Подёнки водоемов Северо-Западного Кавказа
МИНИСТЕРСТВО
ОБРАЗОВАНИЯ И НАУКИ РОССИЙСКОЙ ФЕДЕРАЦИИ
Федеральное
государственное бюджетное образовательное учреждение высшего профессионального
образования
«КУБАНСКИЙ
ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ»
Кафедра
зоологии
ВЫПУСКНАЯ
КВАЛИФИКАЦИОННАЯ РАБОТА БАКАЛАВРА
Подёнки водоёмов северо-западного кавказа
Работу выполнила О. А. Рубанова
Научный руководитель, Г. К.
Плотников
Краснодар
2014
РЕФЕРАТ
Ключевые слова: ПОДЁНКИ (ephemeroptera), ЗАКАЗНИК «КАМЫШАНОВА ПОЛЯНА», фауна, ЧИСЛЕННОСТЬ, ЭКОЛОГИЯ,
СЕВЕРО-ЗАПАДНЫЙ КАВКАЗ.
Цель работы: провести эколого-фаунистическое
исследование подёнок на территории Северо-Западного Кавказа.
В 2013 году было проведено
эколого-фаунистическое исследование отряда Ephemeroptera, по различным водоёмам
Северо-Западного Кавказа. Обнаружено 8 видов, так же определены численность и
распространение подёнок в исследуемых водоёмах.
СОДЕРЖАНИЕ
Введение
. Аналитический обзор
1.1
История изучения отряда Ephemeroptera
.2
Морфологический очерк отряда Ephemeroptera
2. Физико-географическая характеристика исследуемой
зоны
. Материал и методы исследования
. Отряд Ephemeroptera
исследуемых водоёмов
Заключение
Библиографический список
ВВЕДЕНИЕ
Северный Кавказ - самая южная и самая
высокогорная область Российской Федерации. Данный регион, несмотря на
относительную небольшую площадь, характеризуются значительным разнообразием
представленных на его территории ландшафтов и ярко выраженной высотной
поясностью. Наличие мощных ледников позволяет характеризовать Северный Кавказ
как водообеспеченную территорию с относительно высоким коэффициентом густоты
речной сети.
Водотоки являются естественной составляющей
крупных природно-ландшафтных комплексов Северного Кавказа, и в главнейших
чертах они обусловлены региональными, географическими и климатическими
условиями. Зообентос является единственным достаточно развитым и информативным
компонентом биоценозов горных водотоков, на основе данных о составе,
пространственном распределении и количественных показателях развития которого,
возможно проведение гидробиологического мониторинга [Заика, 2009].
Зообентос представляет собой экологическую
группу организмов, характеризующуюся связью с дном водных объектов как
субстратом, на котором (эпибентос) или внутри которого (эндобентос) организмы
проводят свою жизнь. Распределение зообентоса в реках характеризуется
закономерными изменениями видового состава [Байкова, 1970], численности и
биомассы от истоков к устью. Характер этих изменений в реках равнинных и горных
районов не одинаков.
Структура донного сообщества в своей основе
типична для подавляющего большинства горных рек Кавказа. Здесь повсеместно в
речных экосистемах зообентос в большей степени состоит из представителей
четырех отрядов насекомых: Plecoptera,
Trichoptera, Ephemeroptera,
Diptera [Касымов, 1972].
Представители отряда Ephemeroptera
- амфибиотические насекомые, составляющие основную массу гидробионтов в
водотоках, играют важную роль в поддержании устойчивости этих экосистем,
участвуя в процессах на всех уровнях трофической пирамиды. Так же являются
биоиндикаторами водной среды.
В связи с изложенным выше,
целью данной работы стало провести эколого-фаунистическое исследование поденок
на территории Краснодарского края районы Отрадненский и Апшеронский, и
республика Адыгея Майкопский район. Для выполнения цели в дипломной работе
сформулированы следующие задачи: 1) определить видовой состав подёнок; 2)
выявить закономерность распределения подёнок в исследуемых водоёмах; 3)
определить численность особей разных видов.
1. Аналитический обзор
.1 История изучения отряда Ephemeroptera
Кавказский горный хребет, начинается около
Новороссийска невысокими сухими холмами у Черного моря и уходящий на
юго-восток, постепенно повышаясь, расширяясь и отдаляясь от моря.
Соответственно, весь регион естественно делится на Предкавказье - равнины и
предгорья к северу от главного хребта, Большой Кавказ (осевая часть хребта) и
Закавказье - склоны и равнины между хребтом и Черным морем. Чем дальше на
юго-восток и чем выше хребет, тем резче различаются эти зоны.
