Анатомия коллембол
Содержание
Введение
Глава 1. Обзор литературы
.1 Экология коллембол
.1.1 Жизненные формы
.1.2 Питание
.1.3 Жизнь в экстремальных условиях
.2 Морфология коллембол
.2.1 Внешнее строение
.2.2 Внутреннее строение
.2.2.1 Эндоскелет
.2.2.2 Мышечная система
.2.2.3 Пищеварительная система
.2.2.4 Выделительная система
.2.2.5 Дыхательная система
.2.2.6 Кровеносная система
.2.2.7 Нервная система, органы чувств
.2.2.8 Половая система
.3 Миниатюризация животных
.3.1 Эффект миниатюризации у насекомых
.4 Систематическое положение и признаки рода
Mesaphorura
Глава 2. Материалы и методы Глава 3. Результаты
.1 Морфология Mesaphorura
.1.1 Внешнее строение
.1.2 Внутреннее строение
.1.2.1 Эндоскелет
.1.2.2 Мышечная система
.1.2.3 Пищеварительная система
.1.2.4 Нервная система
.1.2.5 Половая система
.1.2.6 Прочие системы
Глава 4. Обсуждение
.1 Эффект миниатюризации у коллембол
.1.1 Сравнение Mesaphorura и ранее изученных
представителей данного отряда
.1.2 Сравнение эффектов миниатюризации коллембол и
насекомых
.1.2.1 Сравнение объёмов органов Mesaphorura и некоторых
насекомых
Выводы
Список литературы
Введение
Повышенный интерес к эффектам миниатюризации проявлялся к позвоночным и
беспозвоночным животным (Hanken J., Wake D.B, 1993), а в последние годы было
сделано много работ по морфологии миниатюрных насекомых (Polilov, 2015). Отряд
Collembola является базальной группой шестиногих (Hexapoda) и включает в себя
небольших представителей (1-3 мм в среднем) (Beutel et al., 2013), одними из
самых маленьких считаются, например, Megalothorax minimus (0,35-0,55 мм)
(Schneider, D’Haese, 2013), Mackenziella psocoides (0,25 мм) (Hopkin, 1997),
что представляет собой интерес не только в исследовании проявления
миниатюризации, но и в их сравнении с насекомыми. Работы предыдущих авторов не
содержат детальных анатомических описаний или щатрагивали только отдельные
системы органов (Lubbock, 1877; Folsom, 1899; Imms, 1939; Bretfeld, 1963;
Wolter, 1963; Shaller,
; Eisenbeis, 1975; Verhoef, 1979; Hopkin, 1997) и все эти работы
посвящены крупным коллемболам.
Целью работы стало изучение внутреннего строения коллембол рода
Mesaphorura.
Были поставлены следующие задачи:
1. Изучить анатомию Mesaphorura sylvatica (Rusek, 1971) с помощью
оптической микроскопии и трёхмерного компьютерного моделирования.
2. Выявить особенности внутреннего строения, связанные с
уменьшением размеров тела.
3. Сравнить особенности строения коллембол, связанные с
миниатюризацией, с данными по миниатюрным насекомым.
Автор выражает благодарность своему научному руководителю Кузнецовой
Наталии Александровне (МПГУ) за помощь в проведении практической части и советы
по написанию данной работы; научному консультанту Полилову Алексею Алексеевичу
(МГУ) за курирование работы; а также научному сотруднику Потапову Михаилу
Борисовичу (МПГУ) за помощь в сборе и определении объектов.
Глава 1. Обзор литературы
1.1 Экология
1.1.1 Жизненные формы
Коллемболы могут быть разделены на жизненные формы, которые отличаются
друг от друга занимаемым почвенным горизонтом и имеют характерные морфологические
особенности. Определение жизненных форм находит применение в сравнительном
анализе экологической структуры сообществ, а также является важным индикатором
состояния почвы. Можно выделить три основных жизненных формы коллембол:
1) эпидафические (поверхностные и
верхнеподстилочные) формы - обитатели поверхности подстилки и верхнего
горизонта подстилки;
2) гемиэдафические (полупочвенные) формы - обитатели нижних слоёв
подстилки и верхнего слоя почвы;
3) эуэдафические (почвенные формы) - обитатели почвенного горизонта
(Karaban et al., 2012).
Стебаева С.К. выделяет также четвёртую форму - специализированные
обитатели пещер, муравейников и термитников (Бабенко и др., 1988).
От эпидафической к эуэдафической форме уменьшается размер тела, частично
или полностью редуцируются пигментация, прыгательная вилка и количество
омматидиев. Однако вертикальное распространение жизненных форм может сильно
варьировать. Существуют два метода определения жизненной формы: по глубине
обитания в почве и по морфологическим признакам, результаты последнего
считается более приближенными к реальному распределению (Karaban et al., 2012).
Вертикальная стратификация коллембол в почве коррелирует с пористостью
почвы, относительной влажностью и распределением пищи. В подзолистых почвах
подстилка медленно перемещается вниз по мере её разложения, и таким образом по
вертикальному расположению отдельных таксонов можно определить фазу разложения
(Rusek, 1988).
1.1.2 Питание
Коллемболы специализируются на питании почвенной микробиоты (грибы, бактерии,
актиномицеты, водоросли), контролируя динамику популяций данных организмов.
Кроме этого, они поглощают мёртвую органику, включая подстилку. Некоторые виды
питаются листьями (исключая жилки), другие же употребляют в пищу частично
разложившимся органическим веществом или экскрементами иной почвенной фауны.
Многие атмобионты (обитатели поверхности рыхлых подстилок) являются фитофагами
и питаются тканями живых растений (например, Sminthurus viridis, Bourletiella
spp.), другие питаются гифами грибов и спорами, а также меняют предпочтения в
еде в течение года (пыльца, водоросли, ткани растений) (Rusek, 1988).
насекомое экстремальный миниатюризация орган
1.1.3 Жизнь в экстремальных условиях
Коллемболы играют определяющую роль в формировании почвы в некоторых арктических,
альпийских регионах и в слабо развитых почвах, находящихся на ранней стадии
сукцессии. В более развитых почвах они участвуют в разложении листового опада и
вторичном расщеплении экскрементов мега- и макрофауны. Самая простая форма
гумуса состоит преимущественно из экскрементов коллембол (таких как из рода
Xenylla); более сложная уже включает в себя, в основном, помёт мегафауны, хотя
по- прежнему экскременты коллембол являются доминирующими элементами почвенного
матрикса данного гумуса. Всё это является важным для формирующих процессов
почвы (Rusek, 1988).
По мере ухудшения климатических условий (в частности, уменьшение
температуры) коллемболы играют всё большую роль в почве. Например, в Антарктике
число коллембол достигает 86% от общей численности шестиногих. Чтобы выживать в
подобных экстремальных условиях, альпийские, арктические и антарктические виды
ногохвосток приспосабливаются к переохлаждению организма. Так, особи
Tetracanthella wahlgreni способны выдерживать понижение температуры до -39C.
При этом холодостойкость уменьшается во время питания, что было показано ещё на
насекомых (Somme, 1981).
Выделяют две категории организмов в зависимости от механизмов адаптации к
холоду: толерантные и не толерантные к замораживанию. Показано, что коллемболы
относятся ко второй категории. Выживанию в подобных условиях им помогает
наличие криопротекторов в гемолимфе (глицерол, сорбитол, маннитол, глюкоза,
трегалоза и др.), а также сокращение объёма воды. Подготовка организма к
повышенной холодостойкости происходит осенью и ранней зимой и происходит в два
этапа: 1) опустошение пищеварительной системы 2) усиление сопротивления к
холоду благодаря накоплению глицерола или иных криопротекторов (Somme, 1981;
Worland, 1996).
1.2 Морфология коллембол
1.2.1 Внешнее строение
Тело коллембол разделено на голову, грудь (3 сегмента) и брюшко (6
сегментов). Крылья и сложные глаза отсутствуют, кутикула частично редуцирована.
Имеется ряд специализированных органов: вентральная трубка (на первом сегменте
брюшка), зацепка (на третьем сегменте брюшка), прыгательная вилка (на последнем
сегменте брюшка).
1.2.2 Внутреннее строение
1.2.2.1 Эндоскелет
Эндоскелет - совокупность впячиваний покровов, которые служат для
крепления мышц и поддержки некоторых органов.
Основным эндоскелетом головы насекомого является тенториум (Imms, 1977).
Но, в отличие от них, у скрыточелюстных (Entognatha) подобная структура
редуцируется. У коллембол эндоскелет головы развит наиболее сильно, тем самым
сильно напоминая тенториум насекомых, однако состоит он не только из впячиваний
экзоскелета, но и из коллагеновых структур (Wolter, 1963; Koch, 2000; Blanke
and Machida, 2015). Чтобы избежать путаницы, различные авторы давали ей иные
термины (архитенториум, псевдотенториум). Было показано, что она не может быть
гомологична тенториуму насекомых, но также и не является неким исходным
образованием, от которого мог бы произойти тенториум (Matsuda, 1965; Wolter,
1963). Руки Фолсома (также “lingual stalks”) считаются гомологичными передним
тенториальным аподемам насекомых, а fulcra (ед. fulcrum), или fulturae, или
sternum, или sternal arms - задним тенториальным аподемам. Они связаны друг с
другом системой сухожилий с которыми и образуют сложный комплекс, названный
псевдотенториумом. Он также имеет свои “руки” - выросты сухожилий.
Псевдотенториум крепится к экзоскелету при помощи тенториальных аподем,
сухожилий и особых подвешивающих мышц. Его вид сильно варьируется у различных
видов (Manton and Harding, 1964; Bitsch and Bitsch, 2002).
Эндоскелет груди не менее сложен и может быть поделён на эндотергиты,
эндоплевриты и эндостерниты. Эндотергиты возникают как складки межсегментных
склеритов и служат для увеличения площади поверхности крепления дорсальных
продольных мышц. Эндоплевриты образуются между плевритами. Эндостерниты
насекомых возникают либо как парные апофизы (фурки), либо как непарные аподемы
(спина (spina) на межсегментном склерите - спинастернуме). У коллембол
наблюдаются похожие эндостерниты, но по аналогии со структурами насекомых они
не называются (Imms, 1977; Bitsch and Bitsch, 2002). Вероятно, структуры,
внешне напоминающие таковые фурки у насекомых, у коллембол больше схожи со
спинами по происхождению (Imms, 1977).
В брюшке эндоскелет развит слабее. Помимо типичных аподем (Рис.1) для
крепления брюшных мышц, могут быть развиты специализированные аподемы,
связанные с половой системой (Imms, 1977).
1.2.2.2 Мышечная система
Все мышцы можно разделить на три большие категории: скелетные мышцы,
висцеральные и мышцы сердца. Скелетные мышцы прикрепляются к кутикуле и
перемещают одни части тела относительно других. Висцеральные мышцы связаны с
внутренними органами, например, имеются круговые мышцы вокруг кишечника и
половых протоков (Chapman, 1998).
