Популяционная структура некоторых представителей семейства Coccinellida
Министерство
образования и науки Российской Федерации
Федеральное
государственное бюджетное образовательное учреждение
высшего
профессионального образования
«КУБАНСКИЙ
ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ»
(ФГБОУ
ВПО «КубГУ»)
Кафедра
генетики, микробиологии и биотехнологии
ВЫПУСКНАЯ
КВАЛИФИКАЦИОННАЯ РАБОТА
БАКАЛАВРА
Популяционная
структура некоторых представителей семейства Coccinellida
Работу выполнила
В.В. Пирожкова
Научный
руководитель
д-р биол. наук,
доцент В.В.Тюрин
Краснодар
2014
РЕФЕРАТ
Выпускная квалификационная работа бакалавра
выполнена на 32 страницах машинописного текста. В работу включено восемь
таблиц. Список используемой литературы включает 30 источников.
Ключевые слова: ФЕНЕТИКА, ПОПУЛЯЦИОННАЯ
СТРУКТУРА, ФЕНОТИП, МАТЕМАТИЧЕСКАЯ ОБРАБОТКА, ПОЛИМОРФИЗМ.
Объектом исследования являлись представители
божиих коровок, собранных на территории Краснодарского края станицы Каневской и
прилегающих к ней реках Мылый Челбас, Сухой Челбас, Мигуты.
Целью работы: на основе анализа фенетического
разнообразия в пределах различных видов сем. Сoccinellida выявить полиморфизм в
различных биотопах. coccinellida
Анализ частоты встречаемости разных видов
представителей семейства Сoccinelidea позволил их разделить на категории:
субдоминантов Coccinellidae sp, второстепенных: Сoccinelia distincta,
малозначимый вид: четырнадцатиточечная божья коровка, двадцатидвухтточечная
божья коровка. Зависимости частоты распространения нескольких фенов в разных
популяциях можно сделать заключение о том, что в большинстве случаев выявляется
статистически достоверная сопряженность между частотами фенов и местами
обитания божьих коровок. Показатель внутрипопуляционного разнообразия,
вычисленный для выборок из каждого местообитания, варьировал от 1,19 до 5,98.
coccinellida божья коровка
популяционная
СОДЕРЖАНИЕ
Определение,
обозначения, сокращения
Введение
.
Аналитический обзор
.1
Фенетика как методология изучения структуры популяции
.2
Классификация
.3
Краткая характеристика представителей семейства Coccinellidae
.3.1
Coccinelia septempunctata
.3.2
Coccinelia guinguepunctata
.3.3
Coccinelia distinct
.3.4
Adalia bipunctata
.3.5
Propylaea guatuordecimpunctata
.3.6
Harmonia axiridis
.3.7
Hippodamia tredecimpunctata
.3.8
Exochomus quadripustulatus
.
Географическое описание района исследования
.
Материал и методы исследования
.
Популяционная структура некоторых представителей семейства Сoccinellida
Заключение
Библиографический
список
ОПРЕДЕЛЕНИЯ, ОБОЗНАЧЕНИЯ, СОКРАЩЕНИЯ
ЗГР
- заднегрудь
НДКР
- надкрылья
РОТ
- ротовой, ротовые
ПСП
- переднеспинка
СГР
- среднегрудь
ВВЕДЕНИЕ
Кокцинеллиды (Coleoptera, Coccinellidae)
относится к числу эффективных энтомофагов многих вредителей
сельскохозяйственных культур таких как тля, листоблошек, щитовок, червецов и
клещей, чем приносят громадную пользу сельскому хозяйству и представляют
значительный интерес для программ биологической защиты растений. Божьи коровки
являются одним из самых заметных компонентов огромного количества экосистем.
Своим широким распространением они обязаны рядом морфофизиологических и
экологических особенностей, которые позволяют им адаптироваться к самым
разнообразным условиям окружающей среды. Использование кокцинеллид для
подавления вредителей особенно актуально в наши дни, когда остро стоит вопрос
ограничения применения пестицидов и замены их биологическими средствами.
Мировая фауна содержит более 5000 видов божьих
коровок, выделенных в одноименное семейство в отряде Жесткокрылых [Семьянов,
1999]. Их можно обнаружить на всех континентах, кроме Арктики и Антарктиды.
Подавляющее большинство божьих коровок - ненасытные хищники и лишь немногие
виды растительноядные.
На территории России встречается 152 вида.
Большинство кокцинеллид приурочено к определенным ландшафтно-географическим
зонам, а в них -
к определенным биотопам. Для кокцинеллид характерна привязанность к
определенным стациям в пределах биотопа. Однако некоторые виды обитают во
многих зонах [Савойская, 1974].
Многие, в том числе двуточечная коровка (Adalia
bipunctata), используются в биологической защите сельскохозяйственных и лесных
культур. В Закавказье австралийские виды Rodolia cardinalis и Cryptolae
musmontrouzieri подавили размножение вредителей цитрусовых - червецов
желобчатого (Icerya purchasi) и мучнистого (Pseudococcus gahani), а также
чайной пульвинарии (Chloropul vtnariaaurantii).
