Таксономическая группа слизевиков
СОДЕРЖАНИЕ
ВВЕДЕНИЕ
. ИСТОРИЯ ИЗУЧЕНИЯ И
КЛАССИФИКАЦИ
. СТРОЕНИЕ И ХИМИЧЕСКИЙ
СОСТАВ
. БИОЛОГИЯ СЛИЗЕВИКОВ
.1 Трофическая (вегетативная) стадия
.2 Расселительная (генеративная)
стадия
. ОТДЕЛ МИКСОМИЦЕТЫ
(MYXOMYCOTA) ИЛИ НАСТОЯЩИЕ СЛИЗЕВИКИ
.1 Особенности строения
.2 Цикл развития
.2.1 Типы спороношения
.3 Распространение
. ОТДЕЛ ПЛАЗМОДИОФОРОВЫЕ
(PLASMODIOPHOROMYCOTA)
.1 Особенности строения
.2 Цикл развития
.3 Распространение
.4 Плазмодиофоромицеты как
возбудители заболеваний культурных растений
5.4.1 Plasmodiophora brassicae
Woronin
.4.2 Spongospora subterranea
(Wallr.) Lagerheim
. ОТДЕЛ
ДИКТИОСТЕЛИДЫ
(DICTYOSTELIOMYCOTA)
6.1 Особенности строения
.2 Цикл развития
.3 Распространение
.4 Значение
. ОТДЕЛ АКРАЗИЕВЫЕ СЛИЗЕВИКИ
(ACRASIOMYCOTA)
.1 Особенности строения
.2 Цикл развития
.3 Распространение
ЗАКЛЮЧЕНИЕ
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
ВВЕДЕНИЕ
Миксомицеты, или слизевики (лат. Mycetozoa от
греч. μύκητος
- «гриб» и ζωον -
«животное» или Myxomycota от μύξα -
«слизь» и μύκητος) -
группа наземных организмов, обычно относимых к псевдогрибам, насчитывающая
около 1000 видов.
Название слизевики связано с обликом
вегетативного тела, представленного слизистым голым многоядерным протопластом -
плазмодием, размеры которого могут быть микроскопическими либо
макроскопическими, достигающими нескольких десятков сантиметров в диаметре.
Слизевики - одна из наиболее своеобразных и
уникальных по своей природе групп организмов. Целый ряд присущих им свойств
отражает ранние стадии эволюции эукариотов, что делает их изучение не просто
увлекательным хобби, но и серьёзной научной задачей. Они широко распространены
в природе и стали известны науке более двух веков назад. При этом вплоть до
настоящего времени, слизевики оставались загадочной группой, систематическое положение
и родственные связи которой не были полностью ясны.
Цель работы: изучить таксономическую группу
слизевиков, их строение, химический состав, распространение в природе и
значение.
.
ИСТОРИЯ ИЗУЧЕНИЯ И КЛАССИФИКАЦИЯ
Длительное время слизевики рассматривались в
качестве отдельного таксона либо в царстве грибов, либо среди простейших
животных. Таксономическое единство слизевиков при этом не подвергалось
сомнению. Лишь в 70-80-е годы ХХ века была показана полифилетичность этой
группы. Установлено, что слизевики представляют собой самостоятельную жизненную
форму, отличающуюся как от животных, так и от грибов. Как и многие другие
жизненные формы, слизевики объединяют несколько неродственных групп, которые в
процессе эволюции независимо друг от друга приобрели сходные черты организации
[7].
Представители слизевиков были описаны ещё в
середине XVIII века (Battara, 1755; Haller, 1768). Первыми науке стали известны
крупные формы с макроскопическими плодовыми телами. В этот период были описаны
слизистые массы, которые превращаются в плодовые тела со спорами. Эти плодовые
тела внешне очень напоминали грибы-дождевики, поэтому в 1801 году Христиан
Персон (Christian Henrik Persson) в своей монографии «Synopsis methodica
Fungorum» относит слизевиков к гастеромицетам. В 1829 году Фриз изучает
строение плодовых тел огромного количества макроскопических слизевиков и
создаёт лучшее для своего времени их описание. Уже через несколько лет после
публикаций Фриза, Донк (Donk) обнаружил, что сходство слизевиков и дождевиков весьма
условно. Он предложил выделить слизевиков в самостоятельную группу грибов -
Myxomycetes (Слизистые грибы).
В 50-е годы XIX века слизевики привлекли
внимание Антуана де Бари. В лабораторных условиях, проращивая споры из плодовых
тел различных видов слизевиков, де Бари обнаружил, что они дают начало не
мицелию, а амебоидам с активным движением и голозойным типом питания. В 1887
году он убедительно доказал, что между грибами и слизевиками существует лишь
внешнее сходство, и предположил наличие родственных связей с саркодовыми
простейшими. Де Бари отнёс слизевиков к царству животных и предложил для них
новое название Mycetozoa (грибы-животные), тем самым, обратив внимание на их
двойственную природу [3].
С животными их сближают зоотрофное питание,
наличие вегетативной амебоидной стадии, присутствие в циклах развития подвижных
стадий (с двумя апикальными гладкими жгутиками), строение митохондрий. С
грибами - сходство спороношений слизевиков с плодовыми телами грибов. По этой
причине слизевиков долго включали в состав царства грибов, а теперь - в состав
грибоподобных организмов [6, 9].