При описании головы основной акцент делается на мышцах челюстного
аппарата. В голове коллембол выделяется 25 пар челюстных мышц. 10 из них
связаны с максиллами, 10 - с мандибулами, 5 - с лабиумом. В зависимости от
выполняемого действия делятся на: абдуктор, который выполняет отведение,
аддуктор - приведение, вращатель - вращение, ректрактор - оттягивание назад,
протрузор - вытягивание вперёд (Folsom, 1899). Антенны имеют пар 9 мышц,
которые крепятся к различным аподемам внутри членико. (Imms, 1939), От начала
груди до конца брюшка проходят три пары продольных мышц: две дорсальных и
вентральная (Bretfeld, 1963). Они отвечают за сближение сегментов при
сокращении (Шванвич, 1949).
В груди количество мышц различается в зависимости от вида. У двух
изученных представителей (Neanura muscorum, Orchesella cincta) насчитывается
23-49 пар. В брюшке - 20-53 пары. Они могут быть поделены на четыре большие
группы:
1) сегментные дорсо-вентральные мышцы, которые располагаются в границах
одного сегмента;
2) латеральные дорсо-вентральные мышцы, которые находятся сбоку от
предыдущих и в груди часто связаны с конечностями;
) межсегментные дорсо-вентральные мышцы, которые проходят через
два сегмента;
) поперечные мышцы, лежащие на вентральной стороне тела.
От места прикрепления мышцы к конечности зависит её функция: сгибание,
разгибание, отведение, приведение. Мускулатура переднегруди заметно отличается
от мускулатуры остальных сегментов (Шванвич, 1949; Bretfeld, 1963). В брюшке
наличие специализированной мускулатуры связано с вентральной трубкой, которая
насчитывает 14 пар мышц (Eisenbeis, 1975), а также зацепкой и прыгательной
вилкой, если таковые имеются.
1.2.2.3 Пищеварительная система
Пищеварительная система типичного строения и состоит из передней, средней
и задней кишок. Передняя и задняя кишки - производные инвагинации эктодермы и
выстланы кутикулой. Средняя кишка имеет энтодермальное происхождение.
Передняя кишка: pharynx (глотка), esophagus (пищевод), crop (зоб).
Передняя кишка представляет собой длинную изогнутую трубку, переход между
глоткой и пищеводом выражен слабо. Спереди глотка связана с ротовой полостью, а
пищевод сзади отгорожен от средней кишки с помощью клапана.
Средняя кишка: ventriculus (желудок), midgut (собственно средняя кишка).
Задняя кишка: pylorus, ileum, rectum/caecum (Lubbock, 1873).
Наиболее полное деление встречается только в одной работе и приведено в
виде цитат более ранних авторов. В остальных работах упоминания о ряде отделов
не встречаются. Отсутствие ясных границ и принципиальной разницы между
некоторыми из них (например, пилорического отдела и тонкой кишки) приводит к
невозможности достоверного разделения и возможных несовпадений различных
мнений.
Глотка формирует четыре кармана, по два с каждой стороны, путём
выпячивания. В дорсальной паре карманов расположены мандибулы, в вентральной -
максиллы (Рис.2). Нижняя часть глотки образована дорсальной поверхностью
лабиума, а верхняя - лабрумом (Folsom, 1899). В неё впадают протоки двух пар
слюнных желёз, не всегда развитых. У представителей с колюще-сосущим ротовым
аппаратом они достигают больших размеров (заходят в первый сегмент груди), чем
у видов с грызущим типом ротового аппарата. (Wolter, 1963; Shaller, 1970).
Пищевод имеет собственные мышцы-расширители, подходящие к одной круговой
мышце, расположенной по всей длине передней кишки. Он начинается небольшим
расширением в передней части глотки и тянется до среднегруди в виде тонкой
трубки единого диаметра, где заканчивается клапаном (Folsom, 1899; Wolter,
1963).
Средняя кишка представляет собой простую расширенную трубку без
каких-либо придатков за исключением редко встречающихся мешковидных образований
(дивертикул; “слепых кишок”), которые связаны с сосущим типом питания. Описаны
у Protanura (в количестве двух), Neanura muscorum, Pseudachorutes (Wolter,
1963; Shaller, 1970).
Задняя кишка представляет собой тонкую короткую трубку, которая
заканчивается анальным отверстием. Ректум, последний отдел задней кишки, не
имеет дифференцировки, но, тем не менее, как и вентральная трубка принимает
важную роль в осморегуляции и в регуляции ионного обмена (Verhoef, 1979).
Во время линьки обновляются все три отдела. Обновление средней кишки, как
и период линьки, подвергается циклическим изменениям, что позволяет ей
выполнять выделительную функцию взамен утраченных мальпигиевых сосудов
(Humbert, 1979).
1.2.2.4 Выделительная система
Мальпигиевы сосуды отсутствуют, за редким случаем на их месте могут
находиться небольшие бугорки (Shaller, 1970). В ранних источниках ещё имеются
упоминания о мальпигиевых сосудах в количестве до 3 пар (Lubbock, 1873). Но,
скорее всего, речь идёт о тех же рудиментах. Структуры с выделительной функцией
у коллембол представлены в большом разнообразии:
1) лабиальные нефридии (Рис. 3),
2) ацинарные, или гроздевидные, железы,
) 3 пары клеточных скоплений, или ‘рудиментарные нефридии’ (в
голове, в 1 и 3 сегментах абдомена),
) средняя кишка,
) жировое тело.
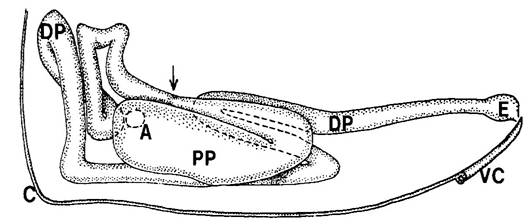
Рис. 1. Лабиальный нефридий (O.cinta). Отверстие указывает на соединение
с мешочком (не нарисован). С - кутикула, DP - дистальная часть, PP -
проксимальная часть, E - концевой мешок, VC - вентральный канал (из Verhoef,
1979).
Парные лабиальные нефридии лежат по бокам от нервной системы и кишечника
и доходят до переднегруди, почти достигают дорсальной стороны головы (Wolter,
1963). Они состоят из двух частей: целомического мешочка (“саккулюс”),
выстланного подоцитами, и двудольного канала нефридия, имеющего проксимальную
(лабиринт) и дистальную части. Стенки проксимального канала кардинально меняют
свой состав в зависимости от стадии линьки, их клетки, определённые как лизосомы,
принимают участие в фагоцитозе. Дистальный канал выстлан базальным лабиринтом,
который участвует в активном всасывании воды и ионов. Дистальные части обоих
канальцев нефридия соединяются и впадают единым протоком на вентральной стороне
лабиума. Клетки мешочка выстланы подоцитами, он участвует в фильтрации
гемолимфы и формировании первичной мочи, а за её реабсорбцию отвечает
нефридиальный канал. Тем не менее, до конца не понятно, происходит ли секреция
конечных веществ клетками лабиринта и мешочка или же имеет место гетерофагиии и
автофагии (Altner, 1968; Verhoef, 1979).
Ацинарные железы парные, расположены в вентральной части головы по бокам
от центральной линии. Состоят из множества одноклеточных желёз, чьи протоки
сливаются в единый канал, который открывается наружу позади протока лабиальных
нефридиев (Wolter, 1963).
Клеточные скопления расположены во всех отделах тела. Они имеют родство с
подоцитами, лишены выводных протоков и выполняют функцию “почек накопления”.
Те, что находятся в голове у основания антенн также называют антеннальными
нефридиями, или головными почками (Shaller, 1970). Структуры в первом сегменте
брюшка находятся по бокам от вентральной трубки, в третьем - по бокам от
зацепки. Их состав изучен слабо, но показано отсутствие кристаллов мочевой
кислоты (Humbert, 1975; Verhoef, 1979).
Выделительная функция также выполняется при помощи средней кишки, которая
обновляется при линьке, как это уже было упомянуто ранее. Таким образом из
организма удаляются токсические элементы, совершается минеральный обмен. Не
исключено, что вне линьки некоторые процессы регенерации также участвуют в
выделении (Humbert, 1979).
Не менее важную роль играют клетки жирового тела, которые, в отличие от
клеточных структур, накапливают кристаллы мочевой кислоты (Шванвич, 1949).
1.2.2.5 Дыхательная система
Трахеи, как правило, отсутствуют, встречаются лишь у нескольких видов.
Основное дыхание происходит за счёт диффузии газов через кутикулу, которая
лишена сплошного слоя эпикутикулы. Около трети необходимого для жизнедеятельности
кислорода поступает через поверхность вентральной трубки (Shaller, 1970).
Трахейная система состоит из двух частей, которые между собой не связаны.
Каждая открывается единственным дыхальцем в передней части переднегруди.
Механизмы, закрывающие дыхальца, отсутствуют. Благодаря тому, что дыхальце с
каждой стороны всего одно, воздух поступает в трахейную систему постепенно, в
отличие от насекомых, у которых при наличии нескольких дыхалец воздух быстро
заполняет главный ствол и оттуда перемещается далее по ответвлениям. Трахейные
стволы расходятся и ветвятся в голове, в груди, в брюшке, не заходя в области
расположения зацепки, анального отверстия и вентральной трубки. Тенидии
(спиральные утолщения) встречаются во всех ответвлениях диаметром до 1,5 мкм.
Трахеолы достигают диаметра в 0,25 мкм (Davies, 1927).
Кутикула, обладая гидрофобностью, вместе с хетами способствует
образованию вокруг тела коллемболы воздушной плёнки, которая рассматривается в
качестве пластрона. Практически на всём протяжении кутикулы имеется чередование
гидрофобных эпикутикулярных гранул с гидрофильными “впадинами”, через которые и
происходит газообмен. Его интенсивность повышается при высокой влажности, при
истирании кутикулы (после линьки транспирация вновь достигает нормального уровня).
Целиком гидрофильными участками являются везикулы вентральной трубки и когти
лапки. Помимо участия в газообмене и ионном обмене, вентральная трубка также
применяется для передвижения, к примеру, по вертикальным поверхностям за счёт
сил адгезии. Гидрофильность когтей позволяет коллемболам передвигаться по
жидкости, при этом погружённые когти образуют точку опоры, а остальная часть
лапки не даёт ноге потонуть, придавая плавучесть (Noble-Nesbitt, 1963).
1.2.2.6
Кровеносная система
Трубчатое сердце расположено в четвёртом сегменте брюшка и имеет от 2 до
6 пар остий. Спереди оно продолжается в виде аорты, сопровождается “крыловой”
мускулатурой. Состав гемолимфы и её клеточных элементов неизвестен (Shaller,
1970; Imms, 1977).
1.2.2.7 Нервная система, органы чувств
Нервная система по своему строению схожа с таковой насекомых. У коллембол
она состоит из пяти ганглиев (Hopkin, 1997). Надглоточный и подглоточный
ганглии (Рис.4) располагаются в голове и связаны между собой короткими
комиссурами, между которыми проходит пищевод. Остальные ганглии располагаются
по одному в каждом сегменте груди на вентральной тороне. Бывшие абдоминальные
ганглии слиты с ганглием заднегруди, который может заходить в первый сегмент
брюшка (Lubbock, 1873).
