
Закладка 8
|
14 лучей
|
14 лучей
|
|
плавник
|
скопление
|
ные почки
|
ные почки
|
мезенхимных
|
не до кон
|
не до кон
|
|
|
мезенхимы
|
|
в конце
|
|
лучей
|
ца плавни
|
ца плавни
|
|
|
|
|
этапа
|
|
|
ковой
|
ковой
|
|
|
|
|
|
|
|
складки
|
складки
|
|
Грудные
|
-
|
-
|
-
|
Скопление
|
Закладыва
|
8 лучей в
|
10 лучей
|
|
плавники
|
|
|
|
мезенхимы
|
ются 4 ме
|
верхней
|
не до кон
|
|
|
|
|
|
|
зенхимных
|
части
|
ца плавни
|
|
|
|
|
|
|
луча
|
плавников
|
ковой
|
|
|
|
|
|
|
|
|
складки
|
|
Глубина вы
|
От слабо
|
Седловидная,
|
Vобразная
|
В конце
|
-
|
-
|
-
|
|
емки на
|
выражен
|
до ⅓ ширины
|
выемка до
|
этапа по
|
|
|
|
|
спинной
|
ной до
|
плавниковой
|
⅔ ширины
|
чти полное
|
|
|
|
|
стороне
|
седловид
|
складки
|
плавнико
|
разделение
|
|
|
|
|
плавниковой
|
ной
|
|
вой складки
|
жирового и
|
|
|
|
|
складки
|
|
|
|
спинного
|
|
|
|
|
|
|
|
плавников
|
|
|
|
|
Плавательный
|
-
|
-
|
-
|
-
|
В конце этапа
|
-
|
-
|
|
пузырь
|
|
|
|
|
заполняется
|
|
|
|
|
|
|
|
воздухом
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
В эмбриональном развитии сиговых рыб выделяют нечувствительные стадии,
когда икринки можно перевозить, перебирать и исследовать без опасения массовой
гибели партии икры: после оплодотворения и до стадии мелкоклеточной морулы; от
стадии пигментации глаз и до появления и образования жаберных лепестков
(Саковская, 1991).
ГЛАВА 2. ВЛИЯНИЕ РАЗЛИЧНЫХ ФАКТОРОВ СРЕДЫ НА СИБИРСКУЮ
РЯПУШКУ
.1 Влияние параметров водной среды (температура, гидрохимические
показатели)
Температура воды является одним из важнейших факторов, оказывающих
воздействие на отправление жизненных функций рыбы, определяющих ее рост и
развитие. Этот фактор действует на рыбу как непосредственно - изменяя
интенсивность ферментативных процессов, происходящих в организме, активность
потребления пищи, характер обмена веществ, ход развития половой системы и
прочие, так и косвенно, оказывая влияние на улучшение или ухудшение состояния
кормовой базы (Иванов, 1988).
Рассматривая сиговых рыб по отношению к температуре можно отметить, что
это холодолюбивые, зачастую стенотермные формы.
Так, сибирская ряпушка - холодолюбивая, пойкилотермная рыба. Крайние
температуры, при которых она способна выживать 0-250С. Ряпушка
перестает питаться при 18-200С, а максимальный рост наблюдается при
температуре воды 8-150С. Повышение температуры свыше 230С
переносится плохо и зачастую вызывает гибель организма (Сергиенко, 2015).
Нерест ряпушки обычно начинается при температурах воды 6-80С и
заканчивается при охлаждении воды до 0-20С.
Хотя влияние температуры воды на миграции рыб очевидно, необходимо
указать, что она при этом не является единственным и определяющим фактором, а
воздействует в совокупности с другими условиями среды.
Продолжительность инкубационного периода и время выклева также напрямую
зависят от температуры. Минимальная температура во время развития эмбрионов, в
зависимости от типа нерестилища, может составить от 0,1 до 30С. Инкубационный
период проходит без отклонений от нормы, если температура воды соответствует
8-100С, а повышение ее ускоряет развитие только до определенной
границы. Если значительное повышение температуры пришлось на период
гаструляции, то резко возрастает % уродств, в худшем случае, происходит гибель
организма на ранних стадиях эмбрионального развития или ранних этапах
постэмбриогенеза. При повышении температуры личинки выходят из яйца меньших
размеров и морфологически недоразвитыми.
В зависимости от температуры воды изменяется и количество растворенного в
воде кислорода, который необходим для дыхания рыб. При понижении температуры
содержание кислорода в воде увеличивается, а при повышении уровень кислорода
снижается. Обмен веществ и потребление кислорода увеличивается
прямопропорционально повышению температуры воды и, наоборот, уменьшается с ее
понижением. Но это происходит только в определенных температурных пределах
(Шкорбатов, 1966).
По количеству кислорода, необходимого для нормального развития, всех рыб
делят на несколько групп, в которых сиговые занимают положение рыб, нуждающихся
в очень больших количествах кислорода (7-11 мг/л). При содержании его до 5 мг/л
дыхание их делается, практически, невозможным. То есть, представители лососевых
относятся к оксигенофильным рыбам, что объясняет их требовательность к
кислородному режиму.
Количество растворенного в воде кислорода зависит от температуры,
солености, ледового покрова, развития растительности, процессов распада
органических веществ и других факторов.
Немаловажной является и активная реакция среды. Влияние степени
концентрации водородных и гидроксильных ионов на гидробионтов установлено.
Жизнедеятельность каждого вида рыб протекает нормально лишь в определенных
границах концентраций. Как правило, активная реакция среды зависит от
соотношения растворенного в воде кислорода и свободной углекислоты, и
изменяется в зависимости от суточного и сезонного хода фотосинтеза.
Так как жизненный цикл ряпушки складывается из нескольких периодов, то и
воздействие активной реакции среды на организм в каждом из них будет различным.
Как правило, пресные воды подвержены большим колебаниям рН что связано, главным
образом, с цветением воды и массовым развитием планктонных водорослей. В свою
очередь, выделившиеся в большом количестве гуминовые кислоты, угнетают рост
рыбы. Также избыток СО2 в пресных водах вызывает увеличение
кислотности.
Как для сиговых, так и для других видов рыб, изменение границ рН вызывает
нарушения в обмене веществ, так как снижается способность организма поглощать
кислород. Оптимальная величина рН для сиговых составляет от 7 до 8.
Оптимальная рН для ряпушки входит в эти пределы (7-8), то есть
предпочтение отдается нейтральной или слабощелочной среде. Кислая среда, в
любых своих проявлениях, угнетающе действует на молодь.
Если же рН составляет меньше 5,6, ряпушка уже не может нормально
размножаться, а реакция среды со значениями выше 9 и более приводит к ее гибели
(Сергиенко, 2015).
