Фитопланктон реки Щара и канала Огинского
МИНИСТЕРСТВО
ОБРАЗОВАНИЯ РЕСПУБЛИКИ БЕЛАРУСЬ
УЧРЕЖДЕНИЕ
ОБРАЗОВАНИЯ «ГРОДНЕНСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ ИМЕНИ ЯНКИ КУПАЛЫ»
Факультет
биологии и экологии
Кафедра
ботаники
Курсовая
работа
ФИТОПЛАНКТОН реки ЩАРА и каналА ОГИНСКОГО
Гродно, 2016
Оглавление
фитопланктон биологический водоросль
водоем
Введение
. Литературный обзор
.1 Понятие «планктон»
.2 Оценка состояния водных экосистем
.3 Связь биологического разнообразия
водорослей с трофностью водоема
.4 Изменение видового состава фитопланктона
при эвтрофировании водоемов
.5 Фитопланктон некоторых водоемов
.5.1 Фитопланктон реки Лососна
.5.2 Фитопланктона реки Городничанка
.5.3 Альгофлоры Августовского канала
.5.4 Состав фитобентоса на примере
каналов Украины: Днепр-Кривой Рог и Сев.Донец-Донбасс
. Материалы и методы исследования
.1 Место проведения исследований
.2 Выбор станции и методика отбора
проб фитопланктона
.3 Консервация, концентрирование
проб, обработка результатов
. Результаты исследований
.1 Видовой состав фитопланктона канала
Огинского и реки Щара
.2 Динамика видового разнообразия
фитопланктона
.3 Таксономический анализ видового
состава фитопланктона
Выводы
Библиографический список
Приложение А
Приложение Б
Введение
Водоросли (Algae), представляют огромную группу
организмов, широко распространенную на нашей планете и имеющую огромное
значение в круговороте веществ в природе и жизни человека [1].
Термин «водоросли» дословно означает «растущие в
воде». Однако растения, которые мы обычно видим растущими в реках и прудах, не
относятся к водорослям, а представляют собой высшие растения, поскольку большая
часть пресноводных водорослей имеет микроскопические размеры. Крупные водоросли
(макрофиты) преобладают в морях, причем часто обитают на больших глубинах.
Многие водоросли обитают не в воде, а в почве, на деревьях, скалах и т.п.
следовательно, водное местообитание, хотя и характерно для большинства
водорослей, но не может быть включено в их определение [2].
В отличие от многообразных водных высших
растений тело водорослей не имеет органов, дифференцированных на листья,
стебель и корни, а представлено своеобразным талломом или слоевищем, который
может быть очень разнообразным.
В настоящее время общепризнано учеными, что
водоросли не представляют собой монолитной группы организмов, объединенных
единством строения и происхождения, а представляют совокупность обособленных
отделов растений, происходящих от первичных сапрофитных организмов [1].
Водоросли играют огромную роль в природе. Они
являются основными производителями органической пищи и кислорода в водных
экосистемах Земли, и, кроме того, играют большую роль в общем балансе кислорода
на планете. В наземных местообитаниях почвенным водорослям наряду с другими
микроорганизмами принадлежит роль пионеров растительности. Водоросли учувствуют
в процессах формирования примитивных почв на субстратах, лишенных почвенного
покрова, а также в процессах восстановления почв, нарушенных сильными
загрязнениями. Водоросли принимают участие в строительстве коралловых рифов -
наиболее грандиозных геологических образований, созданных живыми организмами.
Трудно переоценить значение водорослей и для
жизни человека. Водорослям отводят важную роль в решении ряда глобальных
проблем, волнующих все человечество, в том числе продовольственной,
энергетической, охраны окружающей среды, изыскания новых источников
промышленного сырья, строительных материалов, фармацевтических препаратов,
биологически активных веществ и новых объектов биотехнологии [24].
Альгофлористические исследования в настоящее
время приобретают все большую актуальность. Значительное внимание уделяется
исследованиям альгофлоры крупных рек, однако флоры малых рек практически не
изучены. Вместе с тем обобщение и анализ данных по этим водным объектам
позволили бы выявить не только флористическое богатство водорослей в
разнотипных водоемах и водотоках, но и особенности экологии и распространения
отдельных видов и групп водорослей [25].
Фитопланктон является первым звеном трофической
цепи и одним из основных продуцентов органического вещества в водоемах. Его
структура и функциональные особенности во многом определяют структуру и
функционирование водных экосистем в целом.
Цель нашей работы - изучение сезонной динамики
планктонных водорослей реки Щары и канала Огинского.
Для достижения поставленной цели следовало
решить следующие задачи:
. изучить видовой состав фитопланктона в
отобранных пробах;
. проанализировать сезонную динамику
видового состава фитопланктона за 2015 год;
. составить список водорослей планктона
реки Щары и канала Огинского и провести таксономический анализ.
. Литературный обзор
.1 Понятие «планктон»
Фитопланктон - совокупность мелких,
преимущественно микроскопических водорослей, свободно плавающих в толще воды.
Это основная экологическая группа водорослей, продуцирующая первичное органическое
вещество, без которого невозможно представить все живое в водоеме. В процессе
эволюции планктонные водоросли выработали ряд приспособлений, позволяющих им
достаточно долгое время находиться в воде во взвешенном состоянии. У
планктонных водорослей, не имеющих жгутиков, увеличение плавучести достигается
в значительной мере соответствующей формой тела и наличием разнообразных
выростов и придатков, щетинок, роговых отростков, перепонок и др. Иные формы
планктонных водорослей представлены плоскими или полыми колониями, которые
обильно выделяют слизь. Многие водоросли накапливают в клетках вещества с
удельным весом меньше единицы (например, жир, масло) либо образуют газовые
вакуоли. Одна из особенностей планктонных водорослей, позволяющих им существовать
в толще воды во взвешенном состоянии, - мелкие размеры тела. Благодаря мелким
размерам, а, следовательно, и небольшой массе, планктонные водоросли не так
быстро опускаются на дно водоема [3].
Планктонные водоросли обитают в самых
разнообразных водоемах - от озер, водохранилищ до небольших луж. Типичный
фитопланктон особенно характерен для крупных водоемов.
В зависимости от размеров фитопланктонные
водоросли подразделяются на мезо-, микро- и нанопланктонные.