Кавказ - это один из центров сосредоточения
эндемичной фауны планетарного масштаба. Особое положение данного региона среди
высокогорных районов России по степени эндемизма доказано на примере изучения
таких отрядов как, Ephemeroptera
[Синиченкова, 1979], Plecoptera
[Жильцова, 1964; Черчесова, Жильцова, 2003] Trichoptera
[Мартынов, 1934].
Специальные исследования отдельных районов
Северного Кавказа все более четко подтверждают вышеуказанное. Так, в бентофауне
рек Северной Осетии доля эндемиков ручейников составляет 50 %, веснянок 79 %,
подёнок 62 %. Подобная картина характерна и для рек Кабардино-Балкарии
[Хатухов, Якимов, 2007].
В количественном отношении и по числу видов в
бентофауне горных рек Северного Кавказа доминируют представители класса Insecta
[Касымов, 1972; Цалолихин, 2001] до 96 % от общей численности остальных групп
зообентоса. Такое распределение характерно в основном для рек с ледниковым
питанием.
Представители различных систематических и
экологических групп зообентоса в условиях горных водотоков проявляют достаточно
четкую приуроченность к определенным биотопам и могут служить индикаторами. В
отличие от протяженных широтных физико-географических зон равнин, высотные зоны
горных областей достаточно узки и тесно сомкнуты по линии вертикального
профиля. При таких условиях водные беспозвоночные животные, обладая высотной
пластичности, могут пересекать все ярусы и проникать даже из
равнинно-предгорных зон в высокогорные области и наоборот.
Основными экологическими факторами, влияющими на
формирование биоценозов дна в горных потоках, в том числе и в условия Северного
Кавказа, считаются:
температура воды;
скорость течения;
характер субстрата (гладкие камни предпочитают
подёнки рода Ereorus,
камни со сложным рельефом подёнки родов Rhithrogena,
Ecdyonurus).
Состав и структура зообентоса меняется от
истоков к устью, четко прослеживается в условиях высотной поясности Северного
Кавказа. Так, для многих рек региона характерно ледниковое питание.
Особенностью этих водоемов является наличие «мертвой зоны» [Жильцова, 1989] и
первые представители амфибиотических насекомых появляются на расстоянии 3-5 км
от ледников, где температура воды + 5 °С.
Экологические условия, определяющие модели
зообентосных сообществ в горных потоках с ледниковым питанием рассмотрены в
ряде работ [Панов, 1971], предложил концептуальную модель, отражающую
размещение водных беспозвоночных в водотоках по мере изменения условий и
удаления от ледника. Согласно данной модели, вниз по течению фаунистические
изменения связаны с двумя основными факторами: температурой воды, которая
возрастает с расстоянием вниз по течению от края ледника, и стабильностью
русла.
Также считается основным фактором скорость
течения, формирующим речные сообщества донных беспозвоночных [Жадин, 1940].
Высокие скорости течения, типичные для горных потоков Кавказа, благоприятствуют
высокой аэрации последних, однако это не сопровождается увеличением бентоса. На
участках рек со скоростью течения рек 3-5 м/сек в Северной Осетии, отмечены
представители отряда поденки семейств Heptageniidae
и
Baetidae. В реках со
скоростью течения 1-2 м/сек, а так же в быстротекучих ручьях горного типа
подёнки - род Ecdyonurus.
К морфологическим адаптациям, способствующим
фиксации организмов на субстрате в условиях быстрого течения, относятся
[Бродский, 1976]:
уплощенная, гладкая, обтекаемая форма тела;
небольшой размер тела, что позволяет некоторым
организмам по толщине не превышать толщину пограничного слоя;
положительный реотаксис.
Следует отметить, что степень специализации к
обитанию в условиях быстрого потока различается у разных видов. Наиболее
специализированными обитателями бурных горных водотоков при скорости 3-4 м/сек
являются ползающие поденки родов Epeorus
и Rhithrogena. Как правило,
количество таксонов имеет обратную корреляцию со скоростью течения, и их обилие
выше на участках с малой скоростью течения и седиментации.
