|
Знак
|
Типвзаємодії
|
Підтип
|
|
α
|
β
|
|
|
|
-
|
-
|
Конкуренція
|
Експлуатаційна
(без витрат енергії на взаємодії)
|
|
|
|
Інтерференційна
(з витратою енергії)
|
|
+
|
-
|
Експлуатація
|
Голофагія
або істинне хижацтво
|
|
|
|
Мерофагія
або пасовищне хижацтво (харчування частинами)
|
|
|
|
Паразітоїдність
|
|
|
|
Паразитизм
|
|
+
|
+
|
Симбіоз
|
|
|
|
Мутуалізм
(облігатне, нерозривна взаємодія)
|
|
-
|
0
|
Аменсалізм
|
|
+
|
0
|
Комменсалізм
|
|
0
|
0
|
Нейтралізм
|
Оригінальна класифікація
відносин між видами запропонована російським зоологом В.М. Беклемішевим. Їм
виділялися топічні зв'язки (виражаються в зміні середовища проживання),
трофічні зв'язки (харчування особин одного виду особинами іншого, а також їх
залишками і продуктами життєдіяльності), фабричні зв'язки (пов'язані з наданням
середовища або притулку) і форичні зв'язки (перенесення особинами одних видів
особин інших).
Розділ 2. Симбіоз
Симбіоз - взаємодія і співіснування різних
біологічних видів. Наука про симбіоз називається симбіологія.
У природі зустрічається широкий спектр прикладів
взаємовигідного симбіозу. Від шлункових і кишкових бактерій, без яких було б
неможливе травлення, до рослин (часто орхідеї), чий пилок може поширювати лише
один, певний вид комах. Такі відносини завжди успішні, коли вони збільшують
шанси обох партнерів на виживання. Здійснювані в ході симбіозу дії або
вироблювані речовини є для партнерів істотними і незамінними. У узагальненому
розумінні такий симбіоз - проміжна ланка між взаємодією і злиттям.
У ширшому науковому розумінні симбіоз є будь-якою
формою взаємодії між організмами різних видів, зокрема паразитизм - відносини,
вигідні одному, але шкідливі іншому симбіонту. Обопільно вигідний вид симбіозу
називають мутуалізмом. Коменсалізмом називають відносини, корисні одному, але
байдужі іншому симбіонту, а аменсалізмом - відносини, шкідливі одному, але
байдужі іншому.
Різновид симбіозу - ендосимбіоз, коли один з
партнерів живе усередині клітини іншого.
.1 Мутуалізм
Мутуалізм - це облігатні
взаємовигідні відносини між популяціями. Мутуалізм - поширене явище. Розвинене
наземне життя існує лише завдяки мутуалістичним стосункам. Саме заселення
вищими рослинами суші відбувалося завдяки їх тісній взаємодії з грибами. На
залишках девонських рослин знайдені сліди мікоризи. Мінеральне живлення
наземних рослин - складний процес, і в багатьох випадках воно неможливе без
грибів. При мікориі гіфи грибів охоплюють чохлом коріння рослин, проникають
всередину цих коренів або навіть всередину окремих клітин. Гриби мають значно
більш ефективний механізм отримання мінеральних речовин, ніж рослини, але
обмежені нестачею органіки. Рослини передають частину синтезованої ними
органіки гіфам гриба, отримуючи від них необхідні солі.
У мінеральному живленні
беруть участь і ендосімбіотичні азотфіксатори, на зразок бактерій Rhizobium. На
коренях ряду рослин (бобових, вільхи і т.д.) формуються спеціальні губчасті
розростання, які населяються бактеріями, здатними зв'язувати атмосферний азот.
Такий процес приносить користь не тільки самій рослині-симбіонту, а й іншим
рослинам, оскільки призводить до збагачення грунту азотом.
Отже, розвинений наземний
рослинний покрив - результат мутуалізму рослин з грибами і бактеріями. Але і
споживання рослинної біомаси в істотному ступені пов'язано з процесами цієї ж
категорії. Всі рослиноїдні ссавці в тій чи іншій мірі використовують
ендосимбіотічну мікрофлору своїх кишок. Найвищого розвитку ендосимбіотічні
комплекси досягають у жуйних парнокопитних. Шлунок цих тварин складається з
кількох відділів, один з яких - рубець - є, по суті, мікробіологічним
ферментером - ємністю для вирощування мікроорганізмів.