К мезопланктонным фитопланктерам относят
водоросли размером 1-5 мм. Это немногочисленная группа колониальных организмов (Sphaeronostoc
kihlmani
и др.). Водоросли с размером тела от 50 мкм до 1 мм относятся к группе
микропланктонных организмов. Нанопланктонные организмы имеют тело размером
менее 50 мкм. При отборе проб планктонной сетью они легко проходят через
мелкоячеистую ткань.
В планктоне пресных водоемов наибольшим
разнообразием отличаются зеленые, диатомовые водоросли и цианеи. Из зеленых
обильно представлены одноклеточные, ценобиальные и колониальные вольвоксовые
(виды родов Chlamydomonas,
Gonium, Pandorina,
Eudorina,
Volvox)
и хлорококковые (виды родов Pediastrum,
Scenedesmus, Oocystis,
Golenkinia,
Sphaerocystis,
Chlorella,
Kirchneriella,
Ankistrodesmus
и др.). Характерными представителями диатомовых водорослей в планктоне являются
виды родов Melosira,
Fragilaria,
Tabellaria,
Asterionella,
Cyclotella.
Из цианей часто и обильно встречаются в качестве планктеров Microcystis,
Anabaeпа,
Aphanizomenon,
Gloeotrichia
и др. Из жгутиковых форм в пресноводном планктоне обычны динофитовые - Ceratium
и Peridiniит;
из золотистых - виды родов Dinobryon,
Mallomonas,
Uroglena,
Synura
и др.; из эвгленовых - виды родов Trachelomonas,
Phacus,
Euglena
и др. Последние обильно развиваются в мелких, хорошо прогреваемых водоемах [3].
В мягкой воде заболоченных водоемов и болот
развиваются многочисленные представители десмидиевых: виды родов Closterium,
Cosmarium,
Euastrum,
Staurastrum,
Micrasterias,
Xanthidium,
Desmidium
и др.
Всего в различных водоемах и водотоках Беларуси отмечено
около 1000 видов планктонных водорослей.
Видовой состав фитопланктона и его численность
разнообразны в разных водоемах и даже в одном водоеме в разное время года, он
зависит от совокупности многих факторов. Важнейшими из них являются световой,
температурный и химический режим, а также антропогенное воздействие. Последнее
в одних случаях приводит к обеднению фитопланктона, в других - к значительному
повышению его продуктивности. При попадании в воду большого количества
биогенных веществ наблюдается бурное развитие планктонных водорослей,
окрашивающих воду в зеленый, сине-зеленый и другие цвета. Такое явление
получило название «цветение» воды, при котором в 1 л воды содержатся миллионы
клеток планктонных водорослей. В результате их массового разложения выделяются
сероводород и другие токсические вещества, что может привести к гибели
зооценозов водоема. Следует учесть и тот факт, что токсические вещества
выделяются некоторыми водорослями (например, видами Мicrocystis)
в процессе их жизнедеятельности.
Действие освещенности как экологического фактора
наглядно проявляется в вертикальном распределении фитопланктона. В озёрах,
например, планктонные водоросли обитают обычно в верхних слоях воды, но могут
развиваться и на глубине 10-15 м. Очень требовательны к освещению зеленые
водоросли и большинство видов цианей, развивающихся наиболее интенсивно в
летний период. Так, водоросли родов Microcystis,
Anabaena,
Aphanizomenon
в массе развиваются только у самой поверхности воды. Менее требовательны к
свету диатомовые водоросли. Большинство из них в мало прозрачных водах озер и
водохранилищ более интенсивно развивается на глубине 2-3 м.
Температурный фактор - также один из наиболее
важных, влияющих на состав и распределение фитопланктона. Известны виды,
которые развиваются только в холодноводных водоемах; есть виды, существующие в
водоемах с теплой водой. Многие водоросли способны обитать в водоемах, где
диапазон колебаний температуры весьма велик.
Так как температурный оптимум у разных видов
планктонных водорослей не совпадает, происходит смена видового состава по
сезонам (сезонная сукцессия). Вегетационный цикл фито планктона начинается в
марте-апреле. В это время массовыми планктерами являются мелкие жгутиковые - Chromulina,
Сrурtomonas,
повышается численность холодноводных видов диатомовых - Melosira,
Diatoma.
Во второй половине весны бурно развивается холодноводный комплекс диатомовых.
Летом появляются умеренно тепловодные диатомовые - Asterionella,
Tabellaria,
интенсивнее развиваются зеленые и синезеленые водоросли. Во второй половине
лета максимального развития достигают синезеленые и зеленые водоросли, которые
могут вызвать «цветение» вoды.
Из диатомовых в этот период отмечаются тепловодные представители родов Fragilaria
и Melosira
granulata.
Осенью снова начинают более интенсивно развиваться холодноводные диатомовые
совместно с продолжающими развитие синезелеными водорослями.
Вода природных водоемов содержит различные
химические соединения, необходимые для развития фитопланктона. Важнейшие из них
минеральные соли (биогенные элементы). Из минеральных солей для развития
фитопланктона (и водорослей в целом) необходимы соли азота и фосфора. Как
правило, в водоемах этих соединений явно недостаточно. Элементами питания
водорослей являются железо и кальций. К железолюбивым водорослям относятся
многие диатомовые, десмидиевые. Кремний нужен для формирования панциря
диатомей. Магний, калий и сера - также необходимые элементы для водорослей, но
в воде их всегда достаточно.
Планктонные водоросли являются основными, а
нередко и единственными продуцентами первичного органического вещества, которое
необходимо для существования всего живого в водоемах. Планктонные водоросли
принимают активное участие в самоочищении водоемов. Из отмирающих планктонных
водорослей формируются илы, сапропели и другие отложения. Планктонные водоросли
используются в качестве индикаторов загрязненности воды. Они могут быть
источником белков, витаминов и сырья для многих отраслей промышленности [3].
1.2 Оценка состояния водных экосистем
Водные экосистемы - это природный объект,
который является единством взаимозависимых среды и обитающей в ней биоты.