Нитриты (NO2) накапливаются при повышенном уровне аммиака, могут вызывать
окисление двухвалентного железа гемоглобина крови в трехвалентное железо
метгемоглобина, неспособного переносить кислород. При этом кровь приобретает
коричневый цвет.
Для сиговых порог токсичности нитритов колеблется от 0,1 до 1 мг. При
хорошей аэрации нитриты окисляются до нитратов. Нитраты становятся токсичными
при концентрации 100-300 мг/л.
2.2 Влияние освещенности, уровня и течения воды на сибирскую
ряпушку
Свет в жизни рыб играет огромную роль. В зависимости от стадии
онтогенеза, физиологического состояния, а также видоспецифических особенностей
организм по-разному реагирует на действие светового фактора. Уже в период
эмбриогенеза действие светового фактора оказывает значительное влияние на
формирование будущего организма. Доказано, что у осенненерестующих рыб
наблюдается более быстрое развитие икры в темноте, вследствие чего происходит
более ранний выклев. Свет оказывает отрицательное влияние на развитие скелета у
эмбрионов данных видов (Коровина и др., 1965).
На ранних этапах постэмбриогенеза для сибирской ряпушки свет является
основным фактором, обеспечивающим пищепотребление, оборонительные реакции,
вертикальные и горизонтальные миграции, способствующие сохранению и
распространению вида. Личинки сибирской ряпушки проявляют положительный
фототаксис и более низкую пороговую чувствительность к свету по сравнению с
особями, находящимися на более поздних этапах развития. В дальнейшем, при
переходе мальков на тип питания, характерный для взрослых особей данного вида,
и в начале функционирования других рецепторных систем, участвующих в процессе
пище добывания, отношение рыбы к действию светового фактора изменяется (Иванов,
1988).
Особую роль играет свет в жизни ряпушки в период их полового созревания.
Фотопериодизм является одним из наиболее важных факторов окружающей среды,
обеспечивающих созревание половых продуктов у многих видов рыб (Потапова,
1978).
Для большинства видов сиговых рыб течение воды в реке является
стимулирующим фактором для созревания половых продуктов и при нересте. Вместе с
тем уровень воды в реке также имеет большое значение для этих рыб. Течение
облегчает скат рыбы, с другой определяет направление миграции. Это означает,
что чем сильнее течение, тем благоприятнее условия миграции с точки зрения
энергетических затрат рыбы и тем однозначное для нее направление перемещения
(Иванов, 1988).
Одним из наиболее перспективных холодноводных объектов аквакультуры
является сибирская ряпушка. Зрительная система этого вида пока не подвергалась
детальному изучению. Учитывая, однако, что ряпушка является типичным
планктофагом, можно предположить, что ей в определенной степени свойственны
закономерности, установленные для других планктофагов, которые остаются
наиболее чувствительными к свету на протяжении всего периода онтогенеза. Этих
рыб называют зрительными, так как все виды их активности связаны со светлым
периодом суток. Они обитают в верхних, наиболее освещенных слоях воды. По
сравнению с рыбами других видов планктофаги имеют наибольшую контрастную
чувствительность сетчатки глаза (Гирса, 1981). Известно, что у ранних личинок
сибирской ряпушки проявляется ярко выраженный положительный фототаксис и что
интенсивность потребления кислорода у них на свету возрастает (Шкорбатов,
1966).
ГЛАВА 3. УПРАВЛЕНИЕ ПОЛОВЫМИ ЦИКЛАМИ У СИБИРСКОЙ РЯПУШКИ
РАЗЛИЧНЫМИ МЕТОДАМИ
Методы стимулирования полового созревания производителей при
искусственном выращивании рыб получают все большее распространение в связи с
ростом масштабов рыбоводства и нарастающими потребностями населения в пищевой
продукции. Они позволяют сократить время выдерживания рыб до момента получения
зрелых половых продуктов, сдвигать сроки инкубации икры и увеличивать период
выращивания молоди в первый вегетационный сезон. Для получения качественных
половых продуктов необходимо учитывать биологические и физиологические
особенности разводимых видов рыб, в соответствии с которыми выбирается один из
трех разработанных учеными рыбоводами методов стимулирования: экологический,
физиологический, комбинированный.
Процесс созревания сиговых рыб во многом зависит от температурных и
гормональных факторов, соответственно, в рыбоводстве применяют экологическую
стимуляцию сиговых.
Сущность экологического метода заключается в следующем:
из естественной популяции или искусственно созданного маточного стада
отбираются здоровые рыбы, от которых планируется получить половые продукты в
ранние сроки;
отобранные рыбы размещаются в бассейны или садки (самки и самцы
раздельно);
в рыбоводных емкостях устанавливаются условия, близкие к нерестовым;
регулируются температурный режим, проточность и освещенность.
В этих условиях происходит интенсивное созревание половых продуктов и
рыбы нерестятся на 2-3 месяца раньше, чем в естественных условиях обитания
(Кучко, 2015).
Среди сиговых сибирская ряпушка - одна из выносливых к абиотическим факторам
среды вид, обладающий высокими потенциальными возможностями роста, отличающийся
скороспелостью.
Производителей сиговых рыб для рыбоводных целей отлавливают на
нерестилищах или на подходах к ним. В реках в безледный период на чистых
тоневых участках производителей отлавливают закидными неводами (Черняев, 1987).
В реках с большой скоростью течения или с заболоченными берегами
применяют различные типы ловушек (вентеря, моды и т. д.). Зрелых производителей
используют в первую очередь, рыб с недозревшими половыми продуктами оставляют в
садках для выдерживания. Сиговые хорошо переносят перевозку в живорыбных лодках
при температуре не выше 10°С. Возможность транспортировки производителей в
живорыбных лодках позволяет начинать их отлов на две недели раньше и дает
возможность заготовить наибольшее их количество.
Отлов производителей сиговых следует начинать при снижении температуры
воды в водоеме до 10 °С и заканчивать при наступлении ледостава. За указанный
период одна неводная бригада (6 чел.) 500-метровым неводом способна обловить
около 500 га глубоководных маточных озер и заготовить до 50 тыс.
производителей.
Поскольку нерест у сиговых очень растянут (более 3 мес.), производителей
до созревания половых продуктов выдерживают в садках. Высокую выживаемость производителей
можно обеспечить при создании благоприятных условий в садках (эластичность
стенок, достаточная глубина, хороший водообмен и благоприятный гидрохимический
режим).
Для размещения русловых садков следует использовать небольшие речки и
ручьи, связывающие озера. Ручей нужно расширить и углубить, превратив его в
канал, длина которого будет зависеть от необходимой площади русловых садков:
ширина - до 9м, глубина - до 1,5 м. На концах канала строят водонапорные
деревянные плотины с открытыми водосливами. Верхняя плотина должна обеспечивать
подъем уровня воды в головном озере до 1 м и необходимый водообмен в канале в
течение всего периода выдерживания. Нижняя плотина дает возможность
регулировать уровень воды в канале.