Адаптация личинок некоторых видов поденок, к
существованию в горных потоках, проявляется в ряде морфологических
особенностей, в частности в строении жаберного аппарата. Так, интересным
моментом в поведении личинок семейства Heptageniidea
является использование некоторыми из них брюшных жабр для прикрепления к
субстрату. В наиболее развитой форме «прикрепительный жаберный диск» имеется у Rhithrogena
и у многих Epeorus
[Шиолашвили, 2008]. Жаберные листки у этих личинок малоподвижны, расположены
параллельно оси тела и плоскости субстрата, жабры первой пары сильно расширены
вперед и обычно смыкаются под грудью, жабры последней пары имеют продольную
складку, которая позволяет им загибаться под последние сегменты брюшка. Все
жабры, смыкаясь краями, образуют сплошной диск. М. Н. Шиолашвили и С. К.
Черчесовой [2002] в ряде экспериментов было показано, что у исследуемых личинок
рода Rhithrogena,
жаберный диск представляет собой настоящую присоску и развивается отрицательное
давление между личинкой и субстратом. Функционирование жаберного диска в
качестве настоящей присоски, подтверждается значительным развитием жаберных
мышц у личинок с жаберным диском.
Помимо функций присоски, жаберный диск, бесспорно,
выполняет и гидродинамическую роль, заставляя поток воды прижимать личинку к
субстрату, а не сбрасывать её. У личинок Epeorus
жаберный диск, кроме того, увеличивает поверхность трения с субстратом за счет
многочисленных микроскопически крючков. Вероятно, это не относится к Rhithrogena,
имеющих гладкие жабры.
Сложные многообразные адаптации, возникшие у
личинок амфибиотических насекомых (в частности, жаберный аппарат), тесно
связаны с экологическими особенностями среды обитания и экологической эволюцией
последних.
Кроме морфологических адаптаций, возникшие у
личинок Ephemeroptera
указывается наличием миграции взрослой, крылатой фазы развития в верховья и
откладки там яиц. Именно это, по мнению авторов, предохраняет популяцию
гидробионтов от полного вымывания из водотока [Голубков, 1986].
Скорость течения оказывает влияние как
непосредственно на гидробионтов, так и на сам биотоп, в значительной степени
формируя фракционный состав грунта.
Увеличение мутности и накопление осадков на дне
оказывают влияние на все элементы речной экосистемы и приводят к её изменениям
[Горкин, 1983]. Особенно опасно поступление взвесей в горные реки, поскольку
населяющие их сообщества гидробионтов, в частности зообентос, приспособлены к
существованию при очень низкой естественной мутности и чистом не заиленном
грунте. Распределение подёнок в реках характеризуются закономерным изменением
видового состава, численности и биомассы от истоков к устью и от берега к
середине. Характер этих изменений в реках разного типа и их различных участков
неодинаков. В горных реках, где преобладают литореофильные организмы, бентос
поперек русла распределяется довольно равномерно как по видовому составу, так и
по количеству.
Наиболее признанной теорией, объясняющей причины
смены сообществ, вдоль по течению, является концепция речного континуума [Kluge,
2009] которая рассматривает реку как целостную систему, каждый участок которой
обусловлен взаимодействием вышележащих, а вся система представляет собой цепь
непрерывных взаимообусловленных изменений.
Противоположная теория функционирования речной
экосистемы представлена в концепции «динамика пятен», по которой они
располагаются в речной системе хаотично, и структура сообществ каждого участка
реки формируется случайным образом [Zwick,
1971]. На практике, в условиях горных водотоков Кавказа, наблюдается сочетание
континуальных и дискретных свойств речных экосистем, описанных в этих двух
концепциях.
Большой вклад в познание фауны подёнок Кавказа
внесла О. А. Чернова [1964]. Для быстрых горных рек она указывает 6 семейств
подёнок Heptageniidae, Ametropodidae, Baetidae, Leptophlebiidae (к которому
относит единственный кавказский вид), Ephemerellidae, Prosopistomatidae. Она
отмечает, что семейство Heptageniidae остается слабо изученным.
В 1973 г. Н. Д. Синиченкова [1976] описала по
имаго новые виды Rhithrogena decolorata и Rhithrogena stackelbergi. В 1976 г.
[Синиченкова, 1979] впервые описаны личинки рода Iron с Кавказа: Iron
znojkoi и Iron caucasicus, кроме того установлены 2 новых вида по личинкам Iron
fuscus, Iron nigripilosus из бассейна р. Терек. В работе приведены
определительные таблицы личинок. В своей работе [Синиченкова, 1973] первые два
вида описанных ею нимф соответствуют двум видам подёнок в состоянии имаго,
которые в свое время были описаны [Чернова, 1941] под названиями «Ecdyonurus
znojkoi» и «Cinygma caucasicus».