Поэтому, для того, чтобы охарактеризовать состояние водной экосистемы,
необходимо знать показатели воды как среды обитания и показатели биотической
(организменной) части экосистемы. Каждая водная экосистема находится в
определенном состоянии, которое выражается в определенном сочетании показателей
средовой и организменной составляющих водной экосистемы. Для оценки состояния
водной экосистемы необходимо знать не только ее гидрохимические параметры, но и
параметры ее биотической части [5]. Биотическая часть экосистемы организована в
виде трофической пирамиды, основу которой составляют первичные продуценты. В
водной экосистеме это водоросли, которые дают органическое вещество для
формирования второго трофического уровня - беспозвоночных консументов, которые,
в свою очередь, являются базисом для рыб - верхнего звена трофической пирамиды
в водных объектах. Зная состояние первого трофического уровня, можно
предполагать состояние всей трофической пирамиды. Таким образом, по состоянию
первого трофического звена возможна оценка всей биотической составляющей водной
экосистемы. Способность поверхностных вод к самоочищению также определяется
эффективностью функционирования биотической составляющей экосистемы. Таким
образом, по состоянию первого трофического звена возможна оценка состояния всей
биотической составляющей водной экосистемы. Способность поверхностных вод
самоочищению также определяется эффективностью функционирования биотической
составляющей экосистемы.
Оценка состояния первого трофического звена
возможна с помощью показателей численности и биомассы, биохимического
потребления кислорода (БПК), насыщенности воды кислородом, прозрачности воды и
связанной с прозрачностью первичной продукции, а также индекса сапробности [6].
Однако, монотонность изменения гидрохимических показателей с ростом классности
воды не дает возможности выделить какие-либо критические уровни, значения,
интервалы, что подтверждает необходимость привлечения показателей, более тесно
связанных с биотической частью экосистемы.
Одним из компонентов в оценке состояния водных
экосистем является совокупность организмов и результатов проявления их
жизнедеятельности. Качество вод по обитающим в них организмам определяется уже
в течение сотни лет и метод носит название - биоиндикация. Его следует отличать
от биотестирования как метода оценки реакции тест-организма на среду, в которую
его поместили.
Пока нет представления о едином механизме
функционирования водной экосистемы, а для оценки ее состояния необходимо
понимание функционирования экосистемы в целом как взаимозависимого единства
среды и обитающей в ней биоты и выявления тех показателей, которые изменяются
не монотонно и позволяют определить емкость экосистемы и пределы ее
восстановительных способностей.
1.3 Связь биологического разнообразия водорослей
с трофностью водоема
Биоиндикационные методы на основе видового
состава сообществ и обилия водорослей дают интегральную оценку результатов всех
природных и антропогенных процессов, протекавших в водном объекте. Кроме того,
биоиндикация по сообществам водорослей - дешевый экспресс-метод, в то время как
химические анализы дорогостоящи, а основным преимуществом автотрофов является
то, что водоросли первыми в трофической цепи реагируют на загрязнители, не
успевая их накапливать. Реакцией на изменение условий среды является изменение
состава и обилия водных организмов, причем смена сообщества водорослей может
произойти за несколько часов при смене условий среды [7].
Методы биоиндикации по высшему трофическому
звену наземных экосистем бассейна водосбора еще не достаточно разработаны.
Биоиндикационные оценки по низким трофическим уровням используются довольно
широко [8, 9]. Хотя есть много попыток сопоставить среду и биоту, но пока нет
единой системы. Единая система оценки качества вод и состояния водных экосистем
по сообществам водорослей-индикаторов дает оценку качества воды, устанавливает
интервалы важнейших химических, физических и продукционных показателей в соответствии
с состоянием биоты, дает возможность восстановить недостающие показатели по
имеющимся, позволяет определить, подвержена ли экосистема антропогенному
воздействию, на какой стадии естественной или антропогенной сукцессии она
находится и каковы перспективы ее изменений, вплоть до оценки кризисности
ситуации.
.4 Изменение видового состава фитопланктона при
эвтрофировании водоемов
Структура и продуктивность фитопланктона зависит
не столько от климатических особенностей региона, сколько от уровня поступления
биогенных элементов и условий перемешивания, связанных с морфометрией водоемов
и их проточностью.
В процессе эвтрофирования водоемов наряду с
общим повышением биомассы фитопланктона и сменой доминирующих видов происходит
изменение и сезонной сукцессии фитопланктона.
Сезонное развитие фитопланктона связано с тем,
что массовое развитие какого-либо вида вызывает изменение среды (например,
истощение биогенных и иных элементов, изменение рН, поступление в среду тех или
иных органических соединений и др.), что является предпосылкой для развития
других водорослей. Внезапное увеличение численности одного из доминирующих
видов или даже субдоминантов, свидетельствует о нарушении стабильности
экосистемы и может служить показателем начальной стадии эвтрофирования [6].
По мере повышения трофности водоемов происходит
увеличение биомассы синезеленых, а также повышается их доля в общей массе
фитопланктона. То же самое происходит и с эвгленовыми водорослями; их биомасса
максимальных значений достигает в высокотрофных озерах, а наибольшая доля в
биомассе планктона - в загрязненных водоемах. Возрастает роль и значение
зеленых, в основном, хлорококковых водорослей.
Наоборот, доля диатомовых и золотистых в общей
массе фитопланктона снижается, хотя абсолютные величины их биомасс в эвтрофных
водоемах все же выше, чем в мезотрофных [10].
.5 Фитопланктон некоторых водоемов
1.5.1 Фитопланктон реки Лососно
Лососно, река в Гродненском районе, левый приток
Немана. Относится к категории малых рек, ее длина составляет 46 км. Площадь водосбора
- 468 км. Средний наклон водной поверхности - 1,1 %. Начинается в Польше,
недалеко от деревни Брузги, пересекает государственную границу, впадает в реку
Неман в западной окраине города Гродно. Долина извилистая, пойма прерывистая, в
верхнем течении открытая [4].
Всего в перифитоне реки Лососно было выявлено
137 видов водорослей из 6 отделов, 11 классов, 19 порядков, 32 семейств, 49
родов. Bacillariophyta - группа не только разнообразна флористически
(общее число видов - 72,2%), но и постоянно преобладает численно. Наибольшим
видовым разнообразием отличаются роды Navicula (15), Pinnularia и
Cymbella - по 9 видов, Achnanthes - 7 видов. Chlorophyta -
вторая по таксономическому разнообразию группа (15,3%). Наиболее богаты видами
рода Closterium и Scenedesmus (по 3 вида). Отделы Cyanophyta
и Euglenophyta включают в себя 5 родов (9 видов) и 3 рода (5 видов)
соответственно. Отделы Cryptophyta и Xanthophyta представлены
двумя и одним видом соответственно [6].
Выявлена тройка классов с самой высокой видовой насыщенностью.