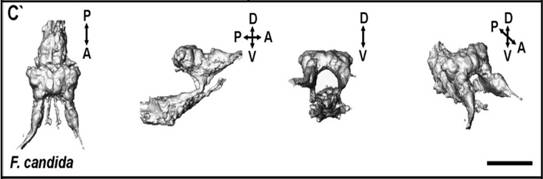
Рис. 2. Головные ганглии. P - задняя, A - передняя, V - нижняя, D -
верхняя оси. На второй проекции виден надглоточный (сверху) и подглоточный
(снизу) ганглии. Шкала: 50 мкм (из Kollmann, 2011).
Надглоточный ганглий состоит из протоцеребрума, дейтоцеребрума и
тритоцеребрума. Центральное тело - совокупность несвязанных нейропилей
(область, в которой сосредоточены синапсы) - не дифференцируется и лежит в
середине протоцеребрума с дорсальной стороны. Все функции до конца не
установлены, но одна из них - обработка визуальной информации. Ниже него
описаны структуры, похожие на узелки (нодули). Их назначение неизвестно, но, по
всей видимости, они отсутствуют у бескрылых насекомых. Есть предположение, что
их наличие связано с эволюцией крыльев. Ниже центрального тела располагается
вытянутый протоцеребральный мост. Грибовидные тела, располагающиеся в передней
части протоцеребрума, также состоят из нейропилей и являются важнейшими
ассоциативными центрами, принимая участие в обонятельном и зрительном
запоминаниях, процессах принятия решения. Грибовидные тела упрощённого
строения, с малым количеством интернейронов наименьшего размера (клеток
Кеньона) и отсутствием вертикальных долей (образования аксонов клеток Кеньона).
Антеннальные доли входят в состав дейтоцеребрума. Это области нейропиля,
ассоциированного с антеннами, которые состоят из отдельных гломерул -
шарообразные структуры из плотного синаптического нейропиля. Число гломерул
различается в зависимости от вида, у Folsomia candida было подсчитано 27. Эти
доли принимают обонятельную информацию (Шванвич, 1949; Chapman, 1998; Kollmann,
2011).
Нейросекреторная система в данной работе не рассматривалась.
Тело целиком покрыто хетами и чешуйками. Хорошо развиты органы чувств,
расположенные на антеннах. Имеются и трихоботрии - длинные хеты, регистрирующие
направление ветра и вибрации, и хемосенсорные хеты (сенсиллы). Совокупность
сенсилл может образовывать сенсорный комплекс, примером которого является
антеннальный орган, расположенный на дорсальной стороне третьего сегмента
антенны и участвующий в механо- и хеморецепции. Он состоит и 5 сенсилл. Другой
пример - это сложный сенсорный комплекс на конце антенны (описан у Allacma
fusca), возможные функции которого - восприятие механических раздражений
(механорецептор), в частности, восприятие сигналов от антеннальных мышц
(проприорецептор). Функции проприорецепторов также выполняют сколопии сложные
внутренние структуры в первом сегменте антенны. На конце антенн могут
располагаться хетоидная сенсилла и округлая папилла (везикула, или
ретрактильная колба). На вентральной стороне последнего сегмента антенны могут
быть найдены “сенсорные щётки” - поля коротких “шпильковидных” хет (Altner,
1978; Бабенко и др., 1988; Altner, 1988).
Постантеннальный орган - уникальная структура, которая расположена у
основания антенны и устроена по-разному в зависимости от таксона. Состоит из
различного количества бугорков (от 1 до розеток из 30 и больше), погружённых в
поверхностную ямку. Его функция до конца не известна; возможно, он является
хеморецептором (восприятие химических веществ), терморецептором (восприятие
температуры) или гидрорецептором (восприятие влажности). Предполагается, что у
группы Symphypleona он редуцирован до хетоподобной структуры, которая выше была
включена в сложный сенсорный комплекс. У группы Neanurinae и у некоторых
Entomobryomorpha он вовсе отсутствует. (Karuhize, 1971; Altner & Thies,
1976; D’Haese, 2003)
Фасеточные глаза отсутствуют, имеются глаза, состоящие максимум из 8
омматидиев каждый. Отмечена реакция коллембол на воздействие света (Jordana,
2000).
Для группы Onychiuridae характерны ложные глазки, или псевдоцелли, среди
которых выделяют три основных типа на основе строения. Были выдвинуты следующие
гипотезы об их функции: выделительная, защитная (выделение отпугивающих
хищников репеллентов).
Ещё одни структуры, найденные в разных группах, - псевдопоры. Есть
предположения, что они гомологичны ложным глазкам (D’Haese, 2003).
Существуют так называемые овальные органы, которые воспринимают
воздействия, оказываемые на экзоскелет (Бабенко и др., 1988).
1.2.2.8 Половая система
Мужская половая система представлена непарными или парными мешковатыми и
иногда слитыми семенниками, которые располагаются по бокам от кишечника на
нижней стороне тела либо с первого (второго) по пятый сегмент брюшка, либо
занимают его целиком, а иногда могут начинаться в среднегруди и продолжаться до
конца тела (Shaller, 1970). Они не разделены на фолликулы (Dallai, 2000).
Концы семяпроводов, или семяпротоков, отходят от семенников и на конце
образуют расширение - семенные пузырьки, в которых накапливается сперма. Далее
семяпротоки впадают в непарный семяизвергательный канал, который открывается
наружу на пятом сегменте брюшка продольным мужским половым отверстием (Shaller,
1970; Matsuda, 1976). Около канала располагаются придаточные железы, секрет
которых участвует в образовании сперматофора. Внутри каналов происходит
разделение секрета желёз и спермы друг от друга при помощи поперечной септ
(Dallai, 2000).
Женская половая система представлена парными мешковатыми, часто
лопастными яичниками, которые могут проходить от среднегруди до 5 сегмента
брюшка. Яйцеводы очень короткие, они объединяются для формирования непарного
влагалища, которое открывается наружу на пятом сегменте брюшка поперечным
женским половым отверстием. Деление на овариолы отсутствует, но у ряда видов
яичники могут вторично делиться на две несимметричные доли (Shaller, 1970;
Matsuda, 1976).
Размножение происходит с помощью сперматофора, который состоит из стебелька
и верхушечной капли со спермой. Самка подбирает его половым отверстием, где
сперматофор попадает в сперматеку (Рис.5). Сперматека - эктодермальный мешок,
который располагается между двумя другими похожими мешками рядом с влагалищем.
Две боковых структуры являются придаточными железами, функция секрета которых
до конца не выяснена. Существуют предположения, что он защищает яйца от
хищников, функционируя в качестве репеллента. Cперматека у самки
репродуктивного возраста достигает величины большей, чем придаточные железы. В
остальное время она малых размеров и запрятана между придаточными железами и
окружена жировым телом, поэтому её обнаружение затруднено. Способность
изменения размера сперматеки связана с тонким слоем кутикулы, который не
препятствует её растяжению во время захвата сперматофора. Напротив, придаточные
железы обладают толстой кутикулой и не меняют своих размеров (Dallai, 2008).

Рис. 3. Три эктодермальных кармана после препарирования дистальной части
половой системы ювенильной самки. go - половое отверстие, sp - сперматека, ag -
придаточные железы (из Dallai, 2008).
1.3 Миниатюризация животных
Миниатюризация - эволюция чрезвычайно маленьких размеров тела в ряде
поколений. Помимо маленького размера, миниатюризация подразумевает изменение
анатомии, физиологии, экологии, поведения. “Критическим размером”
миниатюризации считается такой размер, при котором затронуты важные
физиологические или экологические функции, например, локомоция, размножение,
питание (Hanken and Wake, 1993).
Миниатюризация у позвоночных часто встречается среди рыб, амфибий и
рептилий, реже - у птиц и млекопитающих. Появлению данного эффекта способствуют
особые условия среды, куда можно отнести островную фауну, Антарктику, мейофауну
(донные организмы размером от 60 мкм до 2 мм). Одним из механизмов
миниатюризации считается педоморфоз, играющий огромную роль в эволюции
мейофауны (Hanken and Wake, 1993; Rundell and Leander, 2010).
Наиболее частый эффект миниатюризации - упрощение и редукция структур,
начиная от недоразвитости и заканчивая потерей органов или целых систем
органов. Среди позвоночных наиболее часты примеры, связанные со скелетом.
Потеря целых систем органов характерна для беспозвоночных (Hanken and Wake,
1993).
Практически все эволюционные изменения видов сопровождаются изменением
размеров. Если в ходе эволюции организм становится меньше, то количество
успешных клеточных делений в тканях, клетки которых способны постоянно
делиться. Из-за этого могут исчезнуть целые органы, которые не способны
функционировать в связи с малым количеством клеток. Биологические параметры,
как и физиологические системы, широко варьируют и по-разному зависят от размера
тела, поэтому довольно редко удаётся определить критический размер для какой-либо
группы близкородственных организмов. Нижняя граница размера тела может
определяться лимитом количества яиц, максимальным заполнением тела органами
(Rensch, 1948; Hanken and Wake, 1993).
1.3.1 Эффект миниатюризации у насекомых
Существуют параллелизмы, характерные большинству микронасекомых, часть из
которых:
· упрощение тенториума и эндоскелета;
· редукция мускулатуры кишечника;
· сокращение числа мальпигиевых сосудов;
· упрощение кровеносной системы;
· упрощение трахейной системы (сокращение числа дыхалец);
· олигомеризация и концентрация ЦНС, асимметрия ЦНС;
· сокращение числа овариол.
Также существуют параллелизмы, характерные для части микронасекомых
(например, смещение брюшных ганглиев в брюшной отдел у Liposcelididae,
Ptiliidae, Corylophidae) и уникальные черты, присущие отдельным семействам или
микронасекомым (например, непарные гонады у Ptiliidae) (Polilov, 2015).
1.4 Систематическое положение и признаки рода
Mesaphorura
Надсемейство Onychiuroidea, семейство Tulbergiidae.
Описание рода:
Вытянутое тело длиной 0,4 - 1,2, чаще 0,6 - 1 мм. Пигмент отсутствует.
Антенны короче диаметра головы. Четвёртый сегмент антенны с пятью сенсиллами
различной толщины. Антеннальный орган (3 сегмент антенны) состоит из двух
больших сенсорных бугорков, между которыми проходят два сенсорных ряда.
Постантеннальный орган расположен в продолговатой или овальной полости, в 1,5-2
длиннее ближайших псевдооцеллей. Состоит из 18- 55 простых везикул, лежащих в
два ряда. Хорошо развитые ложные глазки имеют розетчатую форму и центр в виде
звезды.