Исследование семейства Heptageniidae на Кавказе
началось с публикации работы [Чернова, 1938], в которой она описала 3 новых
вида рода Ecdyonurus: Ecdyonurus znojkoi, Ecdyonurus frater, Ecdyonurus
ornatipennis и 1 вид рода Iron: Iron
znojkoi, для которого были описаны самец и самка известный по самцам имаго.
В 1993 г. вышла работа Н. Ю. Клюге [1993], в
которой проведена ревизия рода Habroleptoides, впервые описаны 4 новых и
переописаны 7 известных ранее видов. Всех представителей семейства
Habroleptoides он объединил в 7 групп по признакам имаго. В группу
"caucasica" были включены виды: Нabroleptoides
caucasica, Habroleptoides
pauliana, Habroleptoides
filipovicae.
В 1994 г. [Kluge,
1994] опубликована работа, в которой автор рассматривает филогению семейства
Leptophlebiidae и описывает нимф и имаго кавказских видов Habroleptoides
pontica и Нabroleptoides
caucasicа. Новый вид Habroleptoides pontica сильно отличается по строению
гениталий от Нabroleptoides
caucasica, однако по остальным параметрам оба вида очень схожи, что позволило
объединить их в группу "caucasica". Н. Ю. Клюге в работе [1997] дает
описание 3-х новых подродов, среди которых - подрод Caucasiron subgen подрода
Epeoruss (Heptageniidae), который включает некоторые кавказские и
среднеазиатские виды, прежде относящиеся к роду Iron: Eron
caucasicus, Еron znojkoi, Еron
alpestris, Еron
magnus, Еron
soldani, Еron
guttatus.
Вторым по значимости в структуре литореофильных
биоценозов Кавказа является семейство Baetidae, которое долгое время оставалось
слабо изученным. В 1964 г. О. А. Чернова [1964] описала Baetis petrovi из
Армении. В 1972 г. А. Г. Касымов [1972] в составе пресноводной фауны для всего
Кавказа указывает только 2 вида Вaetis rhodani, Baetis
niger. В 1981 г. выходит работа В. Циммерманна [Zimmermann, 1981], в которой
обобщены сведения о распространении подёнок Baetis на Кавказе. В основу работы
положены сборы материала, которые велись на протяжении 11 лет (1967-1978 гг.)
во время 12 отдельных поездок по Кавказу. В. Циммерманн с коллегами обследовал
около 60 пунктов на северных склонах Большого Кавказа, в Грузии и Армении.
Всего же им установлено 11 видов. Впервые указал присутствие еще 3 видов Вaetis
luteri, Вaetis digitatus, Вaetis fuscatus.
Н. Ю. Клюге [1995] по материалам экспедиционных
поездок на Кавказ описал ряд видов многие из которых описаны по имаго и
личинкам, ассоциированным путем выведения: Еcdyonurus ornatus, Rhithrogena
cdyonurus binerva, Rhithrogena eugeniae, Baetis kodori, Baetis stipposus,
Вaetis petrovibotanicus, Baetis digigradlis, Baetis hexabranchis; им также
подтверждена правильность определения следующих видов: Rhithrogena caucasica,
Rhithrogena joostia,
Rhithrogena antumnalis,
Ecdyonurus ornatipennis,
Ecdyonurus sguamatus,
Ecdyonurus expectata.
В 1987 г. Клюге [1987] описал из Армении новый
вид подёнок рода Ephemera - Еphemera romantzovi по имаго самца и самки.
Основные виды которые встречаются в водоёмах Северо-Западного Кавказа
предствленны в таблице 1.
Таблица 1 - Виды подёнок встречающиеся в
водоёмах Северо-Западного Кавказа
|
Семейства,
виды
|
Наличие,
отсутствие вида
|
|
Baetidae
|
|
|
Baetis
muticus
|
*
|
|
Baetis
rhodani
|
*
|
|
Baetis
vardarensis
|
*
|
|
Baetis
baksan
|
*
|
|
Caenidae Caenis
luctuosa
|
*
|
|
Ephemeridae
Ephemera
danica
|
*
|
|
Oligoneuriidae
Oligoneuriella
rhenana
|
*
|
|
Prosopistomadae
Prosopistoma orhanelicum
|
*
|
|
Heptageniidae Ecdyonurus venosus
Electogenia sp. Epeorus alpestris Epeorus sp. Rhithrogena caucasica
Rhithrogena decolorata Serratella ignitta Heptagenia samochau
|
*
* * * * * * *
|
|
Leptophlebiidae Paraleptophlebia
cincta Habroleptoides caucasica Choroterpes sp.
|
* * *
|
|
Примечание
- * Виды встречающиеся в водоёмах
|
Таксономия подёнок Кавказа разработана
недостаточно - это проявляется в резко противоречивых взглядах на видовую
самостоятельность описанных форм, поэтому целесообразно анализировать материалы
по всему списку существующих на сегодняшний день таксономических описаний
кавказских поденок.