Окончательная классификация слизевиков еще не
разработана.
Согласно одной из классификаций, таксономическая
группа слизевиков по уровню организации плазмодия, особенностям цикла развития,
способу питания и другим признакам делится на 4 отдела и 6 классов (таблица 1).
Эта классификация будет использоваться в данной работе при характеристике
отделов слизевиков [2, 13, 14].
слизевик вегетативный спора
митохондрия
Таблица 1 - Отделы и классы слизевиков (Л. Н.
Гарибова и др., 2005)
|
Отдел
|
Классы
|
|
Mycomycota
|
Myxomycetes
Protosteliomycetes
|
|
Plasmodiophoromycota
|
Plasmodiophoromycetes
|
|
Dictyosteliomycota
|
Dictyosteliomycetes
|
|
Acrasiomycota
|
Acrasiomycetes
|
.
СТРОЕНИЕ И ХИМИЧЕСКИЙ СОСТАВ
Вегетативные тела слизевиков, в основном,
представлены плазменной массой с большим количеством ядер, не одетой оболочкой.
Такие тела называют плазмодиями.
Плазмодий - сложное
образование. В его составе около 75% воды, а из остальной части около 30%
белков; кроме того, в нем содержится гликоген, или животный крахмал, и
пульсирующие вакуоли. Некоторые слизевики характеризуются наличием большого
количества извести (до 28%) или других включений [10].
У большинства слизевиков в
плазмодии находятся пигменты, придающие им самые различные окраски:
ярко-желтую, розовую, красную, фиолетовую, почти черную. При этом окраска
плазмодия постоянна для данного вида слизевика, но на ее интенсивность очень
влияют реакция среды, освещение, температура, питание и другие факторы окружающей
среды. Предполагают, что некоторые пигменты представляют собой фоторецепторы,
играющие важную роль в развитии слизевиков. Для слизевиков с окрашенными
плазмодиями свет необходим для формирования спороношения, которое образуется
после периода вегетативного роста [7].
Размеры плазмодиев у слизевиков
разных видов могут быть самые различные, от микроскопически малых (например,
Echinostelium minutum, Clastoderma debaryanum) до очень больших. Так у фулиго
(Fuligo) плазмодий может вырасти до нескольких десятков сантиметров в диаметре
(рисунок 1). Вообще плазмодий почти всех слизевиков в течение жизни
увеличивается в размерах, и иногда очень быстро, конечно, при наличия
благоприятных условий. Так, плазмодий слизевика многоголового (Physarum
polycephalum) диаметром 1 см за неделю достигает размера 25 см2.
Плазмодий активно перемещается
в направлении источников пищи, т. е. обладает положительным трофотаксисом. Он
движется в направлении более влажных мест и навстречу току воды (положительные
гидро- и реотаксисы). Пользуясь этой особенностью плазмодия, его можно
«выманить», например, из пня.
Для этого нужно поместить от края пня вглубь его наклонно полоску стекла, а
сверху нее положить фильтровальную бумагу, конец которой погрузить в сосуд с
водой. Ток воды может вызвать вползание плазмодия по стеклу, тогда можно не
только рассмотреть его под микроскопом, но и проследить, с какой скоростью он
перемещается.
Движущие силы токов плазмы в
плазмодии еще сравнительно мало изучены. Есть предположение, что движение
связано с изменением вязкости белка - миксомиозина - при взаимодействии с АТФ.
В прозрачном прикраевом слое
цитоплазмы, свободном от органелл, с помощью электронного микроскопа были
обнаружены чрезвычайно тонкие нити, находящиеся в непосредственном контакте с
оболочкой. Было высказано предположение, что сокращение этих нитей также
связано с токами цитоплазмы и движением плазмодия. Токи цитоплазмы в плазмодии
можно непосредственно наблюдать под микроскопом. При этом в направлении
движения у плазмодия возникают выросты, напоминающие псевдоподии простейших
животных, и общий объем цитоплазмы всегда оказывается большим на переднем по
движению конце плазмодия. Такая полярность плазмодия, по-видимому, тесно
связана с концентрацией калия, т. е. большие концентрации возникают на переднем
конце мигрирующего плазмодия. Измерена скорость движения плазмодия. Она
довольно значительна, достигая 0,1-0,4 мм в минуту [6].
Основную массу видов слизевиков
составляют сапрофитные формы. Плазмодий находится в темноте и питается, главным
образом впитывая всей своей поверхностью органические вещества из окружающей
его влаги. Но, кроме этого, плазмодий может активно захватывать и твердые
пищевые частицы, и живых бактерий, амеб, жгутиковых, мицелий и споры грибов.
Также известно большое количество паразитических форм [1].
3.
БИОЛОГИЯ СЛИЗЕВИКОВ
В жизненном цикле слизевиков выделяют две
стадии: трофическую (вегетативную) и расселительную (генеративную). На
трофической стадии происходит рост организма, накопление биомассы и
вегетативное размножение, преимущественно путем бинарного деления клеток. На
расселительной стадии осуществляется бесполое размножение и распространение с
помощью спор [5].
3.1 Трофическая
(вегетативная) стадия
Вегетативная стадия представлена или одиночными
клетками, или надклеточными структурами.
