Сравнительный анализ компонентов растительного покрова
Содержание
экология
анализ растительный покров
Введение
Глава 1.
Схема анализа данных: основные методы
.1
Историко-методологические аспекты сравнительного анализа в экологии
.2 Выбор
элементарного описания
.3
Составление серии описаний
.4 Меры
разнообразия и близости, используемые в сравнительном анализе компонентов
растительного покрова
.4.1 Унарные
индексы
.4.1.1
Коэффициент вариации
.4.1.2
Параметрические семейства унарных мер
.4.1.3 Другие
индексы для оценки разнообразия
.4.2 Бинарные
индексы
.4.2.1 Множественная
интерпретация
.4.2.2
Дескриптивная интерпретация
.4.2.3
Вероятностная интерпретация
.4.2.4
Информационная интерпретация
.4.3
Определение n-арных отношений
.4.3.1
Множественная интерпретация
.4.3.2
Дескриптивная интерпретация
.4.3.3
Вероятностная интерпретация
.4.3.4
Информационная интерпретация
.4.4 Другие
интерпретации мер конвергенций и их применение
.4.5 Меры
близости, не удовлетворяющие системе аксиом
.4.6 Матрица
абсолютных мер конвергенции
.4.7 Матрица
относительных несимметричных мер конвергенции
.4.8 Матрица
относительных симметричных мер конвергенции
.4.9
Алгоритмы упорядочивания данных
.4.9.1
Кластерный анализ
.4.9.2
Диаграмма Чекановского
.4.9.3
Неориентированные графы
.4.9.4
Ориентированные графы
.4.10
Интерпретация результатов
Глава 2.
Сравнительный анализ компонентов растительного покрова
.1 Флоры
водорослей-макрофитов дальневосточного побережья России
.2
Субассоциации лесов поймы реки Амур
.3 Временнáя динамика сообществ
водорослей-макрофитов
Глава 3.
Биоинформационная система «Биоразнообразие»
.1 Базы
данных, содержащие компоненты биоразнообразия
.2 Программы
для обработки биологических данных
.3
Информационно-справочная система по биоценометрии с комментариями
Заключение
Список
литературы
ПРИЛОЖЕНИЯ
Приложение А.
Математические основания теории мер близости
А.1
Аксиоматический подход к введению мер. Теория S-функций
А.1.1
Дивергенция и конвергенция
А.1.2
Метрические относительные симметричные субаддитивные функции
А.1.3
Семейство относительных мер конвергенций
А.1.4
S-функции многих переменных
А.1.5
Дивергенция и конвергенция многих переменных
А.2
Интерпретации мер конвергенции и дивергенции
А.2.1
Интерпретации направленных мер конвергенции и дивергенци
А.2.1.1 Меры
включения и невключения двух множеств
А.2.1.2 Меры
включения и невключения двух дескриптивных множеств (наборов)
А.2.1.3 Меры
односторонней совместимости и несовместимости событий
А.2.1.4 Меры
односторонней зависимости и независимости случайных величин
А.2.2
Интерпретации симметричных мер конвергенции и дивергенции
А.2.2.1 Меры
сходства и различия двух множеств
А.2.2.2 Мера
сходства и различия двух дескриптивных множеств
А.2.2.3 Мера
совместимости и несовместимости двух событий
А.2.2.4 Меры
взаимной зависимости и взаимной независимости двух случайных величин
Приложение Б.
Примеры сравнительного анализа
Приложение В.
Биоинформационная система «Биоразнообразие»
Введение
Актуальность темы. Начиная с XX
века (особенно его второй половины), в рамках многих научных направлений, таких
как сравнительная флористика, геоботаника, экология и многих других можно
отметить устойчивую тенденцию активного внедрения математических методов
сравнительного анализа количественных показателей (Oosting, 1953; Greig-Smith, 1957; 1964; 1983; Savage, 1960;
Sokal, Sneath, 1963; Kershaw, 1964; Lambert, Dale, 1964; Williams, Dale, 1965; Грейг-Смит, 1967; Roux, Roux,
1967; Peters, 1968; Александрова, 1969;
Василевич, 1969; Cancella
da Fonseca, 1969; Cheetham, Hazе1, 1969; Pielou, 1969; 1975; 1977; Сёмкин, 1973 а,
б; 1975; 1977; 1987 а, б; Терентьев, 1975; Goodall, 1973; Sneath, Sokal, 1973; Шмидт, 1980; Юрцев,
Сёмкин, 1980; Песенко, 1982; Legendre, Legendre, 1983; Kershaw, Looney,
1985; Digby, Kempton, 1987; Magurran, 1988; 2004; Миркин, Розенберг, Наумова, 1989; Мэгарран, 1992; Уланова,
1995; Юрцев и др., 1999; Малышев, 1987; 1999; Chao et al., 2000; Koleff,
Gaston, Lennon, 2003; Шитиков, Розенберг, Зинченко, 2005; Diserud,
Ødegaard, 2007 и др.), компьютерных методов
обработки биологической информации, создания тематических биоинформационных
систем и баз данных по компонентам биоразнообразия (Сёмкин, 1994; Сёмкин и др.,
1995; Сёмкин, Варченко, Орешко, 1995; Сёмкин и др., 1997; Компьютерные…, 1995;
1997; Пяк, Зверев, 1997; Зверев, 1998; Марина, Марин, 1998; Юрцев, 1998;
Малышев, 2002; Крестов, 2005 и др.).
Первые попытки формализации таксономического состава сообществ (флор,
фаун и др.) множествами и определение с их помощью отношений (общности,
сходства, различия, включения, невключения), отдельных фракций видов (общие
виды двух флор (фаун), дифференциальных видов одной флоры (фауны) относительно
другой, дифференциальных видов двух флор (фаун) и т.д.) были осуществлены ещё в
70-х годах прошлого века (Тамарин, Шмидт, 1975; Сёмкин, 1977; 1983 а, б; Юрцев,
Сёмкин, 1980; Юрцев, 1982; 1987; Юрцев, Камелин, 1987 а, б; 1991). Эта проблема
имеет большое значение для разработки количественных показателей необходимых
для сравнительного анализа таксономических (главным образом видовых) списков, а
также даёт возможность строгого обоснования биологических наук в целом за счёт
строгой формализации связей системы. Любая система состоит из множества
элементов, объединённых соответствующими связями, которые в совокупности дают
структуру системы. В экологии и других науках в настоящее время отсутствуют
адекватные методики исследования структуры биологических систем, которые бы
учитывали особенности естественного континуума, требования теории измерений,
включали бы полный цикл исследования (от постановки цели исследования до
интерпретации результатов) и др. Многие количественные и качественные
показатели и методы используются специалистами вне общих методик и методологии.
В данной диссертационной работе нами приведены методология и методика
сравнительного анализа биологических данных, учитывающие вышеперечисленные и
многие другие проблемы математизации биологических наук.
Под растительным покровом нами понимается «совокупность особей зеленых
автотрофных растений, населяющих сушу и акваторию нашей планеты» (Галанин,
1991). Внимание акцентировано, главным образом, на компонентах видового
разнообразия растительных сообществ (флор, фитоценозов и др.). Отметим, что
рассмотренные математические методы универсальны и могут успешно использоваться
для сравнения и других биологических объектов (геномов, особей, экосистем,
таксонов), т.к. основаны на теории измерений и формализации мер близости в
биологии.
Современный уровень развития информационных технологий позволяет
реализовать практически любой алгоритм, представленный с помощью математических
методов. Сотрудничество биологии и математических методов в своё время породило
биометрию, биоматематику, биоценометрию и другие аналогичные направления
(Трасс, 1971 : 208), а на современном этапе - биоинформатику. У этого
научного направления довольно широкая область применения, включающая математическое
и компьютерное моделирование живых систем, включая экосистемы, и эволюционных
процессов в живой природе, распознавание и синтез изображений в биологических и
медицинских исследованиях; развитие бионических подходов; создание
информационных систем и баз биологических данных; интеллектуальные системы
анализа и прогнозирования свойств биологических объектов и др.
Цель исследования. Цель данного исследования - развитие общего подхода в
исследованиях структурной организации растительного покрова с использованием
биоинформационных технологий.
Задачи исследования:
1. Оптимизировать методику схемно-целевого подхода к сравнению
компонентов растительного покрова и адаптировать её для программной реализации
посредством создания биоинформационной системы «Биоразнообразие»;
2. На основе рассматриваемой методики получить новые результаты
ботанического районирования дальневосточных альгофлор, выделения оригинальных
флор, оценки антропогенного воздействия на флору, количественно подтвердить
характер распределения субассоциаций вдоль градиента увлажненности почв,
провести анализ временнóй структуры таксоцена.
Защищаемые положения:
1. Решена проблема выбора показателя для сравнения различных
биологических спектров (нормированных дескриптивных наборов), разновеликих и
равновеликих по числу признаков биологических объектов (ненормированных
дескриптивных наборов);
2. Схемно-целевой подход позволяет обеспечить более точные
интерпретации при исследовании структуры сообщества или отдельных таксоценов.
Научная новизна. Предложена современная методика сравнительного анализа
видового состава компонентов растительного покрова в рамках схемно-целевого
подхода: от постановки цели до интерпретации. На основе аксиоматического
подхода и формализации отношений между объектами в виде аналитических
интерпретаций осуществлена ревизия существующих мер близости, которые
используются в биологических и смежных науках, описаны основные классы мер,
которые позволяют упорядочить большинство известных мер близости и на этой
основе решена проблема выбора меры близости для нормированных, разновеликих и
равновеликих дескриптивных множеств. Алгоритм метода реализован в виде
компьютерной программы.
На основе схемно-целевого подхода пересмотрены результаты
альгофлористического районирования и выявлены центры видового разнообразия
водорослей-макрофитов ДВ РФ, приведен метод непрямой ординации на основе n-арных
мер для определения однородности условий увлажнения для серии описаний,
рассчитана теоретико-графовая модель внутригодовой динамики таксоцена водорослей-макрофитов.
Практическая значимость работы. Результаты исследования могут быть
применены в мероприятиях по сохранению биологического разнообразия растительных
сообществ, для количественной оценки данных биологического мониторинга, оценки
изменения видовой структуры сообщества при антропогенном воздействии, при
геоботаническом районировании, для непрямой ординации описаний, для
математико-картографического моделирования.
Рассматриваемые в работе методы были частично включены в учебные курсы
«Экологический мониторинг», «Геоинформационные системы», «Статистическая
обработка экологических данных», «Экосистемный анализ» для студентов
специальностей «Экология» и «Природопользование».
Апробация работы. Основные положения и результаты работы обсуждались: на сессии
ТИГ ДВО РАН (г. Владивосток, 2005 г.), на II-ом Молодежном конкурсе инновационных проектов по
Дальневосточному федеральному округу (г. Владивосток, 2007 г.), на III экологическом форуме «Природа без
границ» (г. Владивосток, 2008 г.), на научной конференции «Природопользование и
охрана окружающей среды» (г. Пекин, 2008 г.), на Комаровских чтениях (г.
Владивосток, 2008 г.).
Публикации. По теме диссертации опубликовано 14 печатных работ, в том
числе 4 в изданиях из списка ВАК.
Личный вклад. Оптимизирована методика сравнительного анализа видового
состава компонентов растительного покрова. Решена проблема выбора меры близости
для нормированных, разновеликих и равновеликих дескриптивных множеств.
При личном участии автора создана биоинформационная система «Биоразнообразие»,
которая реализуется на конкретном биологическом материале. Автором лично
создана программа для сравнительного анализа биологических данных
«BioComparison». Все расчёты и графическое представление в работе произведены
лично автором.
Структура и объём работы. Диссертация состоит из введения, 3 глав, заключения,
библиографического списка источников (включает 581 источник, 174 на иностранных
языках) и приложения. Работа изложена на 219 стр., содержит 161 стр. основного
текста, 58 стр. приложений.
Объекты, материалы и методы исследования. Наше внимание акцентировано,
главным образом, на компонентах видового разнообразия растительных сообществ
(флор, фитоценозов). Взят материал по видовому составу ДВ альгофлор (Клочкова,
1997) и прирусловых лесов р. Амур (Ахтямов, 2001), сезонному распределению
водорослей-макрофитов (Макиенко, 1975; Гусарова, 1988; Кафанов, Жуков, 1993).
Рассмотрены математические методы сравнительного анализа на основе общего
аксиоматического подхода и теоретико-графовых методов, а также вопросы
реализации данных методов с помощью информационных технологий.
Благодарности. Автор благодарит за всестороннюю поддержку и чуткое
руководство своего учителя - Б.И. Сёмкина. Автор выражает благодарность И.С.
Гусаровой, А.П. Орешко, Л.И. Варченко за консультативную помощь в сложных
вопросах, а также д.б.н., профессору Н.Г. Клочковой за предоставленный материал
для математической обработки.
Глава 1. Схема анализа данных: основные методы
1.1 Историко-методологические аспекты
сравнительного анализа в экологии
С начала XX века в
биологические науки активно проникают количественные математические методы
(Александрова, 1969; Применение…, 1972; Трасс, 1976). В рамках настоящего
исследования обратим внимание, главным образом, на внедрение количественных
методов в тех научных направлениях, которые связаны с исследованием
закономерностей в структуре растительных сообществ.
Несмотря на то, что исследование структуры биологических объектов - одна
из центральных проблем многих биологических наук, таких как экология,
геоботаника, фитоценология и др., о создании общей содержательной концепции
самого биологического исследования до сих пор идут споры. Когда дело касается
количественных оценок, привлечение математических приемов неизбежно, особенно
если исследователь работает в некой новой междисциплинарной области, особенно в
случае изучения структуры таких сложных объектов природы, как сообщества.
П. Грейг-Смит в своей известнейшей работе «Количественная экология
растений» (Грейг-Смит, 1967 : 13) писал: «Переход от качественного к
количественному подходу характерен для развития любой отрасли науки. Когда
достигается определённая степень понимания явлений в общих чертах, внимание
исследователей естественно сосредотачивается на более тонких деталях структуры
или поведения, наблюдаемые различия которых менее значительны и могут быть
оценены лишь с помощью измерения. Не удивительно, что количественный подход
развился раньше в большинстве отраслей наук о неживой природе, чем в
биологической науке. По-видимому, одно из наиболее значительных различий между
физикой и химией, с одной стороны, и биологией, с другой, состоит в том, что в
первых обычно можно временно изолировать одну переменную величину для изучения,
в то время как в биологии это редко бывает осуществимо».
Знаковой для количественной биологии, стала работа П. Жаккара (Jaccard, 1901). Этим швейцарским
исследователем был предложен коэффициент флористической общности (coefficient de communaute; Gemlinschaftskoefficient), который и по настоящее время не
потерял своей актуальности. В своём историческом исследовании, Х.Х. Трасс (75
лет спустя!) отметит: «Не без оснований Жаккара считают одним из первых
представителей количественно-статистического направления в фитоценологии»
(Трасс, 1976 : 155-156).
В дальнейшем, было опубликовано огромное количество теоретических
разработок и работ, касающихся применения количественных методов в конкретных
предметных областях. Они возникали независимо друг от друга в различных научных
центрах. Особенно преуспели британские математики - Р.Э. Фишер, К. Пирсон и В.
Уэлдон, которых считают основателями биометрии (Лакин, 1990). В США работал
палеонтолог Дж.Г. Симпсон, который среди прочего предложил широко известный
коэффициент (Simpson, 1947). В Европе математические
методы для исследования растительности разрабатывал и активно использовал,
помимо вышеупомянутого П. Жаккара (Jaccard, 1901), легендарный французский ботаник Ж. Браун-Бланке (Braun-Blanquet, 1928). Больших успехов добились
восточно-европейские ботаники - Ж. Кульчинский, К. Флорек с сотрудниками и др.
(Florek et al., 1951).
В нашей стране можно отметить работы А.А. Любищева (например, Любищев,
1923) с использованием коэффициента вариации и его критики, во многих
отношениях интересную книгу по исследованию изменчивости Ю.А. Филипченко
(Филипченко, 1929), публикации И.И. Шмальгаузена (Шмальгаузен, 1935). В
некоторых работах отечественных ботаников изредка применялись коэффициенты
общности, например в работах В.В. Алёхина, А.Е. Жадовского, В.Н. Сукачёва, В.Н.
Беклемишева и др. (Жадовский, 1925; Беклемишев, 1931. Сукачёв, 1931; Алёхин,
1938). Но, как правило, отечественные исследователи использовали в основном
описательную одномерную статистику. Это было связано с тем, что применение
математических методов осуждалось и даже преследовалось благодаря деятельности
Т.Д. Лысенко и его последователей (см., например, материалы электронного
журнала «Биометрика»: #"881651.files/image001.gif">m путём
сравнения объектов, исходя из признаков, (наиболее распространённый в
биологических науках) называется Q-типом анализа, а в случае
сравнения признаков, на основе объектов - R-типом
анализа. Существуют попытки использования гибридных типов анализа (например, RQ-анализ),
но данная методология ещё должным образом не разработана. Один из весьма
интересных вариантов классификации типов анализа представлен в работе Т.Э.-А.
Фрея (Фрей, 1972).
1.4
Меры разнообразия и близости, используемые в сравнительном анализе компонентов
растительного покрова
Прежде
чем перейти к блоку анализа данных в рамках схемно-целевого подхода, необходимо
чётко представлять себе суть используемых аналитических методов. Методика
сравнительного анализа объектов позволяет определять отношения «целого-части»,
которые могут быть равноправными (определяются симметричные отношения
конвергенции или дивергенции), неравноправные (определяются отношения
направленной конвергенции или направленной дивергенции) и различные комбинации
отношений, требующие определения симметричных и несимметричных отношений
конвергенции или дивергенции. Математическое обоснование всем перечисленным
отношениям дано в Приложении А.
Отношения,
как правило, представляются в виде матриц различного типа, а визуализируются с
помощью графов, а также посредством диаграмм Эйлера-Венна. Выбор оптимальной
меры, оценивающей близость объектов, представляет одну из наиболее важных
проблем в систематике, биогеографии и экологии (Sokal, Sneath,
1963; Бейли, 1970; Sneath, Sokal, 1973; Orloci, 1978). Однако, несмотря на большое количество работ,
посвященных рассмотрению и упорядочиванию индексов близости (Sokal, Sneath,
1963; Roux, Roux, 1967; Василевич, 1969; Cheetham, Hazе1, 1969; Чернов, 1971;
Сёмкин, 1972; 1973 а, б; 1979; 1987 а; Goodall, 1973; Ванчуров, 1975; Orloci,
1975; 1978; Розенберг, 1977; Снетков, Вавилин, 1977; Хеллоуэл, 1977; Миркин,
Розенберг, 1978, 1979, 1983; Шмидт, 1980; Юрцев, Сёмкин, 1980; Песенко, 1982;
Максимов, 1984; Федоров, Смирнов, 1986; Гайдышев, 2001 и др.) проблема создания
общей теории мер близости остается открытой.
Кроме
того, постоянно создаются новые меры для определения отношений сходства
(различия). По этому поводу весьма иронично высказывается Э. Мэгарран: «При
измерении разнообразия всегда кажется, что одного термина будет недостаточно, а
нужно ещё полдюжины» (Мэгарран, 1992 : 41). Следовательно, общая теория мер
близости должна выполнять, кроме прочего, «санитарную» функцию, т.е. не
допускать создания дублирующих и ошибочных индексов, которые не привносят в
общую теорию ничего нового или наносят вред.
Приведём
обзор мер, используемых в биологических и географических науках для
аналитических интерпретаций на основе аксиоматического подхода, математическое
обоснование которым дано в Приложении А. При рассмотрении соответствующих
интерпретаций мы будем придерживаться следующих обозначений - если в
исследовании использовалась формула близости хотя бы в одной из четырех
интерпретаций (при n ≥
2), то мы рассматриваем
соответствующие формулы для всех остальных интерпретаций. Например, формула
Кульчинского была известна для трех интерпретаций - множественной,
дескриптивной и вероятностной, следовательно, мы можем привести соответствующее
аналитическое выражение и для информационной интерпретации.
1.4.1 Унарные индексы
При изучении растительных сообществ и флор широко используются меры
изменчивости, как отдельных признаков, так и частот распределения случайных
величин (Филипченко, 1929; Василевич, 1960; 1969; 1971; 1972; 1983; Работнов,
1961; Работнов, Саурина, 1971; Ипатов, 1962; Росс, Власова, 1966; Грейг-Смит,
1967; Гиляров, 1969; Журавлева, Горшкова, 1971; Алексеенко, 1976; Миркин,
Розенберг, 1978, 1983; Миркин, Розенберг, Наумова, 1989; Шмидт, Малышев, 1983;
Зайцев, 1984; Злобин, 1984). Наиболее полный обзор по применению индексов
изменчивости в биологических исследованиях приведен в следующих работах
(Яблоков, 1966; Айвазян, 1968; Мамаев, 1972; Левич, 1980; Песенко, 1982).
Унарные коэффициенты весьма многочисленны и разнообразны. Большинство мер
можно упорядочить в два коэквивалентных (взаимодополняющих) класса (Сёмкин,
1987 б, Песенко, Сёмкин, 1989): меры однородности (также известные как
меры концентрации) и меры разнообразия. Рассмотрим наиболее известные
меры.
1.4.1.1 Коэффициент вариации
При оценке изменчивости некоторых количественных показателей видов
растений и животных широко используется коэффициент вариации (Йогансен, 1933;
1935; Шмальгаузен, 1935; Алпатов, 1956; Берг, 1956; 1958; Рогинский, 1959;
Урбах, 1963; 1964; Шмидт, 1964; Яблоков, 1966; 1968; Артемьев, 1969; Егоров,
1969; Смирнов, 1971; Дмитриев, 1972; Мамаев, 1972; Дюльдин, 1973; Котенко,
1980; Snedecor, 1989; Уланова, 1995). Несмотря на
критику отдельных свойств коэффициента вариации некоторыми исследователями
(Duncker, 1908; Любищев, 1923; Philiptschenko, 1923 цит. по: Филипченко, 1929;
Haldane, 1955 цит. по: Яблоков 1966; Ефимов, 1976) он остаётся одной из часто
используемых мер изменчивости.
Некоторые проблемы интерпретации закономерностей изменчивости признаков,
установленные с помощью коэффициента остались нерешёнными. Так по-разному
решается вопрос о зависимости коэффициента вариации от среднего значения. Одни
исследователи (Василевич, 1960; 1969; Ипатов, 1962) на различных объектах
(ассоциации верховых болот и широколиственного леса) установили, что численное
значение коэффициента вариации для проективного покрытия доминантных видов
имеет меньшее знание, чем для малообильных видов. Эта закономерность
упомянутыми авторами считается естественной. Однако некоторые зоологи (Котенко,
1980), исследовавшие зависимость коэффициента вариации от абсолютных значений
признака установили, что он не зависит от абсолютной величины признаков, а
встречающаяся тенденция увеличения коэффициента вариации с уменьшением
абсолютной величины признаков может быть объяснена наличием падения точности
его измерения, а при исключении влияния степени точности промеров он отражает
лишь величину разнообразия признака. Существует мнение (Песенко, 1982), что
показатели статистического разнообразия, такие как среднеквадратическое
отклонение и коэффициент вариации не могут являться мерами видового
разнообразия. Отмеченная проблема, видимо, обусловлена неудовлетворительными
знаниями авторов о математических свойствах коэффициента вариации (Сёмкин,
Горшков, 2009 б).
Приведём ряд аналитических выражений коэффициента вариации и дадим его
геометрическую интерпретацию (Сёмкин, Горшков, 2009 б). Коэффициент вариации,
предложенный К. Пирсоном (Pearson 1896 цит. по Филипченко, 1929) был записан в
следующем виде:
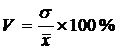 ,
,
где
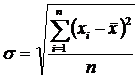 ;
; 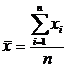 ,
, 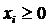 ,
, 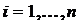 .
По-видимому, этот безразмерный коэффициент изменчивости был получен как
показатель среднего квадратического отклонения для нормированного вариационного
ряда
.
По-видимому, этот безразмерный коэффициент изменчивости был получен как
показатель среднего квадратического отклонения для нормированного вариационного
ряда 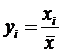 ,
, 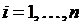 .
Известно, что коэффициент вариации может быть записан в следующем виде (Крамер,
1975 : 392):
.
Известно, что коэффициент вариации может быть записан в следующем виде (Крамер,
1975 : 392):
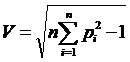 ,
,
где
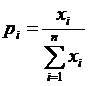 , тогда
, тогда 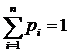 . Эту же
формулу получили М.М. Юзбашев и Т.М. Агапова (Юзбашев, Агапова, 1988) подставив
в формулу коэффициента вариации значения долей.
. Эту же
формулу получили М.М. Юзбашев и Т.М. Агапова (Юзбашев, Агапова, 1988) подставив
в формулу коэффициента вариации значения долей.
Рассмотрим
геометрическую интерпретацию коэффициента вариации. Представим выборку долей 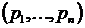 как точку в n-мерном
евклидовом пространстве. Тогда легко установить следующее соотношение
графически (см. рис. 1):
как точку в n-мерном
евклидовом пространстве. Тогда легко установить следующее соотношение
графически (см. рис. 1): 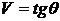 , где
, где 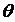 - угол в
начале координат
- угол в
начале координат 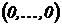 и заключённый между лучами, проходящими через точки
и заключённый между лучами, проходящими через точки 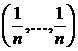 и
и 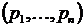 .
Следовательно, коэффициент вариации измеряет близость распределения долей
.
Следовательно, коэффициент вариации измеряет близость распределения долей 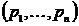 к эталону (абсолютно однородному распределению
долей), причём близость оценивается через
к эталону (абсолютно однородному распределению
долей), причём близость оценивается через 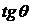 .
.
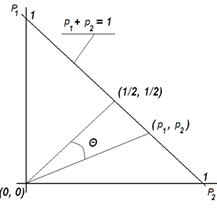
Рис.
1 Геометрическая интерпретация
коэффициента вариации (при n = 2).
Исходя
из геометрической интерпретации коэффициента вариации, легко установить
следующие его свойства: 1) значения коэффициента могут быть больше 1; 2) он
зависит от числа измерений (эталон изменяется в зависимости от n);
3) значения его не зависят от среднего арифметического.
В
качестве эталона можно брать различные распределения долей и частот. Например,
если рассматривать площадное равномерное распределение различных по качеству
выделов растительности, то в этом случае эталон будет характеризовать наибольшее
разнообразие растительности. В случае распределения долей или частот
встречаемости видов, эталон 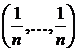 будет
характеризовать максимальное видовое разнообразие. Можно определить также
двойственную меру для коэффициента вариации (Сёмкин, 1971 а):
будет
характеризовать максимальное видовое разнообразие. Можно определить также
двойственную меру для коэффициента вариации (Сёмкин, 1971 а):
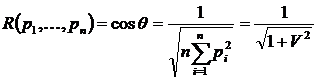 .
.
Эта
мера может служить для определения видового разнообразия и принимает
максимальное значение (R = 1) при
максимальном разнообразии. Двойственная ей мера (отличная от коэффициента
вариации) предложена Г.С. Розенбергом (Миркин, Розенберг, 1978) и называется
мерой диссонанса (Розенберг, 1975), которая определяет различие между реальным
и эталонным распределением видов и может быть записана в виде:
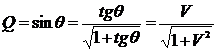 .
.
Эта
мера однородности (концентрации) распределения долей эквивалентна коэффициенту
вариации и имеет одно из преимуществ перед ним - она ограничена сверху 1.
Однако можно произвести нормировку коэффициента вариации на максимально
возможное его значение при заданном n, чтобы
получить верхнюю его границу равную 1:
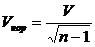 .
.
Действительно,
можно найти расстояние между заданным распределением долей и эталоном, а также
нормировать его на максимально возможное расстояние. Тогда получим меру однородности
распределения частот:
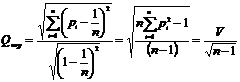
В
случае вероятностного подхода, рассматриваются распределения частот pi по S классам и в вышеприведённых формулах все n
следует заменить на S. Для меры разнообразия распределения частот можно
рассчитать дисперсию (Песенко, Сёмкин, 1989):
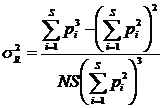 ,
,
где
N - число измерений; S - число
классов; pi - частоты. Таким же способом можно рассчитать
дисперсии и для других мер разнообразия и однородности.
1.4.1.2 Параметрические семейства унарных мер
Первое обобщение для мер разнообразия было предложено Реньи (Rényi, 1961; Песенко, Сёмкин, 1989):
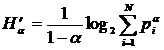 ,
, 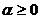 .
.
При
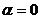 получаем
получаем 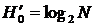 ; при
; при 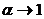 индекс идентичен формуле Шеннона, при
индекс идентичен формуле Шеннона, при 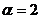 получаем
получаем 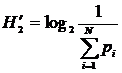 ; при
; при 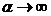 получаем
получаем 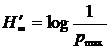 , здесь в
знаменателе индекс Бергера-Паркера (Berger, Parker, 1970),
который определяется как максимум из всех рассматриваемых долей. В дальнейшем в
экологии чаще использовались формулы на основе натуральных логарифмов. Для
оценки разнообразия сообщества весьма часто используется индекс Шеннона (Shannon,
1948 а, б):
, здесь в
знаменателе индекс Бергера-Паркера (Berger, Parker, 1970),
который определяется как максимум из всех рассматриваемых долей. В дальнейшем в
экологии чаще использовались формулы на основе натуральных логарифмов. Для
оценки разнообразия сообщества весьма часто используется индекс Шеннона (Shannon,
1948 а, б):
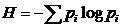 ;
; 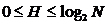 .
.
Н-функцию часто называют индексом Шеннона-Уивера, хотя
правильно называть её индексом Шеннона (Shannon, 1948 а,
б). В совместной книге К. Шеннона и У. Уивера (Shannon, Weaver,
1949), в которую вошли статьи У. Уивера, популяризируются работы Р. Хартли в
области теории информации. В связи с этим, эту работу часто неправильно
цитируют как совместную работу К. Шеннона и У. Уивера (Шеннон, 1963). Функцию H
с обратным знаком использовал Н. Винер (Wiener, 1948). В
связи с чем, эту функцию называют формулой Шеннона-Винера. Она не адекватна
вышеприведённой формуле, как порою ошибочно считают (например, Уланова, 1995).
По-видимому, первым аналог индекса Шеннона в экологии привёл Р. Мак-Артур (MacArthur, 1955). Теоретически Н-функция принимает максимальное значение
тогда, когда имеет место полная выравненность распределения (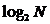 ), что соответствует наибольшему разнообразию системы.
Иногда, чтобы избавиться от непривычной для биолога единицы измерения «бит»
производят нормировку индекса, например так:
), что соответствует наибольшему разнообразию системы.
Иногда, чтобы избавиться от непривычной для биолога единицы измерения «бит»
производят нормировку индекса, например так: 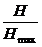 . Индекс
Шеннона получил широкое распространение в биологии (Bertalanffy von, 1950;
Бартков, Сёмкин, 1973; Одум, 1975; Свирежев, Логофет, 1978; Фёдоров, Гильманов,
1980; Песенко, 1982; Мэгарран, 1992; Протасов, 2002 и др.).
. Индекс
Шеннона получил широкое распространение в биологии (Bertalanffy von, 1950;
Бартков, Сёмкин, 1973; Одум, 1975; Свирежев, Логофет, 1978; Фёдоров, Гильманов,
1980; Песенко, 1982; Мэгарран, 1992; Протасов, 2002 и др.).
На
основе формулы Реньи М. Хиллом был предложен континуум мер выравненности (evenness)
на основе унифицированной формулы (Hill, 1973), определенной как
антилогарифм от 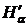 :
:
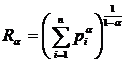 ,
, 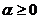 .
.
