Изменение пространственной структуры популяции лесной мыши в антропогенном ландшафте
Содержание
Введение
. Анализ литературных данных о
пространственно-демографической структуре популяций мелких млекопитающих
.1 Структура и понятие о популяции популяций
.2 Семейство мышиные (Muridae)
. Район, материалы и методы исследования
.1 Характеристика района исследования
.2 Материалы и методы исследования
. Объект исследования
. Динамика численности и демографическая структура населения
лесной мыши
. Пространственная структура населения лесной мыши
Заключение и выводы
Список использованных источников
Введение
Актуальность темы. Антропогенное воздействие в последнее время
приобретает все более угрожающий характер. Каждый вид в процессе эволюции
приспособился к определенному сочетанию факторов среды, в пределах которых он
может нормально существовать. Изменение параметров, обусловленное
бесконтрольной хозяйственной деятельностью приводит к нарушениям сначала
скрытым (биохимическим, физиологическим), потом визуально наблюдаемым -
морфологическим, биоритмическим, а при летальных концентрациях к гибели
организмов, популяций отдельных видов и в целом биоценозов [1].
Важнейшими показателями достаточно полно характеризующими степень
благополучия существования популяций мелких млекопитающих, их биологический
успех и приспособительную пластичность являются популяционно-экологические и
демографические, такие как численность, плотность, характер пространственного
распределения особей, амплитуда динамики численности, возрастная и половая
структура, рождаемость, смертность [2,3,4,5,6,7].
Исследования популяций мелких млекопитающих в условиях антропогенного
пресса, нарушающего естественные жизненные циклы организмов, свидетельствуют,
что популяционно-экологические и демографические характеристики в ряду
биологических показателей являются наиболее чуткими, достаточно хорошо
улавливающими изменение среды обитания [2,3,4,5,6,7]
Центральной проблемой экологии остается познание взаимодействий
организмов и их группировок со средой обитания. В рамках эволюционной
проблематики изучение механизмов формирования и функционирования
пространственной структуры необходимо, прежде всего, потому, что именно
пространственная дифференциация населения - может являться первым этапом в
становлении популяции как единицы эволюционного процесса. Она служит основой,
на которой возникает дифференциация населения по биологическим свойствам и
определяет генетическую неоднородность популяции [2,3,4,5].
Изучение пространственной структуры популяций позволяет решать сложные
проблемы теории и практики, существенно повысить уровень знаний о популяционных
процессах и связать изучение популяций с решением практических задач
эпидемиологии, лесоведения, рационального природопользования. Исследование
распределения животных по территории необходимо для экологического мониторинга,
повышения эффективности борьбы с вредителями, улучшения мер охраны редких
видов, для более рационального использования запасов промысловых видов.
Цель исследования. Цель работы состоит в комплексном анализе
пространственно-демографической структуры лесной мыши (Apodemus sylvaticus Linnaeus, 1758), обитающей в условиях
ландшафта, подверженного антропогенным изменениям.
Задачи исследования.
.Исследовать сезонные изменения пространственно-демографической структуры
населения лесной мыши двух биотопов.
.Выявить особенности демографической структуры и пространственного
распределения населения лесной мыши в условиях антропогенного воздействия.
1. Анализ литературных данных по пространственно - демографической
структуре поплуляций мелких млекопитающих
1.1 Структура и понятие о популяции
Жизнь невозможна не только в виде изолированных клеток и тканей, но и в
форме отдельных особей. Невозможна жизнь и непосредственно в форме биоценозов,
т.к. биоценотические связи - это связи между видовыми популяциями, а не связи
между отдельными особями.
Биологический вид - группа родственных организмов, занимающих общий
ареал, имеющая общее происхождение и обладающая общими морфологическими и
физиологическими свойствами, свободно скрещивающихся между собой особей с
образованием плодовитого потомства и обособленная от других нескрещевамостью в
природных условиях.
Любой вид животных, растений, микроорганизмов утверждает себя во внешней
среде, приспосабливается к ее меняющимся условиям не как простая сумма особей,
а в форме своеобразных группировок организмов, представляющих единое
функциональное целое. Вид - это сложная система, состоящая из группировок
организмов, обладающих характерными особенностями морфологического строения,
физиологии и поведения. Группа организмов одного вида, занимающая длительное
время определенную территорию, имеющая общий генофонд, сходную морфологию,
единый жизненный цикл, представляет собой популяцию. С экологической позиции
четкого общепринятого определения популяция не существует. Но наибольшего
признания заслуживает определение, предложенное С.С. Шварцем: под популяцией
целесообразно понимать элементарную совокупность особей, которая обладает всеми
необходимыми условиями для поддержания своей численности на характерном для
данного вида уровне в течение длительного периода и обладает известными общими
свойствами, определяющими единство жизнедеятельности слагающих популяцию
особей.
Популяция как биологическая система характеризуется новыми специфическими
свойствами, которыми не обладают отдельные организмы данного вида. Только на
популяционном уровне проявляются такие свойства, как численность и плотность
населения, половая и возрастная структура, уровень рождаемости и смертности и
другие. Особи, составляющие популяцию, с одной стороны, являются частями
системы более высокого порядка, сохраняют в то же время ярко выраженную
самостоятельность. Отдельный организм относительно недолговечен, популяция же,
как биологическая система, при том, что условия среды обитания не выходят за
пределы толерантности, практически бессмертна.
Вместе с тем популяция обладает и определенными чертами сходства с
организмом как биологической системой: структурированность, интегративность
составных частей, способность к саморегуляции и одно из главных свойств
биологических систем - способность к адаптациям.
Вид занимает тот или иной пространственно структурированный ареал, а
каждая видовая популяция обитает на его определенной части, которая
характеризуется отличающимися условиями. В свою очередь популяция
пространственно и функционально структурирована. Пространственная
структурированность определяется главным образом неоднородностью условий
обитания, а функциональная разнокачественность особей, входящих в состав
популяции, - требованиями условий ее длительного сохранения в данных условиях.
Пространственно-функциональная структурированность формирует систему
внутрипопуляционных взаимоотношений как между отдельными особями, так и их
группировками, определяя тем самым возможность наиболее эффективного
использования ресурсов среды и устойчивость популяции как системы на фоне
изменяющихся условий среды.
Каждый вид распространен в определенном ареале на той или иной
территории, различные части которой характеризуются отличающимися размерами
условиями существования. Н.П. Наумов предложил классификацию популяций в
зависимости от размеров занимаемой ими территории:
· элементарная (локальная) популяция - совокупность особей одного вида,
занимающих небольшой участок однородной площади. Количество таких популяций
пропорционально разнообразию условий среды;
· экологическая популяция - совокупность элементарных
популяций, т.е. внутривидовые группировки, приуроченные к конкретным типам
местообитаний;
Географическая популяция - совокупность экологических популяций,
заселяющих значительную территорию с географически однородными условиями, в
пределах которой наблюдается единый ритм жизненных процессов и другие
функциональные особенности, создающие морфологический тип, отличающий данную
популяцию от соседних, находящихся в иных географических условиях. Структура
популяций Популяция как биологическая система обладает структурой и выполняет
определенные функции. Структура популяции характеризуется составляющими ее
особями и их распределением в пространстве. Функции популяции - рост, развитие,
способность поддерживать существование неопределенно длительное время в
постоянно меняющихся условиях. Групповое распределение (агрегации). Для
внутренней структуры большинства популяций в различные фазы популяционного
цикла характерно образование групп разных размеров. Такие группы представляют
собой результат агрегирования особей, происходящего по следующим причинам:
·
вследствие
локальных различий в местообитаниях;
·
в связи с
процессами размножения;
·
в результате
социального привлечения (у высших животных).
Агрегация может усиливать конкуренцию между особями за ресурсы (пищу или
пространство), но это часто более чем уравновешивается повышением
жизнеспособности группы, поскольку группа обладает большими возможностями для
своей защиты, использования ресурсов или изменения микроклимата.
В целом для растений и, вероятно, для некоторых низших животных применимо
экологическое правило, согласно которому тенденция к агрегации находится в
обратной зависимости от подвижности стадий расселения (семян, спор и т.д.). В
насаждениях кедра, хурмы и других видов растений, с тяжелыми семенами, у
которых семена не имеют приспособлений, способствующих их переносу на большие
расстояния, молодые побеги чаще всего сгруппированы около родительских растений
или в местах, где птицы или животные запасают семена. И, напротив, у видов,
имеющих легкие семена и различные приспособления для их переноса (многие виды
злаков, семена сосны, семена липы) они разносятся ветром на дальние расстояния,
и потому могут заселять территории сравнительно равномерно.
Одна из причин агрегирования у животных - социальное поведение.
Социальная агрегация характеризуется определенной организацией, в частности,
социальной иерархией и специализацией особей. Т.е. особи в таких агрегациях
связаны между собой отношениями лидерства, доминирования и кооперации.
Удивительны поселения общественных насекомых, т.к. составляющие их особи крайне
специализированы. К числу наиболее высокоорганизованных общественных насекомых
относятся термиты, муравьи, пчелы. В сообществах этих животных существует как
минимум три касты: производители (например, матка у пчел), рабочие и солдаты.
Каждая из каст морфологически специализирована для выполнения соответствующих
функций: размножения, заготовки пищи или защиты (пчелы, муравьи и другие виды).
Высокая выживаемость в группе - важный признак, который может быть
результатом агрегации. Группа растений способна лучше противостоять ветру или
эффективнее уменьшать потери воды, чем отдельные особи. У. Олли провел
множество соответствующих экспериментов: например, он обнаружил, что рыбы в группе
могут лучше переносить введенную в воду определенную дозу яда, чем отдельные
особи, т.к. слизь и другие выделения, противодействуя яду, позволяют снизить
эффективность действия яда. Агрегации многих видов птиц и млекопитающих снижают
давление хищников. Другой пример положительного влияния объединения в группу -
повышение выживаемости у пчел. Пчелы в улье (скоплении) выделяют и сохраняют
достаточно тепла для выживания всех особей при температуре, при которой гибнут
изолированные особи. Большинство природных популяций распадается на
микропопуляции - более мелкие группы особей, могущие иметь различные частоты
встречаемости генотипов. Они могут быть временно изолированны (в отдельные
сезоны года), но вследствие миграций и расселения молодняка и других процессов
связаны между собой и имеют единый генофонд. В большинстве случаев между
соседними популяциями осуществляется постоянный обмен особями, а значит и
генетической информацией. Этим определяется их генетическая близость, но при
этом различия по морфологическим и физиологическим особенностям между ними
сохраняются.
Генетическое единство - основное свойство популяции, определяющее все ее
особенности и пути ее развития.
Фенотипические характеристики. Фенотип - это совокупность различных
морфологических и физиологических особенностей растений и животных, которая
формируется под влиянием конкретных условий существования.
К фенотипическим характеристикам популяции в первую очередь относятся
скорость роста и развития, общие размеры тела (вес), плодовитость, пропорции
тела, окраска и т.д. Даже в близко расположенных популяциях, но обитающих в
неодинаковых условиях, могут достаточно ярко проявляться морфологические и
физиологические различия.
Если популяции обитают в среде с постоянным однонаправленным действием
каких - либо факторов среды, живые организмы, составляющие ее, приобретают
специфические фенотипические и генетические особенности. Именно эти особенности
и определяют в последующем направления естественного отбора и во многом -
эволюцию вида.
Особый тип агрегации был Гамильтоном назван "образованием безопасных
поселений ". В этом случае обширные, социально организованные группы
животных обосновываются в благоприятном центрально расположенном участке,
откуда они регулярно расходятся для удовлетворения потребностей и куда
возвращаются вновь. Это наблюдается, например, в колониях скворцов или
гнездовых колониях морских птиц. Очевидно, что городская агрегация человека
организована таким же образом. Разработка горючих ископаемых фактически
уничтожила границы зон жизнеобеспечения; и поскольку города не имеют больших
запасов энергии, величина популяции в них ограничивается доступными источниками
топлива и пищи. Вероятно, городская агрегация благоприятна для человека, но
лишь до определенного предела. По мере роста плотности населения все большую
роль лимитирующего фактора начинает играть загрязнение окружающей среды
отходами жизнедеятельности. С точки зрения экологических принципов ошибочно
поддерживать или субсидировать город, который стал слишком большим для своего жизнеобеспечения.
Концепция чистой энергии биологической системы как целого основывается на
распределении энергии между продукцией (размножением) и поддержанием. Отношение
энергии размножения к энергии поддержания варьирует не только в соответствии с
размерами организмов и характером его жизненного цикла, но также и в
соответствии с плотностью популяции и емкостью среды.
Изоляция и территориальность. Факторы, способствующие изоляции и
разделению особей, пар или мелких групп, составляющих популяцию в пространстве,
важны для оптимизации использования ресурсов и улучшения приспособленности
популяций к условиям среды. Обычно изоляция возникает как следствие:
конкуренции между особями за ресурсы; прямого антагонизма, включающего у высших
животных поведенческие реакции, а у растений и микроорганизмов - химические
изолирующие механизмы (антибиотики, и аллелопатические вещества). В обоих
случаях это может привести к равномерному или случайному распределению,
поскольку ближайшие соседи уничтожаются или изгоняются.Многие животные
ограничивают свою основную деятельность определенной областью или
индивидуальным участком, площадь которого может варьировать от нескольких см2,
до многих квадратных километров (таковы размеры участка пумы). Владельцы
территорий заявляют о своих правах на участок с помощь песни или
демонстративного поведения, и потенциальный захватчик, в общем, избегает
вторжения в занятое владение. Многие птицы, а также рыбы и пресмыкающиеся
обладают заметной, бросающейся в глаза окраской головы, тела и т.д., которую
они демонстрируют для отпугивания незваного гостя. У большинства перелетных
птиц самцы прилетают в места гнездования раньше самок и проводят время,
утверждаясь на своей территории и заявляя об этом громким пением. Особи,
занимающие более выгодные территории, успешнее размножаются, что особенно важно
в суровых условиях (при неблагоприятной погоде или недостатке пищи). Полагают,
что территориальное поведение предотвращает распространение болезней вследствие
пространственного разобщения особей; кроме того, оно способствует разделению и
сохранению ресурсов, и облегчает встречу особей при размножении. Для популяции
каждого вида всегда имеются верхние и нижние пределы плотности, за границы
которых она выходить не может. При благоприятном сочетании всех условий
плотность популяции удерживается на каком-то оптимальном уровне, незначительно
отклоняясь от него в ту или другую сторону. Такие колебания численности носят
регулярный характер и четко отражают реакцию популяции на конкретные условия
среды. Падение плотности ниже оптимальной приводит к ослаблению защитных
реакций популяции, снижает ее плодовитость, могут привести к вымиранию
конкретной популяции и вызывать ряд других отрицательных явлений. Повышение
плотности сверх оптимальной также неблагоприятно сказывается на популяции,
поскольку при этом иссякает кормовая база, сокращается жизненное пространство,
появляются вспышки различных заболеваний и т. д.
Различают непериодические, редко наблюдаемые и периодические, постоянные,
колебания численности естественных популяций. К непериодическим колебаниям
численности, а соответственно и плотности популяции можно отнести вспышки
массового размножения среди насекомых (непарный шелкопряд в южной части России
1879 г.), грызунов и т. д. Резкий подъем численности наблюдается у популяций,
оказавшихся в новом местообитании. Классическими примерами этому может служить
массовое размножение кроликов в Австралии, колорадского жука в Европе.
Периодические изменения численности популяций совершаются обычно в
течение нескольких лет или одного сезона. Циклические изменения численности с
подъемами в среднем через четыре года зарегистрированы у таких животных как
песец, полярная сова, у леммингов. Как правило, циклические изменения
численности полярной совы коррелируют с периодическими изменениями численности
леммингов - основной пищи сов. Численность сов, возрастающая по мере увеличения
количества пищи, затем также быстро снижается по мере ее уменьшения. При этом
численность популяция сов резко уменьшается в результате миграции. Эти колебания
численности так регулярны, что орнитологи США могут точно предсказать нашествие
полярных сов каждые три или четыре года. Поскольку эти птицы хорошо заметны и
появляются повсюду вокруг городов, они привлекают к себе большое внимание.
Хорошо известные примеры циклических изменений численности у жуков-короедов
(Питермен, 1978). Благодаря деятельности жуков, которые изреживают загущенный
древостой, малопригодный для лесозаготовок и обитания в нем животных, создаются
более ценные для человека леса. Деятельность этих жуков можно рассматривать как
одно из средств ведения хозяйства, а не как вредителей леса. В изолированных
лесах, по-видимому, следовало бы допускать вспышки численности этих жуков.
Если агрегация может усилить конкуренцию за жизненно необходимые ресурсы,
то разобщение особей в популяции может уменьшить ее. В природе часто
встречается организация обоих типов, в популяциях некоторых видов одна форма
поведения сменяется другой. Дрозды, например, занимают изолированные территории
в период размножения и собираются в стаи к зиме, получая, таким образом,
преимущества от обоих типов распределения. Кроме того, особи разного возраста и
пола могут вести себя по-разному в один и тот же период (например, взрослые
живут изолированно, а молодые образуют скопления). Она никогда не бывает
постоянной и определяется соотношением рождаемости и смертности, а также
процессов эмиграции и иммиграции. В процессе размножения происходит рост
популяции, смертность же приводит к сокращению численности. Экологическая
плотность - число особей (или биомасса) на единицу заселенного пространства
(т.е. доступной площади или объема, которые реально могут быть заняты
популяцией).
Интересен вопрос о том, насколько человеку свойственно врожденное
территориальное поведение, и о том, в каких пределах он может научиться
планировать землепользование так, чтобы избежать перенаселения. В поведении
человека обнаруживаются определенные черты территориальности, такие как
императив личной собственности, законы и обычаи, предписывающие ему защищать
свой дом, если нужно, то и с оружием в руках. Роберт Ардри в своей книге
"Территориальный императив " утверждает, что человеку от рождения
присущи черты территориального поведения, и поэтому он неизбежно будет
противостоять чрезмерной скученности, избегнув тем самым гибели от
перенаселения. К сожалению, человек слишком хорошо адаптируется к чрезмерной
скученности, как это можно видеть на примере Гонконга, одной из самых
густонаселенных точек Земли.
Характеристики популяционной группы
Популяция характеризуется рядом признаков, причем единственным носителем
этих признаков является группа, а не особи. Вот некоторые из таких свойств
популяции: численность, рождаемость, смертность, возрастная структура,
распределение в пространстве. Численность популяции непостоянна. В природе не
отмечено ни одной популяции, которая бы оставалась стабильной даже на
протяжении короткого отрезка времени. Основными причинами ее изменения являются
изменения условий существования. Это связано с воздействием абиотических
факторов на интенсивность роста и размножения, на смертность, на скорость
развития. Например, известно, что динамика популяций белки, зайца, многих видов
птиц зависит в большинстве случаев от урожая кормов. В настоящее время
изменения численности популяции нередко зависят от деятельности человека.