Садки длиной 30 м, шириной до 9 м и глубиной не менее 1 м отделяют друг
от друга водонепроницаемыми стенками из досок, обитых полиэтиленовой пленкой
или забетонированных, а дно засыпают песком. Отловленных производителей
выпускают в канал.
Шандоры между садками в это время открыты, и рыбы имеют возможность
свободно передвигаться по всему каналу. Когда температура воды в садках
понижается до 3-4°C, производители концентрируются в верхних садках (до 60
экз./м3). Для этого в водосливах садков устанавливают конусообразные
решетки. В вершине конуса решетки имеется вертикальная щель, через которую при
создании проточности 10-20 см/с сиговые поднимаются по течению и скапливаются в
верхних садках. После этого в водосливах устанавливают вертикальные решетки и
проводят осмотр производителей. Самцов отсаживают в нижние садки, а самок - в
верхние.
При последовательном водоснабжении поступление воды в верхний садок
должно быть не менее 20 л/с, при этом полный водообмен в садках происходит в
течение 4 ч. Содержание растворенного в воде кислорода в осенний период при
таком водообмене в верхнем садке составляет около 10 мг/л, а в нижнем падает до
8,2 мг/л, что вполне удовлетворяет потребности сиговых. При снижении
температуры воды в садках до 3°С проводят осмотр самок. Для облегчения работы с
рыбой каждый садок делят на 3 секции. В процессе проверки зрелых самок
переносят в рыбоводное помещение, где отцеживают икру, а незрелых помещают в
свободную секцию садка. В дальнейшем такие осмотры в зависимости от условий
проводят через 1-2 дня.
В начале рыбоводной кампании, когда температура воды высокая (8-12оС),
плотность посадки сиговых рыб в садки не должна превышать 20 кг/м3.
По мере снижения температуры (2-4оС) количество рыбы доводят до 75
кг/ м3.
Продолжительность выдерживания производителей зависит как от времени их
подхода к нерестилищам, так и от конкретных гидрологических условий. При
длительном выдерживании отход рыбы может составить 25-30 %.
Переборка самок в садках производится ежедневно. Недопустимо
передерживание созревших самок, так как это ведет к ухудшению качества икры. На
протяжении всего периода выдерживания из садков удаляют заболевших и
травмированных производителей. Целесообразно самок, выставленных в разные сроки
нерестового хода, т. е. с различным темпом созревания икры, помещать в разные
садки. Это поможет сократить количество просмотров садков и снизить тем самым
отход рыбы, получать в дальнейшем однородные поколения личинок (Козлов, 1998).
Для получения качественных половых продуктов вблизи мест отлова
производителей устраивают временные рыбоводные пункты. Текучих самок с мест
отлова или из садков доставляют в цех в емкостях с водой. Самки, у которых
произошла овуляция, и зрелые ооциты освободились от стромы гонад и находятся в
свободном состоянии в полости тела, имеют мягкое брюшко, при легком нажатии на
которое икра свободно вытекает ровной струей из полового отверстия (Черняев,
1987).
ГЛАВА 4. БИОЛОГИЧЕСКИЕ ОСНОВЫ КОРМЛЕНИЯ СИБИРСКОЙ РЯПУШКИ
Сиговые рыбы - довольно многочисленная группа объектов холодноводного
рыбоводства. Одним из видов относящихся к ним является сибирская ряпушка. Все
виды сиговых являются объектами воспроизводства и пастбищной аквакультуры, а
некоторые из них (в том числе гибриды - пелсиг, пелчир) и товарного
рыбоводства, которое стало интенсивно развиваться в последнее время (Зайцев,
1988).
Сибирская ряпушка обладает достаточно высоким темпом роста, могут
достигать товарной массы уже на втором году жизни. Оптимальная температура воды
для них составляет 15-17 градусов, однако они питаются и неплохо растут и при
более высокой температуре воды. Оптимальное содержание кислорода составляет 6-7
мг/л, хотя рыбы выдерживают понижение уровня растворенного кислорода до 2,5-3
мг/л.
Основу питания ряпушки составляют планктонные организмы, преимущественно
копеподы, но у крупных рыб спектр питания расширяется и включает крупных мизид,
бокоплавов, личинок хирономид, олигохет, икру и молодь рыб (Пономарев, 2012).
Характер питания сибирской ряпушки зависит от состояния кормовой базы
озер и рек, которые по уровню развития зоопланктона можно разбить на три
группы. В первой группе, где высока биомасса зоопланктона (более 2 г/м3),
а биомасса бентоса крайне мала, в питании преобладает рачковый плактон. Во
второй группе с высокой биомассой бентоса и низкой биомассой зоопланктона
питается смешанной пищей. Третья группа характеризовалась высоким уровнем как
зоопланктона, так и бентоса. Кроме того ряпушка питается остракодами, водяными
клещами, растительностью (остатки макрофитов).
По спектру питания сибирская ряпушка - эврифаг. Однако по характеру добывания
пищи ряпушка имеет свою специфику, что связано с ее поисковыми способностями.
Ряпушка, питаясь смешанной пищей, потребляет из зообентоса чаще всего формы,
мигрирующие в толщу воды (хирономиды, хаоборусы и др.), и поэтому по типу
питания больше относится к планктофагам.
Молодь ряпушки имеет широкий спектр питания. По значимости в питании
доминировали роды Bosmina, Daphnia. Среди бентосных организмов наиболее важную
роль в питании сеголетков играют личинки хирономид и поденок.
В процессе жизнедеятельности сиговые рыбы нуждаются в энергии, которую
они получают из корма. Установлено, что для рыб важными являются 10 незаменимых
аминокислот. Отсутствие или дефицит этих аминокислот в пище в течение первых
двух недель вызывают у рыбы потерю аппетита и снижение темпов роста, а в
дальнейшем приводят к возникновению болезней. Потребность в аминокислотах
меняется в зависимости от температуры воды. Так, если при температуре 10-12 %
белка корм для молоди сиговых должен содержать 40-42 % белка, то при 15 оС
- 52-55 %. Потребности в жирах у сиговых наилучшим образом удовлетворяются при
содержании в корме 0,5 % высших жирных кислот. Увеличение содержания жира в
рационе при постоянном содержании белка приводит к росту эффективности питания.
Рекомендуемый уровень содержания углеводов для сиговых 20-30 %, причем в пище
для молоди их должно находиться меньше, чем в пище для взрослой рыбы.
Личинки сибирской ряпушки имеют слаборазвитую систему пищеварения. Так в
возрасте до 20-30 суток у них отсутствует сформированный желудок, снижена
активность протеолитических ферментов. Использование типовых комбикормов в
ранний постэмбриональный период развития в виде монодиеты приводит к гибели
личинок, которые не способны усвоить высокомолекулярные белковые соединения,
свойственные подавляющему большинству кормовых компонентов животного и
растительного происхождения. В этой связи на первых этапах подращивания ранней
молоди сиговых рыб рацион их питания должен включать живые корма - науплии
артемии, мелкие формы зоопланктона. Одновременно должны вноситься искусственные
корма, обогащающие рацион питательными и биологически активными веществами, и
способствующие формированию у рыб адаптивной реакции и их интенсивному росту.