Приведем
примеры для некоторых случаев: 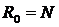 ;
; 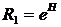 ;
; 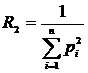 (здесь
просматривается индекс Симпсона:
(здесь
просматривается индекс Симпсона: 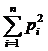 , также
отметим сходный индекс Макинтоша:
, также
отметим сходный индекс Макинтоша: 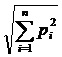 );
); 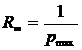 . Позднее, на основе данной формулы был создан ряд мер
(Sheldon, 1969; Heip, 1974; Alatalo, 1981; Molinari, 1989). Отметим также известное семейство мер
. Позднее, на основе данной формулы был создан ряд мер
(Sheldon, 1969; Heip, 1974; Alatalo, 1981; Molinari, 1989). Отметим также известное семейство мер 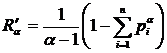 (Patil, Taillie, 1979). Ранее (Песенко, Сёмкин, 1989) было показано,
что семейства
(Patil, Taillie, 1979). Ранее (Песенко, Сёмкин, 1989) было показано,
что семейства 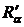 и имеют
плохо интерпретируемый диапазон значений. Также заметим, что меры разнообразия
и имеют
плохо интерпретируемый диапазон значений. Также заметим, что меры разнообразия 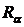 и
и 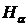 эквивалентны
друг другу
эквивалентны
друг другу 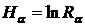 .
.
Особенно
отметим семейство мер концентрации (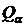 ) А.Н.
Колмогорова. Его меры коэквивалентны мерам семейства Хилла как
) А.Н.
Колмогорова. Его меры коэквивалентны мерам семейства Хилла как 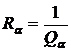 . Стоит отметить, что индексы без логарифмов позволяют
не рассматривать преимущества различных логарифмов с определённым основанием.
. Стоит отметить, что индексы без логарифмов позволяют
не рассматривать преимущества различных логарифмов с определённым основанием.
1.4.1.3 Другие индексы для оценки разнообразия
Следует
отметить, что различные меры разнообразия были известны и до работ К. Шеннона
(см. например, Yule, 1944). Без привязки к параметричсеким семействам
используются следующие меры: индекс Глисона: 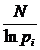 ; индекс
Маргалефа:
; индекс
Маргалефа: 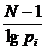 ; индекс Менхиника:
; индекс Менхиника: 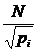 , где N
- число видов; pi - число особей в образце. Рассмотрению особенностей
этих мер посвящено немало публикаций (Hurlbert, 1971; Левич, 1980;
Сёмкин, 1980; Песенко, 1982; Мэгарран, 1992; Протасов, 2002; Frosini,
2003; Magurran, 2004; Шитиков, Розенберг, Зинченко, 2005 и др.). К
информационным индексам разнообразия также относят индекс Бриллюэна:
, где N
- число видов; pi - число особей в образце. Рассмотрению особенностей
этих мер посвящено немало публикаций (Hurlbert, 1971; Левич, 1980;
Сёмкин, 1980; Песенко, 1982; Мэгарран, 1992; Протасов, 2002; Frosini,
2003; Magurran, 2004; Шитиков, Розенберг, Зинченко, 2005 и др.). К
информационным индексам разнообразия также относят индекс Бриллюэна: 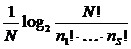 (его аналог для экологии был приведен Р. Маргалефом),
который можно использовать при большом количестве сравниваемых показателей
(Сёмкин, Ралько, 1996).
(его аналог для экологии был приведен Р. Маргалефом),
который можно использовать при большом количестве сравниваемых показателей
(Сёмкин, Ралько, 1996).
Подход
к флорам и фаунам как дескриптивным множествам видов или их групп (крупных
таксонов, элементов флоры) позволяет использовать для сравнительного изучения
флористических систем весьма простой, но вполне строгий математический аппарат
алгебры дескриптивных множеств, приведённый нами в Приложении А. Он отражает
(при сравнении различных структур флоры) пропорциональный вклад каждого таксона
и элемента, а не только его порядковый номер в нисходящем ряду (Сёмкин,
Клочкова, Гусарова, Горшков, 2010 в). Напротив, коэффициенты ранговой
корреляции, часто используемые при сравнении таксономических и иных структур
флоры (Малышев, Байков, 1998; Шмидт, 1980), при большей трудоемкости их расчета
огрубляют выражение соотношений таксонов или элементов, переводя его из шкалы
отношений в более слабую порядковую шкалу (Орлов, 2004).
В
литературе проблема оценки таксономического разнообразия рассмотрена достаточно
подробно (Мэгарран, 1992). При оценке разнообразия внутри конкретной флоры или
фауны, в простейшем случае инвентаризационное разнообразие можно оценить
видовым богатством, или числом видов. Подобные меры разнообразия были
предложены Б.И. Семкиным в 1971 году (Семкин, 1971б; 1973а), а также Р.А.
Акоффом и Ф.Ф. Эмери в 1972 году (Акофф, Эмери, 1974). В 1973 году Б.И. Семкин
(Сёмкин, 1973б) предложил абсолютную меру разнообразия, основанную на сравнении
исследуемого весового множества с эталоном, имеющим максимальное разнообразие:
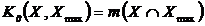 ,
,
где
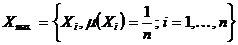 ; X - весовое множество, разнообразие которого
определяется; n - число таксонов. Легко произвести нормировку этой
меры и получить относительную меру разнообразия:
; X - весовое множество, разнообразие которого
определяется; n - число таксонов. Легко произвести нормировку этой
меры и получить относительную меру разнообразия:
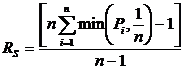 .
.
Некоторые аналогичные идеи были изложены в работе Ю.А. Песенко и Б.И.
Сёмкина (Песенко, Сёмкин, 1989). Большинство исследователей (Акофф, Эмери,
1974) сравнивали определённые объекты с идеальным объектом, но существует идея
использования нескольких естественных эталонов. Одним из таких эталонов может
являться климаксное сообщество и, следовательно, упомянутые меры можно
использовать для оценки сукцессионной динамики, а также для определения уровня
антропогенной нагрузки. Эта область, видимо, будет активно развиваться в связи
с интересом к оценке инвентаризационного биологического разнообразия
(География…, 2002; Сёмкин, Клочкова, Гусарова, Горшков, 2010 в).
.4.2 Бинарные индексы
Самыми распространёнными мерами близости в биологии являются меры
близости двух объектов (см. например, Chao et al., 2000). Приведем для них обзор на
основе аксиоматического подхода по четырём аналитическим интерпретациям
(математическое обоснование приведено в Приложении А.2). Обозначения мер по
табл. 1 Приложения А.
1.4.2.1 Множественная интерпретация
В основе данной интерпретации лежит известное утверждение о том, что
любой видовой список можно представить в виде конечного множества (Сёмкин,
Куликова, 1981). В некоторых случаях используются эквивалентное представление
списков видов в форме булевых наборов или векторов (Сёмкин, 1978 а, б).
Рассмотрим абсолютные меры:
Абсолютная мера сходства (7М): n(ХÇY) - число видов общих для обеих флор X и Y
(мера пересечения двух множеств X и Y). Обычно используется в
промежуточных расчетах при вычислении относительных мер (Андреев, Решетников,
1978; Андреев, 1979; 1980; Юрцев, 1978; Шмидт, 1980; Баркалов, 1981; Борзова,
Сёмкин, Ковалюх, 1981; Сёмкин, Куликова, 1981).
Несимметричные меры различия (4М, 5М): n (X\Y) = n(X) -
n (XÇY),
n (Y\X) = n(Y) - n (X\Y) - число дифференциальных видов флоры Х относительно Y и
флоры Y относительно Х соответственно.
Эти меры используются для характеристики флор на основе дифференциальных
видов одной флоры относительно другой (Юрцев, 1978; Юрцев, Сёмкин, 1980;
Сёмкин, Куликова, 1981).
Симметричная мера различия (6М): d(Х,Y) = n(X) + n(Y) - 2n(XÇY)), где d(X,Y) - число дифференциальных видов
двух флор X и Y.
Абсолютная мера различия может быть представлена в следующей форме:
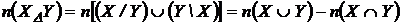 .
.
Функция
(6М) является обобщённым расстоянием Хэмминга (Сёмкин, Куликова, 1981).
Приводится в ряде работ (Godron, 1968; Stephenson, Williams, Lance, 1968;
Юрцев, 1978; Юрцев, Сёмкин, 1980; Сёмкин, Куликова, 1981; Андреев, Козлов,
1983; Гайдышев, 2001 и др.).
Аналогично рассмотрим относительные меры.
Меры включения (16M, 17М):
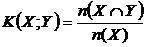 ,
, 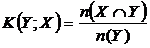
означают
число общих видов двух флор по отношениям к числу видов одной из флор. Впервые,
указанные меры под названием коэффициентов «неспецифичности» одной флоры
относительно другой использовал во флористике Б.А. Юрцев (Юрцев, 1968; Сёмкин,
2007). Подобные меры, под названием коэффициентов «полноты» и «точности» нашли
широкое применение в информационно-поисковых системах (Clevardon, 1963;
Сэлтон, 1973; Шульц, 1973).
После
работ Б.И. Семкина и Т.А. Комаровой (Сёмкин, 1973; Сёмкин, Комарова, 1977;
Комарова, Сёмкин, 1976 а, б) меры включения стали широко использоваться в
синэкологии и биогеографии (Zakharov, 1974; Андреев, 1978; Андреев, Решетников,
1978; 1981; Андреев, Козлов, 1983; Сёмкин, 1978 а; 1980; Юрцев, 1978;
Борисовец, Тараканова, 1979; Петровский, Королева, 1979; Афонина, 1981;
Баркалов, 1981; Полозова, Юрцев, 1981; Максимова, 1982; Максимова, Нечаева, Сёмкин,
1982; Песенко, 1982; Седельников, 1982; Кушнарёв, 1983; Михалева, Петухова,
1983; Филатова, 1983; Хоментовский, 1983).
Меры
невключения (12М, 13М): 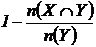 ,
, 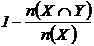
коэффициенты
специфичности флор или фаун (Голлербах, 1936; Юрцев, 1968; 1978; Толмачев,
1974; Ребристая, 1977; Алексахина, Штина, 1984; Сёмкин, 2007).
Приведём
относительные меры сходства.
Коэффициент общности Жаккара (27М):
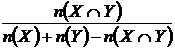 ,
,
где
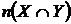 - число видов общих в двух списках видов, n(Х)
и n(Y) - число видов в первом и втором сравниваемых списках
соответственно. Формулу Жаккара можно записать иначе:
- число видов общих в двух списках видов, n(Х)
и n(Y) - число видов в первом и втором сравниваемых списках
соответственно. Формулу Жаккара можно записать иначе:
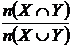 или
или 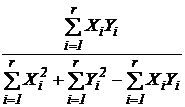 ,
,
где
Xi, Yi - компоненты булевых векторов Х и
Y (т.е. компоненты, принимающие только два значения 0 и 1).
Под названием коэффициента Танимото (или расширенный коэффициент Жаккара)
используется в хемоинформатике (Tanimoto, 1957; Рагойша, URL и др.).
Коэффициент
Жаккара уже более 100 лет используется в сравнительном анализе флор и фаун
(Jaccard, 1901; 1902; 1908; 1912; 1928; Harper, 1915; Алёхин, 1925; Беклемишев,
1931; 1961; 1970; Сукачев, 1931; Быков, 1953; 1957; 1973; 1978; 1983;
Жадовский, 1953; Sokal, Sneath, 1963; Шенников, 1964; Greig-Smith, 1964;
Hagmeier, Stults, 1964; Кац и др., 1965; Češka, 1966; Hagmeier, 1966; Maddocks, 1966; Valentine,
1966; Грейг-Смит, 1967; Макаревич, 1967; 1971; Roux, Roux, 1967; Mello,
Buzas, 1968; Александрова, 1969; Василевич, 1969; Gounot, 1969; Ниценко, 1971;
Levandovsky, Winter, 1971; Красилов, 1972; Сёмкин, 1972 а; 1978 а; Воронов,
1973; Goodall, 1973; Sneath, Sokal, 1973; Толмачев, 1974; Mueller-Dombois, Ellenberg,
1974; Дажо, 1975; Лащинский, Рогинская, 1975; Тамарин, Шмидт, 1975; Чернов,
1975; Шаткаускас, 1975; Леме, 1976; Трасс, 1976; Галанин, 1977; 1979; 1980;
Ребристая, 1977; Андреев, 1979, 1980; Норин, 1979; Сергиенко, 1979; Неронов,
Лушекина, 1980; Шмидт, 1980; Юрцев, Сёмкин, 1980; Сапегин, 1981; Кусакин,
Растомов, 1982; Песенко, 1982; Алексахина, Штина, 1984; Уланова, 1995).
Индекс сходства Сёренсена (22М):
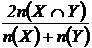 .
.
Эта
мера сходства эквивалентна коэффициенту Жаккара (Сёмкин, Двойченков, 1973). В
синэкологии и биогеографии индекс Сёренсена (Sörensen, 1948) широко используется (Braun-Blanquet, 1951; Dagnelie,
1960; Greig-Smith, 1964; Макфедьен, 1965; Williams, Lance, 1965; Češka, 1966; Roux, Roux, 1967; Василевич, 1969; Шеляг-Сосонко, Куковица,
1969; Gounot, 1969; Макаревич, 1971; Заки, Шмидт, 1972; Красилов,
1972; Romaniszyn, 1972; Воронов, 1973; Goodall, 1973;
Кожевников, 1974; Mueller-Dombois, Ellenberg, 1974; Дажо, 1975; Одум, 1975; Тамарин, Шмидт, 1975;
Чернов, 1975; Goldsmith, Harrison, 1976; Семкин, Комарова, 1977; Семкин, 1979;
Абакумов, Филиппова, 1978; Быков, 1978; Захаров, 1978; Миркин, Розенберг, 1978;
Dzwonko, 1978; Норин, 1979; Сергиенко, 1979; Трасс, 1976;
Уиттекер, 1980; Шмидт, 1980; Юрцев, Сёмкин, 1980; Юрцев, Секретарёва, 1983;
Баркалов, 1981; Гусарова, Семкин, 1982; Марина, 1982; 1983; Несис, 1982;
Песенко, 1982; Пийн, 1982; Суханов, 1982; Зайцев, 1984; Кафанов, Жуков,
Федотов, 1984; Пушкарь, Короткий, Семкин, 1984; Уланова, 1995).
Индекс
сходства Кульчинского (24М):
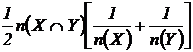 .
.
Индекс
редко используется при сравнении конкретных флор и фаун, но часто приводится в
теоретических и обзорных работах по математическим методам в таксономии и
экологии (Kulczinsky, 1927; Sokal, Sneath, 1963; Макфедьен, 1965; Češka, 1966; 1968; Roux,
Roux, 1967; Peters, 1968; Александрова, 1969; Василевич, 1969; Cheetham, Hazel,
1969; Gounot, 1969; Семкин, Комарова, 1977, 1985; Сёмкин, 1979; Миркин,
Розенберг, 1978; Пименов и др., 1978; Юрцев, Семкин, 1980; Песенко, 1982).
Индекс
сходства Охаи (23М):
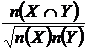 .
.
Индекс
в основном упоминается в теоретических разработках (Ochiai, 1957; Barkman,
1958) и в обзорных работах (Sokal, Sneath, 1963; Roux, Roux, 1967; Peters, 1968; Cheetham,
Hazel, 1969; Sepkovski, 1974; Ванчуров, 1975; Миркин, Розенберг, 1978;
Сёмкин, Комарова, 1985). Также встречается под названием косинусный коэффициент
(cosine similarity coefficient).
Индекс
сходства Шимкевича-Симпсона (25М):
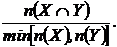
Индекс можно записать в эквивалентных формах (Сёмкин, Комарова, 1977):
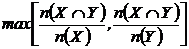 или
или 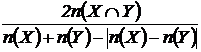 .
.
Д.
Шимкевич (Szymkiewicz, 1934) предложил «показатель родового сходства»,
который в математической записи идентичен индексу сходства Симпсона.
Коэффициент сходства Симпсона широко используется во флористике и фаунистике (Simpson,
1943; 1947; 1960; Long, 1963; Hagmeier, Stults, 1964; Peters, 1968; Cheetham, Deboo, 1969; Cheetham, Hazel, 1969; Oltz, 1969; Красилов, 1972; Whittington, Hughes,
1972; Urbani, Collingwood, 1976; Сёмкин, Комарова, 1977; Ashmole,
1978; Песенко, 1982). Его часто путают с мерами включения. Также встречается
под названием «коэффициент перекрытия».
Индекс
Браун-Бланке (21М):
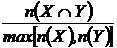
Эквивалентные записи индекса:
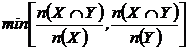 или
или 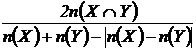
Поучительна
история открытия этого индекса. Ж. Браун-Бланке (Braun-Blanquet, 1928; 1932)
ошибочно записал коэффициент общности Жаккара в виде отношения числа общих
видов к числу видов большей флоры (эту ошибку критиковал Л. Кох (Koch,
1957)). Однако в переизданной в 1951 году книге Браун-Бланке исправил свою
допущенную ошибку, убрал приводимый пример расчета коэффициента общности двух
флор, и привел формулу коэффициента сходства Сёренсена (Braun-Blaquet, 1951).
Несмотря на все это, ошибка проникла в книгу по экологии растений Х.И. Остина
(Oosting, 1953), а затем и в сводки по мерам сходства А. Читама и Дж. Хейзела
(Cheetham, Hazel, 1969).
Индекс
сходства Браун-Бланке вписывается в теорию мер сходства (Юрцев, Сёмкин, 1980;
Сёмкин, Комарова, 1985) и вполне обоснованно может быть использован в
сравнительном анализе флор и фаун. Он значительно реже, чем индекс
Шимкевича-Симпсона приводится в обзорах по мерам сходства (Savage, 1960; Peters,
1968; Песенко, 1982) и его также часто путают с мерами включения.
Индекс
сходства Сокала и Снита (31М):
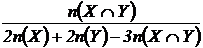
Данный
индекс эквивалентен коэффициентам сходства Жаккара и Сёренсена (Сёмкин,
Двойченков, 1973). Приводится в сводках по численной таксономии и биогеографии
(Sokal, Sneath, 1963; Roux, Roux, 1967; Бейли, 1970; Миркин, Розенберг, 1978;
Сёмкин, 1979; Андреев, 1980; Песенко, 1982).
1.4.2.2 Дескриптивная интерпретация
Описания сообществ можно представить с помощью дескриптивных весовых
множеств (или дескриптивных наборов), введённых Б.И. Сёмкиным (1973 а).
Дескриптивные множества обозначаются следующим образом:
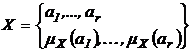 ,
,
где
a1,…,ar - элементы; mX(a1),…,mX(ar) - соответствующие им веса, неотрицательные числа. В
случае фиксированного для определённого исследования элементов 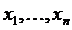 множества X, можно
использовать вместо дескриптивного множества эквивалентное понятие
«дескриптивный набор», т.е. вектор, компонентами которого являются веса
(Сёмкин, 1978 а, б; 1979; Сёмкин, Куликова, 1981; Сёмкин, Комарова, 1985).
Основное требование, предъявляемое к дескриптивным наборам теорией измерений
(Суппес, Зиннес, 1967) - однородность компонентов набора, т.е. каждый компонент
набора должен быть измерен в одной и той же шкале отношений. Это свойство
дескриптивных наборов позволяет находить сумму его компонентов. Обозначая
дескриптивный набор как
множества X, можно
использовать вместо дескриптивного множества эквивалентное понятие
«дескриптивный набор», т.е. вектор, компонентами которого являются веса
(Сёмкин, 1978 а, б; 1979; Сёмкин, Куликова, 1981; Сёмкин, Комарова, 1985).
Основное требование, предъявляемое к дескриптивным наборам теорией измерений
(Суппес, Зиннес, 1967) - однородность компонентов набора, т.е. каждый компонент
набора должен быть измерен в одной и той же шкале отношений. Это свойство
дескриптивных наборов позволяет находить сумму его компонентов. Обозначая
дескриптивный набор как 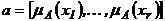 или
или 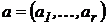 , свойство
его однородности можно записать как
, свойство
его однородности можно записать как 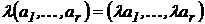 , λ>0.
, λ>0.
Рассмотрим
абсолютные меры в указанной интерпретации.
Абсолютная
мера сходства (7Д):
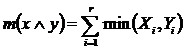 .
.
Важным
случаем является абсолютная мера сходства нормированных дескриптивных наборов:
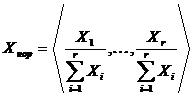 и
и 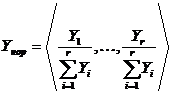 .
.
В
подобных случаях значения меры сходства заключены между 0 и 1. В
синэкологических исследованиях эти индексы нашли широкое применение как меры процентного
сходства при использовании относительных единиц измерения (в процентах) и как
меры пересечения в промежуточных расчетах относительных мер сходства (Renkonen,
1938; Шорыгин, 1939; Whittaker; 1952; 1970; Whittaker, Fairbanks, 1958; Bray, Curtis, 1957; Clausen, 1957; Dahl, 1957; Ramsay, 1964; Василевич, 1969; Константинов, 1969;
Ковалевская и др., 1970; Mauchline, 1972; Сёмкин, 1973 а; Сёмкин, Куликова,
1981; Сёмкин, Комарова, 1985; Чернов, 1975; Заки, Шмидт, 1977; Уиттекер, 1980;
Юрцев, Сёмкин, 1980; Баркалов, 1981; Марина, 1982, 1985; Быков, 1983; Пааль,
Сёмкин, 1983; Zangerl, Bazzar, 1983; Кобелева, 1984; Сличенкова, 1984; Соколова,
1984).
Абсолютная
мера различия (6Д):
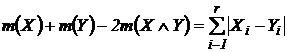 .
.
Известное
в математике обобщённое расстояние Хэмминга, называемое «ситиблок-метрикой» или
«l-нормой», приводится в ряде работ по экологии и систематике (Sokal,
Sneath, 1963; Василевич, 1969; Sneath, Sokal, 1973; Айвазян, Бежаева,
Староверов, 1974; Orloci, 1975; 1978; Дюран, Оделл, 1977; Сёмкин, Куликова,
1981; Штейнгауз, 1981; Песенко, 1982; Миркин, Розенберг, 1983; 1989).
Для дескриптивной интерпретации рассмотрим относительные меры.
Меры включения (16Д, 17Д):
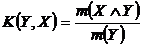 ,
, 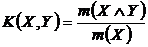
или
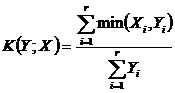 ,
, 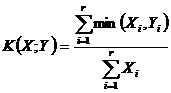 .
.
В
практику синэкологических исследований были введены Б.И. Сёмкиным и Т.А.
Комаровой (Сёмкин, 1973 а; 1978 а, б; Семкин, Комарова, 1977; 1985; Комарова,
Сёмкин, 1976 а, б).
Меры
включения используются в других разделах биологии и таксономии для сравнения
дескриптивных наборов (Сэлтон, 1973; Андреев, 1979, 1980; Сёмкин, Куликова,
1981; Суханов, 1982; Михалева, Петухова, 1983; Пааль, Сёмкин, 1983; Юрцев,
1983; Сёмкин, Орешко, Горшков, 2009 б).
Меры невключения (12Д, 13Д):
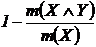 ,
, 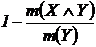 .
.
В
практике синэкологических исследований ещё не нашли применения.
Приведём относительные меры сходства:
Мера сходства Ружечки (27Д):
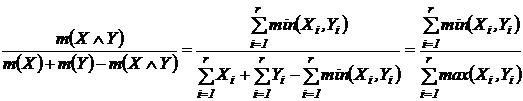 .
.
В
сравнительном анализе сообществ растений и животных широко использовался (Ružička, 1958; Наумов, 1964; Равкин, 1967; Чернов, 1971,
1978; Сёмкин, 1973 а; Сёмкин, Комарова, 1977; Шаткаускас, 1975; Комарова, 1978;
Федоров, 1979; Фёдоров, Гильманов, 1980; Песенко, 1980, 1982; Цыбулин, 1980;
Полозова, 1981; Шадрина, 1982). Иногда используют меру различия, являвшуюся
дополнением до единицы коэффициента сходства Ружечки (Marcsewski, Steinhaus,
1958; Комарова, 1978; Федоров, 1979; Песенко, 1980), которая удовлетворяет
аксиомам метрики (Сёмкин, Двойченков, 1973).
Коэффициент
сходства Чекановского (22Д):
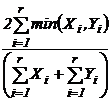
Широко
используется в сравнительном анализе дескриптивных наборов (Czekahowski,
1909; 1932; Dahl, Hadač, 1949 цит. по Kershaw, Looney, 1985; Odum,
1950; Ярошенко, 1953; 1961; Culberson, 1955; Bray, Curtis, 1957; Greig-Smith, 1964; Golf, Gottam, 1967; Миркин, 1970; Дыренков, Волков, 1971; Чернов,
1971; 1975; Васильев, Сёмкин, 1974; Zakharov, 1974; Фридланд, Рожков,
1975; Корчагин, 1976; Goldsmith, Harrison, 1976; Заки, Шмидт, 1977; Семкин, 1973 а, б; 1979;
Сёмкин, Комарова, 1977, 1985; Townsend, Hildrew, 1979; Песенко, 1980, 1982; Уиттекер, 1980; Быков,
1983; Пааль, Сёмкин, 1983; Кобелева, 1984; Пушкарь, Короткий, Сёмкин, 1984).
Коэффициенты сходства Ружички и Чекановского эквивалентны (Сёмкин, Двойченков,
1973).
Индекс
сходства Кульчинского (24Д):
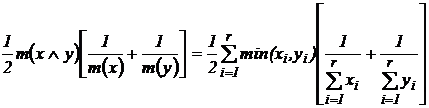 .
.
Используется
сравнительно редко в исследовании сообществ растений и животных (Kulczynsky,
1928; Сукачев, 1931; Szafer et al., 1964; Александрова, 1969; Василевич,
1969; Сёмкин, 1973 а, б; 1979; Сёмкин, Комарова, 1977; 1985; Андреев, 1979, 1980;
Песенко, 1982).
Индекс
сходства (21Д):
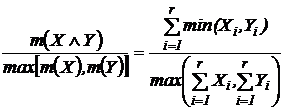 .
.
Предложен Б.И. Сёмкиным (1979).
Индекс сходства (25Д):
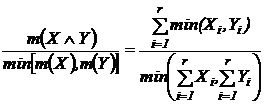 .
.
Использовался
в системах поиска информации (Сэлтон, 1973). В биоценологию введен Б.И.
Сёмкиным и Т.А. Комаровой (Сёмкин, Комарова, 1977, 1985; Сёмкин, 1979). Для
булевых наборов этот индекс совпадает с индексом Шимкевича-Симпсона (28М).
Индекс
сходства (23Д):
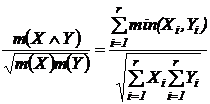 .
.
Аналог
коэффициента Охаи (28М) для данной интерпретации (Сёмкин, 1979; Сёмкин,
Комарова, 1985; Песенко, 1982;).
Индекс
сходства (31Д):
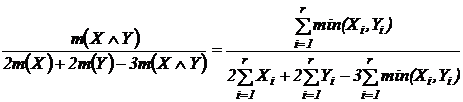 .
.
Аналог
коэффициента Сокала и Снита (34М) на случай использования дескриптивных
множеств (Сёмкин, 1979).
1.4.2.3 Вероятностная интерпретация
Если данные получены в результате статистического эксперимента, мы имеем
полное право производить расчёты, используя меры в рамках данной интерпретации.
Обычно, события, и их вероятности (частоты) представляют в виде таблицы
сопряженности 2×2 (Sokal, Sneath, 1963; Василевич, 1969; Кендалл, Стюарт,
1973; Goodall, 1973; Сёмкин, 1978; Песенко, 1982; Kershaw, Looney, 1985).
Отметим, что таблицы сопряжённости или таксономические таблицы, которые
приводятся у ряда авторов (Sokal, Sneath, 1963; Василевич, 1969; Sneath, Sokal,
1973; Дюран, Оделл, 1977; Песенко, 1982; Digby, Kempton, 1987) для
множественной и дескриптивной модели, содержательно неверны и не могут
использоваться для этих интерпретаций. Данная таблица может быть использована
только для вероятностных интерпретаций.
Рассмотрим варианты абсолютных мер.
Абсолютная
мера совместимости событий (7В): 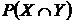 . Оценка меры производится с помощью частот:
. Оценка меры производится с помощью частот: 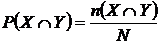 , где
, где 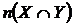 - число
совместных встреч X и Y; N - число проведенных
экспериментов. Использовалась в промежуточных расчетах при определении
относительных мер (Коломыц, 1976; Сёмкин, Варченко, 1978).
- число
совместных встреч X и Y; N - число проведенных
экспериментов. Использовалась в промежуточных расчетах при определении
относительных мер (Коломыц, 1976; Сёмкин, Варченко, 1978).
Абсолютная
мера различия двух событий (6В):
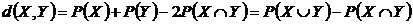
Мера
является расстоянием между событиями. Используется только при расчетах
относительных мер.
Рассмотрим относительные меры в рамках настоящей интерпретации. Стоит
отметить, что часто меры совместимости событий (т.е. вероятностные меры) иногда
путают с мерами сходства (Sokal, Sneath, 1963).
Меры включения событий (несимметричные меры Дайса) (16В, 17B):
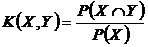 ,
, 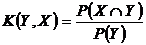 .
.
Меры
Дайса - условные вероятности двух событий (Яглом, Яглом, 1973; Колмогоров,
1974). Использовались рядом исследователей (Hacker, 1921; Dice, 1945;
Cassagnan, 1961; Василевич, 1969; Goodal, 1973; Сёмкин, 1979; Сёмкин, Варченко,
1978).
Рассмотрим
относительные меры совместимости событий.
Коэффициент
Иверсена (27B):
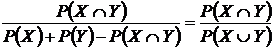 .
.
Использовался
рядом исследователей (Agrell, 1945; Iversen, 1954; Ellenberg, 1956; Whittaker,
Fairbanks, 1958; Шенников, 1964; Василевич, 1969; Чернов, 1971, 1975; Goodall,
1973; Порк, 1974; Варченко, 1978; Сёмкин, 1978 а, б, 1979; Семкин, Варченко,
1978).
Коэффициент
Дайса-Брея (22В):
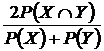 .
.
Индекс
введен в экологию Л. Дайсом (Dice, 1945), в дальнейшем использовался Р. Бреем (Bray,
1956) и другими исследователями (Василевич, 1969; Миркин, 1970; Чернов, 1971;
1975; Goodall, 1973; Pielou, 1977; Варченко, 1978; Сёмкин, Варченко, 1978;
Сёмкин, 1979). В некоторых работах используется под названием индекса Дайса или
индекса Брея-Кёртиса.
Широко
использовался при расчетах парциальных мер совместимости по данным таблиц
сопряженности (Петропавловский, Сёмкин, Худяков, 1982; Семкин, Усольцева, 1982;
Усольцева, 1985).
Коэффициент
совместимости событий (24В):
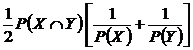 .
.
Часто отождествляется с коэффициентом сходства Кульчинского (Sokal,
Sneath, 1963; Sneath, Sokal, 1973). Приводится как мера совместимости событий
Б.И. Сёмкиным (1979) и встречается в промежуточных расчетах у B.C. Ипатова с
сотрудниками (Ипатов, Самойлов, Тархова, 1974).
Коэффициент совместимости событий (23В):
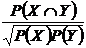 .
.
Предложен
в работе Б.И. Сёмкина (1979). Его часто отождествляют с коэффициентом сходства
Охаи (Sokal, Sneath, 1963; Sneath, Sokal, 1973).
Коэффициент
совместимости событий Гудолла (25В):
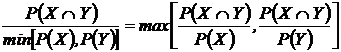 .
.
Предложен
Д. Гудоллом (Goodall, 1973). Рассматривался в работе Б.И. Семкина (1979).
1.4.2.4 Информационная интерпретация
Данная интерпретация описывает отношения категоризованных данных, т.е.
данных, представленных в виде частот наблюдений, попавших в некоторые категории
или классы (Кендалл, Стьюарт, 1973; Елисеева, Рукавишников, 1977; Аптон, 1982).
Рассмотрим абсолютные меры.
Абсолютная мера зависимости (7И):
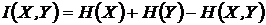 ,
,
где
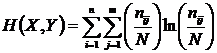 ;
;
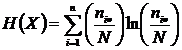 ,
, 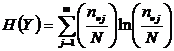 .