Первое место занимает класс Pennatophyceae - сюда входит 92 вида, что
составляет 72,7% от всех выявленных видов (они относятся к 22 родам, 10
семействам и 2 порядкам). На втором месте класс Hormogoniophyceae с 9
видами. А на третьем месте - Conjugatophyceae (8 видов) [15].
.5.2 Фитопланктона реки Городничанка
В реке Городничанка за все время исследования
было выявлено 40 видов водорослей, которые относятся к 6 отделам, 11 классам,
16 порядкам, 24 семействам, 40 родам. Наиболее представительным классом
является Pennatophycea, в него входит 2 порядка, 7 семейств, 19 родов, 19
видов.
К отделу Bacillariophyta относится - 20
видов (50% от общего количества), Chlorophyta - 14 видов (35% от общего
количества) Euglenophyta - 2 вида (5% от общего количества), Cyanophyta
- 2 вида (5% от общего количества), Xanthophyta - 1 вид (2,5% от общего
количества), Dynophyta - 1 вид (2,5% от общего количества) [14].
.5.3 Альгофлора Августовского канала
За все время исследования Августовского канала
было выявлено 41 вид водорослей [12].
Таксономический анализ результатов отбора проб
различных экологических группировок представлен в таблице 3.
Таблица 3
Таксономическая структура видового состава
альгофлоры
|
Отдел
|
Класс
|
Порядок
|
Семейство
|
Род
|
Вид
|
|
Bacillariophyta
|
Centrophyceae
|
3
|
3
|
3
|
3
|
|
|
Pennatophyceae
|
2
|
8
|
13
|
22
|
|
Chlorophyta
|
Protococcophyceae
|
2
|
3
|
3
|
3
|
|
|
Ulothrichophyceae
|
1
|
2
|
2
|
2
|
|
|
Conjugatophyceae
|
2
|
3
|
3
|
3
|
|
Euglenophyta
|
Euglenophyceae
|
1
|
1
|
2
|
3
|
|
Chrysophyta
|
Chrysophyceae
|
1
|
1
|
1
|
1
|
|
Cyanophyta
|
Chroococcophyceae
|
1
|
1
|
1
|
2
|
|
|
Hormogoniophyceae
|
1
|
2
|
2
|
2
|
|
Общее
количество:
|
14
|
24
|
30
|
41
|
Класс Pennatophyceae обладает самой
высокой видовой насыщенностью видами - в него входит 22 вида, что составляет
почти половину от всех выявленных видов (они относятся к 13 родам, 8 семействам
и 2 порядкам). Остальные классы имеют по одинаковому числу видов. Это следующие
классы: Centrophyceae, Protococcophyceae, Conjugatophyceae, Euglenophyceae
- по 2-3 вида.
В альгофлоре преобладают представители отделов Bacillariophyta
- 25 видов (% от общего количества) и Chlorophyta - 8 видов (% от общего
количества). Представители отдела Cyanophyta, Euglenophyta, Chrysophyta
выявлены в меньшем количестве.
.5.4 Состав фитобентоса на примере каналов
Украины: Днепр-Кривой Рог и Северный Донец-Донбасс
Формирование флористического состава фитобентоса
в каналах зависит как от поступления водорослей из водоисточников, так и
особенностей самих каналов: их местоположения, характера ложа, почв окружающих
территорий и т.п., причем специфика каналов имеет даже большее значение, чем
водоисточник, как это видно на примере каналов Украины. В облицованных каналах
Днепр-Кривой Рог и Северный Донец-Донбасс, имеющих совершенно различные
водоисточники, коэффициент флористической общности достигает 61,3%, тогда как в
каналах Днепр-Кривой Рог и Краснознаменском, имеющих один водоисточник, но
отличающихся характером ложа, он значительно ниже - 52,8%. Примерно такая же
степень сходства фитобентоса каналов с разными водоисточниками и разным
характером ложа - Сев.Донец-Донбасс и Краснознаменского. В каналах различных
географических областей нередко встречаются одни и те же виды. Вместе с тем
коэффициент флористической общности все-таки не очень велик (около 20-30%).
На разных твердых покрытиях - бетоне и щебне - в
условиях одного канала поселяются одинаковые виды водорослей, что обусловливает
высокую флористическую общность группировок водорослей на них (коэффициент
свыше 80%). В то же время видовой состав донных сообществ водорослей,
развивающихся на твердых субстратах и грунте в одном и том же канале,
отличается значительно сильнее [13].
Степень изменчивости флористического состава
фитобентоса по трассе каналов различна. При сохранении примерно одинаковых
условий на всем протяжении канала видовой состав фитобентоса остается в
значительной мере идентичным. Такая картина характерна для облицованных каналов
(Днепр-Кривой Рог и Сев.Донец-Донбасс), где коэффициент общности на разных
участках достигает 78-80%. В необлицованных (Краснознаменском и Ингулецком),
где сильнее сказывается влияние местных грунтов, видовой состав фитобентоса
варьирует в большей степени: коэффициент колеблется в пределах 58-68%. Из
ведущих отделов наиболее вариабилен видовой состав зеленых, особенно
протококковых и десмидиевых.
Сезонная динамика видового разнообразия фитобентоса
существенно отличается в разных каналах. Что касается флористических спектров
фитобентоса, то во многих каналах проявляется общераспространенная тенденция к
увеличению роли синезеленых и зеленых водорослей в летний период, а диатомовых
- весной и осенью.
Каналам свойственна интенсивная вегетация как
макро-, так и микрофитобентоса. Из макроскопических водорослей ведущее место
занимают представители зеленых: виды родов Cladophora, Vaucheria,
Chara, Spirogyra,
Enteromorpha, Stigeoclonium, Oedogonium, Ulothrix, Hydrodictyon
и т.д., реже встречаются другие водоросли, например желтозеленые (виды рода Tribonema),
красные [13].
Чрезвычайно широко распространены в каналах
различного назначения и географического положения виды рода Cladophora (C.
fracta, C. glomerata и др.). Обильная вегетация кладофор отмечается как в
равнинных, так и в горных и предгорных каналах. Виды рода Spirogyra
(S. crassa, S. varians, S. weberi Kuetz. и многие другие) также очень
характерны для каналов, особенно для участков с замедленным течением воды: они
интенсивно развиваются в прибрежьях, среди высших водных растений, в дренах и
т.п. Вегетации спирогир способствует высокое содержание биогенных веществ в
воде и накопление органических соединений в грунтах. Поэтому в тех каналах, где
в процессе эксплуатации происходит накопление ила, может наблюдаться
постепенное возрастание их количества. Обильное развитие Hydrodictyon reticulatum
в каналах имеет место обычно также при слабой поточности и большом количестве
биогенных элементов. Энтероморфа предпочитает каналы с повышенной
минерализацией воды (например, Северный Донец-Донбасс, устьевые области
коллекторов и т.п.). Вошерия достигает максимального развития в дренах и
начальных участках коллекторов с небольшой глубиной воды и слабой проточностью.