После завершения формирования дефинитивных органов пищеварения и продуцирования
эндопротеаз в необходимом количестве живые корма могут быть исключены, и рыбы в
возрасте свыше 1 месяца полностью переходят на питание комбикормами без ущерба
для роста и развития.
У личинок желточный мешок сохраняется до 30-40 дней. В течение этого
времени при низкой температуре личинок содержат без кормления. Затем, при
повышении температуры воды до 4-6°С, перед рассасыванием желточного мешка,
личинки переходят на активное питание живым кормом. В это время расход корма
составляет 20-30% массы. Наилучший рост личинок отмечен при кормлении их
смешанным кормом (молодь моин = науплии артемий). Выживаемость личинок - в
пределах 80-85%. В качестве добавочного возможно применять замороженный корм,
но не раньше чем спустя 7-10 дней после начала кормления.
Основным фактором, определяющим рост и развитие личинок, является
температура воды. Поэтому первое условие нормального роста личинок -
оптимальная температура воды (в пределах 10-15°C). Вторым условием,
положительно влияющим на рост личинок, является наличие живого корма. При
отсутствии живого корма временно можно использовать яичный порошок и сухие
дафнии. Сухой корм растирают в ступке с небольшим количеством воды. Корм дается
личинкам не менее 4-5 раз в сутки. При температуре воды 15-20° С применять яичный
порошок или сухие дафнии не рекомендуется: они сильно загрязняют воду и
ухудшают кислородный режим. Основной отход личинок при подращивании в заводских
условиях связан с загрязнением воды остатками корма и илом, поэтому вода должна
быть чистой, а количество корма - соответствовать оптимальной норме. Важно,
чтобы корм как можно дольше находился во взвешенном состоянии, тем самым
создаются оптимальные условия для его большей поедаемости. В 35-40-дневном
возрасте у сиговых рыб заканчивается личиночный и начинается мальковый период
развития. Отход за личиночный период достигает 30-40%, при подращивании же
личинок исключительно на живом корме отход снижается в 2-3 раза (Козлов, 1998).
При выращивании личинок сиговых рыб необходимо использовать сухие гранулированные
корма, отвечающие потребностям рыб в питательных веществах.
По завершении личиночного и наступлении малькового периода жизни в
возрасте 20-30 суток при массе тела 25-50 мг следует применять стандартные
комбикорма.
Стартовый комбикорм для личинок сибирской ряпушки отличается своеобразным
качественным составом. Его протеин имеет повышенную доступность (табл. 4).
С начала личиночного периода следует использовать комбикорм РГМ-СС на
протяжении 25-50 суток выращивания, затем применяют стартовый комбикорм РГМ-ПС.
Стартовый комбикорм для сибирской ряпушки производят в виде крупки (частиц
многоугольной формы). Размер крупки должен соответствовать массе выращиваемой
рыбы (табл. 5).
Таблица 4 - Стартовый комбикорм для личинок (РГМ-СС) и молоди (РГМ-ПС)
сибирской ряпушки, %
|
Компоненты
|
РГМ-СС
|
РГМ-ПС
|
|
Мука рыбная
|
19-30
|
31,5-50
|
|
Крилевая
|
0-10
|
0-10
|
|
Пшеничная
|
0-5
|
5-13
|
|
Водорослевая
|
-
|
1-2
|
|
Обрат сухой
|
5-8
|
5-10
|
|
Дрожжи этаноловые
|
0-50
|
5-15
|
|
Кормовой рыбный белок
(КРБ), гидролизат
|
8-10
|
10-15
|
|
Метионин
|
1,5
|
-
|
|
Жир рыбный
|
7
|
7-10
|
|
Премикс ПФ-1М
|
1,5
|
1,5
|
|
Протеин не менее
|
50
|
45
|
|
Жир
|
9
|
8
|
|
Минеральные вещества
|
12
|
13
|
|
Энергия, МДж/кг
|
12-13
|
11-12
|
Активность питания и активный пищевой рефлекс в начале кормления низкий.
Личинки захватывают частицы корма, находящиеся только непосредственной близости
к головной части. При массе тела 10-12 мг личинки плавают сформировавшейся
стаей, активность питания увеличивается. Частоту раздачи суточной нормы можно
уменьшить до 10-12 раз.
Периодичность кормления личинок и ранних мальков сибирской ряпушки -
через 0,5-1,0 ч в светлое время суток. Корм вручную или с помощью механических
кормораздатчиков разбрасывают по поверхности воды.
Таблица 5 - Размер крупки и гранул в зависимости от массы тела молоди
сибирской ряпушки
|
Масса молоди, г
|
Размер частиц корма, мм
|
Номер крупки и гранул
|
|
крупка
|
гранулы
|
|
|
до 0,02
|
0,1-0,2
|
-
|
1
|
|
0,02-02
|
0,2-0,4
|
-
|
2
|
|
0,2-1
|
0,4-0,6
|
-
|
3
|
|
1-3
|
0,6-1
|
-
|
4
|
|
3-7
|
1,1,5
|
-
|
5
|
|
7-10
|
1,5-2,5
|
-
|
6
|
|
10-20
|
-
|
3,2
|
7
|
|
более 20
|
-
|
4,5
|
8
|
По достижении малькового периода развития в возрасте 15-25 суток
отмечается максимальная активность питания и утилизация корма. Время
наступления этого периода зависит от температуры воды, условий выращивания и
питательности корма.
Кормление личинок, мальков и сеголеток следует производить по
определенным нормам в зависимости от массы тела и температуры воды (табл. 6).
Первые 10 дней пищевая реакция личинок еще низкая и потери комбикорма велики.
Суточную норму в этот период следует увеличить на вероятную величину потерь,
которая составляет до 30% раздаваемого корма.
Это избыточное кормление, компенсирующее потери, требует соответствующего
повышения затрат, однако эти затраты оправданы повышением скорости роста и
выживаемости молоди.
Для повышения эффективности кормления личинок, особенно в первые дни,
можно добавлять организмы зоопланктона в количестве 20% основного рациона
(науплии артемии салина, моины, босмины, коловратки) (Пономарев, 2013).