.
Мера
широко используется в исследованиях при определении зависимости признаков,
(Godron, 1968; Juhasz-Nagy, 1967; Orloci, 1968; Пузаченко, Мошкин, 1969; Сёмкин,
1973 а; Conant, 1973; Yasuichi, 1973; Арманд, 1975, 1979; Басаргин, Горовой,
Сёмкин, 1978).
Метрика
Шеннона (6И):
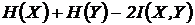 .
.
Или
в эквивалентной записи:
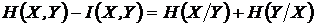 ,
,
где
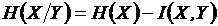 ,
, 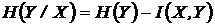 -
условные энтропии. Доказательство метричности функции
-
условные энтропии. Доказательство метричности функции 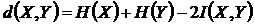 можно найти в работах (Сёмкин, 1973 в; Yasuichi,
1973).
можно найти в работах (Сёмкин, 1973 в; Yasuichi,
1973).
Интересны
в данной интерпретации и относительные меры.
Относительные
меры односторонней зависимости (16И, 17И):
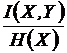 ,
,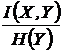
Меры
односторонней зависимости используются во многих разделах биологии и географии
(Nakahama, Nishioka, 1966; Nakahama et al.,
1966; Фрей, 1969; Красникова, Антомонов, 1970; Соnаnt, 1973; Арманд, 1975,
1979; Сёмкин, Ююкин, 1976; Сёмкин, Наумова, 1981; Сёмкин, Усольцева, 1983;
Pielou, 1977; Пузаченко, Мошкин, Сёмкин, 1979; Усольцева,
1985).
Относительные
меры односторонней независимости (12И, 13И):
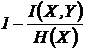 ,
, 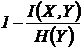
Эти
меры коэквивалентны мерам (16И, 17И).
Рассмотрим
относительные меры взаимозависимости.
Мера
взаимозависимости Райского (27И):
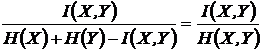 .
.
Коэквивалентная
ей мера 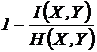 - есть метрика (Сёмкин, 1973 в; Yasuichi, 1973). Мера
широко используется в экологии и систематике (Raijski, 1961,
a, б; Hawkaworth, Estabrook, Rogers, 1968; Orloci, 1968; Сёмкин, 1973, в;
Семкин, Ююкин, 1976; Yasuichi, 1973; Пярна, 1976; Басаргин, Горовой, Сёмкин,
1978).
- есть метрика (Сёмкин, 1973 в; Yasuichi, 1973). Мера
широко используется в экологии и систематике (Raijski, 1961,
a, б; Hawkaworth, Estabrook, Rogers, 1968; Orloci, 1968; Сёмкин, 1973, в;
Семкин, Ююкин, 1976; Yasuichi, 1973; Пярна, 1976; Басаргин, Горовой, Сёмкин,
1978).
Мера
взаимозависимости (22И):
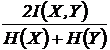 .
.
Эта
мера взаимозависимости впервые предложена Б.И. Сёмкиным (1972 а, б; 1973 в) и
впоследствии её независимо использовали чешские авторы (Řehak, Řehakova, 1973).
Как и предыдущая мера, она применяется в научных исследованиях для определения
степени взаимозависимости двух признаков (Басаргин, Горовой, Сёмкин, 1978;
Сёмкин, 1979, 1981; Сёмкин, Усольцева, 1982; Коломыц, 1984; Усольцева, 1985).
Мера
взаимозависимости (25И):
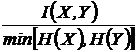 .
.
Использовалась
в климатологии и систематике растений (Conant, 1973).
Меры
взаимозависимости (21И, 23И, 24И):
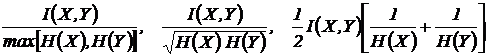
Эти
меры предложены Б.И. Сёмкиным (1979) и еще не нашли применения при оценке
взаимозависимости признаков.
Дополнительно
рассмотрим оценки выборочности мер зависимости. Для расчета дисперсий мер
зависимости использовалась формула Фишера (Фишер, 1958; Ли, 1978; Сёмкин,
Наумова, 1981). Приведем дисперсии для наиболее используемых мер зависимости:
а)
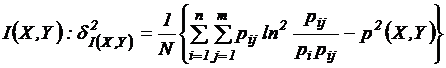 ,
,
б)
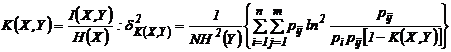 ,
,
в)
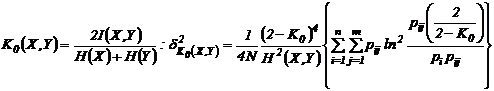 ,
,
г)
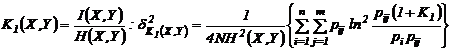 ,
,
где
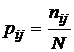 ;
; 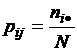 ;
; 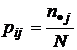 , N - число измерений.
, N - число измерений.
Приведенные
дисперсии использовались рядом исследователей для оценки ошибки выборочности
мер зависимости (Сёмкин, Ююкин, 1976; Сёмкин, Наумова, 1981; Семкин, Усольцева,
1982; Басаргин, Горовой, Сёмкин, 1978; Баранов, Басаргин, 1979; Ююкин, Сёмкин,
1979 а, б).
Приведенные
четыре интерпретации не исчерпывают всего многообразия S-функций. Однако
большинство используемых в синэкологии и таксономии индексов сходства,
совместимости и зависимости были получены с помощью указанных интерпретаций.
Неохваченные
теорией S-функций оказались индексы различия и сходства, основанные на
евклидовом расстоянии. Расширение S-функций за счет отказа от
выполнимости некоторых из четырех аксиом, по-видимому возможно, но при этом
теряются некоторые из свойств несимметричных индексов (Сёмкин, Горшков, 2008).
1.4.3 Определение n-арных отношений
Во многих биологических науках, особенно в экологии, зачастую приходится
иметь дело с n-арными
отношениями (в литературе также встречаются названия «multiple-site similarity measure» (Baselga et al., 2007; Diserud, Ødegaard, 2007), «multidimensional coefficient» (Сёмкин, Горшков, 2009 а), «multiple-community measure» (Chao et al. 2000). Система аксиом для n-арных показателей приведена в Приложении
А. Используя тот же подход, что и при рассмотрении бинарных отношений (бинарные
отношения являются, по сути, частным случаем n-арных, при n = 2), представим аналогичные аналитические
интерпретации для n-арных
мер (Сёмкин, Горшков, 2009 а; 2010).
1.4.3.1 Множественная интерпретация
Один из первых индексов для сравнения совокупности n-видовых
списков был предложен Л. Кохом (Koch, 1957) под названием «индекс биотической
дисперсии».
Индекс Коха (46М):
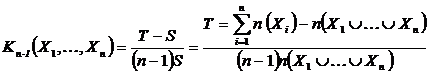 ,
,
где
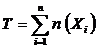 ,
,  ,
, 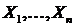 - совокупность n множеств, n(•) - число
элементов множества. Этот индекс широко используется в геоботанике и экологии
(Koch, 1957; Greig-Smith, 1964; Грейг-Смит, 1967; Василевич, 1969; Зарубин,
1971; Лысенко, 1971; Чернов, 1971, 1975; Красилов, 1972; Сёмкин, 1972 а, б;
1973 а; Воронов, 1973; Песенко, 1982). При n = 2, индекс Коха совпадает
с коэффициентом общности Жаккара. В некоторых случаях используют меру различия n
множеств, дополняющую до единицы меру сходства (Сёмкин, 1972 б; Воронов,
Арустамова, 1975):
- совокупность n множеств, n(•) - число
элементов множества. Этот индекс широко используется в геоботанике и экологии
(Koch, 1957; Greig-Smith, 1964; Грейг-Смит, 1967; Василевич, 1969; Зарубин,
1971; Лысенко, 1971; Чернов, 1971, 1975; Красилов, 1972; Сёмкин, 1972 а, б;
1973 а; Воронов, 1973; Песенко, 1982). При n = 2, индекс Коха совпадает
с коэффициентом общности Жаккара. В некоторых случаях используют меру различия n
множеств, дополняющую до единицы меру сходства (Сёмкин, 1972 б; Воронов,
Арустамова, 1975):
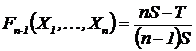 .
.
Индекс (44М):
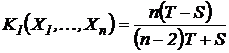
Эквивалентный
индексу Коха индекс (48M), при n = 2 совпадает с коэффициентом
общности Жаккара.
Индекс (43М):
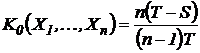 .
.
Индекс
сходства, обобщающий меру сходства Сёренсена на случай сравнения n
множеств предложен Б.И. Сёмкиным (Сёмкин, 1972 а, б; 1973 а, б; 1977). Этот
индекс приводит в своих работах палеозоолог Ю.Д. Захаров (Zakharov, 1974;
Захаров, 1978).
1.4.3.2 Дескриптивная интерпретация
Впервые обобщение индекса Коха на случай сравнения дескриптивных множеств
было предложено Б.И. Семкиным (1972 а, б; 1973 а).
Индекс (46Д):
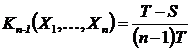 ,
,
где
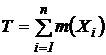 ;
; 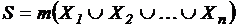 .
.
Дополнение
индекса (46Д) до единицы (Сёмкин, 1972 а, б) для дескриптивных множеств,
в несколько иной записи приводятся в работе В.Д. Фёдорова (1979):
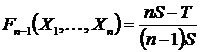 .
.
На
случай дескриптивных множеств можно обобщить и другие индексы, рассмотренные
для множественной интерпретации (43Д), (44Д).
1.4.3.3 Вероятностная интерпретация
Индексы совместимости n событий были предложены в 1972 году Б.И.
Сёмкиным (Сёмкин, 1972 а, б). Формула расчета дисперсий для таких индексов
приводится в работе Л.И. Варченко (1978). В табл. 1 Приложения А
рассматриваются в основном формулы 38В-46В, которые обобщают на
многомерный случай, индексы Дайса-Брея (22В) и Иверсена (27B), а
также дополнения этих индексов до единицы. Приведем только две формулы из них:
Индекс (43В):
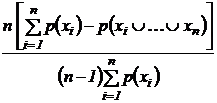 .
.
Индекс (46В):
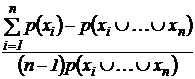
При
n = 2 индекс (43В) совпадает с индексом Дайса-Брея, а индекс (46В)
с индексом Иверсена. Все рассматриваемые индексы (38В-46В) или
эквивалентны, или соответственно коэквивалентны. На практике можно использовать
любой из указанных индексов.
1.4.3.4 Информационная интерпретация
Индексы взаимозависимости n признаков также впервые были
предложены Б.И. Сёмкиным в 1972 году (Сёмкин, 1972 а, б). Рассматриваемые
индексы (38И-46И) обобщают на случай определения взаимозависимости n признаков хорошо известные меры
взаимозависимости двух признаков Райского (27И) и
Сёмкина-Рехак-Рехаковой (22И), а также дополнение этих индексов до
единицы. Например, индекс (43И):
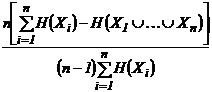 ,
,
при
n = 2 преобразуется в следующий индекс взаимозависимости:
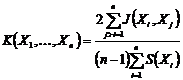 или
или 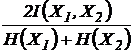 .
.
Все
рассматриваемые индексы (38И-46И) эквивалентны или коэквивалентны
(Сёмкин, Двойченков, 1973) т.е. дают одни и те же результаты при оценке
взаимозависимости n признаков. Следует отметить, что известный в
кибернетике коэффициент избыточности (Быховский, Вишневский, 1971; Пузаченко,
1983), может быть преобразован путем нормировки в индекс (43И).
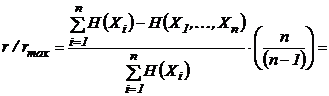
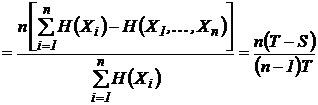 , т.к.
, т.к. 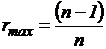 .
.
Рассмотренные
n-арные индексы строятся на основе функций n переменных. Однако в
экологических исследованиях для оценки сходства n описаний или оценки
корреляции n признаков часто используют меры, полученные путем
осреднения 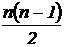 относительных коэффициентов от двух переменных
(Песенко, 1982). Возможно также получить меру сходства n описаний путем
осреднения абсолютных мер сходства (Сёмкин, Степанова, Щербова,
1973). В последнем случае, в обозначении S-функций, получается следующий
индекс:
относительных коэффициентов от двух переменных
(Песенко, 1982). Возможно также получить меру сходства n описаний путем
осреднения абсолютных мер сходства (Сёмкин, Степанова, Щербова,
1973). В последнем случае, в обозначении S-функций, получается следующий
индекс:
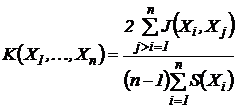 .
.
Например,
для множественной интерпретации, указанный индекс запишется в виде:
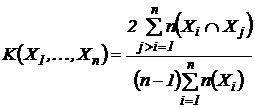 .
.
В
работе (Сёмкин, Степанова, Щербова, 1973) приводится формула, получаемая путем
осреднения 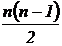 абсолютных мер сходства
абсолютных мер сходства 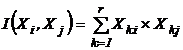 (Xki, Xkj - k-я
компонента i и j векторов xi и xj
соответственно, r - число компонент).
(Xki, Xkj - k-я
компонента i и j векторов xi и xj
соответственно, r - число компонент).
Стоит
также отметить критику существующих n-мерных
показателей Б.И. Сёмкиным (Сёмкин, 2001), а также обзор классов эквивалентных
мер сходства n дескриптивных множеств (Сёмкин, Горшков, 2010).
1.4.4 Другие интерпретации мер
конвергенций и их применение
Обобщая рассмотренную концепцию мер, можно сказать, что основная задача
введения системы S-функций в
том, чтобы объединить все известные меры, используемые в экологии и смежных
науках, под «крышей» одной теории. В этом ракурсе также стоит упомянуть
неохваченные нами в данной работе, но удачно развиваемые другими
исследователями, интерпретации мер близости.
Например, Г.С. Лбовым, сотрудником института математики имени С.Л.
Соболева развивалась логическая интерпретация мер близости (см.
например, Лбов, 1998; Викентьев, Лбов, 1998). В приведённых работах
опубликована система аксиом для логических мер.
Известным отечественным ботаником Б.А. Юрцевым (Юрцев, 1983)
рассматривались отношения между ареалами, фитохоориями с использованием
диаграмм Эйлера. Этот подход можно назвать пространственной интерпретацией
мер близости.
Существует интерпретация экологической ниши как гиперобъёма. Основная
идея, разработанная Дж. Хатчинсоном (Hutchinson, 1957; 1965), получила позднее дополнительное математическое
выражение в книге В.Д. Фёдорова и Т.Г. Гильманова (Фёдоров, Гильманов, 1980).
Упомянутые авторы приводят меру перекрывания экологических ниш двух видов (i и j):
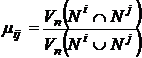 .
.
Здесь
несложно увидеть аналитическое выражение известной меры (27S)
для данной интерпретации. Кроме того, подразумевается, что мера может
использоваться и для многомерного случая. В этой связи интересна работа Б.
Грюнбаума по комбинаторной геометрии (Грюнбаум, 1971). Приведённые в указанной
работе метрики (абсолютная и относительная) могут быть также использованы в
экологии при определении пересечения многомерных экологических ниш. Эти метрики
справедливы и для определения расстояния для других множеств (двумерных фигур и
«ареалов»), флористических множеств (множеств видов) и др.
По
всей видимости, существует ещё огромное число приложений общей теории мер
близости. Мы предполагаем возможность применения (после некоторой адаптации)
нашей методики в сравнительной геномике, биохимии, морфологии и др., т.е. в тех
областях, где уже применяются вышеупомянутые меры (достаточно задать в
параметрах поиска какой-нибудь крупной базы данных по биологическим публикациям
(например, PubMed Central) названия рассмотренных
коэффициентов, чтобы увидеть насколько высока потребность в подобных мерах).
Возможно и другие специалисты найдут применение рассматриваемой методике в
своей профессиональной области.
1.4.5 Меры близости, не
удовлетворяющие системе аксиом
Системы аксиом, приведённые в Приложении A, позволяют получить континуум различных мер и индексов. Ряд
индексов связан между собой монотонными функциями и являются эквивалентными или
коэквивалентными (см. Приложение А.1.3). Неэквивалентные относительные меры обладают
важным свойством. В силу того, что они получены путем осреднения двух
несимметричных мер, для совпадающих или близких мер, они дают совпадающие или
близкие результаты.
Следовательно, для существенно несимметричных объектов достаточно
использовать только одну несимметричную меру, а для существенно симметричных
также достаточно всего одной меры для оценки близости. Для контроля можно
определить две неэквивалентные относительные меры, например, K0
и K¥. Если значения этих мер близки, то все промежуточные меры
имеют также близкие значения в силу, того, что средняя степенная является
монотонно возрастающей функцией параметра h. В этом случае достаточно взять любую из этих мер.
Обычно выбирается наиболее простая из них, которая лучше интерпретируется и визуально
графически изображается. Простота меры важна не для быстроты расчетов, а для её
лучшей интерпретируемости. Это свойство имеет существенное значение при
использовании мер для определения отношений на данных.
Как видно из обзора по использованию различных бинарных индексов в
геоботанике и экологии, преимущество имеют только те из них, которые входят в
семейство мер, получаемых из формулы средней обобщенной степенной двух
несимметричных мер. Наиболее часто использовались из этого семейства следующие средние:
гармоническая, геометрическая, арифметическая.
Из рассмотренных мер сходства и различия только мера сходства
Браун-Бланке и двойственная ей мера различия (расстояние) Юрцева удовлетворяют
условию согласования меры сходства (различия) с соответствующими мерами
включения, что позволяет корректно представлять отношения включения-сходства
(или двойственные им отношения) и графически изображать в виде смешанных
графов, имеющих как дуги для характеристики включения, так и рёбра,
характеризующие отношения сходства (Сёмкин, Орешко, Горшков, 2010). Причём,
последние приведённые меры являются частными случаями мер процентного сходства
(различия). Для парного сравнения флористических признаков (при условии их
равновеликости) вполне достаточно использовать только одну меру процентного
сходства или двойственную ей меру различия, удовлетворяющую аксиомам расстояния
(см. Приложение A).
Но существует большое количество мер, которые используются абсолютно
некорректно. Первая попытка «закрыть» некорректные и дублирующие меры близости
была предпринята в обзоре Ю.А. Песенко (1982). Автором были приведены
эквивалентные коэффициенты, показаны способы создания «новых» коэффициентов, но
при этом автором необоснованно были введены новые классы (Песенко 1982 :
152-154).
Весьма
известным способом создания нового индекса является преобразование: 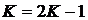 . Например, таким способом можно получить
трансформированный коэффициент Дайса (Миркин, Розенберг, 1978):
. Например, таким способом можно получить
трансформированный коэффициент Дайса (Миркин, Розенберг, 1978):
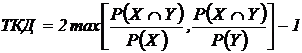 .
.
Коэффициент
измеряет совместимость событий и не измеряет независимость. Он эквивалентен
коэффициенту Гудолла, т.е. 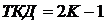 , где K
- коэффициент Гудолла (25В). Аналогично получен индекс Маарела из
индекса Сёренсена-Чекановского (Песенко, 1982).
, где K
- коэффициент Гудолла (25В). Аналогично получен индекс Маарела из
индекса Сёренсена-Чекановского (Песенко, 1982).
Следует
отметить, что понятие сходства является двойственным понятию различия. Понятие
двойственности мер конвергенции и дивергенции исходит из самой системы аксиом
мер конвергенции и дивергенции, приведённой нами в Приложении А. Например,
очевидно что, если два объекта сходны на 60%, то они различаются на 40%. В
связи с этим стоит отметить эклектичные коэффициенты, в которых некоторые
исследователи пытаются объединить меру сходства и меру различия. Одним из таких
коэффициентов является коэффициент Стугрена-Радулеску (Stugren, Radulescu,
1961):
где
X - число видов, встречающихся в одной флоре, но
отсутствующее в другой; Y - число видов, встречающихся во второй флоре, но
отсутствующее в первой; Z - число видов, встречающихся в обеих флорах.
Коэффициент Стугрена-Радулеску изменяется в пределах от - 1 до + 1,
в связи с чем утверждается, что он обладает высокой разрешающей способностью.
Проще говоря, любой коэффициент сходства можно преобразовывать, чтобы его значения
изменялись между - 1 и + 1. Например, K` = 2K - 1, 0 ≤ K ≤ 1.
Существует
аналогичный коэффициент Малышева (Малышев, 1972):
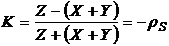 .
.
Представим
в современных теоретико-множественных обозначениях:
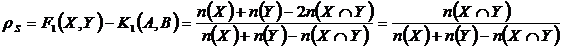 .
.
Следовательно,
коэффициент Малышева равен:
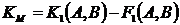
Эти коэффициенты не дают ничего нового в сравнении с коэффициентом
Жаккара, т.к. они ему эквивалентны:
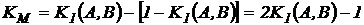 .
.
Отдельно
отметим индекс Маунтфорда (по: Песенко, 1982):
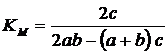 .
.
Этот
индекс некорректен и не может являться мерой сходства, потому что в частном
случае совпадения числа видов в обоих сообществах (т.е. когда а =b= c)
в знаменателе получаем 0. Не корректен также индекс Фейджера (Fager,
1957; 1972):
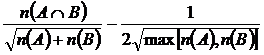 ,
,
т.к.
он не удовлетворяет аксиоме мер сходства (см. Приложение A): 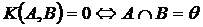 . Далее рассмотрим индекс Вишарта (Wishart, 1969),
использующийся в геоботанике (Bouxin, 1975; Jannsen, 1975; Песенко, 1982):
. Далее рассмотрим индекс Вишарта (Wishart, 1969),
использующийся в геоботанике (Bouxin, 1975; Jannsen, 1975; Песенко, 1982):
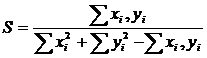 , xi ≥
0, yi ≥
0.
, xi ≥
0, yi ≥
0.
Этот
индекс при значениях xi равных 0 или 1, совпадает с коэффициентом общности
Жаккара. Следовательно, в этом случае мера различия F1 = 1 - S будет
удовлетворять аксиомам расстояния. Для случая xi ≥
0 и yi ≥
0 дополнение индекса Вишарта до 1 не
удовлетворяет аксиомам расстояния. Отметим также коэффициент Симпсона. Этот
индекс при всех его достоинствах не является мерой сходства в строгом смысле,
т.к. не удовлетворяет всем аксиомам S-функций и,
следовательно, является мерой квазисходства (Сёмкин, 2007).
Большую
проблему представляет различия в обозначениях коэффициентов.
Теоретико-множественная запись коэффициентов сходства (различия) появилась в
70-х годах с появлением общих принципов введения этих мер (Сёмкин, Двойченков,
1973). Классическая запись коэффициентов была представлена у П. Жаккара (Jaccard,
1901). Например, коэффициент Жаккара в первоначальной форме имеет следующий
вид:
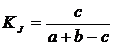 ,
,
где
c - общее количество видов для двух описаний; a
- количество видов в первом описании; b -
количество видов во втором описании. Этот коэффициент переписывался разными
обозначениями, что приводило к большой путанице. Тенденцию к ошибочному
написанию коэффициентов усилили работы Р. Сокала и П. Снита (Sokal,
Sneath, 1963; Sneath, Sokal, 1973), в которых была приведена таблица
сопряжённости (т.е. только на стохастический случай) с отличными от
классических обозначениями. Например, для того же коэффициента Жаккара запись
следующая:
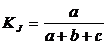 ,
,
где
a - общее количество видов для двух описаний; a
+ b - количество видов в первом описании; a
+ c - количество видов во втором описании. К сожалению,
эта форма записи проникла и в смежные направления (Malde, Jonassen, 2008).
Коэффициент Жаккара в различных модификациях и эклектичных записях активно
используется в молекулярной биологии, биоинформатике, геномике, протеиномике и
других направлениях (например, Snitkin et al.,
2006; Weirauch et al., 2008); аналогично другие
известные в экологии и смежных науках коэффициенты - коэффициент Симпсона
(Aguilar, Oliva, 2008), коэффициент Сёренсена (Bryant et al.,
2009). Во всех этих статьях коэффициенты приведены некорректно. Например, в
работе Д. Голдберг и Ф. Рота (Goldberg, Roth, 2003) приведёны: коэффициент
Жаккара (в теоретико-множественной записи), коэффициент Симпсона (названный
«meet/min coefficient») и возведённый в квадрат коэффициент Охаи (назван
авторами «geometric coefficient»). Таких работ очень много.
Отдельно
стоит сказать про коэффициенты, которые используют показатель d
таксономической таблицы сопряжённости Сокала-Снита (т.е. отсутствие видов)
матрицы сопряжённости (Sokal, Sneath, 1963, Песенко, 1982; География…, 2002). Это,
например, индекс Сокала-Майченера, индекс общности Барони-Урбани, индекс
Бюссера, коэффициент Хаманна, коэффициент Бравэ-Пирсона, коэффициент Юла,
коэффициент Пирсона, расстояние Сокала и др. (многие из этих индексов приведены
в известном обзоре Р. Сокала и П. Снита (1963) или в аналогичном обзоре А. Читама и Дж.
Хезела (1969) и др. (Wolda, 1981)). Видимо авторы подобных коэффициентов желают
учитывать какие-то латентные эффекты, но практика применения данных мер
показала, что это иллюзия. Упомянутые коэффициенты не содержат ничего нового и
при изменении объёма выборки теряют свою ценность в отличие от коэффициентов на
основе пересечения (т.е. числа общих видов), которые отличаются зачастую только
масштабом.
1.4.6 Матрица абсолютных мер
конвергенции
После того как исследователь определил тип данных и предполагаемый
результат, т.е. экстраполировал конкретную цель на конкретный объект
исследования, можно переходить непосредственно к блоку анализа данных. В
экологии на данный момент существует несколько примеров реализации
рассматриваемой схемы на конкретном материале (Сёмкин, Куликова, 1981;
Филатова, 1983; Сёмкин и др., 2007; Сёмкин, Клочкова, Гусарова, Горшков, 2010
а, б и др.).
Ha
основе матрицы данных легко можно рассчитать матрицу абсолютных мер
конвергенций, что, например, для множественной и дескриптивной интерпретаций
соответствует матрице абсолютных мер сходства (матрице пересечений) размером n2 (табл. 2). В вероятностной
интерпретации аналог этой матрицы носит название «матрицы совместных
вероятностей», а в информационной интерпретации это «матрица информационных
функций».
Таблица 2
Матрица пересечения
|
№ флоры
|
1
|
2
|
. . .
|
j
|
. . .
|
n
|
|
1
|
m11
|
m12
|
. . .
|
m1j
|
. . .
|
m1n
|
|
2
|
m21
|
m22
|
. . .
|
m2j
|
. . .
|
m2n
|
|
. . .
|
. . .
|
. . .
|
. . .
|
. . .
|
. . .
|
. . .
|
|
i
|
mi1
|
mi2
|
. . .
|
mij
|
. . .
|
min
|
|
. . .
|
. . .
|
. . .
|
. . .
|
. . .
|
. . .
|
. . .
|
|
n
|
mn1
|
mn2
|
. . .
|
mnj
|
. . .
|
mnn
|
Этот тип матрицы является основным «документом» исследования (после самых
списков видов), т.к. матрица пересечений содержит информацию о числе видов в
каждом списке и число видов общих для каждой пары сравниваемых списков.
Достоинство данной матрицы состоит в том, что по данным этой матрицы можно
рассчитать другие типы матриц (матрицы включения, сходства, транзитивного
замыкания и др.), т.е. реализовать принцип воспроизводимости в узком смысле
(Сёмкин, 1983). Элементы матрицы пересечений определяются по формуле:
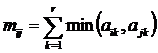 ,
,
где
mij - элемент, находящийся в i-ой строке
и j-ом столбце; 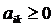 ;
; 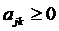 ;
; 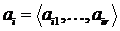 ;
; 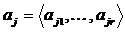 , (i, j = 1, …, n;
k = 1, …, r).
, (i, j = 1, …, n;
k = 1, …, r).
Матрица
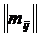 симметрична относительно главной диагонали, т.е.
симметрична относительно главной диагонали, т.е. 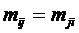 . В связи с этим свойством для дальнейших расчётов
используют только часть элементов, расположенных по диагонали и под диагональю
(общее количество этих элементов равно
. В связи с этим свойством для дальнейших расчётов
используют только часть элементов, расположенных по диагонали и под диагональю
(общее количество этих элементов равно 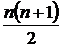 ).
Наддиагональные элементы можно не приводить, т.к. их легко восстановить в силу
симметрии матрицы.
).
Наддиагональные элементы можно не приводить, т.к. их легко восстановить в силу
симметрии матрицы.
Несмотря
на достаточно компактную форму матриц пересечения (количество столбцов и строк
равно количеству сравниваемых объектов), размеры таблиц иногда бывают
достаточно велики. Для частных случаев подобные способы компактной записи
матриц ранее приводились в литературе (см. например (Андреев, Козлов, 1983)).
Рассмотрим здесь общий способ компактной записи приводимых матриц пересечений
(без потери информации). Разобьём квадратную матрицу пересечений на две
прямоугольные матрицы. При чётном n первая прямоугольная
матрица имеет размер 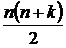 , а вторая
, а вторая 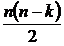 . Возьмём
первую из них, имеющую большую размерность и рассмотрим показатель,
характеризующий полноту заполнения этой матрицы
. Возьмём
первую из них, имеющую большую размерность и рассмотрим показатель,
характеризующий полноту заполнения этой матрицы 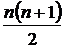 элементами:
элементами:
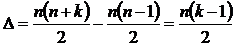 .
.
) Пусть
n и k чётные числа. Тогда при k = 0 и k = 2 имеем  и
и 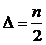 соответственно.
В первом случае невозможно включить всю необходимую информацию из матрицы
пересечений в прямоугольную матрицу, а во втором останутся незаполненными ещё
соответственно.
В первом случае невозможно включить всю необходимую информацию из матрицы
пересечений в прямоугольную матрицу, а во втором останутся незаполненными ещё 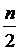 ячеек.
ячеек.
) Пусть
n и k нечётные числа. Тогда при k = 1 и k = 3 имеем 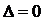 и
и 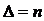 соответственно.
В первом случае квадратная матрица включает всю необходимую информацию из
матрицы пересечений (пустых мест нет), а во втором останутся незаполненными ещё
n ячеек.
соответственно.
В первом случае квадратная матрица включает всю необходимую информацию из
матрицы пересечений (пустых мест нет), а во втором останутся незаполненными ещё
n ячеек.
Для иллюстрации метода приведём примеры прямоугольных таблиц:
 →
→  ,
,
 →
→  .
.
В
случае нормирования меры пересечения к 1, получаем меру процентного сходства
(одна из мер сходства). Это означает, что матрицу мер пересечения можно также
использовать и как матрицу мер сходства (только если мы имеем дело с долями -
на диагонали 100 % (Ковалевская и др., 1970)).
1.4.7 Матрица относительных
несимметричных мер конвергенции
Для ненормированных данных матрица пересечений легко
преобразуется в матрицу мер включения (табл. 3). Для этого необходимо элементы
каждой строки матрицы пересечения разделить на соответствующий этой строке
диагональный элемент. Например, элементы i-ой строки mi1, mi2,…, mij,…, mir делятся на диагональный элемент mii и получается следующая строка:
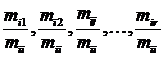 .
.
В
результате получаем матрицу включения (табл. 3).