Одиночные клетки - это первый этап трофической
стадии слизевиков. Условно их делят на 3 типа:
• амебоидные клетки;
• мастигамебы.
Амебоидные клетки - наиболее распространенный
тип трофических особей слизевиков. Они в основном одноядерны, имеют
микроскопические размеры, передвигаются с помощью псевдоподий и осуществляют
фаготрофное питание. Амебоидные клетки слизевиков принято называть миксамебами.
Внешне они часто неотличимы от амебоидных простейших.
Жгутиконосные или флагеллятные клетки
присутствуют в жизненном цикле многих слизевиков. Это видоизмененные миксамебы,
образующие жгутик при повышении влажности субстрата.
Мастигамёбы или амёбофлагелятные клетки образуют
как жгутики, так и псевдоподии. Они являются переходной формой между
миксамебами и жгутиковыми клетками. Как правило, жгутиковые клетки, миксамебы и
мастигамебы, способны к взаимным превращениям.
Надклеточные структуры образуются из одиночных
клеток, и составляют второй этап трофической стадии жизненного цикла
слизевиков.
Известно два основных типа межклеточных
агрегатов:
• плазмодии;
• псевдоплазмодии.
Плазмодий - наиболее характерный для слизевиков
тип вегетативного тела. Он представляет собой гигантскую многоядерную клетку
амебоидной морфологии, и образуется из миксамеб и жгутиковых клеток в ходе их
развития. Пути формирования плазмодия различны. Он образуется либо в результате
множественных делений ядра амебоидной клетки, либо при слиянии отдельных
амебоидных особей.
Псевдоплазмодий - сообщество амебоидных клеток,
не теряющих своей индивидуальности. Он образуется в результате агрегации -
тесного сближения отдельных амебоидных особей с объединением их в единый
организм. Псевдоплазмодий структурно подобен колонии или даже многоклеточному
организму, но принципиально отличается от них тем, что не происходит от одной
материнской клетки, а объединяет особей различного происхождения.
На протяжении трофической стадии, амебоидные и
флагеллятные клетки слизевиков образуют плазмодии и псевдоплазмодии. Последние,
продолжая питаться, достигают иногда значительных размеров, после чего
распадаются на отдельные клетки - споры. Переход к образованию спор знаменует
начало расселительной стадии жизненного цикла [3].
3.2 Расселительная (генеративная) стадия
Процесс образования спор у слизевиков называют
споруляцией. Споры могут образовываться из различных вегетативных структур: из
одиночной амебоидной клетки, из плазмодия, из отдельных сегментов плазмодия, из
псевдоплазмодия
Споруляция слизевиков может происходить двумя
способами: споры либо образуются в виде массы свободных клеток, либо формируют
плодовые тела. Массы спор не образуют морфологически-единой структуры, и
связаны между собой только общностью происхождения. У паразитических организмов
они сохраняют целостность лишь пока находятся в оболочке клетки хозяина.
Плодовые тела слизевиков - это
специализированные структуры, служащие для созревания и распространения спор.
Они обладают структурной цельностью, и являются более прогрессивной формой
споруляции по сравнению с массами несвязанных клеток. Характерны почти для всех
групп слизевиков.
Споры в плодовых телах могут образовываться
двумя способами - эндогенно (внутри плодового тела) и экзогенно (на его
поверхности). При этом плодовые тела могут состоять только из спор, или же
включать стерильные структуры (стебелек, капиллиций и др.), способствующие
распространению спор.
В соответствии с этим различают три основных
типа плодовых тел слизевиков:
• Сорусы - споры образуются экзогенно, плодовое
тело целиком состоит из спор (характерны для плазмодиофорид и акразид);
• Сорокарпы - споры образуются экзогенно,
плодовое тело помимо спор содержит стерильные структуры
• Спорофоры - споры образуются эндогенно,
плодовое тело помимо спор обычно содержит стерильные структуры (характерны для
физаровых).
Размеры плодовых тел у слизевиков колеблются в
значительных пределах - от нескольких микрометров до десятков сантиметров, а
количество спор в них варьирует от одной до сотен тысяч. Строение плодовых тел
весьма разнообразно, и, наряду с морфологией спор, служит важнейшим
диагностическим признаком при определении видовой принадлежности слизевиков [1,
3, 5].
. ОТДЕЛ МИКСОМИЦЕТЫ (MYXOMYCOTA) ИЛИ
НАСТОЯЩИЕ СЛИЗЕВИКИ
Миксомицеты (иногда их также называют физаровыми
слизевиками) - наиболее крупный и хорошо изученный отдел слизевиков, включающий
около 400 видов. Это свободноживущие организмы, обитающие в различных сырых
местах, богатых разлагающимися органическими веществами: в сырой лесной почве,
в компостных кучах, в трухлявых пнях и других подобных местообитаниях [5].
В отделе выделяют два класса - Myxomycetes
(миксомицеты) и Protosteliomycetes (протостелиомицеты). В некоторых источниках
к этому отделу относят также третий класс Ceratiomyxomycetes [7].
4.1 Особенности
строения
Таллом представляет собой плазмодий, способный к
амебообразным движениям по поверхности или внутри субстрата. При половом
размножении плазмодии превращаются в плодовые тела, называемые спорокарпами.
Спорокарпы как правило имеют вид спорангиев.