Интенсивная вегетация видов рода Ulothrix, как и в других водоемах,
приурочена к низкой температуре воды: в северных и горных каналах она
наблюдается летом, а в равнинных и предгорных каналах средних и южных широт
главным образом весной или даже зимой (например, каналы Альберта, Средней
Азии). Аналогичная закономерность свойственна и Bangia, также являющейся
холодноводной формой. Обильное развитие харовых зарегистрировано в каналах
Средней Азии и Закавказья при наличии условий, благоприятствующих закреплению
растений в грунте.
Необходимо иметь в виду, что в сообществах
зеленых нитчаток (особенно видов рода Cladophora) развиваются в большом
количестве многочисленные микроскопические формы водорослей из диатомовых,
синезеленых, зеленых и др. Эпифиты поселяются на поверхности нитей, иногда
покрывая их сплошным чехлом [13].
Степень развития зеленых нитчатых водорослей
зависит от специфики каналов. В равнинных облицованных каналах создаются
исключительно благоприятные условия для их вегетации благодаря наличию твердых
субстратов, умеренной скорости течения, достаточной прозрачности воды,
богатства биогенными веществами. Здесь сообщества зеленых нитчаток развиваются
чрезвычайно интенсивно с максимумом в летний период и образуют большую
биомассу. В канале Северный Донец-Донбасс их сырой вес достигал 2,8 кг/м2
(сухой вес 560 г/м2), а разовые запасы на 100 км открытой части
канала составляли 200-300 г/м2. Однако в этом же канале в годы с
повышенной мутностью воды в летний период количество зеленых нитчаток резко падает.
В каналах различных типов интенсивно развиваются
сообщества микрофитобентоса. Следует подчеркнуть, что во многих облицованных
каналах происходит заиливание твердых покрытий и нередко слой иловых отложений
достигает значительных размеров. Это обстоятельство способствует совместному
развитию в донных сообществах перифитонных и собственно бентосных форм
водорослей и влечет за собой сглаживание отличий между водорослевыми
группировками облицованных и необлицованных каналов. Необлицованные каналы с
каменистым ложем аналогичны каналам, облицованным щебнем.
Основную роль в микрофитобентосных группировках
различных каналов, как правило, играют синезеленые (виды родов Oscillatoria,
Phormidium, Lyngbya, Spirulina и др.) и диатомовые (виды родов Navicula,
Nitzschia, Gyrosigma, Cymatopleura, Cymbella, Synedra и многие другие)
водоросли. Как те, так и другие могут образовывать чистые ценозы или же
развиваться совместно. В последующем случае нередко синезеленые преобладают по
численности, а диатомовые по биомассе. Зеленые обычно играют значительно
меньшую роль, хотя встречаются в микрофитобентосе довольно регулярно. Чаще
всего это бывают протококковые и десмидиевые, реже вольвоксовые [11].
Представители остальных отделов водорослей
(эвгленовых, пирофитовых, желтозеленых, золотистых) встречаютя в донных
биоценозах каналов нерегулярно и, как правило, в небольшом количестве. Из них
обращают на себя внимание эвгленовые, которые в некоторых каналах отмечаются
систематически и иногда играют значительную роль, особенно по образуемой
биомассе. Они могут попадать в каналы и придаточных водоемов или со сбросными
водами с полей.
Необходимо отметить, что многие зеленые (в
частности, вольвоксовые), эвгленовые, золотистые, пирофитовые являются
обитателями в основном придонного слоя воды, и в пробах грунта, взятых без
этого слоя, их несравненно меньше. Особо следует остановиться на протококковых,
экология которых изучена крайне недостаточно, и среди которых, несомненно,
имеются, как планктонные, так и бентосные формы. Об их обильной вегетации на
дне водоемов различного типа неоднократно упоминается в литературе. И в каналах
некоторые протококковые (виды родов Pediastrum, Scenedesmus, Coelastrum)
нередко встречаются в бентосе даже в большом количестве, чем в планктоне.
Доминирующие комплексы микрофитобентоса каналов
представлены преимущественно синезелеными и диатомовыми водорослями.
Спорадически в их состав включаются также представители зеленых (вольвоксовых,
протококковых и десмидиевых), а иногда и других отделов (например, эвгленовых).
Синезеленые в целом являются постоянными компонентами ведущих комплексов
микрофитобентоса. Однако состав этих водорослей варьирует в различных каналах.
2. Материалы и методы исследования
.1 Место проведения исследований
Щара - второй по протяженности приток Немана
после Вилии и первый подлине в пределах Беларуси. Щара образует речной бассейн
площадью 6990 км2. Общее падение реки относительно небольшое и составляет 77,7
м. Соответственно и средний уклон водной поверхности имеет среднее значение - 0,2
промилле. Водосборный бассейн имеет треугольную форму с широким (более 100 км)
основанием в южной части. Эта часть границы водосбора Щары является
Черноморско-Балтийским водоразделом. Щара не имеет крупных притоков, самые
протяженные из них - Мышанка и Исса (справа) и Гривда (слева). Общая
протяженность речной системы Щары составляет 2650 км. Река относится к
Неманскому гидрологическому району. По среднему показателю уровня загрязнения
поверхностных вод Щара относится к категории относительно чистой (ИЗВ менее 1),
однако для отдельных участков речной долины характерен сравнительно небольшой
уровень загрязнения цезием-137 (Рисунок 1 в ПРИЛОЖЕНИИ А).