Сотрудником ЗИН РАН В.П. Семьяновым разработаны
методики разведения, длительного (до 1 года) хранения и способы применения
инкубированных яиц, личинок и имаго тропической божьей коровки Leisdimidiata
(Fabr.) для борьбы с тлями в теплицах. Создана также оригинальная технология
для быстрого подавления локальных очагов тлей в теплицах (даже при очень
высокой численности вредителя) путем использования личинок Leisdimidiata. Этот
вид кокцинеллид с успехом может быть использован для борьбы с тлями и в
открытом грунте (при температуре воздуха не ниже +20 градусов), а также для
уничтожения тлей на комнатных растениях в жилых помещениях и офисах, где
применение пестицидов нежелательно.
Цель работы: на основе анализа фенетического
разнообразия в пределах различных видов сем. Сoccinellida выявить полиморфизм в
различных биотопах.
Основными задачами данной работы являлись:
Сбор представителей Coccinellidae на территории
ст. Каневской.
Определения видового разнообразия Coccinellidae.
Выявление популяционной структуры Соссinellidae.
1. АНАЛИТИЧЕСКИЙ ОБЗОР
.1 Фенетика как методология изучения структуры
популяции
Фенетика это - распространение генетических
подходов и принципов на формы, собственно генетическое изучение которых
затруднено или невозможно.
Предметом фенетики является внутривидовая
изменчивость, доводимая в конечном итоге до рассмотрения дискретных
альтернативных признаков - фенов. Методами фенетики являются вычисление
изучаемых в изменчивости изучаемых форм различных фенов, количественное и
качественное изучение фенов в популяциях и других группах особей. Целью
фенетики является разработка вопросов микроэволюции, теоритической систематики,
практической биотехнии и других проблем, связанных с популяционным
исследованием видов в природе. [Тимофеев-Ресовский, Яблоков, 1973].
В основе фенетики лежит задача популяционного
исследования видов в природе для разработки проблем микроэволюции. Одним из
центральных понятий в фенетики является «Фен». Фен - это дискретный, обычно
альтернативный, простой признак, отражающий особенности данного генотипа.
Фен является достаточно широким маркером
генотипа: присутствие того или иного фена должно свидетельствовать не о
присутствии какого-то определенного аллеля единственного гена, а о присутствии
одного из аллелей нескольких генов. Проигрывая в точности маркирования, они
выигрывают в широте охвата генов.
На пути реализации наследственной информации от
гена к фену для фенетики существует несколько сложностей, такие как полифения и
псевдофены «онтогенетический шум». Псевдофены это так называемые ложные фены
т.е. такие дискретные признаки фенотипа, которые не могут маркировать данный
генотип.
При анализе получаемых данных феногеографии
обычно встает задача выделения качественных границ в распределении концентраций
того или иного фена. Например, в разных популяциях концентрация фена колеблется
от 0 до 80%.
Начиная с первых этапов развития фенетики, в
наше время выяснение структуры популяции рассматривалось как важное направление
фенетических исследований. Во всех изучаемых случаях популяционное население
оказывалось неоднородным: и у полевок [Турутина, 1979], и у некоторых
китообразных [Яблоков,1979] и у многих других.
Проблема структурированности популяции тесно
связана с проблемой определения популяционных границ. В простейшем случае, при
сравнении двух или более популяционных выборок из заведомо удаленных друг от
друга частей видового населения, можно определить, относятся ли они к одной
общей совокупности или нет. Если они различаются достаточно устойчиво и
статистически достоверно, то вывод о популяционной самостоятельности очевиден.
При этом граница популяции должна проходить где-то между точками взятия наших
популяционных выборок и точнее быть определена, не может.
Иное дело при использовании на сплошь заселенном
видом участке, где множество мелких и мельчайших группировок оказываются
изолированными друг от друга лишь расстоянием. Зная средний радиус
индивидуальной активности [Тимофеев-Ресовский, 1973], можно рассчитать в первом
приближении возможную степень изоляции любой мелкой группе особей от любой
другой такой же группы. Однако такой расчет не имеет особого биологического
смысла, если мы не будем сравнивать реально существующие в природе мелкие
группы. Возможность выделения мелких естественных группировок дает фенетический
подход. После нанесения на карту распределения отдельных редких фенов во всех
исследованных случаях возникала возможность выделение естественных группировок
особей.
.2 Классификация божьей коровки
Царство животные -Animalia
Подцарствобилотеральные - Bilateria
Тип членистоногие - Arthropoda
Подтип - Mandibulata
Надкласс - Panhexapoda
Класс насекомые - Insecta
Отряд жесткокрылые - Polyphaga
Семейство божьи коровки- Coccinellidae
Подсемейство - Coccinellinae
.3 Краткая характеристика встречаемых на
территории ст. Каневской видов божьих коровок
.3.1 Семиточечная божья коровка (Coccinelia
septempunctata)
Эпимеры сгр. белые. Надкр. красные с черными
пятнами. Эпимеры згр. черные. Надкр. вместе с семью точками. Баковая каемка
надкр. впереди сильно утолщена, валекообразная 5,5-8мм. Семиточечная божья
коровка наиболее многочисленный, транспалеарктический вид божьих коровок.