Мікрофлора і мікрофауна
рубця жуйних складна і до кінця не вивчена. Її суттєвими компонентами є як
найпростіші (наприклад, джгутиконосці і інфузорії), так і бактерії. Результатом
роботи цього комплексу є глибока переробка целюлози.
Які організми можуть
стати мутуалістичними? Обов'язково дуже різні. Якщо екологічні ніші двох
популяцій подібні, тобто цим популяціям необхідні однакові ресурси, між ними
виникне конкуренція. Умова, за якої може розвинутися мутуалізм і
протокооперація, - розходження ніш. Найчастіше мутуалізм розвивається тоді,
коли кожен член пари видів віддає напарнику менш цінний для себе ресурс, а
отримує більш цінний. Оптимальною є ситуація, при якій ресурсом для одних
організмів є відходи інших, і навпаки.
Як частинний випадок
мутуалізму можна розглядати протокооперацію. Це необов'язкові взаємовигідні
відносини між двома популяціями. Пов'язані цими відносинами види можуть зустрічатися
як разом, так і окремо.
Найчастіше грань між
протокооперацією і мутуалізмом провести нелегко. Якщо спостерігається глибока
спеціалізація між певними видами, такі відносини треба класифікувати як
мутуалістичні. Якщо, наприклад, одні й ті ж рослини можуть запилюватися різними
комахами, а ті харчуються пилком і нектаром різних видів рослин і можуть
використовувати іншу їжу, слід говорити про протокооперацію. Іноді
спостерігаються проміжні між цими двома категоріями відносини, коли один з
видів може зустрічатися і без другого, а другий у своєму поширенні цілком
залежить від першого.
2.2 Коменсалізм
Коменсалізмом називаються
прямі або опосередковані через середовище відносини між двома популяціями, від
яких одна - популяція комменсала - отримує вигоду (збільшує свою чисельність у
відповідь на збільшення чисельності партнера), а інша - популяція господаря -
індиферентна до цих відносин і не залежить від чисельності комменсала. З цього
визначення випливає, що вигода, яку отримує комменсал, не пов'язана з якимись
спеціальними витратами з боку господаря, а є наслідком його нормальної
активності або життєдіяльності. Найчастіше комменсализм пов'язаний майже
безперервними переходами з протокооперацією (якщо господар починає отримувати
вигоду від свого партнера) або з паразитизмом (якщо господар відчуває шкоду).
Класифікувати коменсалізм
можна за чотирма проявами:
.2.1 Їжа
Комменсал може
харчуватися відмерлими господарями або їх частинами, а також відходами їх
життєдіяльності. Так, копрофаги харчуються відходами більших тварин, а
некрофаги - їх трупами. Песці харчуються недоїдками білих ведмедів, а
риби-лоцмани - недоїдками акул.
.2.2 Притулок і захист
У норах бабаків живуть
різноманітні жуки, метелики, жаби, мокриці і багато інших тварин. Горобці в'ють
свої гнізда поруч з гніздами орлів та інших великих хижих птахів - ті не
підпустять до своїх (а заодно і до гороб'ячих) кладок ворін, які розоряють
гнізда. Риби морські качечки ховаються між голками морських їжаків. У прикладах
з цієї дуже численної категорії комменсал отримує від хазяїна саме безпечний
притулок.
.2.3 Переміщення
Переміщуючись, тварини
здатні переносити на собі дрібні організми. Будинкові мухи і прусаки, що
харчуються нашою їжею, переносять на своїх тілах цілий комплекс бактерій і
грибів.
До цієї ж категорії слід
відносити явище лоцманювання - супроводу дрібними тваринами більших. Справа в
тому, що навколо великого тіла, що пливе під водою, виникають турбулентності,
що полегшує плавання. І риби (в тому числі риби-лоцмани, що дали назву цьому
явищу), що супроводжують акулу, і дельфіни, що пливуть поряд з швидкохідним
кораблем, використовують цей ефект.
.2.4 Зоохорія
Широко відомий феномен
зоохорії - перенесення тваринами насіння рослин. Іноді рослини «зацікавлюють»
тварин, надаючи плодам м'ясисті частини, які потенційні переносники
використовують в їжу, а іноді просто використовують тварин, формуючи чіпкі або
клейкі плоди та насіння.