Огинский канал
<#"896319.files/image001.gif">
Рисунок 3.1 Динамика видового разнообразия
фитопланктона канала Огинского
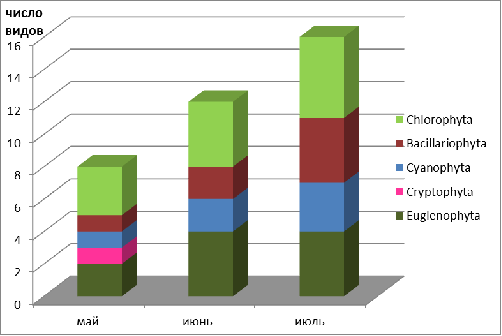
Рисунок 3.2 Динамика видового разнообразия
фитопланктона реки Щара
3.3 Таксономический анализ видового состава
фитопланктона
В исследуемых образцах фитопланктона выявлено 48
и 21 виды водорослей. Систематический список представлен в ПРИЛОЖЕНИИ Б.
Систематическая принадлежность водорослей указана в соответствии с
«Таксономическим каталогом» Т.М. Михеевой [23].
Таксономический анализ результатов обработки
проб водорослей канала Огинского в таблице 3.3.
Таблица 3.3
Таксономический спектр
фитопланктона канала Огинского
|
Отдел
|
Класс
|
Порядок
|
Семейство
|
Род
|
Вид
|
|
Chlorophyta
|
Protococcophyceae
|
1
|
6
|
12
|
22
|
|
|
Volvocophyceae
|
2
|
3
|
4
|
4
|
|
|
Conjugatophyceae
|
1
|
1
|
1
|
1
|
|
|
Siphonocladophyceae
|
1
|
1
|
1
|
1
|
|
Bacillariophyta
|
Pennatophyceae
|
2
|
3
|
5
|
8
|
|
Cyanophyta
|
Hormogoniophyceae
|
2
|
2
|
3
|
3
|
|
|
Сhroococcophyceae
|
1
|
1
|
1
|
1
|
|
Cryptophyta
|
Cryptophyceae
|
1
|
1
|
1
|
1
|
|
Chrysophyta
|
Chrysophyceae
|
1
|
1
|
1
|
1
|
|
Euglenophyta
|
Euglenophyceae
|
1
|
1
|
3
|
5
|
|
Общее
количество:
|
13
|
20
|
30
|
47
|
Класс Protococcophyceae обладает самой
высокой видовой насыщенностью - сюда входит 22 вида, что составляет почти
половину от всех выявленных видов (они относятся к 12 родам, 6 семействам и 1
порядку). Самым многочисленным является порядок Chlorococcales.
Он включает в себя 6 семейств, среди которых наиболее часто встречались виды
родов Scenedesmus
и
Ankistrodesmus.
Следующим по видовой насыщенности является класс: Pennatophyceae - 8
видов, большинство представителей данного класса относится к таким родам как Fragilaria.
Далее следует Euglenophyceae
-
5 видов и Volvocophyceae - 4 вида. Hormogoniophyceae - 3 вида.
Самыми немногочисленными оказались: Chrysophyceae, Cryptophyceae,
Сhroococcophyceae, Siphonocladophyceae, Conjugatophyceae по 1
виду каждый.
Разнообразие различных классов в фитопланктоне
канала Огинского представлено на рисунке 3.4.
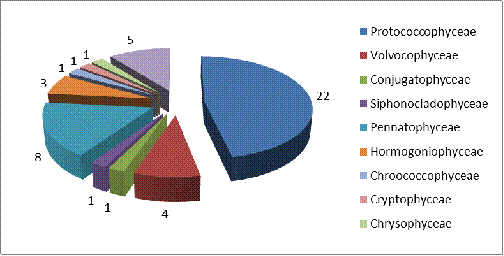
Рисунок 3.4 Соотношение видового разнообразия
различных классов в фитопланктоне канала Огинского
Таксономический анализ результатов обработки
проб водорослей реки Щара в таблице 3.5.
Таблица 3.5
Таксономический спектр фитопланктона реки Щара
|
Отдел
|
Класс
|
Порядок
|
Семейство
|
Род
|
Вид
|
|
Chlorophyta
|
Protococcophyceae
|
1
|
3
|
3
|
3
|
|
|
Ulothrichophyceae
|
1
|
1
|
1
|
1
|
|
|
Siphonocladophyceae
|
1
|
1
|
1
|
2
|
|
Bacillariophyta
|
Pennatophyceae
|
2
|
3
|
3
|
4
|
|
Cyanophyta
|
Hormogoniophyceae
|
1
|
1
|
1
|
1
|
|
|
Сhroococcophyceae
|
1
|
2
|
2
|
2
|
|
Cryptophyta
|
Cryptophyceae
|
1
|
1
|
1
|
1
|
|
Euglenophyta
|
Euglenophyceae
|
2
|
3
|
5
|
7
|
|
Общее
количество:
|
10
|
15
|
17
|
21
|
Класс Euglenophta
обладает самой высокой видовой насыщенностью - сюда входит 7 видов (они относятся
к 5 родам, 3 семействам и 2 порядкам). Наиболее часто встречались виды родов Trachelomonas
и
Lepocinalis.
Следующим по видовой насыщенности является класс: Pennatophyceae - 4
вида, большинство представителей данного класса относится к таким родам как Navicula.
Далее следует Protococcophyceae - 3 вида, Siphonocladophyceae и
Сhroococcophyceae по 2 вида каждый. Самыми немногочисленными оказались: Cryptophyceae,
Hormogoniophyceae и Ulothrichophyceae по 1 виду каждый.
Разнообразие различных классов в фитопланктоне
реки Щара представлено на рисунке 3.6.
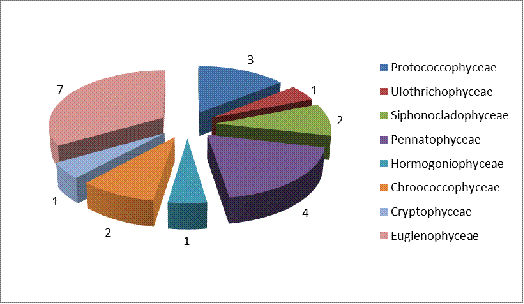
Рисунок 3.6 Соотношение видового разнообразия
различных классов в фитопланктоне реки Щара
Выводы
1. За период исследования
в фитопланктоне канала Огинского и реки Щара было выявлено 47 ( из 6 отделов 10
классов 13 порядков 20 семейств 30 родов) и 21 (из 5 отделов 8 классов 10
порядков 15 семейств 17 родов) вид соответственно.
. На канале Огинского и
реке Щаре выявлены наиболее частые представители фитопланктона, которые относятся
к отделу Chlorophyta, немного меньше к отделу Bacillariophyta,
далее следуют отделы Euglenophyta и Cyanophyta.