Таблица
3
Матрица
включения
|
®
|
1
|
2
|
. . .
|
j
|
. . .
|
n
|
|
1
|
m11
/ m11
|
m12
/ m11
|
. . .
|
m1j
/ m11
|
. . . . . .
|
m1n
/ m11
|
|
2
|
m21
/ m22
|
m22
/ m22
|
. . .
|
m2j
/ m22
|
. . .
|
m2n / m22
|
|
. . .
|
. . .
|
. . .
|
. . .
|
. . .
|
. . .
|
. . .
|
|
i
|
mi1 / mii
|
mi2 / mii
|
. . .
|
mij / mii
|
. . .
|
min / mii
|
|
. . .
|
. . .
|
. . .
|
. . .
|
. . .
|
. . .
|
. . .
|
|
n
|
mn1 / mnn
|
mn2 / mnn
|
. . .
|
mij / mnn
|
. . .
|
mnn / mnn
|
Стрелка (®)
указывает направление включения одной флоры в другую. Например, первая строка
матрицы означает степень включения флоры №1 в остальные флоры под №2,
..., n. Элементы матрицы включения,
находящиеся симметрично относительно главной диагонали не равны друг другу,
т.е. матрица включения несимметрична относительно диагонали, а на самой
диагонали значения равны 1 (или 100%).
Матрица включения является весьма полезным источником информации об
отношениях «целого-части» и адекватно показывает «роль» каждого объекта в общей
системе (Сёмкин, Орешко, Горшков, 2009 б). Из этой матрицы мы можем получить
ориентированные графы включения-сходства (рассмотрим ниже).
При рассмотрении большого количества списков видов и наличии многих
односторонних и двухсторонних связей между этими списками, графы
включения-сходства теряют геометрическую наглядность. В подобных случаях
целесообразно использовать индексы преобладания полустепеней захода дуг графа
над полустепенями исхода дуг для вершины графа. Этот индекс рассчитывается по
соответствующей формуле (Сёмкин, Куликова, 1981):
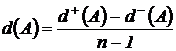 ,
,
где
d (A) - индекс «преобладания» вершины A
графа; d+ (A) - число дуг, входящих в вершину A
графа; d- (A) - число дуг, исходящих из вершины A
графа; n - число вершин графа. Полученные данные удобно
представить в виде таблицы. Этот метод позволяет достаточно чётко выделять
наиболее «оригинальные» и наиболее «банальные» флоры. Рассмотренный метод уже
применяется в экологических исследованиях (Кафанов, 1994; Сёмкин и др., 2007;
Сёмкин, Селедец, Майоров, Горшков, 2009; Сёмкин, Гарбузова, Майоров, Горшков,
2009; Сёмкин, Клочкова, Гусарова, Горшков, 2010 а, б).
Следует
особо отметить, что меры включения более информативны, чем меры сходства, т.к.
последние, по сути, представляют собой усреднённые показатели и, следовательно,
теряют часть информации об объектах, а меры включения адекватно оценивают более
распространённые в природе нетранзитивные отношения. Например, один список
может включаться на 100% в другой список, а другой список в свою очередь
включаться только на 10%. При этом коэффициент сходства не сможет адекватно
отразить эти отношения, т.к., например, 10 общих видов значительны для одного
списка в 10 видов, но не столь значимы для большого списка в (к примеру) 100
видов. Мера сходства Сёренсена в данном случае будет равна около 20%.
Можно
даже построить аналог дендрограммы, используя меры включения (тогда сходство
будет определяться по наименьшему из включений). Но с точки зрения идеологии
схемно-целевого подхода приведённые манипуляции не имеют смысла, т.к. там, где
мы для разновеликих множеств используем только меры включения, не имеет смысла
строить дендрограмму, которая отражает симметричные отношения сходства.
1.4.8 Матрица относительных
симметричных мер конвергенции
Существует два способа получения матрицы сходства (Сёмкин, Орешко,
Горшков, 2010). Классический способ - это получение матрицы сходства напрямую
путём расчёта конкретной меры сходства для каждой пары объектов.
Второй способ (Сёмкин, 2007) заключается в симметризации матрицы
включения (см. Приложение А). Данный способ позволяет обеспечить требуемую
преемственность между мерами включения и мерой сходства по классу τ. Например, мере сходства Сёренсена (K0,-1 (A,B)) соответствуют меры включения K0,-1 (A;B) и K0,-1
(B;A), а мере сходства Жаккара (K1,-1 (A,B)) соответствуют меры включения K1,-1 (A;B) и K1,-1
(B;A).
Последовательно преобразуя матрицу включения (табл. 3), получаем типичную
матрицу сходства (табл. 4).
Таблица 4
Матрица сходства
|
№ флоры
|
1
|
2
|
. . .
|
j
|
. . .
|
n
|
|
1
|
1
|
K12
|
. . .
|
K1j
|
. . .
|
K1n
|
|
2
|
K21
|
1
|
. . .
|
K2j
|
. . .
|
K2n
|
|
. . .
|
. . .
|
. . .
|
. . .
|
. . .
|
. . .
|
. . .
|
|
i
|
Ki1
|
Ki2
|
. . .
|
Kij
|
. . .
|
Kin
|
|
. . .
|
. . .
|
. . .
|
. . .
|
. . .
|
. . .
|
. . .
|
|
n
|
Kn1
|
Kn2
|
. . .
|
Knj
|
. . .
|
1
|
Следует отметить, что данный приём удобен именно при схемно-целевом
подходе (Сёмкин, 2009). Матрица сходства является информационной основой для
получения закономерностей сходства списков. На данном этапе можно использовать
и матрицу различия (расстояний), которая будет двойственной матрице сходства
(подробнее в работе: Сёмкин, Орешко, Горшков, 2010 и в Приложении A).
1.4.9 Алгоритмы упорядочивания данных
Исследователями отношений близости разработано весьма
солидное количество методов упорядочивания данных. Самыми распространёнными
являются методы теории графов (Оре, 1969, 2002; Харари, 2003, Робертс, 1986) и
кластерного анализа (Дюран, Оделл, 1977; Мандель, 1978; Айвазян и др., 1989).
Обзор методов упорядочивания данных можно посмотреть в следующих работах
(Орлов, 2009; Бериков, Лбов, 2008).
Практически все алгоритмы упорядочивания данных
исходят из того, что некое множество объектов характеризуется определённой
степенью связности. Здесь сразу же появляются первые «подводные камни». Между
объектами может просто не оказаться никакой связи, но математический аппарат
нечувствителен к таким «мелочам» и в любом случае выдаёт классификационное
разбиение.
На эту проблему неоднократно указывали исследователи
методов кластерного анализа и теоретико-графовых методов (Дюран, Оделл, 1977;
Сокал, 1980; Соломон, 1980; Кафанов, Борисовец, Волвенко, 2004; Кафанов, 2005
а). А.И. Кафанов с соавторами в одной из своих последних работ по кластерному
анализу (Кафанов, Борисовец, Волвенко, 2004), после довольно резкой и подробной
критики методов, отметили: «Таким образом, исследователь, выбирая тот или иной
метод обработки эмпирических данных, предварительно должен для себя решить,
должна ли получаемая им схема в принципе быть иерархической». Эта мысль чётко
коррелирует с основной идеей схемно-целевого подхода и заставляет исследователя
чётко сформулировать цель до применения математических методов.
Кроме того, следует заметить, что почти все алгоритмы
упорядочивания данных имеют графическую форму представления, что позволяет
получить такие преимущества как выразительность, доходчивость, лаконичность,
универсальность и обозримость. «Достоверно и то, что теория графов служит
математической моделью для всякой системы, содержащей бинарное отношение»
(Харари, 2003).
Рассмотрим основные методы упорядочивания данных,
применяемых в биологических науках и формы их графического представления. В
настоящей диссертационной работе нами не рассматривались методы ординации, т.к.
они преследуют другие цели (Goodall,
1954; Александрова, 1969; Kershaw,
Looney, 1985; Миркин, Наумова, Соломещ,
2002).
1.4.9.1 Кластерный анализ
Впервые термин «кластерный анализ» (cluster (англ.) -
гроздь, сгусток, пучок) был предложен математиком К. Трионом (по Мандель,
1978). Метод широко используется в геоботанике, экологии и численной таксономии
и др. (Goodall, 1970; 1973; Jardine, Sibson, 1971; Austin, 1972; Sneath, Sokal,
1973; Дюран, Оделл, 1977; Pielou,
1977; Джефферс, 1981; Сёмкин, 1987 в). Можно выделить основные подходы,
используемые в кластерном анализе (по: Бериков, Лбов, 2008):
1. Вероятностный подход. Предполагается, что каждый
рассматриваемый объект относится к одному из k классов (в терминологии Орлова (2004) это методы
сортировки, распознавания образов, обучения с учителем и т.п.). К алгоритмам этого
подхода относится: метод k-средних,
метод FOREL.
2. Подходы на основе систем
искусственного интеллекта. Кластер представляется в виде нечёткого множества (алгоритм нечётких C-средних), в виде нейрона
искусственной нейронной сети (самоорганизующиеся сети Кохонена), в виде
условных хромосом (генетические алгоритмы) и др.
3. Логический подход. Построение дендрограммы
осуществляется с помощью дерева решений.
4. Теоретико-графовый подход. К алгоритмам этого подхода
относится: алгоритм кратчайшего незамкнутого пути, алгоритм ближайшего соседа и
др.
5. Иерархический подход. Предполагается наличие вложенных
групп (кластеров различного порядка). Алгоритмы в свою очередь подразделяются
на агломеративные (объединительные) и дивизивные (разделяющие). По количеству признаков
иногда выделяют монотетические и политетические методы классификации. Последние
были хорошо рассмотрены в обзорной работе Ю.А. Песенко (Песенко, 1982).
В данной диссертационной работе будем использовать
иерархические агломеративные методы. Результаты группировки указанными методами
представляется в форме дендрограммы, которая является графом без циклов,
напоминающим дерево. Дендрограмма позволяет изобразить взаимные связи между
объектами из заданного множества. Для создания дендрограммы требуется матрица
сходства (или различия), которая определяет уровень сходства между парами
объектов. Далее необходимо выбрать метод построения дендрограммы, который
определяет способ пересчёта матрицы сходства (различия) после объединения
очередных двух объектов в кластер.
В работах по кластерному анализу описан довольно
внушительный ряд способов построения дендрограмм. Рассмотрим основные:
1. Метод одиночной связи (single linkage). Также известен, как «метод ближайшего соседа».
2. Метод полной связи (complete linkage). Также известен, как «метод дальнего соседа».
. Метод средней связи (pair-group method using arithmetic averages).
3.1. Невзвешенный (unweighted).
3.2. Взвешенный (weighted).
4. Центроидный метод (pair-group method using
the centroid average).
4.1. Невзвешенный.
4.2. Взвешенный (медианый).
5. Метод Уорда (Ward`s method).
Для первых трёх методов существует общая формула,
предложенная А.Н. Колмогоровым (по Айвазян и др., 1989):
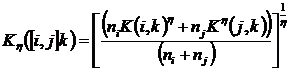 ,
, 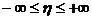 ,
,
где
[i,
j] - группа из двух объектов (кластеров) i и j;
k - объект (кластер), с которым ищется сходство
указанной группы; ni - число элементов в i
кластере; nj
- число элементов в j кластере. Как видим, формула схожа с формулой
осреднения мер включения из Приложения А.
Центроидный
метод использует для пересчёта матрицы расстояний (Sneath and Sokal, 1973). В
качестве расстояния между двумя кластерами в этом методе берется расстояние
между их центрами тяжести. Для этих методов также есть общая формула Ланса-Уильямса
(Lance, Willams, 1967).
В
методе Уорда в качестве расстояния между кластерами берется прирост суммы
квадратов расстояний объектов до центров кластеров, получаемый в результате их
объединения (Ward, 1963). В отличие от других методов кластерного анализа для
оценки расстояний между кластерами, здесь используются методы дисперсионного
анализа. На каждом шаге алгоритма объединяются такие два кластера, которые
приводят к минимальному увеличению целевой функции, т.е. внутригрупповой суммы
квадратов. Этот метод направлен на объединение близко расположенных кластеров.
В 1987 году Б.И. Сёмкиным (Сёмкин, 1987 в) было проведено исследование ряда
методов построения дендрограмм. Наиболее оптимальным был признан метод средней
взвешенной связи. Хотя, следует отметить, что явные связи между мерами близости
и методами построения дендрограмм ещё только предстоит выявить.
Отдельно
следует сказать о моменте, которому не уделяется почти никакого внимания. Речь
идёт о выборе значения близости для двух объектов, объединяющихся в кластер.
Традиционно производится поиск максимального элемента по матрице сходства (или
минимального для матрицы расстояний) и на объекты, на пересечении которых стоит
искомое значение, объединяются в кластер. Однако, иногда возникают ситуации
когда в матрице содержится несколько значений, которые удовлетворяют данному
условию. Возникает неопределённость математического алгоритма. Как правило, в
компьютерных программах задаётся определённое правило (например, поиск вести
слева направо, сверху вниз), но в литературе особого указания на сей счёт не
дано. В общем, такая ситуация означает, что необходимо приводить все случаи
дендрограмм, которые возникают в подобных точках бифуркации.
1.4.9.2 Диаграмма Чекановского
Метод «диагонализации» матрицы различия и графическое изображение
кластеров вдоль главной диагонали матрицы различия (диаграмма Чекановского)
впервые предложен И. Чекановским в 1909 году (Czekanowski, 1909). В то время, в математике был
известен метод приведения матриц к блочно-диагональному виду для «чётких
отношений» (отношений эквивалентности), а И. Чекановский впервые распространил
этот метод на «нечёткие» отношения «сходства и различия» (см. Сёмкин, 1987 б,
в; 1988). Метод Чекановского получил распространение в геоботанике и флористике
для выделения однородных групп (Ružička, 1953, 1958; Fager, 1957; Roux, Roux,
1967; Александрова, 1969; Василевич, 1969; Шмидт, 1980). В своём обзоре П.
Грейг-Смит (Greig-Smith, 1967) приводит диаграмму Чекановского для диаграмм
сходства, но ссылается на работу 1962 года П. Снита и Р. Сокала.
Приведём описание методики: «Сущность этого метода заключается в том, что
всю амплитуду полученных величин сходства разбивают на ряд классов, а затем в
матрице величин сходства заменяют эти величины штриховкой, различной для
каждого класса, причём обычно для более высоких классов сходства применяют
более тёмную штриховку. Затем, меняя порядок описаний в таблице, добиваются
того, чтобы более сходные описания оказались рядом» (Василевич, 1969 : 159).
Приведём гипотетический пример получения вышеуказанной диаграммы. Основой
метода является построение матрицы транзитивного замыкания (Tamura et al., 1971). Для построения матрицы транзитивного замыкания
возьмём простую матрицу сходства (рис. 2 а) и умножим её саму на себя:
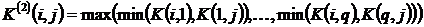 ,
,
где
K(i,
j) - элемент, стоящий на пересечении i-ой
строки и j-го столбца в новой (второй) матрице, полученной после
первой итерации; q - общее количество строк (столбцов) матрицы сходства.
Данную процедуру необходимо продолжать пока матрица не станет самоподобной: 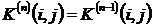 , где n - число итераций. Для этого
сначала сравниваем элементы первой строки с первым столбцом, определяя
минимальное значение. В нашем случае самоподобной стала матрица А2
(рис. 2 б), т.е. при последующих итерациях она даёт саму себя. Далее мы
преобразовываем матрицу таким образом, чтобы близкие числовые значения
находились рядом (рис. 2 в). Если каждому числовому значению присвоить
какой-либо цвет или оттенок цвета (как в нашем случае), то мы получим
классическую диаграмму Чекановского (рис. 2 г). Традиционно более тёмный цвет
соответствует большему сходству, а более светлый меньшему.
, где n - число итераций. Для этого
сначала сравниваем элементы первой строки с первым столбцом, определяя
минимальное значение. В нашем случае самоподобной стала матрица А2
(рис. 2 б), т.е. при последующих итерациях она даёт саму себя. Далее мы
преобразовываем матрицу таким образом, чтобы близкие числовые значения
находились рядом (рис. 2 в). Если каждому числовому значению присвоить
какой-либо цвет или оттенок цвета (как в нашем случае), то мы получим
классическую диаграмму Чекановского (рис. 2 г). Традиционно более тёмный цвет
соответствует большему сходству, а более светлый меньшему.

Рис. 2. Схема расчёта и построения диаграммы Чекановского. Обозначения: а-б -
расчётные матрицы транзитивного замыкания; в - преобразованная матрица
транзитивного замыкания; г - диаграмма Чекановского.
Стоит отметить, что приоритет использования этого метода И. Чекановским
оспаривается (Макаревич, 1971). В.Н. Макаревич считает, что метод построения
диаграммы Чекановского по своей сути является модификацией метода
дифференциального анализа, разработанного К. Пирсоном.
1.4.9.3 Неориентированные графы
Широко применяются в геоботанике и флористике. Их
часто называют корреляционными плеядами (Терентьев, 1959, 1960; Выханду, 1964;
Василевич, 1969; Миркин, Розенберг, 1979; Шмидт, 1980). Частным случаем
является метод, известный как метод построения оптимальных деревьев
(дендритов), который был предложен Г. Штейнгаузом (1981), впоследствии метод
был развит математиками вроцлавской таксономической школы (Florek et al., 1951; Василевич, 1969; Шмидт, 1980). Имеется достаточно
большое количество исследований, в которых был использован данный метод (Matuszkiliewicz, Polakowska, 1955; Falińsky, 1960; Kowal, 1965).
Обычно дендрит строят, используя метод одиночной связи
(Сёмкин, 1979), но известно (Сёмкин, 1987 в), что эквивалентную структуру можно
получить и при использовании матрицы транзитивного замыкания, т.к. оба метода
основаны на учёте единственной, наиболее сильной из связей между сравниваемыми
объектами.
Изредка неориентированные графы применяются для
методов оптимизации (Сёмкин, 1978 б; Выханду, 1964). В частности Л.К. Выханду
(1964) занимался вопросами использования в биологии метода построения
максимального корреляционного пути.
1.4.9.4 Ориентированные графы
Ориентированные графы (орграфы) были впервые
использованы Б.И. Сёмкиным и Т.А. Комаровой в фитоценологии при изучении
сукцессий растительности (Сёмкин, Комарова, 1971). В последующем эти методы
нашли широкое применение в биогеографии и экологии (Юрцев, Сёмкин, 1980;
Сёмкин, Куликова, 1981), т.к. удачно применяются для визуализации
несимметричных отношений.
Например, на основе матрицы включения, возможно
построение графа включения при условии предварительного выбора порога
(например, d ³ 50%). Просматривая строки матрицы
включения, отмечаем все значения, которые превышают пороговую величину. Вершину
графа с номером строки соединяем с вершиной графа номера столбца, на пересечении
которого и стоит отмеченное число, а стрелка указывает направление включения.
Граф включения показывает связь флористических участков (по сходству видов).
Рассмотрим гипотетический пример. Возьмём некую условную матрицу включения
(табл. 5).
Таблица 5
Матрица включения для гипотетического примера
|
→
|
1
|
2
|
3
|
4
|
5
|
|
1
|
100
|
44
|
35
|
48
|
17
|
|
2
|
46
|
100
|
32
|
32
|
18
|
|
3
|
38
|
34
|
100
|
43
|
29
|
|
4
|
50
|
32
|
41
|
100
|
27
|
|
5
|
31
|
31
|
46
|
46
|
100
|
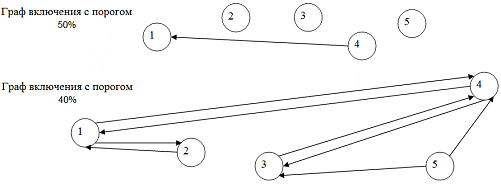
Рис. 3. Гипотетический пример
ориентированного графа включения-сходства для пяти объектов при разных порогах.
Согласно вышеприведённому алгоритму построим граф
включения-сходства (при порогах δ ≥ 50% и δ ≥ 40%). Если между вершинами графа
включения наблюдаются стрелки, направленные в двух направлениях, то списки,
характеризующие эти вершины, будут сходны при заданном пороге (рис. 3). В нашем
случае при пороге δ ≥ 40% (рис. 3), списки под номерами 1, 3 и
5 включаются в список 4 по видовому составу. При необходимости можно построить
граф невключения, обратный приведённому графу включения.
В заключение раздела 3.4 хотелось бы остановиться на некоторых проблемах
упорядочения данных. Выделяют две основные проблемы: проблема качества
разбиения и проблема устойчивости разбиения. Для решения проблемы устойчивости
предлагают плюралистический подход (если разбиение повторяется при
использовании разных методов (Шитиков, Розенберг, Зинченко, 2005)),
бутстреп-метод, предложенный Б. Эфроном в 1977 г., метод «складного ножа» и
«скользящего контроля» (Количественные методы…, 1984). Несколько способов
предлагает В.Л. Леонов (Леонов, URL):
«исходную выборку случайным образом разделить на две примерно равные части,
провести кластеризацию обеих частей и затем сравнить полученные результаты.
Более трудоемкий путь предполагает последовательное исключение вначале первого
объекта и кластеризацию оставшихся (N - 1) объектов. Далее
последовательно проводя эту процедуру с исключением 2, 3 и т.д. объектов
анализируется структура всех N полученных кластеров. Другой алгоритм
проверки устойчивости предполагает многократное размножение, дублирование
исходной выборки из N объектов, затем объединение всех дублированных
выборок в одну большую выборку (псевдогенеральную совокупность) и случайное
извлечение из нее новой выборки из N объектов. После этого проводится
кластеризация этой выборки, далее извлекается новая случайная выборка и вновь
проводится кластеризация и т.д. Очевидно, что это также достаточно трудоемкий
путь».
Проблеме оценки качества разбиения также уделяют немало внимания.
Специалисты предлагают множество критериев оптимальности разбиения (Ward, 1963; Мандель, 1988 и др.), но
зачастую рекомендуют диверсифицировать исследования с помощью других методов. В
естествознании, кроме того, проблема обостряется вследствие континуальности
многих естественных объектов, а, следовательно, дискретного перехода быть не
может и структуру системы можно определить, только комбинируя методы. В общем,
качество кластеризации обычно определяется через некоторый функционал, т.е.
требуется при выборе кластеров найти оптимум (минимум внутрикластерных
расстояний и максимум внекластерных расстояний) функции (Загоруйко, 1999) или
через сравнение с реальными данными (если это возможно). Первый случай
рассматривается в рамках таких направлений как функциональный анализ,
распознавание образов, робастная статистика и др. Второй случай обычно
применяется при апробации новой методики классификации.
В биологии, как правило, определяется свойствами исследуемого объекта.
Например, объект континуальной природы по определению не может дать адекватного
разбиения.
1.4.10 Интерпретация результатов
Исходя из системного понимания территории и
взаимосвязей внутри объекта, исследователь может адекватно интерпретировать
полученные результаты. Из-за высокой сложности исследуемых объектов к процессу
исследования, как правило, подключаются специалисты разных направлений -
биогеографы, геоботаники, экологи, математики, программисты и др.
Рассмотренный индуктивный схемно-целевой подход
позволяет разрешить основные концептуальные, методологические и методические
проблемы, а также проблемы интерпретации полученных результатов, которые с
неизбежностью возникают при использовании математических методов.
В этом разделе стоит особо рассмотреть обобщённую
функцию «связующего звена» между вычислительными мощностями и прикладной
областью - аналитика. В нашем понимании, аналитик это специалист по формальным
методам решения задачи, т.е. прикладной математик, знакомый с предметной
областью пользователя. Последнее свойство аналитика очень важно, иначе он бы
просто не понимал бы конечного пользователя. Задачу пользователя,
сформулированную на естественном языке, аналитик представляет в математической
форме и разрабатывает алгоритм ее решения.
Составленную формализованную задачу и алгоритм ее
решения он передает программисту, который составляет программу решения этой
задачи на компьютере в виде текста на одном из компьютерных языков, вводит ее в
компьютер и отлаживает. Компьютерная техника решает задачу. Программист
расшифровывает результат, передает аналитику, и тот переводит решение на язык
конечного пользователя. По сути, аналитик является промежуточным звеном между
конечным пользователем (экологом, геоботаником, флористом и др.) и специалистом
в области информационных технологий (программистом).
Можно, разумеется, передать функции аналитика
конечному пользователю, чтобы он сам формализовал и программировал свою
задачу, работая непосредственно на компьютере, но очевидно, что этот путь
требует серьёзного и зачастую длительного обучения конечного пользователя. В
процессе обучения он должен научиться формализировать задачу, которую
собирается решить на ПК, т.е. выполнить функцию прикладного математика в своей
области и прикладного программиста, чтобы составить нужную программу. Именно
так зачастую приходится поступать сейчас конечному пользователю, если он хочет
решить свою задачу с помощью информационно-компьютерных технологий (ИКТ).
На первых этапах специалисту приходится тратить
слишком много дополнительного времени на формализацию своей задачи и
программирование, не говоря уже об отладке программы, на другие неизбежные
программистские заботы. Некоторые специалисты (Миркин, Наумова, Соломещ, 2002)
считают, что компьютер по большей части вредит специалисту: «компьютер не может
мыслить геоботанически (Вальтер, 1982)» и там же: «студент или аспирант,
который начал осваивать эколого-флористическую классификацию с компьютерной
обработки, никогда не станет натуралистом, т.к. будет отравлен «компьютерным
опиумом» в понимании Р. Маргалефа (1992)».
Многие разработчики математических методов в
прикладных областях ратуют за активное внедрение своих разработок и массовое
обучение специалистов. Разумеется, грамотное использование любых методов всегда
приветствуется, но одна из фундаментальных задач науки - оптимизация.
По-видимому, самым приоритетным направлением, на данном этапе, является
подготовка системных аналитиков и прикладных математиков, а также развитие
систем искусственного интеллекта (см. например, Шитиков, Розенберг, Зинченко,
2005).
Другое способ решения проблемы это «искусственный
интеллект», цель которого в формализации задачи технической системой с
последующим решением и представлением результата в виде, доступном пониманию
пользователя (т.е. реализация оптимального интерфейса). Несмотря на то, что
создание таких интеллектуальных компьютеров, работающих в определенной
предметной области, дело чрезвычайно сложное, на это придётся пойти, чтобы не
готовить из каждого специалиста профессионального программиста или
программирующего пользователя.
В заключение раздела отметим, что даже самые мощные
инструменты анализа не заменят специалиста, который способен грамотно поставить
задачу и выбрать чёткую схему исследования, т.е. любому специалисту на
современном этапе стоит выбрать «золотую середину» при использовании
вычислительных методов и технологий, а также всегда руководствоваться чёткой
схемой исследования.
Глава 2. Сравнительный анализ компонентов растительного
покрова
В настоящей главе мы решим ряд биологических задач с помощью методов на
основе схемно-целевого подхода. Большая часть первичных данных взята из
опубликованных работ по альгологии, флористике, геоботанике, экологии и др.
2.1 Флоры водорослей-макрофитов
дальневосточного побережья России
В данном разделе рассмотрена совокупность флор водорослей-макрофитов
российского Дальнего Востока. Нашей целью явилось разделение естественного
альгофлористического континуума на дискретные единицы (районирование), с
последующим анализом пространственной структуры с формальных позиций.
Материал взят из работ по сравнительной альгологии побережья Дальнего
Востока России (Гусарова, 1975; Гусарова, Сёмкин, 1982; 1986; Клочкова, 1997 и
др.). Огромная протяжённость и сильная расчлёнённость береговой линии
дальневосточных морей, их значительная меридиональная вытянутость, влияние
Азиатского материка, широкая связь с Тихим океаном, холодные и тёплые течения,
различные типы приливов, а также амплитуда приливно-отливных колебаний создают
предпосылки для развития здесь богатой и разнообразной альгофлоры макрофитов,
состоящей из арктических, бореальных и субтропических элементов.
С формальной точки зрения, сложность сравнительного изучения таких
конкретных альгофлор макрофитов побережий обусловлена тем, что первичные
флористические выделы существенно разновелики по числу видов, что не позволяет
при их сравнении адекватно определять сходство (Сёмкин, 2007). В этих случаях
необходимо использовать методы оценки несимметричных флористических связей,
основанные на мерах включения. При выделении сообществ литорали на основе
флористико-фаунистических признаков (Погребов, Горянина, 1988) было
установлено, что «выделяемые сообщества представляют собой более или менее
однородные части непрерывно изменяющейся биоты». Границы между ними нечёткие
(группировки переходят друг в друга, не образуя хорошо разграниченных зон, а
число сообществ зависит от избранного масштаба расчленения континуума). К
аналогичным результатам пришли А.И. Кафанов и В.Е. Жуков (1993), изучавшие
прибрежные сообщества водорослей-макрофитов залива Посьет, которые установили,
что «растительный покров литорали и верхней сублиторали выраженно континуален и
составлен определённым числом вертикальных поясообразующих флористических
группировок». Ситуацию «флористического континуума» (Юрцев, 1987) для флор
водорослей-макрофитов, вообще говоря, ещё не рассматривали. Дело в том, что для
таких исследований необходимы не только достаточно полные списки видов участков
побережий, но и расположение этих участков в виде географических рядов
соответствующих направлениям флористического обмена, причём целесообразно
обеспечение «примыкания» участков в рассматриваемых рядах. К районированию
побережья Дальнего Востока России обращались неоднократно. Использовались
методы «зонально-географического и альгофлористического фитогеографического
анализа» (Клочкова, 1997), причём, как отмечает Н.Г. Клочкова до её работы
«дальневосточная флора водорослей-макрофитов никогда не анализировалась как
единый комплекс». В полной мере для исследования флористического континуума
подходят данные Н.Г. Клочковой (1997). Автором было выделено, для
дальневосточных морей России (по гидрологическим, климатическим (границы
участков хорошо совпадают с положением средне августовских изотерм),
геоморфологическим, а также по палеогеологическим признакам), 40 участков
сублиторали, примыкающих друг к другу в географическом ряду (рис. 1 Приложения
Б), составлены списки видов водорослей-макрофитов (общий список включает 551
вид) и предварительно участки были объединены в четыре крупных района.
Нами была произведена математическая обработка общего видового списка
водорослей-макрофитов: рассчитана матрица пересечений (табл. 1 Приложения Б),
матрица мер включения (табл. 2 Приложения Б) и таблица индексов преобладания
(табл. 6 Приложения Б).
Проанализируем полученные результаты. Матрица пересечений приводится в
компактной форме в виде прямоугольной таблицы 40×21 (табл. 1 Приложения Б). При
рассмотрении значений главной диагонали матрицы пересечений можно увидеть, что
число видов на участках меняется от 32 видов (мыс Наварина-мыс Олюторский) до
269 видов (залив Петра Великого). Отметим, что полнота видовых списков
существенно возросла в сравнении со списками, составленными И.С. Гусаровой
(Сёмкин, Гусарова, 1986). Так, например, для о-ва Симушир список увеличился на
47 видов, для о-ва Парамушир на 37 видов, для о-ва Кунашир на 44 вида, для
Командорских о-вов на 81 вид. На Командорских островах по данным Н.Г. Клочковой (1997) составляет 209 видов водорослей-макрофитов.
На основе матрицы включения построено ориентированное
оптимальное дерево (дендрит) односвязывающим методом (Сёмкин, Клочкова,
Гусарова, Горшков, 2010 а). Структура графа наглядно показывает непрерывный
переход флор разных районов Северо-западной Пацифики в пределах бореальной
области (рис. 2 Приложения Б), причём односторонние связи весьма существенны
(80-90%). По схеме схеме зонально-географического членения К.Л. Виноградовой
(цит. по: Клочкова, 1997) бореальная зона в Тихом океане делится на две
подзоны: высокобореальную и низкобореальную. Высокобореальная зона выделяется
от Берингова пролива до мыса Терпения (о-в Сахалин), нижняя граница
низкобореальной зоны выделяется значительно южнее зал. Петра Великого (по
территории о-ва Хонсю).
Проведенное сравнение по общему списку видов
полученных нами районов и ранее выделенных Н.Г. Клочковой (1997) показывает
достаточно большую сходимость этих двух схем, но имеются некоторые различия. В
нашем случае в районе I появляется 11 участок (мыс Лопатка-мыс Утхолокский).
Этот участок является обеднённым дериватом флоры юго-восточной Камчатки и
видимо отделяется от II
района за счёт увеличения приливной зоны, нарастающего похолодания в
северо-западной части Камчатки (12 участок).
Во II-ом районе других изменений, кроме отмеченного,
нет. Условия обитания в данном районе формируются течениями, песчаными
грунтами, воздействием Амурского лимана.
Район III был нами разбит на два подрайона: III A (Курильские острова) и III B (Восточный Сахалин), причём первый
тяготеет к I району и включается через 10 участок
(Командорские острова) на 81% (данный участок является источником американской
флоры с видами преимущественно островного генезиса). По данным Гусаровой И.С.