Глава 2. Материалы и методы
Объекты были собраны на песчаном берегу Пироговского водохранилища города
Москвы в сентябре 2015 года. Для сбора использовался метод флотации (Потапов,
Кузнецова, 2011): берут сосуд, ёмкостью несколько литров (низкое ведро или
тазик), засыпают порцию песка, наливают воды и аккуратно перемешивают. Через
несколько минут микроартроподы всплывают, и их переносят иглой или тонкой
кисточкой во флаконы.
Живые объекты были зафиксированы спиртовым Буэном, после чего хранились в
70% спирте.
Для получения изображений внешнего строения часть образцов была отснята
при помощи сканирующего электронного микроскопа Jeol JSM-6380 после высушивания
в критической точке.
Для изучения внутреннего строения зафиксированные образцы были залиты в
аралдит, и из полученных блоков были сделаны полные срезы поперечных и
продольных срезов (1 и 0,5 мкм толщиной) на микротоме Leica RM2255 с алмазным
ножом. Полученные срезы окрашены толуидиновым синим и пиронином.
Для построения трёхмерной модели серия срезов была сфотографирована под
микроскопом Motic BA410 на камеру Toupteck. Трёхмерная компьютерная
реконструкция проведена в программе Bitplane Imaris 7.2.1, а дальнейшая
обработка моделей - в Autodesk Maya 2015.
Глава 3. Результаты
3.1 Морфология
Mesaphorura
3.1.1 Внешнее строение

Рис. 4. Внешний вид Mesaphorura sylvatica. A - вид сбоку, B - вид снизу,
C - вид сверху. Условные обозначения:ao -анальное отверстие, cx1 - тазик
первого сегмента груди, cx2 - тазик второго сегмента груди, cx3 - тазик третьего
сегмента груди, fu - анальные шипы, go - половое отверстие, PAO -
постантеннальный орган, sx1 - субкокса первого сегмента груди, sx2 - субкокса
второго сегмента груди, sx3 - субкокса третьего сегмента груди.
Размер взрослых особей около 0,4 мм (рис. 4). На первом сегменте брюшка
расположена вентральная трубка, на последнем - анальные шипы. На голове
расположен постантеннальный орган в продолговатой полости с двумя рядами
везикул. Ложные глазки тела хорошо развиты и представляют собой розетчатую структуру.
Антенны короче диаметра головы. Тергиты хорошо развиты, склериты и плевриты
плохо различимы.
3.1.2 Внутреннее строение
3.1.2.1 Эндоскелет
Толщина кутикулы в среднем - 0, 57 мкм. Максимальное значение - 1,24 мкм,
минимальное - 0,31 мкм.

Рис. 5. Эндоскелет. Вид сверху. Условные обозначения: en - эндостернит,
fur1 - первая фуркоподобная структура, fur2 - вторая фуркоподобная структура;
fur3 - третья фуркоподобная структура; pst - псевдотенториум.
Эндоскелет (рис. 5) сложно устроен, имеются расположенный в голове
псевдотенториум и три фуркоподобные структуры, которые располагаются на границе
между первым и вторым, вторым и третьим сегментах груди и между третьим
сегментом груди и первым сегментом брюшка. Псевдотенториум разветвлённый,
сложно устроенный, имеет передние и задние руки тенториума, а также различные
собственные выросты. Фуркоподобные структуры также разветвлённые, разной формы,
вторая имеет отростки меньшего размера. Между третьим сегментом груди и первым
сегментом брюшка имеется короткий прямоугольный эндостернит.
3.1.2.2 Мышечная система
Описаны 22 пары мышц в голове (рис. 6, табл. 1), 53 - в груди (рис. 7,
табл. 2) и 62 - в брюшке (и 1 непарная поперечная мышца) (рис. 8, табл. 3). В
голове имеются челюстные (18 пар), глоточные (3 пары) и антеннальная мышцы
(табл. 1). В груди и брюшке имеются дорсо-вентральные, латеральные
дорсо-вентральные, межсегментные дорсо-вентральные, вентральные и дорсальные
продольные мышцы. 15 пар стерно-коксальных мышц в груди (табл. 2). 13 пар мышц
брюшка связаны с вентральной трубкой (табл. 3), 4 пары связаны с ректумом
(табл. 3). Номенклатура проводилась на основе работ предыдущих авторов (Folsom,
1899; Bretfeld, 1963; Eisenbeis, 1975).
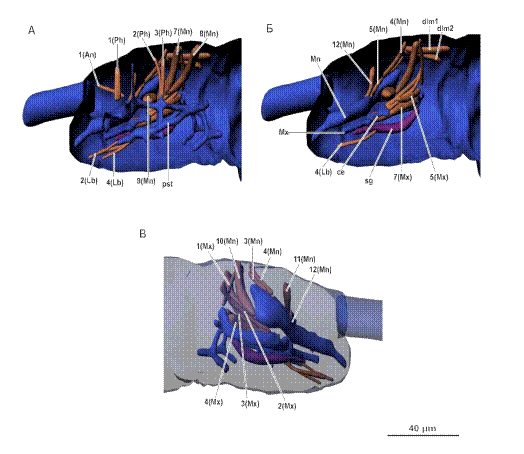
Рис. 6. Мускулатура головного отдела. А, Б - вид слева (изнутри), Б - вид
справа. Условные обозначения: ce - хитиновое расширение максиллы, pst -
псевдотенториум, sg - слюнная железа. Мускулатуру см. в тексте.
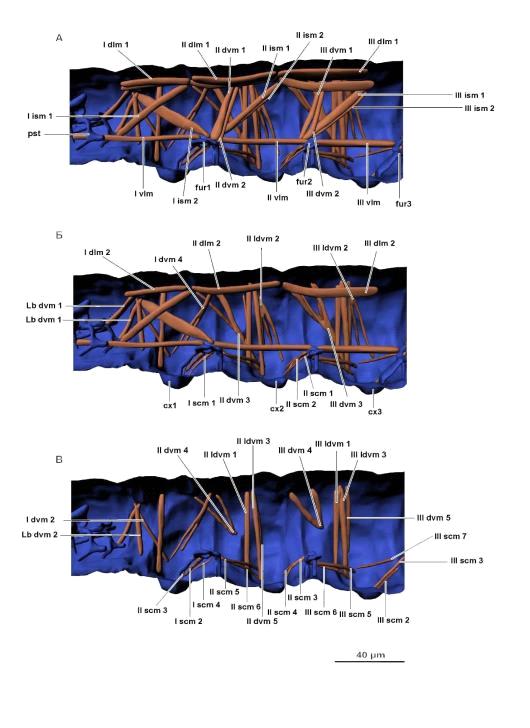
Рис. 7. Мускулатура грудного отдела. А, Б, В - вид слева (изнутри).
Условные обозначения: cx1 - тазик первого сегмента груди, cx2 - тазик второго
сегмента груди, cx3 - тазик третьего сегмента груди, fur1 - первая фуркоподобная
структура, fur2 - вторая фуркоподобная структура; fur3 - третья фуркоподобная
структура; pst - псевдотенториум. Мускулатуру см. в тексте.
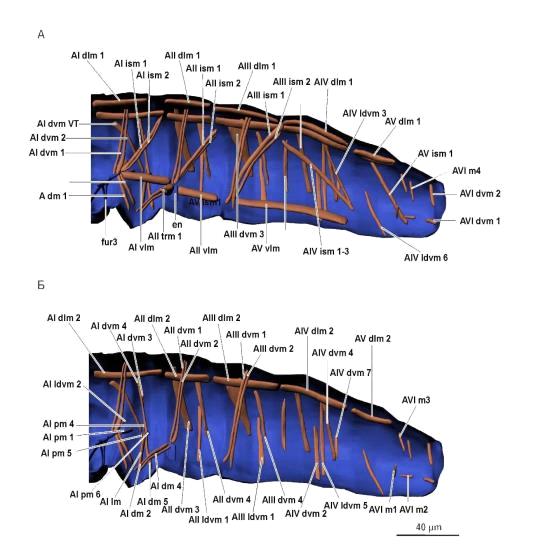
Рис. 8. Мускулатура брюшного отдела. А, Б - вид слева (изнутри). Условные
обозначения: en - эндостернит, fur 3 - третья фуркоподобная структура.
Мускулатуру см. в тексте.
Табл.1. Мускулатура головы.
|
Номенклатура в данной
работе
|
Отхождение
|
Прикрепление
|
|
2(Lb)
|
внутренний край нижней губы
|
слюнная железа, спереди от
Lb4, Lb5
|
|
4(Lb)
|
нижняя поверхность нижней
губы, медиально к Lb5
|
слюнная железа, сзади от
Lb2, спереди от Lb5
|
|
5(Lb)
|
нижняя поверхность нижней
губы, латерально к Lb4
|
слюнная железа, сзади от
Lb2 и Lb4
|
|
1(Mx)
|
медиальный верхний
хитиновый выступ рядом с основанием головы и Mn7
|
вогнутость кардо, выше Mx5
|
|
2(Mx)
|
задняя верхнебоковая часть
головы, ниже Mx3
|
хитиновое расширение
максиллы, медиально к Mx3
|
|
3(Mx)
|
задняя верхнебоковая часть
головы, выше Mx2
|
хитиновое расширение
максиллы, латерально к Mx2
|
|
4(Mx)
|
внутренняя поверхность
кардо
|
нижняя поверхность
хитинового расширения максиллы
|
|
5(Mx)
|
основание тенториума
|
вогнутость кардо, ниже Mx1
|
|
7(Mx)
|
основание стипеса
|
основание хитинового
расширения максиллы
|
|
3(Mn)
|
верхнебоковая часть головы,
сзади от её середины
|
внутренняя сторона
базального кольца мандибулы, сзади от Mn4
|
|
4(Mn)
|
латерально к середине
верхней поверхности головы, медиально к Mn 3
|
внутренняя сторона
базального кольца мандибулы, спереди от Mn3, сзади от Mn5
|
|
5(Mn)
|
передняя рука
псевдотенториума
|
верхняя поверность мандибулы,
спереди от Mn4
|
|
7(Mn)
|
медиальный верхний
хитиновый выступ рядом с основанием головы и Mx1, медиально к Mn10
|
нижний задний край
мандибулы вместе с Mn8, Mn10
|
|
8(Mn)
|
верхняя поверхность головы
рядом с основанием, сзади от Mn7, пересекая медианную плоскость
|
нижний задний край
мандибулы вместе с Mn7, Mn10
|
|
9(Mn)
|
основание псевдотенториума
|
внутренняя поверхность
мандибулы задней части мандибулы
|
|
10(Mn)
|
медиальный верхний
хитиновый выступ рядом с основанием головы, латерально к Mn7
|
нижний задний край мандибулы
вместе с Mn7, Mn8
|
|
11(Mn)
|
передний боковой край
головы у основания антенны, снизу от Mn12
|
наружный край мандибулы и
тенториум, латерально к Mn12
|
|
12(Mn)
|
передний боковой край
головы у основания антенны, сверху от Mn11
|
наружный край мандибулы и
тенториум, медиально к Mn11
|
|
1(An)
|
боковая поверхность первого
сегмента антенны
|
псевдотенториум
|
|
1(Ph)
|
медиальная поверхность
переднего края головы, спереди от ph2, ph3
|
пищевод
|
|
2(Ph)
|
медиальная поверхность
переднего края головы, латерально к ph3
|
пищевод, латерально к ph3
|
|
3(Ph)
|
медиальная поверхность
переднего края головы, медиально к ph2
|
пищевод, медиально к ph2
|
Табл. 2. Мускулатура груди.