Представители подёнок, в условия горных
водотоков проявляют достаточно четкую приуроченность к определенным биотопам и
могут служить индикаторами.
Таксономия поденок Кавказа разработана
недостаточно - это проявляется в резко противоречивых взглядах на видовую
самостоятельность описанных форм, поэтому целесообразно анализировать материалы
по всему списку существующих на сегодняшний день таксономических описаний
кавказских подёнок.
.2 Морфологический очерк отряда Ephemeroptera
подёнка видовой водоём фаунистический
Среди обитателей водных экосистем бентосные
беспозвоночные и, в первую очередь, личинки амфибионтных насекомых,
составляющие основную массу гидробионтов в водотоках, играют важную роль в
поддержании устойчивости этих экосистем, участвуя в процессах на всех уровнях
трофической пирамиды. Личинки всех видов подёнок развиваются в пресных водоемах
и дышат растворенным в воде кислородом, крылатые стадии развития (имаго и
субимаго) обитают в воздушной среде.
Строение личинок: пара фасеточных глаз и три
простых глазка всегда достаточно развиты. Антенны щетинковидные, с
неопределенным числом члеников. Ротовой аппарат либо грызущий [Байкова, 1972],
либо в той или иной степени модифицирован.
Зачатки крыльев направлены вершинами назад,
верхней стороной дорсально и костальным краем латерально (в отличие от личинок
стрекоз и прямокрылых). Зачатки задних крыльев не менее чем вдвое меньше
зачатков передних крыльев, либо отсутствуют; могут быть видны сверху только у
молодых личинок, а у личинок последнего возраста всегда скрыты под зачатками
передних крыльев. Лапка подвижно сочленена с голенью, не расчленена; иногда
видны следы слияния 5 члеников [Клюге, 1983]. Каждая лапка имеет лишь один
коготок (который у личинок рода Behningia редуцирован). На рисунке 1 показаны
личинки подёнок [Новикова, 1987].
Брюшко состоит из десяти явных сегментов. На
1-7-м сегментах брюшка имеется по паре подвижно причлененных придатков - тергалий
(трахейные жабры, или жаберные листки). Тергалии обычно причленяются на
дорсальной стороне по бокам заднего края тергита (лишь у Behningia и на первом
сегменте брюшка у Oligoneuriinae их основания смещены на вентральную сторону, а
на задних сегментах Ephemeridae и Caenoidea - в переднюю половину тергита)
[Мамаев, Медведев, Правдин, 1976].
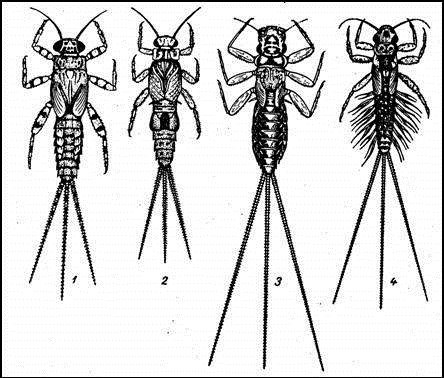
Рисунок 1 - Личинки подёнок 1 - личинка Heptagenia
fuscogrisea; 2 - личинка Heptagenia
sulphurea; 3 - личинка Heptagenia
flava; 4 - личинка Ephemerella
ignita
Сзади от десятого сегмента брюшка отходят три
длинные хвостовые нити, состоящие из неопределенного числа мелких члеников:
пара церков и непарный парацерк; церки развиты у всех видов подёнок, а парацерк
развит у личинок большинства подёнок (в том числе в тех таксонах, где у имаго
парацерк редуцирован), но у личинок некоторых специализированных реофильных
видов и родов парацерк укорочен или рудиментарный (одночлениковый).