Личинка голубовато-серого цвета с жёлтыми или оранжевыми пятнами. Яйцо крупное,
оранжевое, заострённое на вершине, расширенное к середине и резко суживающееся
к основанию [Полякова, 1969]. Взрослые жуки зимуют в защищённых местах близ
полей, где они питаются и размножаются. Весной только появившиеся жуки начинают
питаться тлёй прежде, чем начать откладывать яйца. В сезон от одного до трёх
месяцев (весной и ранним летом) самка откладывает от 200 до 1000 яиц. Яйца
сравнительно крупных размеров (длиной 0,8-1,2 мм) обычно откладываются близ
добычи, небольшими кучками (от 25 до 85 штук) в защищённых местах на листьях
или веточках (Cornell University Biological Control)
Личинка с одного миллиметра за 10-30 дней
вырастает до 4-8 мм в длину, в зависимости от достатка добычи - тли. Личинка
может подниматься на высоту до 12 метров, как и их жертвы. Личинка четвёртого
(последнего) возраста длиной 8-10 мм. В куколочной стадии развития коровка
проводит до 12 дней, в зависимости от температуры окружающей среды. Развитый
взрослый жук в длину достигает 5-8мм.
В северо-восточных Соединённых Штатах коровка
даёт два поколения за год. Развитие из яйца до имаго может занимать только две
или три недели. Пик численности имаго выпадает на середину и конец лета.
Продолжительность жизни имаго от нескольких недель до двух-трёх месяцев, в
зависимости от места обитания, достатка пищи и времени года.
На территории ст. Каневской было собрано 85 шт.
Место обитания берег реки Сухие Челбасы, Большие Челбасы, Малые Челбасы,
окрестности ст. Каневской, лиманы, искусственные пруды. Растения, на которых
было встречено: яблоня (Macrosiphoneilla oblonga), осот, осока, камыш лесной
(Scirpus sylvaticus), тростник южный (Phragmites australis).
.3.2 Двадцатидвухточечная божья коровка
(Coccinelia guinguepunctata)
Эпимеры загр. темноватые, заднее пятно лежит к
бокового края далеко от шва. Встречается весьма часто на лугах, по побережью
озер, реже в лесу. Количество собранных особей в ст. Каневской 4. Место
обитания берег реки Сухие Челбасы, Большие Челбасы, Малые Челбасы, окрестности
ст. Каневской. Растения, на которых было встречено: яблоня (Macrosiphoneilla
oblonga), осот, осока, камыш лесной (Scirpus sylvaticus), тростник южный
(Phragmites australis), крапива.
1.3.3 Coccinelia distincta
Эпимеры срг. и згр. белые. Длина не менее 6 мм.
Надкр. с общим прищитковым и каждое еще с 3 или 4 пятнами, из которых лежащие
на диске очень крупные. Очень выпуклая боковая каемка надкр., равномерно
тонкая.6-8мм. Встречается повсюду в степи, лесу, на лугах, в лесополосах,
посевах. На территории станицы Каневской было собрано 25 представителей. Место
обитания берег реки Сухие Челбасы, Большие Челбасы, Малые Челбасы. Растения, на
которых было встречено: камыш лесной (Scirpus sylvaticus), тростник южный
(Phragmites australis), лопух (Arctium sp.), крапива.
.3.4 Двух точечная коровка (Adalia bipunctata.)
Активный хищник - афидофаг из отряда
жесткокрылых (Coleoptera), семейства коровок (Coccinellidae). Эпимеры сгр.
черные. Надкр. без складки. Когти с широкими крупными зубцами при основании.
Шов у светлых форм не наблюдается. Надкр. красные с одним черным пятном.
Рисунок на надкр. очень изменчив. Длина тела имаго достигает 5 мм. Ноги и грудь
чёрные, у светлых форм иногда буро-чёрные. Рот. части и усики желтовато-бурые.
Длина тела 3,5-5,5 мм. В некоторых популяциях наблюдается аномальное
соотношение полов, выражающееся в преобладании самок (до 80-90%). Это явление
связано с влиянием симбиотических внутриклеточных бактерий (Wolbachia,
Rickettsia, Spiroplasma), живущих внутри гамет жуков. Яйца желто-оранжевые,
размером 0.4 мм. Самка откладывает их группами по 10-40 штук, в колонии тлей с
нижней стороны листьев. Продолжительность яйцекладки около 30 дней.
Плодовитость самки за жизненный цикл - до 500 яиц. Преимагинальное развитие при
температуре 25˚С длится около 19 дней. Средняя продолжительность жизни
взрослой особи - 56 дней. Соотношение полов близко 1:1
(#"869762.files/image001.gif">
Рисунок 2 - Карта Каневского района [Борисов
2006].
3. МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Работа была выполнена в Краснодарском крае ст.
Каневской. Территории, с которых производились выборки: окрестности ст.
Каневской, берега рек Сухие Челбасы, Большие Челбасы, искусственные пруды,
берег реки Мигуты, берег реки Малый Челбас. Исследование проводилось в период с
июня по август 2013 года.
Объектами исследования для данной работы
послужили сборы за время
летней практики в Краснодарском крае станице
Каневской. Число особей, собранных за этот период составило 336.
Среди них были обнаружены следующие виды:
семиточечная божья коровка, коровка тринадцатиточечная, изменчивая божья
коровка, двух точечная коровка, четырнадцати точечная божья коровка,
двадцатидвухточечная божья коровка, сoccinelia distincta, Exochomus
quadripustulatus.
Сбор насекомых производился с использованием
стандартных методик энтомологических исследований, главным образом это кошение
энтомологическим сачком и ручной сбор на растениях.
Собранных насекомых определяли в соответствии с
таксономическими ключами [Савойская, 1983; Кузнецов, 1992]. Определение
растений проводили по определителю Косенко И.С. (1970).
Рисунок окраски пронотума изучали с помощью
бинокуляра при 16-24 кратном увеличении. Определение велось на основе
морфометрических показателей, таких как окраска, количество точек, расположение
точек, длина голени, кайма псп. , количество члеников и др.
В пределах каждого вида было проведено выделение
фенов - различных вариантов окраски надкрылий. Так, например, у коровки
двухточечной были встречены следующие фены: черная окраска надкр с красными
пятнами (рисунок 1а), красная окраска надкр. с черными пятнами (рисунок 1б),
черная окраска надкр. с желтыми пятнами (рисунок 1в).



а) б) в)
Рисунок 1 - Коровка двухточечная. а) черная
окраска с красными пятнами б) красная окраска с черными пятнами в) черная
окраска с желтыми пятнами.
В популяционно-фенетическом анализе
использовалась дискретное состояние признака (вариации) окраски надкр.
Выделение состояния признака надкр. проводилось по количеству, окраске и
положению пятен, а также окраски самих надкр.
Для оценки фенетического разнообразия
использовались следующие общепринятые показатели. Внутрипопуляционное
разнообразия дискретных состояний анализируемых признаков оценивалось с помощью
показателя µ(1)
µ= (√р1+√р2+…. +√рn) 2,(1)
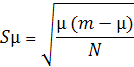 , (2)
, (2)
где µ - среднее число вариаций в выборке, р1,
р2…, рn - выборочное значение частот вариации в долях от единицы (р1+р2…+рn=1),
N - объем выборки, S - статистическая ошибка, m - количество морф. [
Животовский Л.А. 1982]
Статистическая обработка проводилась с помощью
стандартных биометрических методов.
Для статистической формы отклонения признака
нами был применен метод хи-квадрат. Расчеты производились с помощью
статистической программы биостат.
По методу хи-квадрат можно определить
вероятность того, насколько данные отклонения являются случайными или,
наоборот, закономерными. Хи-квадрат дает возможность сравнивать различные
численные отклонения при разных объемах выборки в одном масштабе, что очень
важно для оценки опытных данных. На основе критерия «хи-квадрат» определяли
показатель степени тесноты связи - коэффициент взаимной сопряженности и А.
Чупрова, который позволяет учесть число групп по каждому признаку и
определяется следующим образом:
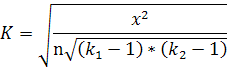 (3)
(3)
где к1 и к2 - число строк и столбцов в таблице,
n - число степеней свободы [Кендалл, 1973]
4. ПОПУЛЯЦИОННАЯ СТРУКТУРА НЕКОТОРЫХ
ПРЕДСТАВИТЕЛЕЙ СЕМЕЙСТВА СOCCINELLIDA
Число выборки составило 336 особей. Были
выделены следующие биотопы:
− Берег реки Сухие Челбасы.
− Берег реки Мигуты.
− Окрестность ст. Каневской (огороды).
− Берег реки Большие Челбасы.
− Окрестность ст. Каневской (улицы).
− Искусственные пруды.
− Берег реки Малый Челбас.
При организации исследования методом выборочного
обследования численность видов (при прочих равных условиях) пропорциональна
количеству собранных проб, т.е. обследованной площади, поэтому для выделения
классов доминирования по численности мы используем шкалу Г.Н. Зайцева [1979]
(таблица 1)
Таблица 1 - Шкала выделения классов
доминирования божьей коровки
|
Балл
|
Границы
классов по численности
|
Название
степени доминирования
|
|
1
|
0<N
<4
|
Малозначимый
вид
|
|
2
|
4<N<16
|
Второстепенный
|
|
3
|
16<N<36
|
Субдоминант
|
|
4
|
36<N<64
|
Доминант
|
|
5
|
64<N<100
|
Абсолютный
доминант
|
Использую данную шкалу мы провели классификацию
видов по частоте их встречаемости в районе исследования (таблица 2).