Немалую роль в этом процессе играет уничтожение хищниками жертв, конкурентные
взаимоотношения, влияние паразитов на жизнедеятельность хозяев и т. д.
Популяции обладают также генетическими и фенотипическими характеристиками, непосредственно
связанными с условиями обитания. Каждая особь в популяции отличается от другой,
но все они вместе представляют сложный биологический комплекс особей,
распределенных по своим индивидуальным качествам по кривой, близкой к
нормальному распределению.
Основные характеристики популяции можно разделить на две самостоятельные
группировки:
· характеристики, определяющие предпосылки для самостоятельного
существования и развития популяции;
· характеристики, определяющие биологическую специфичность
(морфологическую и физиологическую) слагающих популяцию особей.
Генетические характеристики. Генотип - совокупность всех генов данного
организма. Еще в начале прошлого века популяцию рассматривали как совокупность
генетически неоднородных особей, противопоставляя ее генетически чистым линиям.
В дальнейшем этот подход приобрел генетико-эволюционный смысл и в современном
представлении популяция рассматривается как элементарная единица эволюционного
процесса. С этих позиций главный критерий популяции - способность к свободному
обмену генетической информацией. Размножение. Динамика численности и плотности
популяции находятся в тесной зависимости от размножения и смертности, а так же
от способности слагающих их особей совершать миграции.
Рождаемость (плодовитость) - характеризует процесс появления на свет
новых особей любого организма независимо от того, рождаются ли они, вылупляются
из яиц, прорастают из семян или появляются в результате деления, что приводит к
увеличению численности популяции. Этот термин используется также и при анализе
популяций человека (в демографии). Обычно рождаемость выражают либо как
скорость, определяемую путем деления общего числа вновь появившихся особей на
время (общая рождаемость), либо как число вновь появившихся особей в единицу
времени на 1 особь в популяции (удельная рождаемость).
В живых организмах заложена огромная потенциальная возможность к
размножению. Подсчитано, что бактерии делятся каждые 20 минут. При таком темпе
одна клетка за 36 часов дает потомство, которое может покрыть сплошным слоем
всю планету. Один одуванчик менее чем за 10 лет способен заселить своими
потомками земной шар, если все семена прорастут. В действительности же такая
громадная плодовитость организмов никогда не реализуется. В связи с этим
различают максимальную или потенциальную рождаемость и экологическую или
реализованную рождаемость.
Потенциальная рождаемость - это теоретически возможная максимальная
скорость образования новых особей в идеальных условиях, когда отсутствует
действие лимитирующих экологических факторов, а размножение сдерживается только
физиологическими возможностями организма.
Экологическая или реализованная рождаемость означает увеличение
численности популяции при фактических условиях среды. Эта величина не постоянна
и изменяется в зависимости от размерного и возрастного состава популяции и
физических условий среды.
Например, в колонии чайки отложено 510 яиц (потенциальная рождаемость) из
них вылупилось 265 птенцов, реализованная плодовитость составила 52% от
потенциальной. Мучные хрущики отложили 1200 яиц - вылупилось 713 особей, что
составило всего 60%. У капустной белянки в результате гибели яиц, гусениц и
куколок по разным причинам (болезни, паразиты, хищники и др.) на свет
появляется лишь 0,32 % взрослых бабочек от числа отложенных яиц.
Рождаемость каждого вида определяется как приспособление, обеспечивающее
пополнение популяций, а значит, ее жизнестойкость. Поэтому естественно, что у
менее приспособленных к неблагоприятным условиям видов высокая смертность в
молодом возрасте компенсируется значительной плодовитостью. В зависимости от
взаимоотношений между родителями и потомками существует две противоположные
стратегии размножения. Виды животных и растений, у которых забота о потомстве
выражена явно - смертность их потомства на ранних стадиях развития низка, а
сами они имеют низкую плодовитость. Потомство видов, у которых отсутствует
забота о детенышах, имеют высокую смертность, а их родители - высокий
репродуктивный потенциал (как компенсацию за высокую смертность потомков). Для
видов, которые не охраняют яйца и не заботятся о потомстве, характерна высокая
потенциальная и низкая экологическая рождаемость. В благоприятных условиях
рождаемость обычно низкая а в неблагоприятных - высокая. Этим объясняется более
высокая рождаемость животных в зонах умеренного климата по сравнению с
тропическими видами.
Смертность. Численность и плотность популяции зависят также от ее
смертности. Смертность популяции - это количество особей, погибших за
определенный период. Она, как и рождаемость, изменяется в зависимости от
условий среды, возраста и состояния популяции и выражается в процентах к
начальной или чаще к средней ее величине. Выделяют: экологическую смертность -
гибель особей, наблюдаемую в конкретных условиях среды (эта величина
непостоянна и изменяется в зависимости от условий среды и состояния самой
популяции); теоретическую минимальную смертность - величина, постоянная для
популяции; она представляет собой гибель особей в идеальных условиях, при
которых популяция не подвергается действию лимитирующих факторов. Даже в самых
лучших условиях особи будут умирать от старости. Этот возраст определяется
физиологической продолжительностью жизни, которая часто превышает среднюю
экологическую продолжительность жизни. На все популяции оказывают воздействие
как зависящие, так и не зависящие от плотности факторы, однако не у всех видов
демографические реакции одинаковы. Численность некоторых видов довольно
постоянна. Плотность других видов резко изменяется под действием внешних
факторов, причем колебания численности нерегулярны. Наконец, плотность
некоторых видов (полярная сова, лемминг, заяц-беляк) подвержена годовым
циклическим колебаниям. Чем объясняются эти различия между видами? Вероятно,
существуют различные способы, при помощи которых численность того или иного
вида приходит в соответствие с условиями среды, и эти различные способы есть
один из видов адаптаций на популяционном уровне.
Таким образом любой фактор, лимитирующий или благоприятный (отрицательный
или положительный), может быть: независимым от плотности, если его влияние не
зависит от величины популяции, или зависимым от плотности, если его влияние на
популяцию есть функция плотности. К первым относятся факторы, действующие на
популяцию постоянно. Это абиотические и, прежде всего климатические факторы. Хорошо
известно влияние температуры, освещенности, влажности на продолжительность
жизни, рождаемость и смертность. Причем на пойкилотермных животных
климатические факторы оказывают непосредственное и более сильное воздействие,
чем на гомойотермных. Действие климатических факторов не всегда проявляется
сразу, немедленно. Например, в тайге благоприятные погодные условия
способствуют высокому урожаю семян через год, а подъем численности популяции
животных при обильном корме наблюдается лишь через два года. При этом погодные
условия действуют независимо от плотности популяций. Независимо от плотности
популяции проявляют себя и другие факторы. К примеру, количество белой
куропатки и ряда видов млекопитающих резко сокращается, если они не находят
подходящих местообитаний даже при благоприятном сочетании остальных факторов.
Зависимые от плотности факторы, как правило, воздействуют на скорость
роста популяции. При определенных условиях скорость роста уменьшается. Это
явление широко распространено и позволяет объяснить относительную устойчивость
популяций животных. Как показали лабораторные эксперименты с крысами, когда
плотность популяции достигает определенной величины, плодовитость животных
сильно снижается, даже если нет недостатка в пище. Происходят различные гормональные
сдвиги, которые влияют на половое поведение; все чаще встречается неспособность
к спариванию, бесплодие, выкидыши, поедание детенышей родителями. Родительская
забота о потомстве ослабевает, и детеныши покидают гнездо в очень раннем
возрасте, что снижает вероятность их выживания. Усиливается агрессивность
животных. Напротив, падение плотности популяции ниже оптимального уровня,
например, при усиленном истреблении крыс, вызывает повышение плодовитости и
симулирует их более раннее половое созревание. Подобные изменения характерны
для ряда млекопитающих и могут происходить также в природных условиях вне
лаборатории, например в популяциях полевок.
Регуляция численности популяции. Вся сложная совокупность факторов среды
постоянно изменяется и создает определенные условия для популяции. Численность
популяции зависит от многих факторов. Существенное влияние могут оказывать как
изменения климатических условий (например, температуры, количества осадкой
осадков и др.) так и пищевые ресурсы, враги и т.д. Размеры популяции могут
возрастать в результате иммиграции из соседних популяций или за счет
размножения особей. Численность популяции может уменьшаться в результате
эмиграции или смертности.
1.2 Семейство
Мышиные (Muridae)
Изучение динамики численности млекопитающих и управление ею остается
одной из важнейших проблем популяционной экологии. Вопросы познания механизмов
взаимодействия животных и их группировок со средой обитания, роли
пространственных взаимоотношений в формировании структуры популяций млекопитающих
актуальны как для решения практических задач эпидемиологии, лесоведения,
рационального природопользования, так и для развития теоретических положений
биоценологии и эволюционной теории.
Познание закономерностей, определяющих функционирование группировок
животных на уровне популяции, их биологическую целостность и единство, их
автономность во времени и пространстве, открывает широкие перспективы для
решения ряда теоретических и практических вопросов. Многие эволюционные
процессы, и особенно те, которые относятся к области микроэволюции, могут быть
поняты только на основе четких представлений о начальных этапах дифференциации
видового населения, о путях этой дифференциации и о причинах, их
обусловливающих. С другой стороны, такие в значительной мере практические
проблемы, как регулирование численности животных, регламентация промысла,
эпидразведка и оздоровление природных очагов инфекций, повышение биологической
продуктивности угодий и, наконец, охрана природы, требуют для своего разрешения
глубоких и всесторонних знаний обо всех аспектах популяционной структуры [3].
Познанию пространственной структуры популяций принадлежит особая роль.
Именно пространственная дифференциация видового населения является первым
этапом в становлении каждой популяции как основной единицы эволюционного
процесса, той базой, на которой возникает дифференциация по другим
биологическим показателям, генетическая неоднородность популяции в целом [6].
В настоящее время наиболее широко обсуждаются вопросы пространственного
распределения по территории не отдельных особей, а их группировок разного
уровня. В.Н.Беклемишев (1960) понимал под пространственной структурой популяции
классификацию ее по самым различным признакам - автономности, протяженности,
устойчивости [8].
Н.П.Наумов (1971) идентифицирует пространственную структуру с
биохорологической, а Флинт как типичную и закономерную для данной популяции
пространственно-временную динамику мерусов, слагающих ее [3,9].
В настоящее время в науке сложилось достаточно определенное представление
о том, что биологический вид не является беспорядочной совокупностью особей, а
представляет собой функциональную систему, состоящую из естественных
территориальных группировок, построенную по иерархическому принципу и
отражающую уровни интеграции. Такая система получила название биохорологической
структуры вида. Реальность территориальных группировок и комплексов группировок
животных разного масштаба не вызывает сомнения и, если речь идет о возможно
более дифференцированном подходе к анализу биологических особенностей вида,
выделение таких группировок оправдано, а число их не ограничено. Однако далеко
не каждая территориальная группировка (совокупность особей), несмотря на ее
реальность и объективность, составляет таксон биохорологической структуры вида
[10]. Для обозначения территориальных группировок, слагающих популяцию, т.е.
для таксонов биохорологической структуры В.Е. Флинт (1977) предложил новый и
ранее не использовавшийся термин мерус. Здесь мерус - совокупность особей,
обладающих фенотипическим единством и представляющих собой таксон
биохорологической структуры вида. Фенотипическое единство животных основано на
тонкой адаптированности к конкретным и неустойчивым условиям ограниченного
участка земной поверхности на коротком отрезке времени. Цикличность изменений
биологических параметров в каждом отдельном мерусе носит не многолетний, а
сезонный характер. В целом пространственное выражение меруса представляется как
элементарное поселение. Территориальные группировки животных, образующих
элементарное поселение, выделяются по общности всего комплекса биологических
параметров (биотопическая приуроченность, сезонная динамика численности,
характеристика питания, суточная активность, типы убежищ, морфофизиологические
особенности) и представляют собой достаточно четко отграниченное единство. Шилов
И.А. (1977) описывает отдельные группировки животных внутри популяции, в
которых особи находятся в устойчивых информационных и функциональных контактах.
Исследование топографической структуры популяций млекопитающих показало,
что пространственная организация популяций определяется сложными,
слабоизученными взаимоотношениями между отдельными особями и их группировками.
И.А.Шилов (1977) подчеркивал, что характер пространственной структуры,
различной у отдельных видов, во всех случаях определяет возможность самого
"выгодного" для популяции в целом использования ресурсов среды, а
также наиболее эффективного осуществления внутрипопуляционных взаимоотношений
особей и их групп. Такие группы в литературе называются демами или парцеллами
[9,11,12]. В зависимости от расположения мерусов выделяют различные типы
пространственного распределения животных. Так, В.Е. Флинт (1977) описал четыре
типа пространственной структуры популяций: эквальный (диффузный), инсулярный
(мозаичный), циклический (переложный), пульсирующий.
Другой подход к описанию пространственной структуры населения реализован
на признании ее как совокупности агрегаций животных. В отличие от мерусов,
агрегации могут и не иметь признаков фенотипического единства и являются
совокупностью особей, проживающих вместе. Исходя из этого, пространственная
структура населения нами рассматривается как закономерное чередование пятен,
характеризующихся определенным усилием животных. Каждое такое пятно - это
площадь территории, занятая частью популяции, причем независимо от уровня
численности все такие части имеют один и тот же иерархический
внутрипопуляционный ранг [13].
В соответствии с расположением групп животных по территории и степенью их
дифференциации рассматриваются различные типы пространственного распределения
организмов. Наиболее часто встречается разделение на: агрегированное, случайное
и равномерное распределения животных и их группировок в пространстве [14,15].
Степень агрегированности можно оценить путем статистического анализа
распределения животных по территории [16,17,18].
Равномерный тип распределения в идеале характеризуется равным удалением
каждой особи от всех соседних; величина расстояния между особями соответствует
порогу, за которым начинается взаимное угнетение. Таким образом, теоретически
этот тип распределения в наибольшей степени соответствует задаче полного
использования ресурсов при минимальной степени конкуренции. В действительности
же равномерное распределение особей встречается в природе достаточно редко.
Невозможность реализации этого "идеального" типа распределения
определяется прежде всего неоднородностью среды обитания; нарушают
равномерность распределения и свойственные многим видам формы взаимосвязей.
Диффузный (случайный) тип распределения особей, встречается в природе
значительно чаще, при нем особи распределены в пространстве неравномерно,
случайно. В этом случае расстояния между особями неодинаковы, что определяется,
с одной стороны, вероятностными процессами, а с другой - определенной степенью
неоднородности среды. Диффузное распределение характерно для животных, у
которых социальная связность в пространстве выражена относительно слабо.
Агрегированный (мозаичный) тип распределения выражается в образовании
группировок особей, между которыми остаются достаточно большие незаселенные территории.
Биологически это связано либо с резкой неоднородностью среды, либо с выраженной
социальной структурой, действующей на основе активного сближения животных. Во
всех случаях тип пространственного распределения организмов в популяциях
определяется, с одной стороны, гетерогенностью среды, а с другой - соотношением
процессов пространственной дифференциации, снижающей степень конкурентности, и
функциональной интеграции особей, обеспечивающей целостность популяции как
функциональной системы. Особенно четко это выявляется при изучении популяций
животных [15].
Пространственная структура популяции теснейшим образом связана с главными
особенностями ландшафта и его подразделений (морфологических частей), причем
все компоненты ландшафта (рельеф, климат, почвы, растительность и т.д.) в той
или иной мере налагают свой отпечаток на специфику формирования определенного
типа пространственной структуры популяции. Также очевидно, что при формировании
определенного типа использования территории огромную роль играет биологическая
характеристика самого животного, его потребности и возможности, его
биологические параметры (тип питания, физиологические особенности, характер
размножения и динамики численности, строение убежищ и т.д.). Именно как
взаимодействующая этих двух групп явлений и может рассматриваться в каждом
конкретном случае основная тенденция к возникновению совершенно определенного
типа пространственной структуры популяции [3].
Распределение животных по территории неравномерно и во многом
определяется биотопическими особенностями [1,19,20,21,22,23,24,25,26]. При этом
неоднородность местообитаний входит в число ведущих факторов, определяющих
пространственную структуру популяции, в зависимости от ее выраженности можно
наблюдать весь спектр распределений животных на территории (от равномерного до
агрегированного).
Конкретно в экологии мелких млекопитающих необходимость учета фактора
неравномерности прямо следует и из показанной многими авторами приуроченности
поселений мелких млекопитающих разных видов к определенным элементам
экологической мозаики с оптимальными и зачастую видоспецифичными условиями
обитания [3,19,22]. В связи с этим выделяются зоны, существенно различающиеся
по степени их использования животными [9,25,26,27,28,29,30,31]. К таким
территориям относятся: резервации, временные поселения и транзитные участки.
Резервации или донорные участки характеризуются наиболее благоприятными для
проживания и размножения условиями среды, животные обитают на них постоянно.
Временные поселения позволяют размножаться и существовать в них только на
протяжении определенных сезонов. Транзитные участки используются для
проживания, но практически непригодны для размножения.
Совмещая представления о структуре пространства обитания и характере
перемещений мышевидных грызунов, можно сформулировать следующее общее
положение. В годы низкой численности животные концентрируются в резервациях и
вне их встречаются эпизодически, во время разведочных выходов. При повышении
численности сначала половозрелые особи низкого ранга, а затем неполовозрелые
животные начинают выселяться во временные поселения, выбирая для образования
гнездового участка наилучшую из свободных в данный момент территорий.
Постепенно все пригодные для размножения и обитания участки заполняются и в
действие вступают плотностно-зависимые механизмы регуляции численности. Впервые
подобное представление было сформулировано Дж. Хестбеком (1982) и названо
гипотезой социальных барьеров [32]. Согласно этой гипотезе, численность группы,
занимающей один из популяционных рефугиумов, в процессе роста численности (от
одного до нескольких лет) регулируется эмиграцией животных в сопредельные
биотопы, где формируются временные поселения с исходно невысокой плотностью и,
следовательно, с высокой проницаемостью для новых мигрантов. С течением времени
проницаемость связанных с резервацией временных поселений снижается, что
обусловлено ростом в них плотности и формируется "социальный барьер",
который начинает ограничивать расселение. В результате плотность в центральной
группе и во временных поселениях превышает емкость среды, что приводит к
включению плотностно-зависимых механизмов на фоне возможного истощения пищевых
ресурсов. Менее строго такой эффект описал В.Лидикер (1975) [33].