(Гусарова, Сёмкин, 1986) также выделялся район, аналогичный III A, но с некоторыми расхождениями.
Четвёртый район изменений не претерпел.
Для выделения наиболее «оригинальных» и «банальных» по видовому составу
альгофлор по матрице мер включения можно построить ориентированный граф, но
графическое изображение не является наглядным из за большого числа дуг графа,
поэтому мы отразим структуру графа с помощью индексов преобладания (Сёмкин,
Куликова, 1981; Кафанов,1994; Сёмкин и др., 2009 б) в табл. 6 Приложения Б. Из
таблицы видно, что самой «банальной» альгофлорой является 3 участок (м.
Наварина - м. Олюторский), а самой «оригинальной» - 40 участок (залив Петра
Великого). Наиболее оригинальные флоры по районам (в скобках приведены значения
индексов преобладания): I - участки 10 (0,51) и 8 (0,38); II - 17 участок (0,33);
III A - участки 25 (0,44), 22 (0,38), 23 (0,36); III B - участки 28 (0,41); IV
- участки 40 (0,76), 30 (0,54), 38 (0,49), 39 (0,49), 31 (0,44), 37 (0,41).
Наиболее банальные флоры по районам (в скобках приведены значения индексов
преобладания): I - участки 3 (-1), 1 (-0,87), 5 (-0,49), 4 (-0,46), 2 и 11
(-0,41), 6 (-0,33), 7 и 9 (-0,23); II - участки 18 (-0,82), 13 (-0,62), 14
(-0,54), 12 (-0,51), 15 (-0,33), 16 (-0,13); III A - участки 21 (-0,03); III B
- участки 26 (-0,56); IV - участки 32 (-0,05). Наиболее оригинальные флоры
совпадают с основными центрами видообразования водорослей-макрофитов (Клочкова,
1997), кроме IV района, хотя очевидно, например,
высокое видовое разнообразие флоры 40 участка (зал. Петра Великого).
Макрофитобентос рассматриваемого района формируется представителями
зелёных (Chlorophyta), бурых (Phaeophyta) и красных (Rhodophyta) водорослей,
которые достаточно сильно различаются уровнем организации, жизненными циклами,
реакциями на условия среды и другими характеристиками. Известно, что
соотношение численности зелёных, бурых и красных водорослей зависит от широты,
и в альгофлорах разных районов Мирового океана оно достаточно устойчиво
(Feldman, 1937; Druehl, 1981, Segawa,
1962). В умеренных и относительно холодных водах по числу видов доминируют
багрянки (46%, в тёплых водах повышается до 58%), зелёные водоросли являются
самыми малочисленными (12-13%, в тёплых до 24%), бурые водоросли занимают
промежуточное положение (40-43%, в тёплых падает до 18%) (Жизнь растений…, 1977
: 361; Перестенко, 1982). В дальневосточных морях России (Клочкова, 1997)
наибольшим числом видов представлен отдел Rhodophyta - 319 видов (57,9%), за ним следует
отдел Phaeophyta - 159 видов (28,9%) и наименьшим по
числу видов является отдел Chloropyta - 73 вида (13,2%), т.е., в общем, подтверждается характер распределения
видов по отделам при значительном усилении Rhodophyta.
Исходя из существенных количественных соотношений водорослей-макрофитов в
рассматриваемых отделах и их различных флористических ролей, целесообразно
провести сравнительный анализ по видовым спискам для каждого отдела (Сёмкин,
Клочкова, Гусарова, Горшков, 2010 б). Привёдем матрицы пересечения (табл. 3-5
Приложения Б) и оптимальные деревья (рис. 3-5 Приложения Б). Для зелёных
водорослей (табл. 3 Приложения Б) наибольшее число видов (50 и 42)
определено в 40 участке (залив Петра Великого) и на 17 участке (Шантарские
о-ва) соответственно. Наименьшее число видов представлено на участке 3 (мыс
Наварина - мыс Олюторский) - всего 5 видов, т.е. на порядок меньше чем на 40
участке. По таблице 4 (Приложение Б) можно увидеть, что наибольшее число бурых
водорослей (78) определялось на 40 участке (залив Петра Великого), второе
место по числу видов занимает 30 участок (мыс Крильон - мыс Перепутье) - 64
вида. Наименьшее число видов обнаружено на участке 3 (13 видов). Максимально
насыщена красными водорослями (табл. 5 Приложения Б) флора залива Петра
Великого (141 вид), Командорских островов (128 видов) и участок 30 на юге
Сахалина между мысами Крильон и Перепутье (118 видов). Минимум устойчиво
определяется на участке 3 (14 видов).
Рассмотрим связи флор отделов Chlorophyta, Phaeophyta и Rhodophyta между районами по оптимальным
ориентированным деревьям. Как можно видеть из рис. 3 (Приложение Б) оптимальное
ориентированное дерево для зелёных водорослей имеет достаточно простую
структуру: в центрах находятся флоры Авачинского залива (участок 8), Шантарских
островов (17) и Приморья (38, 40). В эти флоры включаются флоры остальных
участков и формируются 4 флористических района. Район I включает флоры западного побережья Берингова моря, восточной
Камчатки, Командорских островов и юга западной Камчатки (участки 1-11). В район
II объединились флоры западного
побережья Камчатки, материкового побережья Охотского моря, севера Сахалина
(участки 12-18, 26). Самый маленький район (III) включает всего 3 участка (19-21) и объединяет флоры
северных Курильских островов (Парамушир, Симушир, Уруп). Район IV самый насыщенный по числу участков
(17) и по видовому составу флоры и объединяет флоры всего Приморья с заливом
Петра Великого, материковое побережье Татарского пролива, островов Большой
(Итуруп, Кунашир) и Малой Курильских гряд, острова Монерон и юга Сахалина.
Флоры западной Камчатки (участок 12) и острова Уруп (21) являются связующим звеном
между условно дискретными флористическими объединениями районов. Внутри районов
наблюдаются односторонние сильные (на уровне 100%), за редким исключением,
связи, особенно характерные для северных флор (I-III). В IV районе связи также односторонние, но
уровни несколько ослаблены (79-100%). Флоры зелёных водорослей северо-западной
Пацифики слабо дифференцированы и явно отличаются от структурирования флор по
общему списку видов (рис. 2 Приложения Б).
На рис. 4 Приложения Б приведено дерево для бурых водорослей.
Выделяется пять крупных дискретных единиц. Флоры северо-восточной части
российского сектора Тихого океана до Курильских островов и западной Камчатки до
мыса Баджедомова на севере объединились в I район (участки 1-12). Во II район включились флоры материкового побережья Охотского
моря, Шантарских островов и небольшого участка северо-западного Сахалина
(13-18). Флоры всех Курильских островов объединились в район IIIA (участки 19-25). Следующее
объединение (IIIB) включает флоры восточного, южного и
юго-западного Сахалина, а также острова Монерон (участки 26-31). В IV район объединились флоры всего
Приморья, материкового и северо-восточного побережий Татарского пролива
(участки 32-40). Внутри районов наблюдаются связи разных уровней (79-92%),
максимальные связи (96-100%) встречаются редко. Связь между районами (I-IV) осуществляются через флоры участка между мысом Ламанон и
мысом Китоуси (33), северо-восточного Сахалина и Амурского лимана (26), флору
участка между мысом Александра и мысом Лазарева (18), острова Парамушир (19) и
северо-западной Камчатки (12). В целом, флора более дифференцирована по
сравнению с флорой зелёных водорослей и районы объединяют преимущественно
участки, расположенные в географической последовательности. При сравнении
дендритов, построенных для конкретных флор по общему составу видов и по видам
бурых водорослей, наблюдаются некоторые расхождения в объединении участков по
полному списку видов. Так в дендрите для бурых водорослей флора 12 участка
(между бухтой Квачина и мысом Баджедомова) проявляет свойства переходного
района с равными значениями сходства 82% и к I району (11 участок) и к II району (17 участок). Это связано с
промежуточным температурным оптимумом для Phaeophyta между условиями
относительно тёплого 11 участка и более сурового по гидрологическому режиму и
мутности воды 13 участка. Флоры острова Монерон (29) и юго-западного Сахалина
(30-31) объединились с флорой района IIIB, но данный факт проинтерпретировать не удалось.
Флора красных водорослей формируется в 5 крупных дискретных
районов (рис. 5 Приложения Б), границы которых почти полностью совпадают с
границами районов, объединённых по общему списку видов. Исключением является
только флора самого южного участка западного Сахалина между мысом Крильон и
посёлком Перепутье, который включается во флору IV района по общему составу флоры, то в район IIIB по красным
водорослям. Причиной может являться температурная аномалия, известная как
«холодноводное пятно Макарова», которая мешает развитию тепловодных видов
(преимущественно красных водорослей). Связь между районами осуществляется через
флоры участков 3 (между мысами Наварина и Олюторским), 18 (между мысом
Александра и Лазарева), 20 (остров Симушир) и 32 (между мысом Слепиковского и
Штернберга).
Индексы преобладания для водорослей трёх отделов позволяют выявить
наиболее оригинальные и банальные флоры отдельных участков (табл. 6 Приложения
Б). Анализ индексов преобладания по отделам показывает, что многие флоры
северных участков (1-7, 9, 11-15, 18, 26) относятся к банальным по видовым
спискам всех трёх отделов. Это может быть связано с выраженным арктическим
влиянием (1-3 участки), недостаточной изученностью (3, 9, 13 участки),
опресняющим влиянием Амурского лимана (18 и 26 участки) и другими причинами.
Флоры Авачинского залива (8) и Шантарских островов (17) оригинальны по всем
трём отделами (0,05-0,38). Командорские острова (10) высокооригинальны по
красным водорослям (0,62), что объясняется связью с американской флорой
(Селиванова, 2004), но слабее по зелёным (0,38). Остальные участки,
расположенные к югу, имеют флору оригинальную на разном уровне (от 0,03 до
0,67) по видам всех отделов. На некоторых участках отмечена банальность флоры
по бурым водорослям (остров Парамушир), по зелёным (остров Уруп), по зелёным и
красным (участок между мысом Слепиковского и Штернберга). Самой оригинальной по
бурым и зелёным является флора залива Петра Великого (40 участок).
Поскольку абсолютная численность видов разных флор весьма различна и
отражает разный уровень их богатства, а косвенно - протяжённость пространства
занимаемого каждой флорой, представляется более целесообразным оперировать не с
абсолютными значениями, а с показателями соотношений между отдельными группами,
выраженными в процентах (или долях) от общей численности соответствующих флор
(Толмачёв, 1974). Исходя из этого, систематическая структура флоры
дополнительно определена на основе DS-спектров (относительные таксономические
спектры типа «отдел-вид»: D - divisio (отдел), S - species
(вид)) посредством расчёта меры процентного сходства и унарных мер
(разнообразия, концентрации). DS-спектры
характеризуют систематическую структуру крупных хорологических единиц рангов
флористической области и выше. В связи с этим, для флор водорослей-макрофитов
дальневосточного побережья морей России DS-спектры должны быть близки друг другу, а отклонения,
возможно, могут быть вызваны антропогенными воздействиями, недостаточной
изученностью флоры, слабой представленностью набора экотопов данного района или
переходным характером участка, тем самым, нарушая дискретность крупных
хорологических единиц (Сёмкин, Клочкова, Гусарова, Горшков, 2010 в).
Для определения DS-спектров
взяты диагональные элементы матриц пересечения видовых списков водорослей
соответствующих отделов. Сходство спектров флор двух районов будем определять с
помощью меры процентного сходства. В альгофлористических исследованиях на
основе DS-спектров рассчитывались различные
отношения между компонентами спектра. Наиболее часто используются отношения
между числом видов отделов Rhodophyta и Phaeophyta (R/P
или p3/p2), а также между числом видов Chlorophyta и Phaeophyta (C/P
или p1/p2), которые являются показателями
широтной принадлежности флор (Feldman,
1937; Клочкова, 1996; Клочкова, Березовская, 2001). Иногда используется (для
оценки экологического состояния водоёмов) флористический коэффициент, равный
сумме двух приведённых отношений (Cheney, 1977; Калугина-Гутник, 1989; Селиванова, 2004):
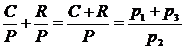 .
.
Можно
использовать также индексы концентрации, зависящие только от одной доли,
максимальной или минимальной (Berger, Parker, 1970; May, 1975; Песенко, Сёмкин,
1989; Сёмкин, 1987 а; Magurran, 1988; 2004) - соответственно 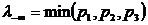 или
или 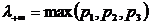 . В нашем
случае, почти у всех DS-спектров
. В нашем
случае, почти у всех DS-спектров 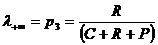 , т.е.
определяется как доля числа видов красных водорослей. Этот индекс не зависит от
числа высших таксонов (здесь отделов) и, как отмечает Мэй (May,
1975), является лучшей мерой видового разнообразия. Следует также отметить, что
индексы R/P, C/P, (C+R)/P определяются как отношения «часть к части» для
спектра, а индекс R/(C+R+P) есть отношение «части к целому» и потому является
более предпочтительным для определения отношений величин (Сёмкин, Горшков, Варченко,
2008; Semkin, Gorshkov, Varchenko 2009).
, т.е.
определяется как доля числа видов красных водорослей. Этот индекс не зависит от
числа высших таксонов (здесь отделов) и, как отмечает Мэй (May,
1975), является лучшей мерой видового разнообразия. Следует также отметить, что
индексы R/P, C/P, (C+R)/P определяются как отношения «часть к части» для
спектра, а индекс R/(C+R+P) есть отношение «части к целому» и потому является
более предпочтительным для определения отношений величин (Сёмкин, Горшков, Варченко,
2008; Semkin, Gorshkov, Varchenko 2009).
На
основе DS-спектров флор сублиторали 40 участков побережья ДВР
рассчитана матрица мер процентного сходства. DS-спектры всех
рассматриваемых 40 участков сходны на высоком уровне, несмотря на
разновеликость. Мера процентного сходства является оптимальной мерой для
нормированных дескриптивных множеств (здесь DS-спектров). Матрица мер
процентного сходства показывает, что DS-спектры всех рассматриваемых 40
участков сходны на высоком уровне. Например, DS-спектр 40 участка (зал. Петра
Великого) сходен на уровне 99% со спектрами участков 39, 35, 28, 20, 15, причём
эти участки разновелики по числу видов: 269, 189, 164, 189, 150 и 117
соответственно. Есть заметные отклонения: спектры арктических флор (1-3
участки) сильно отличаются от остальных дальневосточных флор (для всех спектров
значения сходства ниже в сравнении с другими спектрами), высокое сходство
(0,92-0,95) наблюдается только с 26 участком (северо-восточный берег о-ва
Сахалин, Амурский лиман), возможно, по причине опресняющего действия р. Амур.
Следует отметить, что два DS-спектра имеющих высокое сходство всегда равны или
близки по индексам разнообразия, но обратно, вообще говоря, неверно, т.е.
близкие или равные значения индексов разнообразия могут определяться для весьма
различных DS-спектров. В связи с этим, при сравнении DS-спектров
для резко изменяющегося флористического состава прибрежных сообществ при
естественных или антропогенных катастрофических воздействиях нельзя
использовать индексы разнообразия (Сёмкин, Клочкова, Гусарова, Горшков, 2010
в). Рассмотрим индексы разнообразия
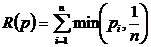 и
и 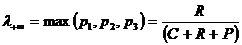 .
.
Индекс
концентрации λ+∞
принимает максимальные значения в случае, когда индекс разнообразия имеет минимальное
значение и наоборот. Отсюда следует, что все DS-спектры имеют
сходные значения индексов разнообразия и индексов концентрации (за редкими
исключениями). Например, наибольшие значения индекса разнообразия имеют
DS-спектры участков 2 (0,93), 26 (0,88), 17 (0,87), а наиболее выровненными по
отделам являются участки 10 (0,72), 29 (0,73), 14 (0,76), что слабо коррелирует
с представлениями о центрах видового разнообразия данного региона. Эти же
DS-спектры выделяются и по индексам концентрации и между ними определяется
наибольшее сходство.
Для
проверки интерпретируемости вышеописанных мер рассмотрим изменения
систематической структуры альгофлоры при антропогенном воздействии на примере
Авачинской губы на основе DS-спектров. Для этих целей возьмём 9 спектров,
характеризующих три части Авачинской губы: вся губа, горло и внутренняя часть в
различные годы (1970 (1-3 спектры), 1991 (4-6 спектры), 1999 (7-9 спектры) гг.)
по данным Н.Г. Клочковой и В.А. Березовской (2001) и два DS-спектра
флор в качестве контрольных эталонных районов: Авачинский залив до мыса
Безымянного на юге (10 спектр) и о-в Парамушир с прилежащими островами (11
спектр) из вышерассмотренных данных (табл. 7 Приложения Б). Ранее имела место
попытка оценить экологическое состояние Авачинской губы с помощью коэффициентов
(C+R)/P (Селиванова, 2004), но она не привела к успеху.
Однако оценка той же системы с помощью отношений C/P
и R/P позволила установить существенные изменения состояния
участков Авачинской губы в период с 1970 по 1999 гг. (Клочкова, Березовская,
2001).
Повторно
проведём обработку данных Н.Г. Клочковой и В.А. Березовской (2001), используя
меру разнообразия и меру процентного сходства. Рассмотрим полученные
результаты. Индексы разнообразия R(p) для отдельных районов будем обозначать как R(x), где x или y
- номер района (аналогично и для меры процентного сходства Jx,y).
Индексы разнообразия DS-спектров имеют наибольшее численное значение во
внутренней части Авачинской губы во все рассматриваемые годы, а также в горле и
во всей губе в 1999 г. Численные значения мер разнообразия колеблются от 0,85
до 0,89 (R(3) = 0,855; R(6) = 0,848; R(9) = 0,864; R(7) = 0,886; R(8) = 0,869). Для
остальных DS-спектров индекс разнообразия изменяется в пределах от
0,75 до 0,81, т.е. имеет значения близкие к эталонным DS-спектрам
(табл. 3). Отсюда следует, что наибольшей антропогенной нагрузке подвергались
участки внутренней части губы в разные годы, а в 1999 г. вся губа и горло. Из
расчётов также следует, что в 1970 г. DS-спектры всей губы (1) были
сходны с контрольными эталонными зонами (10, 11) более чем на 98% (J1,10 = 98,2%; J1,11 = 98,3%), горло
более чем на 95% (J2,10 = 95,5%; J2,11 = 95,1%).
Внутренняя часть губы уже значительно отличалась от эталонов (J3,10 = 93,9%; J3,11 = 94,3%). Это
подтверждается и исследованиями Н.Г. Клочковой и В.А. Березовской (2001),
которые утверждают, что уже в 1970 г. начался процесс полисапробизации. К 1991
г. сходство для всей губы и горла по-прежнему высокое (J4,10 = 98,0%; J4,11 = 98,3%; J5,10 = 98,0%; J5,11 = 97,6%). Для
внутренней части губы отмечается небольшой скачок в сторону увеличения сходства
(J6,10 = 94,6%; J6,11 = 95,0%). К 1999
г. происходит существенное изменение структуры DS-спектров во
внутренней части Авачинской губы (J9,10 = 72,5%; J9,11 = 72,8%), в горле (J8,10 = 92,5%; J8,11 = 92,9%) и в целом в губе (J7,10 = 90,3%; J7,11 = 90,6%). Разнообразие DS-спектров
внутренней части Авачинской губы в разные годы почти не менялось. Меры сходства
были попарно различны, кроме меры сходства (около 99%) между DS-спектрами
внутренней части губы в 1970 и 1991 гг. (J3,6 = 98,8%; J3,9 = 75,6%; J6,9 = 76,1%).
Наименьшее сходство всех DS-спектров наблюдается со спектром внутренней части
губы в 1999 г. (J1,9 = 72,5%; J2,9 = 68,6%; J3,9 = 75,6%; J4,9 = 72,7%; J5,9 = 70,7%; J6,9 = 76,1%; J7,9 = 82,2%; J8,9 = 78,9%; J10,9 = 72,5%; J11,9 = 72,8%) и,
следовательно, эта часть губы в большей степени была нарушена антропогенными
воздействиями. С 1970 по 1991 гг. существенных изменений в систематической
структуре Авачинской губы не обнаружено (J1,4 = 99,8%; J2,5 = 97,8%; J3,6 = 98,8%).
Значительные изменения отмечаются в период с 1991 по 1999 гг. (J4,7 =90,5 %; J5,8 = 90,5%; J6,9 = 76,2%).
В
общем, можно сказать, что методика сравнения флоры Авачинской губы с
контрольными участками по мерам процентного сходства, рассчитанных на основе DS-спектров
лучше интерпретируема для выделения этапов антропогенной деструкции
макрофитобентоса Авачинской губы и её частей. Предложенные нами математические
методы могут заменить ранее используемые методы Фельдмана (Feldman,
1937) и Чини (Cheney, 1977).
2.2 Субассоциации лесов поймы реки
Амур
В своей работе М.Х. Ахтямов (2001) исследует структуру приречных лесов
поймы р. Амур. Автором были рассмотрены особенности классификации
растительности приречных лесов поймы р. Амур, с помощью традиционного метода
Браун-Бланке выделяются синтаксоны прирусловых лесов (всего установлено - 1
класс, 4 порядка, 10 ассоциаций и 46 субассоциаций).
Цель данного раздела - с помощью метода непрямой ординации на основе
использования n-арных
показателей сходства подтвердить достоверность априорно формулируемых гипотезы
о выравненности описаний вдоль градиента увлажненности.
В таблицах, характеризующих субассоциации сообществ, обычно приводятся
такие данные, как число описаний сообществ (n), число видов в каждом описании (S) и общее число видов в субассоциации
(T), на основе которых можно рассчитать
меры сходства и различия n
описаний. М.Х. Ахтямов для характеристики субассоциаций по градиенту
увлажненности каждой ассоциации предлагает определить индекс биотической
дисперсии Коха (Kn-1) и
меру разнообразия Уиттекера (Iw), используя для их расчёта параметры n, S и T.
Первый индекс интерпретируется автором как мера однородности условий при
формировании видового состава насаждений, а второй индекс используется для
«количественной оценки степени дифференцированности ценофлоры числом полных
смен видов в экологическом диапазоне субассоциаций» (Ахтямов, 2001 : 52). При
этом следует отметить, что у автора наблюдается некоторое непонимание математического
смысла этих мер. Так, например, имеет место неверная интерпретация значений
этих индексов: «из-за большей выравненности условий увлажнения и характеристик
состава аллювиальных наносов в местообитаниях показатели сходства видового
состава (IBD) в рассматриваемых сообществах выше,
а значения IW - ниже» (Ахтямов, 2001 : 64). На самом деле индекс Коха
является мерой сходства, а индекс Уиттекера мерой различия серии описаний,
отсюда и различия в их значениях.
В
качестве примера корректного расчёта мер сходства и различия серии описаний
возьмём данные по ассоциации Salicetum udensis. Сообщества данной
ассоциации находятся в прирусловой части поймы и подвергаются весьма
длительному затоплению, что создает преимущества для развития влаголюбивых
видов на супесчано-суглинистых аллювиальных отложениях (Ахтямов, 2001).
Приведём расчёты в таблице 8 Приложения Б. Интерпретация полученных значений
мер вызвала затруднения у М.Х. Ахтямова: «причины более высоких значений Iw насаждений на этих стадиях в S.u.
swidietosum пока трудно интерпретировать» (Ахтямов, 2001 : 74).
Как можно видеть в таблице 8 Приложения Б на участках A4 и A5
происходит явный скачок значений, который невозможно объяснить с исключительно
биологических позиций, так как данные субассоциации являются рефренами, т.е.
формируются в сходных условиях и отличаются только признаку доминирования
различных видов ив. Очевидно, что в ряду не должно быть значительных колебаний
значений соответствующих мер. С нашей точки зрения, использовать указанные меры
для определения сходства или различия серии описаний нежелательно. Мера
разнообразия Уиттекера так сложно интерпретируется, потому что значения этой
«меры» не нормированы (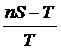 ), т.е. в формуле пропущен сомножитель (n-1) в знаменателе. Сложности возникают и с индексом
Коха, который можно использовать только в случае сравнения одинаковых по числу
описаний серий (n), следовательно индекс зависит от n
и при различных n производится расчёт по разным мерам K0,
…, Kn, значения которых определяются в разных шкалах.
), т.е. в формуле пропущен сомножитель (n-1) в знаменателе. Сложности возникают и с индексом
Коха, который можно использовать только в случае сравнения одинаковых по числу
описаний серий (n), следовательно индекс зависит от n
и при различных n производится расчёт по разным мерам K0,
…, Kn, значения которых определяются в разных шкалах.
Этих
проблем можно избежать, используя численные значения только одной меры сходства
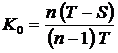 , т.к. мера K1 эквивалентна K0, а меры различия F0 и F1
коэквивалентны мерам сходства K0, и K1 соответственно (см.
таблицу). Сходство описаний согласно мере сходства K0 возрастает от субассоциаций S.u.
typicum (A1) к
субассоциации S.u. spiraetosum salicifoliae (A3), затем
в субассоциациях S.u. swidietosum (A4) и S.u. caricetosum appendiculatae (A5)
стабилизируется и возрастает к ассоциации S.u.
naumburgietosum thyrsiflorae
(A6).
, т.к. мера K1 эквивалентна K0, а меры различия F0 и F1
коэквивалентны мерам сходства K0, и K1 соответственно (см.
таблицу). Сходство описаний согласно мере сходства K0 возрастает от субассоциаций S.u.
typicum (A1) к
субассоциации S.u. spiraetosum salicifoliae (A3), затем
в субассоциациях S.u. swidietosum (A4) и S.u. caricetosum appendiculatae (A5)
стабилизируется и возрастает к ассоциации S.u.
naumburgietosum thyrsiflorae
(A6).
В
итоге получена хорошо интерпретируемая мера для целей непрямой ординации, т.е.
можно отметить отсутствие перепадов на участках A4 и A5, что согласуется с
априорными предположениями М.Х Ахтямова (2001).
2.3 Временнáя динамика сообществ
водорослей-макрофитов
Одним из важнейших направлений в биологии является изучение динамики
экосистем. Нахождение регулярной структуры во временнóм масштабе весьма редкое явление в
силу большого числа факторов необходимых для её возникновения. Фенологические
наблюдения ведутся в двух направлениях: наблюдения за неживой природой по
инструментальным метеорологическим данным (различают наступление климатических
сезонов), по изменениям физико-химических факторов водоёма (определяются
гидрологические сезоны) и др.; наблюдения за живой природой: биоиндикационные
исследования (дает нам информацию о биологических сезонах). Обширные данные
экологического мониторинга позволяют спрогнозировать наступление соответствующих
периодов (общая фенология) или же отметить изменение временнóй структуры сообществ (сравнительная
фенология), что может быть использовано в различных областях человеческой
деятельности: экономике, экологии, заповедном деле.
В данном разделе мы остановимся на биофенологических аспектах
исследования временнóй динамики сообществ и покажем возможности
теоретико-графовых методов для исследования временнóй динамики сообществ, а также оценим
возможности по выделению биологических сезонов на основе данных только по
видовому составу таксоцена водорослей-макрофитов с определёнными периодами
вегетации (такие данные также известны как фенологические спектры (Шенников,
1964)), что видимо ранее не производилось по причине, главным образом,
описательного характера фенологических наблюдений. Явление сезонности
подразумевает наличие определенной закономерности, связи некоторых периодов по
определенным критериям, причём периоды воспринимаются человеком дискретно (дни,
месяцы, сезоны и т.п.), но, по своей сути, являются континуальными. Цель -
исследование фенологической структуры таксоценов водорослей-макрофитов на
предмет выявления регулярной временнóй структуры, а также нахождение
соответствующего аналитического описания и графического представления
обнаруженных зависимостей.
В данном примере нас интересует возможность применения алгоритмов
построения оптимального графа к флористическим данным, причём объектами будут
списки видов соответствующих временных интервалов (в данном случае - месяцев).
Рассмотрим задачу построения оптимального графа для данных по временной
динамике экосистем. Для построения интересующего нас графа (кратчайшего пути)
требуется в матрице расстояний найти для каждого объекта (например, по строке)
наименьшее значение различия (исключая диагональный элемент). Определённые
таким образом объекты (столбец и строка) на графе соединяем дугой. Объекты не
должны образовывать цикла до тех пор, пока не будут пройдены все вершины графа
(получаем разновидность неориентированного графа). В качестве источника данных
о близости объектов нами будут использоваться матрицы различия (расстояния),
т.к. мы рассматриваем графы, для построения которых традиционно используются
расстояния. В частности, здесь будем использовать меру различия (расстояние)
Юрцева, которое является наиболее приемлемым для определения подобных отношений
(Сёмкин, 2007):
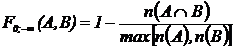 ,
,
где
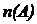 и
и 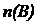 - число
видов в списке A и в списке B
соответственно;
- число
видов в списке A и в списке B
соответственно;  - общее число видов для списка A
и для списка B.
- общее число видов для списка A
и для списка B.
Для
расчётов взяты литературные данные по вегетации в течение года
водорослей-макрофитов залива Восток (Японское море), проведённые И.С. Гусаровой
(1988) и В.Ф. Макиенко (1975) и бухты Витязь проведённые А.И. Кафановым и В.Е.
Жуковым (1993).Залив Восток и бухта Витязь очень хорошо изучены (Коженкова,
2008) и (Суханов, Жуков, 2010) соответственно, что позволяет успешно
использовать их для моделирования.
В
работе И.С. Гусаровой (Гусарова, 1988) были приведены данные по периодам
вегетации видов водорослей-макрофитов: всего 111 вегетирующих видов, из них 13
зелёных, 33 бурых и 65 красных. Для оценки отношений мы взяли таблицу, в
которой представлены периоды вегетации водорослей. Отдельно отметим виды
вегетирующие круглый год (константные виды). На основании первичной
матрицы для периода с января (I) по декабрь (XII) нами была
рассчитана матрица пересечений (табл. 9 сверху Приложения Б). Данные матрицы
указывают на значительную равновеликость рядом расположенных диагональных
элементов, что позволяет применять для них методы, определяющие отношения
сходства (различия).
Последовательно
(согласно детерминистской схеме (Сёмкин, Орешко, Горшков, 2009 а, б; 2010))
рассчитываем матрицу включения и симметричную матрицу различия (расстояний) мер
Юрцева (табл. 9 снизу Приложения Б). Используя данные матрицы мер различия
Юрцева построим граф по алгоритму получения кратчайшего пути (рис. 7 Приложения
Б). В результате нами был получен оптимальный (гамильтонов) цикл. В теории
графов получение гамильтонова цикла является архисложной задачей (эта задача
известна также как задача коммивояжёра, т.е. задача построения цикла с учётом
весовых значений). В литературе имеется опыт решения таких задач методом ветвей
и границ (например, Рейнгольд, Нивергельт, Део, 1980). В нашем случае «природа
сама решила» её, т.е. нам удалось выявить цикл связных объектов (месяцев)
формальными (математическими) методами. Отметим некоторые сложности, которые
возникли при построении данного оптимального графа. Объект XI
тяготеет к месяцу III в большей степени (28.6%), чем к соседнему XII
(31.3%), но III месяц уже включен в граф между II
(16.9%) и III (7.8%), т.е. значения расстояния меньше.
Следовательно, получаем последовательную связность компонентов годового
цикла. Оценим влияние константных видов на общую временную структуру
экосистем залива. Для этого мы удалим из первичных данных константные виды.