Таблица 6 - Суточная норма кормления личинок, мальков и сеголеток
сибирской ряпушки сухим гранулированным кормом, % к массе тела
|
Т, оС
|
Масса молоди, г
|
|
до 0,02
|
0,02-0,05
|
0,05-0,1
|
0,1-0,2
|
0,2-0,5
|
0,5-1,0
|
1-2
|
2-5
|
5-12
|
более 12
|
|
2
|
14,0
|
9,4
|
7,8
|
2,2
|
3,9
|
2,7
|
2,3
|
1,8
|
1,5
|
0,9
|
|
3
|
15,2
|
10,1
|
8,4
|
5,6
|
4,2
|
2,9
|
2,4
|
1,9
|
1,6
|
1,0
|
|
4
|
16,7
|
11,2
|
9,3
|
6,2
|
4,6
|
3,2
|
2,6
|
2,1
|
1,8
|
1,2
|
|
5
|
17,8
|
11,9
|
9,9
|
6,6
|
4,8
|
3,4
|
2,8
|
2,3
|
1,9
|
1,3
|
|
6
|
19,4
|
13,0
|
10,8
|
7,2
|
4,9
|
3,7
|
3,1
|
2,5
|
2,2
|
1,4
|
|
7
|
21,1
|
14,0
|
11,7
|
7,8
|
5,4
|
4,0
|
3,3
|
2,7
|
2,3
|
1,5
|
|
8
|
22,7
|
15,1
|
12,6
|
8,4
|
6,3
|
4,4
|
3,6
|
2,9
|
2,6
|
1,6
|
|
9
|
24,3
|
16,2
|
13,5
|
9,0
|
6,7
|
4,7
|
3,9
|
3,2
|
2,8
|
1,8
|
|
10
|
26,5
|
17,6
|
14,7
|
9,8
|
7,3
|
5,1
|
4,4
|
3,4
|
3,0
|
1,9
|
|
11
|
28,6
|
19,1
|
15,9
|
10,6
|
7,9
|
5,6
|
4,7
|
3,8
|
3,3
|
2,0
|
|
12
|
30,8
|
20,5
|
11,4
|
8,5
|
6,0
|
5,0
|
4,1
|
3,5
|
2,1
|
|
13
|
33,5
|
22,3
|
18,6
|
12,4
|
9,3
|
6,5
|
5,5
|
4,4
|
3,8
|
2,4
|
|
14
|
36,2
|
24,1
|
20,1
|
13,4
|
10,1
|
7,0
|
5,9
|
4,7
|
4,2
|
2,5
|
|
15
|
38,9
|
25,9
|
21,6
|
14,4
|
10,8
|
7,6
|
6,3
|
5,1
|
4,6
|
2,8
|
|
16
|
41,6
|
27,7
|
23,1
|
15,5
|
11,5
|
8,0
|
6,7
|
5,4
|
5,1
|
3,1
|
|
17
|
44,8
|
29,9
|
24,9
|
16,6
|
12,4
|
8,6
|
7,1
|
5,8
|
5,5
|
3,4
|
|
18
|
47,5
|
31,7
|
26,4
|
17,6
|
13,2
|
9,1
|
7,6
|
6,2
|
6,0
|
3,5
|
|
19
|
50,2
|
33,5
|
27,9
|
18,7
|
13,9
|
9,6
|
8,1
|
6,6
|
6,1
|
3,6
|
|
20
|
53,5
|
35,6
|
29,7
|
19,8
|
14,9
|
10,1
|
8,4
|
7,1
|
6,3
|
3,7
|
|
|
|
|
|
|
|
|
|
|
|
|
|
По завершению личиночного периода развития суточную норму снижают до
предусмотренной в кормовой таблице.
ГЛАВА 5. ТРАНСПОРТИРОВКА ИКРЫ, ЛИЧИНОК, МОЛОДИ И ВЗРОСЛЫХ
ОСОБЕЙ СИБИРСКОЙ РЯПУШКИ
Оплодотворенную икру транспортируют на начальных или конечных стадиях
развития, когда эмбрион менее всего чувствителен к механическим воздействиям
(Иванов, 1988).
Икру следует перевозить на рамках с туго натянутой марлей. На бортиках
рамок делают отверстия диаметром около 0,5 см для доступа воздуха к икре. На
каждую рамку кладут марлевую салфетку, на которой размешают икру в 1,5 - 2
слоя. Икру закрывают краями салфетки, затем марлей в 1 - 5 слоев, чтобы при
упаковке рамок в стопки икра на верхних рамках слегка прижимала нижние. Стопки
с пустыми нижней и верхней рамками перетягивают шпагатом и укладывают в
изотермические контейнеры. Поверх стопки рамок по бокам на дно контейнера
кладут увлажненную губку или другой подобный материал, препятствующий
перемещению рамок внутри контейнера при перевозке. Норма загрузки икры 7 тыс.
икринок.
Температура при перевозке должна быть постоянной и не должна превышать
4-5°С (Пономарев, 2012).
В сухих стерильных пробирках следует перевозить сперму, установленных в
термосе со льдом. Необходимо учитывать сроки ее активности. Длительность
оплодотворяющей способности спермы сиговых 2-3 суток при температуре в
контейнере равной 2 градусам. Хранению в перевозке подлежит свежеотобранная
сперма, помещенная в сухие пробирки отдельно для каждого самца с плотно
прижатыми пробками во избежание попадания воды. Пробирки с этикетками
заворачивают в марлю и опускают в термос, наполненный мелко наколотым льдом.
Для транспортировки личинок и молоди сиговых лучшим выбором будут
стандартные полиэтиленовые пакеты (рис. 3) объёмом 40 л, высотой 65 см и
шириной рукава 50 см. Перед перевозкой их упаковывают в стандартные картонные
коробки (65х35х35 см) и наливают 10-12 л воды, свободно пространство заполняют
кислородом, затем закрывают при помощи зажима Мора или резинового шланга.
Оптимальная температура для перевозки сиговых в летнее время составляет 6-8°С,
а весной и осенью -3-5° С. В один пакет помещают до 166 тыс. личинок сиговых
(Пономарев и др., 2006).
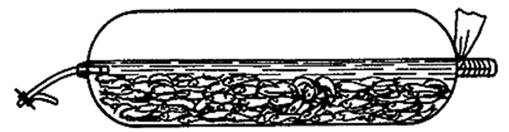
Рисунок 3 - Полиэтиленовый пакет для перевозки живой рыбы
При расчете плотности посадки личинок и молоди рыб в герметически
закрытый стандартный полиэтиленовый пакет, заполняемый водой и кислородом,
необходимо учитывать следующее:
степень растворения кислорода в воде в зависимости от механического
перемешивания в пути, температуры и парциального давления;
пороговое содержание кислорода для предличинок, личинок и молоди (1-3
мл/л);
потребление кислорода и выделение свободной углекислоты предличинками,
личинками и молодью в зависимости от температуры, их массы и видовой
принадлежности;
предельно допустимые концентрации углекислоты для предличинок, личинок и
молоди;
коэффициент растворения углекислоты в воде в зависимости от температуры;
фактор пространственного расположения предличинок, личинок и молоди;
длительность перевозки предличинок, личинок и молоди.