Нередко пространственная структура рассматривается как система
индивидуальных участков [34,35,36,37,38,39,40,41,42]. С количеством свободных
участков часто связывают репродуктивную активность животных. В размножении, как
правило, принимают участие особи, обладающие собственной территорией. Зверьки,
которые не имеют индивидуального участка, составляют "резерв"
популяции и приступают к размножению только тогда, когда смогут завладеть
гнездовым участком [42,43,44,45].
В настоящее время накопилось множество сведений о видовых особенностях,
сезонных и географических изменениях участков обитания. При этом отмечается
высокая степень вариабельности их размеров и конфигураций. Причинами таких
изменений могут быть как факторы внешней среды (сезон года, кормность и
защищенность биотопа), так и внутрипопуляционные. Современными исследованиями
выявлены механизмы образования индивидуальных участков, половые и возрастные
особенности размеров и степени перекрывания участков обитания мелких
млекопитающих в разные сезоны [46,47,48,49,50]. Так, например, сравнение данных
разных исследователей показали, что размеры участков обитания мышевидных
грызунов очень изменчивы и колеблются от нескольких сот квадратных метров до 7
га и более.
Анализируя работы по пространственному распределению мелких
млекопитающих, мы старались подчеркнуть тесную связь пространственной структуры
с целым комплексом факторов, вытекающих в первую очередь из свойств биотопа и
биологических особенностей вида.
Познание закономерностей становления, существования и развития
пространственной структуры популяций дает ценный материал для анализа самих
популяций. Моделирование как метод изучения процессов, протекающих в
популяциях, невозможно без надежного знания основных закономерностей
пространственной структуры. Перспективы в этом плане почти неограниченны.
Например, огромный опыт медицинских зоологов показал, что в основе явления
природной очаговости большинства инфекций лежит феномен элементарной
очаговости. Биологическая сущность его заключается в том, что в силу ряда
обстоятельств возбудитель сохраняется в течение неопределенного долгого времени
(теоретически беспредельно) не на всей территории очага, а лишь в определенных
его точках, в стратегически важных узлах [51]. В.Е. Флинту (1970) удалось
показать, что эти узлы совпадают с совершенно конкретными элементами пространственной
структуры популяций; на основе этого положения разработаны методы оценки
эпизоотологической значимости ландшафта [10]. Вся тактика работы по ликвидации
природных очагов инфекций строится на основе этих принципов. Точно так же
обстоит дело с регулированием численности вредных животных, с
регламентированием промысла, с составлением прогноза численности и т.д.
Семейство тушканчиковых объединяет
небольшую группу грызунов, населяющих степи, полупустыни и пустыни юга
Палеарктики и приспособившихся к специфическим и суровым условиям обитания в
этих ландшафтах. Тушканчики - грызуны от средних до очень мелких размеров:
длина тела от 5 до 26 см. В общем облике тушканчиков характерны очень
крупная голова с сильно притуплённой мордочкой, длинными закругленными ушами,
громадными круглыми глазами и длинными вибриссами, короткое кургузое туловище,
крошечные передние лапки, мощные прыгательные задние конечности и длинный,
часто снабженный кисточкой хвост. Большие уши, глаза и длинные вибриссы
свидетельствуют о высоком развитии слуха, сумеречного зрения и осязания,
которые необходимы тушканчикам при поиске пищи и защите от врагов в ночных
условиях. Маленькие передние лапки служат для схватывания и удержания пищи, а
также для рытья нор, в чем тушканчики достигают большого мастерства. Задние
конечности - прыгательные, и в связи с этой функцией они сильно видоизменены:
ступня удлиненная и 3 средние плюсневые косточки срастаются в одну общую кость,
называемую цевкой. Боковые пальцы задних ног развиты слабее средних или полностью
отсутствуют. Хвост, обычно превышающий по длине туловище, играет важную роль
при движении: он служит для поддержания равновесия тела при прыжках, особенно
при резких поворотах на быстром скаку. Черно-белая кисточка, имеющаяся на конце
хвоста у многих видов, называется "знамя" и, очевидно, служит важным
сигнальным средством при внутривидовом общении.
Коренных зубов (вместе с предкоренными) у
тушканчиковых 3(4)/3 Резцы тушканчиков, помимо разгрызания пищи, служат
основным орудием разрыхления почвы при рытье нор, в то время как конечности
употребляются главным образом уже для отгребания разрыхленного грунта.
Большинство тушканчиков приурочено в своем
распространении к полупустынным и пустынным ландшафтам низменностей, лишь
отдельные виды населяют степную зону, а некоторые проникают в горы выше 2000 м. У разных видов выработались
приспособления к обитанию на рыхлых или на плотных грунтах, и поэтому
тушканчиков можно встретить и в песчаных, и в глинистых, и в щебнистых
полупустынях и пустынях.
Тушканчики - типично ночные животные,
появляющиеся на поверхности только с наступлением темноты. Перед рассветом они
прячутся в норы, которые строят сами. Нора тушканчика устроена так: главный
ход, идущий наклонно под поверхностью, с одним или несколькими слепыми запасными
ходами, подходящими почти к поверхности. Главный ход на день закупоривается
земляной пробкой, которая называется копеечкой. По этой копеечке, ранним утром
еще не просохшей, можно обнаружить нору тушканчика. Если обитаемую нору начать
раскапывать, то зверек вышибает крышу одного из запасных ходов и выскакивает
через него. В дальней части главного хода тушканчик выкапывает отнорок с
округлой жилой камерой, которую выстилает мелко разгрызенными травинками.
Зимний период тушканчики проводят в
глубокой спячке в своих норах. Весной и летом происходит размножение зверьков,
самка рождает 1-8 детенышей, (обычно 2-5). Пищей тушканчикам служат семена
различных растений, луковицы лилейных, которые они выкапывают из грунта,
оставляя характерные копанки. В питание входят также и зеленые части, и корни
различных растений, а у некоторых видов значительную долю в рационе составляют
животные корма (мелкие насекомые и их личинки).
Тушканчиковые играют важную роль в
пустынных биоценозах, они оказывают значительное воздействие на почвенный и
растительный покров, служат пищей для многих пустынных хищников. Во многих
районах по численности тушканчики оказываются фоновыми животными. В настоящее
время известно около 26 видов тушканчиков, объединенных в 11 родов. Область
распространения семейства простирается от Северной и Северо-Восточной Африки,
Юго-Восточной Европы, Малой и Передней Азии через Закавказье, Среднюю Азию,
Казахстан, крайний юг Сибири (Алтай, Тува, Забайкалье) до Северо-Восточного
Китая и Монголии. Почти все роды и большинство видов тушканчиков имеются в
фауне нашей страны, и отечественным ученым принадлежит ведущая роль в изучении
этой своеобразной группы животных. Остается классическим фундаментальное
исследование Б. С. Виноградова (1937), а в недавние годы молодые ученые внесли
много нового в наши познания по систематике, распространению и биологии
тушканчиков.
Длинноухий тушканчик (Euchoreutes naso) - зверек длиной 8-9 см,
с хвостом до 16 см и ступней в половину длины туловища.
Примечательны его удлиненная коническая мордочка, громадные уши,
достигающие задней части спины, и длинные вибриссы, которые, будучи заложены
назад, достают до основания хвоста. Характерны сравнительно маленькие глаза.
Задние конечности пятипалые, причем боковые пальцы укорочены. В строении скелета
много своеобразных и примитивных черт.
Длинноухий тушканчик сверху
рыжевато-серый, с боков и на брюхе - белый. Черно-белая кисточка на конце
хвоста выражена хорошо, но не уплощена, как у других видов, а круглая в
сечении.
Обитает длинноухий тушканчик в песчаных
пустынях Синцзяна и Алашани, где распространен среди редких зарослей саксаулов;
иногда забегает в юрты кочевников.
Пятипалый карликовый тушканчик (Cardiocranius paradoxus) имеет длину тела всего
5-6 см, а хвоста - 7-8 см., непропорционально большую голову с
довольно крупными глазами (однако меньшими, чем у типичных тушканчиков) и с
короткими трубчатыми ушами. Задние конечности - пятипалые, все пальцы хорошо
развиты, на плюсне и ступне есть щетка из краевых жестких волос длиной до 5 мм.
Хвост тушканчика имеет отложения жира в подкожной клетчатке и поэтому
заметно утолщен, главным образом в основной и срединной части. Сверху пятипалый
карликовый тушканчик серовато-охристый с темно-серыми тонами, а на брюхе -
чисто-белый.
До недавнего времени было известно лишь 4
экземпляра этого вида из Тань-шаня, Северной и Центральной Гоби, и лишь в 1961
г. группа исследователей под руководством Д. И. Бермана обнаружила этот
интереснейший вид на территории нашей страны, на юге Тувинской АССР. Держатся
карликовые пятипалые тушканчики в щебнистой пустыне с редкой растительностью из
тас-биюргуна и ковыля, активны ночью, день проводят в норах, причем на твердых
грунтах вынуждены селиться в норах сибирских тушканчиков, делая себе короткие
отнорки. Вероятно, в иных районах этот вид живет и в песчаных пустынях, так как
в неволе зверьки умело рыли собственные норки в песчаном грунте. Питаются
зверьки семенами ковыля, келерии, реже - семенами и другими частями разных
растении, а также, возможно, и насекомыми. В поисках семян зверьки могут лазать
по стеблям трав и ветвям кустарника.
Трехпалые карликовые тушканчики (род Salpingotus)- самые мелкие
представители семейства. Известно 5 видов этих тушканчиков, из которых 3 вида
описаны по единичным экземплярам из Афганистана, Пакистана и Гоби, один вид
описан в 1969 г. по двум экземплярам из Приаральского Кызыл-кума. Обычен и
сравнительно хорошо изучен лишь один вид - жирнохвостый карликовый тушканчик
(S.
crassicauda). Этот зверек имеет
длину тела всего 4-5 см, хвост - 9-10 см. На крупной сравнительно
с туловищем голове - трубкообразные уши длиной около 5 мм. Глаза более
крупные, чем у пятипалого карликового тушканчика. Задние конечности трехпалые,
с хорошо развитой щеткой жестких волос по краям ступни. Хвост покрыт короткими
волосами, на конце хвоста иногда бывает слабовыраженная кисточка. В основной
трети хвост часто заметно утолщен за счет подкожных жировых отложений. Окраска
зверька сверху серовато-желтоватая, снизу - белая.
Жирнохвостый карликовый тушканчик
распространен в Южном Прибалхашье, в Зайсанской котловине, в Монгольском Алтае
и Гоби. По новейшим данным Н. Н. Воронцова и его сотрудников, жирнохвостый
карликовый тушканчик - фоновый вид в закрепленной песчаной ковыльной степи,
реже встречается среди полузакрепленных песков и по окраинам такыров.
Питается зверек семенами различных злаков,
насекомыми, изредка - вегетативными частями растений. При содержании в неволе
зверьки, посаженные в одну клетку, устраивают ожесточенные драки, и не раз
отмечались случаи каннибализма.
Жирнохвостые карликовые тушканчики активны
ночью. Днем они забираются в норы, которые достигают длины 3 м, с
отнорками и гнездовой камерой. Вход в нору зверек забивает песчаной пробкой,
выталкивая песок изнутри головой.
Самки в конце мая - июне приносят по 2-4
детеныша. С наступлением заморозков зверьки залегают в спячку. Все остальные
виды тушканчиков, составляющие типичное ядро семейства, распадаются на две
четкие группы - пятипалых и трехпалых тушканчиков. Пятипалые тушканчики, к
которым относятся земляные зайцы, земляные зайчики и толстохвостые тушканчики,
имеют более длинные уши и пятипалые задние конечности без щетки на ступне. Все
эти тушканчики обитают на твердых грунтах - в глинистых и щебнистых пустынях,
полупустынях и степях, на такырах и солончаках. Трехпалые тушканчики, к которым
принадлежат емуранчик, мохноногий, гребнепалып и песчаные тушканчики,
характеризуются более короткими ушами и трехпалыми задними ногами с плотной
щеткой из жестких щетинистых волос на ступнях. Излюбленные места обитания
трехпалых тушканчиков - песчаные пустыни (лишь емуранчик в противоположность
остальным своим ближайшим родичам предпочитает глинистые и щебнистые пустыни).
Самая обширная группа пятипалых
тушканчиков - это земляные зайцы (род Allactaga), из 9 видов которых 6 встречаются в
нашей стране.
Большой тушканчик, или земляной
заяц (A.
major), - самый крупный из
всех видов семейства. Длина его туловища 19-26 см, хвоста до 30 см; на
конце хвоста - яркое и широкое, расчесанное на две стороны "знамя" с
черным основанием и белой вершиной. Сверху большой тушканчик окрашен в
буровато-серый, бледный песчано-серый цвет, снизу окраска белая.
Большой тушканчик не только самый крупный,
но и самый северный из всех тушканчиков. Он распространен в полупустынях,
степях и даже лесостепях Юго-Восточной Европы, Казахстана и юга Сибири, доходя
на запад до Крыма и Днепра, на север до Оки и Камы, а на восток до верховьев
Оби. Поселяется он и в луговой степи, и в глинистой полупустыне, выбирая места
с плотным грунтом, любит жить на выгонах и вдоль грунтовых дорог. Норы большого
тушканчика имеют 1-2 запасных выхода, забитых земляными пробками или просто не
доведенных на несколько сантиметров до поверхности. Кроме больших и сложных
постоянных нор, у большого тушканчика имеются также более простые и короткие
временные норы, а также зимовальные норы, уходящие в глубь почвы на два с
лишним метра.
Пищу большого тушканчика составляют семена
различных растений, луковицы и клубни, которые он выкапывает из земли, при
случае поедает также насекомых. На сельскохозяйственных землях этот зверек
нередко собирает посеянные семена арбузов и дынь, кормится также зернами
культурных злаков, семенами подсолнуха, горохом.
Весной, вскоре после пробуждения из
спячки, большие тушканчики приступают к размножению. Самки в конце апреля
приносят по 1-4 детеныша, но отмечены и более поздние пометы (возможно,
некоторые особи рождают по два выводка в год).
Тушканчик-прыгун (A. saltator) имеет длину тела 13-17
см, хвоста - 19-22 см. Мех его охристо-буроватый или
желтовато-серый. Черная часть "знамени" на хвосте отделена от
основной части хвоста светлым колечком.
Тушканчик-прыгун обитает в различных
степях, а также в глинистых, щебнистых и отчасти песчаных пустынях, от
Прикаспийской низменности, Казахстана, Северного Узбекистана до Юго-Восточного
Алтая, Тувы, Забайкалья, Монголии и Северного Китая. В горы он поднимается до
высоты 2000 м и более. Питается этот зверек семенами, зелеными частями и
луковицами растений, а также насекомыми и их личинками. По сведениям.В. Е.
Флинта, в выводке может быть 1-7 детенышей (чаще 2-3); продолжительность жизни
в природе, как правило, не превышает трех лет.
Малоазийский тушканчик (A. euphratica) немного мельче
тушканчика-прыгуна, с буроватой спиной, он населяет предгорные и горные степи,
полупустыни и пустыни от Южного Закавказья и Малой Азии до Сирии и Ирака.
Малый тушканчик (A. elater) еще мельче: длина его
тела всего 9-12 см, хвоста - 15-18 см. Он живет в глинистых и
щебнистых полупустынях и пустынях, от Нижнего Поволжья и Прикаспийской
низменности через весь Казахстан и Среднюю Азию до Южного Закавказья, Ирана и
Афганистана. Своеобразный четырехпалый тушканчик (A. tetradactyla) имеет лишь по 4 пальца
на задних конечностях. Нередко его выделяют в отдельный род. Живет этот
тушканчик в Египте и Ливии.
Земляной зайчик (Alactagulus асопtion) по размерам сходен с
малым тушканчиком, но отличается заметно более короткими ушами и менее ярким
"знаменем". От представителей предыдущего рода этот зверек отличается
отсутствием предкоренного зуба в верхней челюсти. Земляной зайчик широко
распространен по всей равнинной Средней Азии, Казахстану и Нижнему Поволжью,
живет на такырах и солончаках, роет норы до 4 ж длиной в твердом и сухом
грунте, с запасным ходом, который закрывает земляной пробкой (копеечкой).
Размножается он дважды в год, при этом в
каждом выводке бывает до 6 детенышей. Земляной зайчик найден и во Внутренней
Монголии.
Толстохвостые тушканчики (род Руgerethmus) - мелкие зверьки, с
более короткими, чем у земляных зайчиков, ушами, без выраженного
"знамени" на конце толстого вальковатого хвоста. В хвосте есть
подкожные жировые отложения. Тушканчик Житкова (P. zhitkovi) имеет длину тела 9-12
см, хвоста - 10-13 см, уши у него длиной 3-3,5 см. На конце
хвоста - небольшая кисточка из более длинных волос, темного цвета, со светлыми
вершинками. Тушканчик Житкова водится в глинистых пустынях и полупустынях
Прибалхашья, местами многочислен, живет в норах длиной до 6 м с
запасными ходами.
Другие толстохвостые тушканчики по
размерам немного меньше: тушканчик Виноградова (P. Vinogradovi) и приаральский
тушканчик (P. platyurus). Среди трехпалых тушканчиков обособленное положение по своей
биологии занимает емуранчик (род Stylodipus). S. telum, в отличие от остальных
видов этой группы, населяет глинистые и щебнистые пустыни и ковыльные степи,
хотя встречается и на песках. Щетка на ступнях емуранчика развита слабо,
"знамя" выражено плохо. Длина тела зверька 9-12 см, хвоста -
13-16 см. Распространен емуранчик от Приднепровья, Среднего и Нижнего
Поволжья через весь Казахстан до Прииртышья. В Монголии обитает близкий вид -
монгольский емуранчик (S. andrewsi).
В род Dipus включается небольшой
мохноногий тушканчик (D. sagitta), который имеет длину тела 10-14 см, хвоста до 15 см, сравнительно
короткие уши (1,5-2 см). Окраска его спины бывает от бледно-песчаной до
охристо-коричневатой, низ и бока белые, с широкой размытой желтой полосой по
бедру. "Знамя" на хвосте развито хорошо. Мохноногий тушканчик живет в
барханных, грядовых, развеянных и полузакрепленных песках. Он заходит по массивам
песков далеко на север, до Среднего Поволжья, распространенно всему Казахстану,
в Средней Азии, Северном Афганистане, Монголии и Северном Китае. Поедает семена
и зеленые части растений, выкапывает луковицы и клубни, часто ест и насекомых.