Хвостовые нити могут быть опушены первичными
[Клюге, 1997] плавательными волосками: это расположенные в одной горизонтальной
плоскости волоски, образующие по одному продольному ряду на внутренней стороне
каждого церка и пару продольных рядов на боковых краях парацерка. Иногда
бывают, развиты также вторичные плавательные волоски (на наружных сторонах
церков), либо мутовки волосков на вершине каждого членика, либо иное опушение.
Личинки самцов обычно (кроме Caenidae и многих
Baetidae) отличаются от личинок самок формой заднего края 9-го стернита брюшка
(несущего зачатки гоностилей); обычно также (кроме Caenidae) более крупными
глазами; у личинок самцов Baetidae и некоторых других медиальные глаза, а так
же имеется пара зачатков тюрбанных глаз у имаго [Клюге, 1987]. У семейства
Caenidae личинки самцов отличаются от личинок самок вздутыми основаниями
хвостовых нитей.
Полет осуществляется четырьмя перепончатыми
густо-сетчатыми, крыльями. Пара задних крыльев гораздо меньше передних, а
иногда (у Cloeon diptenim) вовсе отсутствует. Кишечник подёнки заполнен
воздухом и играет роль аэростатического приспособления. У подёнки, вследствие
ненужности для неё во взрослом состоянии пищи, пищеварительный канал перестал
выделять пищеварительные соки и взял на себя другую функцию - облегчать этому
насекомому полет в воздухе, то есть в функциональном отношении стал органом
[Хаберман, 1952] обслуживающим движение, хотя по внешности сохранил все черты
типичного анатомического строения.
По типу питания подёнки детритофаги, у личинки
хорошо развит грызущий ротовой аппарат. Личинка активно питается в основном
растительными остатками, в свою очередь служит пищей для веснянок, а так же
велико значение поденок в пище рыб, особенно горных и предгорных рек. По данным
[Чернова, 1958], в рационе таких рыб, как ленок, хариус, осетр, подкаменщик, личинки
поденок в летний период составляют от 81 до 100 % от общего числа потребленных
животных.
Во взрослом состоянии у подёнок наблюдается
недоразвитие ротовых частей, служащих для принятия и пережевывания пищи, это
объясняется тем, что подёнка во взрослом состоянии ничего не ест и не может
есть. Не только недоразвит её ротовой аппарат, но и внутренние пищеварительные
органы, её желудочно-кишечный канал не работает по своей специальности, а
является воздушным пузырем, облегчающим насекомому парение в воздухе. Кишечник
подёнки заполнен воздухом и играет роль аэростатического приспособления
облегчая насекомому полет в воздухе. Воздушные танцы подёнок являются их
брачным полётом - в это время они спариваются. На летающую самку набрасываются
несколько самцов, и одному из них удается схватить её; самец при этом
помещается под самкой. Он держит её за переднегрудь своими длинными передними
ногами, а генитальными щипцами за кончик брюшка [Клюге, 1985]; хвостовые нити
самца находятся в это время между крыльями самки. Во время спаривания летит
только самка, самец пассивно висит на ней; самке трудно нести самца и пара во
время своего полёта несколько снижается. Копуляция обычно продолжается всего
несколько секунд; у некоторых видов дольше 6-7 минут. Самки на лету опускаются
на поверхность воды, куда и сбрасывают выступающие из заднего конца брюшка
комочки яиц. Некоторые виды подёнок (Baetis) для той же цели ныряют под воду и
откладывают яйца на нижнюю поверхность камня [Синиченкова, 1976]. При этой
операции они дышат под водой воздухом, который уносят с собой при погружении.
Развитие происходит в воде, взрослая особь
откладывает яйца в воду. Личинка 1-го возраста не имеет тергалий, ее жгутики
антенн, церки и парацерк состоят из небольшого числа члеников. Со второго или
более позднего возраста появляются тергалии; при этом форма тергалий, их
относительные размеры и количество могут существенно различаться у личинок
разных [Макаров, 2000] возрастов, число личиночных линек большое до 23, иногда
неопределенное. На рисунке 2 показан цикл развития подёнок [Кустрева, 1984].
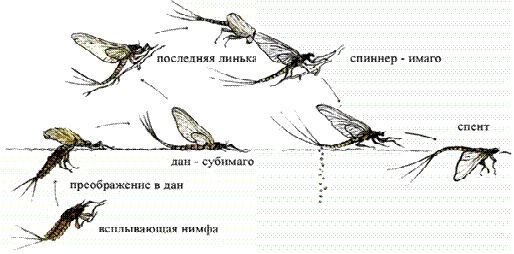
Рисунок 2 - Цикл развития подёнок
Развитие личинки продолжается обычно один или
несколько месяцев, у некоторых видов - до трех лет. За этот период личинка
линяет до 23-х раз. Личинка последнего возраста превращается в крылатое
насекомое, перелиняв на поверхности воды, либо под водой, либо на выступающем
из воды предмете.