Перевозка личинок и молоди рыб в полиэтиленовых пакетах удобнее и
экономичнее, чем в других емкостях. Пакеты, доставленные к месту назначения,
помещают на некоторое время в водоем или заполненный водой бассейн, в который
будет выпущена рыба. После того как температура воды в пакетах и в водоеме
(бассейне) становится одинаковой, пакеты вскрывают и выпускают привезенный
материал.
На выживаемость перевозимой рыбы влияет содержание в воде кислорода,
накопление продуктов жизнедеятельности и свободное пространство. Для перевозки
живой рыбы необходимо использовать воду только из естественных водоемов. Очень
важно, чтобы перевозимая рыба не испытывала резких колебаний температуры. Так
же важно, чтобы перевозимая рыба была подготовлена к длительной перевозке. С
этой целью ее отсаживают в специальные бассейны с постоянным водообменом. Во
время выдерживания рыбу не кормят.
Для перевозки производителей следует использовать авто- или
авиатранспорт, применяют баки из оцинкованного железа длиной 140 см, шириной 60
см, высотой 125 см и вместимостью до 600 л. Баки наполняют на 2/3 объема речной
водой, а затем в каждый из них сажают по 3 - 5 производителей.
Продолжительность транспортировки на автомашинах не должна превышать 4 - 6
часов. При перевозке рыбы осенью, когда температура воды в баках около 2 - 5°С,
а содержание кислорода составляет 10 - 11 мг/л, воду в пути следования не
меняют. Плотность посадки производителей при транспортировке сибирской ряпушки
2 - 4 шт/м3 (Иванов, 1988).
Благодаря авиации масштабы транспортировки рыбы увеличиваются, и это
позволяет расширить возможности акклиматизации ценных видов рыб, в том числе и
сибирской ряпушки.
ГЛАВА 6. БИОЛОГИЧЕСКИЕ ОСНОВЫ АККЛИМАТИЗАЦИИ
В отличие от европейской ряпушки и ее крупной формы, рипуса, сибирская
ряпушка Coregonus sardinella (Valenciennes, 1948) для акклиматизации
использовалась в ограниченных объемах. Опытные работы выполнялись в Тюменской
области. Она завозилась из Норильских озер в оз. Большое Красноярского края
(1964-1969 гг.) и в Красноярское водохранилище (1968 г.). Результаты
акклиматизации не установлены (Кудерский, 2015).
Я планирую вселить сибирскую ряпушку в Новосибирское водохранилище.
Новосибирское водохранилище искусственный водоем на реке Обь возле города
Новосибирск (рис. 4). Водохранилище возникло после завершения строительства
плотины Новосибирской ГЭС в 1957-1959 годах. Находится водохранилище на
территории Новосибирской области и Алтайского края. На берегах водохранилища
расположены города: Бердск, Камень-на-Оби, Новосибирск. Площадь Новосибирского
водохранилища составляет - 1082 км², объем - 8,8 км3, длина -
около 200 км, наибольшая ширина - 22 км, наибольшая глубина - 25 м, зимой
замерзает (Васильев, 2000).
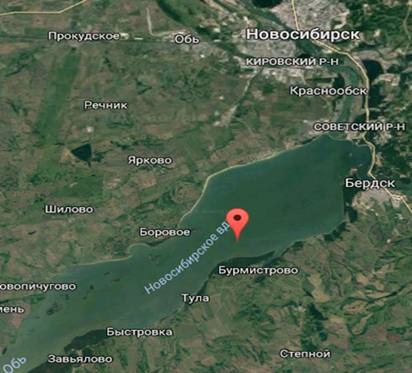
Рисунок 4 - Карта Новосибирского водохранилища
Температура воды в Новосибирском водохранилище в поверхностном слое на
всех участках в третьей декаде мая равняется в среднем 12,6 оС, в
июне - 17,0, в июле - 21,6, в августе - 19,6, в сентябре - 14,0. В октябре: от
9,5 до 3,1 - озерной части, от 13 до 2,0 - в речной и от 14 до 2,6 оС
- в среднем по всему водохранилищу (Оборемко, 1980).
Содержание растворенного в воде кислорода стабильное: максимальные
концентрации кислорода (9-12 мг/л) наблюдается осенью, что обусловлено
увеличением его растворимости при низких температурах, а также снижением его
расхода на дыхание гидробионтов. Минимальные концентрации кислорода в воде
(5,1-8,5 мг/л) наблюдаются зимой и связано это с нарушением газового обмена с
атмосферой при образовании ледяного покрова. Кислородный режим водохранилища
считается благоприятным и стабильным для жизнедеятельности гидробионтов.
Значения pH в открытый период в пределах 7,7-8,2, зимой- 7,4-7,5. Зимой
значение pH несколько меньше, чем в открытый период. Рост величин pH в
весенне-летний период объясняется усилением фотосинтетической деятельности
фитопланктона, что ведет к снижению содержания свободной углекислоты.
Содержание нитратов составляет: зимой - 0,23-0,29; весной - 0,13-0,15;
летом - 0,08-0,11; осенью - 0,06-0,11 мг N/л.
Зоопланктон насчитывает 16 видов кладоцер и 11 видов копепод. Летом на
всех участках водохранилища доминируют ветвистоусые ракообразные, которые
являются главными фильтраторами органоминеральной взвеси, весь осенне-зимний
период доминируют копеподы. В верхней зоне водохранилища биомасса зоопланктона
достигает 4,5 г/м3. В многоводные годы численность зоопланктона
составляет в летний период от 10 до 41, в маловодные - до 100 тыс. экз./м3
(при биомассе 4,3 г/м3). Из зообентоса в водохранилище
доминируют хирономиды, олигохеты, моллюски. Биомасса зообентоса составляет 10,3
г/м2 (Васильев, 2000).
В Новосибирском водохранилище обитает 1 вид круглоротых и 26 видов рыб.
Объектами рыбной ловли являются: судак, сазан, налим, линь, лещ, щука, плотва,
карась, окунь.
Основными конкурентами сибирской ряпушки в водохранилище являются
серебряный карась, лещ, пескарь.
Серебряный карась имеет длинный спинной плавник, крупную чешую. Тело
высокое, сжато с боков. Питается личинками насекомых и низшими водорослями.
Половой зрелости достигает в двух - четырехлетнем возрасте, при длине более 12
см.
Лещ держится группами, преимущественно в глубоких местах, поросших
растениями. Личинки питаются зоопланктоном. Мальки по достижении длины 30 мм
переходят на питание бентосом.
Пескарь имеет веретеновидное тело, чешуя крупная, рыло удлиненное, рот
нижний. Относится к бентофагам (http://www.sevin.ru/).