2.3 Протокооперація
Форма симбіозу, при якій спільне існування
вигідне для обох видів, але не обов'язкове для них. У цих випадках відсутній
зв'язок саме цієї, конкретної пари партнерів.
Прикладом протокооперації є взаємини дрібних
рибок сімейства Губанових і великих хижих мурен. Серед Губанових є так звані
риби-чистильники, що звільняють великих риб від зовнішніх паразитів, що
знаходяться на шкірі, в зябровій і ротовій порожнинах. Великі хижаки, в тому
числі мурени, які страждають від паразитів, припливають в місця проживання
губанів і дають їм можливість знищувати паразитів навіть у себе в роті, хоча
могли б з легкістю їх проковтнути.
Розділ 3. Антібіоз
.1 Різноманітність форм експлуатації
Виділяють 4 форми
експлуатації: справжнє хижацтво (голофагія), пасивне хижацтво (мерофагія),
паразитоїдність і паразитизм.
Голофагія. Голофагі, істинні хижаки, вбивають
жертву відразу і з'їдають за своє життя безліч жертв (лев, вовк, дафнія,
кашалот, росичка).
Мерофагія. Мерофагі, пасивні хижаки (корова,
ґедзь, медична п'явка) зазвичай з'їдають тільки частину жертви, завдаючи їй при
цьому певний, але не обов'язково смертельний збиток. За своє життя мерофагі
експлуатують багатьох жертв, з якими не мають особливо тісних зв'язків.
Паразитизм. Паразити (солітер, ВІЛ, омела біла,
тля) тісно пов'язані зі своїм господарем, забирають у нього лише частину його
ресурсів і не обов'язково спричинюють смерть. Їх зв'язок з господарем дуже
тісний; в типовому випадку особина паразита все своє життя пов'язана з єдиним
господарем.
Паразитоїдність. Паразитоіди, до яких відносяться
перетинчастокрилі і деякі двокрилі, ведуть вільний спосіб життя, але відкладають
свої яйця в жертву, на неї або біля неї. Личинки поїдають жертву живцем, а
потім вбивають її: смерть жертви неминуча, але відстрочена. З урахуванням
чисельності комах взагалі і перетинчастокрилих зокрема, така стратегія не є
рідкісною.
3.2 Хижацтво
Найважливішою
характеристикою хижаків є те, наскільки вони спеціалізовані на харчуванні
певними видами жертв. За цим фактором їх можна розділити на генералистів, тобто
пластичних хижаків, і спеціалістів, які можуть харчуватися лише певними видами.
До цієї класифікації близьким є поділ хижаків на монофагів (пристосованих до
харчування одним видом жертв), олігофагів (орієнтованих на кілька близьких
категорій жертв) і поліфагів (здатних харчуватися дуже різноманітними
жертвами).
Ці класифікації не цілком
ідентичні, хоча, звичайно, спеціалізовані хижаки зазвичай є оліго- і
монофагами, а генералісти - поліфагами. Втім, буває так, що якесь досить вузьке
пристосування допомагає освоїти харчування групою схожих за своїх особливостям
жертв. Наприклад, гігантський мурахоїд високоспеціалізований на харчуванні
мурахами й термітами: його кігті дозволяють пробивати стінки термітників або
добиратися до мурашок, а довгий липкий язик дає можливість підхоплювати і
ковтати одночасно сотні комах. Нічим іншим, крім мурах і термітів, мурахоїд
харчуватися не здатний, але практично будь-який з досить численних видів цих
соціальних комах може стати жертвою цього хижака-олігофага.
Спеціалізований хижак
може бути більш ефективним, ніж неспеціалізований, і це дозволяє частково
уникати конкуренції. З іншого боку, поліфаги можуть мати більш збалансовану
дієту, легше знаходити корм, мати більш стійку популяційну чисельність.
Хижацтво - один з
найважливіших механізмів передачі енергії від одних популяцій до інших і є тому
важливим механізмом, що робить екосистему єдиним цілим.