Таблица 2 - Классификация видов по частоте их
встречаемости
|
Вид
|
%
из общего числа
|
Границы
классов по численности
|
Название
степени доминирования
|
|
Божья
коровка семиточечная
|
27,8
|
16<N<36
|
Субдоминант
|
|
Coccinelia
distincta
|
7,44
|
4<N<16
|
Второстепенный
|
|
Двух
точечная коровка
|
8,92
|
4<N<16
|
Второстепенный
|
|
Коровка
четырнадцати точечная
|
2,68
|
0<N
<4
|
Малозначимый
вид
|
|
Коровка
тринадцатиточечная
|
11,01
|
4<N<16
|
Второстепенный
|
|
Двадцатидвухточечная
божья коровка
|
4,16
|
0<N
<4
|
Малозначимый
вид
|
|
Изменчивая
божья коровка
|
20,83
|
16<N<36
|
Субдоминант
|
|
Соссinellidae
sp.
|
18,15
|
16<N<36
|
Субдоминант
|
К числу видов субдоминантов следует отнести:
изменчивую божью коровку, Coccinellidae sp, семиточечная божья коровка.
К числу видов второстепенных следует отнести:
сoccinelia distincta , двухточечная божья коровка, тринадцатиточечная божья
коровка.
К числу малозначимых видов следует отнести:
четырнадцатиточечная божья коровка, двадцатидвухтточечная божья коровка.
Дальнейшие исследования были посвящены анализу
каждого вида отдельно. В таблице 3 представлены частоты встречаемости особей
Coccinelia septempunctata в разных популяциях.
Таблица 3 −Биотоп божьей коровки
семиточечной (Coccinelia septempunctata), абсолютные и относительные частоты.
|
Фенотип
|
Популяция
|
Всего
|
|
1
|
2
|
3
|
4
|
5
|
6
|
7
|
|
|
Красные
с 3 парами точек по бокам и 1 центральной.
|
9
|
11
|
13
|
36
|
1
|
4
|
17
|
91
|
|
Процент
|
9,9
|
12,1
|
14,3
|
39,6
|
1,1
|
4,4
|
18,7
|
100
|
Для данного вида был встречен только один фен -
красные с 3 парами точек по бокам и 1 центральной. Однако встречаемость особей
в разных популяциях была неодинакова. Так, наибольшей частотой вида
характеризовался биотоп 4 (Коровка четырнадцати точечная) 39,6%. Данная частота
статистически достоверно отличалась от частот в других биотопах, о чем
свидетельствуют результаты, представленные в таблице 4.
Таблица 4 - Сравнение частот особей божьей
коровки семиточечной (Coccinelia septempunctata) в разных биотопах с
использованием критерия Стьюдента
|
Популяция
|
1
|
2
|
3
|
4
|
5
|
6
|
7
|
|
1
|
|
0,47
|
0,91
|
4,92
|
2,9
|
1,47
|
1,72
|
|
2
|
|
|
0,44
|
4,44
|
3,38
|
1,95
|
1,24
|
|
3
|
|
|
|
4
|
3,82
|
2,38
|
0,8
|
|
4
|
|
|
|
|
7,82
|
6,39
|
3,19
|
|
5
|
|
|
|
|
|
1,43
|
4,63
|
|
6
|
|
|
|
|
|
|
3,19
|
Примечание: Курсивом выделены достоверные
значения критерия Стьюдента (p<0,05).
Наблюдаются и другие факты достоверной разницы
численности особей.
Этот результат указывает на неоднородность
распределения особей в пределах изучаемого региона.
В таблице 5 представлены данные по частоте
встречаемости Coccinelia distincta. Данный вид определен нами как
второстепенный.
Таблица 5 - Биотоп Coccinelia distincta
абсолютные и относительные частоты
|
Фенотип
|
Популяция
|
Всего
|
|
1
|
2
|
3
|
4
|
5
|
6
|
7
|
|
|
Надкрылья
с общим прищитковым и каждое еще с 3 пятнами
|
0
|
0
|
1
|
1
|
0
|
0
|
23
|
25
|
Во всех изученных биотопах численность вида
мала. Исключение составляет биотоп 7 (Coccinellieade sp.), где численность вида
высока и статистически достоверно превышает частоты в остальных биотопах.
Особое внимание было уделено той части данных,
где наблюдалось одновременное присутствие нескольких фенов. Сюда можно отнести
данные по численности изменчивой божьей коровки (Harmonia axiridis).