План работ по акклиматизации сибирской ряпушки:
. Выбор водоёма вселения, его изучение (Новосибирское водохранилище);
. Отлов производителей на путях миграции в конце осени (ноябрь);
. Получение личинок в Тобольском региональном рыбопитомнике;
. Транспортировка самолетом личинок сибирской ряпушки в полиэтиленовых
пакетах из Тобольского регионального рыбопитомника в Новосибирское
водохранилище;
. Вселение объекта: начало весны (март) при массе 5-6 мг.
Перед вселением сибирской ряпушки, необходимо выяснить, какой тип
акклиматизации подходит для данного объекта. Выделяют 5 типов акклиматизации:
) Акклиматизация внедрения - проводится при наличии относительно
свободной пищи, в которой переселенец занимает свое место, используя имеющиеся
в водоеме резервы корма и не вступает или почти не вступает в конкурентные
отношения с видами аборигенами.
) Замещения - предусматривает замещение малоценных аборигенов более
выгодными в хозяйственном отношении видами. Нужно, чтобы вселяемый объект был
достаточно ценным и конкурентоспособным, чтобы выселить из водоема виды
аборигены.
) Отторжения - это вселение в водоем нового вида, который по своей
жизнестойкости и конкурентоспособности не может противостоять аборигенам, но
его особи могут образовывать малочисленную популяцию которая займет
ограниченный ареал располагаясь на окраинах местного биоценоза.
) Пополнения - это пополнение переселенцем бедной ихтиофауны водоема,
который находится в изоляции.
) Акклиматизация конструирования - предусматривает целенаправленное
формирование кормовой базы и ценный промысел ихтиофауны в водоеме, который
только что образован или в котором произошли глубокие изменения в экосистеме
под влиянием климатического или антропогенного воздействия.
Рассматривая типы акклиматизации можно сделать вывод, что для сибирской
ряпушки подходит акклиматизация внедрения, так как имеет стабильная кормовая
база водоема, вследствие этого конкурентные отношения между вселяемым видом и
видами аборигенами будут минимальными.
Задачей акклиматизационных работ являются повышение продуктивности и
хозяйственной ценности водоемов, улучшение видового состава фауны, а также
сохранение и увеличение численности отдельных ценных видов водных организмов за
счет расширения ареала.
Различают пять основных понятий акклиматизации.
Интродукция - любое переселение особей вида в водоем, не освоенный ранее
ими. Интродукция всегда является первым этапом процесса акклиматизации, но не
всегда интродукция заканчивается акклиматизацией интродуцента.
Акклиматизация - процесс приспособления интродуцированных особей вида к
новым условиям среды, а также формирование в этих условиях новой популяции.
Вселение - переселение особей вида в водоем, условия среды в котором мало
или совершенно не отличаются от условий жизни данного вида в материнском
водоеме. Вселенные особи вида успешно размножаются на новом месте обитания без
какой-либо предварительной внутренней перестройки организма. Биологические
особенности потомства переселенных особей вида не изменяются.
Зарыбление - это регулярный выпуск молоди одного и того же вида на нагул
в апробированные водоемы.
Натурализация - конечный высший этап акклиматизации, когда определились
ареал вида в новом водоеме, его взаимоотношения со средой и возможность
использования вселенца.
На данный момент существует 4 метода акклиматизации: пассивный, активный,
радикальной акклиматизации, ступенчатой акклиматизации (Пономарева, 2013).
Для акклиматизации сибирской ряпушки наиболее приемлем пассивный метод,
сущность которого заключается в том, что человек осуществляет лишь выбор и
перенос объекта акклиматизации в новый водоем. Процесс акклиматизации переселенца
проходит без вмешательства человека и зависит от природы интродуцента.
Фазы процессов акклиматизации переселенца
Б.Г. Иоганзен выделяет 3 фазы в соответствии с изменением численности
акклиматизантов: 1) малой численности, когда происходит физиологическая и
биологическая адаптация особей к новым условиям; 2) быстровозрастающей и
высокой численности популяции; 3) колеблющейся численность переселенцев
изменяется в соответствии с новыми условиями среды.
А.Ф. Карпевич выделяет пять условных фаз процесса акклиматизации и
натурализации вида в новых условиях:
I фаза - выживание переселенных особей в новых для них условиях.фаза -
размножение и начало формирование популяции.фаза - максимальная численность
переселенца (фаза взрыва).фаза - обострение противоречий переселенца с
биотической средой.фаза - натурализация в новых условиях (Пономарева, 2013).
Рассчитываем промысловый эффект по фазам Карпевича А.Ф. (1975):
Первая фаза от момента вселения особей до появление потомства, 6 лет.
Вторая фаза, размножение и формирование популяции, 4 года. Третья фаза,
максимальная численность переселенца, 4 года. Четвертая фаза, обострение
противоречий переселенца с биотической средой, 2 года. Пятая фаза,
натурализация вида, 16 лет.
При перевозке необходимо поддерживать благоприятную температуру воды 3-7 0С.
Выпуск посадочного материала в водохранилище будет производится в хорошо
аэрированные места.
Нерест сибирской ряпушки происходит на глубинах от 25 до 65 м на
заиленных грунтах, что особенно перспективно, так как аборигенные виды
Новосибирского водохранилища сосредоточены в основном на глубинах от 2 до 15 м.
Таким образом, сибирская ряпушка займет эту экологическую нишу водохранилища.
Обилие кормовой базы в Новосибирском водохранилище позволяет сибирской ряпушке
натурализоваться и сформировать свою популяцию в условиях конкуренции с другими
рыбами. Сибирская ряпушка имеет большое промысловое значение, в связи с
вкусовыми качествами мяса.
БИБЛИОГРАФИЧЕСКИЙ СПИСОК
1. Берг Л.С.
Рыбы пресных вод СССР и сопредельных стран / Л. С. Берг. - М.: Академия наук
СССР, 1948. - 466 с.
. Богданов
В.Д. Морфологические особенности развития и определитель личинок сиговых рыб р.
Оби / В. Д. Богданов. - Екатеринбург: УрО РАН, 1998. -56 с.
. Богданов В.
Д. Эмбриональное развитие сиговых рыб на естественных нерестилищах в уральских
притоках Нижней Оби / В. Д. Богданов. Екатеринбург: Институт экологии растений
и животных УрО РАН, 2006. - 14 с.
. Валюшок Д.
С. Качественный состав и локализация пигментов в икре сиговых рыб / Д. С.
Валюшок. Тольятти: Всесоюз. совещ. по лососевидным рыбам, 1988. -С. 46-48.
. Васильев О.
Ф. Экологическое состояние Новосибирского водохранилища / О. Ф. Васильев, В. М.
Савкин, С. Я. Двуреченская, С. Я. Тарасенко, П. А. Попов, А. Ш. Хабидов. -
Новосибирск: СО РАН, 2000. - С. 149-163.
. Гирса И. И.
Освещенность и поведение рыб / И. И. Гирса. М.: Наука, 1981. - 167 с.