.2.1 Зв'язок
"хижак-жертва"
Важлива характеристика
хижака-поліфага, здатного переключатися з однієї жертви на іншу, - його реакція
на зміну відносної частоти жертв (рис.1). Хижак з реакцією I типу вибірково
споживає певний вид жертв, навіть якщо вони рідкісні, і підвищує їх споживання
із зростанням їх поширеності. Хижак з реакцією II типу споживає жертв певного
виду пропорційно їх чисельності, поки вони рідкісні, а після якогось рівня
стабілізує їх експлуатацію (насичується).
Найпоширенішою, однак, є
реакція III типу: рідкісна жертва споживається непропорційно мало, і хижак
звертає на неї «увагу» тільки тоді, коли її чисельність сягає певної межі.
Поширеність третього типу реакцій хижаків-поліфагів призводить до того, що
хижаки вирівнюють чисельність своїх жертв, сильніше експлуатуючи більш поширені
види і даючи «перепочинок» рідким.
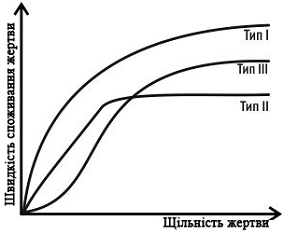
Рис. 1 Типи реакції
хижака-поліфага на зміну чисельності однієї з видів його жертв
У типовому випадку
популяції хижака стабілізують чисельність популяцій своїх жертв. Коли жертв
стає мало, хижаки-поліфаги шукають інші види для харчування, а хижаки-олігофаги
знижують свою чисельність слідом за жертвами в результаті недокорму. Оскільки в
більшості випадків хижаки розмножуються повільніше своїх жертв, відновлення їх
чисельності відбувається повільніше, ніж відновлення жертв.
Таким чином, у типовому
випадку хижаки просто не можуть знищити своїх жертв без залишку. Повністю
знищити певний вид своїх жертв може лише той хижак, чисельність якого мало
залежить від чисельності цього виду жертв. Значить, цей хижак повинен бути
поліфагом і не бути спеціалізованим на даному виді жертв.
3.3 Паразитизм
Справжні паразити тісно
пов'язані зі своїми господарями. Вони живуть усередині організму господарів або
прикріплені до їх поверхні. Зазвичай протягом життя (або етапу життєвого циклу)
паразит пов'язаний з одним господарем. Для того, щоб використовувати господаря,
паразитові зовсім не потрібно його вбивати (іноді господар гине, але це не є
необхідною умовою його вживання).
Господар є для паразита
не тільки джерелом енергії, а й середовищем. Наслідком цього є те, що паразит
відчуває набагато сильнішу зміну власної будови, ніж хижак. Така перебудова
може бути пов'язана зі спрощенням одних систем органів і, навпаки,
удосконаленням інших.
У більшості випадків
спеціалізовані паразити досить ефективно «обходять» захисні системи своїх
господарів. Це пов'язано з різною метою відбору: для паразита побороти
господаря - питання життя і смерті, а для господаря - лише економії деяких
ресурсів. Еволюційно старі паразити, добре пристосовані до своїх господарів,
зазвичай викликають неважкі захворювання. Однак якщо паразити зовсім не
експлуатуватимуть своїх господарів, вони не зможуть розмножуватися і заражати
нові організми. Оптимальна для паразита стратегія найчастіше полягає в тому,
щоб інтенсивно використовувати деяких особин хазяїна (слабких, старих, хворих,
а також представників інших видів, крім тих, з яким тісно пов'язаний паразит),
а інших особин експлуатувати слабо. Пристосуванням для втілення такої стратегії
виявляється здатність паразита знаходитися в двох фазах: активній і пасивній.
Перехід до активної фази може бути викликаний, наприклад, гормональними змінами
в організмі господаря (його тривалим важким стресом).
Найчастіше
найнебезпечніші паразити - ті, які перейшли до нового виду господарів і не
мають пристосувань для збереження його життя. Дуже показові результати отримані
в дослідах, в яких спільно вирощували культуру мух та експлуатуючих їх
оспаразитоїдів (рис. 2). Якщо разом зсаджували мух і ос, які раніше не
контактували один з одним, в експериментальній установці спостерігалися різкі
скачки чисельності обох видів. Якщо два взаємодіючих виду вирощувалися разом
протягом двох років, їх чисельність залишалася досить стабільною. Такий
результат відображає пристосування двох взаємодіючих популяцій одна до одної.