. Сезонная динамика
видового богатства фитопланктона канала Огинского и реки Щара характеризуется
пиком развития в июле.
Библиографический список
1. Миловидова, Л.С. Водоросли /
Л.С. Миловидова. Томск: Томск. ун-та, 1982. 164 с.
. Дьяков, Ю.Т. Введение в
альгологию и микологию / Ю.Т. Дьяков. Москва: МГУ, 2000. 192 с.
. Альгология и микология:
учеб.пособие / А.С. Шуканов [ и др.]; под общ. ред. А.С. Шуканова. Минск: БГУ,
2009. 423 с.
. Капкова, В.И. Практическая
гидробиология / под ред. В.Д. Федорова и В.И. Капкова. М.:Изд-во «ПИМ», 2016.
367 с.
5. баринова,
С.С. Полиморфизм соединительных структур диатомовых водорослей / С.С. Баринова.
Владивосток: ДВО АН СССР, 1988. 205 с.
. Садчиков, А.П. Методы
изучения пресноводного фитопланктона / А.П. Садчиков. «Университет и школа»,
2003. 157 с.
. Баринова, С.С. Атлас
водорослей - индикаторов сапробности (российский Дальний Восток) / С.С.
Баринова, Л.А. Медведева. Владивосток: Дальнаука, 1996. 364 с.
. Макрушин, А.В.
Биологический анализ качества вод / А.В. Макрушин. ЗИН АН СССР, 1974. 58 с.
. Совещание руководителей
водохозяйственных органов стран - членов СЭВ. Унифицированные методы
исследования качества вод. Москва, 1977. 91 с.
. Вассер, С.П. Водоросли.
Справочник / С.П. Вассер [ и др.]. Киев: Наук. думка, 1989. 608 с.
. Кутикова, Л.А. Определитель
пресноводных беспозвоночных Европейской части СССР (планктон и бентос) / под
ред. Л.А. Кутиковой и Я.И. Старобогатовой. Л.: Гидрометиоиздат, 1977. 233 с.
. Августовский канал / А.М.
Лосминский [и др.] // Оформил УП «Издательство Беларусь». 2006. 74 с.
. Липин, А.Н. Пресные воды и
их жизнь / А.Н. Липин. М.:Учпедгиз, 1950. 247 с.
. Максимова, С.Е. Таксономическая
и эколого-географическая характеристика фитопланктона реки Муховец / С.Е.
Максимова, Н.С. Прибыловская // Актуальные научно-технические и экологические
проблемы сохранения среды обитания: научные статьи Международн. науч.-практ.
конф., Брест 23-25 апр. 2014.: в 4-х частях. Ч. Ш. / УО «Брестск. гос. техн.
ун-т.»; под ред. А.А. Волчека [и др.]. Брест, 2014. С. 184-186.
. Жизнь растений, т. 3:
Водоросли и лишайники. М., Просвещение, 1977.
16. Мешкова, Н.А. Зеленые водоросли.
Желтозеленые водоросли. Определитель пресноводных водорослей СССР. Л.: Наука,
1986. 360 с.
17. Топачевский, А.В. Пресноводные
водоросли Украинской ССР / А.В. Топачевский, Н.П. Масюк // под ред. А.В.
Топачевского - Киев: Вища школа, 1984. 336 с.
. Царенко, П.М. Краткий определитель
хлорококковых водорослей Украинской ССР / П.М. Царенко. Киев: Наук. думка,
1990. 208 с.
. Голлербах, М.М. Синезеленые
водоросли. Определитель пресноводных водорослей СССР. Вып. 2. / М.М. Голлербах,
Е.К. Косинская, В.И. Полянский. Москва, 1953. 652 с.
. Криштофович, А.Н. Диатомовый
анализ. Кн. 2. Определитель ископаемых и современных диатомовых водорослей.
Порядки Centrales и Mediales. // Под общей ред. А.Н. Криштофовича. Госуд.
изд-во геологич. лит-ры, 1949.
. Криштофович, А.Н. Диатомовый
анализ. Кн. 3. Определитель ископаемых и современных диатомовых водорослей.
Порядок Pennales. // Под общей ред. А.Н. Криштофовича. Госуд. изд-во геологич.
лит-ры, 1950.
. Михеева, Т.М. Методы
количественного учета нанофитопланктона (обзор) / Т.М. Михеева // Гидробиол.
журнал. 1989, Т. 25, № 4. С. 3-21.
. Михеева, Т.М. Альгофлора Беларуси.
Таксономический каталог / Т.М. Михеева. Мн.: БГУ, 1999. 396 с.
. Экологическая ситуация как фактор
влияющий на тенденцию формирования фитопланктона [Электронный ресурс]. Режим
доступа: http//lifeuderwater.ru/?p=156. Дата доступа: 06.03.2016.
Приложение А
Рисунок 1 Географическое положение станций
отбора проб

Рисунок 2 Месторасположение станции №1

Рисунок 3 Месторасположение станции №2
Приложение Б
Сводный список водорослей,
выявленных в фитопланктоне канала Огинского
отд. Chlorophyta
кл. Protococcophyceae
пор. Chlorococcales
сем. Scenedesmaceae
Oltmanns
1. Scenedesmus quadricauda
(Turp.) Breb. var. quadricauda (=Sc. quadricauda var. maximus W. et G.S.West)
2. Scenedesmus acuminatus
var. bernardii (Smiss.) Deduss
3. Scenedesmus denticulatus
Lagerh. var. denticulatus
4. Tetrastrum
staurogeniaeforme (Scrod.) Lemm. var. staurogeniaeforme
5. Tetrastrum puncatatum
(Schmidle) Ahlst. et Tiff.
6. Crucigenia apiculata
(Lemm.) Schmidle
7. Crucigenia rectangularis
(A.Br.) Gay
8. Crucigenia tetrapedia
(Kirchn.) W. et G.S.West
9. Monoraphidium contortum
(Thur.) Kom.Legn.
10. Monoraphidium irregular
(G.M.Smith) Kom.-Legn.
11. Monoraphidium obtusum
(Korsch.) Kom.-Legn.
сем.
Coelastraceae
12. Coelastrum microporum Nag.
сем.
Parmellaceae
13. Sphaerocystis
schroeteri Chod.
сем.