Распределение частот представлено в таблице 6
Таблица 6 - Встречаемость различных морф
изменчивой божьей коровки (Harmonia axiridis)
|
Фенотип
|
Популяция
|
Всего
|
|
1
|
2
|
3
|
4
|
5
|
6
|
7
|
|
|
Без
точечные
|
0
|
5
|
0
|
3
|
0
|
1
|
1
|
10
|
|
Хорошо
развитая одна пара точек.
|
0
|
0
|
0
|
4
|
0
|
0
|
2
|
6
|
|
Две
пары точек
|
2
|
0
|
0
|
3
|
0
|
0
|
8
|
13
|
|
Черная
окраска с 2 парами красных точек
|
0
|
0
|
0
|
4
|
0
|
0
|
4
|
8
|
|
Черная
окраска с 2 парами желтых точек
|
0
|
0
|
0
|
3
|
0
|
0
|
1
|
4
|
|
С
3 и более плохо сформировавшимися пятнами
|
1
|
2
|
0
|
10
|
0
|
0
|
6
|
19
|
|
желтые
|
0
|
0
|
0
|
4
|
2
|
1
|
10
|
Основой полиморфизма природных биотопах, как
сосуществования в них нескольких генетически разных групп, является
естественный отбор, поддерживающий разные генотипические варианты, исходя из
изменяющихся условий среды. Группы, формирующие полиморфизм должны обладать
разной адаптивностью.
Оценка адаптивности была выполнена в анализе
сопряженности численности фенов с условиями местообитаний. Значение критерия
Хи-квадрат, оценивающего сопряженность, оказалось достоверным (х²=51,3;
р=0,00). Поскольку данный критерий служит только для оценки достоверности
связи, её сила измерена по критерию сопряженности (Чупрова). Он оказался равным
0,35, что для популяционных исследований, где собственно генетические различия
скрыты за высокой индивидуальной изменчивостью, является большой величиной.
Иными словами, было показано, что адаптивность разных фенов в разных биотопах
не только различна, но эти различия весьма велики.
Аналогично был выполнен анализ распределения
частот фенов следующих видов:
Таблица 7 − Распределение абсолютных и
относительных частот двух точечной божьей коровки (Adalia bipunctata)
|
Фенотип
|
Популяция
|
Всего
|
|
1
|
2
|
3
|
4
|
|
|
Красная
окраска
|
7
|
2
|
12
|
1
|
22
|
|
Черная
окраска с красными пятнами
|
0
|
1
|
3
|
1
|
5
|
|
Черная
окраска с желтыми пятнами.
|
0
|
1
|
2
|
0
|
3
|
|
Процент
|
23.3
|
13.3
|
56.7
|
6.7
|
100
|
В данном случае критерий Хи-квадрат был равен
5,67 (p>0,05), что указывает на отсутствие сопряженности распределения фенов
в разных популяциях. Действительно, для данного вида обнаружено превалирование
фена «красная окраска» над всем остальными. Сила сопряженности в данном случае
составила 0,28
Таблица 8 − Распределение абсолютных и
относительных частот божьей коровки четырнадцатиточечной (Propylaea
guatuordecimpunctata)
|
Фенотип
|
Популяция
|
Всего
|
|
1
|
2
|
3
|
|
|
Желтая
окраска, точки сливаются друг с другом
|
1
|
6
|
0
|
7
|
|
Желтая
окраска, точки раздельные
|
1
|
0
|
1
|
2
|
|
Процент
|
11.1
|
66.7
|
11.1
|
100
|
Для четырнадцатиточечной божьей коровки связь
между частотами разных фенов и популяциями установлена (Χ2=6,1;
p<0,05). Сила сопряженности равна 0,69, что для популяционных исследований,
где собственно генетические различия скрыты за высокой индивидуальной изменчивостью,
является большой величиной. Иными словами, было показано, что адаптивность
разных фенов в разных биотопах не только различна, но эти различия весьма
велики.
Таблица 9 − Распределение абсолютных и
относительных частот божьей коровки тринадцатиточечной (Hippodamia
tredecimpunctata)
|
Фенотип
|
Популяция
|
Всего
|
|
1
|
2
|
3
|
4
|
5
|
|
|
Желтая
окраска
|
3
|
12
|
6
|
1
|
2
|
23
|
|
Фенотип
|
Популяция
|
Всего
|
|
1
|
2
|
3
|
4
|
5
|
|
|
Оранжевая
окраска
|
0
|
12
|
0
|
0
|
2
|
14
|
|
Процент
|
8.1
|
32.4
|
16.2
|
2.7
|
10.8
|
100
|
Анализ сопряженности встречаемости фенов в разных
биотопах установил статистически достоверную зависимость (Χ2=8,9;
p<0,05). Сила сопряженности равна 0,37
Таблица 10 − Распределение абсолютных и
относительных частот изменчивой божьей коровки (Harmonia axiridis)
|
Фенотип
|
Популяция
|
Всего
|
|
1
|
2
|
3
|
4
|
5
|
6
|
|
|
Без
точечные
|
0
|
5
|
3
|
0
|
1
|
1
|
10
|
|
Хорошо
развитая одна пара точек.