Рассчитаем матрицу пересечений (табл. 10 сверху Приложения Б), матрицу
расстояний Юрцева (табл. 10 снизу Приложения Б) и построим граф по алгоритму
кратчайшего пути (рис. 8 Приложения Б). Отметим, что матрица пересечений для
полного списка и матрица пересечений без константных видов связаны простой
зависимостью:
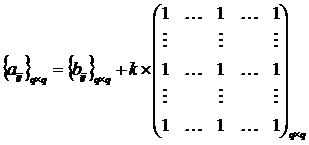 ,
,
где
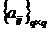 - матрица пересечений для полного списка;
- матрица пересечений для полного списка; 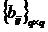 - матрица пересечений без константных видов; k
- число константных видов; q - число списков. Или, говоря проще, для того чтобы
нам из матрицы пересечений без константных видов вновь получить матрицу
пересечений с полным списком видов надо к каждому элементу первой матрицы
прибавить число равное количеству удалённых из первичной матрицы константных
видов. Анализ оптимального графа (рис. 8 Приложения Б) показывает, что
константные виды размывают представление о близости объектов, уменьшая значение
метрического расстояния. Полученный граф изоморфен предыдущему.
- матрица пересечений без константных видов; k
- число константных видов; q - число списков. Или, говоря проще, для того чтобы
нам из матрицы пересечений без константных видов вновь получить матрицу
пересечений с полным списком видов надо к каждому элементу первой матрицы
прибавить число равное количеству удалённых из первичной матрицы константных
видов. Анализ оптимального графа (рис. 8 Приложения Б) показывает, что
константные виды размывают представление о близости объектов, уменьшая значение
метрического расстояния. Полученный граф изоморфен предыдущему.
В
работе В.Ф. Макиенко (1978) исследования проводились в течение периодов:
июнь-декабрь 1971, апрель-ноябрь 1972, февраль-март и май-август 1973 годов. В
первичной таблице представлены: зеленые (10 видов), бурые (19) и красные (37)
водоросли-макрофиты. Всего 66 видов. Произведём аналогичные расчёты для полного
списка (табл. 11 Приложения Б) и граф (рис. 9 Приложения Б). Удаление
константных видов (21 вид) даёт характерную картину изоморфной структуры с
явным гамильтоновым циклом. Как и в предыдущем случае меняется лишь абсолютное
значение расстояния, при сохранении масштаба близости.
Анализ
графов по заливу Восток позволяет достаточно явно выделить весенне-летний (III-VII),
осенний (VIII-XI) и зимний (XII-II) периоды (по
данным И.С. Гусаровой); весенне-летний (III-VI),
летне-осенний (VII-X) и зимний (XI-XII) периоды (по
данным В.Ф. Макиенко). Распределение объектов близко, но есть расхождения.
Возможно, они обусловлены выпадением января (I) в данных В.Ф.
Макиенко, а также тем, что в работе Гусаровой И.С. приведён дополненный видами
список (были добавлены виды, найденные в заливе Восток за период с 1975 по 1988
гг.).
По
водорослям-макрофитам бухты Витязь (Японское море) взяты исследования А.И.
Кафанова и В.Е. Жукова (1993) за период с июня 1979 г. по август 1980 г. Был
обнаружен 101 вид водорослей (по данным (Суханов, Жуков, 2010) видовый состав
близок к насыщенному - 103±3): 15 зеленых, 35 бурых и 51 красных. Авторами была
подробно изучена фенологическая структура таксоцена макрофитов бухты на основе
теоретико-графовых методов: построены дендрограммы, ориентированный граф
включения. В результате: 1) были выделены четыре периода: зимний, весенний,
летний и осенний; 2) выявлено несовпадение биологических и гидрологических
сезонов. Мы, по аналогии с предыдущими примерами рассчитали матрицу пересечения
(табл. 12 сверху Приложения Б) и матрицу расстояний Юрцева (табл. 12 снизу
Приложения Б). Значения диагонали матрицы пересечений равновелики, не считая
некоторых разрывов вследствие выпадения месяцев III и XI.
На основе матрицы различия построим граф (рис. 10 Приложения Б).
Рассмотрим
спорные ситуации. Объект XII на 26.8% тяготеет к I и II
объектам. Включение II месяца в XII больше (93.8%) чем включение I
месяца в XII (90.9%). Кроме того, в спорных моментах более сильным
считается включение в меньший по числу видов список. Здесь это снова II
месяц. Если убрать константные виды (26 видов), то замкнуть цикл не
представляется возможным (табл. 13 Приложения Б) и мы можем получить лишь
оптимальный путь (рис. 11 Приложения Б).
Объект
VIII тяготеет к VI на уровне 37.2%, также как и V к VI,
но в V месяце меньше видов. Объект X ближе к VII, VI, V (но
они тяготеют больше к соседним месяцам). Наиболее проблемным является XII
месяц: он тяготеет к IX месяцу, хотя ближе к соседям, но его близость к IV
месяцу (на уровне 62.5%) разрушает цикл, потому что IV месяц
тяготеет ко II только на 79.2%. Даже если отбросить эту проблему, то
ситуация осложняется еще и тем, что XII тяготеет одинаково к I и к
II месяцу, как и в случае полного списка.
Очевидно,
что разрыв цикла связан с недостатком данных по видовому составу III и
XI месяцев. Причём, более явно это влияние проявляется
без размывания уровня близости константными видами.
Глава 3. Биоинформационная система «Биоразнообразие»
В данной главе мы непосредственно обратимся к такому научному направлению
как биоинформатика. В основе биоинформатики лежит понятие «биологической
информации», которая обладает рядом весьма специфических свойств. Основной
объект нашего интереса это компоненты биологического разнообразия, а в
частности компоненты растительного покрова (виды растений, растительные
сообщества, спектры, биоморфы и т.д.). «Биологическое разнообразие не есть
только простое множество объектов разного типа, но это внутреннее свойство
системы, которое раскрывается нами с помощью разных на нее взглядов, создания
разных моделей фитоценоза. И позволяет это делать объективно существующее
разнообразие качеств, свойств, сторон растительных организмов и наши различные
представления о наиболее функционально значимых из них» (Ганнибал, 2007).
Понятие «биологическое разнообразие» за весьма короткий отрезок времени
получило большое количество толкований. В биологическом смысле рассматриваются
представления о генетическом, видовом и ценотическом и надценотическом
разнообразии жизни. Однако в дальнейшем, сначала деятели охраны природы, а
затем и ученые стали говорить об экосистемном и ландшафтном разнообразии как
объектах сохранения, а, соответственно, выделения в природе и изучения (см.
например, Юрцев, 1991; Бочарников, 1998; География…, 2002; Сохранение…, 2002;
Шитиков, Розенберг, Зинченко, 2005). В 1992 году на Конференции ООН по
устойчивому развитию была принята «Конвенция о биологическом разнообразии», к
которой присоединилось большинство стран на планете (в том числе Россия), а,
следовательно, сложнейшая и многообразнейшая проблема биоразнообразия приобрела
еще и политическое звучание (География…, 2002). Большинство исследователей
сходятся во мнении, что словосочетание «биологическое разнообразие» впервые
применил Г. Бэйтс в 1892 г. в работе «Натуралист на Амазонке» (рус. перевод:
Бейтс, 1958), когда описывал свои впечатления от встречи около 700 разных видов
бабочек за время часовой экскурсии (см. например, Лебедева, Дроздов,
Криволуцкий, 1999; Протасов, 2002; Шитиков, Розенберг, Зинченко, 2005). Однако
основные научные концепции биоразнообразия были сформулированы лишь в середине
ХХ века, что напрямую связано с развитием количественных методов в биологии.
Особенно отметим работы Р. Уиттекера (Whittaker, 1970; 1972), в которых была предложена организация уровней
экосистемного разнообразия и исследованы зависимости биоразнообразия от
факторов окружающей среды. В дальнейшем идеи Р. Уиттекера были развиты и в
настоящее время схематически совокупность уровней биоразнообразия F(R)
можно представить в следующем виде (География…, 2002):
Всё
это типологическое многообразие сводится к двум типам разнообразия -
инвентаризационному, т.е. разнообразию внутри системы и дифференцирующему, т.е.
разнообразию между системами. Инвентаризационное разнообразие обычно
оценивается с помощью унарных индексов, а дифференцирующее с помощью n-арных
(чаще бинарных) мер. Здесь нижний ряд греческих букв обозначает инвентаризационное
разнообразие. Например, для растительного покрова: w(омега) - разнообразие
планетарной флоры; c(хи) -
разнообразие флористического царства; u(ипсилон) - разнообразие флористической области; s(сигма) -
разнообразие флористической провинции; p(пи) - разнообразие флористического округа; x(кси) -
разнообразие флористического района; m(мю) - разнообразие конкретной флоры. Верхний ряд букв
соответствует дифференцирующему разнообразию. Для растительного покрова
соответственно: y(пси) -
разнообразие между флористическими царствами в пределах флоры планеты; f(фи) -
разнообразие между флористическими областями в пределах царства; t(тау) -
разнообразие между флористическими провинциями в приделах флористической
области; r(ро) -
разнообразие между флористическими округами в пределах флористической
провинции; o(омикрон) -
разнообразие между флорами районов в пределах флористического округа; n(ню) -
разнообразие между конкретными флорами в пределах флористического района.
Исследование различных аспектов биоразнообразия является одним из
магистральных направлений концепции устойчивого развития (Loreau et al., 2001). Устойчивое развитие определяют как управляемый
процесс коэволюционного развития природы и общества. Как деятельность,
устойчивое развитие нуждается в поддержке мощных информационных баз данных по
экологическим, социально-демографическим, производственно-экономическим
факторам и тенденциям их развития. На основе этих данных строятся модели
экономического благополучия и проводятся научные исследования. Здесь мы
сталкиваемся с рядом проблем. Во-первых, объемы первичной информации для
устойчивого развития территорий, даже локального уровня, очень велики (Крестов,
2001; 2005; 2006; Сёмкин, Гарбузова, Майоров, Горшков, 2009). В связи с этим
проявляется проблема накопления, целенаправленного структурирования и обработки
колоссального объема информации (Горшков, 2008). Во-вторых, существует
проблема получения качественно новой информации на основе первичной (здесь
следует отметить, что на данный момент накопилось большое количество
тематической информации в различных форматах) (Остроумова, 2000; Батурина,
Чернобаева, 2007). Анализ информации невозможен без адекватного математического
аппарата и на его разработку в различных областях тратится «львиная доля»
ресурсов. В-третьих, необходима единая концептуальная схема,
объединяющая этапы сбора, обработки, анализа данных и прогноза.
В настоящее время существует несколько попыток реализации информационных
систем с экологическим наполнением. Как правило, подразумевается создание
некоторой ЭИС (экологическая информационная система) (Основы геоинформатики…,
2004; Шитиков, Розенберг, Зинченко, 2005).
Для решения этих проблем нами предлагается использовать биоинформационную
систему «Биоразнообразие», основные идеи которой представлены нами в ряде работ
(Горшков, 2008; Сёмкин, Орешко, Горшков, 2005 а, б) и на Интернет-ресурсе,
посвящённом сравнительному анализу в биологии (www.biocomparison.ucoz.ru). Биоинформационная система (БИС) «Биоразнообразие»
используется для целей сравнительного анализа флор, фаун, сообществ и других
биологических объектов. БИС содержит математическое обеспечение для анализа
компонентов биоразнообразия на уровне видов, ценозов и сообществ. Структура БИС
«Биоразнообразие» нами определена как совокупность трёх основных подсистем.
1. Базы данных, содержащие компоненты биоразнообразия;
2. Программы для обработки компонентов биоразнообразия;
. Информационно-справочная система по биоценометрии.
3.1 Базы данных, содержащие
компоненты биоразнообразия
Экологи и биологи в процессе научно-практической деятельности собирают
большое количество фактического материала, характеризующего различные аспекты
жизнедеятельности животного и растительного мира. В настоящее время большая
часть этого оригинального материала находится в виде бумажных архивов и не
представляет собой организованной информационной среды, которая является
необходимой основой для современных научных исследований.
Первичные данные, переведённые в электронный вид (нами для расчётов
используется формат Excel), являются
информационной основой БИС «Биоразнообразие». С использованием баз данных по
компонентам биоразнообразия и специализированных компьютерных программ
появилась возможность сравнивать фитохории рангов провинций и областей, при
этом обрабатывать видовые списки размером в несколько тысяч (Крестов, 2001;
2005; 2006; Малышев, 2002). Базы данных содержат информацию по различным
компонентам биоразнообразия (списки видов животных и растений, таксономические
спектры, экологические спектры, географические спектры и др.). Кроме того, базы
данных могут содержать обобщённую информацию в виде матриц пересечений,
включения, сходства и др. Зачастую исследователи не выдают своих первичных
данных, накопленных с большим трудом до тех пор пока сами не получат из них всю
возможную информацию. Но здесь возникает проблема воспроизводимости результатов,
т.е. такие исследователи публикуют данные о закономерностях, полученных из
«чёрного ящика», в который никто кроме них самих не может заглянуть и оценить
адекватность выявленных закономерностей а, следовательно, теряется сам смысл
таких публикаций и выводы автора приходится принимать на веру. Возможно,
компромиссным вариантом будет публикация матриц пересечения, которые позволяют
производить обработку данных, но не требуют представления первичных данных в
открытой печати.
Для обработки открытых первичных данных необходимо создать
биоинформационную систему, содержащую первичные данные (в виде электронных
таблиц) и всю необходимую информацию для проверки предложенных в статье методов
обработки данных. При налаживании быстрых информационных коммуникаций (Шокин,
Федотов, 1998; Мещеряков, Хомоненко, 2001) можно произвести расчёты по
первичным данным и установить достоверность выводов (воспроизводимость в
широком смысле слова). Современные информационные технологии в настоящее время
позволяют приступить к широкомасштабному переводу накопленной информации в
электронную форму и созданию принципиально новых видов информационных ресурсов,
какими, например, являются биологические базы данных, электронные коллекции и
т.п. (Шокин, Федотов, 1998).
Но из-за сложности формализации выбора описания возникают принципиальные
затруднения: «на пути к эффективной компьютеризации процесса обработки
геоботанических материалов» (Боряков, Воротников, Борякова, 2005). В первую
очередь необходимо определиться с целью использования данных. Если их функция
сугубо информирующая (сводные списки) и не подразумевает обработки, то тогда
автор будущей базы данных выбирает максимально простой текстовый формат, в
дальнейшем его можно будет массово представить с помощью Интернет-технологий
(более сложным вопросом будет представление этих данных с помощью полноценной
базы данных). Если исследователь желает обрабатывать эти данные, то здесь
необходимо обратиться к редакторам электронных таблиц. Обзор существующих
программных средств данной специализации показывает, что существуют
коммерческие продукты Lotus
1-2-3, Quattro Pro и Excel
(явный доминирующий продукт рынка) и некоммерческие редакторы, представляющие
компоненты офисных пакетов «с открытым кодом» Open Office и Star Office. Отметим,
что некоторые современные пакеты математических и статистических программ имеют
табличную структуру (почти без возможности применения формул) и, зачастую,
совместимы с MS Excel. Кроме того, в MS Excel предусмотрена возможность достаточно мощной обработки
данных, которые усиливаются возможностями встроенного языка программирования VBA, что по достоинству оценили и
биологи (Новаковский, 2006).
На данный момент уже накоплено некоторое количество данных в
информационных системах различных уровней, связанных с экологическими
исследованиями (Бочарников, 1998; Bisby, 2000; Edwards, Lane, Nielsen, 2000; Федотенков, 2005). Отметим основные
и наиболее крупные информационные системы, содержащие информацию о компонентах
биоразнообразия:
1. Информационная система GRID (Global Resource Information Database, Глобальная база данных о ресурсах).
Создана UNEP. В задачи GRID входит организация,
анализ и хранение данных об окружающей среде, полученных из различных
источников. В системе используется информация, собираемая в результате
деятельности ГСМОС (системы глобального мониторинга) при осуществлении
различных программ UNEP, находящаяся в распоряжении международных и
национальных, правительственных и научных организаций. Конечной целью сбора и
анализа данных является получение информации, которая может использоваться для
принятия решений. Данные в GRID хранятся электронном виде в формате
геоинформационной системы ArcInfo.
2. Информационная система ИНФОТЕРРА. Также создана UNEP для содействия международному обмену
экологической информацией.
. Программа по исследованию биоразнообразия Diversitas (Dirzo, Loreau, 2005).
4. Информационная
система BioDat. Пример известной российской информационной системы
экологической направленности (http://www.biodat.ru).
Несмотря на впечатляющие вычислительные возможности, огромные финансовые
и человеческие ресурсы, брошенные на решение этих задач (Redfearn, 1999), в
настоящее время не существует адекватной концептуальной схемы создания базы
данных компонентов биологического разнообразия, которая бы использовалась
повсеместно (Горшков, 2008).
3.2 Программы для обработки
биологических данных
Биологи и прикладные математики имеют достаточно большой опыт
использования пакетов статистических и математических программ в обработке
своих данных (Новаковский, 2006). Более последовательным является рассмотрение
используемых в настоящее время программных продуктов и публикаций, которые
подтверждают использование этих программ. Очевидно, что пользователи склонны
использовать те методы и критерии, которые представлены в популярных
статистических и математических пакетах (Пузаченко, 2004; Шитиков, Розенберг,
Зинченко, 2005).
Рассмотрим
основные программные продукты, применяемые в области обработки экологических
данных. Весьма условно можно разделить их на математические и статистические
пакеты. Несколько в стороне находятся средства визуального представления
зависимостей, т.к. многие крупные пакеты включают в себя, кроме прочего,
возможности графического представления данных. Большая часть математических
программ обеспечивает вычисления в области линейной алгебры и
интегрально-дифференциального исчисления, численных методов и математического
моделирования. Можно выделить из коммерческих Mathematica, Maple, Mathcad,
Matlab (есть свободные аналоги - Scilab и Octave); из некоммерческих - Axiom, Maxima, Sage.
Основные проблемы таких программ для биологов в сложности интерфейса (как
правило, работа с такими программами напоминает программирование) и в
ориентированности на теоретические конструкции математики (перечислены выше),
адаптировать которые пользователю-биологу под свои задачи предлагается
самостоятельно. Пакеты статистических программ значительно проще для конечного
пользователя, т.к. зачастую обладают интуитивным интерфейсом. Это пожалуй,
самый популярный статистический пакет Statistica. Пакет содержит весьма
солидный перечень аналитических процедур и активно используется многими
исследователями (Шитиков, Розенберг, Зинченко, 2005). Модуль «Кластерный
анализ» предлагает весьма ограниченный перечень мер близости, представленный
только мерами различия, построить какие-либо графы кроме дендрограммы не
представляется возможным. Пакет SPSS (<http://www.spss.com/>) является
одной из старейших систем статистического анализа данных и основном конкурентом
пакета Statistica. Намного реже используются: SAS, S-plus,
NCSS, Statgraphics. К свободным продуктам относится язык
программирования для статистической обработки R и его
оболочки.
Отметим специализированные программы для построения графов. К наиболее
удачным относятся коммерческие Ilog, GoView, Lassalle AddFlow, LEDA (есть
бесплатная редакция
(http://www.algorithmic-solutions.com/leda/ledak/index.htm)). Из бесплатных
можно отметить Boost Graph Library (http://www.boost.org/). Для визуализации
графов можно использовать Graphviz (http://graphviz.org/) (по мнению экспертов,
она хорошо работает для орграфов) или LION Graph Visualizer
(http://lion.disi.unitn.it/intelligent-optimization/visualizer.html). Особо
отметим программу «Графоанализатор» (http://grafoanalizator.unick-soft.ru) -
это русскоязычная программа, с весьма простым пользовательским интерфейсом.
Также рассмотрим специальные программы для биологических расчётов. В той
связи интересна обзорная работа А.Б. Новаковского (Новаковский, 2006), в
которой рассматриваются современные программы для обработки геоботанических и
экологических данных. Множество ссылок дано на сайте В.П. Леонова
(http://www.biometrica.tomsk.ru/list/other.htm), программы учитывающие
специфику сравнительного анализа в биологии (http://www.exetersoftware.com),
огромное количество программ для филогенетических расчётов можно найти на
Интернет-портале Вашингтонского университета (http://evolution.genetics.washington.edu), тайваньскими специалистами создана программа Spade (http://chao.stat.nthu.edu.tw). Подобных
«малых» программ, созданных для частных целей, довольно много, но широкое
распространение получили лишь немногие. Многие исследователи отмечают, что для
биолога весь этот огромный перечень сложнейших методов недостаточен. Редко в
каком-нибудь пакете можно найти расчёт меры Жаккара или меры Сёренсена
(например, Graphs, NTSYS, BIODIV, PAST и др.). Но при использовании классов
мер смысл приведения огромного количества мер теряется.
По причине того, что математический аппарат, предложенный нами для оценки
компонентов биоразнообразия ещё не получил должного распространения, ни в одном
из рассмотренных программных продуктов не может быть соответствующих процедур.
Для решения этой проблемы нами предлагается программа «BioComparison» (замена ранее объявленной в наших
публикациях, но не законченной программы «Биоценометрия»), которая
предназначена для сравнительного анализа биологических данных (Горшков, 2010).
Алгоритмы программы основаны на изложенном в данной работе математическом
аппарате и реализованы с помощью языка VBA (Visual Basic for Applications) в среде MS Excel 2003.
Возможности программы позволяют произвести сравнительный анализ
биологических объектов, имеется возможность графической визуализации (программа
и инструкция к ней выложена на сайте www.biocomparison.ucoz.ru).
Программа «BioComparison» имеет ряд блоков (рис. 1 Приложения
В):
1. Блок «Матрицы». Состоит из трёх вкладок:
· Вкладка «Бинарные» (рис. 2 Приложения В). Позволяет
рассчитать матрицы пересечений, включения, сходства, различия, причём расчёт
может начинаться с матриц наблюдения, пересечений, включения, сходства,
различия. Предусмотрено задание индексов τ и η.
· Вкладка «Многоместные» (рис. 3 Приложения В). Позволяет
рассчитать первичные компоненты n-арных мер (S,
T, n) по матрице наблюдений; возможен расчёт индекса Коха,
K0 и F0.
· Вкладка «Унарные» (рис. 4 Приложения В). Позволяет
рассчитывать матрицу долей из матрицы значений. Возможен расчёт мер семейства
Реньи, семейства Хилла и семейства Колмогорова; абсолютной и относительной мер
разнообразия R(p), индекса концентрации Бергера-Паркера, коэффициента
вариации и его нормированной формы, меры диссонанса Розенберга.
2. Блок «Несимметричные графы» (рис. 5 Приложения В). Позволяет (при
заданном пороге) построить ориентированный граф (на отдельном листе) или
рассчитать таблицу индексов преобладания на основе матрицы включения.
Программа «BioComparison»
будет развиваться, и дополняться в дальнейшем, реализуя новейшие разработки в
области теории близости. Все расчёты в данной диссертационной работе были
произведены полуавтоматическим способом: в среде MS Excel 2003 рассчитывались матрицы, с помощью графических
редакторов создавались графы. Все примеры, приведённые в разделе 4, были
вторично использованы для всестороннего тестирования программы «BioComparison».
3.3 Информационно-справочная система
по биоценометрии с комментариями
Ранее было показано, что во многих работах по сравнительной флористике,
количественной экологии и других научных направлениях используются различные
показатели близости для обработки данных и методы определения отношений в виде
графов, дендрограмм и др. Обзоры по использованию методов сравнительного
анализа в биологии имеют методологические или методические ошибки (Песенко,
1982; Legendre, Legendre, 1983; Kershaw, Looney,
1985; Digby, Kempton, 1987; Мэгарран, 1992; Уланова, 1995; Magurran, 2004; Шитиков, Розенберг, Зинченко,
2005). Для консультационных целей и целей обучения нами предлагается
информационно-справочная система с комментариями.
Информационно-справочная система по биоценометрии как часть БИС
«Биоразнообразие» реализована на основе базы библиографических данных
Тихоокеанского института географии (на основе MS Access 2003), созданной А.П. Орешко (рис. 6-8 Приложения В).
Предложенная нами информационно-справочная система имеет и ряд особенностей
(Сёмкин, Орешко, Горшков, 2005 а, б). В частности это наличие рефератов с
комментариями по предметной области (рис. 8 Приложения В). В комментариях к
источникам отмечены следующие компоненты: 1) Особенности данного источника и
его вклад в общую теорию мер близости; 2) Ошибки (если таковые имеются) и их
корректировка; 3) Представление формул в современных теоретико-множественных
обозначениях. Пользователь имеет возможность сортировать список источников в
системе по автору, названию, году издания и ключевым словам (рис. 7 Приложения
В). Окна для выбора критерия сортировки позволяют производить иерархическую
сортировку в различном порядке (рис. 6 Приложения В). При этом можно задавать
выполнение всех фильтров сразу, либо любого из фильтров. По каждому фильтру
можно строить логическую структуру с использованием операторов «И» и/или «ИЛИ».
Для любого фильтра можно задавать любую его часть. Это удобно в случае, если
пользователь не знает точного написания поискового значения. В конце списка
источников имеется две гиперссылки: «реферат» и «доп. реферат». Щелчок левой
клавишей мыши на первой ссылке приведет к открытию реферата в отдельном окне,
одном и том же для разных изданий. Щелчок на второй гиперссылке приведет к
открытию реферата в новом дополнительном окне. Это дает возможность сравнивать
рефераты между собой. Планируется регулярное дополнение базы данных новыми
источниками, сопровождаемых комментариями, а также интеграция базы с Интернет.
В настоящее время информационную функцию на себя частично взял сайт Горшкова
М.В. на платформе Ucoz (www.biocomparison.ucoz.ru).
Информационно-справочную систему по биоценометрии с комментариями
целесообразно использовать для обучения студентов и аспирантов в области
биоценометрии, а также для консультационной помощи специалистам различных
научных направлений, связанных с биоценометрией, биоинформатикой и
математической биологией.
Заключение
В настоящей диссертационной работе, на основе схемно-целевого подхода
нами решены некоторые методологические и методические задачи математической
биологии:
1. Представлена схема сравнительного анализа биологических объектов,
с аксиоматическими основаниями использования мер близости (сходства,
совместимости, взаимозависимости, включения и двойственных им мер), определены
основные классы мер близости и на их основе предложена для использования только
одна (оптимальная) мера для каждого класса мер, а как следствие и для каждого
класса формализованных биологических объектов. С учётом приведённых методологических
принципов создана биоинформационная система «Биоразнообразие», в частности
автором посредством языка VBA в
среде MS Excel адаптированы и реализованы основные математические
алгоритмы в виде программы BioComparison.
2. Уточнены границы некоторых районов дальневосточного побережья РФ
по видовому составу водорослей-макрофитов, отражена роль зелёных, бурых и
красных водорослей в создании общей видовой структуры (структура построенных
графов показывает непрерывный переход флор районов Северо-западной Пацифики в
пределах бореальной зоны), как наиболее оригинальные флоры выделены центры
видового разнообразия. Показана лучшая интерпретируемость мер сходства в
сравнении с мерами разнообразия и концентрации для определения систематической
структуры сообществ.
. Для определения однородности условий на основе градиента
увлажненности совокупности связанных субассоциаций прирусловых лесов р. Амур
приведена методика непрямой ординации на основе n-арных мер.
4. Рассчитана теоретико-графовая модель временнóй структуры таксоцена
водорослей-макрофитов, на основе которой показано, что виды, вегетирующие в
течение года, сильно размывают представление о сходстве объектов, и их удаление
из общего списка не меняет общую структуру графа. Выявлено, что биологические
сезоны только по видовым спискам с помощью анализа оптимального пути для
годового цикла и дендрограммы невозможно выделить однозначно в силу
континуальной природы исследуемых объектов
Мы убеждены, что все рассмотренные методы будут широко применяться в
биологии для оценки сходства, разнообразия видов, ценозов, особей, генов,
надценотических структур и др. Об этом косвенно говорят факты использования
вышеупомянутых мер и их модификаций (зачастую необоснованных) в гидробиологии,
биохимии, сравнительной геномике, молекулярной биологии, биометрии,
биогеографии, а также в других биологических и географических науках.
Список литературы
1. Абакумов
В.А., Филиппова Л.М. Гидробиологические методы контроля загрязнения водной
среды // Опыт и методы экологического мониторинга. Пущино: Науч. центр биол.
исследований АН СССР, 1978. С. 96-101.
2. Айвазян
С.А. Статистическое исследование зависимостей. - М.: Металлургия, 1968. - 228
с.
3. Айвазян
С.А., Бежаева З.И., Староверов О.В. Классификация многомерных наблюдений. - М.:
Статистика, 1974. - 240 с.
. Айвазян
С.А., Бухштабер В.М., Енюков И.С., Мешалкин Л.Д. Прикладная статистика:
Классификация и снижение размерности. - М.: Финансы и статистика, 1989. - 607
с.
. Акофф
Р.А., Эмери Ф.Ф. О целеустремленных системах. - М.: Сов. радио, 1974. - 272 с.
. Александрова
В.Д. Классификация растительности. Обзор принципов классификации и
классификационных систем в различных геоботанических школах. - Л.: Наука, 1969.
- 275 с.
. Алексахина
Т.И., Штина Э.А. Почвенные водоросли лесных биогеоценозов. - М.: Наука, 1984. -
148 с.
. Алексеенко
Л.Н. Водный режим луговых растений в связи с условиями среды. - Л.: ЛГУ, 1976.
- 200 с.
. Алёхин
В.В. География растений. - М.: Учпедгиз, 1938. - 328 с.
. Алёхин
В.В. География растений. - М.: Советская наука, 1944. - 455 с.
. Алёхин
В.В. Фитосоциология (учение растительных сообществах) и её последние успехи у
нас и на Западе // Методика геоботанических исследований. Л.-М.: Пучина, 1925.
С. 7-95.
. Алпатов
В.В. По поводу статьи Никольского «Об изменчивости организмов» // Зоол. ж. Т.
35. вып. 3. 1956. С. 373-375.
. Андреев
В.Л. Использование мер включения в анализе гидробиологических данных //
Гидробиол. ж. 1978. Т. 14. № 6. С. 34-41.
. Андреев
В.Л. Классификационные построения в экологии и систематике. - М.: Наука, 1980. -
142 с.
. Андреев
В.Л. Системы-классификации в биогеографии и систематике (детерминистские
методы) // Иерархические классификационные построения в географической экологии
и систематике. Владивосток: ТИГ. ДВНЦ АН СССР, 1979. С. 3-59.
. Андреев
В.Л., Козлов В.И. Предварительные списки видов рыб и рыбообразных пресных вод
Понто-Каспия // Теоретико-графовые методы в биогеографических исследованиях.
Владивосток: ДВНЦ АН СССР, 1983. С. 102-115.
. Андреев
В.Л., Решетников Ю.С. Анализ состава пресноводной ихтиофауны северо-восточной
части СССР на основе методов теории множеств // Зоол. ж. 1978. Т. 7. вып. 2. С. 165-175.
. Андреев
В.Л, Решетников Ю.С. Классификационные построения с использованием списков
видов пресноводных рыб Чукотки и Аляски // Зоол. ж. 1981. Т. 10. вып. 9. С. 1285-1296.
. Аптон
Г. Анализ таблиц сопряжённости. - М.: Финансы и статистика, 1982. - 144 с.
. Арманд
А.Д. Информационные модели природных комплексов. - М.: Наука, 1975. - 126 с.
. Арманд
А.Д. Особенности переходной зоны между лесным и болотным ландшафтом в области
Валдайского поозёрья // Организация экосистем ельников южной тайги. М.:
Институт географии АН СССР, 1979. С. 194-213.
. Артемьев
Ю.Т. К трактовке коэффициента вариации в учебниках биологической статистики //
Науч. докл. высш. шк. Биол. науки. № 11. 1969. С. 141-144.
. Афонина
О.М. Парциальная флора окружения горячих ключей: мхи и лишайники. Мхи //
Экосистема термальных источников Чукотского полуострова. Л.: Наука, 1981. С.
71-87.
. Ахтямов
М.Х. Ценотаксономия прирусловых ивовых и ивово-тополёвых и уремных лесов поймы
реки Амур. - Владивосток: Дальнаука, 2001. - 138 с.
. Баранов
В.И., Басаргин Д.Д. К систематике Betula fructosa
Pall. (Betulaceae) // Бот. ж. 1979. Т. 64. № 4. С. 514-524.
26. Баркалов
В.Ю. Сосудистые растения острова Шумшу и Парамушир (северные Курилы) //
Автореф. канд. дисс. биол. наук. Томск: ТГУ, 1981. - 26 с.
. Бартков
Б.И., Сёмкин Б.И., Беликов И.Ф. Количественные методы в исследовании
разнокачественности семян // Физиология растений. Т. 20. вып. 2. 1973. С.