|
Номенклатура мышц в данной
работе
|
Отхождение
|
Прикрепление
|
|
III dlm1
|
антекоста II, медиально к
III dlm2
|
антекоста III, медиально к
III dlm2
|
|
III dlm2
|
антекоста II, латерально к
III dlm1
|
антекоста III, латерально к
III dlm1
|
|
III vlm
|
заднебоковой участок
мезофуркоподобной структуры
|
переднебоковой участок
метафуркоподобной структуры
|
|
III ism1
|
боковой участок
мезофуркоподобной структуры, cпереди от III ism2
|
антекоста III, рядом с III
dlm1, спереди от III ism2
|
|
III ism2
|
боковой участок
мезофуркоподобной структуры, сзади от III ism1
|
антекоста III, рядом с III
dlm1, сзади от III ism1
|
|
III dvm1
|
верхнебоковой участок
мезофуркоподобной структуры, спереди от III dvm2
|
середина бокового участка
метанотума, спереди от III dvm2
|
|
III dvm2
|
верхнебоковой участок
мезофуркоподобной структуры, сзади от III dvm1
|
середина бокового участка
метанотума, сзади от III dvm1
|
|
III dvm3
|
передняя часть бокового участка
метанотума, сзади от антекосты II, вместе с III dvm4
|
верхний передний участок
метакоксы, сзади от III ldvm1, вместе с III ldvm3 и III scm5
|
|
III dvm4
|
передняя часть бокового
участка метанотума, сзади от антекосты II, вместе с III dvm3
|
верхний край метасубкоксы
|
|
III dvm5
|
cередина бокового участка
метанотума, вместе III ldvm1, III ldvm2, III ldvm3
|
боковой край метакоксы,
спереди от III ldvm2
|
|
III ldvm1
|
cередина бокового участка
метанотума, вместе с III ldvm2, III ldvm3, III dvm5
|
передний край метакоксы,
спереди от III ldvm3, III dvm3
|
|
III ldvm2
|
cередина бокового участка
метанотума, вместе с III ldvm1, III ldvm3, III dvm5
|
боковой край метакоксы,
сзади от III dvm5, и задний край метакоксы
|
|
III ldvm3
|
cередина бокового участка
метанотума, вместе с III ldvm1, III ldvm2, III dvm5
|
верхний передний участок
метакоксы, сзади от III ldvm1, вместе с III dvm3 и III scm5
|
|
II dlm1
|
антекоста I, медиально к II
dlm2
|
антекоста II, медиально к
II dlm2
|
|
II dlm2
|
антекоста I, латерально к
II dlm1
|
антекоста II, латерально к
II dlm1
|
|
II vlm
|
заднебоковой участок
профуркоподобной структуры
|
переднебоковой участок
мезофуркоподобной структуры
|
|
II ism1
|
верхнебоковой участок
профуркоподобной структуры, cпереди от II ism2
|
антекоста II, cпереди от II
ism2, латерально к II dlm1
|
|
II ism2
|
верхнебоковой участок
профуркоподобной структуры, cзади от II ism1
|
антекоста II, cзади от II
ism1, латерально к II dlm1
|
|
II dvm1
|
заднебоковой участок
профуркоподобной структуры, сзади от II dvm2
|
середина бокового участка
мезонотума, спереди от II dvm2, рядом с II dvm1
|
|
II dvm2
|
заднебоковой участок
профуркоподобной структуры, спереди от II dvm1
|
середина бокового участка
мезонотума, сзади от II dvm1
|
|
II dvm3
|
середина бокового участка
мезонотума (рядом с II dvm1) и передний участок мезонотума (вместе с II dvm4
и I dvm4)
|
верхний передний участок
мезококсы, сзади от II ldvm1, вместе с II ldvm3 и II scm5
|
|
II dvm4
|
передний участок мезонотума
(вместе с II dvm3 и I dvm4) и антекоста I, рядом с I ism1
|
верхний передний край
мезосубкоксы
|
|
II dvm5
|
cередина бокового участка
мезонотума, вместе с II ldvm1, II ldvm2, II ldvm3
|
нижний край мезосубкоксы,
спереди от II ldvm2
|
|
II ldvm1
|
cередина бокового участка
мезонотума, вместе с III ldvm2, III ldvm3, II dvm5
|
верхний передний край
мезококсы, спереди от II dvm3
|
|
II ldvm2
|
cередина бокового участка
мезонотума, вместе с III ldvm1, III ldvm3, II dvm5
|
нижний край мезосубкоксы и
задний край мезококсы
|
|
II ldvm3
|
cередина бокового участка
мезонотума, вместе с III ldvm1, III ldvm2, II dvm5
|
верхний передний участок
мезококсы, сзади от II ldvm1, вместе с II dvm3 и II scm5
|
|
I dlm1
|
верхний край заднезатылка,
медиально к I dlm2
|
антекоста I, медиально к I
dlm2
|
|
I dlm2
|
верхний край заднезатылка,
латерально к I dlm1
|
антекоста I, латерально к I
dlm1
|
|
I vlm1
|
середина псевдотенториума
|
передний участок
профуркоподобной структуры
|
|
I ism1a
|
антекоста I
|
задний вырост
псевдотенториума, вместе с Lb dvm1
|
|
I ism2
|
боковой край заднезатылка,
вместе с Lb dvm1, Lb dvm2, I dvm2
|
верхнебоковой участок
профуркоподобной структуры
|
|
I dvm2
|
боковой край заднезатылка,
вместе с Lb dvm1, Lb dvm2, I ism2
|
переднебоковой край
прококсы, вместе с I dvm4
|
|
I dvm4
|
передний участок
мезонотума, вместе с II dvm3, II dvm4
|
переднебоковой край
прококсы, вместе с I dvm4, и боковой край прококсы
|
|
I ldvm1
|
задний участок пронотума
|
верхний передний участок
прококсы
|
|
Lb dvm1
|
боковой край заднезатылка,
вместе с Lb dvm2, I dvm2, I ism2
|
задний вырост
псевдотенториума, вместе сI ism1a
|
|
Lb dvm2
|
боковой край заднезатылка,
вместе с Lb dvm1, I dvm2, I ism2
|
нижняя граница головы и
проторакса
|
|
I scm1
|
передняя поверхность
профуркоподобной структуры
|
задняя поверхность
прококсы, рядом с I scm2, медиально к I scm4
|
|
I scm2
|
нижняя медиальная
поверхность профуркоподобной структуры
|
задняя поверхность
прококсы, рядом с I scm1, медиально к I scm4
|
|
I scm3
|
нижняя боковая поверхность
профуркоподобной структуры, вместе с I scm4
|
боковая поверхность
прококсы
|
нижняя боковая поверхность
профуркоподобной структуры, вместе с I scm3
|
задняя поверхность
прококсы, латерально к I scm2, I scm1
|
|
II scm1
|
передняя поверхность
мезофуркоподобной структуры, медиально к II scm3
|
задняя поверхность
мезококсы, рядом с I scm2, медиально к I scm4
|
|
II scm2
|
нижняя боковая поверхность
мезофуркоподобной структуры, вместе с II scm4
|
задняя поверхность
мезококсы, рядом с I scm1
|
|
II scm3
|
передняя поверхность
мезофуркоподобной структуры, латерально к II scm1
|
задняя или боковая
поверхность мезококсы, рядом с I scm4
|
|
II scm4
|
нижняя боковая поверхность
мезометафуркоподобной структуры, вместе с II scm2
|
задняя или боковая
поверхность мезококсы, рядом с I scm4
|
|
II scm5
|
задняя поверхность
профуркоподобной структуры, латерально к II scm6
|
верхний передний участок
мезококсы, сзади от II ldvm1, вместе с II dvm3 и II ldvm3
|
|
II scm6
|
задняя поверхность
профуркоподобной структуры, медиально к II scm5
|
боковая поверхность
мезококсы
|
|
III scm2
|
нижняя медиальная
поверхность метафуркоподобной структуры
|
задняя поверхность
метакоксы
|
|
III scm3
|
нижняя боковая поверхность
метафуркоподобной структуры
|
боковая или передняя
поверхность метакоксы
|
|
III scm5
|
задняя поверхность
мезофуркоподобной структуры, латерально к III scm6
|
верхний передний участок
метакоксы, сзади от III ldvm1, вместе с III dvm3 и III ldvm3
|
|
III scm6
|
задняя поверхность
мезофуркоподобной структуры, медиально к III scm5
|
боковая поверхность
метакоксы, спереди от III scm7
|
|
III scm7
|
задняя поверхность
метафуркоподобной структуры
|
боковая поверхность
метакоксы, сзади от III scm7
|
|
Lb dlm1
|
верхний край заднезатылка,
медиально к Lb dlm2
|
латерально к середине
верхней поверхности головы, медиально к Lb dlm2
|
|
Lb dlm2
|
верхний край заднезатылка,
латерально к Lb dlm1
|
латерально к середине
верхней поверхности головы, латерально к Lb dlm1
|
Табл.3. Мускулатура брюшка.
