Особенности глюконеогенеза млекопитающих
Министерство
образования и науки Российской Федерации
Федеральное
Государственное
бюджетное
образовательное учреждение
Высшего
профессионального образования
Воронежский
Государственный Педагогический Университет
Реферат
на тему:
«Особенности глюконеогенеза млекопитающих»
Введение
Глюконеогене́з
- метаболический путь, приводящий к образованию глюкозы из неуглеводных
соединений (в частности, пирувата). Наряду с гликогенолизом, этот путь
поддерживает в крови уровень глюкозы, необходимый для работы многих тканей и
органов, в первую очередь, нервной ткани и эритроцитов. Он служит важным
источником глюкозы в условиях недостаточного количества гликогена, например,
после длительного голодания или тяжёлой физической работы. Глюконеогенез
является обязательной частью цикла Кори (совокупность биохимических
ферментативных процессов транспорта лактата из мышц в печень, и дальнейшего
синтеза глюкозы из лактата, катализируемое ферментами глюконеогенеза), кроме
того, этот процесс может быть использован для превращения пирувата,
образованного при дезаминировании аминокислот аланина и серина.
Глюконеогенез происходит у животных, растений,
грибов и микроорганизмов. Его реакции одинаковы для всех тканей и биологических
видов. Важными предшественниками глюкозы у животных выступают трёхуглеродные
соединения, такие как лактат, пируват, глицерол, а также некоторые
аминокислоты. У млекопитающих глюконеогенез происходит главным образом в
печени, в меньшей степени - в корковом слое почек и эпителиальных клетках,
выстилающих тонкую кишку. Образованная в ходе глюконеогенеза глюкоза уходит в
кровь, откуда доставляется к другим тканям. После интенсивной физической работы
лактат, образовавшийся при анаэробном гликолизе в скелетных мышцах,
возвращается в печень и превращается там в глюкозу, которая снова поступает в
мышцы или превращается в гликоген (этот круговорот известен как цикл Кори). У
проростков растений запасённые в семени жиры и белки превращаются, в том числе
и через глюконеогенез, в дисахарид сахарозу, который транспортируется по всему
развивающемуся растению. Глюкоза и её производные служат предшественниками для
синтеза растительной клеточной стенки, нуклеотидов, коферментов и многих других
жизненно важных метаболитов. У многих микроорганизмов глюконеогенез начинается
с простых органических соединений, содержащих два или три атома углерода,
например, ацетата, лактата и пропионата, которые содержатся в питательной
среде.
Хотя реакции глюконеогенеза одинаковы у всех
организмов, соседние метаболические пути и регуляторные пути глюконеогенеза
отличаются у различных видов и в различных тканях. Рассмотрим подробнее
особенности глюконеогенеза млекопитающих.
Глава 1. Особенности глюконеогенеза
млекопитающих
.1 Понятие глюконеогенеза
Глюконеогенез - синтез глюкозы из неуглеводных
продуктов. Такими продуктами или метаболитами являются в первую очередь
молочная и пировиноградная кислоты, так называемые гликогенные аминокислоты,
глицерол и ряд других соединений. Иными словами, предшественниками глюкозы в
глюконеогенезе может быть пируват или любое соединение, превращающееся в
процессе катаболизма в пируват или один из промежуточных продуктов цикла
трикарбоновых кислот.
Глюконеогенез и гликолиз не являются полностью
идентичными процессами, протекающими в противоположных направлениях, хотя
несколько стадий являются общими для обоих процессов: 7 из 10 ферментативных
реакций глюконеогенеза обратны соответствующим реакциям гликолиза. Однако 3
реакции гликолиза необратимы и не могут использоваться в глюконеогенезе:
образование глюкозо-6-фосфата из глюкозы под действием фермента гексокиназы,
фосфорилирование фруктозо-6-фосфата до фруктозо-1,6-бифосфата
фосфофруктокиназой-1, а также превращение фосфоенолпирувата в пируват под
действием пируваткиназы. При глюконеогенезе три необратимые стадии гликолиза
заменены «обходными» реакциями, катализируемыми другими ферментами, и эти
реакции также необратимы. Таким образом, в клетках как гликолиз, так и
глюконеогенез являются необратимыми процессами. У животных гликолиз происходит
только в цитозоле, как и большая часть реакций глюконеогенеза, хотя некоторые
его реакции происходят в митохондриях и эндоплазматическом ретикулуме. Это даёт
возможность для их координированной и взаимно обратной регуляции. Регуляторные
механизмы, различающиеся у гликолиза и глюконеогенеза, действуют на
ферментативные реакции, уникальные для каждого процесса.
Основные стадии гликолиза:
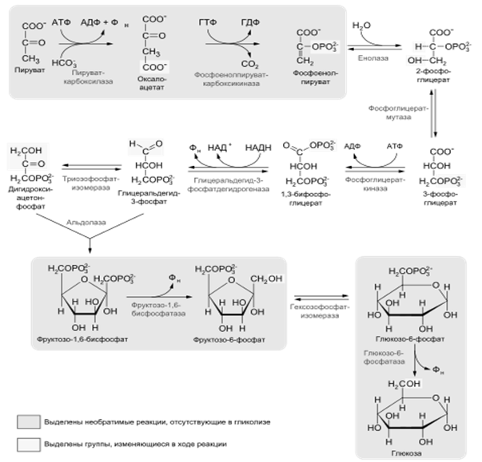
.2 Обходные реакции глюконеогенеза
Образование фосфоенолпирувата из пирувата.
Синтез фосфоенолпирувата осуществляется в несколько этапов. Первоначально
пируват под влиянием пируваткарбоксилазы и при участии СО2 и АТФ
карбоксилируется с образованием оксалоацетата:

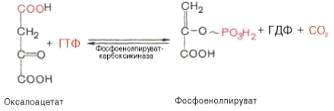
Установлено, что в процессе образования
фосфоенолпирувата участвуют ферменты цитозоля и митохондрий.
Первый этап синтеза протекает в митохондриях.
Пируват-карбоксилаза, которая катализирует эту реакцию, является
аллостери-ческим митохондриальным ферментом. В качестве аллостерического
активатора данного фермента необходим ацетил-КоА. Мембрана митохондрий
непроницаема для образовавшегося оксалоацетата. Последний здесь же, в
митохондриях, восстанавливается в малат:

Реакция протекает при участии митохондриальной
НАД-зависимой малатдегидрогеназы. В митохондриях отношение НАДН/НАД+
относительно велико, в связи с чем внутримитохондриальный оксалоацетат легко
восстанавливается в малат, который легко выходит из митохондрии через
митохондриальную мембрану. В цитозоле отношение НАДН/НАД+ очень мало, и малат
вновь окисляется при участии цитоплазматической НАД-зависимой
малатдегидрогеназы:

Дальнейшее превращение оксалоацетата в
фосфоенолпируват происходит в цитозоле клетки.
Превращение фруктозо-1,6-бифосфата во
фруктозо-6-фосфат. Фосфо-енолпируват, образовавшийся из пирувата, в результате
ряда обратимых реакций гликолиза превращается во фруктозо-1,6-бифосфат. Далее
следует фосфофруктокиназная реакция, которая необратима. Глюконеогенез идет в
обход этой эндергонической реакции. Превращение фруктозо-1,6-бифосфата во
фруктозо-6-фосфат катализируется специфической фосфатазой:

Образование глюкозы из глюкозо-6-фосфата. В
последующей обратимой стадии биосинтеза глюкозы фруктозо-6-фосфат превращается
в глюкозо-6-фосфат. Последний может дефосфорилироваться (т.е. реакция идет в
обход гексокиназной реакции) под влиянием фермента глюкозо-6-фосфатазы:

1.3 Другие предшественники глюкозы
Описанный выше биосинтетический путь образования
глюкозы относится к синтезу глюкозы не только из пирувата, но также 4-, 5- и
6-углеродных промежуточных соединений цикла трикарбоновых кислот. Цитрат,
изоцитрат, α-кетоглутарат,
сукцинил-СоА, сукцинат, фумарат и малат - все промежуточные продукты цикла
трикарбоновых кислот могут окисляться до оксалоацетата. Некоторые или все атомы
углерода большей части аминокислот могут быть катаболизированы в пируват или
промежуточные соединения цикла трикарбоновых кислот. Поэтому эти аминокислоты
могут подвергнуться превращению в глюкозу и называются глюкогенными. Аланин и
глутамин - важнейшие молекулы, переносящие аминогруппы в печень из других
тканей - служат особенно важными глюкогенными аминокислотами у млекопитающих.
После того, как эти аминокислоты отдают свои аминогруппы в митохондриях печени,
их углеродные скелеты (пируват и α-кетоглутарат
соответственно) вовлекаются в глюконеогенез. Аминокислоты образуются при
распаде белков мышц и соединительной ткани, их включение в глюконеогенез
происходит при продолжительном голодании или длительной физической нагрузке.
У растений, дрожжей и многих бактерий имеется
путь, позволяющий получать углеводы из жирных кислот - глиоксилатный цикл. У
животных ключевых ферментов этого цикла нет, и, ввиду необратимости
пируватдегидрогеназной реакции, они не могут получать пируват из ацетил-СоА, а
значит, образовывать углеводы из жирных кислот (следовательно, и из липидов).
Тем не менее, они могут использовать для глюконеогенеза те небольшие количества
глицерола, который образуется при распаде жиров. При этом глицерол
фосфорилируется глицеролкиназой, далее следует окисление центрального атома
углерода с образованием дигидроксиацетонфосфата, который является промежуточным
соединением глюконеогенеза.
Глицеролфосфат является необходимым
промежуточным соединением при синтезе жиров (триглицеридов) в адипоцитах,
однако эти клетки лишены глицеролкиназы и поэтому не могут осуществлять
фосфорилирование глицерола. Вместо этого адипоциты могут осуществлять
сокращённый вариант глюконеогенеза, известный как глицеронеогенез:
преобразование пирувата в дигидроксиацетонфосфат через первые реакции
глюконеогенеза, вслед за которым следует восстановление дигидроксиацетонфосфата
до глицеролфосфата.
.4 Регуляция глюконеогенеза
В целом можно выделить два способа регуляции
глюконеогенеза: гормональная при участии гормонов и метаболическая, т.е. при
помощи промежуточных или конечных продуктов обмена глюкозы.
Гормональная активация глюконеогенеза
осуществляется глюкокортикоидами, которые увеличивают синтез
пируваткарбоксилазы, фосфоенолпируват-карбоксикиназы, фруктозо-1,6-дифосфатазы.
Глюкагон стимулирует те же самые ферменты через аденилатциклазный механизм
путем фосфорилирования.
Также имеется метаболическая регуляция, при
которой аллостерически активируется пируваткарбоксилаза при помощи ацетил-SКоА,
фруктозо-1,6-дифосфатаза при участии АТФ.
Гормональные и метаболические факторы,
регулирующие глюконеогенез:
При снижении использования лактата в качестве
субстрата для глюконеогенеза, которое может вызываться дефектом ферментов
глюконеогенеза, концентрация лактата в крови повышается, что приводит к
понижению pH крови и развитию лактатацидоза.
Лакта́тацидо́з
(лакта́тацидоти́ческая
ко́ма,
молочноки́слая ко́ма,
ги́перлакта́тацидеми́я)
- редкое крайне тяжёлое состояние, летальность при котором от 50 до 90% -
неспецифический синдром, развивающийся в ряде тяжёлых патологических состояний
(в том числе, сахарный диабет), когда создаются предпосылки для повышенного
образования и накопления в крови и периферических тканях молочной кислоты.
Обычно развивается остро (в течение нескольких
часов), предвестники, как правило, отсутствуют или не характерны. Пациенты
могут отмечать мышечную боль, боль за грудиной, диспептические явления,
учащение дыхания, апатию, сонливость или бессонницу. Однако превалирующими
симптомами клинической картины лактатацидоза являются проявления
сердечно-сосудистой недостаточности, усугубляющиеся тяжёлым ацидозом, на фоне
которого наступают изменения сократительной способности миокарда. В динамике
состояние пациентов прогрессивно ухудшается: по мере нарастания ацидоза может
появиться боль в животе, рвота. Выявляется разнообразная неврологическая
симптоматика от арефлексии до спастических парезов и гиперкинезов. Перед
развитием комы (потери сознания), несмотря на отсутствие запаха ацетона в
выдыхаемом воздухе (нет кетонемии), наблюдается шумное дыхание Куссмауля,
обычно сопровождающее выраженный метаболический ацидоз. Развивается коллапс с
олиго-, а затем и анурией, гипотермией. На этом фоне развивается ДВС-синдром
(синдром внутрисосудистого свёртывания крови), нередки внутрисосудистые
тромбозы с геморрагическими некрозами пальцев рук и ног. Быстрое развитие
лактатацидоза (несколько часов) не способствует выявлению признаков,
характерных для диабетической комы (сухость кожи, слизистых оболочек и языка).
У 10-30% пациентов с диабетической и гиперосмолярной комой присутствуют
элементы лактатацидоза
Стоит отметить, что кратковременный лактатацидоз
встречается и у здоровых людей при интенсивной мышечной работе, в этом случае
он компенсируется путём гипервентиляции лёгких и ускоренным выведением
углекислого газа.
биохимический глюкоза фермент
Заключение
Так как синтез и окисление глюкозы являются
крайне важными для существования клетки (гликолиз) и всего организма
(глюконеогенез), то регуляция этих процессов отвечает запросам органов и тканей
при различных условиях существования.
Глюконеогенез, как образование глюкозы из
неуглеводных компонентов, необходим:
) При гипогликемии во время мышечной нагрузки -
синтез из молочной кислоты, поступающей из мышц, из глицерола, образующегося
при мобилизации жиров;
) При гипогликемии при голодании - синтез из
аминокислот, образующихся при катаболизме белков.
Таким образом, при голодании или физической
нагрузке глюконеогенез, идущий в печени, обеспечивает глюкозой все остальные
органы (эритроциты, нервная ткань, мышцы и др.), в которых активен гликолиз и
другие процессы, производящие энергию. Наличие глюкозы в указанных клетках
необходимо, чтобы поддержать концентрацию оксалоацетата и обеспечить сгорание
ацетил-SКоА (получаемого также из жирных кислот или кетоновых тел) в цикле
трикарбоновых кислот.
Список использованной литературы
1. Биохимия: Учеб. для вузов, Под
ред. Е.С. Северина. - Москва: Аспект пресс, - 2004. - 779 с.
. Ефимов А.С. Клиническая
диабетология. /А.С. Ефимов, Н.А. Скробонская /Киев: Здоровье, 2008.. - 320 с.
. Кольман Я. Наглядная биохимия./ Я.
Кольман, К. Рём/ - 4-е изд. - Москва: БИНОМ. Лаборатория знаний, 2012. - 469 с.