Плазмодий образуется на диплоидной стадии. Когда субстрат насыщен доступными
органическими веществами, плазмодий достигает значительных размеров. Внутри
плазмодия находятся тысячи ядер. На переднем крае ползущего организма заметны
многочисленные мелкие псевдоподии. Перемещение плазмодия происходит по
градиенту концентрации воды и органических веществ (положительный гидротаксис,
положительный трофотаксис) и в сторону от источника света (отрицательный
фототаксис) [5].
4.2 Цикл развития
Ядра плазмодия диплоидны. В определенный момент
развития плазмодий, достигнув определенных размеров, приобретает положительный
фототаксис, выползает на поверхность субстрата, где его можно обнаружить в виде
бесцветной или окрашенной слизистой массы. Здесь он переходит к генеративной
стадии - образованию спорангиев со спорами.
Обычно в спорангий или в спорангии превращается
весь плазмодий, причем это превращение происходит очень быстро, часто в течение
нескольких часов. Внутри спорангиев образуется масса спор и у многих родов и
видов особые нити - капиллиции с неравномерно утолщенными стенками. Эти нити
образуются из спавшихся при образовании спор вакуолей, находившихся в
плазмодии. Они способны, благодаря утолщениям стенок в виде колец, спиралей,
шипиков и т.д., к гигроскопическим движениям и содействуют, таким образом,
разрыхлению и рассеиванию спор [3].
Образованию спор и спорангиев предшествует
редукционное деление диплоидных ядер плазмодия. Гаплоидные споры, попав на
субстрат, прорастают в гаплоидные зооспоры с двумя гладкими жгутиками на
переднем конце. Зооспоры могут размножаться делением, затем теряют жгутики и
превращаются в гаплоидные миксоамебы, которые также могут размножаться
делением, затем эти миксоамебы попарно сливаются и образуется диплоидная
миксоамеба. У некоторых видов этот цикл укорочен и сливаются попарно зооспоры.
Диплоидная миксоамеба разрастается, число ядер в ней увеличивается за счет
митотических делений диплоидных ядер и таким образом вновь образуется
плазмодий, уходящий вглубь субстрата (рисунок 2) [2].
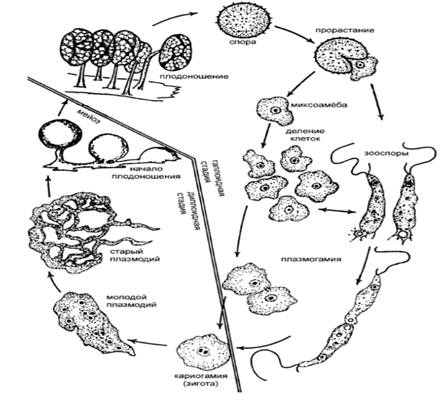
Рисунок 2- Цикл развития миксомицетов (Л. Г.
Переведенцева, 2012)
4.2.1 Типы спороношения
При переходе к расселительной стадии физаровые
слизевики образуют плодовые тела - спорофоры. Они очень разнообразны, могут
иметь различную форму, окраску и размеры. Выделяют четыре основных типа
спорофоров:
• плазмодиокарпы;
• спорокарпы;
• эталии;
• псевдоэталии.
Плазмодиокарпы представляют собой сидячие
плодовые тела неправильной формы. При их образовании плазмодий не претерпевает
морфологических перестроек и лишь покрывается оболочкой (перидием). Встречаются
у низших представителей лициевых, трихиевых и физариформных слизевиков.
Спорокарпы - дифференцированные споровместилища,
состоящие из спороносной головки (спорангия), имеющей округлую, овальную, реже
удлиненную форму. Часто спорангии расположены на ножке (стебельке), имеющей у
различных групп неодинаковое строение. Иногда на одном стебельке располагается
несколько спорангиев (Metatrichia). При образовании спорокарпов крупный
плазмодий обычно дифференцируется на несколько зачатков, каждый из которых
затем образует отдельное плодовое тело. Спорокарп - самый распространённый тип
плодовых тел физаровых слизевиков. У протостелиевых и эхиностелиевых плодовые
тела всегда являются спорокарпами, а у миксогастрид спорокарпы известны у
представителей всех отрядов.
Эталий - группа спорокарпов, слившихся в единое
крупное плодовое тело, покрытое общей оболочкой - кортексом. Обычно эталии не
имеют стебелька. Встречаются нечасто, но известны во всех отрядах миксогастровых
слизевиков.
Псевдоэталий представляет собой группу
спорокарпов, сросшихся боковыми стенками. Внешне псевдоэталий выглядит единым
плодовым телом, но при этом спорокарпы в нем не теряют индивидуальности, и
между отдельными споровместилищами сохраняются перегородки. Псевдоэталии обычно
не имеют общего покрова. Встречаются редко, среди лициевых и физариформных
слизевиков [3, 7, 9].
4.3 Распространение
Миксомицеты довольно обычны в наших лесах, но
плохо заметны на стадии плазмодия. Как правило, обращают на себя внимание их
спороношения, например ярко-розовые одиночные спорангии слизевика ликогала
Lycogala, или группы узких коричневых спорангиев слизевика из рода стемонитис
Stemonitis (рисунок 3).