409-411.
. Басаргин
Д.Д., Горовой П.Г., Сёмкин Б.И. Таксономическая характеристика размеров
мерикарпиев у борщевиков Heracleum L. Дальнего
Востока и Северной Америки // Бот. ж. 1978. Т. 63. № 12. С.
1766-1774.
. Батурина
М.Г., Чернобаева М.Б. Справочно-информационные и справочные сайты по ботанике
// Бот. ж. 2007. Т. 92. № 10. С. 1595-1615.
. Бейли
Н. Математика в биологии и медицине. - М.: Мир, 1970. - 326 с.
. Бейтс
Г.У. Натуралист на реке Амазонке: рассказ о тропических картинах природы, о
нравах животных, о жизни бразильцев и индейцев и о путевых приключениях автора
во время его одиннадцатилетних странствий. - М.: Географгиз, 1958. - 430 с.
. Беклемишев
В.Н. Биоценологические основы сравнительной паразитологии. - М.: Наука, 1970. -
502 с.
. Беклемишев
В.Н. Основные понятия биоценологии в приложении к животным компонентам наземных
сообществ // Труды по защите растений. Т. 1. вып. 2. 1931. С. 278-358.
. Беклемишев
В.Н. Термины и понятия, необходимые при количественном изучении популяций
эктопаразитов и нидиколов // Зоол. ж. 1961. Т. 40. вып. 2. С. 149-158.
. Беликович
А.В. Ландшафтная флористическая неоднородность растительного покрова (на
примере модельных районов Северо-Востока России). Владивосток: БПИ ДВО РАН,
2001. 248 с.
. Берг
Р.Л. Дальнейшие исследования по стабилизирующему отбору в эволюции цветка //
Бот. ж. 1958. Т. 43. № 1. С. 12-27.
. Берг
Р.Л. Стабилизирующий отбор в эволюции цветка // Бот. ж. 1956. № 3. С. 318-334.
. Бериков
В.С., Лбов Г.С. Современные тенденции в кластерном анализе // Всероссийский конкурсный
отбор обзорно-аналитических статей по приоритетному направлению
«Информационно-телекоммуникационные системы», 2008. - 26 с. Режим доступа:
http://www.ict.edu.ru/ft/005638/62315e1-st02.pdf.
. Бешел
Р. Флористические соотношения на островах Неоарктики // Бот. ж. 1969. Т. 54.
№6. С. 872-891.
. Борзова
Л.М., Сёмкин Б.И., Ковалюх Н.Н. Развитие островных растительных сообществ в
голоцене (на примере острова Большой Пелис) // Развитие природной среды в
плейстоцене (юг Дальнего Востока). Владивосток: ДВНЦ АН СССР, 1981. С. 127-134.
. Борисовец
Е.Э., Тараканова Т.Ф. Классификация видовых списков полихет из бореальной
области Тихого океана // Иерархические классификационные построения в
географической экологии и систематике. Владивосток: ДВНЦ АН СССР, 1979. С.
131-142.
. Боряков
И.В., Воротников Е.Е., Борякова Е.Е. Использование информационных технологий
для организации фитоценариев и обработки геоботанических данных // Бот. ж.
2005. Т. 90. №1. С. 95-104.
. Бочарников
В.Н. Биоразнообразие: оценка и сохранение на основе технологий ГИС. -
Владивосток: Дальнаука, 1998. 288 с.
. Быков
Б.А. Геоботаника. - Алма-Ата: АН Каз.ССР, 1953. - 457 с.
. Быков
Б.А. Геоботаника. 2-е изд., испр. - Алма-Ата: АН Каз.ССР, 1957. - 367 с.
. Быков
Б.А. Геоботаника. 3-е изд., перераб. - Алма-Ата: Наука, 1978. - 288 с.
. Быков
Б.А. Геоботанический словарь. 2-е изд., перераб. и доп. - Алма-Ата: Наука,
1973. - 214 с.
. Быков
Б.А. Экологический словарь. - Алма-Ата: Наука, 1983. - 216 с.
. Быховский
М.Л., Вишневский А.А. Кибернетические системы в медицине. - М.: Наука, 1971. -
408 с.
. Ванчуров
И.А. Математический метод сравнения палеонтологических объектов // Палеонтол.
ж. 1975. № 2. С .141-148.
. Варченко
Л.И. Обобщения парных мер ассоциации на многомерный случай // Математические
методы в экологии и географии. Владивосток: ДВНЦ АН СССР, 1978. С. 21-30.
. Василевич
В.И. К методике выделения растительных ассоциаций с помощью математических
методов // Методы выделения растительных ассоциаций. Л.: Наука, 1971. С.
111-124.
. Василевич
В.И. Количественные методы изучения структуры растительности // Итоги науки и
техники. Ботаника. М.: 1972. Т. 1. С.
7-83.
. Василевич
В.И. О применении статистических методов для характеристики растительности //
Вестник ЛГУ. 1960. № 9. вып.
2. С. 64-70.
. Василевич
В.И. Очерки теоретической фитоценологии. - Л.: Наука, 1983. - 248 с.
. Василевич
В.И. Статистические методы в геоботанике. - Л.: Наука, 1969. - 232 с.
. Васильев
Н.Г., Сёмкин Б.И. Сравнение динамики хода роста двух типов лесов из ясеня
маньчжурского // Мат. IV
Всесоюз. совещ. «Количественные методы анализа растительности». Уфа: 1974. С.
159-160.
. Викентьев
А.А., Лбов Г.С. О метризациях булевой алгебры предложений и информативности
высказываний экспертов // Докл. АН. Информатика. 1998. Т. 361. №2. С. 174-176.
. Вилли
К. Биология. М.: Мир, 1966. - 685 с.
. Винберг
Г.Г. Многообразие и единство жизненных явлений и количественные методы в
биологии // Журн. общ. биол. 1981. Т. 42. № 1. С. 5-18.
. Воронин
Ю.А. Введение мер сходства и связи для решения геолого-геофизических задач //
Докл. АН СССР. 1971. Т. 139. №5. С. 64-70.
. Воронин
Ю.А. Начала теории сходства. - Новосибирск: Наука. Сиб. отд-ние, 1991. - 128 с.
. Воронов
А.Г. Геоботаника. - М.: Высш. школа, 1973. - 384 с.
. Воронов
А.Г., Арустамова Д.М. О некоторых коэффициентах сходства и различия // Бюлл.
МОИП. отд. биол. 1975. Т. 80. № 5. С. 119-121.
. Выханду
Л.К. Об исследовании многопризнаковых биологических систем // Применение
математических методов в биологии. Л.: вып. 3. 1964. С. 19-22.
. Гайдышев
И.П. Анализ и обработка данных. Специальный справочник. - СПб.: Питер, 2001. -
752 с.
. Галанин
А.В. Анализ распределения растений по типам местообитаний (на примере района
равнинно-горного ландшафта среднего течения р. Рау-Чуа Западной Чукотки) //
Бот. ж. 1977. Т. 62. № 8. С. 1089-1101.
. Галанин
А.В. Флора и ландшафтно-экологическая структура растительного покрова. -
Владивосток: ДВО АН СССР, 1991. - 272 с.
. Галанин
А.В. Флора и растительность Усть-Чаунского биологического стационара (Западная
Чукотка) // Бот. ж. 1980. Т. 65. № 9. С. 1174-1187.
. Галанин
А.В. Экотопологическая структура флоры хребта Куркуре (Восточный Алтай) // Бот.
ж. 1979. Т. 64. № 10. С. 1041-1413.
. Ганнибал
Б.К. Некоторые проблемы геоботаники в свете современных концепций
биологического разнообразия // III
Всерос. школа-конф. «Актуальные проблемы геоботаники». Ч.1. Петрозаводск: 2007.
С. 129-133.
. Гелашвили
Д.Б., Иудин Д.И., Розенберг Г.С., Якимов В.Н. Степенной характер накопления
видового богатства как проявление фрактальной структуры биоценоза // Журн. общ.
биол. 2007. Т. 67. №3. С. 170-179.
. География
и мониторинг биоразнообразия // Колл. авторов. - М.: Научный и
научно-методический центр, 2002. - 432 с.
. Гиляров
А.М. Перестройка в экологии: от описания видимого к пониманию скрытого //
Вестн. РАН. 2005. Т. 75. №3. С. 214-223.
. Гиляров
А.М. Соотношение биомассы и видового разнообразия в планктонном сообществе //
Зоол. ж. 1969. Т. 48. № 4. С. 485-493.
. Голлербах
М.М. К вопросу о составе и распространении водорослей в почвах // Тр. Бот.
ин-та АН СССР. сер. 2. вып. 3. 1936. С. 99-301.
. Горшков
М.В. Концепция биоинформационной системы «Биоразнообразие» и её использование
для оценки компонентов биоразнообразия // Усп. совр. естествознания. №11. 2008.
С. 61-62.
. Горшков
М.В. Программа для сравнительного анализа биологических объектов
«BioComparison» // Хроники объединенного фонда электронных ресурсов «Наука и
образование». №8 (15) август 2010. С. 29. [Электронный ресурс]. - Режим
доступа: http://ofernio.ru/portal/newspaper/ofernio/2010/8.doc
. Грейг-Смит
П. Количественная экология растений. - М.: Мир, 1967. - 360 с.
. Грюнбаум
Б. Этюды по комбинаторной геометрии и теории выпуклых тел. Математическая
библиотечка. - М.: Наука, 1971. - 96 с.
. Гусарова
И.С. Макрофитобентос залива Восток (Японское море) // Комаровские чтения. Вып. XXXV. Владивосток: ДВО АН СССР, 1988. С.
11-35.
. Гусарова
И.С. Макрофитобентос сублиторальной зоны островов Итуруп, Уруп и Симушир
(Большая Курильская гряда) // Автореф. канд. дисс. Владивосток: 1975. - 23 с.
. Гусарова
И.С., Сёмкин Б.И. Сравнительный анализ конкретных флор сублиторали
Северо-западной части Тихого океана // Тез. докл. 3 Всесоюз. конф. по морской
биол. «Биология шельфовых зон мирового океана». Ч. 1. Владивосток: ДВНЦ АН
СССР, 1982. С. 18-19.
. Гусарова
И.С., Сёмкин Б.И. Сравнительный анализ флор макрофитов некоторых районов
северной части Тихого океана с использованием теоретико-графовых методов //
Бот. ж. 1986. Т. 71. №6. 1986. С. 781-789.
. Дажо
Р. Основы экологии. - М.: Мир, 1975. - 415 с.
. Джефферс
Дж. Введение в системный анализ: применение в экологии. - М.: Мир, 1981. - 256
с.
. Джини
К. Средние величины. - М.: Статистика, 1970. - 448 с.
. Дмитриев
Е.А. Математическая статистика в почвоведении. - М.: МГУ, 1972. - 292 с.
. Дыренков
С.А., Волков А.Д. Выявление степени сходства структуры древостоев с применением
методов теории множеств // Количественные методы анализа растительности. Рига:
Т. 2. 1971. С. 83-86.
. Дюльдин
А.А. Коэффициент вариации и аллометрия // Экология. 1973. № 6. С. 97-99.
. Дюран
Б., Оделл П. Кластерный анализ. - М.: Статистика, 1977. - 128 с.
. Егоров
Ю.Е. Диапазон изменчивости и его связь с абсолютной величиной признака и
формообразовательными процессами // Журн. общ. биол. Т. 30. № 6. 1969. С.
658-663.
. Елисеева
И.И., Рукавишников В.О. Группировка, корреляция, распознавание образов:
(статистические методы классификации и измерения связей). - М.: Статистика,
1977. - 143 с.
. Ефимов
В.М. О некоторых коэффициентах межвидовой сопряжённости // Изв. Сиб. отд. АН
СССР. сер. биол. 1976. вып. 2. № 10. С. 28-30.
. Жадовский
А.Е. Метод Жаккара (Jaccard) в
изучении растительных сообществ // Методика геоботанических исследований.
Л.-М.: Пучина, 1925. С. 97-117.
. Жизнь
растений. В 6 т. Т. 3. Водоросли. Лишайники / гл. ред. А.А. Фёдоров; 3-й том
под ред. М.М. Голлербаха. - М.: Просвещение, 1977. - 487 с.
. Журавлёва
Н.А., Горшкова А.А. Реакция отдельных компонентов степных сообществ Забайкалья
на различные условия увлажнения // Количественные методы анализа
растительности. Рига: Латв. гос. ун-т. 1971. С. 87-93.
. Загоруйко
Н. Г. Прикладные методы анализа данных и знаний. - Новосибирск: ИМ СО РАН,
1999. - 270 с.
. Зайцев
Г.Н. Математическая статистика в экспериментальной ботанике. - М.: Наука, 1984.
- 424 с.
. Заки
М.А., Шмидт В.М. О систематической структуре флор южного Средиземноморья. I. Методика и анализ структуры 5
региональных и 11 локальных флор // Вестник ЛГУ. Биология. 1972. № 9. вып. 2.
С. 57-69.
. Зарубин
С.И. Определение доверительных значений коэффициента сходства фитоценозов //
Количественные методы анализа растительности. Рига: Латв. гос. ун-т. 1971. С.
101-104.
. Захаров
Ю.Д. Раннетриасовые аммоноидеи Востока СССР. - М.: Наука, 1978. - 224 с.
. Зверев
А.А. Сравнительный анализ флор с помощью компьютерной системы «IBIS» // Мат. IV раб. совещ. по сравнительной флористике «Изучение
биологического разнообразия методами сравнительной флористики». Березинский
биосферный заповедник., 1993: СПб.: НИИХ СпбГУ, 1998. С. 284-288.
. Злобин
Ю.А. Ценотические популяции растений. - Владивосток: ДВНЦ АН СССР, 1984. - 52
с.
. Ильина
И.С., Кобелева Н.В. Количественный анализ карт природы с целью районирования:
На примере геоботанической карты Среднего Приобья. - Л.: ЛГУ, 1989. - 276 с.
. Ипатов
В.С. Сравнение методов определения роли вида в структуре травяного покрова
дубового леса // Бот. ж. 1962. Т. 47. № 3. С. 360-368.
. Ипатов
В.С., Самойлов Ю.И., Тархова Т.Н. Двусторонний коэффициент межвидовой
сопряжённости // Бот. ж. 1974. Т. 59. № 11. С. 1596-1602.
. Йогансен
В. Элементы точного учения об изменчивости и наследственности с основами
биологической вариационной статистики. - М.-Л.: Сельколхозгиз, 1933. - 410 с.
. Йогансен
В. О наследовании в популяциях и чистых линиях. - М.-Л.: Огиз.-сельхозгиз,
1935. - 80 с.
. Калугина-Гутник
А.А. Изменение видового состава фитобентоса в бухте Ласпи за период 1964-1985
гг. // Экология моря. Киев: Наукова Думка, Вып. 31. 1989. С. 7-11.
. Кафанов
А.И. Биогеография: география или биология? // Журн. общ. биол. 2009. Т. 70. №
1. С. 46-65.
. Кафанов
А.И. Влияние видового богатства и обилия на меры включения при
теоретико-графовом анализе // Изв. РАН. сер. биол. №3. 1994. С. 428-434.
. а
Кафанов А.И. Историко-методологические аспекты общей и морской биогеографии. -
Владивосток. ДВГУ, 2005. - 208 с.
. б
Кафанов А.И. Континуальность и дискретность геомериды: биономический и
биотический аспекты // Журн. общ. биол. 2005. Т. 66. № 1. С. 28-54.
. Кафанов
А.И. Континуальность и дискретность живого покрова: проблема масштаба // Журн.
общ. биол. 2006. Т. 67. № 4. С. 311-313.
. Кафанов
А.И., Жуков В.Е. Прибрежные сообщества водорослей-макрофитов залива
Посьет (Японское море): Сезонная динамика и пространственная структура. -
Владивосток: Дальнаука, 1993. - 156 с.
117. Кафанов А.И., Кудряшов
В.А. Классики биогеографии: библиографический указатель // 2005. [Электронный
ресурс]: <http://biogeografers.dvo.ru/>
. Кафанов А.И.,
Борисовец Е.Э., Волвенко И.В. О применении кластерного анализа в
биогеографических классификациях // Журн. общ. биол. 2004. Т. 65. №3. С.
250-265.
. Кафанов А.И., Жуков
В.Е, Федотов П.А. Состав и распределение летней фауны амфипод (Amphipoda, Gammaridea) на литорали бухты Витязь // Гидробиологические
исследования заливов и бухт Приморья. Владивосток: ДВНЦ АН СССР, 1984. С.
52-68.
. Кац Н.Я., Кац С.В.,
Кипиани М.Г. Атлас и определитель плодов и семян, встречающихся в четвертичных
отложениях СССР. - М.: Наука, 1965. - 367 с.
. Кендалл М., Стюарт
А. Статистические выводы и связи. - М.: Наука, 1973. - 900 с.
. Клайн М. Математика.
Утрата определенности. - М.: Мир, 1984. - 434 с.
123. Клочкова
Н.Г. Водоросли-макрофиты дальневосточных морей России // Дисс. докт. биол.
наук. Петропавовск.-Камчатский: КИЭП, 1997. - 277 с.
124. Клочкова
Н.Г. Флора водорослей-макрофитов Татарского пролива и особенности её
формирования. - Владивосток: Дальнаука, 1996. - 288 с.
125. Клочкова
Н.Г., Березовская В.А. Макрофитобентос Авачинской губы и его антропогенная
деструкция. - Владивосток: Дальнаука, 2001. - 208 с.
126. Кобелева
Н.В. Количественный метод анализа геоботанических описаний таёжной территории
для крупномасштабного картографирования (на примере Атлымского водораздела) //
Растительность Западной Сибири и её картографирование, Новосибирск: Наука. Сиб.
отд., 1984. С. 106-119.
. Ковалевская
В.Б., Погожев И.Б., Погожева (Кусургашева) А.П. Количественные методы оценки
степени близости памятников по процентному содержанию массового материала //
Советская археология. 1970. № 3. С. 26-39.
. Кожевников
Ю.П. Анализ флоры Телекойской рощи и её окрестностей (Центральная Чукотка) //
Бот. ж. 1974. Т. 59. № 7. С .967-979.
. Коженкова
С.И. Ретроспективный анализ морской флоры залива Восток Японского моря //
Биология моря. 2008. Т.34. №3. С. 159-174.
. Коли
Г. Анализ популяций позвоночных. - М.: Мир, 1979. - 362 с.
. Количественные
методы в исторических исследованиях / Под ред. И.Д. Ковальченко. - М.: Высшая
школа, 1984. - 384с.
. Количественные
методы экологии и гидробиологии / Под ред. Розенберга Г.С. - Тольятти: ИЭВБ
РАН, 2005. - 404 с.
. Колмогоров
А.Н. Основные понятия теории вероятности. - М.: Наука, 1974. - 120 с.
. Коломыц
Э.Г. Кристалло-морфологический атлас снега. - Л.: Гидрометеоиздат, 1984. - 215
с.
. Коломыц
Э.Г. Структура снега и ландшафтная индикация. - М.: Наука, 1976. - 208 с.
. Комарова
Т.А. О горизонтальной неоднородности пихтово-еловых мелкотравно-папоротниковых
лесов северного Сихотэ-Алиня // Экология. 1978. № 1. С. 42-49.
. а
Комарова Т.А., Сёмкин Б.И. Динамика растительного покрова долины р. Амгуэмы
(Центральная Чукотка) // Биологические проблемы Севера. VII Симпозиум. Ботаника. Петрозаводск:
1976. С. 123-125.
. б
Комарова Т.А., Сёмкин Б.И. Первичные сукцессии тундровых сообществ долины реки
Амгуэма. // Комаровские чтения. Владивосток: ДВНЦ АН СССР, 1976. вып. 24. С. 36-61.
. Компьютерные
базы данных в ботанических исследованиях // Тез. докл. II совещания: СПб.: 1995. - 60с.
. Компьютерные
базы данных в ботанических исследованиях // Тез. докл III совещания: СПб.: 1997. - 52 с.
. Константинов
А.С. Использование теории множеств в биогеографическом и экологическом анализе
// Усп. соврем. биол. 1969. Т. 67. вып. 1. С 99-108.
. Корчагин
А.А. Строение растительных сообществ // Полевая геоботаника. 1976. Т. 5. Л.:
Наука. С. 7-313.
. Котенко
Т.И. О коэффициентах вариации // Количественные методы в экологии животных. Л.:
ЗИН АН СССР. 1980. С. 72-73.
. Котов
В.Н. Применение теории измерений в биологических исследованиях. - Киев: Наукова
думка, 1985. - 100 с.
. Крамер
Г. Математические методы статистики. - М.: Мир, 1975. - 848 с.
. Красилов
В.А. Палеоэкология наземных растений (основные принципы и методы). -
Владивосток: ДВНЦ АН СССР, 1972. - 210 с.
. Красникова
Л.И., Антомонов Ю.Г. К применению энтропийных оценок для анализа импульсной
активности Нерона // Биологическая, медицинская кибернетика и бионика. вып. 4.
Киев: 1970. С. 37-50.
. Крестов
П.В. Использование флористической базы данных для фитогеографических сравнений
на Дальнем Востоке Азии // Информационные и телекоммуникационные ресурсы в
зоологии и ботанике. СПб.: 2001. С. 32.
. Крестов
П.В. Предложения к флористическому районированию северной Азии на основе
сравнительного анализа флор на родовом уровне // Комаровские чтения. вып. LI. Владивосток: Дальнаука, 2005. С.
15-56.
. Крестов
П.В. Растительный покров и фитогеографические линии Северной Пацифики //
Автореф. дисс. докт. биол. наук. - Владивосток: 2006. - 42 с.
. Крэйн
М., Лемуан О. Введение в регенеративный метод анализа моделей. - М.: Наука,
1982. - 104с.
. Куликова
Л.С. О подготовке энтомологического материала для математического анализа //
Теоретико-графовые методы в биогеографических исследованиях, Владивосток: ДВНЦ
АН СССР, 1983. С 115-129.
. Кусакин
О.Г., Ростомов С.А. Биогеографическая структура фауны равноногих ракообразных
шельфа западного Сахалина и острова Монерон // Морская биогеография. Предмет,
методы, принципы районирования. М.: Наука, 1982. С. 176-184.
. Кушнарёв
Е.Л. Применение графических методов анализа при изучении видовой структуры
орнитосообществ по экологическому профилю // Теоретико-графовые методы в
биогеографических исследованиях. Владивосток: ДВНЦ АН СССР, 1983. С. 33-48.
. Лакин
Г.Ф. Биометрия. - М.: Высш. шк., 1990. - 352 с.
. Лапинскене
Н.А. Подземная часть травянистых растений и фитоценозов в Литовской ССР. -
Вильнюс: Мокслав, 1986. - 176с.
. Лащинский
Н.Н., Ронгинская А.В. Особенности флористического состава травяных сосновых
лесов Нижнего Приангарья // Изв. Сиб. отд. АН СССР. сер. биол. 1975. Вып. 2. №
10. С. 50-60.
. Лбов
Г.С. Методы разработки разнотипных экспериментальных данных. - Новосибирск:
Наука, 1981. - 161 с.
. Лебедева
Н.В, Дроздов Н.Н., Криволуцкий Д.А. Биоразнообразие и методы его оценки. - М.:
МГУ, 1999. - 94 с.
. Левич
А.П. Структура экологических сообществ. - М.: МГУ, 1980. - 181 с.
. Леме
Ж. Основы биогеографии. - М.: Прогресс, 1976. - 309 с.
. Ленг
С. Введение в теорию диофантовых приближений. - М.: Мир, 1970. - 104 с.
. Леонов
В.П. Кластерный анализ: основы метода и его применение в биомедицине //
[Электронный ресурс] : [Статья] - Режим доступа:
http://www.biometrica.tomsk.ru/cluster.htm.
. Ли
Ч. Введение в популяционную генетику. - М.: Мир, 1978. - 555 с.
. Лысенко
С.В. Флористическая общность микрофитоценозов - как один из критериев
объединения сообществ в ассоциации // Количественные методы анализа
растительности. II. Мат. третьего
Всесоюз. совещ.: Применение количественных методов при изучении структуры
растительности. Рига: Латв. гос. ун-т, 1971. С .185-189.
. Любищев
А.А. О критериях изменчивости организмов // Изв. биол. НИИ Пермск. гос. ун-та.
1923. Т. 1. вып. 7-8.
. Макаревич
В.Н. Применение метода Чекановского при изучении динамики флористического
состава лугового сообщества // Бот. ж. Т. 52. № 11. 1967. С. 1630-1639.
. Макаревич
В.Н. Применение метода Чекановского при первичной обработке геоботанических
описаний // Методы выделения растительных ассоциаций. Л. Наука. 1971. С.
125-140.
. Макиенко
В.Ф. Водоросли-макрофиты залива Восток (Японское море) / В.Ф. Макиенко //
Биология моря. 1975. №2. С. 45-57.
. Максимов
В.Н. Метрологические свойства индексов сходства (в приложениях к биологическому
анализу качества поверхностных вод) // Комплексные оценки качества
поверхностных вод. Л.: Гидрометеоиздат, 1984. С. 77.
. Максимова
В.Ф. Использование некоторых параметров травяного яруса для оценки различий
приморских и континентальных дубрав восточного склона Сихотэ-Алиня //
Современные проблемы биогеографии. М.: МГУ, 1982. С. 152-163.
. Максимова
В.Ф., Нечаева Н.С., Сёмкин Б.И. Сравнительный анализ травяного яруса в лесах
среднего Сихотэ-Алиня с помощью мер включения // Локальный мониторинг
растительного покрова. Владивосток: ДВНЦ АН СССР, 1982. С. 102-110.
. Макфедьен
Э. Экология животных. Цели и методы. - М.: Наука, 1965. - 375 с.
. Малышев
Л.И. Зависимость флористического богатства от внешних условий и исторических
факторов. // Бот. ж. 1969. Т. 54. № 8. С. 1137-1147.
. Малышев
Л.И. Моделирование флористического деления Европы с помощью кластерного анализа
// Бот. ж. 2002. Т. 87. № 7. С. 16-33.
. Малышев
Л.И. Основы флористического районирования // Бот. ж. 1999. Т. 84. № 1. С. 3-14.
. Малышев
Л.И. Синдромы в сравнительной флористике // Мат. конф. «Проблема вида и
видообразование» Т. 1. «Эволюционная биология». Томск: ТГУ, 2001. С. 190-206.
. Малышев
Л.И. Современные подходы к количественному анализу и сравнению флор // Мат. II раб. совещ. по сравнительной
флористике «Теоретические и методические проблемы сравнительной флористики»:
Неринга, 1983. - Л.: Наука, 1987. С. 142-148.
. Малышев
Л.И. Флористические спектры Советского союза // История флоры и растительности
Евразии. Л.: Наука, 1972. С. 17-40.
. Малышев
Л.И., Байков В.М. Таксономические спектры флоры Сибири на уровне семейств //
Бот. ж. 1998. №10. С. 3-17.
. Мамаев
С.А. Формы внутривидовой изменчивости древесной растительности (на примере Pinaceae на Урале). - М.: Наука, 1972. - 284
с.
182. Мандель И.Д. Кластерный
анализ <http://www.biometrica.tomsk.ru/lib/books/mandel.djvu>. - М.:
Финансы и статистика, 1988. - 176 с.
. Марина Л.В. Опыт
сравнительного анализа высокогорных флор речных бассейнов хребта Куркуре
(Восточный Алтай) // Бот. ж. 1982. Т. 67. № 3. С. 285-292.
. Марина Л.В.
Сравнительный анализ двух высокогорных флор Восточного Алтая (по их
внутриландшафтным подразделениям) // Автореф. канд. дисс. биол. наук. - Л.:
1983. - 23 с.
. Марина Л.В., Марин
Ю.Ф. Пример использования информационной системы Висимского заповедника в
ботанических исследованиях // Мат. IV раб. совещ. по сравнительной
флористике «Изучение биологического разнообразия методами сравнительной
флористики»: Березинский биосферный заповедник. 1993: СПб.: НИИХ СПбГУ, 1998.
С. 273-284.
. Мещеряков Е.В.,
Хомоненко А.Д. Публикация баз данных в Интернете. - СПб.: БХВ-Петербург, 2001.
- 560 с.
. Миркин Б.М. Введение
в количественные методы анализа растительности. - Уфа: БГУ, 1970. - 87 с.
. Миркин Б.М. Значение
классификации растительности для современной экологии // Журн. общ. биол. 2004.
Т. 65. №2. С. 167-177.
. Миркин Б.М. Проблема
соотношения непрерывности и дискретности и современная экология // Журн. общ.
биол. 2005. Т. 66. № 6. С. 520.
. Миркин Б.М., Наумова
Л.Г. Метод классификации растительности по Браун-Бланке в России // Журн. общ.
биол. 2009. Т.70. № 1. С. 66-77.
. Миркин Б.М.,
Розенберг Г.С. Количественные методы классификации, ординации и геоботанической
индикации // Итоги науки и техники. Сер. ботаника. М.: ВИНИТИ, 1979. Т. 3. С.
71-137.
. Миркин Б.М., Розенберг
Г.С. Толковый словарь современной фитоценологии. - М.: Наука, 1983. - 134 с.
. Миркин Б.М.,
Розенберг Г.С. Фитоценология. Принципы и методы. - М.: Наука, 1978. - 211 с.
. Миркин Б.М., Наумова
Л.Г., Соломещ А.И. Современная наука о растительности. - М.: Логос, 2002. - 264
с.
. Миркин Б.М.,
Розенберг Г.С., Наумова Л.Г. Словарь понятий и терминов современной
фитоценологии. - М.: Наука, 1989. - 223 с.
. Михалёва Е.В.,
Петухова Е.Л. Сравнительный анализ фауны диплопод (Diplopoda) лесов
Приморского края с помощью мер включения и сходства // Теоретико-графовые
методы в биогеографических исследованиях. Владивосток: ДВНЦ АН СССР, 1983. С.
48-66.
. Мэгарран Э.
Экологическое разнообразие и его измерение. - М.: Мир, 1992. - 184 с.
. Науменко Н.И.
Локальные флоры и флористические границы в лесостепном Зауралье // Мат. IV
раб. совещ. по сравнительной флористике «Изучение биологического разнообразия
методами сравнительной флористики»: Березинский биосферный заповедник, 1993.
СПб.: НИИХ СПбГУ, 1998. С. 54-70.
. Наумов Р.Л. Птицы в
очагах клещевого энцефалита Красноярского края // Автореф. канд. дисс. биол.
наук. - М.: 1964. - 9 с.
. Нееф Э.
Теоретические основы ландшафтоведения. - М.: Прогресс, 1974. - 218 с.
. Неронов В.М.,
Лушекина А.А. Региональная биогеография Монгольской Народной Республики (анализ
опубликованных схем районирования) // Итоги науки и техники. Биогеография. М.:
1980. Т. 2. С. 41-64.
. Несис К.Н.
Зоогеографическое положение Средиземного моря // Морская биогеография. Предмет,
методы, принципы районирования. М.: Наука, 1982. С. 270-306.
. Ниценко А.А.
Растительная ассоциация и растительное сообщество как первичные объекты
геоботанического обследования. - Л.: Наука, 1971. - 183 с.
. Новаковский А.Б.
Обзор программных средств, используемых для анализа геоботанических данных //
Растительность России. 2006. №9. С. 86-95.
. Норин Б.Н. Структура
растительных сообществ Восточно-европейской лесотундры. - Л.: Наука, 1979. -
200 с.
. Одум Ю. Основы
экологии. - М.: Мир, 1975. - 740 с.
. Оре О. Графы и их
применение. - М.: Едиториал УРСС, 2002. - 168 с.
. Оре О. Теория
графов. - М.: Наука, 1969. - 352 с.
. Орлов А.И.
Нечисловая статистика. - М.: МЗ-Пресс, 2004. - 513 с.
. Орлов А.И. О
развитии математических методов теории классификации // Заводская лаборатория.
2009. Т. 75. №7. С. 51-63.
. Основы
геоинформатики: В 2 кн. Учеб. пособие для студ. вузов // Под ред. В.С.
Тикунова. - М.: Академия. 2004. Кн. 1. 352 с. Кн. 2. 480 с.