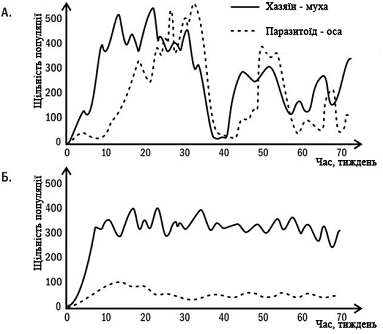
Рис. 2. Пристосування
господаря і паразітоїда один до одного. На графіках показана динаміка
чисельності двох видів двокрилих, яких вирощували в експериментальній установці
з 30 з'єднаних один з одним камер. А. Культури господаря і паразітоїда раніше
не контактували один з одним. Б. Культури господаря і паразітоїда спільно
вирощувались протягом двох років
3.4 Аменсалізм і нейтралізм
Аменсалізм - відносини,
при яких одна популяція відчуває несприятливий вплив іншої, а та ніяк не
залежить від першої. Такі відносини виникають, коли один з конкурентів
виявляється істотно сильніше іншого.
Велике дерево затінює
траву біля його підніжжя, майже не відчуваючи від цієї трави несприятливих
впливів. У тропічних океанах поширене явище, зване «червоним припливом».
Планктонні панцирні джгутикові водорості дінофлагелляти накопичують у собі
токсин, яким пригнічують конкурентів і захищаються від хижаків. Коли цих
водоростей стає багато, вода набуває червонуватого відтінку і стає небезпечна
для більшості живих організмів, включаючи риб і донну фауну.
Одним з типів відносин
популяцій є нейтралізм, при якому популяції не впливають один на одного. Це
означає, що ці дві популяції як би не існують одна для іншої.
Хоча на практиці ідеально
нейтральні відносини існувати не можуть, адже кожен організм споживає ресурс,
розсіює енергію, змінює середовище і тим самим, хоча б в невеликій мірі,
впливає на всі організми навколо себе. Але цей вплив може бути дуже малим -
нижче якогось порогового значення, після якого розглядати його вже недоцільно.
З цієї точки зору
нейтралізм - не відсутність взаємного впливу двох популяцій, а ситуація, коли
таким впливом можна знехтувати. Такі відносини виникають, коли між популяціями
немає прямих зв'язків, коли вони належать до різних трофічних ланцюгів.
Розділ 4. Конкуренція і екологічні ніші
Конкуренцією називається
таке відношення між двома популяціями, при якому зростання кожної з них
викликає зниження чисельності іншої. Втім, це визначення стосується тільки
міжвидової конкуренції, а крім неї існує також внутрішньовидова конкуренція,
яка розгортається всередині однієї популяції.
При конкуренції дві
популяції можуть несприятливо впливати одна на одну двома різними способами.
Перший (непряма, або експлуатаційна конкуренція) пов'язаний з тим, що дві
популяції використовують один і той же ресурс. При зростанні чисельності однієї
з цих популяцій посилиться споживання нею загального ресурсу, і іншій
дістанеться менша його кількість. Другий (пряма, або інтерференційна конкуренція)
пов'язаний з витратами енергії особин на заподіяння один одному шкоди.
Прикладом прямої конкуренції може бути алелопатія - виділення рослинами
речовин, що пригнічують інші види.
Це явище не могло б
спостерігатися, якби такі затрати не вирішували якусь важливу для конкуруючих
популяцій задачу. Це завдання - зменшення збитків від експлуатаційної
конкуренції. Отже, хоча пряма конкуренція, яка може супроводжуватися
конфліктами між особинами різних популяцій, здається більш «явною», вона є лише
наслідком прихованої, непрямої конкуренції.
4.1 Поняття про екологічні ніші
Чим більш подібні види,
тим сильніше перетинаються їхні потреби в ресурсах, тим гостріше буде
конкуренція між ними. Для опису конкуренції дуже корисним є поняття екологічної
ніші.
Вперше словосполучення
«екологічна ніша» ввів Дж. Ґрінелл в 1917 році. Він позначав таким чином
характерне місцепроживання виду, сукупність умов, в яких вид зустрічається в
природі. Ч. Ельтон в 1927 році визначив екологічну нішу як місце виду в
співтоваристві, його положення в структурі харчових взаємин.