В отличие от всех прочих насекомых, имеющих
только одну летающую стадию (имаго), поденки имеют две летающие стадии
развития, разделенные линькой - субимаго (неполовозрелые) и имаго
(половозрелые). По размерам и форме крыльев субимаго не отличается от имаго.
Крылья субимаго покрыты микротрихиями (отчего
выглядят замутненными), у имаго крылья голые. У субимаго и имаго [Кустарева,
1984] поденок антенны короткие, щетинковидные; ротовой аппарат не развит;
крылья в покое не складываются, подняты вверх или распростерты в стороны;
задние крылья не более 1/2 длины передних или отсутствуют; лапки расчленены, с
двумя коготками; сзади от 10 сегмента брюшка отходит пара многочлениковых
церков (обычно церки длиннее тела, лишь у субимаго и самок Caenidae и некоторых
других короче тела), иногда бывает такой же парацерк.
2.
Физико-географическая характеристика исследуемой зоны
Под Северо-Западным Кавказом понимается
территория, ограниченная с севера Кумо-Маныченской впадиной, с запада
побережьем Азовского и Черного морей, с востока бассейнами рек Егорлык, Уруп и
Большая Лаба, с юга - бассейном р. Псоу. Площадь исследуемого региона
составляет примерно 87 000 км, большая его часть приходится на Краснодарский
край [Канонников, 1984].
Лагонакское нагорье это часть Западного Кавказа
между реками Белой и Пшехи. Это Кубанский вариант поясности. В районе
Лагонакского района сохранились следы древних ледников. От общей площади
оледенения Большого Кавказа площадь ледников района составляют 0,06 %. На
территории Лагонакского нагорья выделяют четыре ледника, последний был открыт в
1983 году. Два располагается на Фиште, и два на Пшеха-Су. Общая площадь
Ледников составляет 1 км2, это 0,15 % площади Лагонакского нагорья,
самый крупный находится на северном склоне горы Фишт.
Климат района умеренно-континентальный с
повышенной влажностью. Среднегодовая температура + 10,5 °С. Средняя температура
января минус 14 °С, июля + 25 °С. Число дней со снежным покровом 40-50,
безморозный период составляет 185-195 дней.
Ландшафты представляют собой оригинальное
сочетание лесных, опушечных и полярных сообществ, что позволяет организовывать
места отдыха рекреантов и временные стоянки отдыхающих, места богаты родниками,
чистыми ручьями.
Густота речной сети Лагонакского нагорья
определяется закарстованностью района. Для рек характерны значительные уклоны,
наличие водопадов до 40-60 м и более. Реки Лагонакского нагорья принадлежат к
бассейну второго по длине и самого мощного по водоносности притока Кубани реки
Белой.
Река Белая зарождается на склонах горного
массива Фишт-Оштен и, пробежав 265 км, впадает в Краснодарское водохранилище.
Все притоки Белой являются типичными горными реками с быстрым течением,
многочисленными порогами и водопадами. Белая на своем пути пересекает ряд
различных ландшафтов, поэтому характер реки меняется от верховий к устью
[Борисов, 1976].
Питание р. Белой идет за счет атмосферных
осадков в виде дождя и снега, подземных вод, а также высокогорных снегов и
ледников. В бассейне реки насчитывается 29 ледников. Их общая площадь
составляет 7,6 км.
Река Белая многоводна, минерализация воды
колеблется от 330 мг/л до 24 000 мг/л, а суммарный дебит составляет около 30
тысяч кубических метров в сутки. Вода этой реки мягкая и обладает хорошими
питьевыми и техническими качествами. В ней преобладающими по весу ионами
являются гидрокарбонатные, кальция и сульфатный. По классификации О. А.
Алекина, воды р. Белой и ее притоков во все сезоны года относятся к
гидрокарбонатнокальциевым второго типа.
Река Курджипс начинается на Лагонакском нагорье
с восточного склона хребта Абадзеш 2376 метров над уровнем моря, и из родников
на северных отрогах горы Оштен. Длина реки составляет 100 км. Имеет 82 притока,
самые большие это: Мезмай и Морозка.