|
Номенклатура мышц в данной
работе
|
Отхождение
|
Прикрепление
|
|
AI dlm1
|
антекоста III, медиально к
AI dlm2
|
антекоста IV, медиально к
AI dlm2
|
|
AI dlm2
|
антекоста III, латерально к
AI dlm1
|
антекоста IV, латерально к
AI dlm1
|
|
AI vlm
|
боковой участок
метафуркоподобной структуры
|
эндостернит I
|
|
AI ism1
|
антекоста IV, спереди от AI
ism2
|
боковой участок
метафуркоподобной структуры, спереди от AI ism2
|
|
AI ism2
|
антекоста IV, сзади от AI
ism1
|
боковой участок
метафуркоподобной структуры, сзади от AI ism1
|
|
AI dvm1
|
середина тергита между AI
dlm1 и AI dlm2, спереди от AI dvm2
|
боковой участок
метафуркоподобной структуры, спереди от AI dvm2
|
|
AI dvm2
|
середина тергита между AI
dlm1 и AI dlm2, сзади от AI dvm1
|
боковой участок
метафуркоподобной структуры, сзади от AI dvm1
|
|
AI ldvm1
|
середина боковой
поверхности сегмента
|
середина бокового края
стернита
|
|
AI ldvm2
|
боковая поверхность сегмента,
ниже его середины
|
боковой участок
метафуркоподобной структуры
|
|
AI dvm VT
|
передняя часть верхнего
края тергита
|
сухожилие, вместе с AI Im,
AI pm4, AI pm5, AI pm6, AI dvm3, AI dvm4, AI dm5
|
|
AI dm1
|
заднебоковой участок
мутафуркоподобной структуры , медиально к dm3
|
передняя боковая
поверхность вентральной трубки, медиально к AI dm3
|
|
AI dm2
|
эндостернит I, латерально к
dm4
|
передняя поверхность
вентральной трубки, медиальный желоб, спереди от dm4
|
|
AI dm4
|
эндостернит I, медиально к
dm2
|
передняя поверхность вентральной
трубки, медиальный желоб, сзади от dm2
|
|
AI lm
|
сухожилие, вместе с pm4,
pm5, pm6, AI dvm3, AI dvm4, dm5, AI dvm VT
|
клапан вентральной трубки
|
|
AI pm1
|
боковой участок
мутафуркоподобной структуры
|
передний боковой край
основания вентральной трубки
|
|
AI pm4
|
сухожилие, вместе с AI Im,
AI pm5, AI pm6, AI dvm3, AI dvm4, AI dm5, AI dvm VT
|
боковой край основания
вентральной трубки, сверху от pm5, pm6
|
|
AI pm5
|
сухожилие, вместе с AI Im,
AI pm4, AI pm6, AI dvm3, AI dvm4, AI dm5, AI dvm VT
|
боковой край основания вентральной
трубки, ниже pm4, спереди от pm6
|
|
AI pm6
|
сухожилие, вместе с AI Im,
AI pm4, AI pm5, AI dvm3, AI dvm4, AI dm5, AI dvm VT
|
боковой край основания
вентральной трубки, сзади от pm5
|
|
AI dvm3
|
середина боковой
поверхности сегмента
|
сухожилие, вместе с AI Im,
AI pm4, AI pm5, AI pm6, AI dvm4, AI dm5, AI dvm VT
|
|
AI dvm4
|
верхний край сегмента
посередине
|
сухожилие, вместе с AI Im,
AI pm4, AI pm5, AI pm6, AI dvm3, AI dm5, AI dvm VT
|
|
AI dm3
|
заднебоковой участок
мутафуркоподобной структуры , латерально к AI dm1
|
передняя боковая
поверхность вентральной трубки
|
|
AI dm5
|
сухожилие, вместе с AI Im,
AI pm4, AI pm5, AI pm6, AI dvm4, AI dm5, AI dvm VT
|
задняя поверхность
вентральной трубки
|
|
AII dlm1
|
антекоста IV, медиально к
AII dlm2
|
антекоста V, медиально к
AII dlm2
|
|
AII dlm2
|
антекоста IV, латерально к
AII dlm1
|
антекоста V, латерально к
AII dlm1
|
|
AII vlm
|
эндостернит I
|
межсегментный участок между
стернитами 2 и 3 сегментов
|
|
AII ism1
|
антекоста V, спереди от AII
ism2
|
эндостернит I, спереди и
медиально к AII ism2, латерально к AII dvm1, AII dvm2
|
|
AII ism2
|
антекоста V, сзади от AII
ism1
|
эндостернит I, спереди и
латерально к AII ism1, латерально к AII dvm1, AII dvm2
|
|
AII dvm1
|
середина тергита между AII
dlm1 и AII dlm2, спереди от AII dvm2
|
эндостернит I, медиально к
AII ism1, AII ism2, спереди от AII dvm2
|
|
AII dvm2
|
середина тергита между AII
dlm1 и AII dlm2, сзади от AII dvm1
|
эндостернит I, медиально к
AII ism1, AII ism2, сзади от AII dvm1
|
|
AII dvm3
|
верхний передний край
тергита, латерально к AII dlm1
|
переднебоковой край
вентральной поверхности сегмента
|
|
AII dvm4
|
верхний край сегмента
посередине
|
заднебоковой край
вентральной поверхности сегмента
|
|
AII ldvm1
|
середина боковой
поверхности сегмента
|
середина бокового края
стернита
|
|
AII trm1
|
внутренняя поверхность
эндостернита
|
внутренняя поверхность
эндостернита
|
|
AIII dlm1
|
антекоста V, медиально к
AIII dlm2
|
антекоста VI, медиально к
AIII dlm2
|
|
AIII dlm2
|
антекоста V, латерально к
AIII dlm1
|
антекоста VI, латерально к
AIII dlm1
|
|
AIII ism1
|
антекоста VI, медиально к
AIII ism2
|
межсегментный участок между
стернитами 2 и 3 сегментов, впереди от AIII ism2, латерально к AIII dvm1,
AIII dvm2
|
|
AIII ism2
|
антекоста VI, латерально к
AIII ism2
|
межсегментный участок между
стернитами 2 и 3 сегментов, сзади от AIII ism1, латерально к AIII dvm1, AIII
dvm2
|
|
AIII dvm1
|
середина тергита между AIII
dlm1 и AIII dlm2, спереди от AIII dvm2
|
межсегментный участок между
стернитами 2 и 3 сегментов, медиально к AIII ism1, AIII ism2, спереди от AIII
dvm2
|
|
AIII dvm2
|
середина тергита между AIII
dlm1 и AIII dlm2, сзади от AIII dvm1
|
межсегментный участок между
стернитами 2 и 3 сегментов, медиально к AIII ism1, AIII ism2, сзади от AIII
dvm1
|
|
AIII dvm3
|
верхний передний край
тергита, латерально к AIII dlm1
|
переднебоковой край
вентральной поверхности сегмента
|
|
AIII dvm4
|
верхний край сегмента
посередине
|
заднебоковой край
вентральной поверхности сегмента
|
|
AIII ldvm1
|
середина боковой
поверхности сегмента
|
середина бокового края
стернита
|
|
AIV dlm1
|
антекоста VI, медиально к
AIV dlm2
|
антекоста VII, медиально к
AIV dlm2
|
|
AIV dlm2
|
антекоста VI, латерально к
AIV dlm1
|
антекоста VII, латерально к
AIV dlm1
|
|
АIV vlm
|
межсегментный участок между
стернитами 2 и 3 сегментов
|
межсегментный участок между
стернитами 3 и 4 сегментов
|
|
AIV dvm3
|
верхний передний край
тергита, латерально к AIV dlm1
|
переднебоковой край
вентральной поверхности сегмента
|
|
AIV ism1-3
|
межсегментный участок между
3 и 4 сегментами, верхний край, спереди от AIV dvm4
|
межсегментный участок между
стернитами 4 и 5 сегментов вместе с AIV ldvm3
|
|
AIV dvm2
|
середина боковой
поверхности сегмента
|
стернит, вместе с AIV ldvm5
|
|
AIV dvm4
|
межсегментный участок между
3 и 4 сегментами, верхний край, сзади от AIV ism1-3
|
боковой край вентральной
поверхности межсегментного участка
|
|
AIV ldvm3
|
верхний край сегмента,
сзади от его середины, вместе с AIV ldvm4, медиально к AIV ldvm5
|
межсегментный участок между
стернитами 4 и 5 сегментов вместе с AIV ism1-3
|
|
AIV ldvm4
|
верхний край сегмента,
сзади от его середины, вместе с AIV ldvm3, медиально к AIV ldvm5
|
заднебоковой край
вентральной поверхности сегмента, вместе с AIV ldvm7
|
|
AIV ldvm5
|
верхний край сегмента,
сзади от его середины, латерально к AIV ldvm3, AIV ldvm4
|
стернит, вместе с AIV dvm2
|
|
AIV ldvm7
|
боковая поверхность
сегмента, сзади от его середины
|
заднебоковой край
вентральной поверхности сегмента, вместе с AIV ldvm4
|
|
AV dlm1
|
антекоста VII, медиально к
AV dlm2
|
антекоста VIII, медиально к
AV dlm2
|
|
AV dlm2
|
антекоста VII, латерально к
AV dlm1
|
антекоста VIII, латерально
к AV dlm1
|
|
AV ldvm1
|
середина боковой
поверхности сегмента
|
середина стернита
|
|
AV ism1
|
середина верхнего края
тергита
|
боковой край вентральной
поверхности межсегментного участка или стернита 6 сегмента
|
|
AVI m1
|
нижний боковой край
сегмента, спереди от AV ism1
|
ректум
|
|
AVI m2
|
нижний боковой край
сегмента, рядом с AV ism1
|
ректум
|
|
AVI m3
|
верхний боковой край
сегмента, спереди от AVI m5
|
ректум, спереди от AVI m5
|
|
AVI m4
|
верхний боковой край
сегмента, сзади от AVI m4
|
ректум, сзади от AVI m4
|
|
AVI dvm1
|
нижний боковой край
сегмента
|
боковая анальная лопасть
|
|
AVI dvm2
|
верхний задний край
сегмента
|
верхняя анальная лопасть
|
3.1.2.3 Пищеварительная система
Пищеварительная система (рис. 9) в виде трубки, которая делится на
переднюю, среднюю и заднюю кишки и не образует изгибов и выростов. Тонкая
передняя кишка круглая в поперечном сечении, начинается ротовым отверстием и
продолжается пищеводом. Пищевод диаметром 4,2 мкм, имеет круговую мускулатуру и
три пары глоточных мышц. На уровне второго сегмента груди он переходит в более
толстую среднюю кишку, которая образована одним слоем клеток (6-8 клеток в
слое). Первая половина её круглая в поперечном сечении, диаметром 27,8 мкм, во
второй половине поперечное сечение овальное. Имеется перитрофическая мембрана.
Пара слюнных желёз расположены в голове, их протоки впадают в ротовую полость
за нижней губой, заканчиваются они у конца головы. Переход средней кишки в
заднюю слабо различим на уровне четвёртого сегмента брюшка. Задняя кишка
диаметром 9,34 мкм, с круговой мускулатурой. Ректум заметен на уровне шестого
сегмента брюшка в виде небольшого расширения. К нему подходят четыре пары мышц.
За канчивается пищеварительная система анальным отверстием, которое открывается
на последнем сегменте тремя анальными лопастями.
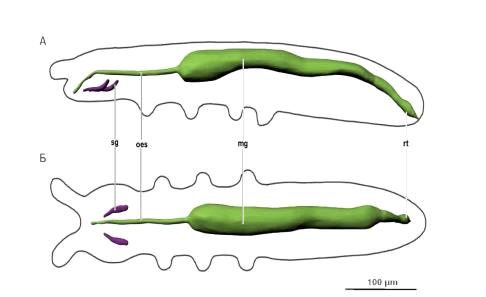
Рис. 9. Пищеварительная система. Условные обозначения: mg - средняя
кишка, oes - пищевод, rt - ректум, sg - слюнные железы.
3.1.2.4 Нервная система
Нервная система (рис. 10) состоит из надглоточного ганглия (мозг),
подглоточного ганглия и трёх грудных ганглиев. Надглоточный ганглий начинается
у основания антенн и заканчивается в середине первого сегмента груди. Дальше
середины головы (позади начала подглоточного ганглия) мозг имеет сужение.
Подглоточный ганглий начинается на середине головы и тянется до конца первого
сегмента груди. Соединение этих ганглиев широкое, расположено на уровне задней
части головы и затылка. Сквозь надглоточный ганглий проходят две пары глоточных
мышц. Ганглии груди крупные, хорошо различимые, расположены в нижней половине
тела и доходят до кутикулы. Переходы между ними в виде сужений расположены в
межсегментных участках и огибают элементы эндоскелета.