Норы бывают до 5 м в длину. Особенно длинные и сложные норы, с рядом
запасных ходов, роют самки. Они приносят один, а иногда и два помета в год,
причем в выводке бывает до 8 детенышей.
Гребнепалый тушканчик (Paradipus ctenodactylus) до 16 см длиной.
На ступне длинная двухъярусная щетка. Живет он в закрепленных песках Кызылкума
и Восточных Каракумов. Примечательно, что гребнепалый тушканчик ловко лазает по
кустам, при беге делает прыжки до 3 м, но может и "бежать" на
задних ногах, ступая на них поочередно.
Гребнепалый тушканчик любит также
"играть" на песке, кружась и прыгая на небольшой площадке, а иногда
валяется по песку.
Песчаные тушканчики (род Jaculus) - зверьки средних
размеров (до 15 см длиной), с щеткой волос на ступне и с хорошо развитым
"знаменем" на хвосте. Тушканчик Лихтенштейна (J. lichtensteini) (длиной 10-11 см), живет
в песчаных пустынях Каракумов, Кызылкума и Прибалхашья.
В нашей стране живет также туркменский
тушканчик (Т. turkmenicus), другие виды песчаных тушканчиков (2 или 3
вида) населяют Северную Африку и Юго-Западную Азию.
Семейство Мышовковые (Zapodidae)
В это семейство выделяются мелкие, внешне
похожие на мышей грызуны, главное отличие которых от мышей заключается в числе
верхних коренных зубов: почти у всех мышовковых их с каждой стороны верхней челюсти
по 4 (в нижней челюсти, как у мышей,- по 3). Хвост обычно значительно превышает
длину тела; он покрыт роговыми чешуйками и редкими короткими волосками, не
образующими кисти на конце. Распространены мышовковые в Европе, Азии и Северной
Америке. К северу в Европе доходят до Полярного круга, в Западной Сибири - до
64-66° с. Дальше до северной половины Байкала и Среднего Приамурья; к югу - до
берегов Черного и Азовского морей, Закавказья, Тянь-Шаня, Западных Гималаев,
Ганьсу и Сычуани; в Америке к северу почти до северных берегов Аляски и
материковой Канады, а к югу - до Калифорнии, Нью-Мексико, Арканзаса и Южной
Каролины.
Мышовки (род Sicista) имеют черную полоску
вдоль спины или одинаковую окраску спины и боков без резкой границы с
желтоватой, охристой или серой (не белой) окраской низа. Длина ступни около 20
мм. 4, 5 или 6 видов мышовок распространены в Евразии от Норвегии, Дании
и Венгрии до Сахалина.
Для всех мышовок характерны зимняя спячка
длительностью до 8-9 месяцев, сумеречная и ночная активность в летнее время,
питание преимущественно семенами растений и насекомыми. Один раз в году самка
рождает от 2 до 7 (обычно 4-5) детенышей.
Степная мышовка (Sicista subtilis) самая мелкая: длина
тела 6-8 см, длина хвоста не более 9 см. На спине (от затылка или
лопаток до основания хвоста) ясная черная полоска, верх и бока серые или
охристо-серые. Распространена по зоне степей, южному лесостепью и северным
полупустыням от Венгрии и Румынии до юго-западного берега Байкала. Наиболее
полные сведения о жизни этой мышовки в степях Казахстана собраны В. Е. Флинтом
(1960). Живет она во всех степных биотопах с хорошо развитым травяным покровом.
Поселяется она в нежилых норах серых полевок или степных пеструшек, в трещинах
и под пластами земли на межах пашен, а сами нор, видимо, не роют. В природе
степные мышовки редко встречаются в светлое время суток, но в неволе бывают
деятельными и днем. Из семян различных растений в неволе они предпочитали
семечки подсолнуха и зерна пшеницы. Но когда им вместе с зернами пшеницы, овса
и гречихи давали насекомых, зверьки ели их в первую очередь. По наблюдениям А.
А. Гаджия, одна мышовка за 26 дней жизни в неволе съела 2154 вредных насекомых.
Размножение степных мышовок, по
наблюдениям В. Е. Флинта, протекает не так, как у других видов мышовок. В конце
мая - начале июня приносят потомство перезимовавшие самки, масса которых более
9,5 г, а в конце июня - начале июля потомство воспитывают самки, масса которых
7-9,5 г; более крупные самки в это время в природе не встречаются. Создается
впечатление, что к концу июня половозрелыми становятся самки того же года
рождения. Перезимовав, они приносят второе потомство, но сами часто гибнут.
Лесная мышовка (S. betulina) с более длинным хвостом
(8,5-10 см), сверху и с боков рыжевато-коричневая, с черной полоской
посередине спины. Живет в пойменных, смешанных и лиственных лесах и в
полезащитных полосах от Центральной Норвегии, Нидерландов, Венгрии и Украинских
Карпат к востоку до Забайкалья. На Алтае поднимается до предгольцовых березовых
редколесий на высоте около 2000 м над уровнем моря, предпочитает лесные
насаждения с подлеском или кустами. Но встречается и на полях поблизости от
леса или полезащитных полос. В лесах зверьки этого вида охотнее всего
поселяются в гнилых, высоких пнях (обычно осиновых или березовых). В гнилой (но
не влажной) древесине, обычно под самой корой, мышовка прогрызает ходы
диаметром около 2 см общей протяженностью до 5 - 6 и даже 10 л. В
эту систему ходов ведет снаружи только одно отверстие, обычно укрытое куском
отставшей коры и расположенное на высоте 0,3-1 м от земли. Характерно, что при
прогрызании ходов труха не выбрасывается зверьком наружу, а заполняет до 80%
протяженности ходов и не мешает мышовке передвигаться по ним. В таких гнездах
обнаруживались только самцы и молодые. В пне, занятом взрослой самкой, над
"голой" камерой с пометом и хитиновыми остатками насекомых
располагается другая камера - с выстилкой из мха и листьев. В таком гнезде,
видимо, воспитываются дети хозяйки. Покинутые гнезда мышовок занимают другие
обитатели - бурозубки, ящерицы или шмели.
Питается лесная мышовка преимущественно
насекомыми, особенно муравьями, их личинками, а также личинками жуков,
проходящих развитие в древесине. Мышовка обнаруживает их на расстоянии и
прогрызает от магистрального хода боковой тупичок к укрывшемуся насекомому.
Активными мышовки бывают преимущественно в
сумерки и ночью; встречаются днем на ветках кустарников, но в полусонном
состоянии. Лишь с наступлением темноты вялость зверьков сменяется активностью.
Одноцветная мышовка (S. сон color и близкие к ней формы)
отличается одноцветной (рыжеватой) окраской верха, без черной полоски
посередине спины. Распространена она в горах и предгорьях Кавказа, Тянь-Шаня,
Алтая, Тарбагатая, Кашмира, Ганьсу, Сихотэ-Алиня и Южного Сахалина. В горах,
окаймляющих Иссык-Куль, эта мышовка встречается в предгорьях котловины и во
всех высотных поясах, от степей до альпийских лугов. Много ее в кустарниковых
зарослях у нижней границы леса, разреженных ельниках с кустарниковым подростом,
на лесных полянах и вырубках. По численности там она уступает только лесным
полевкам. В жилищах человека не поселяется. В спячке находится 8-9 месяцев в
год - с октября до мая. В горах пробуждается даже в последней декаде мая, а
залегает в спячку до второй половины сентября. Часто поселяется в старых,
трухлявых пнях.
Размножение ее начинается очень поздно. До
середины июня в уловах бывают только самцы. Самки появляются с 15 - 20 июня, а
беременные и кормящие - с конца июня до конца июля. В приплоде бывает 3-7
(обычно 4-5) детенышей. Молодые становятся самостоятельными (при массе 5-7 г)
с первой половины августа. В сентябре в уловах бывают только молодые, а
взрослые уже в августе впадают в спячку. Предельная продолжительность жизни
около трех лет. В питании весной преобладают животные корма (насекомые, черви,
моллюски) и зелень растений, а в августе и сентябре - семена и ягоды.
На Юго-Западном Сахалине в ольховнике
около Шебунино добыт взрослый самец одноцветной мышовки в дупле острокрылого
дятла, расположенном на высоте 3,8 м от земли. От дна дупла проложен
ход, засыпанный древесной трухой (как у лесной мышовки). По наблюдениям Л.
Тимофеевой, 7-граммовая одноцветная мышовка за сутки съедает 8-10 г насекомых
(до 140 крупных комнатных мух за 1 час). Когда нет насекомых, она может есть
орехи арахиса, творог и пить молоко. Зверек часто и тщательно чистит передними
лапками свой длинный хвост.
Американские полутушканчики (Zapus hudsonius и Z. princeps) отличаются от мышовок
трехцветной окраской. Хвост двухцветный, с темным верхом до самого конца.
Распространены эти зверьки от реки Юкон на Аляске, озера Невольничьего и
Гудзонова залива в Канаде к югу до Калифорнии, Аризоны и Южной Каролины, от
приморских берегов до альпийского пояса в горах. Встречаются на болотах, лугах,
лесных полянах и в сомкнутых лесах, в зарослях кустарников и т. п. Гнезда
устраивают в гнилых пнях, пустотах бревен, среди травы и кустов на земле или в
норах. Летом бывают активными преимущественно ночью. Передвигаются короткими
прыжками беспорядочным курсом, но напуганный зверек может совершать прыжки
длиной до 2 м и более. Питаются семенами трав, мякотью плодов и
насекомыми. Много пьют. Охотно и хорошо плавают. Детеныши (5-6 или до 8)
рождаются в июне и в сентябре. Беременность длится около 18 дней. Детеныши
родятся голыми, исключая маленькие усы, и слепыми. К 17-му дню жизни
покрываются волосами и уже совершают короткие прыжки, но глаза их открываются
только на 22-25-й день. Зрячие уже осмеливаются выходить из гнезда и едят
твердую пищу. В возрасте 4 недель способны жить независимо от родителей. К
осени накопляют значительные запасы жира и впадают в спячку поодиночке или
парами, в гнездах, расположенных ниже уровня промерзания земли. Пробуждаются
поздней весной. В отдельные годы встречаются очень редко, в другие годы в
подходящих местах бывают многочисленными.
Лесная полу мышь (Napaeozapus insi-gnis) похожа на
полутушканчиков, но не имеет верхних предкоренных зубов, в отличие от всех
мышовковых и подобно настоящим мышиным. Кончик длинного хвоста белый.
Распространена на востоке Америки к югу и юго-востоку от Гудзонова залива.
Живет в равнинных и горных лесах. Прыгает еще лучше, чем полутушканчики.
Входное отверстие в нору на день закупоривает землей. По питанию, годовому
циклу жизни и биологии размножения похожа на полутушканчиков.
Семейство Мышиные (Muridae)
Семейство объединяет очень разнообразных
по размерам, облику и образу жизни животных. Большинство представителей имеет
"мышевидную" или "крысовидную" внешность. Для всех видов
характерно отсутствие предкоренных зубов. С каждой стороны верхней или нижней
челюсти обычно имеется по одному резцу и по 3 коренных зуба. Изредка число
последних сокращается до 2 или даже до 1. Часть зоологов рассматривает
мышевидных грызунов как одно семейство мышиных, а другая часть делит их на
семейство мышиных (Muridae в узком смысле) и хомяковых (Cricetidae). Ископаемые остатки
мышиных в широком смысле этого слова известны в Евразии и Северной Америке
(лучше всего изученные территории) начиная по меньшей мере с олигоцена. Они
найдены также в плиоцене Южной Америки и плейстоцене Мадагаскара.
В настоящее время числится около 1500
видов мышиных, которые группируются примерно в 200 родов и 13 подсемейств.
Подсемейство Древесномышиные (Dendromurinae)
Древесномыши (Dendromus) - небольшие, размером
примерно с домовую мышь, грызуны, встречающиеся в большинстве районов Африки,
южнее Сахары. Длина тела 6-10 см, хвост немного длиннее - 7-12 см. Верх
серовато-коричневатый, часто с темным спинным "ремешком", низ
беловатый или желтоватый. Хвост без волос, покрыт чешуйками, мыши могут
обхватывать им ветки и стебли трав при лазании. На передних ногах всего три
пальца, длинных и хорошо приспособленных к цеплянью за ветви. Верхние резцы с
желобками. Зверьки хорошо лазают в траве, по кустам и деревьям, но в случае
опасности нередко прячутся в норы. Активны ночью. Свое шаровидное гнездо из
растительного волокна и пуха они помещают в траве, кустах (на высоте до 1 м)
либо в норах.
D. melanotis обитает в Восточной
Африке, преимущественно в сухих саваннах. Для нее характерна широкая продольная
темная полоска на спине от плечей до основания хвоста. Под ухом расположено
белое пятно. Другой зверек - D. mesomelas (рис. 120) живет преимущественно во влажной местности,
особенно в зарослях высоких трав и кустов на болотах. В Конго эта мышка нередко
поселяется в хижинах деревень.
D. myslacalis не имеет темного
"спинного ремешка". Обычны в зарослях бананов и в лесах. При этом
часто устраивают гнезда в пазухах их листьев. Самки древесной мыши рождают по
3-5 детенышей. Размножаются древесные мыши в разных частях ареала в различные
месяцы года, однако совсем молодые зверьки, недавно покинувшие родительское
гнездо, встречаются чаще всего между мартом и сентябрем. Древесные мыши -
спокойные и миролюбивые зверьки (может быть, за исключением весьма агрессивного
D. my-stacalis).
Жирные мыши, два вида которых (Stea-tomys pratensis и S. bocagei) объединены в особый
род, распространены по всему пространству Африки, от Судана до Капской
провинции, избегая лишь влажных лесов и болот. Длина тела от 7 до 12 см, хвост
короткий - 4-5 см. Мех мягкий и шелковистый, сверху коричневатый с
отдельными черными волосками, снизу белый. Уши большие и округлые. Ведут ночной
образ жизни. Сооружают норы длиной 1,5-2 м, глубиной около 1 м. Днем
вход закрывается пробкой из земли. В большой камере, в конце хода, они
устраивают гнездо из сухих травинок. Самка рождает за один раз 4-6 детенышей.
Пища их - трава, семена и насекомые. В течение нескольких (до 6) засушливых
месяцев зверьки впадают в спячку, а перед этим нагуливают такое количество
жира, что используются местными жителями в пищу как деликатес.
Подсемейство Деомиины (Deomyinae)
В подсемейство выделяют один вид - деомис
(Deomys ferrugineus) из лесов бассейна
Конго. Это средних размеров зверек (тело 12-16 си, хвост 17-20 см), с
очень длинным и мягким мехом красноватого цвета, с белыми лапками и кончиком
хвоста. Уши большие и округлые. Морда вытянута. Деомис ведет ночной и
преимущественно древесный образ жизни. При движениях в кронах он использует
свой хвост не только как балансир, но и как хватательный орган. Задние ноги
очень длинные, и грызун может отлично прыгать с ветки на ветку и по земле.
Основная его пища - мягкие фрукты и мелкие беспозвоночные животные.
Подсемейство Мышиные (Murinae)
Мышиные объединяют основную часть видов
"настоящих" мышей и крыс с трехрядным расположением бугорков на
верхних коренных зубах. Распространены в Евразии, Африке и Австралии. В Новый
Свет завезены человеком в историческое время.
Обезьяновая мышь (Hapalomys longi-caudatus) считается наиболее
примитивной из мышей. Размером этот коричневатый зверек с крупную мышь
(например, лесную) и имеет очень длинный хвост. Ведет ночной и древесный образ
жизни.
Обитает во влажных тропических лесах
Индонезии, Таиланда и прилежащих территорий. Питается различными плодами н
семенами деревьев. Гнездо устраивает в дуплах. Долгохвостая мышь (Vandeleuria oleracea) при длине тела 6-8 см
имеет хвост длиной 10-13 см, который хорошо опушен. Первый и пятый
пальцы вместо обычных коготков несут плоские ногти. Образ жизни исключительно
древесный. День проводит в гнезде, которое располагает в дуплах или в густых
ветвях. Ночью зверек быстро бегает по ветвям в поисках плодов и семян. Хвост
служит ему балансиром и также может обхватывать ветки, превращаясь в
"пятую конечность". Размножение длится круглый год. В одном выводке
обычно бывает от 3 до 6 мышат. Распространены долгохвостые мыши во влажных
тропических лесах Юго-Восточной Азии, на юге Индии и на Цейлоне, на север
доходят до китайской провинции Юньнань. Эти маленькие грызуны хорошо живут в
неволе.
Мышь-малютка (Micromys minutus) - самый мелкий зверек
из всех видов семейства. Длина тела 6-7 см, хвоста - 5- 1 см, масса
- 5-6 г. Сверху окраска меха коричневатая, на груди и брюхе светлеющая.
Хвост хватательный, может обвивать стебли
трав и кустов при лазании. Среди высоких трав и кустов мышь-малютка сооружает
шарообразное гнездо из стеблей диаметром 6-13 см. Высота гнезда от
поверхности обычно около метра. Гнездо предназначено для выведения детенышей. В
одном выводке бывает от 5-9 до 12 мышат. Беременность длится 21 день. Вне
периода размножения мыши-малютки живут в различных наземных убежищах - среди
валежника, под скирдами и стогами. На зиму делают запасы семян. Летом, кроме
семян, охотно поедают также насекомых. Распространены мыши-малютки в умеренных
широтах Евразии. В СССР на Дальнем Востоке местами может вредить посевам. Эти
маленькие мышки весьма мало пугливы. В неволе они прекрасно живут, питаясь тем
же, чем их хозяин.
В умеренных и субтропических широтах
Старого Света распространены 7 видов рода Apodemus. Это среднего и крупного
размера мыши (длина тела 7-12 см), хвост обычно равен или немного
превосходит длину тела. Масса взрослых от 15 до 30 г.
Беременность длится 21-29 дней; 3-4 раза в
год рождается по 2-9 детенышей (обычно 4-6). Гнездо помещается в норах или в
закрытых наземных убежищах. Некоторые виды селятся в дуплах. Наиболее массовые
и известные виды - лесная и желтогорлая мыши (A. sylvaticus и А. flavicollis), обитающие в смешанных
и лиственных лесах западной половины Евразии. Первый вид проникает также в
Северную Африку. На Дальнем Востоке и в Восточной Сибири их заменяет азиатская
мышь (A.
speciosus). В лесостепной и
степной зонах Евразии встречается полевая мышь (A. agrarius) - красивый зверек
рыжеватой окраски, с черным "ремешком" вдоль спины. Все мыши также
регулярно поедают насекомых и других беспозвоночных животных.