Леокарпус Leocarpus имеет яркий желтый
плазмодий. Плазмодии леокарпуса можно часто увидеть, когда они выползают на
поверхность почвы. За короткое время плазмодий этого слизевика заползает на
невысокие предметы - мхи, травы, основания стволов деревьев, и образует группы
мелких блестящих коричневых спорангиев на коротких ножках. [3],
[4]
.
ОТДЕЛ ПЛАЗМОДИОФОРОВЫЕ (PLASMODIOPHOROMYCOTA)
Виды имеют внутриклеточный (паразитический)
плазмодий. Отличаются сложным циклом развития, включающим продолжительные по
времени гаплоидную и диплоидную стадии. Включает один класс Плазмодиофоромицеты
- Plasmodiophoromycetes [2].
5.1 Особенности
строения
Вегетативное тело в виде многоядерного
протопласта, не способного к самостоятельному движению и находящегося внутри
клетки растенияхозяина. Специальные спороношения не образуются. Зимующая стадия
представлена спорами, образующимися при распаде внутриклеточного плазмодия. При
наступлении благоприятных условий споры прорастают двужгутиковыми зооспорами.
Жгутики гладкие, прикреплены к переднему концу, один из них длинный, другой - короткий.
Пораженные ткани разрастаются и формируют наросты неправильной формы на
подземных органах растений [7].
5.2 Цикл развития
Жизненный цикл плазмодиофорид характеризуется
наличием двух трофических и двух расселительных стадий. Первые трофическая и расселительная
стадии составляют спорогенную фазу жизненного цикла, а вторые трофическая и
расселительная стадии - цистогенную фазу. В ходе спорогенной фазы происходит
лишь бесполое размножение, а на цистогенной фазе осуществляется половой процесс
в виде изогамии [3].
Оказавшиеся в почве споры прорастают, образуя
двужгутиковые зооспоры, которые теряют жгутики и превращаются в гаплоидные
миксоамёбы. Такие миксамебы живут в почве, затем проникают в корневой волосок,
и разрастаясь, образуют первичный гаплоидный плазмодий. Он не проникает глубоко
в ткани корня и не образует опухолей. Затем он выходит из корневых волосков и
формирует гаплоидные зооспоры (которые можно трактовать как гаметы), попарно
копулирующие и образующие сначала двуядерную, а затем, после слияния ядер,
диплоидную зооспору, которая теряет жгутики и превращается в диплоидную
миксоамебу. Последняя заражает клетки корня и разрастается в нем в диплоидный
вторичный плазмодий. После периода вегетативного роста и мейотического деления
ядер вторичный плазмодий распадается на гаплоидные споры (рисунок 4). Именно
вторичные миксамебы способны вызвать заражение [2,
5].
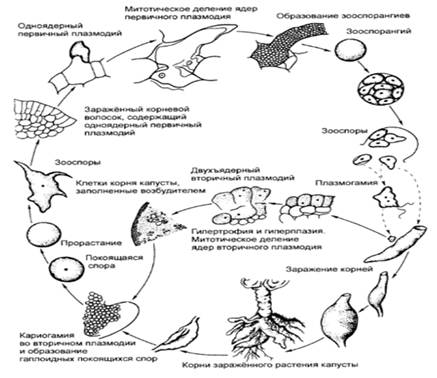
Рисунок 4 - Цикл развития Plasmodiophora
brassicae
(Л. Г. Переведенцева, 2012)
5.3 Распространение
Плазмодиофориды - облигатные внутриклеточные
паразиты зеленых растений (включая зеленые водоросли и сосудистые растения) и
гетероконтных протистов (оомицетов и желтозеленых водорослей). Средой для
распространения плазмодиофорид служит вода. Поэтому они паразитируют либо на
водных организмах, либо, используя почвенную влагу, проникают в подземные, а
через них - и в надземные органы высших растений. Распространение
плазмодиофорид по Земному шару изучено недостаточно. Известно лишь, что многие
виды являются космополитами. [3]
5.4 Плазмодиофоромицеты
как возбудители заболеваний культурных растений
Поражая некоторые культурные растения
(картофель, томаты, капусту) плазмодиофориды наносят значительный вред
сельскому хозяйству. Пораженные ткани, как правило, подвергаются гипертрофии, в
результате чего образуются опухоли и пустулы [3].
5.4.1 Plasmodiophora brassicae Woronin
Этот организм является возбудителем опасного
заболевания крестоцветных, известного под названием кила. Зараженные растения
вырастают слабыми и не образуют кочанов. Их корни покрыты бесформенными
наростами, в которых имеются мертвые клетки капусты с плазмодиями паразита и
активно делящиеся живые клетки, вызывающие разрастания корней (рисунок 5).
Гриб проникает в растения через корневые
волоски. Развитию этого заболевания способствуют температура почвы 18-24°С,
влажность 75-90%, кислотность почвы (рН 5,6-6,5). Часто возбудитель болезни
заносится в поле с рассадой, т.к. поражённые килой растения по внешнему виду
ничем не отличаются от здоровых. В почве споры сохраняют жизнеспособность 4-6
лет и более [8].
5.4.2 Spongospora subterranea (Wallr.) Lagerheim
Этот слизевик является возбудителем порошистой
парши клубней картофеля. Многоядерные плазмодии спонгоспоры развиваются в
периферических клетках клубня. Затем плазмодий распадается на споры, плотно
склеивающиеся в губчатые комочки. Пораженный клубень покрыт язвочками с
коричневым порошистым содержимым, представленным скоплением этих комочков
(рисунок 6). Цикл развития похож на цикл развития Plasmodiophora brassicae.