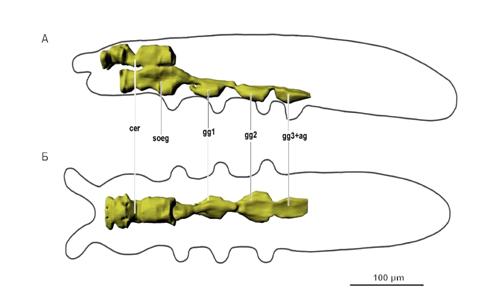

Рис. 10. Нервная система. Условные обозначения: cer - мозг, soeg -
подглоточный ганглий, gg1 - первый грудной ганглий, gg2 - второй грудной
ганглий, gg3 - третий грудной ганглий, gg3+ag - четвёртый грудной ганглий и
брюшной синганглий.
3.1.2.5 Половая система
Половая система (рис. 11) изученной самки начинается слитым непарным
яичником с двумя лопастями. Яичник, заполненный яйцами, начинается между вторым
и третьим сегментом груди и доходит до границы между четвёртым и пятым
сегментами брюшка. Парные лопасти расположены несимметрично относительно друг
друга на уровне второго-четвёртого сегментов брюшка, яйца в них отсутствуют.
Яйцевод также слитый непарный, короткий, начинается на конце четвёртого
сегмента брюшка и переходит в непарное влагалище, границы которого неразличимы.
Влагалище открывается поперечным половым отверстием на пятом сегменте брюшка.
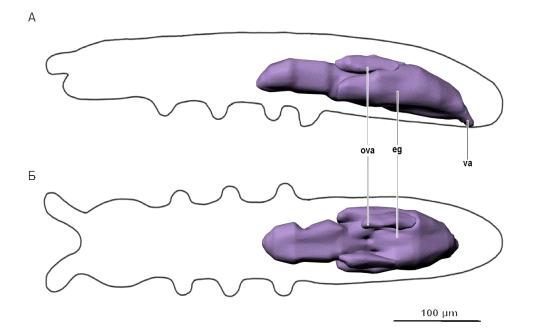
Рис. 11. Половая система. Условные обозначения:eg - яичник с яйцами, ova
- лопасти яичника без яиц, va - влагалище.
3.1.2.6 Прочие системы
Органы дыхательной (трахеи), кровеносной (сердце, сосуды) и выделительной
(лабиальные железы) систем отсутствуют.
Глава 4. Обсуждение
4.1 Эффект миниатюризации у коллембол
4.1.1 Сравнение Mesaphorura и ранее изученных
представителей данного отряда
Наименьшие изменения обнаружены в пищеварительной системе, сохраняется
одна пара слюнных желёз (Lubbock, 1873; Folsom, 1899; Wolter, 1963; Shaller,
1970).
Половая система самок коллембол представлена парными яичниками и
яйцеводами, у Mesaphorura в связи с миниатюризацией эти структуры становятся
парными. Половые железы и сперматека не найдены, что связано, скорее всего, с
их слабой выраженностью и трудностью обнаружения даже у более крупных
представителей (Shaller, 1970; Matsuda, 1976; Dallai, 2000; Dallai, 2008).
Сокращение числа мышц наблюдается во всех отделах. Выделяются 25
челюстных мышц у Orchesella cincta (Folsom, 1899), y Mesaphorura остаётся 18. В
грудном отделе Neanura muscorum (Bretfeld, 1963) выделяются 49 пар мышц (аз
исключением стерно-коксальных), у Mesaphorura их 53 (15 из которых -
стерно-коксальные). В брюшном отделе Neanura muscorum 63 пар мышц (Bretfeld,
1963), не включая 14 пар мышц, связанных с вентральной трубкой, которые описаны
отдельно у Sminthurides aquaticus (Eisenbeis, 1975). У Mesaphorura всего 64
пары мышц, из них 14 связаны с вентральной трубкой и 4 - с ректумом. Упоминания
мускулатуры средней кишки не встречаются.
Элементы нервной системы соответствуют таковым у остальных коллембол, но
строение надглоточного ганглия с проходящими сквозь него мышцами уникально, что
тоже связано с уменьшением размеров тела (Lubbock, 1873; Hopkin, 1997;
Kollmann, 2011).
Кровеносная система коллембол представлена сердцем с 2-6 парами остий и
отходящей вперёд аортой, у Mesaphorura в связи с миниатюризацией эти структуры
отсутствуют (Shaller, 1970; Imms, 1977).
Дыхательная система в виде трахей описана только у Symphypleona и
отдельных представителей Actaletoidea (ныне Entomobryomorpha и Poduromorpha),
поэтому роду Mesaphorura она не характерна (Davies, 1927; Shaller, 1970;
Hopkin, 1997).
Отсутствие лабиальных желёз выделительной системы, вероятно, также можно
считать проявлением эффекта миниатюризации, хотя у данного семейства
выделительная система ранее описана не была (Wolter, 1963; Verhoef, 1979).
4.1.2 Сравнение эффектов миниатюризации коллембол и насекомых
При миниатюризации важные изменения происходят в структуре экзоскелета и
эндоскелета насекомых. Отсутствуют руки тенториума, фурки становятся
неразветвлёнными (Polilov, 2015). У Mesaphorura sp., напротив, псевдотенториум
и фурки сохраняют сложную структуру, и из эндоскелета редуцируются лишь
эндостерниты брюшка (остаётся один).
У микронасекомых кровеносная система сильно упрощается и состоит из
сердца и короткой аорты (Polilov, 2015). Как и у рода Mesaphorura, у жуков
семейства Ptiliidae кровеносная система сильно редуцирована, сердце и аорта
отсутствуют, а гемолимфа занимает лишь небольшую долю полости тела, так как
большая часть полостей занята жировым телом. Вместо кровеносной системы имеет
место диффузия метаболитов, что может быть более эффективным благодаря малому
размеру (Polilov, 2008; Polilov and Beutel, 2009). Такое же упрощение можно
наблюдать у перепончатокрылых семейства Trichogrammatidae (Polilov, 2016).
Как и у рода Mesaphorura, у жуков семейства Ptiliidae половая система
упрощается: семенник и семяпровод самца, а также яичник и яйцевод самки
непарные (Polilov, 2008).
Специализированные органы выделения насекомых - мальпигиевы сосуды - у
микронасекомых полностью не исчезают, но их количество уменьшается (Polilov,
2015). У коллембол от мальпигиевых сосудов могут оставаться лишь небольшие
бугорки (Shaller, 1970). Лабиальные нефридии у рода Mesaphorura отсутствуют. В
отличие от изученной коллемболы, микронасекомым характерна редукция слюнных
желез (Polilov, 2015). Отсутствие мышц средней кишки - ещё один признак
миниатюризации у насекомых, который свойственен и Mesaphorura.
Мышечная система микронасекомых характеризуется исчезновением отдельных
мышц, подобное происходит и у коллембол (на 23,5% из описанных ранее).
Считается, что миниатюризация слабо влияет на мускулатуру, которая подвержена
изменениям гораздо меньше, чем скелет (Polilov, 2015).
4.1.2.1 Сравнение объёмов органов
Mesaphorura и некоторых насекомых
Объём тела Mesaphorura sylvatica - 0,8 нл. Относительный объём (рис.
12) скелета сопоставим с таковым у взрослых особей Paraneoptera, но
значительно меньше, чем у других миниатюрных насекомых того же размерного
класса. Это может быть связано с различием в толщине кутикулы.
Относительный объём мускулатуры значительно меньше остальных изученных
насекомых за исключением личинок Coleoptera, что может быть связано с
достаточной специализацией мышц у Collembola для того, чтобы совершать
необходимые движения без увеличения объёма мышц.
Относительный объём пищеварительной системы схож с таковым у Coleoptera,
значительно меньше, чем у Paraneoptera, но больше, чем у Hymenoptera и
отдельных видов Coleoptera. Подобные изменения, вероятно, связаны с
эффективностью работы кишечника при уменьшении размеров тела (Polilov and
Makarova, 2017).
Относительный объём кровеносной системы и жирового тела высок, его
превышают только объёмы личинок Paraneoptera и Coleoptera. Это можно объяснить
тем, что у коллембол жировое тело играет важную роль в выделении (Shaller,
1970).
Относительный объём половой системы также выше, чем у большинства
насекомых того же размерного класса, и уступает только объёму некоторых
взрослых особей Coleoptera и Hymenoptera. Такие особенности могут быть связаны
с особенностями относительного размера яиц (Polilov, Makarova, 2017).
Относительный объём центральной нервной системы сопоставим с
относительным объёмом взрослых особей Coleoptera, значительно меньше, чем у
Hymenoptera и личинок Paraneoptera и Coleoptera, и больше, чем у взрослых
особей Paraneoptera.
Относительный объём мозга сравним с таковым у взрослых особей Coleoptera,
больше, чем у взрослых особей Paraneoptera и значительно меньше, чем у прочих
миниатюрных насекомых того же размерного класса. Такой маленький объём можно
объяснить более простым поведением (Jordana, 2000), в том числе отсутствием
глаз (Hopkin, 1997).
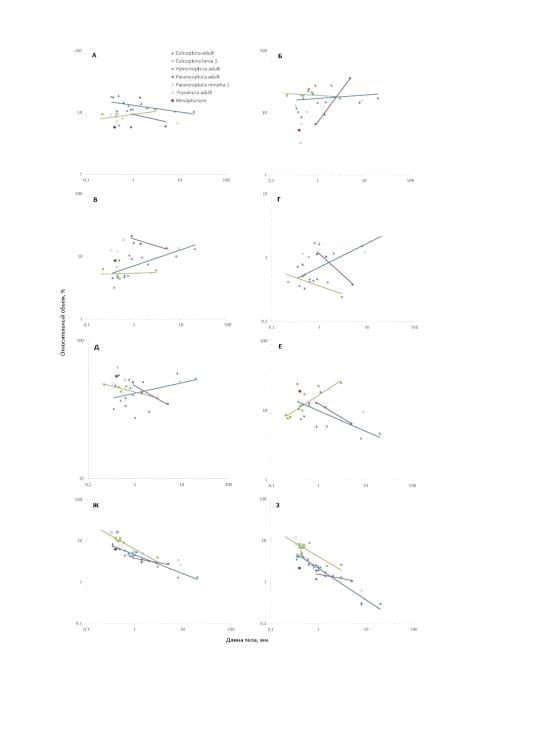
Рис. 12. Изменения относительного объёма органов взрослых и личинок
насекомых разных таксонов и коллембол. (A) - скелет, (Б) - мускулатура, (В) -
пищеварительная система, (Г) - мальпигиевы сосуды, (Д) - кровеносная система и
жировое тело, (Е) - половая система, (Ж) - центральная нервная система, (З) -
мозг (взято из Polilov, Makarova, 2017).