На рис. 3 показана
двовимірна ніша синьо-зеленого мошколова. Його ніша охарактеризована за двома
параметрами: яку здобич ловить розглянутий вид птахів і де (на якій висоті) він
це робить. Можна припустити, що для будь-якого іншого виду птахів подібна
картина виглядала б інакше.
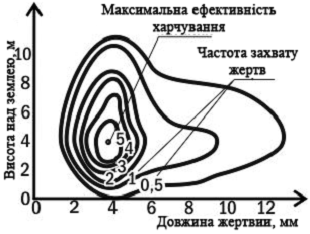
Тож екологічна ніша - це
комплексна характеристика способу життя виду, що включає споживані ним ресурси,
а також ті умови, за яких він може існувати.
Цінність поняття
екологічної ніші полягає в тому, що воно дозволяє порівнювати особливості
способу життя різних видів. Так, якщо два види можуть мешкати в подібних
умовах, але відрізняються за споживаним ними ресурсом (наприклад, за розміром
своїх жертв), ми можемо порівнювати ніші цих видів саме по цьому параметру.
За ту частину ресурсу,
яку споживає тільки один вид, конкуренція неможлива. Зате для тієї частини
ресурсу, яку можуть вживати два види (тобто частина, в якій ніші цих видів
перетинаються), між цими видами виникатиме конкуренція (рис. 4).
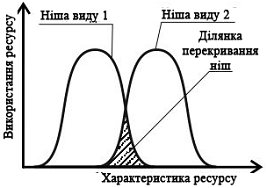
Рис. 4. Порівняння
екологічних ніш двох видів по тому параметру, за яким вони відрізняються
(наприклад, за розміром споживаних хижаками жертв)
Чим більш схожі два види,
тим сильніше перетинаються їхні потреби в ресурсах і тим сильніше конкуренція
між ними. Тому найгостріша конкуренція - внутрішньовидова. Але особини одного
виду, хоча і конкурують дуже гостро за ресурси, спільно формують наступне
покоління.
4.2 Експерименти Гаузе
У 1931-1935 роках
радянський біолог Георг Францевич Гаузе намагався експериментально перевірити
рівняння конкуренції, виведені В. Вольтерра. Гаузе зрозумів, що для
експериментів у галузі популяційної біології добре підходять найпростіші: їм
потрібний зовсім невеликий обсяг середовища, а зміна їх поколінь відбувається
набагато швидше, ніж у багатоклітинних організмів. Експерименти Гаузе, що
прославили його ім'я, виконані на інфузорії туфельці (Paramecium caudatum) і її
найближчих родичах (рис. 5).
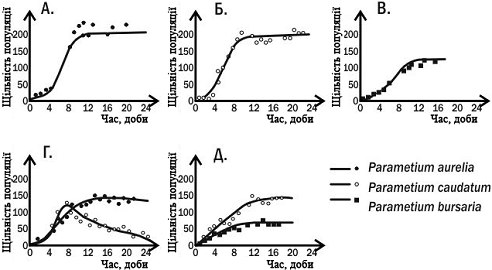
Рис. 5. Динаміка
чисельності трьох видів інфузорій в дослідах Г.Ф. Гаузе. У варіантах
експериментів А., Б. і В. види вирощували поодинці; у випадках Г. та Д. в
пробірку заселяли відразу два види інфузорій
Гаузе вирощував три види
інфузорій роду Paramecium в пробірках, куди він додавав вівсяну муку і звідки
періодично видаляв відходи. На вівсяному борошні розвивалися дріжджі і
бактерії, якими і харчувалися інфузорії. Три види роду Paramecium успішно
розвивалися в монокультурі в таких умовах. При спільному утриманні P. aurelia
завжди повністю витеснялаP. caudatum. Однак при спільному утриманні P. aurelia
і P. bursaria два види могли існувати разом. Як встановив Гаузе, ці два види
відрізняються за своїм способом живлення: перший вид харчувався бактеріями в
товщі рідини, другий - дріжджовими клітинами у дна.