|
0
|
0
|
4
|
0
|
0
|
2
|
6
|
|
Две
пары точек
|
2
|
0
|
3
|
0
|
0
|
8
|
13
|
|
Черная
окраска с 2 парами красных точек
|
0
|
0
|
4
|
0
|
0
|
4
|
8
|
|
Черная
окраска с 2 парами желтых точек
|
0
|
0
|
3
|
0
|
0
|
1
|
4
|
|
С
3 и более плохо сформировавшимися пятнами
|
1
|
2
|
10
|
0
|
0
|
6
|
19
|
|
желтые
|
0
|
0
|
4
|
2
|
1
|
3
|
10
|
|
Процент
|
4.3
|
10
|
44.3
|
2.9
|
2.9
|
35.7
|
100
|
Таблица 11 − Распределение абсолютных и
относительных частот Соссinellidae sp.
|
Фенотип
|
Популяция
|
всего
|
|
1
|
2
|
3
|
4
|
5
|
6
|
|
|
С
3 парами точек
|
2
|
10
|
0
|
15
|
0
|
1
|
28
|
|
С
4 парами точек
|
5
|
0
|
9
|
0
|
0
|
0
|
14
|
|
С
5 парами точек
|
0
|
8
|
0
|
8
|
0
|
0
|
16
|
|
С
6 парами точек
|
0
|
0
|
0
|
0
|
3
|
0
|
3
|
|
Процент
|
11.5
|
30
|
14.8
|
37.7
|
4.9
|
1.6
|
100
|
Для Соссinellidae sp. установлена достоверная
связь между местом сбора материала и выявлеными фенами (Χ2=115,8;
p<0,05). Эту связь следует отнести к категории сильных, исходя из значения
критерия Чупрова (К=0,7).
По результатам исследования зависимости частоты
распространения нескольких фенов в разных биотопах можно сделать заключение о
том, что в большинстве случаев выявляется статистически достоверная
сопряженность между частотами фенов и местами обитания божьих коровок. Сила
связи колеблется от средней (0,32) до высокой (0,7). Можно сделать вывод о
достаточно высоких различиях в адаптации разных генотипов к условиям
существования в разных биотопах.
Была получена количественная оценка
гетерогенности видов божьих коровок в пределах каждого биотопа в виде
коэффициента внутрипопуляционного разнообразия (табл.12).
Таблица 12 - Значение коэффициента
внутрипопуляционного разнообразия (М)
|
Виды
|
Коэффициент
(М)
|
Ошибка
М
|
Число
фенов
|
Число
особей
|
|
Двух
точечная коровка
|
3,46
|
0,062
|
3
|
30
|
|
Коровка
четырнадцати точечная
|
2,19
|
0,1956
|
2
|
9
|
|
Коровка
тринадцатиточечная
|
3,05
|
0,1604
|
2
|
37
|
|
Изменчивая
божья коровка
|
4,52
|
0,0953
|
7
|
70
|
|
Соссinellidae
sp.
|
4,98
|
0,0827
|
4
|
61
|
|
|
|
|
|
|
|
Показатель внутрипопуляционного разнообразия,
вычисленный для выборок из каждого местообитания, варьировал от 1,19 до 5,98.
Если учесть, что его максимальное значение может быть равно числу выделенных
морф, уровень генотипической гетерогенности местообитаний должен быть признан
очень высоким.
ЗАКЛЮЧЕНИЕ
По результатам проделанной работы можно сделать
следующие выводы:
. Анализ частоты встречаемости разных видов
представителей семейства Сoccinelidea позволил их разделить на категории: к
числу видов субдоминантов следует отнести: изменчивую божью коровку,
Coccinellidae sp, семиточечная божья коровка, второстепенных: Сoccinelia
distincta, двухточечная божья коровка, тринадцатиточечная божья коровка,
малозначимый вид: четырнадцатиточечная божья коровка, двадцатидвухтточечная
божья коровка.
. При исследовании видов, представленных одним
феном, в разных биотопах, выявлена их различная частота, что может
свидетельствовать о разной адаптивности в разных экологических условиях.
. По результатам исследования зависимости
частоты распространения нескольких фенов в разных популяциях можно сделать
заключение о том, что в большинстве случаев выявляется статистически
достоверная сопряженность между частотами фенов и местами обитания божьих
коровок. Сила связи колеблется от средней (0,32) до высокой (0,7). Можно
сделать вывод о достаточно высоких различиях в адаптации разных генотипов к
условиям существования в разных популяциях.
4. Показатель внутрипопуляционного разнообразия,
вычисленный для выборок из каждого местообитания, варьировал от 1,19 до 5,98.
Если учесть, что его максимальное значение может быть равно числу выделенных
морф, уровень генотипической гетерогенности местообитаний должен быть признан
очень высоким.
БИБЛИОГРАФИЧЕСКИЙ СПИСОК
Балуева,
Е.Н. Редкие морфотипы Harmonia axyridis Pall. (Coccinellidae) в популяциях
Корейского полуострова // Труды Ставропольского отд. РЭО, №5. Материалы Второй
Международной научно-практической интернет - конференции «Актуальные вопросы
энтомологии». 2009. Ставропольский государственный аграрный университет.
Ставрополь. 2009. C. 40-43.
Богданов
Л.В., Гагальчий Н.Г. Внутривидовая изменчивость азиатской божьей коровки
Harmonia axyridis Pall. в районе Владивостока // Экология. 1986. № 2. С. 56-62.