Внешне сходны с песчанками 5 видов
африканских мышей из рода Thamnomys. Это сравнительно крупные зверьки с длиной тела
10-14 см, хвоста -14-20 см. Спина их каштановая или светло-красновато-коричневая.
Брюхо белое. На конце хорошо опущенного хвоста - небольшая кисточка из волос.
Низ белый или светло-коричневый. Хвост чешуйчатый, почти без волос. Обитают в
лесах и кустарниках от Ганы до Западной Уганды. В горы идут до высоты 4000 м
над уровнем моря. В дуплах или в развилках ветвей устраивают гнездо из
травы и сухих листьев. Ведут исключительно древесный и ночной образ жизни.
Питаются плодами и листьями. Размножаются почти круглый год. В отличие от
большинства других видов мышиных они часто обитают во влажных экваториальных
лесах, например Т. venustus (рис. 121) - в Конго и Западной Уганде. К предыдущей группе
близки кустарниковые крысы, или куру (род Grammomys),8 видов которых
распространены в Африке, от Судана к югу до конца материка. Длина тела 10-16см,
хвоста - 13- 20 см, масса -20-60 г. Верх темно-коричневый,
красноватый или серовато-коричневый, бока и низ светлые. Хвост покрыт чешуйками,
почти лишен волос, приносит 3 детенышей, размножение прерывается в засушливые
сезоны года тает куру в лесах и среди зарослей кустарника. Активен в любое
время суток. Хорошо лазает по ветвям. Гнездо ел среди ветвей. Иногда занимает
птичьи гнезда, причем может при этомзаодно позавтракать их содержимым.
Вообще питается главным образом зелеными частями растений и плодами.
Длиннохвостая куру (G. dolichurus) из Восточной Африки - любимая добыча змеи Boaedon lineatus.
Кроликовые крысы (Conilurus) - крупные (длина тела
16 - 20 см и хвост 18-21 см), черновато-коричневые или песчаные
грызуны. Низ белый или желтый. Хвост покрыт густым мехом известны 2 вида.
Чернохвостая крыса (C. penicillatus) распространена в Северной Австралии и на Новой Гвинее.
Занимает прибрежные заросли кустарников, обитает в лесу и на болотах. Часто на
морском берегу в полосе прибоя coбирает пищу, выброшенную волнами, толстых ветвей
или в дупле самка устраивает гнездо. Новорожденные кроликовые крысята, в
отличие от других представителей мышиных, покрыты мехом, быстро прозревают и
развиваются длина тела 24-35 см немного длиннее тела) обычны в светлых
эвкалиптовых лесах Австралии. Мантбулы Гульда (Mesembriomys gouldi) обитает в северных
частях материка и в Квинселде, другой вид (М. macrurus) - в прибрежных частях
Западной Австралии. Мантбул Гульда сверху окрашен в серовато-желтый цвет, ноги
черные, но пальцы белые. Ведет ночной образ жизни. Большую часть времени
проводит на деревьях, в которых устраивает гнездо из сухих и травы. Очень любит
плоды пандуса. В свою очередь, местные жители считают мантбула большим
лакомством и по этому усиленно на него охотятся
В роде Oenomys ржавоносая (Oenomys hypoxanthus) - единственный
представитель Длина 14-18 см, хвоста - 17-20 см, 60-90 г. Мягкий
и пушистый варьирует от коричневато-красноватого до темно-оливкового оттенка.
Верх носа рыжевато-красный, за что зверек и получил свое название. Низ белый
или светло-коричневый. Хвост чешуйчатый, почти без волос. Распространена
ржавоносая крыса от Кении и Уганды до Ганы и на юг до Анголы и Конго. В горах
она встречается до высоты 3000 м. Многочисленна в расчищенных лесах, на
вырубках, по краям полей, где может приносить вред урожаю. Питается семенами и
зелеными частями растений. Сооружает под землей сложные норы, с многочисленными
туннелями. Больших скоплений не образует, чаще живет поодиночке. Размножение
приурочено к сезону от сентября до апреля. Африканцы ловят ржавоносую крысу
из-за вкусного мяса.
Косматая крыса (Dasymys incomtus) также единственный
представитель отдельного рода. Коричневато-оливкового цвета, примерно такого же
размера, как и ржалоносая крыса, но хвост немного короче. Масса около 60 г. Распространена
от Центральной до Южной Африки. Селится главным образом по болотам и на берегах
водоемов, в зарослях тростника и в высокотравье. Встречается и высоко в горах.
В месте своего обитания многочисленна. Очень хорошо плавает. В пустотах под
берегом или на поверхности в густой растительности, под колодами, в основаниях
кустов устраивает гнездо из переплетенных сухих трав. Самка обычно рождает 2-4
детенышей, размножение длится круглый год, но особенно интенсивно с июня по
октябрь. Активные зверьки встречаются в любое время суток. Питается околоводной
и болотной растительностью, преимущественно зелеными частями трав. Одни из
самых многочисленных африканских грызунов - травяные мыши (Агvicanthis), 4 вида которых распространены
на большей части континента, а также на юге Аравийского полуострова. Особенно
заметны они в Восточной Африке, к югу до Малави.
Размер травяных мышей крупный: длина тела
12-19 см, хвоста 9-16 см, масса - 50-100 г. Окраска
серовато-коричневая, низ слегка светлее. Мех длинный с отдельными колючими
щетинками, у некоторых видов с настоящими тонкими иголками. Населяют саванны,
кустарниковые заросли, светлые леса. Живут в норах, иногда занимают пустые
термитники. Часто образуют колониальные поселения, проторяя в густой траве
дорожки, сходные с ходами полевок. Питаются разнообразной растительной пищей,
часто вредят посевам и запасам зерна в амбарах, могут селиться в жилищах
человека. Активны днем и ночью. В неволе зверек живет до 7-8 лет. В Уганде A. niloticus (рис. 123) размножается
круглый год, но максимум размножения приходится на конец дождливого и начало
сухого сезона. Обычное число новорожденных - 4. Особенно любит кормиться
листьями и стеблями Ameranthus polygamus. Норы чаще роет у куч валежника или у основания
кустов. Гнездовую камеру помещает на глубине 20-60 см. В одной системе
нор обычно обитает не менее 4-5 пар взрослых зверьков. Часто является основным
кормом хищных птиц, особенно черноплечего коршуна (Elanus coeruleus).
Мышь Эллиота (Golunda ellioti) распространена на
Цейлоне, в Западном Пакистане, Пенджабе, Непале и Бутане. Длина тела 11-15 см,
хвост несколько короче. Сверху серовато-желтовато-коричневая, снизу почти
белая. По общему облику напоминает крупную лесную полевку из рода Clethrionomys. Среди густого мягкого
меха на спине есть жесткие колючие щетинки. Верхние резцы - с желобками.
Обитает на болотах, лугах, по краю густых джунглей и около посевов. Активна
утром и вечером. Живет семейными группами. Норы роет редко, гнездо обычно
устраивает в каких-либо наземных убежищах вроде кучи валежника или под стволом
поваленного дерева. Гнездо представляет собой шар диаметром 15-23 см, сделанный
из плотного растительного войлока. Самка рождает обычно по 3-4 детеныша
несколько раз в год. Зверьки добывают корм на земле и на деревьях, по которым
могут передвигаться с большой ловкостью. Питаются исключительно растительной
пищей. На Цейлоне бывают в большом количестве на кофейных плантациях, где
приносят большой вред, поедая почки и цветы на кофейных деревьях.
Ручьевые крысы (Pelomys) внешне сходны с
травяными мышами, но резцы у них желобчатые. Окраска разнообразных оттенков
коричневого цвета, мех жесткий, частично щетинистый. У некоторых видов (всего
известно 9) на спине - узкий продольный "ремешок". Длина тела 12-22
см, хвост может быть длиннее и короче тела. Обычно обитают во влажных
местах у рек, ручьев, озер и на болотах, также могут быть встречены по опушкам
лесов. В Восточной Африке P. fallax может размножаться круглый год, принося в одном помете до 7-9
детенышей. Этот вид в Юго-Западной Африке ведет дневной образ жизни, в
остальных частях ареала преимущественно ночной. В Танзании известен как
вредитель полей. В Анголе сооружает глубокие норы, но в других районах Африки
строит наземные гнезда.
Пестрые мыши (Lemniscomys) обитают на большей
части Африки южнее Сахары, Всего известно 6 внешне сходных видов. Живущая в
Гане и прилежащих странах Западной Африки L. striatus (табл. 16) - характерный
представитель группы.
Длина тела пестрых мышей 10-14 см, хвоста-
10-16 еж. На темном фоне спины и боков протягиваются прерывистые светлые
полоски. Обитают в высокотравной саванне и по опушкам лесов, в горы поднимаются
до высоты 2100 м. Часто поселяются в чужих норах, хотя вообще-то
способны устраивать и свои. В одном помете обычно бывает 2-5 детенышей, хотя
ловились беременные самки даже с 12 эмбрионами. Размножение возможно круглый
год, хотя некоторые виды прекращают размножаться в сухие сезоны. Активны
преимущественно днем. Питаются главным образом растительной пищей, в основном
плодами, корнеплодами, мягкими семенами. Иногда поедают насекомых.
Полосатая мышь (Rhabdomys pumilio) - представитель
монотипического рода. Длина тела около 11 см, хвост примерно такой же
длины. Очень характерная окраска: на спине чередуются темные (коричневые) и
светлые (желтоватые) полосы. Хвост с редкими щетинистыми волосками. Обитает в
высокотравье среди кустарников и сухого буша, вдоль русел высохших водотоков,
по опушкам лесов и у посевов. В ряде мест обычна. Активна днем. В Восточной
Африке селится преимущественно в горах. Может передвигаться среди ветвей
деревьев и кустарников. Роет норы или устраивает гнездо среди корней и в густой
траве. Самка приносит от 3 до 10 детенышей за один раз. Размножение бывает с
сентября по апрель (конец дождливого - начало сухого сезона). Обычны 4 выводка
в год. В возрасте 3 месяцев способна принять участие в размножении. В одном
гнезде часто живет семья в 10-30 особей (родители и разновозрастное потомство).
Акациевые крысы (Thallomys) - весьма многочисленная
группа мышиных в Африке, от Кении до Анголы. Длина тела 12-16 см, хвоста
- 13-21 см. Мех высокий, коричневато-серой окраски. Бока сероватые, низ
белый (рис. 124). Известно 5 видов. Часто акациевых крыс видели и добывали в
куртинах акаций. Гнезда устраивают в дуплах, ветвях и на земле, между корнями
акаций. В больших старых деревьях образуются целые колонии акациевых крыс. День
проводят в гнезде, активны в сумерки и ночью. Могут совсем не пить воды,
довольствуясь влагой трав, семян и плодов. Нередко поедают также смолистые
подтеки на стволах. Размножение приходится на влажный летний период. Самка
рождает от 1 до 4 детенышей.
Настоящие крысы (Rattus) - одни из наиболее известных грызунов, два вида
которых - серая крыса, или. пасюк (R. погvegicus), и черная крыса (R. rattus) - с помощью человека
расселились почти по всем странам мира. Альбиносы черных крыс разводятся в
лабораториях. Кроме того, ряд других видов многочисленны в тропических и
субтропических широтах Старого Света. Длина тела от 8 до 30 см, хвост
может быть короче или длиннее тела. Окраска сверху серая, черная, коричневатая
или рыжеватая, снизу сероватая или белая.
Ведут наземный, роющий, лазающий и
околоводный образ жизни. Соответственно убежища могут располагаться в самых
разнообразных условиях: в норах, среди камней и скал, в дуплах, в древесных
гнездах. Есть виды, ведущие дневную активность (полинезийская крыса - R. exulans), большинство - ночные
зверьки. Пища очень разнообразна, часто со значительной долей животных кормов.
Беременность длится 21-30 дней, большинство видов обладает большим потенциалом
размножения и может давать вспышки высокой численности. Известны как
микробоносители и переносчики различных возбудителей опасных болезней человека
и домашних животных, а также как вредители запасов пищи и урожая.
Пасюк (R. norvegicus) в зоологической,
учебной, популярной, производственной и медицинской литературе называется
просто крысой, обыкновенной, серой, бурой или амбарной, крысой. Все это
касается одного вида. Длина тела пасюка 18-25 (до 27) см; хвост
составляет около 80% от длины тела, а длина уха около 2/3 длины
ступни. Распространен почти по всему свету. В СССР приурочен главным образом к
лесолуговой и степной зонам, а также к морским портам и речным пристаням. В
водораздельных ландшафтах тундры, лесотундры, тайги, полупустынь и пустынь не
встречается. В 40-х годах завезен в Ташкент, где укоренился и расселился по
городу и окрестным селениям.
Живет обычно в постройках человека, но на
юге страны селится и по берегам водоемов, вдали от построек. Размножается в
природных биотопах только в теплые месяцы, а в постройках - круглый год. В
каждом приплоде бывает от 1 до 17 (в среднем 8-10) детенышей. За год самка
приносит 1 или 2 (реже 3) приплода. Питается разнообразной, но преимущественно
животной и в меньшей степени растительной пищей. Приносит вред истреблением и
порчей продовольственных продуктов, порчей построек и мебели. Основной источник
чумы, крысиного риккетсиоза, лептоспирозной желтухи и около трех десятков
других опасных для человека болезней. Самый вредный из грызунов всего мира.
Австралийские мыши (род Gyomys, 8 видов) населяют весь
австралийский континент, за исключением лишь влажного севера. Длина тела 7-13
см, хвоста - 6-14 см. Эти мыши бывают самых различных оттенков:
оливкового, песчаного и пепельного. Низ светлый, часто белый. Австралийские
мыши обитают в высокотравье и эвкалиптовых лесах (в последних G. fumeus - массовый вид).
Населяют также скалистые участки гор, песчаные равнины. Селящийся по пескам G. alboci-nereus роет глубокие, до 1 м,
системы нор с несколькими выходами. Питаются главным образом насекомыми с
добавлением семян и зелени трав. Размножение приурочено к австралийской весне.
В ноябре - декабре рождается по 3-5 детенышей .
В Австралии же обитают 3 вида прутогнездных
крыс (Leporillus): L. conditor и L. apicalis. Длина их тела 14 -20 см. Хвост хорошо опушен, с
небольшой кисточкой на конце. В спокойном положении зверьки напоминают
небольших кроликов с длинным хвостом, так как имеют длинные уши. Недаром
буквальный перевод латинского родового названия означает "зайчик".
Два вида (L.
conditor и L. jonesi) сооружают
"дома" из прутьев. Строение представляет собой сложенную в виде
плоского конуса кучу прутьев диаметром около 6 м и высотой до 1,3 м. Прутья,
идущие на постройку, достигают длины 1 м и толщины 2-3 см. На
постройку убежища тратится много сил и стараний, прутья тщательно монтируются,
и в результате постройка получается очень прочной. "Дом" служит для
устройства внутри гнезд, защищенных от хищников и в особенности от частых
сильных ветров. В зависимости от местных условий он может иметь более или менее
вытянутую форму. Иногда постройку делают вокруг небольшого куста или камня. В
местности без достаточного количества хвороста прутогнездные крысы могут
селиться в выбросах кроличьих нор и среди куч сухой травы. При этом на вершину
таких травяных охапок помещают камни. После этого ветер уже не может разрушить
облюбованных убежищ. Внутри кучи из прутьев или травы они прокладывают много
ходов и устраивают несколько гнезд из стеблей травы и растительного пуха. В
каждом гнезде живет пара взрослых и выводок из 3-4 молодых. L. apicalis - ночной колониальный
зверек, часто занимает дупла и готовые постройки.
Некоторые хищные птицы в безлесной
местности опрометчиво устраивают свои гнезда на "крыше дома" L. conditor. В этом случае грызуны
нередко поедают отложенные яйца и даже маленьких птенцов своего заклятого
врага. Вообще же прутогнездные крысы преимущественно вегетарианцы. Эти зверьки
хорошо живут в неволе, где быстро привыкают и могут размножаться.
В Центральной, Восточной и Южной
Австралии, а также на Тасмании обычные грызуны - представители рода псевдомисов
(Pseudomys), в который объединяют
10 видов. Окраска их сероватых, желтоватых и коричневых тонов. Низ светлый.
Длина тела 7-16 см, хвост примерно равен длине тела. Наиболее типичные
представители рода - австралийский (P. minnie) и тасманийский (P. higginsi) псевдомисы, с
длиной тела около 13 см и массой около 70-80 г. Обитают в
травянистых равнинных и холмистых местах, нередки в зарослях кустарника у русел
пересыхающих водотоков, у болот и небольших соленых водоемов. Роют неглубокие
норы с одним входом, где устраивают свои гнезда. Самка приносит от 2 до 4
детенышей. Двенадцать видов банановых мышей (род Melomys) распространены на Новой
Гвинее и прилежащих островах, на севере Австралии, в Квинсленде, Новом Южном
Уэльсе, на архипелаге Бисмарка и на Соломоновых островах. Длина тела этих
грызунов от 9 до 18 см, хвост длинный (11 -18 см). Мягкий и
длинный мех коричневатой или рыжеватой расцветки. Снизу окраска белая или
кремовая. Хвост голый и чешуйчатый, на каждую чешуйку приходится один волосок.
Поверхность хвоста напоминает поверхность напильника, выложена мелкими
выпуклыми чешуйками в форме своеобразной мозаики. Банановая мышь поселяется на
лугах, болотах, на плантациях сахарного тростника, в зарослях трав и
кустарников, около рек и озер. Хорошо лазает, причем хвост может служить
дополнительной "зацепкой". Гнездо в виде шара диаметром 12-20 см из
травы помещает в кустах, кроне дерева или в густой траве, реже - в просто
устроенной норе с одним входом. Размножается в течение сезона дождей (обычно с
ноября по март). Одно - и двухнедельные детеныши цепляются за мать, которая
носит их с собой на брюхе между сосков. Это обстоятельство вначале породило у
зоологов ошибку - банановых мышей приняли было за сумчатых животных. После двух
недель молодые могут самостоятельно передвигаться и питаться, но при малейшей
тревоге спасаются на брюхе своей родительницы. Зрелость наступает в возрасте 6-7
месяцев. Питаются различными фруктами, ягодами, орехами.
Два вида болотных крыс (Malacomys) водятся в Западной
Африке, от Либерии до Нигерии (М. edwardsi) и от Анголы и Конго до Либерии (М. longipes). Длина тела около 15
см, хвост несколько длиннее, масса - 90-100 г. Зверьки покрыты
густым бархатистым мехом, на спине темно-коричневым, на боках с охристым
оттенком и снизу серым. Характерны длинные ноги. Обитают в густой околоводной
растительности, а также во влажных лесных участках; гнезда они делают на поверхности
земли в виде травяного шара. Нередко (особенно в слишком мокрых местах) гнездо
помещают и над поверхностью в кроне куста или низкого густого дерева.