На підставі описаних
спостережень був сформульований принцип конкурентного виключення, або правило
Гаузе: два види, що займають одну і ту ж екологічну нішу, не можуть стійко
співіснувати в одному місті, співіснування видів можливо завдяки поділу їх ніш.
Правило конкурентного
виключення є однією з можливих причин, що пояснюють надзвичайну різноманітність
видів у природі. Якщо дві популяції в одному місті не можуть займати одну нішу,
їм доводиться її розділяти. З часом кожна з таких популяцій спеціалізується у
своїй ніші і може дати початок новому спеціалізованому виду.
Екологічно перевірити дію
правила Гаузе в природних умовах досить важко, проте деякі приклади його дії
біологам відомі. З кінця XIX століття на більшій частині Європи широкопалий рак
(Astacus astacus) був витіснений довгопалим раком (A. leptodactylus).
Екологічна ніша довгопалого раку перекриває нішу короткопалого, і при цьому
вид-переможець більш плідний. Плотва витісняє в багатьох водоймах червоноперку
і окуня. Екологічні ніші дорослих особин роз'єднані, а мальків -
перекриваються. Мальки плотви виявляються більш конкурентоспроможними. Однак наявність
навіть багатьох прикладів конкурентного витіснення не означає, що воно
відбувається завжди. За той час, протягом якого біологи досліджують дію правила
Гаузе, вдалося знайти як приклади його дії, так і ситуації, коли воно не
«спрацьовує».
Наприклад, в
співтоваристві планктонних водоростей поділу ніш не виявляється. Причиною цього
є середовище, де мінливі умови дають перевагу то одному, то іншому виду, а
також інтенсивний вплив з боку хижаків (зоопланктону і різноманітних
фильтраторів), які не дають конкурентному виключенню стати головним чинником у
формуванні співтовариства. Проте в деяких випадках конкурентне витіснення
відбувається і в планктонному співтоваристві. Це відбувається при так званому
«цвітінні» води, коли планктонні водорості вириваються з-під обмежувальної дії
хижаків. У результаті «цвітіння» води в ній стають масовими всього кілька видів
планктонних водоростей, які відрізняються один від одного за своїм способом
життя.
Деякі види відносин
можуть пов'язувати не тільки особин з різних популяцій, а й родичів, що входять
до складу однієї популяції. Дія внутрішньовидової і міжвидової конкуренції
схожа тим, що і та й інша обмежують кількість ресурсів, доступних для особини.
Проте їх вплив на ширину екологічних ніш протилежний. Внутрішньовидова конкуренція
розширює екологічні ніші: всім особинам не вистачає оптимальних для виду
ресурсів, і деякі витісняються «на край», використовувати менш сприятливі
ресурси. Міжвидова конкуренція, згідно з правилом Гаузе, навпаки, звужує
екологічні ніші. Таким чином, природна ширина ніш є результатом врівноваження
двох протилежних процесів: їх розширення через внутрішньовидову конкуренцію і
звуження внаслідок міжвидової.
4.3 Принцип конкурентного виключення
Цікавий приклад
конкурентного виключення був досліджений в дослідах Т. Парка з борошняним
хрущаком. Два види цих жуків з сімейства чорнотілок (Tribolium confusum і T.
castaneum) розмістили в ящиках з борошном. Ці види конкурували за їжу (борошно)
і, крім того, могли харчуватися один одним, причому жуки обох видів поїдали
переважно особин виду-конкурента, а не свого виду (таке поєднання конкуренції з
хижацтвом називається антагонізмом). Парку вдалося підібрати умови, за яких
закономірно перемагав перший або другий вид. Однак найцікавіше виявилося
досліджувати підсумок конкуренції при проміжних умовах, що не забезпечували
безумовного переваги одного з видів (таблиця 2).
Таблиця 2. Кількість
перемог двох видів борошняних хрущаків в експериментах Т. Парка
|
«Клімат»
(умови експерименту)
|
%
перемог
|
|
Tribolium
confusum
|
Tribolium
castaneum
|
|
Жаркий
вологий
|
0
|
100
|
|
Помірний
вологий
|
14
|
86
|
|
Холодний
вологий
|
71
|
29
|
|
Жаркий
сухий
|
90
|
10
|
|
Помірний
сухий
|
87
|
13
|
|
Холодний
сухий
|
100
|
0
|
Повторюючи дослід при
однакових умовах, експериментатор реєстрував перемогу то одного, то іншого виду
з певною частотою. Зміна умов вирощування змінювала шанси кожного виду на
перемогу, але в досить широкому діапазоні температури і вологості з певною
ймовірністю перемогти могли обидва види. Постійним залишалося тільки одне:
через якийсь час в середовищі залишався лише один вид, а другий зникав.