. Зайцев В.
Ф. Корма и их использование в рыбоводстве / В. Ф. Зайцев. - Владивосток:
Дальнерыбтуз АТИРПИХ, 1988. - 79 с.
. Иванов,
А.П. Рыбоводство в естественных водоемах / А.П. Иванов. -М.: Агропромиздат,
1988. - 367 с.
. Карпевич,
А.Ф. Теория и практика акклиматизации водных организмов / А.Ф. Карпевич. - М.:
Пищевая промышленность, 1975. - 432 с.
. Козлов В.
И. Справочник фермера-рыбовода / В. И. Козлов. -М.: Изд-во ВНИРО, 1998. - 447
с.
. Коровина
В.М. Влияние видимого света и темноты на скорость образования хрящевых
элементов скелта костистых рыб / В. М. Коровина, А. И. Любицкая, Е. А.
Дорофеева. Вопросы ихтиологии. - 1965. - С. 403-410.
. Кудерский,
Л.А. Акклиматизация рыб водоемах России / Л.А. Кудерский // Сборник научных
трудов ФГБНУ «ГосНИОРХ». - Санкт- Петербург, 2015. - 285 с.
. Кучко Т. Ю.
Методы получения половых продуктов от производителей рыб / Т. Ю. Кучко. -
Петрозаводск : Изд-во ПетрГУ, 2015. - 63 с.
. Лебедева О.
А. Температурные адаптации сиговых в период эмбрионального развития / О. А.
Лебедева. Сб. тр. ГосНИОРХ. - 2006. -С. 56-78.
. Литвиненко
А. И. Искусственное воспроизводство ценных видов рыб Урала и Сибири: состояние,
проблемы и перспективы / А. И. Литвиненко, С. М. Семеченко, Я. А. Капустина //
Труды ВНИРО. Том 153. Аквакультура. -М.: ВНИРО, 2015. - С. 77-78.
. Макеева А.
П. Атлас молоди пресноводных рыб России / А. П. Макеева, Д. С. Павлов, Д. А.
Павлов. - М.: Товарищество научных изданий КМК, 2011. - 383 с.
. Мешков М.
М. О некоторых закономерностях перехода от эмбрионального состояния к
личиночному / М. М. Мешков. М.: Теоретические основы рыбоводства, 1965. -С.
165-167.
. Микулин А.
Е. Закономерности динамики изменения каротиноидных пигментов в процессе
эмбрионального развития костистых рыб / А. Е. Микулин, Л. В. Котик, В. Н.
Дубрович. Научн. докл. высш. шк. Биол. науки. №9. - С. 31-37.
. Оберемко Ф.
А. Гидрология, гидрохимия Новосибирского водохранилища и его охрана / Ф. А.
Оберемко, Ю. И. Подлипский // Проблемы использования и охраны природных
ресурсов Сибири. - Новосибирск, 1980. - 353 с.
. Пономарев
С. В. Аквакультура / С. В. Пономарев, Е. Н. Пономарева. -Астрахань: Нова, 2002.
- 264 с.
. Пономарев,
С.В. Индустриальное рыбоводство / С.В. Пономарев, Ю.Н. Грозеску, А.А. Бахарева.
- М.: Колос, 2006. - 320 с.
. Пономарев,
С.В. Лососеводство / С.В. Пономарев. - М.: МОРКНИГА, 2012. - 561 с.
. Пономарев,
С.В. Корма и кормление рыб в аквакультуре / С.В. Пономарев, Ю.Н. Грозеску, А.А.
Бахарева. - М.:МОРКНИГА, 2013. - 417 с.
. Пономарева,
Елена Николаевна. Конспект лекций по дисциплине «Биологические основы
рыбоводства» для студентов очной и заочной форм обучения специальности 311700 /
АГТУ. - Астрахань, 2004. - 80 с.
. Пономарева,
Елена Николаевна. Методические указания к лабораторной работе по курсу
«Биологические основы рыбоводства» на тему «Основы акклиматизации гидробионтов.
Методы, способы и оценка результатов акклиматизации» для студентов
специальности 311700 «Водные биоресурсы и аквакультура» / Е.Н. Пономарева. -
Астрахань: АГТУ, 2013. - 16 с.
. Потапова О.
И. Крупная ряпушка / О. И. Потапова. - М.: Агропромиздат, 1978. - 114 с.
. Привезенцев
Ю. А. Рыбоводство / Ю. А. Привезенцев, В. А. Власов. - М: Мир, 2004. - 456 с.
. Решетников
Ю. С. Экология и систематика сиговых рыб / Ю. С. Решетников. -М.: Наука, 1980.
- 300 с.
. Саковская
В. Г. Практикум по прудовому рыбоводству / В. Г. Саковская, З. П. Ворошилина,
В. С. Сыров, Е. И. Хрусталев. - М.: Агропромиздат, 1991. - 174 с.
. Семеченко
С.М. Состояние запасов и искусственного воспроизводства ценных видов рыб
Сибири. Внутренние водоемы. Сырьевые ресурсы / С. М. Семеченко, Н.В. Янкова.
ФГУП "Госрыбцентр" Журнал "Рыбное хозяйство", 2006. - С.
46-49.
. Сергиенко
Л. Л. Сибирская ряпушка как объект рыбоводства / Л. Л. Сергиенко. - Тюмень:
Вестник рыбохозяйственной науки. Т. 2, 2015. - 9 с.
. Смешливая,
Наталья Владимировна Эколого-физиологические аспекты репродуктивной функции
сиговых рыб Обь-Иртышского бассейна: автореферат дис. канд. биологических наук
/ Н. В. Смешливая. - Калининград, 2015. - 23 с.
. Сытина Л.
А. Периодизация развития осетровых и проблема изменчивости организмов / Л. А.
Сытина, О. Б. Тимофеев. Вопр. ихтиологии, Т. 13, вып. 2, 1973. - С. 275-291.
. Черняев Ж.
А. Эмбриональное развитие озерного сига в связи с вопросом его искусственного
разведения / Ж. А. Черняев. Вопр. ихтиологии, Т. 13, вып. 2, 1973. - С.
252-274.
. Черняев Ж.
А. Методические указания по сбору и хранению икры сиговых рыб на временных
рыбоводных пунктах, ее транспортировке и инкубации / Ж. А. Черняев. - Москва,
1987. - 82 с.
. Шестаков,
Александр Викторович Биология молоди сиговых рыб бассейна р. Анадырь:
автореферат дис. канд. биологических наук / А. В. Шестаков. - Владивосток,
1996. - 114 с.
. Шкорбатов
Г. Л. Избираемая температура и фототаксис личинок сигов / Г. Л. Шкорбатов.
Зоол. журн. - 1966. - С. 1515-1525.
.
www.pravo.gov.ru/
39.
www.sevin.ru/
40.
www.aquacultura.org/