Иногда поражаются стебли и столоны. Кроме картофеля, поражаются томаты и другие
растения семейства пасленовые [7].
Порошистая парша является широко
распространённым заболеванием клубней, которое может причинять существенный
ущерб, в т. ч. снижение товарных качеств клубней и их лёжкости в период
хранения, поскольку через язвы, вызванные паршой, проникают возбудители
различных гнилей - грибов и бактерий [11].
Оптимальные условия для развития заболевания:
Высокая влажность почвы (70% ППВ), температура 12-18°С, слабокислая реакция
среды. Патоген чаще встречается на тяжёлых и сырых почвах, где сохраняется до 5
лет.
Источники инфекции: заражённые клубни, почва,
навоз [12].
. ОТДЕЛ ДИКТИОСТЕЛИДЫ (DICTYOSTELIOMYCOTA)
6.1 Особенности
строения
6.2 Цикл развития
Цикл развития включает в себя бесполый и половой
процесс (рисунок 7). При переходе к бесполому размножению вегетативные клетки
сползаются в группу, не теряя своей обособленности - образуется
псевдоплазмодий. В пределах псевдоплазмодия клеточных делений уже не
происходит. Псевдоплазмодий двигается, выползая на более открытое место. Здесь
отдельные амебы в пределах псевдоплазмодия начинают заползать друг на друга в
строгой последовательности, образуя «ножку», и «головку» - спорокарп. Спорокарп
напоминает спорангий, однако в отличие от настоящего спорангия, каждая клетка
спорокарпа, как и ножки, представляет собой отдельный организм. Амебы в
середине спорокарпа округляются и одеваются плотной оболочкой, превращаясь в
споры. Спорокарп вскрывается, и споры рассеиваются [5].
В условиях, неблагоприятных для образования
плодовых тел (повышенная влажность, темнота, измененный состав среды и др.),
некоторые виды диктиостелид способны к половому процессу и образованию
диплоидной зиготы. В ходе полового процесса, миксамебы попарно сливаются, в
результате чего образуются двуядерные клетки. Параллельно с этим происходит
сначала набухание, а затем и слияние обоих ядер, в результате чего образуется
диплоидное ядро. Клетка, в которой произошло слияние ядер, называется зиготой
или гигантской клеткой. Показано, что одноядерные амебоидные особи, участвующие
в половом процессе, представляют собой специализированные гаметы, и
соответственно, половой процесс диктиостелид можно считать изогамией.
Сформировавшиеся гигантские клетки начинают привлекать к себе гаплоидных
миксамеб, выступая, фактически, в роли центров агрегации. У миксамёб D.
discoideum агрегация связана с положительным хемотаксисом к цАМФ (рисунок 8). В
результате образуется плотный агрегат, в центре которого расположена гигантская
клетка, а на периферии - гаплоидные амебы, служащие, впоследствии для питания
зиготы. Вокруг агрегата образуется так называемая первичная оболочка. Окружение
многоклеточного агрегата оболочкой знаменует его превращение в молодую
макроцисту. После образования первичной оболочки гигантская клетка приступает к
фагоцитозу окружающих ее амебоидных особей, а после окружается целлюлозной
вторичной оболочкой. Во время формирования вторичной оболочки диплоидное ядро
макроцисты делится мейозом с образованием четырех гаплоидных ядер. После этого
макроциста окружается третичной оболочкой и претерпевает множественные деления
ядер. Зрелая многоядерная макроциста, окруженная трехслойной обоблочкой,
вступает в период покоя.
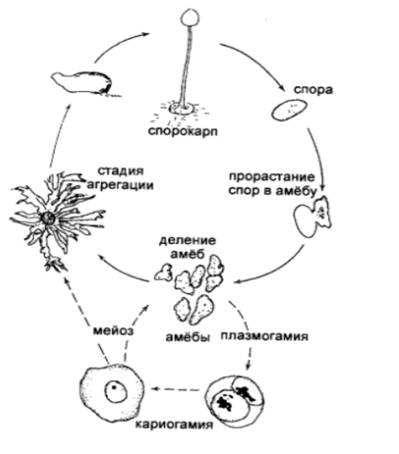
Рисунок 7 - Цикл развития диктиостелид (Л. В. Гарибова
и др., 2005)
В процессе прорастания цитоплазма макроцисты
сперва распадается на крупные одноядерные клетки, называемые проамебами.
Последние многократно делятся, образуя трофические амебоидные особи нормального
размера [2 , 3].
6.3 Распространение
Диктиостелиды - сапротрофы, обитающие в листовой
подстилке, на почве, мертвых частях растений, разлагающихся плодовых телах
грибов или навозе, а также в ризосфере многих растений. Распространены они
повсеместно, но из-за микроскопических размеров, утончённого строения и
эфемерности плодовых тел плохо сохраняются в коллекциях и редко привлекают
внимание специалистов.
Наибольшее число диктиостелиевых слизевиков
обитает в листовой подстилке и почвах листопадных лесов в зоне умеренного
климата. Среди них наиболее часто встречается Dictyostelium mucoroides.