Відмінності між видами
однієї гільдії (сукупності видів, що використовують один і той же ресурс)
можуть бути опосередковані впливом хижака. Так, У Південно-Східній Азії
сітчасті пітони живуть прямо в поселеннях людини, крадуть і їдять кіз, свиней і
собак і вкрай рідко нападають на людину. Це пов'язано з тим, що людина не схожа
на своїх чотирилапих родичів і не сприймається як жертва.
симбіоз зоохорія антібіоз
екологічний
Висновки
В моїй роботі було
розглянуто типи взаємних відносин біологічних організмів при їхньому
співіснуванні. Таким чином, їх можна поділити на симбіоз - тобто в цілому
взаємовигідні відносини, та антибіоз, де хоча б один з видів зазнає шкоди чи
пригнічення.
В свою чергу ці два типи
також діляться в залежності від того, чи обидва організми отримують користь чи
шкоду, або лише один. Тож можемо бачити, що зв'язки між організмами в
екосистемах є досить складними і різноманітними, але необхідними для її
стійкості.
Також був розглянутий
такий тип відносин, як конкуренція. Вона має велике еволюційне значення, адже
організми, конкуруючи за ресурс, мають вдосконалюватись, щоб мати шанс на
існування. Багато вчених працювали над вивченням взаємного впливу конкуруючих
видів і отримали досить наглядні результати. Так бачимо, що вид може бути більш
чи менш конкурентоспроможним за певних умов, що екологічні інші різних видів не
можуть повністю перекриватись.
Я вважаю цю тему досить
цікавою і важливою. Людина може використовувати знання про залежність життєвих процесів
організмів від інших видів, наприклад, в сільському господарстві, при
полюванні, створенні штучних екосистем.
Людина також пов'язана
великою кількістю взаємних відносин з іншими організмами. Безпосередньо з нашим
організмом поєднані як корисні бактерії, так і шкідливі паразити. І ще безліч
видів ми використовуємо для отримання певної користі. Тому, розглядаючи тему
співвідносин між живими організмами, мусимо пам'ятати, що вона стосується
кожного з нас.
Список використаної літератури
1. Анісімова C.,
Риболова О.В., Поддашкін О.В. Екологія. - К.: Грамота, 2001.- 136с.
. Білявський Г.О.,
Фурдуй P.C., Костіков Î.O. Основи
екологічних знань. - К.: Либідь, 2000. - 334 с.
. Гайнріх Д.,
Герат M. Екологія: dtv - Atlas. Пер. з 4-го нім. вид. - К.: Знання - Прес,
2001. - 287 с.
. Горкин А.П.
Біологія. Сучасна ілюстрована енциклопедія. - 2006, 560с.
. Грин Н., Стаут
У., Тейлор Д. Биология: У 3 т. Т.1.: Пер. с англ. /Под. ред. Р. Сопера. - М.:
Мир, 1990. - 368с.
. Дедю И.Л.
Экологический энциклопедический словарь. - Кишинев: Гл. ред. МСЭ, 1989. - 408
с.
. Дорогунцов СЛ.,
Коценко К.Ф., Аблова O.K. Екологія. - К.: КНЕУ, 2001. -162 с.
. Запольсъкий А.К.
Основи екології: Підручник. - К.: Вища шк., 2001. -358 с.
. Реймерс Н.Ф.
Основные биологические понятия и термины: кн. для учителя. - М.: Просвещение,
1988. - 319с.
. Реймерс Н.Ф.
Экология: теории, законы, правила, принципы и гипотезы. - М.: Россия молодая,
1994. - 367 с.
. Шабанов Д.А.,
Кравченко М.А. Материалы для изучения Ш 12 курса общей экологии с основами средоведения
и экологии человека: Учебное пособие для студентов биологических специальностей
университетов. - X.: ХНУ ім. В.Н. Каразіна, 2009. - 292 с.