Размножаются с марта по июнь, но отдельные особи и в другие месяцы. Чаще всего
рождаются 3 детеныша. Кроме зелени трав, сочных стеблей, корневищ, семян и
плодов, охотно поедают насекомых, слизней, даже жаб и лягушек. Наиболее
известны нам домовые мыши (род Mus), а также их одомашненные альбиносы - белые
мыши, которые используются как лабораторные животные. Всего известно 16 видов
этого рода, распространенных в тропических, субтропических зонах и на юге
умеренных широт Евразии и Африки.
Домовая мышь (Mus musculus) вместе с человеком
расселилась по всем континентам, обитая как в постройках человека, так и в
природе. Длина тела ее 6-10 см, хвоста - 5-11 см, масса - 18- 30.
Окраска разных оттенков, с преобладанием серого цвета. 125).
Большинство других видов настоящих мышей
из рода Mus внешне весьма сходны с домовой. Широко распространена в Уганде и
прилежащих странах малая мышь (М. minutoides) массой всего 6 - 6,5 г
(рис. 126).
Весьма близка внешне к домовым мышам
группа австралийских мышей, 7 видов которых объединены в род Leggadinа. Распространены
австралийские мыши преимущественно на севере и востоке континента. Длина их
тела 5-10 см, хвоста - 5-9 см. Окраска коричневатая, иногда
желтовато-коричневая, снизу более светлая, почти белая. Обитают по сухим
травянистым равнинам, у опушек лесов, по морским побережьям. Строят простые
норы, с одним входом. Глубина нор у L. delicatula до 60 см, а длина туннеля не более 2 м.
Другой вид (L. hermannsbur-gensis) роет еще более простые убежища, глубиной не более 20 см и длиной
хода 1 м. В одной норе живет семья до 5 - 6 особей. Гнездо устраивают из
сухой травы и листьев. Особо сильной интенсивности размножение достигает в
сезон дождей, хотя не прекращается круглый год. За один раз самка приносит 3-4
детенышей. Питаются грызуны растительной пищей: семенами, зеленью и кореньями
трав. В засушливых районах несколько семей могут селиться вместе, располагая
свои норы на участке диаметром 1 м. При этом зверьки сооружают у входов
общий плоский курганчик из гальки. На камнях по утрам собираются капли росы,
служащие единственным напитком для обитателей колонии.
Жесткошерстные мыши (Lophuromys) - одни из самых
массовых представителей семейства на обширных пространствах
Африки, от Эфиопии, Кении, Уганды,
Танзании до Камеруна, Ганы, Габона и Анголы. Известно 10 видов этих зверьков.
Длина тела их от 10 до 14,5 см, хвост обычно короче (5 - 11,5см).
Имеются два типа окраски: темная, коричневая или оливковатая, или пестрая,
когда на темном фоне расположены отдельные беловатые, желтоватые или
оранжеватые пестрины. У некоторых видов основание волос меха оранжевого или
тускло-оранжевого цвета. Такая черта уникальна для африканских млекопитающих.
Жесткошерстные мыши обитают в
разнообразных местах: в лесах, зарослях трав и кустарника, на болотах, на
полях, в зарослях тростника. В горы поднимаются до верхней границы леса (до
4000 м). Активны в любое время суток. Гнезда устраивают в норах или
наземных убежищах: под колодами, валежником, в густой растительности. Питаются
в значительной степени насекомыми и другими беспозвоночными животными, а также
жабами, мелкими ящерицами. Из растительной пищи предпочитают сочные корнеплоды
и фрукты.
Кенгуровые мыши (Notomys) по облику напоминают не
столько миниатюрного кенгуру, сколько тушканчика. Это крупные для мышей грызуны
(длина тела 9-18 см, хвоста - 12-26 см), песчаного, пепельного
или коричневого цвета со спины и белые снизу. У них очень большие уши и глаза,
на длинном хвосте - небольшая кисточка, задние ноги сильно удлинены, характерны
большие и сильные резцы. Обычно бегают на всех ногах, но при остановках
опираются только на задние. Известно 10 видов, обитающих на большей части
Австралии: в пустынях, степях, зарослях кустарника и в светлых сухих лесах.
Активны в сумерки и ночью, а день проводят
в гнезде, которое помещают в норах. Обычно самка сама роет нору и устраивает
гнездо, прежде чем произвести на свет 2-5 детенышей. Сумчатые зверьки из рода Antechinomys весьма похожи на
кенгуровых мышеи и ведут сходный образ жизни. Более того, оба
"двойника" нередко занимают одну систему нор и буквально живут бок о
бок. Питаются зеленью трав, семенами и ягодами. Австралийские аборигены охотно
ловят кенгуровых мышей ради их вкусного мяса.
Колючие мыши (Acomys dimidiatus и 3 или 4 близких к ним
вида) - "мышиной" величины (длина тела 7 -12 см, хвоста б-12
см), с колючками среди грубых волос. Их голый чешуйчатый хвост очень
ломок и легко теряется в сложных жизненных ситуациях, поэтому среди обитающих в
природе особей многие имеют благоприобретенную короткохвостость. Уши большие,
стоячие. На верхних резцах - продольные желобки. Распространены почти по всей
Африке Гравии, в Иране и Пакистане. Живут в сухих саваннах и полупустынях на
каменистых или песчаных участках. Укрываются в норах или среди каменных
россыпей и в трещинах скал. В Африке часто занимают термитники. Самки массой
50-90 г с февраля до сентября, после 42 дней беременности, приносят по 1-3
детеныша (реже до 5) массой 5-6 г каждый. Новорожденные имеют открытые глаза и
уже в возрасте двух недель
перестают питаться молоком. Самцы в
возрасте 7 недель уже могут участвовать в размножении. Колючие мыши всеядны, но
предпочитают растительные корма, особенно зерна злаков.
Каирская мышь (A. cahirinus) в Египте живет главным
образом в постройках человека и очень сходна по образу жизни с домовой мышью.
Род бандикотов (Bandicota). Индийская и бенгальская бандикоты (В. indica и В. bengalensis) - крупные крысовидные
грызуны (рис. 129): длина тела 16-36 см, хвоста - 14-26 см, масса
- 600-1100 г. Морда тупая, широкая. На передних лапах длинные и крепкие
когти. Окраска верха от серого и коричневого до почти черного, низ
грязно-белый. Резцы желтые или оранжевые. Распространены в тропической Азии:
Цейлон, Индостан, Непал, Ассам, Бирма, Южный Китай, Таиланд, Индокитай,
Малайзия, Суматра и Ява. Индийская бандикота завезена на Тайвань, где сильно
размножилась. Пластинчатозубая крыса (Nesokia indiса) - единственный
представитель рода Ne.sokia. Коренастого сложения, длина тела 14-21,5 см, хвоста - 9
-13 см, масса около 150 г. Хвост голый. Окраска верха серовато -
или желтовато-коричневая. Ноги с большими когтями. Резцы широкие и крепкие.
Распространена в Египте, Сирии, Израиле, на севере Аравийского полуострова, в
Иране, Ираке, Афганистане, Западном Пакистане, на севере Индии и обычна у нас
на юге Средней Азии. Типичный житель долин рек и оазисов. Ведет роющий образ
жизни. На глубине 15 - 70 см сооружает сложную систему нор. При этом
может вредить ирригационным сооружениям. Питается растительной пищей, главным
образом подземными частями трав. Устраивает запасы пищи. По ночам выходит на
поверх ность на короткое время. В системах нор живет по нескольку семей,
образуя колонию. Иногда может селиться в постройках человека. Самка 3-4 раза в
год приносит по 3-5 детенышей. Может достигать высокой численности и сильно
вредить на поливных землях посевам кормовых трав, риса, бахчевым и огородным
культурам. Размножение длится круглый год, но особо интенсивно с марта по июнь.
В некоторых местах пластинчатозубую крысу ловят и едят, а также специально
разыскивают собранные ею запасы пищи.
Мешотчатые крысы (Beamys) характеризуются
наличием защечных мешков для переноса пищи. В Кении водится В. hin-dei размером 11 см, хвост
чуть короче тела. В Малави и Замбии обитает В. major более крупного размера:
длина тела 14,5- 16 см, хвоста - 13 - 14,5 см. Хвост голый, с
белым концом, без чешуек. Зверек ведет древесный образ жизни, редко ловится.
Самка рождает 4 детенышей.
Короткохвостая крыса (Saccostomus campestris) - коренастый зверек, с
длиной тела 12-15 см, с коротким (3-6 см) хвостом. Масса - 50-70
г Высокий, густой и мягкий мех сверху серый, нижняя половина и лапы белые. Хвост
покрыт короткими волосками, конец хвоста весь белый. Уши короткие, округлые,
глаза небольшие Имеет защечные мешки, которые открываются с внутренней стороны
губ и тянутся назад почти до плечей. Резцы без желобков и бороздок.
Распространена от Кении, Уганды, Танзании на юг до конца материка. Обитает в
редколесьях и светлых лесах, в травянистых и кустарниковых зарослях, на болотах
и на посевных землях. Ночной зверек. Весьма медлительный в движениях. Копает
индивидуальные норы с одним входом. На самом юге Африке, в более умеренных
широтах, устраивает запасы пищи, которые переносит в нору защечных мешках.
Питается зерном, семенами акаций, плодами и насекомыми. Размножение отмечено в
декабре (Юго-Западная Африка), в июле (Восточная Африка), в январе
(Малави). Во многих местах мясо короткохвостых крыс едят.
Хомяковая крыса (Cricetomys gambianus) также представитель
монотипического рода и характеризуется наличием защечных мешков. Распространена
эта крыса на большей части Африки южнее Сахары. Длина тела 25-45 см, хвоста -
36-45 см, масса около 1 кг. Короткий мех сверху коричневато-серый, низ белый
или кремовый. Конец чещуйчатого хвоста также белый. У отдельных особей могут
быть светлые пестрины по бокам. Обитают хомяковые крысы в самых разнообразных
местах, но предпочитают леса и кустарники. Нередко селятся рядом с полями и
постройками человека, а в некоторых городах Западной Африки весьма обычны в
системе канализации, где обитают вместе с черными крысами. Нередко занимают
старые термитники. Каждый зверек сам роет себе индивидуальную нору с
несколькими отнорками и входами. Чаще всего такое убежище устраивается у
основания ствола дерева или густого куста. Гнездовая камера выстилается сухой
травой и листьями. Размножение возможно в любое время года. После 42 дней беременности
самка рождает 2-3 детенышей, хотя имеет 8 сосков и может прокормить больший
выводок. Кормится в основном по ночам, поедает охотнее всего семена и плоды. В
поисках плодов крыса ловко влезает на деревья и передвигается в кронах,
используя свой длинный хвост как балансир. Продолжительность жизни в природе
обычно 3-5 лет. В неволе быстро привыкает к человеку и становится забавным
домашним животным. Туземцы во многих местах охотятся на Хомяковых крыс ради их
вкусного мяса.
2. Район, материал и методы исследования
2.1 Характеристика района исследования
Первая
площадка находилась на территории Аулиекольского района. Аулиекольский район
расположен на севере тургайской впадины в южной части соленого озера Кушмурун.
Территория района характеризуется континентальным климатом, проявляющемся в
резких сменах температур дня и ночи, зимы и лета; а также в быстрых переходах
от зимы к лету.
Зима
продолжительная, малоснежная и холодная, с сильными периодическими ветрами,
метелями. Ввиду маломощности снежного покрова (около 250 мм) почва сильно
промерзает (более 1,5м).
Весна отличается быстрым ростом тепла и высокой контрастностью дневных и
ночных температур, а также сильными ветрами, вызывающими быстрое иссушение
поверхности почвы.
Лето сухое и жаркое. Как и весной, летом довольно часты суховеи и засухи.
Жаркий период со среднесуточной температурой выше +200С длится не
более месяца.
Осень характеризуется постепенным похолоданием, первые заморозки
наступают в конце сентября.
Среднегодовое количество осадков для Аулиекольского района - 284мм, что
указывает на дефицит влаги. Поэтому, для выращивания сельскохозяйственных
культур в регионе, необходимо проводить агротехнические мероприятия,
направленные на накопление и сохранение влаги.
Аулиекольский район расположен вдоль Тургайской ложбины в центре
Тургайского прогиба. На севере и западе района расположена Северо-Тургайская
равнина, рельеф которой плоскоравнинный или слабоволнистый.
На юге Аулиекольского района расположено Терсекско-Адаевское плато,
которое имеет плоскую поверхность и ограничено с севера высоким уступом,
изрезанными балками. С востока к району примыкает Восточно-Тургайское плато,
которое представлено равниной, слабо расчлененной руслами мелких пересыхающих
речек, а также террасовидные уступы вдаются в Тургайскую ложбину и сильно
изрезаны глубокими балками. В районе поселка Аманкарагай в 5 км от центра
района расположен Аманкарагайский бор с огромным песчаным массивом. В
приборовой полосе на границе леса со степью расположены высоко бугристые пески.
В целом по району рельеф равнинных массивов способствует применению
сельскохозяйственной техники. Почвенный покров Аулиекольского района
разнообразен в преобладающем большинстве это темно-каштановые почвы с небольшим
процентом других почв. Чаще всего это примеси солончаков, так как реки и озера
здесь в основном соленые. В районе Аманкарагайского бора почвы
дерново-подзолистые по механическому составу песчаные. Примерно половина всех
земель составляют почвы легкосуглинистые. Очень небольшой процент занимают
черноземы южные карбонатные и южные карбонатные луговые.
На вышеперечисленных почвах пахотные угодья не целесообразно располагать.
Поэтому в Аулиекольском районе занимаются скотоводством и частично выращиванием
бахчевых культур в северо-восточной части района.
Так как район расположен в зоне засушливой степи, а почвенный покров в
основном не пригоден для выращивания зерновых культур, то большая часть
территории занята пастбищными угодьями. Распаханы не большие участки вокруг
поселков, озер, рек. Наиболее распространенными растительными комплексами
являются разнотравно-красноковыльные степи и разнотравно-песчаноковыльные степи
- темнокаштановых почвах, а на каштановых почвах и солонцах комплексами
являются типчаково-ковыльные степи с типчаково-полынными сообществами. Основу
травостоя образуют: ковыль волосатик, типчак, тонконог, полынь австрийская.
Использовать данные пастбища рекомендуется вечной, летом и осенью для выпаса
всех видов скота.
Животный мир представлен наиболее встречающими видами, такими как -
ящерица живородящая, жаба обыкновенная, воронок полевой, перепел, трясогузка
белая, каменка обыкновенная, воробей полевой, ласточка деревенская, коршун
черный, кобчик обыкновенный, лунь луговой, полевка серая, полевая мышь, суслик,
заяц беляк, лесная мышь, полевка обыкновенная, бурозубка обыкновенная.
Гордостью района является Аманкарагайский бор, состоящий из соснового и
березово-соснового леса на песках и слабо сформированных дерново-подзолистых
почвах.
Засушливый климат и равнинный рельеф Аулиекольского района определяет
слабое развитие речной сети. Основная река Убаган имеет снеговое питание. Для
нее характерны резкие колебания среднегодовых расходов воды в различные по
водности годы. До 80% - 90% годового стока приходится на весну (апрель - май).
Большинство озер летом пересыхают, и образуется множество плесовых блюдец, в
которых вода держится весь пастбищный период. Уровень воды поддерживается
родниками, атмосферными осадками и грунтовыми водами, которые находятся на
глубине 3 - 3,5 м.
Исследуемый нами участок 1 расположен в ольшанике. Древесный ярус
представлен ольхой, осиной и березой. Средний - это ива, черемуха, дикая вишня,
шиповник, ирга. Нижний - лопух, лапчатка, крапива, папоротник, а завершает все
ярус мхов и лишайников.
На почве всегда есть остатки растений, опавших листьев, сухие ветки,
образующие лесную подстилку, богато населенную микроорганизмами и грибами,
вызывающими минерализацию опада. Поэтому в ольшанике почва плодородная, рыхлая,
влажная. Наличие обильной стабильной кормовой базы делает фауну богатой и
разнообразной.
Вторая площадка располагалась в 15 км от г. Рудного. (Костанайский
район). Характерной чертой климата является резко выраженная континентальность:
жаркое и сухое лето сменяется холодной и малоснежной зимой. Годовая амплитуда
температуры воздуха в среднем составляет 75° в отдельные годы достигает 880
Зимой минимальная температура воздуха нередко падает до 400С, а
абсолютный минимум составляет 530С мороза. Абсолютная максимальная
температура равна 430С. Теплый период со средней суточной
температурой выше О0С длится 195-201 день: с 7-12 апреля до 19-28
октября. Продолжительность безморозного периода колеблется от 108 до 130 дней.
Средняя годовая температура воздуха 0,3-2,3 повышается в отдельные годы до
4,5-50С или снижается до О-1,20С.
Характерным признаком континентальности климата является существенное
преобладание осадков теплого периода (май - октябрь), когда выпадает 60-80% от
годовой нормы. Осадки теплого периода распределяются неравномерно. Весна и
начало лета отличаются малым количеством осадков, что в сочетании с большими
скоростями ветра (в среднем 5-6 м/с), наблюдающимися в это время, обусловливает
быстрое иссушение почвы. Максимум осадков на большой части территории
приходится на вторую половину лета, чаще всего на июль. Несмотря на это летом
преобладает ясная погода. Среднее число пасмурных дней в теплый период не
превышает 5-8 в месяц. Продолжительность дня летом составляет 16-17 часов.
Зима холодная, малоснежная и продолжительная (около 5 мес.), с сильными
ветрами (чаще юго-западными) и метелями. Средние скорости ветра в зимние месяцы
составляют 4,5-5,5 м/с. Снежный покров образуется с 9 по 26 ноября и лежит
устойчиво 132-157 дней. Но он неглубок (до 22 см), поэтому почва зимой
значительно промерзает (до 2 м.). Годовая сумма осадков - 290-350 мм, за период
вегетации выпадает 170-200 мм.
С глубинными разломами и древними интрузиями складчатого фундамента на
левобережье р. Тобола связана основная часть железорудного пояса. Наиболее
богатые месторождения магнетитовых железных руд - Соколовское, Сарбайское,
Качарское, Коржункульское. На базе двух первых создан мощный
горнообогатительный комбинат в городе Рудном [52].