Oocystaceae Bohlin
14. Oocystis solitaria Wittr.
var. solitaria
15. Oocystis submarina Lagerh.
16. Oocystis pussilla Hansg.
17. Lagerheimia genevensis
Chod. var. Genevensis
сем. Ankistrodesmaceae
Korschik.
18. Ankistrodesmus
bibrianus(Reinsch) Korschik.
19. Didymogenes inconspicua
Korsch.
20. Kirchneriella obese (West)
Sehmidle var. obesa
сем.
Hydrodictiaceae (O. F. Gray) Dumortier
21. Pediastrum duplex Meyen
var. duplex (=P. duplex var. clathratum (A. Br.) Lagerh.)
22. Pediastrum duplex Meyen
var. Duplex
кл.
Volvocophyceae
пор.
Volvocales
сем.
Volvocaceae
23. Gonium sociale (Duj.)
Warming
пор. Ulothrichales
сем. Elakatothrichaceae
24. Elakatothrix genevensis
(Reverd.) Hindak
25. Koliella longiseta
(Vischer) Hindak
сем. Ulothrichaceae
26. Clorhormidium flaccidum
(Kutz.) Fott
кл.
Conjugatophyceae
пор.
Desmidiales
сем.
Closteriaceae Nitzsch.
27. Closterium kuetzingii
Breb. var. kuetzingii
кл. Siphonocladophyceae
пор. Cladophorales
сем. Cladophoraceae
28. Rhizoclonium
hieroglyphicum (Ag.) Kutz. var. hieroglyphicum
отд.
Bacillariophyta
кл.
Pennatophyceae
пор.
Raphales
сем.
Naviculaceae
29. Pinnularia viridis
(Nitzsch.) Ehr. var. viridis
30. Pinnularia borealis var.
brevicostata Hust.
31. Navicula Bory sp.
32. Navicula radiosa Kütz.
var. radiosa (= N. radiosa var. acuta (W. Sm.) Grun.)
пор.
Araphales
cем.
Fragilariaceae (Kütz.)
D. T.
33. Fragilaria
bicapitata A. Mayer var. bicapitata
34.
Fragilaria bicapitata var. lineolata Moiss.
35.
Fragilaria virescens var. elliptica Hust.
cем.
Tabellariaceae Schütt
36.
Tabellaria fenestrata (Lyngb.) Kütz.
var. fenestrata (=T. Fenestrata var. asterionelloides Grun.)
отд.
Cyanophyta
кл.
Hormogoniophyceae
пор.
Nostocales
сем.
Rivulariaceae
37. Gloeotrichia
natans (Gedw.) Rabenh
пор.
Oscillatoriales
сем.
Oscillatoriaceae (Kirchn.) Elenk.
s. strict.
38. Oscillatoria
limosa Ag. f. Limosa
39. Lyngbya
aestuarii (Mert.) Liebm. f. aestuarii
кл. Сhroococcophyceae
пор. Chroococcales
сем. Microcystidaceae
40. Microcystis
pulverea (Wood.) Elenk. f. pulvera
oтд.
Cryptophyta
кл.
Cryptophyceae
пор.
Cryptomonadales
сем.
Cryptomonadaceae
41. Cryptomonas
Ehr. sp
отд.
Chrysophyta
кл.
Chrysophyceae
пор.
Chromulinales
сем.
Chromulinaceae
42.
Chrysococcus rufescens Klebs. var.
rufescens
отд.
Euglenophyta
кл.
Euglenophyceae
пор.
Euglenales
сем.
Euglenaceae Klebs
43. Euglena gracilis Klebsf.
Gracilis
44. Euglena pisciformis Klebs
45. Lepocinalis ovum (Ehr.) Mink.
var. ovum
46. Lepocinalis ovum var.
dimidio-minor Defl.
47. Urceolus
cyclostomus (Stein) Mereschk.
Сводный список водорослей,
выявленных в фитопланктоне реки Щара
отд. Chlorophyta
кл.
Ulothrichophyceae
пор. Ulothrichales
сем. Ulothrichaceae
1.Clorhormidium flaccidum (Kutz.)
Fott
сем. Elakatothrichaceae
2.Koliella longiseta (Vischer)
Hindak
сем. Microsporaceae
3.Binuclearia tectorum (Kuetz.)
Beger
кл.
Protococcophyceae
пор.
Chlorococcales
сем.
Parmellaceae
4. Planctococcus
sphaerocystistiformis Korsck.
кл. Siphonocladophyceae
пор. Cladophorales
сем. Cladophoraceae
5.Cladophora gromerata (L.)
Kuetz. var. Glomerata
6. Closteum lunula
(Mull.) Nitzsch. f. Lunula
отд.
Bacillariophyta
кл.
Pennatophyceae
пор.
Raphales
сем.
Naviculaceae
7. Navicula interrupta W. Sm.
8. Navicula pupyla Kuetz.
пор.
Araphales
cем.
Fragilariaceae (Kütz.)
D. T.
9. Fragilaria
bicapitata A. Mayer var. bicapitata
сем.
Diatomaceae Dumortier
10. Diatoma vulgaris Bory
Morphotyp vulgaris
отд.
Cyanophyta
кл.
Hormogoniophyceae
пор.
Oscillatoriales
сем.
Oscillatoriaceae (Kirchn.) Elenk.
s. strict.
11. Oscillatoria
limosa Ag. f. Limosa
кл. Сhroococcophyceae
пор. Chroococcales
сем.
Woronichiniaceae Elenk.
12. Woronichia naegelina (Ung.)
Elenk. f. naegeliana
сем.
Coelosphaeriaceae Elenk.
13. Coelosphaerium kuetringianum
Nag. f. kuetzingianum
oтд.
Cryptophyta
кл.
Cryptophyceae
пор.
Cryptomonadales
сем.
Cryptomonadaceae
14. Cryptomonas curvata Ehr.
отд.
Euglenophyta
кл.
Euglenophyceae
пор.
Euglenales
сем.
Euglenaceae Klebs
15. Trachelomonas intermedia
Dang. f. intermedia
16. Trachelomonas volvocina Ehr.
var. volvocina
17. Lepocinalis ovum (Ehr.) Mink.
var. ovum
18. Lepocinalis ovum var.
dimidio-minor Defl.
19. Euglena viridis Ehr. f.
viridis
сем.
Menoidiaceae
20. Menoidium minimum Matvienko
пор.
Peramatales
сем.
Petalomonadaceae
21. Notosolenus apocamptus Stokes
em. Skyja sp.