Основным фактором влияющим на встречаемость и распространение диктиостелид
служит наличие пищевых ресурсов в виде почвенных бактерий, составляющих основу
их пищевого рациона. Впрочем, известны виды, способные поглощать не только
бактерий, но и миксамеб других слизевиков, нападая даже на клетки более
крупные, чем они сами, и поглощая их фрагменты. Миксамебы Dictyostelium
сaveatum способны привлекать с помощью аттрактантов клетки других видов и
поедать их. Нередки и случаи каннибализма [3].
6.4 Значение
Диктиостелиды легко культивируются на бактериях
в агаровых средах. Кроме того, ряд видов (например Polysphondilium pallidum и
Dictyostelium discoideum) удается культивировать в питательных средах, лишенных
бактерий. Диктиостелиды используются в качестве модельных объектов при изучении
таксисов, межклеточных взаимодействий, возникновения гетерокариона и других
вопросов клеточной биологии [16].
.ОТДЕЛ АКРАЗИЕВЫЕ СЛИЗЕВИКИ
7.1 Особенности
строения
Трофическая стадия жизненного цикла акразид
представлена амебоидными клетками (миксамебами). Это одиночные клетки овальной
формы, изредка образующие дольчатые псевдоподии, более широкие, чем у
диктиостелид. Передний край миксамебы содержит прозрачную цитоплазму, лишенную
включений - гиалоплазму, а задний конец представлен уроидной зоной - концевой
лопастью, которая содержит выделительные вакуоли. жгутиконосные клетки известны
не у всех видов. Миксамебы способны образовывать неподвижный псевдоплазмодий.
Псевдоплазмодий формирует плодовые тела - сорокарпы, в которых происходит
экзогенное спорообразование. Сорокарпы состоят либо из шаровидных спороносных
головок, либо из ветвящихся цепочек спор; при этом они могут быть стебельчатыми
или сидячими. При созревании, сорокарп полностью распадается на споры. Стенки
спор содержат целлюлозу. Споры прорастают новыми миксамёбами или жгутиконосными
клетками. Жизненный цикл проходит в гаплофазе. Половой процесс не обнаружен
[2].
7.2 Цикл развития
Трофическая стадия: миксамебы передвигаются по
субстрату, питаясь бактериями, простейшими и миксамебами слизевиков, включая
представителей своего вида. Их образ жизни мало отличается от такового у других
свободноживущих слизевиков. В процессе вегетации, трофические клетки акразид
активно делятся путем митоза. При исчерпании запасов пищи, наступлении
неблагоприятных условий среды, или достижении «критической плотности»
трофических клеток, они приступают к образованию псевдоплазмодия путем
агрегации.
В результате агрегации миксамебы образуют псевдоплазмодий.
В отличие от диктиостелид, псевдоплазмодий акразид не способен передвигаться
как единый организм, и поэтому сохраняет неподвижность. Неподвижность
псевдоплазмодиев свидетельствует о том, что у этих слизевиков не выработаны
механизмы согласованного движения клеток, свойственные диктиостелидам.
Псевдоплазмодии акразид не способны к фаготрофному питанию, и являются
кратковременной структурой, переходной между трофической и расселительной
стадиями.
Расселительная стадия: вскоре после своего формирования,
псевдоплазмодий начинает приподниматься над субстратом, приобретая
полусферическую форму. На стадии формирования плодового тела псевдоплазмодий
называют сорогеном. Иногда один псевдоплазмодий образует несколько сорогенов. У
некоторых видов (Copromyxa) полусферический сороген не меняя формы превращается
в примитивное плодовое тело - сорус. У других акразид сороген развивается в
более дифференцированное плодовое тело - сорокарп. При этом миксамебы,
сосредоточенные в верхней части сорогена, образуют сферический спороносный
отдел, а клетки основания сорогена формируют цилиндрическую ножку. У
большинства видов развитие прекращается на этом этапе, и сороген становится
зрелым сорокарпом. Спороносный отдел при этом остается сферическим, и образует
головку, состоящую из большого числа округлых спор. Ножка также сохраняет
первоначальную цилиндрическую форму. У представителей рода Acrasis, развитие
плодового тела не прекращается на этапе сферической головки. Напротив,
спороносный участок сорогена начинает формировать лопасти, направленные
радиально от его центра. Постепенно лопасти становятся все тоньше и длиннее, и
начинают дихотомически ветвиться. Ветвление происходит до тех пор, пока не
образуются ветви толщиной в одну клетку - цепочки спор, напоминающие бусы. В
результате образуется древовидный сорокарп. Степень его разветвленности зависит
от числа клеток, составлявших псевдоплазмодий (рисунок 9) .
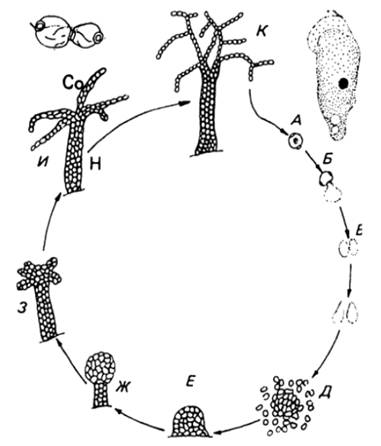
Рисунок 9 - Цикл развития акразиевых на примере Acrasis
rosea: А-Г-прорастание
споры миксамебой, инцистирование ее и новое прорастание, Д- агрегация миксамеб,
Е-И - образование сорокарпа с дифференциацией на ножку (Н) и сороген (Со), З -
разветвление сорогена, И - формирование из миксоамеб цепочек спор, К - зрелый
сорокарп. (Л. В. Гарибова и др., 2005)
Образование плодовых тел у акразид по-видимому
преследует единственную цель - переселение части особей в новые местообитания.