Костанайскии район - засушливая степь, преимущественно с южными
малогумусными черноземами. Гумусный горизонт у них менее мощный (35-50 см),
перегноя содержит 4,0-5,5%. По агроклиматическим условиям входит в засушливый
теплый район. Костанайскии район относится к подзоне - разнотравно-ковыльная
(засушливая) степь на южных черноземах. В этой подзоне коренной растительный покров
представлен разнотравно-ковыльными степями и их вариантами. Основу
растительности слагают дерновинные злаки: красный ковыль, иногда ковыль
Лессинга, а также типчак, тонконог. Примесь разнотравья еще значительна, но ее
состав намного беднее, чем в более северных степях. Облик фауны определяют
всего несколько наиболее многочисленных (фоновых) видов. Среди птиц это
жаворонки (полевой, белокрылый и черный). Весьма характерны для степи степной и
луговой луни. Многочисленны мелкие сокола (кобчики, пустельги, чеглоки) в
лесных островах, расположенных на песчаных массивах и по склонам степных плато.
Среди млекопитающих в степях господствуют грызуны: сурки, большой суслик, малый
суслик, ушастый еж, степная пеструшка, полевки узкочерепная и обыкновенная,
хомяк обыкновенный. Широко распространены лесная мышь, хомячок Эверсманна,
большой тушканчик и слепушонка, тушканчик-емуранчик. Из четвероногих хищников
наиболее многочисленным в годы обилия мышевидных грызунов бывает небольшая
степная лисица - корсак. Есть также степной хорь, горностай, ласка, барсук,
волк.
Вторая площадка располагалась в степи рядом с отвалами ССГПО. С
северо-запада ее ограничивали отвалы, с северо-востока - лесопосадки. Возраст
лесополосы 6-7лет. Основной ее древесной породой был вяз высотой до 3-х метров.
Кое-где встречались клены, тополя, ивы. Трансекты располагались на участке с
растительностью, характерной для разнотравноковыльной степи. На площадке были
представлены следующие растения: клевер, козлобородник, укропник, козелец,
полынь, ковыль, тысячелистник, конопля, молочай, очиток. Бурьян был высотой до
1 м. По рельефу площадка представляет холмистую равнину. Основной тип почвы -
деградированные черноземы. Около 7 лет назад земля на площадке была вспахана.
Температурный режим и влажность были подвержены резким изменениям вследствие
интенсивного солнечного облучения и колебались в летние месяцы в широких
пределах. Основу населения грызунов на площадке составляли лесная мышь и
обыкновенная полевка [53,54].
2.2 Материалы
и методы исследования
Исследования проводились в мае-сентябре 2008 -2009г. Основной метод сбора
материала отлов мелких млекопитающих стандартными линиями давилок (ловушек
Геро). За указанный период отработано 1200 ловушко-суток. Пойманных зверьков
взвешивали, определяли пол, репродуктивное состояние, возраст согласно методики
Н.В. Тупиковой.
Таблица 1 Календарные даты и периоды отловов лесной мыши в 2008 г
Площадка1
|
Тур
|
Дата
|
|
1
|
11.05.-14.05.
|
|
2
|
10.06.-13.06.
|
|
3
|
12.07.-15.07.
|
|
4
|
11.08.-14.08.
|
|
5
|
07.09.-10.09.
|
Календарные даты и периоды отловов лесной мыши в 2008 г
Площадка2
|
ТурДата
|
|
|
1
|
10.05.-13.05.
|
|
2
|
09.06.-12.06.
|
|
3
|
11.07.-14.07.
|
|
4
|
12.08.-15.08.
|
|
5
|
05.09.-08.09.
|
Для характеристики обилия и пространственной структуры населения животных
использовали следующие показатели [55,56].
. Общее обилие (I),
выражаемое в числе отловленных особей на 100 л/с, рассчитывали обычным способом
по формуле: I = (c/d)*100, где c - число отловленных особей за 4
суток, d - число отработанных ловушко-суток (d=a*t, где a - общее число ловушек, t - число суток отлова, в данном
случае t=4). Этот показатель отражает
совокупное обилие животных на территории, включающей все типы микроучастков,
часть из которых непригодна для обитания вида и не заселена им. Общее обилие
выражает в некотором смысле емкость и пригодность среды обитания в целом для
населения вида.
. Частное обилие (А) - обилие вида на микроучастках, заселенных
животными, выражено числом отловленных особей на 100 л/с, рассчитано по
формуле: А = (с/f)*100, где с -
количество отловленных животных за 4 суток, f - число отобранных ловушко-суток в микроучастках (f = b*t, где b - частное число ловушек из общего
количества d, отлавливавших животных, отражает
число микроучастков территории, занятых зверьками, t - количество суток отлова). Этот показатель отражает обилие
животных на подходящих микроучастках территории, которые они реально заселяют.
. Заселенность территории (F)
отражает долю (%) от обследованной территории, заселенную животными.
Заселенность рассчитывается по формуле: F=b/a*100, где а - общее количество
ловушек, b - частное число ловушек,
отлавливавших животных. Этот показатель отражает совокупную емкость всей
территории для особей данного вида.
. В качестве меры агрегированности (Ag), или степени скученности населения мы используем индекс
Уитфорда [16], который в данном случае равен отношению частного обилия (А) к
заселенности территории (F): Ag=A/F.
Этот показатель принимает наименьшее значение при выраженном размещении
животных по территории и возрастает с увеличением мозаичности (когда небольшое
число ловушек попадает на микроучастки, плотно заселенные животными, а
оставшиеся - на территорию, не заселенную зверьками).
Для оценки доверительных интервалов, в пределах которых лежат
действительные значения параметров, мы вычисляли стандартные ошибки показателей
обилия и пространственной структуры.
Статистическая обработка проведена посредством прикладного
статистического пакета "Statistica". Достоверность различия средних
оценивалась с помощью t-критерия. Различия считались достоверными на уровне значимости
5% (р=0.05).
3. Объект исследования
В качестве объекта исследования мы остановились на мелких млекопитающих.
Выбор этот не случаен: среди всех наземных позвоночных именно мелкие
млекопитающие (преимущественно грызуны) наиболее многочисленны и доминируют в
любом ландшафте. С высокой численностью мелких млекопитающих связано и другое
не менее важное их качество как объекта исследования: доступность и массовость
материала, что обеспечивает объективность и достоверность полученных данных,
Наконец, небольшая продолжительность жизни, а следовательно, высокая скорость
обновления популяции, позволяют на сравнительно ограниченном отрезке времени
проследить специфику основных биологических процессов в популяции у ряда
поколений. Таким образом, данная группа млекопитающих отвечает основным
требованиям, предъявляемым к объектам, выбранным для решения задач
экологического мониторинга [57,58,59,60]. Мелкие млекопитающие, являясь важным
компонентом как естественных, так и нарушенных экосистем, а также сельскохозяйственных
ландшафтов широко используются в качестве модельных объектов в экологических
исследованиях, включая мониторинг среды обитания [19,61].
Мелкие млекопитающие играют большую роль в процессах биологического
круговорота. При этом как в продукционных, так и в деструкционных процессах они
выступают в качестве биокатализаторов, в существенной мере определяя
самоочистительные свойства экосистем [62,63].
Велико значение роющих видов мелких млекопитающих в почвообразовательном
процессе. В результате их деятельности стабилизируется минеральный запас
элементов и улучшается минеральное питание растений, что ведет к увеличению и
стабилизации продукции растений. Роль мелких млекопитающих в агроценозах по
сравнению с естественными экосистемами существенно дополняется тем, что они
являются одними из основных вредителей сельскохозяйственных культур.
Исходя из теоретических представлений экологии животных можно ожидать,
что для этой группы организмов наиболее важными эколого-популяционными
показателями, определяющими степень воздействия, а также качество окружающей
среды, в первую очередь, будут являться ценотические, демографические и
морфофизиологические [14,64].
В качестве объекта исследования нами была выбрана лесная мышь Apodemus sylvaticus Linnaeus (1758). Лесная мышь распространена в
лесной зоне большей части Евразии (включая Британские острова), кроме ее южной
части; водится также в Северной Африке. В СНГ обитает на всей территории
европейской части (кроме северных районов и большей части Эстонии), включая
Карпаты, Крым, Кавказ, Средний и Южный Урал и Зауралье. Южная граница ареала
лесной мыши в Казахстане проходит в 60 км южнее с. Фурманово (Западный
Казахстан), через с. Калмыкове (р. Урал), верховья Эмбы, Улутау, горы Бектауата
(Северное Прибалхашье), Аягуз и Алакольскую котловину. К северу от этой границы
лесная мышь распространена всюду, где имеются подходящие условия для ее
обитания. Она населяет также весь Восточный Казахстан к востоку от реки Иртыш.
Кроме того, обитает на Сауре, Монраке, Тарбагатае, Джунгарском Алатау, Северном
и Западном Тянь-Шане, а также Каратау, Чу-Илийских горах и в долинах всех рек
Семиречья, Во многих местообитаниях, особенно в среднегорье южных гор, является
доминирующим видом, и численность бывает значительной [54,65]. Длина тела лесной
мыши до 115, хвоста до 96 мм. Морда заостренная, уши и глаза сравнительно
крупные. Задняя ступня длинная (18-23 мм). Окраска мехового покрова тела
песчаная, буровато-охристая или коричневая без темной полосы по хребту. Брюшко
беловатое или белое, резко отграниченное от темных боков. На груди между
передними лапами иногда бывает небольшое охристое пятно. Сосков 6. Длина хвоста
составляет около 86% длины тела [66].
В Северном Казахстане (Костанайская обл.), как и в других районах
республики, в питании мыши главную роль играет семенной корм. Грызуны поедают
семена всех древесных и кустарниковых пород, составляющих лесные полосы и
естественные насаждения. У зверьков, отловленных в посевах проса, пшеницы и
кукурузы, содержимое желудков всегда состояло из семян названных культур. Из
диких растений грызун охотно поедает семена бодяка, василька, осота. К
предпочитаемым кормам можно также отнести орешки бобовника или горького
миндаля. На севере Казахстана лесная мышь в большом количестве поедает зеленые
части растений во все месяцы тёплого периода. Из ягод предпочитает клубнику и
плоды боярки, но весьма редко, даже в местах обильного урожая. Животный корм
грызуна представлен насекомыми. Влагу, необходимую для утоления жажды, зверёк
получает, поедая сочные корма [67].
Норы этой мыши в Северном Казахстане без выбросов земли и тропинок,
малозаметны, поэтому их довольно трудно обнаружить среди травы и кустов. Реже
они устраиваются на открытом месте. Так, одна из найденных нор находилась на
краю старого арыка, совершенно лишенного растительности. Ход был проложен по
засыпавшейся трещине и затем расширен в шарообразную камеру диаметром 15см.
Гнездо, находившееся в этой камере, было сооружено из лоскутов бумаги, листьев
и сухой травы, внутри была более мелкая подстилка из шерсти и тонких
размочаленных стеблей трав. Временные норы на посевах устроены тоже довольно
просто; короткий ход оканчивается небольшим расширением со скудной выстилкой.
Имеется камера для запасов; она таких же размеров, что и гнездовая. Входных
отверстий 1-3. Использует лесная мышь и чужие брошенные убежища.
Летом мыши на поверхности появляются перед закатом солнца и энергично
передвигаются между кустами в поисках корма. Через час после захода солнца
примерно одна треть зверьков ночного улова уже попадает в ловушки, то есть
период наибольшей активности наблюдается в первые 1,5-2ч после вечерних сумерек
и ночи, во второй половине ночи она заметно снижается [67].
По данным В.А. Борисенко (1970) на Севере Казахстана мышь зимой не
размножается. Начало размножения отмечено в первой декаде апреля; самки со
второй беременностью стали встречаться с 20-х чисел мая; в это время уже
ловились молодые первого выводка [53].
Перезимовавшие самки размножаются в течение всего лета и в начале осени,
хотя уже в августе и особенно в сентябре количество их заметно снижается. Таким
образом, размножение перезимовавших зверьков в Северном Казахстане длится 5-6
месяцев. Наиболее интенсивно оно протекает в мае, когда у большинства самок
беременность сочетается с лактацией.
Во второй половине лета наступление очередной беременности несколько
затягивается, и поэтому уже в июне встречаются временно яловые самки с
плацентарными пятнами от предыдущих беременностей. Но тем не менее перерыва в
размножении их в середине лета не наступает, хотя в жаркое и сухое лето оно
прекращается несколько раньше. В связи с этим число выводков у перезимовавших
зверьков равно 3-5. Особенно много выводков за сезон бывает во влажные годы.
Сеголетки становятся половозрелыми на втором месяце жизни. Молодые,
родившиеся в первой половине лета, участвуют в размножении, а родившиеся позже
в этот сезон уже не размножаются. Число детенышей в помете, судя по эмбрионам,
у лесной мыши в Северном Казахстане колеблется от 4 до 10. Средняя величина
выводка равна 6,5 детеныша, Размеры выводка в начале размножения меньше, чем
летом. К осени они повышаются. Эти мыши живут в одиночку и семьями, не образуя
колоний, Их норы располагаются более или менее равномерно по местообитанию.
В Костанайской области лесная мышь попадается в самых разнообразных
биотопах, но наиболее многочисленна там, где имеются деревья или кустарники.
Всюду в степной и сухостепной полосах она наиболее многочисленна в поймах рек,
колках и лесополосах. Из лесных биотопов менее всего заселены ею боры. На
посевах и залежах эта мышь встречается чаще, чем домовая, особенно вблизи
колков и лесных полос. Мыши известны как хранители возбудителя туляремии.
Многие авторы отмечали спонтанную зараженность мышей клещевым энцефалитом [68].
В Северном Казахстане у этих грызунов известен лептоспироз. В степных и
особенно в лесостепных районах лесная мышь становится заметным вредителем
посевов. Ее вред на полях проявляется не только в период созревания хлебов, но
и весной. Основные районы вредоносности в Казахстане - предгорная полоса на юге
и юго-востоке и северная часть целинных земель. Однако зверек приносит и
определенную пользу, уничтожая насекомых - вредителей сельскохозяйственных
культур и леса [69]. Наконец, лесная мышь-корм ценных пушных зверей.
4. Динамика численности и демографическая структура населения лесной мыши
Наиболее высокая численность лесной мыши на протяжении всего периода
исследований 2004 г. была отмечена на площадке 1 (19,5 экз. на 100л.с.), обилие
исследуемого вида на 2 площадке всегда оставалось на более низком уровне (табл.
2, 3). Средние за сезон размножения численности составляли соответственно 12,3
и 3,7 экз. на 100л.с.
Динамика численности лесной мыши на первой площадке характеризуется
возрастанием обилия с мая до августа более чем в два раза (табл. 3) с
последующим снижением к концу репродуктивного периода. Численность лесной мыши
на 2 площадке увеличивалась в течение сезона.
За период размножения численность лесной мыши на первой площадке
увеличилась в 2,8 раза, а на второй только в 1,8 раза.
Динамика возрастной структуры в рассматриваемых биотопах характеризуется
синхронным снижением численности перезимовавших животных и одновременным
увеличением доли прибылых. Однако на второй площадке вплоть до сентября среди
прибылых преобладают половозрелые особи, а общая доля участвующих в размножении
животных в среднем за сезон 2004 г. составляет 61,3 %. На первой площадке на
протяжении всего сезона размножения среди сеголеток преобладают неполовозрелые
зверьки, а общая доля участвующих в размножении особей в среднем за лето - 44
%.
Существенно различается и динамика половой структуры. В населении лесной
мыши на первой площадке в направлении от весны к осени наблюдаются колебания
вокруг средней за сезон доли самок - 55.4%. На второй площадке доля самок
всегда ниже, чем самцов (в среднем за сезон размножения составляет 46.8%).
Таким образом, более высокой репродуктивной активности лесной мыши на
второй площадке соответствуют малые численности и низкие скорости их прироста,
тогда как на первой площадке прослеживается обратная картина (при низкой доле
половозрелых животных, наблюдается быстрый и значительный рост численности)
Таблица 2 Демографическая структура лесной мыши на площадке 2
|
Тур
|
Общее обилие, экз.
на100л.с.
|
Доля неполовозрелых, %
|
Доля половозрелых, %
|
Доля половозрелых
перезимовавших, %
|
Доля самцов, %
|
Доля самок, %
|
|
1
|
2,8
|
7,0
|
10,3
|
82,7
|
52,0
|
|
2
|
3,0
|
22,0
|
30,0
|
48,0
|
57,0
|
43,0
|
|
3
|
3,7
|
40,5
|
54,5
|
5,0
|
52,0
|
48,0
|
|
4
|
4,0
|
44,5
|
50,9
|
4,6
|
53,0
|
47,0
|
|
5
|
5,0
|
79,0
|
21,0
|
0
|
56,0
|
44,0
|
Таблица 3
Демографическая структура лесной мыши на площадке 1
|
Тур
|
Общее обилие, экз.
на100л.с.
|
Доля неполовозрелых, %
|
Доля половозрелых, %
|
Доля половозрелых
перезимовавших, %
|
Доля самцов, %
|
Доля самок, %
|
|
1
|
7,0
|
11,3
|
2,5
|
86,2
|
32,0
|
68,0
|
|
2
|
11,0
|
29,0
|
17,0
|
54,0
|
41,0
|
59,0
|
|
3
|
14,0
|
67,0
|
22,0
|
11,0
|
51,0
|
49,0
|
|
4
|
19,5
|
82,0
|
10,3
|
7,7
|
50,0
|
50,0
|
|
5
|
10,2
|
90,6
|
9,4
|
0
|
49,0
|
5. Пространственная структура населения лесной мыши
Характерной особенностью пространственной структуры лесной мыши на первой
площадке является постепенное увеличение доли используемой территории до
августа, наряду с ростом общей численности населения (табл.4). В результате к
достижению сезонного пика численности доля используемой территории составила 23
%.
С мая до июня у лесной мыши на первой площадке отмечалось увеличение
частного обилия и степени агрегированности при незначительном снижении доли
используемой территории (рис.1,2). В июле общая численность значительно
возросла это сопровождалось увеличением как доли используемой территории, так и
плотности на занятых микроучастках, которая достигла в это время максимальных
значений.
В августе численность лесной мыши на первой площадке достигла сезонного
пика при увеличении доли используемой территории. В последующий период, к
осени, отмечалось снижение общего обилия, которое произошло в результате
естественного отхода перезимовавших и прибылых животных, снижения
репродуктивной активности половозрелой части населения и ухода с площадки части
сеголеток, условия обитания для которых, по-видимому, ухудшились вследствие
перенаселения территории. Все это привело к уменьшению частного обилия и доли
заселенной территории. Можно отметить более равномерное заселение территории
площадки в сентябре.