Борисов
В.И. Реки Кубани. Краснодар, 2005 134 с.
Блехман,
А.В. Изменчивость рисунка протонума у божьей коровки Harmonia axyridis Pallas
(Coleoptera, Coccinnellidae) // Генетика популяции и эволюции. 2007. Т. 5, №2.
С. 25 - 36.
Воронцов
Н. Н., Блехман А. В. Феногеография и геногеография окраски надкрылий в
популяциях восточноазиатской коровки Harmonia axyridis Pall. (Coleoptera,
Coccinellidae) //Энтомология. 1986. Т. 286, № 1. С. 205-208.
Гордеева
И.С., Попов И.Б. К фауне кокцинеллид (Coleoptera, Coccinellidae) Таманского
полуострова // Россия и славянский мир в интеллектуальном контексте времени:
сборник материалов международной научно-практической конференции, посвященной
135-летию начала Русско-турецкой войны 1877 - 1878 гг. Славянск-на-Кубани,
2012. №1. С. 327-330.
Гордеева
И.С. Кокцинеллиды (Coleoptera, Coccinelidae) центральной зоны Краснодарского
края // Биологическая защита растений - основа стабилизации агроэкосистем.
Материалы Международной научно-практической конференции «Современные мировые
тенденции в производстве и применении биологических и экологически малоопасных
средств защиты растений» Краснодар, 2012. №7. С. 90-91.
Гордеева
И.С., Попов И.Б. Некоторые аспекты формирования фауны жуков жужелиц в
центральной зоне Краснодарского края. // Труды Кубанского государственного
аграрного университета. 2012. №2. С. 197-201.
Горностаев
Г.Н. Насекомые СССР / под редакцией док. биол. наук Мазохиной-Поршняковой Г.А.
М. 1970. 372с.
Животовский
Л.А. Показатели сходства популяционной изменчивости по полиморфным признакам.
Фенетика популяций. М., 1982. С 38-48
Зайцев
Г.Н. Построение шкал балльной оценки // Биометрические методы. М., 1974. 123с.
Косенко
И.С. Определитель высших растений Северо-западного Кавказа и Предкавказья. М.,
1970. 753 с.
Кендалл
М.С., Стьюард А.Л. Статестические выводы и связи. М., 1973. 65 с.
Кобзарь
А.И. Прикладная математическая статистика. М., 2006. 816 с.
Кузнецов,
В.Н. Жуки-кокцинеллиды (Coleoptera Coccinellidae) Дальнего Востока России //
Энтомология Владивосток. 1993. №1. С. 34-84. №2. С. 243- 246.
Лотышев
И.П. География Кубани. Энциклопедический словарь. Майкоп, 2006. 230 с.
Лус
Я.Я. О наследовании окраски и рисунка у божьих коровок Adalia оbipunctata L. и
Adalia decempunctata L. М., 1928. 173с.
Лера
П.А. Определитель насекомых Дальнего Востока СССР Т.3. Жесткокрылые, или жуки
Л., 1992. 704 с.
Полякова
Г. М. Определитель жуков трибы Coccinellini (Coleoptera, Coccinellidae)
Среднего Поволжья // Вестник Куйбышевский государственный педагогический
институт имени В.В.Куйбышева. Куйбыш,1969. С 370-372.
Савойская
Г.И. Кокцинеллиды: (систематика, применение в борьбе с вредителями сельского
хозяйства). Алма-Ата, 1983. 248с.
Семьянов
В.П. Кокцинеллиды (Coleoptera, Coccinellidae) опытного хозяйства Всероссийского
научно-исследовательского института биологической защиты растений, как
достижение энтомологии на службе агропромышленного комплекса, лесного хозяйства
и медицины. Краснодар. 2007. №5. С. 194-193.
Семьянов
В.П., Семейство Coccinellidae - божьи коровки или кокцинеллиды (включая
Epilachninae) 1999. Краснодар, 284с.
Тимофеев-Роевский
Н.В., Яблоков А.В. Фены, фенетика и эволюционная биология. // Природа. 1973.
№3. С.40-51.
Тимофеев-Роевский
Н.В., Яблоков А.В.Глотов Н.В. Очерк учения о популяциях. М., 1973. 287с.
Турутина
Л.В Особенности пространственно-генетической внутрипопуляционной структуры
высших позвоночных: Автореф. канд. биол. наук. М.: Ин-т биологии развития им.
Н.К.Кольцова АН СССР, 1979. 85с.
Украинский.А.С.
Азиатская божья коровка Harmonia axyridis Pall. (Coccinellidae) на Северном
Кавказе. М., 632.7, 95с.
Филатова
И. Т. Коровки (Coleoptera, Coccinellidae) Обь-Енисейского междуречья 1993. М.,
58 с.
Яблоков
А.В. Фенетика популяций. М., 1982. 177 с.
Яблоков
А.В. Фенетика: Эволюция, популяция, признак. Москва 1976. 136 с.