Выводы
1. Впервые изучена анатомия миниатюрной коллемболы Mesaphorura
sylvatica, и показано, что несмотря на чрезвычайно малые размеры тела (400 мкм
длиной), заметного упрощения уровня организации большинства систем органов не
наблюдается.
2. Эффекты миниатюризации внутреннего строения Mesaphorura
sylvatica проявляются в слиянии парных элементов половой системы (яичники,
яйцеводы), сокращении числа мышц, изменении нервной системы (появление
отверстий для сквозного прохождения мышц), а также в отсутствии
специализированных органов выделительной (лабиальные железы) и кровеносной
(сердце и сосуды) систем.
3. Внутреннее строение изученного вида имеет как черты
миниатюризации, которые являются параллелизмами с особенностями строения других
микронасекомых (непарная половая система, сокращение мышечной системы, редукция
кровеносной и выделительной систем), так и уникальные черты (отверстия в
надглоточном ганглии).
Список литературы
4.2 Бабенко
А.Б., Кузнецова H.А., М.Б. Потапов, Стебаева С.К., Ханисламова Г.М., Чернова
Н.М. Определитель коллембол фауны СССР // М: Наука, 1988. 214 с.
.3 Потапов
М.Б., Кузнецова Н.А. Методы исследования обществ микроартропод // М.: Т-во
научных изданий КМК. 2011. 84 с.
.4 Шванвич
Б.Н. Курс общей энтомологии. Введение в изучение строения и функций тела
насекомых // М.Л.: Советская наука, 1949. 899 с.
.5 Altner,
H. Die Ultrastruktur der Labialnephridien von Onychiurus quadriocellatus
(Collembola) // J. Ultrastructure Research, 1968. Vol. 24. P. 349- 366.
.6 Altner,
H., Thies, G. The Postantennal Organ: A Specialized Unicellular Sensory Input
to the Protocerebrum in Apterygotan Insects (Collembola) // Cell Tiss., 1976.
Res. 167. P.97-110.
.7 Altner,
H. The scolopidial organs in the first antennal segment in Allacma fusca
(Collembola, Sminthuridae) // Zoomorphology (berlin), 1988. Vol 108(3). P.
173-181.
4.8 Beutel
RG, Friedrich F, Ge S-Q, Yang XK. Insect Morphology and Phylogeny. De
Gruyter, Berlin. 2013.
4.9 Bitsch
C., Bitsch J. The endoskeletal structures in artropods: cytology, morphology
and evolution // Arthropod Structure & Development, 2003. Vol. 30. P.
159-177.
.10 Blanke
A., Machida R. The homology of cephalic muscles and endoskeletal elements
between Diplura and Ectognatha (Insecta), Organisms Diversity & Evolution,
2015. Pp. 17.
.11 Bretfeld,
G. Zur Anatomie und Embryologie der Rumpfmuskulatur und der
abdominalen Anha¨nge der Collembolen // Zoologische Jahrbu¨cher
(Anatomie), 1963. Vol. 80. P. 309-384.
.12 Chapman
R. F. The Insects: Structure and Function // Cambridge University Press,
Nature, 1998. Pp. 770.
.13 Cook,
C.E., Yue, Q.-Y. et Akam, M. Mitochondrial genomes suggest that hexapods and
crustaceans are mutually paraphyletic // Proceedings of the Royal Society B,
2005. Vol. 272. P. 1295-1304.
.14 D’Haese,
C.A. Homology and morphology in Poduromorpha (Hexapoda, Collembola) // Eur. J.
Entomol, 2003. Vol. 101. P. 385-407.
4.15 Dallai,
R, Cavallo, V, Falso, L.F, Fanciulli, P.P. The fine structure of the male
genital organs of Allacma fusca (L.) (Collembola, Symphypteona) // Pedobiologia, 2000. Vol. 44
(3-4). P. 202-209.
4.16 Dallai,
R., Zizzari, Z.V. & Fanciulli, P.P. Fine structure of the spermatheca and
of the accessory glands in Orchesella villosa (Collembola, Hexapoda). Journal of Morphology, 2008. Vol.
269(4). P. 464-478.
4.17 Davies,
W.M. On the tracheal system of Collembola, with special reference to that of
Sminthurus viridis Lubb // Quart. J. micr. Sci., N.S., 1927. Vol. 281. P. 15- 30.
.18 Eisenbeis,
G., Wichard, W. Feinstruktureller und histochemischer Nachweis des
Transportepithels am Ventraltubus symphypleoner Collembolen (Insecta,
Collembola) // Z. morph. Tiere, 1975. Vol. 81. P. 103-110.
4.19 Faddeeva, A., Studer, R.A., Kraaijeveld, K.,
Sie, D., Ylstra, B., Mariën, J., op den Camp, H.J.M., Datema, E., den
Dunnen, J.T., van Straalen, N.M. et Roelofs, D. Collembolan transcriptomes
highlight molecular
evolution of hexapods and provide clues on the adaptation to terrestrial life
// PLoS ONE, 2015. Vol. 10(6). P. 1-18.
.20 Folsom,
J.W. The Anatomy and Physiology of the Mouth-Parts of the Collembolan,
Orchesella cincta L // Bull. Mus. Comp. Zoology, 1899. Bd. 35. Pp. S.
.21 Hanken,
J., & Wake, D. B. Miniaturization of body size: organismal consequences and
evolutionary significance // Annual Reviews of Ecology and Systematics, 1993.
Vol. 24. Pp. 501-519.
.22 Hopkin,
Stephen, P. Biology of the springtails (Insecta: Collembola) // Oxford
University Press, Oxford, New York etc., 1997. Pp. 330.
.23 Humbert,
W. Ultrastructure des Nephrocytes cephaliques et abdominaux chez Tomocerus
minor (Lubbock) et Lepidocyrtus curvicollis Bourlet (Collemboles) // Int. J.
Insect Morphol. & Embryol., 1975. Vol. 4. P. 307-318.
.24 Humbert,
W. The midgut of Tomocerus minor Lubbock (Insecta: Collembola): ultrastructure,
cytochemistry, ageing and renewal during a moulting cycle // Cell And Tissue
Research, 1979. Vol. 196(1). P. 39-57.
4.25 Imms,
A.D. On the antennal musculature in insects and other arthropods // Quart. J. micr. Sci., 1939. Vol.2. P.
273-320.
.26 Imms,
A. D. A general textbook of Entomology, 10th edn., revised by O. W. Richards
and R. G. Davies // London: Methuen, 1957.
4.27 Jordana,
R, Baquero, E, Montuenga, L.M. A new type of arthropod photoreceptor //
Arthropod structure & development, 2000. Vol. 29 (4). P. 289-293.
.28 Karaban
K., Karaban E., Uvarov A. V.. Determination of life form spectra in soil
Collembola communities: a comparison of two methods // Polish Journal of
Ecology, 2012. Vol. 60 (1). P. 201-206.
4.29 Karuhize,
G.R. The Structure of the Postantennal Organ in Onychiurus sp. (Insecta: Collembola) and its
Connection to the Central Nervous System // Z. Zellforseh, 1971. Vol. 118. P.
263-282.
.30 Koch,
M. The cuticular cephalic endoskeleton of primarily wingless hexapods:
ancestral state and evolutionary changes // Pedobiologia, 2000. Vol. 44. P.
374-385.
.31 Kollmann,
M., Huetteroth, W. & Schachtner, J. Brain organization in Collembola
(springtails) // Arthropod Structure & Development, 2011. Vol. 40(4). P.
304-316.
.32 Lubbock,
J. Monograph of the Collembola and Thysanura. // 1873. Pp. Kopie.
.33 Matsuda,
R. Morphology and evolution of the insect head // Mem Am Ent Inst, 1965. Vol.
4. Pp.334.
4.34 Manton
S.M., Harding J.P. Mandibular Mechanisms and the Evolution of Arthropods //
Phil. Trans. R. Soc.
Lond. B, 1964. Vol. 247. Pp.183.
.35 Matsuda,
R. Morphology and Evolution of the Insect Abdomen // Oxford: Pergamon Press,
1976. Pp. 534.
.36 Noble-Nesbitt,
J. A site of water and ionic exchange with the medium in Podura aquatica L.
(Collembola, Isotomidae) // Journ. exp. Biol., 1963. P. 701-711.
4.37 Polilov
A.A. Anatomy of the smallest of the Coleoptera, feather-winged beetles from tribe
Nanosellini (Coleoptera, Ptiliidae) and limits to insect miniaturization //
Zool Zh, 2008. Vol.
87(2). P. 181-188 [Entomol Rev. 2008; 88(1): 26-33].
4.38 Polilov,
A.A. Small is beautiful: Features of the smallest insects and limits to
miniaturization // Annual Review of Entomology, 2015. Vol. 60. P. 103-121.
4.39 Polilov,
A.A. “Features of the Structure of Hymenoptera Associated with Miniaturization.
2. Anatomy of
Trichogramma evanescens (Hymenoptera, Trichogrammatidae)” // Zoologicheskii
Zhurnal. 2016. Vol. 95 (6). P. 699-711) [Entomological Review 96 (4), 419-431
(2016)].
.40 Polilov,
A. A. and Makarova, A. A. The scaling and allometry of organ size associated
with miniaturization in insects: A case study for Coleoptera and Hymenoptera //
Sci. Rep. 7, 43095; doi: 10.1038/ srep43095, 2017.
.41 Rensch,
B. Histological changes correlated with evolutionary changes of body size
evolution // 1948, 2:2 1. P. 8-30.
.42 Rundell,
R. J., & Leander, B. S. Masters of miniaturization: Convergent evolution
among interstitial eukaryotes // BioEssays, 2010. Vol. 32(5). P. 430-437.
.43 Rusek,
J. Biodiversity of Collembola // Biodiv. Conserv., 1988. Vol. 7. P. 1207-1219.
4.44 Schaller, F. I. Uberordnung und I. Ornung
Collembola (Springschwanze) // Handb. Zool. Berlin, 1970. Vol. ½. P.
1-72.
.45 Schneider,
C. & D’Haese, C. Morphological and molecular insights on Megalothorax: the
largest Neelipleona genus revisited (Collembola) // Invertebrate Systematics,
2013. Vol. 27. P. 317-364.
4.46 Sømme, L. Invertebrates in Hot and
Cold Arid Environments // Springer- Verlag, Berlin, 1995.
4.47 Verhoef,
H.A., Bosman, C, Bierenbroodspot, A, Boer, H.H. Ultrastructure and function of
the labial nephridia and the rectum of Orchesella-cinta (L) (Collembola) //
Cell and tissue research, 1979. Vol. 198 (2). P. 237-246.
.48 Wolter,
H. Vergleichende Untersuchungen zur Anatomie und Funktionsmorphologie der
stechend-saugenden Mundwerkzeuge der Collembolen // Zool. Jb. (Anat.), 1963.
Bd. 81. Pp. 74.
4.49 Worland
M.R. The relationship between water content and cold tolerance in the arctic
collembolan Onychiurus arcticus (Collembola: Onychiuridae) // Eur J Entomol,
1996. Vol. 93. P.
341-348.