Численность лесной мыши на второй площадке находилась на более низком
уровне по сравнению с первой (табл. 2).
С мая до июня отмечалось снижение численности половозрелых особей лесной
мыши а так же доли используемой территории (табл 4.). В следующий
период, к июлю, наблюдалось увеличение численности, уровня частного обилия и
доли используемой территории лесной мышью на второй площадке (рис.1, табл 4.)
на фоне снижения агрегированности животных (рис.2). Рост частного обилия
продолжался до августа, достигнув наибольшего значения за сезон. Рост частного
обилия к августу вызывает увеличение степени агрегированности. В последующий
период, к осени, отмечалось снижение общего обилия, которое произошло в
результате естественного отхода перезимовавших и прибылых животных, снижения
репродуктивной активности половозрелой части населения и ухода с площадки части
сеголеток, условия обитания для которых, по-видимому, ухудшились вследствие
перенаселения территории. Все это привело к уменьшению частного обилия и доли
заселенной территории. Можно отметить более равномерное заселение территории
площадки в сентябре.
К сентябрю на второй площадке наблюдалось уменьшение плотности населения
на занятых участках за счет увеличения размеров используемой территории, в
результате чего индекс агрегированности снизился (рисунок 2).
Таблица 4
Доля используемой территории населением лесной мыши
|
Тур
|
Доля используемой
территории, %
|
|
Площадка 1
|
Площадка 2
|
|
1
|
12
|
6
|
|
2
|
9
|
3
|
|
3
|
17
|
9
|
|
4
|
23
|
8
|
|
5
|
19
|
10
|
Частное
обилие лесной мыши на площадках 1, 2 рисунок 1
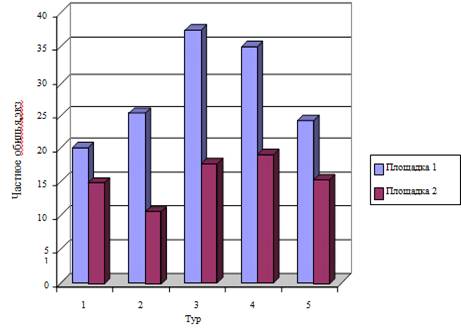
Агрегированность населения лесной мыши на площадках 1 и 2 рисунок 2
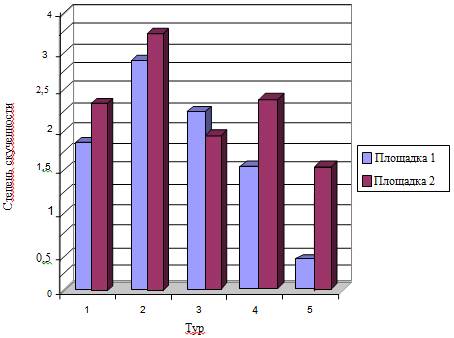
Заключение
В результате антропогенных воздействий каждый вид, как и отдельная
популяция, приспосабливается к новым, измененным условиям обитания. Происходит
этот процесс различными путями. Ведущим фактором, позволяющим зверькам заселять
местообитания антропогенного происхождения, является пластичность основных
экологических параметров. В условиях антропогенизации у многих видов отмечено
изменение пространственно-этологических структур, следствием чего является
возможность существования на территории, измененной хозяйственной деятельностью
[12, 56].
Популяционно-экологические и демографические показатели всесторонне характеризуют
жизненный цикл и экологическую стратегию видовых популяций. Эти показатели
являются интегральными, отражающими, с одной стороны, степень
сбалансированности процессов рождаемости, смертности, миграции в популяциях, а
с другой - указывающими на емкость и пригодность местообитаний для данного вида
организмов [14,33,61,64,70].
Исследование по влиянию антропогенных факторов свидетельствует об
изменении основных популяционных характеристик вида. Антропогенные факторы
через разрушение основных местообитаний лесной мыши приводят к существенному
снижению общего обилия вида, а также экологического обилия животных в
осваиваемых микростациях. В наибольшей степени снижается заселенность животными
территории, что связано с сокращением числа микростаций. Опережающее снижение
числа микроучастков по сравнению с их емкостью приводят к повышенной
агрегированности населения. Использование пространства в значительной степени
сокращено по сравнению с тем, как оно используется в естественном биотопе.
Зверьки, поселяющиеся в местообитаниях, нарушенных в результате антропогенного
вмешательства, образуют локальные, относительно изолированные поселения.
Подобные изменения пространственной организации населения мелких млекопитающих
являются прямым следствием перестроек в структуре природных биоценозов в
результате вмешательства человека.
Антропогенные воздействия какого то ни было типа могут приводить к
различным изменения структуры популяций грызунов.
Появление лесной мши вблизи отвалов, лимитируется скудной кормовой базой,
ограниченным числом укрытий и других, необходимых для обитания условий.
На популяционном уровне такой процесс приводит к сдвигу соотношения
полов, в сторону самцов, усилению интенсивности размножения (как компенсаторной
реакции на снижение плотности) и, как следствие последнего, к изменению
возрастной структуры в направлении увеличения доли сеголеток. Интенсификация
воспроизводства достигается значительным увеличением доли половозрелых
сеголеток в популяции.
На основании комплексного изучения сезонных изменений демографической
структуры и пространственного распределения лесной мыши, обитающей в различных
биотопических условиях можно сделать следующие выводы:
. На территории, измененной хозяйственной деятельностью отмечается
существенное снижение как общего обилия вида, так и частного обилия.
. В результате антропогенных воздействий значительно снижается
заселенность животными территории и увеличивается агрегированность.
. В биотопе, измененном хозяйственной деятельностью отмечается тенденция
к увеличению доли самцов, что свидетельствует о неблагоприятной ситуации.
. Антропогенные воздействия приводят к изменению возрастной структуры
населения в направлении увеличения доли сеголеток, что подтверждает усиление
интенсивности размножения.
. Принимая как аксиому тот факт, что динамика численности наряду с
популяционными механизмами саморегуляции зависит от условий окружающей среды,
мы можем рассматривать особенности ее в качестве критерия оценки этой среды,
подверженной трансформациям под действием антропогенных факторов.
Список использованных источников
1. Hestbeck J.B. Population regulation of cyclic
mammals: The social fense hypothesis // Oikos. 1982, Vol. 39, p. 157-163.
. Lidicker W.Z.(Jr.). The role of dispersal in the
demography of small mammals // Small Mammals: Their Production and Population
Dynamic. London, 1975. P. 103-128.
. Whitford P.B. Distribution of woodland plants in
relation to succession and clonal growth //Ecology. 1949. Vol. 30. P.
199-208.
. Баженов А.В., Куликова И.Л., Садыков О.Ф. Радионуклидные
методы группового мечения // Методы исследования пространственной структуры
популяций мелких млекопитающих в естественной среде и агроценозах. Свердловск,
1983. С. 25-38.
. Башенина Н.В. Пути адаптации мышевидных грызунов. М.:
Наука, 1977. 355 с.
. Бекенов А. Млекопитающие пустынь Казахстана (Фауна,
биология, рациональное использование и охрана)//Дис.в виде научного доклада на
соискание уч.степени д.б.н. Свердловск, АН СССР ИЭРиЖ, 1991.
. Беклемишев В.Н. Пространственная и функциональная структура
популяций // Бюлл. МОИП, отд. биол., 1960, вып. 2, с. 5-6.
. Бердюгин К.И., Дороватовский С.А. Вертикальное
распределение грызунов в горах Урала // Млекопитающие Уральских Гор.
Свердловск, 1979. С. 4-5.
. Большаков В.Н. Высотное распределение мелких млекопитающих
Уральских Гор и отличия биопродуктивности // 3 Всесоюз. Совещ. "Вид и его
продуктивность в ареале" (Паланга, 18-19 сент. 1980 г). Вильнюс, 1980. С.
117-118.
. Большаков В.Н. Пути приспособления мелких млекопитающих к
горным условиям. М.: Наука, 1972. 200 с.
. Большаков В.Н., Баженов А.В. Радионуклидные методы мечения
в популяционной экологии млекопитающих. М.: Наука, 1988. 158 с.
. Большаков В.Н., Баженов А.В. Радионуклидные методы мечения
в популяционной экологии млекопитающих. М.: Наука, 1988. 158 с.
. Большаков В.Н., Балахонов В.С. Мелкие грызуны полярного
Урала и аналогичных ландшафтных зон // Грызуны: Материалы 5 Всесоюз. Совещ.
(Саратов, 3-5 дек. 1980 г). М., 1980. С. 162-163.
. Борисенко В.А. Влияние распашки целинных земель на
распространение и численность мышей и полевок в Кустанайской области / Автореф.
канд. дис. Алма-Ата. 1970, с. 2-18.
. Гашев С.Н. Влияние антропогенных факторов на устойчивость
сообществ наземных позвоночных животных среднетаежной зоны Западной Сибири. В
сб.: Проблемы взаимодействия человека и природной среды.Тюмень: ИПОС СО РАН,
2000 б, с. 122-116.
. Гашев С.Н. Млекопитающие в системе экологического
мониторинга (на примере Тюменской области). Тюмень:ТюмГУ, 2000 а, 200 с.
. Гашев С.Н. Млекопитающие в системе экологического мониторинга
(на примере Тюменской области)//Автореф. дис. на соискание уч.степени д.б.н.
Тюмень, 2001.
. Гвоздецкий Н.А., Николаев В.А. Казахстан. Очерк природы.
М.: Мысль, 1971, с. 145-149.
. Громов И.М. и др. Млекопитающие фауны СССР. М.-Л., 1963,
ч.2, с. 79.
. Груздев В.В. Территориальные центры и районы благоденствия
и прозябания популяций // Популяционная структура вида у млекопитающих. М.,
1970. С. 7-10.
. Демидов В.В., Хлебалов И.Я. Наблюдения за использованием
территории мелкими грызунами в южной тайге Камского Приуралья. Перм. гос. ун-т.
Пермь, 1981. 18 с. Деп. в ВИНИТИ 20.04.81, N1737-81 Деп.
. Жигальский О.А. Механизмы динамики популяций мелких
млекопитающих: Автореф. дис. д-ра биол. наук. Свердловск, 1989. 49 с.
. Жигальский О.А., Бернштейн А.Д. Популяционные факторы
регуляции размножения рыжей полевки (Clethrionomys glareolus)// Докл. АН СССР. 1986. Т. 291, N1. С. 250-252.
. Кошкина Т.В., Коротков Ю.С. Популяционная регуляция
численности красной полевки в оптимуме ареала // Фауна и экология грызунов. М.,
1975. Вып. 11. С. 5-61.
. Крылов Д.Г. Использование территории Clethrionomys glareolus, Schreber (Rodentia) в условиях средней полосы
Европейской части СССР//I
Междунар. териол. конгр. М., 1974. Реф. докл. Т.1. С. 317.
. Кутенков А.П. Об особенностях использования территории
рыжей полевкой в заповеднике "Кивач"// Фауна и экология птиц и
млекопитающих таежного Северо-Запада СССР. Петрозаводск, 1978. С. 120-127.
. Литвинов Ю.Н. Сообщества и популяции мелких млекопитающих в
экосистемах Сибири.-Новосибирск: ЦЭРИС, 2001, 128 с.
. Лукьянов О.А. Феноменология и анализ миграций в популяциях
мелких млекопитающих: Автореф.дис.док. биол. наук. Екатеринбург, 1997. 46 с.
.Лукьянова Л.Е., Лукьянов О.А. Характеристика обилия и
пространственной структуры населения рыжей полевки на техногенных территориях
//Животные в условиях антропогенного ландшафта. Екатеринбург, 1992, с. 85-95.
. Макфедьен Э. Экология животных (цели и методы). М.: Миир,
1965, 375 с.
. Мариковский П.И. Влияние мышевидных грызунов на численность
лунки серебристой // Изд. АН КазССР, серия биол., 1970, №1, с. 5-15.
. Марин Ю.Ф. Абсолютная численность красно-серой и красной
полевок и использование ими территории в среднегорье Северо-Восточного Алтая //
Изв.Сиб. Отд. АН СССР. Сер. биол. Наук.1976, т.15, вып. 3, с. 80-85.
. Млекопитающие Свердловской области. Справочник-определитель
/В.Н. Большаков, К.Н. Бердюгин, И.А, Васильева, И.А. Кузнецова.- Екатеринбург,
Изд-во "Екатеринбург", 2000, 240 с.
. Наумов Н.П. Изучение подвижности и численности мелких
млекопитающих с помощью ловчих канавок // Вопросы краевой, общей и
экспериментальной паразитологии и медицинской зоологии. М., 1955. Т. 9. С.
179-202.
популяция
млекопитающий мышь антропогенный
. Наумов Н.П. Мечение млекопитающих и изучение их
внутривидовых связей // Зоол. журн. 1956. Т. 35, вып. 1. С. 3-15.
. Наумов Н.П. Новый метод изучения экологии мелких лесных грызунов
// Фауна и экология грызунов. М., 1951, Вып. 4, с. 3-21.
. Наумов Н.П. Пространственная структура вида мелких
млекопитающих // Зоол. журн. 1971, т.50, вып. 7, с. 965-980.
. Наумов Н.П. Структура популяций и динамика численности
наземных позвоночных // Зоол. журн. 1967. Т. 46, вып. 10. С. 1470-1486.
. Наумов Н.П. Экология животных. М.: Высш. шк., 1963. 618 с.
. Никитина Н. А., Меркова М. А. Использование территории
мышами и полевками по данным мечения //Бюл. МОИП. Отд. биол. 1963. Т. 68,
вып.5. С.18-36.
. Никитина Н.А. К понятию "индивидуальный участок"
у мелких млекопитающих //I Междунар. териол. конгр.: Реф. докл. М., 1974. Т. 2.
С. 71-72.
. Никитина Н.А. О размерах индивидуальных участков грызунов
фауны СССР //Зоол. журн. 1972. Т. 51, вып. 1. С. 119-126.
. Никитина Н.А. Рыжие полевки // Итоги мечения млекопитающих.
М., 1980. С. 189-219.
. Никитина Н.А., Карулин Б.Е., Литвин Ю.В. и др. О размерах
суточной территории и вероятном строении индивидуальных участков у некоторых
видов грызунов // Зоол. журн. 1977. Т. 56, вып. 12. С. 1860-1869.
. Одум Ю. Экология: Пер. с англ. М.: Мир, 1986. Т. 2. 376 с.
. Окулова Н.М., Аристова В.А., Кошкина Т.В. Влияние плотности
популяции на размер индивидуальных участков у мелких грызунов в тайге Западной
Сибири и на Дальнем Востоке // Тез. докл. 3 Всесоюз. териолог. съезда. М.,
1982. Т. 1. С. 260-265.
. Окулова Н.М. Биологические взаимосвязи в лесных
экосистемах: (На прим. природ. очагов клещевого энцефалита). М.: Наука, 1986.
248 с.
. Павловский Е.Н. и др. Клещевой энцефалит //
Природно-очаговые болезни человека. М., 1960, с.67-69.
. Садыков О.Ф. Значение исследований пространственной
структуры популяций // Методы исследования пространственной структуры популяций
мелких млекопитающих в естественной среде и агроценозах. Свердловск, 1983.
с.3-8.
. Садыков О.Ф. О зимних стациях переживания мышевидных
грызунов в горах Южного Урала // Фауна, экология и изменчивость животных.
Информ. Мат-лы зоол. музея Ин-та экологии растений и животных УНЦ АН СССР.
Свердловск, 1978, с.8-9.
. Садыков О.Ф. Пространственная структура горных популяций
полевок рода Clethrionomys // Популяционная экология и
морфология млекопитающих. Свердловск, 1984. С. 20-36.
. Садыков О.Ф., Бененсон И.Е. Динамика численности мелких
млекопитающих: концепции, гипотезы, модели. М.: Наука, 1992. 190 с.
. Садыков О.Ф., Большаков В.Н., Баженов А.В. Пространственная
структура горных популяций лесных полевок // Экология. Свердловск, 1984, №4, с.
58-64.
. Самойлова Н.М. Сравнительный анализ сообществ позвоночных
сосняков городского лесопарка и заповедника//Автореф.дис. на соискиние уч.
Степени к.б.н., Екатеринбург, 2003
. Слудский А.А. и др. Млекопитающие Казахстана. В 4-х томах.
Алма-Ата, "Наука" КазССР, 1978, т.1, ч. 3, с.383-407.
. Слудский А.А. и др. Млекопитающие Казахстана. В 4-х томах.
Т.I, ч. 2. Алма-Ата, "Наука"
КазССР, 1977, с.431-445.
. Смуров А.В. Новый тип статистического пространственного
распределения и его применение в экологических исследованиях // Зоол. журн.
1975а. Т. 54, вып. 2. С. 283-289.
. Смуров А.В. Статистические методы в исследовании
пространственного размещения организмов // Методы почвенно-зоологических
исследований. М., 1975б. С. 217-240.
. Смуров А.В., Полищук Л.В. Количественные методы оценки
основных популяционных показателей: Стат. и динам. аспекты. М.: Изд-во МГУ,
1989. 208 с.
. Флинт В.Е. Пространственная структура популяций мелких
млекопитающих. М.: Наука, 1977. 183 с.
. Флинт. В.Е. Типы пространственной структуры популяций у
мелких млекопитающих. М.: МГУ, 1970, с. 10-15.
. Хляп Л.А. Основные проблемы изучения связей мелких
млекопитающих с окружающим пространством // Фауна и экология грызунов. М.,
1980. Вып. 14. С. 44-62.
. Шварц С.С. Популяционная структура вида // Зоол. журн.
1967. Т. 46, вып. 10. С. 1456-1469.
. Шварц С.С. Эволюционная экология животных. Свердловск,
1969,200 с.(Тр. Ин-та экологии растений и животных УФАН СССР; Вып. 65.)
. Шварц С.С. Экологические закономерности эволюции. М.:
Наука, 1980. 277 с
. Шилов И. Эколого-физиологические основы популяционных
отношений животных. М.: Изд-во МГУ, 1977, 262 с.
. Шилов И.А. Принципы организации популяций у животных //
Популяционные проблемы в биогеоценологии. М., 1988. С.5-23.
. Шилов И.А. Экология. М.: Высш.шк., 1997, с. 245-249.
. Шилов И.А., Калецкая М.П., Ивашкина И.Н., Солдатова А.Н.
Пространственная структура популяции полевок-экономок (Microtus oeconomus) в
Дарвиновском заповеднике // Зоол. журн. 1977. Т. 56, вып.2. С.257-267.