При этом, с одной стороны уменьшается конкуренция между миксамебами в исходном
местообитании, а с другой стороны - происходит их распространение [3].
7.3 Распространение
Акразиды ведут типичный для слизевиков образ
жизни. Они обитают в лесной подстилке, на влажной разлагающейся древесине, а
также на почве и испражнениях животных. Вопрос распространения акразид по
Земному шару недостаточно изучен. Характерный представитель - Acrasis rosea,
образует оранжевые ветвистые сорокарпы (рисунок 10) на разлагающихся
растительных остатках [3].
ЗАКЛЮЧЕНИЕ
Несмотря на немногочисленность
видов и недостаточную изученность, слизевики имеют очень важное значение. В
последние десятилетия интерес к этим организмам возрос благодаря многочисленным
цитологическим, биохимическим, биофизическим и генетическим исследованиям, в
которых слизевики использовались как модельные организмы [13].
Плазмодии слизевиков, чьи
размеры позволяют изучать тончайшие процессы, проистекающие в цитоплазме
клетки, даже при небольшом увеличении - настоящая находка для цитологов, а
сотни синхронно делящихся ядер обеспечивают массовый материал для исследований
генетиков [18].
Изучение миксомицетов имеет
большое значение для эволюционных и филогенетических построений, а также для
систематики грибов и протистов.
Миксомицеты активно поедают
бактерии, играя значительную роль в регуляции численности и состава
бактериальной флоры почв.
Таксономия миксомицетов
базируется главным образом на морфологических признаках плодовых тел
(спорофоров), которые относительно легко гербаризируются и поддаются
количественному учёту. Присутствие вида в природе оценивается по наличию
спорофоров на изучаемой площади или в образцах субстратов. Эти особенности
миксомицетов и уникальная для протистов комбинация в жизненном цикле
микроскопических амёб, плазмодиев и часто хорошо заметных спорофоров, позволяют
использовать их в качестве модельного объекта для экологических и
географических исследований.
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
1. Билай
В.И. Основы общей микологии. Киев: Вища школа, 1980
-
360 с
. Гарибова
Л.В. Основы микологии. Морфология и систематика грибов и грибоподобных
организмов. / Л. В. Гарибова, С. Н. Лекомцева. Учебное пособие. М.:
Товарищество научных изданий КМК,
2005 -220 с
. Глущенко
В. И. Слизевики: учеб. пособие для студентов биологических специальностей. /
В.И. Глущенко, Д.В. Леонтьев, А. Ю. Акулов, Харьков: ХНУ, 2002- 135 с
. Евстигнеев
О. И. Царство грибы. Настоящие грибы, слизевики и лишайники заповедника
«Брянский лес». / О. И. Евстигнеев, Ю. П. Федотов, Е. Ф. Ситникова. Брянск,
2007- 56 с
. Ефимов
П. Г. Альгология и микология: учебное пособие. - Москва: Товарищество научных
изданий КМК , 2011- 120 с
. Жизнь
растений. Грибы Т.2 / Под ред. М. В Горленко. М.: Просвещ., 1991- 478с.
. Переведенцева
Л.Г. - Микология: грибы и грибоподобные организмы. Изд. «Лань», 2012
- 272 c
8. Кила
капусты. Источник: --http://www3.syngenta.com/country/ru/ru/crops/vegetables/cabbagediseases/Pages/plasmodiophorabrassicae.aspx
<http://www3.syngenta.com/country/ru/ru/crops/vegetables/cabbage-diseases/Pages/plasmodiophora-brassicae.aspx>
. Отдел
Myxomycota (mycetozoa). Миксомикота, или настоящие слизевики. Источник:
http://www.activestudy.info/otdel-myxomycota-mycetozoa-miksomikota-ili-nastoyashhie-slizeviki/
© Зооинженерный факультет МСХА
. ОТДЕЛ
СЛИЗЕВИКИ (MYXOMYCOTA)
<http://dic.academic.ru/dic.nsf/enc_biology/1461/ОТДЕЛ>
. Парша
порошистая. Источник: <https://ru.wikipedia.org/wiki/Парша_порошистая>
. Парша
порошистая. Источник :
--http://www3.syngenta.com/country/ru/ru/crops/potato/potatodiseases/Pages/spongosporasubterranea.aspx
<http://www3.syngenta.com/country/ru/ru/crops/potato/potato-diseases/Pages/spongospora-subterranea.aspx>
. <https://ru.wikipedia.org/wiki/Миксомицеты>
. <https://ru.wikipedia.org/wiki/Слизевики>
. <http://educationally.narod.ru/myxomycotaphotoalbum.html>
. <https://ru.wikipedia.org/wiki/Dictyostelium_discoideum>
. <http://en.wikipedia.org/wiki/Acrasis_rosea>
18. http://www.vokrugsveta.ru/vs/article/2701/