. Остроумова Т.А.
Интернет и ботанические коллекции // Бот. ж. 2000. Т. 85. №1. С. 123-124.
. Пааль Я.Л., Сёмкин
Б.И. Анализ растительности средней тайги при помощи мер включения и сходства //
Бот. ж. 1983. Т. 68. № 10. С. 1341-1350.
. Перестенко Л.П. О
принципах зонального биогеографического районирования шельфа Мирового океана и
о системах зон // Морская биогеография. М.: Наука, 1982. С. 99-114.
. Песенко Ю.А. О
классификации известных мер соответствия и об эквивалентности некоторых из них,
выводимых на основе концепций общности, расстояния, корреляции и разнообразия
// Количественные методы в экологии животных. Л.: 1980. С. 103-107.
. Песенко Ю.А.
Принципы и методы количественного анализа в фаунистических исследованиях. - М.:
Наука, 1982. - 287 с.
. Песенко Ю.А., Сёмкин
Б.И. Измерение видового разнообразия сообществ и ширины экологических ниш
видов: индуктивный и плюралистический подход // Экосистемные исследования:
историко-методологические аспекты. Владивосток: ДВО АН СССР, 1989. С. 133-159.
. Петровский В.В.,
Королёва Т.Н. К флоре дельты реки Колымы // Бот. ж. 1979. Т. 64. № 1. С. 19-31.
. Петропавловский Б.С.
Опыт и перспективы геоботанического картографирования на основе
математико-картографического моделирования с применением ГИС-технологий,
интегрированных картографических программ // Картография на рубеже тысячелетий:
1-я всероссийская научная конференция по картографии. М.: 1997. С. 552-556.
. Петропавловский Б.С.
Принципы составления корреляционных эколого-фитоценотических карт мира и ее
использование с целью оптимального размещения биосферных станций мониторинга
окружающей среды // Разработка и внедрение на комплексных станциях методов
биологического мониторинга. Т. 1. Материалы первой международной школы
биологического мониторинга. Рига. Зинатне, 1983. С. 99-104.
. Петропавловский
Б.С., Сёмкин Б.И., Худяков Г.И. Принципы и методы анализа взаимосвязей между
компонентами геосистем для задач оптимизации охраны и рационального
использования природных ресурсов // Тез. докл. VII совещ.
географов Сибири и Дальнего Востока «Задачи географов в реализации планов XI
пятилетки». Иркутск: Институт географии Сибири и Дальнего Востока СО АН СССР,
1982. С. 89-90.
. Пийн Т.Х.
Напочвенные лишайники и их местообитания на мысе Челюскин (Таймыр) // Структура
растительности полярных пустынь и болот. Труды по ботанике. Тарту: ТГУ, 1982.
Т. 59. С. 22-38.
. Пименов М.Г.,
Терёхин А.Т., Девяткова Г.Н., Баранова Ю.В. Классификация видов рода Ferula L. (Umbelliferae) с помощью метода иерархического кластер-анализа //
Вопр. кибернетики. вып. 47. Математико-статистические методы анализа и
планирования эксперимента. М.: 1978. С. 98-113.
. Плохинский Н.А.
Математические методы в биологии. - М.: МГУ, 1978. - 226 с.
. Погребов В.Б.,
Горянин О.О. Литораль полузамкнутой акватории Белого моря в условиях
распреснения. 2. Нумерическая классификация биоты // Вестник ЛГУ. Сер. 3 вып.
3. 1988. С. 21-28.
. Полозова Т.Г.
Жизненные формы сосудистых растений в различных подзонах таймырской тундры //
Жизненные формы: структуры, спектры и эволюция. М.: Наука, 1981. С. 265-281.
. Полозова Т.Г., Юрцев
Б.А. Парциальная флора окружения горячих ключей: сосудистые растения //
Экосистемы термальных источников Чукотского полуострова. Л.: Наука, 1981. С
.94-121.
. Порк К.М.
Исследование горизонтальной микроструктуры лугов путём анализа межвидовой
сопряжённости // Материалы IV Всесоюзного совещания «Количественные методы анализа
растительности». Уфа: 1974. С. 111-114.
. Применение
количественных методов при изучении структуры фитоценозов. - М.: Наука, 1972. -
176 с.
. Прозоров М.М.
Влияние удобрений на общий запас органического вещества и соотношение подземных
и надземных органов луговых фитоценозов // Растительные ресурсы. 1967. Т.3.
вып.2. С. 212-223.
. Протасов А.А.
Биоразнообразие и его оценка. Концептуальная диверсикология. - Киев: Наукова
думка, 2002. - 105 с.
. Пузаченко Ю.Г.
Инвариантность геосистем и их компонентов // Устойчивость геосистем. М.: Наука,
1983. С. 32-41.
. Пузаченко Ю.Г.
Математические методы в экологических и географических исследованиях. - М.:
Академия, 2004. - 416 с.
. Пузаченко Ю.Г.,
Мошкин А.В. Информационно-логический анализ в медико-географических
исследованиях // Итоги науки и техники. Серия медицинской географии. М.:
ВИНИТИ, 1969. вып. 3. С. 5-74.
. Пушкарь В.С.,
Короткий А.М., Сёмкин Б.И. Кластерный анализ флористических и фитоценотических
описаний ископаемых диатомовых водорослей сообществ голоцена Сихотэ-Алиня //
Бот. ж. 1984. Т. 69. № 11. С. 1532-1542.
. Пфанцагль И. Теория
измерений. - М.: Мир, 1976. - 249 с.
. Пяк А.И., Зверев
А.А. Опыт сравнительного анализа локальных флор с помощью прикладного
статистического пакета Biostat // Бот. ж. 1997. Т. 82. № 5. С. 64-75.
. Пярна К.А. Алгоритм
группировки для объектов, задаваемых номинальными признаками // Тр. Выч. центра
Тартуского ун-та. Тарту: 1976. вып. 36. С .81-94.
. Работнов Т.А. Выбор
опытного участка и подготовка к проведению опытов // Методика опытных работ на
сенокосах и пастбищах. М.: Сельхозгиз, 1961. С. 10-13.
. Работнов Т.А.,
Саурина Н.И. К методике определения площади выявления численности особей Ranuculus acris L. и R. aurucomis L. // Количественные методы анализа растительности. II.
Мат. третьего Всесоюз. совещ: Применение количественных методов при изучении
структуры растительности. Рига: Латв. гос. ун-т. 1971. С. 325-240.
. Равкин Ю.С.
Структурные особенности населения птиц Северо-Восточного Алтая // Орнитология.
1967. № 8. С. 175-191.
. Рагойша А.А.
Информационные технологии в химии // [Электронный ресурс]. - URL.
- http://www.abc.chemistry.bsu.by/it/Lecture-3-np.pdf.
. Раменский Л.Г. О
некоторых принципиальных положениях современной геоботаники // Бот. ж. Т. 37. №
2. 1952. С. 181-202.
. Раушенбах Г.В. Меры
близости и сходства // Анализ нечисловой информации о социологических
исследованиях. М.: Наука, 1985. С. 169-203.
. Ребристая О.В. Флора
востока Большеземельской тундры. -Л.: Наука, 977. - 34 с.
. Ревушкин А.С.
Высокогорная флора Алтая. -Томск: ТГУ, 1988. - 320 с.
. Рейнгольд Э.,
Нивергельт Ю., Део Н. Комбинаторные алгоритмы. Теория и практика. - М.: Мир,
1980. - 478 с.
. Робертс Ф.С.
Дискретные математические модели с приложениями к социальным, биологическим и
экологическим задачам. - М.: Наука, Физматлит, 1986. - 496 с.
. Рогинский Я.Я. О
некоторых результатах применения количественного метода к изучению
морфологической изменчивости // Архив АГ и Э. 1959. Т. 36. № 1. С. 83-89.
. Розенберг Г.С. Обзор
методов статистической геоботаники. 3. Методы автоматической классификации. М.:
1977. 83 с. Деп. В ВИНИТИ 11.04.1977. № 1321-77.
. Розенберг Г.С. О
сравнении различных методов автоматической классификации // Автоматика и
телемеханика. 1975. №9. С. 145-148.
. Росс Ю.К., Власова
М.П. Биометрическая характеристика и динамика развития посева кукурузы //
Фотосинтезирующие системы высокой продуктивности. М.: Наука, С. 78-95.
. Сапегин Л.М.
Структура и изменчивость луговых фитоценозов. - Минск: БГУ, 1981. - 100 с.
. Свирежев Ю.М.,
Логофет Д.О. Устойчивость биологических сообществ. - М.: Наука, 1978. - 352 с.
. Свирежев Ю.М.,
Пасеков В.П. Основы математической генетики. - М.: Наука, 1982. - 512 с.
. Седельников В.П. К
применению мер включения в сравнительной флористике // Нетрадиционные методы в
исследованиях растительности Сибири. Новосибирск: Наука. Сиб. отд., 1982. С.
32-35.
. Селиванова О.Н.
Макрофиты российского шельфа Берингова моря, Командорскиз островов и
юго-восточной Камчатки // Автореф. докт. дисс. биол. наук. - Владивосток: ДВГУ,
2004. - 38 с.
. Сергиенко В.Г.
Материалы к анализу конкретных флор полуострова Канин // Вестник ЛГУ. 1979. №
21. вып. 4. С. 37-45.
. Сёмкин Б.И. Анализ
структур фитоценотических данных // Автореф. канд. дисс. биол. наук., Тарту:
Тартуский гос. ун-т. 1977. 49 с.
. а Сёмкин Б.И.
Дескриптивные множества и их приложения // Исследование систем. Т. 1. Анализ
сложных систем. Владивосток: ДВНЦ АН СССР, 1973. С 83-94.
. Сёмкин Б.И. К
методике анализа разновеликих множеств в сравнительной флористике //
Комаровские чтения. Вып. LVI. 2008. C. 179-195.
. Сёмкин Б.И.
Количественная оценка дифференцирующего разнообразия для целей сравнительного
анализа растительных сообществ и их классификации // Классификация и динамика
лесов Дальнего Востока. Дальнаука, 2001. С. 286-287.
. Сёмкин Б.И.
Количественные показатели для оценки односторонних флористических связей,
предложенных Б.А. Юрцевым // Бот. ж. 2007. Т. 92. №4. С. 114-127.
. Сёмкин Б.И.
Математические методы в фитоценологических исследованиях // Тез. докл. XII
Междунар. бот. конгресса. Л.: Наука, 1975. С. 200.
. Сёмкин Б.И.
Методологические принципы исследования сложных систем в экологии и географии //
Математическое моделирование природных систем. Владивосток: ДВНЦ АН СССР, 1981.
С. 5-9.
. Сёмкин Б.И. О
математических методах выделения растительных ассоциаций // Эмпирические методы
исследования и моделирования растительных сообществ. Владивосток: ДВО РАН СССР,
1988. С. 3-28.
. а Сёмкин Б.И. О мере
сходства между растительными сообществами // Тез. докл. совещ. по классиф.
растит. Л.: Наука, 1971. С. 85.
. а Сёмкин Б.И. О
постановке задач в структурной биоценологии // Теоретико-графовые методы в
биогеографических исследованиях. Владивосток: ДВНЦ АН СССР, 1983. С. 5-12.
. Сёмкин Б.И. О связи
между средними значениями двух мер включения и мерами сходства // Бюлл. БСИ ДВО
РАН [Электронный ресурс] : науч. журн. / Ботан. сад-институт ДВО РАН. -
Владивосток, 2009. Вып. 3. С. 91-101. - http://www.botsad.ru/
journal/number3/02.pdf.
. б Сёмкин Б.И. О
теоретико-множественных методах изучения растительных сообществ // Тез. докл. V
делегатского съезда Всесоюз. бот. о-ва. Киев: Наукова Думка, 1973. С. 210-211.
. а Сёмкин Б.И. Об
аксиоматическом подходе определению мер различия и квазиразличия на семействах
множеств // Информационные методы в системах управления измерения и контроля.
Т. 1. Владивосток: ДВНЦ АН СССР, 1972. С 23-26.
. в Сёмкин Б.И. Об
информационных мерах и метриках // Исследование систем. Т. 1. Анализ сложных
систем. Владивосток: ДВНЦ АН СССР, 1973. С. 105-117.
. а Сёмкин Б.И. Об
использовании математических методов для оценки состояния окружающей среды //
Методы оценки состояния природной среды Владивосток: ДВО АН СССР, 1987. С.
163-165.
. Сёмкин Б.И. Об
использовании метода анализа разнообразий в исследовании водных экосистем //
Проблемы морской биогеографии. Владивосток: ДВНЦ АН СССР, 1980. С. 106-107.
. б Сёмкин Б.И. Об
одном подходе к определению разнообразия растительных сообществ //
Количественные методы анализа растительности. Том II. Мат.
третьего Всесоюз. совещ.: Применение количественных методов при изучении
структуры растительности. Рига: Латв. ун-т, 1971. С. 266-267.
. Сёмкин Б.И. Об
эколого-географической структуре фиторазнообразия Дальнего Востока России //
Природоохранные территории и акватории Дальнего Востока и проблемы сохранения
биологического разнообразия: Мат. 2-ой науч. конф., посвящ. 60-летию со дня
организации Уссурийского заповедника. Владивосток: Дальнаука, 1994. С. 32-33.
. б Сёмкин Б.И. Общие
принципы введения мер различия, сходства и разнообразия в биоценологии //
Принципы и методы экспериментального изучения растительных сообществ. Л.:
Наука, 1972. С. 12-16.
. а Сёмкин Б.И. Основы
методов систематизации географических данных // Математические методы в
экологии и географии. Владивосток: ДВНЦ АН СССР, 1978. С. 3-11.
. б Сёмкин Б.И.
Предисловие // Теоретико-графовые методы в биогеографических исследованиях.
Владивосток: ДВНЦ АН СССР, 1983. С. 3-5.
. б Сёмкин Б.И.
Принципы автоматической систематизации биоценотических описаний // Третья
Всесоюз. конф. по биол. и мед. кибернетике. 1978. Т. 3. М.: ВЦ АН СССР, С.
325-328.
. Сёмкин Б.И. Рецензия
на книгу Песенко Ю.А. «Принципы и методы количественного анализа в
фаунистических исследованиях». М.: Наука, 1982. 278 с. // Известия АН СССР. се.
биол. №1. 1986. С. 158-159.
. б Сёмкин Б.И.
Сравнительный многомерный анализ структурной организации растительного покрова
// Автореф. дисс. докт. биол. наук. Л.: ДВО АН СССР, 1987. 34 с.
. в Сёмкин Б.И.
Теоретико-графовые методы в сравнительной флористике // Теоретические и
методические проблемы сравнительной флористики: Мат. II раб. совещ.
по сравнительной флористике. Неринга. 1983. Л.: Наука. 1987. С. 149-163.
. Сёмкин Б.И.
Эквивалентность мер близости и иерархическая классификация многомерных данных //
Иерархические классификационные построения в географической экологии и
систематике. Владивосток: ДВНЦ АН СССР, 1979. С. 97-112.
. Сёмкин Б.И.,
Варченко Л.И. Асимптотические оценки мер ассоциации Дайса и Иверсена //
Математические методы в экологии и географии. Владивосток: ДВНЦ АН СССР, 1978.
С. 12-20.
. а Сёмкин Б.И.,
Горшков М.В. Аксиоматическое введение мер сходства, различия, совместимости и
зависимости для компонентов биоразнообразия в многомерном случае // Вестник
КрасГАУ. 2009. №12. С. 18-24.
. б Сёмкин Б.И.,
Горшков М.В. О геометрической интерпретации коэффициента вариации и её
использование для определения мер разнообразия // Леса российского Дальнего
Востока: 150 лет изучения. Мат. Всерос. конф. посвящ. 100-летию со дня рожд.
члена-корр. РАН Колесникова Б.П. Владивосток: Дальнаука, 2009. С. 180-183.
. Сёмкин Б.И., Горшков
М.В. Об оценке сходства и различия в серии флористических и фитоценотических
описаний // Комаровские чтения. 2010. Вып. LVII. С. 203-220.
. Сёмкин Б.И., Горшков
М.В. Система аксиом симметричных функций двух переменных и меры, измеряющие
отношения сходства различия, совместимости и зависимости для компонентов
биоразнообразия // Вестник ТГЭУ. 2008. №4. С. 31-46.
. Сёмкин Б.И.,
Двойченков В.И. Об эквивалентности мер сходства и различия // Исследование
систем. Т. 1. Анализ сложных систем. Владивосток: ДВНЦ АН СССР, 1973. С 95-104.
. Сёмкин Б.И.,
Комарова Т.А. Анализ фитоценотических описаний с использованием мер включения
(на примере растительных сообществ долины реки Амгуэмы на Чукотке) // Бот. ж.
1977. Т. 62. № 1. С. 54-63.
. Сёмкин Б.И.,
Комарова Т.А. Использование мер включения при изучении вторичных сукцессий (на
примере послепожарных сообществ Сихотэ-Алиня) // Бот. ж. 1985. Т. 70. № 1. С.
89-97.
. Сёмкин Б. И.,
Куликова Л.С. Методика математического анализа списка видов насекомых в
естественных и культурных биоценозах. Владивосток: ТИГ ДВНЦ АН СССР, 1981. 73
с.
. Сёмкин Б.И., Наумова
В.В. Использование формулы Фишера для оценки дисперсий мер зависимости // Тез.
докл. Всесоюз. конф. «Теория классификаций и анализ данных». Ч. 2. Новосибирск:
ВЦ СО АН СССР, 1981. С. 9-11.
. Сёмкин Б.И., Ралько
В.Д. Моделирование ландшафтообразующих процессов переходных зон Дальнего
Востока // Исследование и конструирование ландшафтов Дальнего Востока и Сибири.
Владивосток: ДВО РАН, 1996. С. 150-163.
. Сёмкин Б.И.,
Соболева Т.А. Оценка темпов изменения общей численности населения городов
Приморского края // География и природные ресурсы. №4. 2005. С. 118-123.
. Сёмкин Б.И.,
Усольцева Л.А. Об использовании метода анализа разнообразий при исследовании
взаимосвязи растительности и среды // Локальный мониторинг растительного
покрова. Владивосток: ДВНЦ АН СССР, 1982. С. 54-60.
. Сёмкин Б.И., Ююкин
Н.А. Общие принципы определения зависимости явлений // Докл. на симп. XXIII
Международного конгр. «Количественные методы в географии». М.: МГУ, 1976. С.
8-12.
. Сёмкин Б.И.,
Варченко Л.И., Орешко А.П. Теоретические и методические подходы к учёту и
охране биологического разнообразия Дальнего Востока с помощью баз данных //
Тез. докл. II совещания «Компьютерные базы данных в ботанических
исследованиях». СПб.: 1995. С. 52.
. Сёмкин Б.И., Горшков
М.В., Варченко Л.И. Об изменениях содержания воды в однолетних побегах хвойных
древесных растений в умеренной климатической зоне // Сибирский экол. журн.
2008. №4. Т. 15. С. 537-544.
. Сёмкин Б.И., Майоров
И.С., Горшков М.В. Микрорезерваты биоразнообразия в береговой зоне Японского
моря (Приморский край) и их включение в государственный кадастр особо
охраняемых территорий // III экол. форум «Природа без границ». Владивосток: Рея,
2008. С. 108.
. а Сёмкин Б.И.,
Орешко А.П., Горшков М.В. БИС «Биоразнообразие» и её использование при изучении
структурной организации растительного покрова // Мат. XXXIII Всерос. семинара
«Актуальные вопросы охраны окружающей среды. Программное обеспечение для
экологов». М.: 2005. С. 14-16.
. б Сёмкин Б.И.,
Орешко А.П., Горшков М.В. Биоинформационная система (БИС) «Биоразнообразие» и
её использование // Мат. Всерос. науч.-практ. конф. «Информационные технологии
в науке, образовании и экономике». Якутск: 2005. С. 6.
. Сёмкин Б.И., Орешко
А.П., Горшков М.В. Биоинформационная система "Биоразнообразие" и её
использование в научных и учебных целях // Мат. XIII науч. совещ. географов
Сибири и Дальнего Востока. Т.1. Иркутск: Ин-т геогр. им. В.Б. Сочавы СО РАН,
2007. С. 195.
. а Сёмкин Б.И.,
Орешко А.П., Горшков М.В. Об использовании биоинформационных технологий в
сравнительной флористике. I. Схемно-целевой подход. Абсолютные меры сходства и
различия // Бюлл. БСИ ДВО РАН [Электронный ресурс] : науч. журн. / Ботан.
сад-институт ДВО РАН. - Владивосток, 2009. Вып. 3. С. 102-111.
http://www.botsad.ru/journal/ number3/02.pdf.
. б Сёмкин Б.И.,
Орешко А.П., Горшков М.В. Об использовании биоинформационных технологий в
сравнительной флористике. II. Меры включения дескриптивных множеств и их
использование // Бюлл. БСИ ДВО РАН [Электронный ресурс] : науч. журн. / БСИ ДВО
РАН. - Владивосток, 2009. Вып. 4. С. 58-70.
http://www.botsad.ru/journal/number4/number4_58-70.pdf.
. Сёмкин Б.И., Орешко
А.П., Горшков М.В. Об использовании биоинформационных технологий в
сравнительной флористике. III. Меры сходства и различия дескриптивных множеств
// Бюлл. БСИ ДВО РАН [Электронный ресурс] : науч. журн. / Ботан. сад-институт
ДВО РАН. - Владивосток, 2010. Вып. 6. С. 76-89.
http://botsad.ru/journal/number6/76-89.pdf.
. Сёмкин Б.И.,
Степанова К.Д., Щербова М.А. Влияние скашивания на видовую структуру
крупнотравного сообщества на Камчатке // Бот. ж. 1973. Т. 58. № 5. С. 665-670.
. Сёмкин Б.И., Гарбузова
Г.Ф., Майоров И.С., Горшков М.В. Использование информационных системных
береговых кадастров многоцелевого назначения для управления и мониторинга
побережья залива Петра Великого // Региональная экономика: теория и практика.
2009. №8 (101). С. 9-13.
. Сёмкин Б.И., Киселёв
А.Н., Бочарников В.Н., Краснопеев С.М. Бассейновый подход в оценке
биоразнообразия на основе ГИС-технологий // Тез. докл. II совещания
«Компьютерные базы данных в ботанических исследованиях». СПб.: 1995. С. 53.
. а Сёмкин Б.И.,
Клочкова Н.Г., Гусарова И.С., Горшков М.В. Дискретность и континуальность флор
водорослей-макрофитов дальневосточных морей России. I. Общий видовой состав //
Изв. ТИНРО. 2010. Т.160. С. 57-70.
. в Сёмкин Б.И.,
Клочкова Н.Г., Гусарова И.С., Горшков М.В. Дискретность и континуальность флор
водорослей-макрофитов дальневосточных морей России. III. Таксономические
спектры // Изв. ТИНРО. 2010. Т. 163. С. 217-227.
. Сёмкин Б.И., Селедец
В.П., Майоров И.С., Горшков М.В. Использование сравнительного
анализа растительного покрова для кадастровых оценок ботанических памятников
природы // Вестник КрасГАУ. 2009. №4. С. 115-120.
. Сёмкин Б.И.,
Тимофеев И.В., Варченко Л.И., Орешко А.П. Сравнительный анализ флоры с
использованием баз данных // Тез. докл. III совещания
«Компьютерные базы данных в ботанических исследованиях». СПб.: 1997. С. 90-93.
. Семкин Б.И.,
Петропавловский Б.С., Кошкарев А.В., Варченко Л.И., Усольцева Л.А. О методе
многомерного анализа соотношения растительности с экологическими факторами //
Бот. ж., 1986. Т. 71. № 9. - С. 1167-1981.
. Сёмкин Б.И., Селедец
В.П., Горшков М.В., Дробот Я.А., Борзова Л.М. Сравнительный анализ ботанических
памятников природы в береговой зоне Японского моря (Приморский край) // Мат.
XIII науч. совещ. географов Сибири и Дальнего Востока. Т.2. Иркутск: Ин-т
геогр. им. В.Б. Сочавы СО РАН, 2007. С. 40.
. Сёмкин Б.И., Селедец
В.П., Борзова Л.М., Варченко Л.И., Майоров И.С., Горшков М.В. Методы
сравнительного анализа компонентов биоразнообразия ботанических памятников
природы // Бот. ж. 2010. №3. С. 408-421.
. Слинченкова Е.Ф.
Криофитно-степные сообщества среднего течения р. Амгуэмы (перешеек Чукотского
полуострова) // Бот. ж. 1984. Т. 69. № 11. С. 1509-1519.
. Смирнов В.С.
Изменчивость биологических явлений и коэффициент вариации // Журн. общ. биол.
1971. Т. 32. № 2. С .152-161.
. Снетков М.А.,
Вавилин В.А. Оценка степени загрязненности водоёмов по интегральным показателям
качества воды // Научные основы контроля вод по гидробиологическим показателям.
Л. Гидрометеоиздат, 1977. С. 65-78.
. Сокал Р.Р.
Кластер-анализ и классификация: предпосылки и основные направления //
Классификация и кластер. Под ред. Дж. Вэн Райзина. М.: Мир, 1980. С. 7-19.
. Соколова М.В.
Количественные сравнения восьми конкретных флор Таймыра по их таксономической
структуре (Арктическая Средняя Сибирь) // Бот. ж. 1984. Т. 69. № 6. С. 840-849.
. Соломон Г. Зависящие
от данных методы кластер-анализа // Классификация и кластер. Под ред. Дж. Вэн
Райзина. М.: Мир, 1980. С. 129-148.
. Сохранение и
восстановление биоразнообразия / Колл. авторов. - М.: Научный и
учебно-методический центр, 2002. - 286 с.
. Степанов В.В. Курс
дифференциальных уравнений. - М.: Физматлит, 1959. - 468с.
. Сукачёв В.Н.
Руководство к исследованию типов лесов. - М.-Л.: Сельколхозгиз, 1931. - 328 с.
. Суппес П., Зиннес Д.
Основы теории измерений // Психологические измерения. М.: 1967. С. 9-110.
. Суханов В.В. Модель
распределения видового обилия на литорали островной гряды // Морская
биогеография. Предмет, методы, принципы районирования. М.: Наука, 1982. С. 52-75.
. Суханов В.В., Жуков
В.Е. Оценивание видового богатства сообщества по его видовой структуре на
примере макрофитов бухты Витязь Японского моря // Биология моря. 2010. Т.36.
№3. С. 221-227.
. Сэлтон Г.А.
Автоматическая обработка, хранение и поиск информации. - М.: Сов. радио, 1973.
- 560 с.
. Тамарин П.В., Шмидт
В.М. Сравнительный анализ некоторых коэффициентов сходства // Тр. Ленингр.
общ-ва естествоиспытателей «Успехи биометрии». Л.: ЛГУ, 1975. С. 45-54.
. Терентьев П.В.
Дальнейшее развитие метода корреляционных плеяд // Применение математических
методов в биологии. Т. 1. Л.: 1960. С 42-58.
. Терентьев П.В.
Математизация биологии // Тр. Ленингр. общ-ва естествоиспытателей «Успехи
биометрии». Л.: ЛГУ, 1975. С. 5-8.
. Терентьев П.В. Метод
корреляционных плеяд // Вестн. ЛГУ. № 9. 1959. С. 35-43.
. Толмачёв А.И.
Введение в географию растений. - Л.: ЛГУ, 1974. - 244 с.
. Толмачёв А.И. К
методике сравнительно-флористических исследований. 1. Понятие о флоре в
сравнительной флористике // Журн. Рус. бот. о-ва. 1931. Т. 16. № 1. С. 111-124.
. Толмачёв А.И. Методы
сравнительной флористики и проблемы флорогенеза. - Новосибирск: Наука, 1986. -
196 с.
. Трасс Х.Х.
Геоботаника. История и современные тенденции развития. - Л.: Наука, 1976. - 252
с.
. Трасс Х.Х. Развитие
количественно-статистических методов изучения растительного покрова // Бот. ж.
1971. Т. 56. №4. С. 457-464.
. Уиттекер Р.
Сообщества и экосистемы. - М.: Прогресс, 1980. - 327 с.
. Уланова Н.Г.
Математические методы в геоботанике. - M.: МГУ, 1995. - 109 с.
. Урбах В.Ю.
Биометрические методы. - М.: Наука, 1964. - 416 с.
. Урбах В.Ю.
Математическая статистика для биологов и медиков. - М.: Наука, 1963. - 324 с.
. Усольцева Л.А.
Моделирование растительности на картах с использованием средств автоматизации
// Автореф. дисс. канд. геогр. наук. Институт геофизики АН СССР. - Киев: 1985.
- 20 с.
. Устинов В.А.,
Фелингер А.Ф. Историко-социальные исследования, ЭВМ и математика. - М.: Мысль,
1974. - 325 с.
. Фёдоров В.Д. О
методах изучения фитопланктона и его активности. - М.: МГУ, 1979. - 168 с.
. Фёдоров В.Д.,
Гильманов Т.Г. Экология. - М.: МГУ, 1980. - 464 с.
. Фёдоров В.В.,
Смирнов Н.А. Измерение экологического сходства // Изв. АН СССР. сер. биол. № 5.
1986. С. 762-772.
. Федотенков А.В.
Информационные основы сохранения биологического разнообразия на территориях
проживания коренных малочисленных народов России // Автореф. канд. дисс. биол.
наук. - Владивосток: 2005. - 26 с.
. Филатова Л.Д.
Использование мер включения для сравнения комплексов стафилинид в разных
местообитаниях // Теоретико-графовые методы в биогеографических исследованиях.
Владивосток: ДВНЦ АН СССР, 1983. С. 66-77.
. Филипченко Ю.А.
Изменчивость и методы её изучения. - М.-Л.: Наука, 1929. - 275 с.
. Фихтенгольц Г.М.
Курс дифференциального и интегрального исчисления. Т.1. - М.-Л.: Гостехлит,
1947. - 680 с.
. Фишер Р.А.
Статистические методы для исследователей. - М.: Госсметиздат, 1958. - 286 с.
. Фрей Т.Э.-А.
Биоценометрия - область науки, нуждающаяся в определении // Количественные
методы анализа растительности. Тарту: ТГУ, 1969. С. 95-97.
. Фрей Т.Э.-А.
Использование некоторых принципов техники факторного анализа при выделении
классификационных единиц // Применение количественных методов при изучении
структуры фитоценозов. М.: Наука, 1972. С. 11-16.
. Фридланд В.М.,
Рожков В.А. Применение математических методов и ЭВМ для классификации почв //
Геогр. сборник. № 5. М.: 1975. С. 336-369.
. Харари Ф. Теория
графов. - М.: Едиториал УРСС, 2003. - 296 с.
. Харвей Д. Научное
объяснение в географии. - М.: Прогресс, 1974. - 503 с.
. Хеллоуэл Д.М.
Сравнительный обзор методов анализа данных в биологическом надзоре // Научные
основы контроля вод по гидробиологическим показателям. Л. Гидрометеоиздат,
1977. С. 108.
. Хинчин А.Я. Цепные
дроби. - М.: Физматлит, 1961. 112 с.
. Хоментовский П.А.
Насекомые ксилофаги хвойных пород Камчатки. - Владивосток: ДВНЦ АН СССР, 1983.
- 176 с.
. Хорева М.Г. Флора
островов Северной Охотии. - Магадан: ИБПС ДВО РАН, 2003. - 173 с.
. Цыбулин С.М.
Зимне-весеннее население птиц некоторых ландшафтов Приобской лесосистемы //
Проблемы зоогеографии и истории фауны. Новосибирск: Наука, 1980. С. 170-190.
. Чернов Ю.И. О
некоторых индексах, используемых при анализе структуры животного населения суши
// Зоол. журн. Т. 50. вып. 7. 1971. С 1079-1091.
. Чернов Ю.И.
Природная зональность и животный мир суши. - М.: Мысль, 1975. - 224 с.
. Чернышев В.Д. О
спектральном диффузном отражении света хвои древесных видов Дальнего Востока //
Физиология растений. Т. 20. вып. 2. 1973. С. 321-325.
. Шадрина В.И.
Пространственно-типологическая структура населения птиц Забайкалья //