Границы заказника совпадают с внешними границами
образующих его лесных кварталов, поэтому на значительном протяжении они
условны, а на севере и на юге территории, к тому же субширотны и идеально
прямолинейны. Заказник располагается на пологом северо-западном склоне хребта
Азиш-Тау и, малой частью, - на правом, крутом берегу глубоковрезанной долины
Курджипса (Лагонакское нагорье, Западный Кавказ). Территория заказника в целом
расположена в среднем горном поясе на высотах 800-1450 м над ур. м. и
подразделяется на две основные морфодинамические ландшафтные структуры:
вершинные поверхности наклонных плато на моноклинальных структурах,
бронированных карбонатными породами, ландшафты глубоко врезанных крутосклонных
долин, буковых с кленами, разнотравных, папоротниковых, выработанных в
известняково-доломитовой и песчано-глинистой толщах. В настоящее время имеют
место антропогенные ландшафты.
Речные долины охватывают хребет Азиш-Тау с юга, запада
и севера, Река Мезмай и не большие притоки Курджипса разрезают хребет. Питание
рек осуществляется за счет карстовых источников. Для рек характерны
значительные уклоны. Реки имеют многочисленные водопады. Самый крупный водопад
это - Университетский, расположен в приустьевой части реки Мезмай - Горелой
балке. Высота водопада составляет 60 метров, имеются несколько уступов.
Вода в реках слабо минерализована. В половодье
она содержит до 200-300 мг/л растворимых солей. Вода мягкая и обладает хорошими
питьевыми качествами.
Отрадненский район находится в юго-восточной
части Краснодарского края на северном склоне Главного Кавказского Хребта.
Климат характеризуется небольшими годовыми перепадами температур. Лето обычно
не жаркое, средняя температура + 19 оС, с дождями, зима теплая
средняя температура минус 10 оС. Среднегодовое количество осадков
560-770 мм. Рельеф района представляет собой предгорные равнины, высота над
уровнем моря составляет от 300 до 1500 метров в лесной горной части.
Район имеет густую речную сеть. Главной рекой
района, является Уруп, с притоками Аргош, Малый и Большой Тегинь. Уруп в своем
верхнем и среднем течении горная река. Долина реки в этом районе узка и
глубока, склоны ее круты и местами она имеет вид отвесных неприступных каньонов,
в которых стремительно несется водный поток.
В местах прорыва через хребты долина реки имеет
вид ущелья. Водный режим р. Урупа неустойчив. Уровень воды в реке и расходы ее
значительно колеблются в течение года. Половодье наступает летом, когда тают
снега высокогорных вершин Передового хребта. Наивысшие подъемы воды в реке
наблюдаются в июне - июле. В конце лета, осенью и зимой Уруп сильно мелеет.
Питается река за счет атмосферных осадков и грунтовых вод. Причем в долине р.
Урупа выбивается ряд минеральных источников.
В своих верховьях р. Уруп имеет воду малой и
средней минерализации 100-200 мг/л. Вниз по течению она повышается и происходит
обогащение вод сульфатами, потому что в бассейне р. Урупа имеются залежи гипса.
Так, у города Армавира минерализация может достигать более 1000 мг/л, жесткость
воды также повышается, а ее питьевые и технические качества ухудшаются. В воде
преобладают по весу ионы гидрокарбонатный, сульфатный и кальция.
3. Материал и методы
исследования
Объектами исследования выбраны бентосные
амфибионтные насекомые из отряда подёнки. Амфибионтные насекомые это те виды, у
которых пред имагинальные фазы (яйцевая и личиночная) развиваются в водной
среде, а имаго (взрослая фаза) обитает на суше.
Сборы проводили в летний период 2013 года. В водоёмах
заказника «Камышанова Поляна» (ручей на Шанхае, родник Камышановой поляны) а
так же в реках Курджипс, Мезмай, правый приток реки Горелая Балка, были взяты
пробы с Малого и Университетского водопадов, обследованы стоячие водоемы -
лесные озера и мочаки, расположенные в различных местах заказника. Было
собранно более 400 личинок подёнок различных видов.
А так же сборы проводили на территории
Отрадненского района. Были исследованы такие водоемы как река Уруп, рука Аргош,
река Малый Тегинь, ручьи и Озеро. На рисунке 3 изображены точки сбора
материала.
Рисунок 3 - Места сбора материала (Отрадненский
район)
В ходе исследований было собрано 171 особь из
отряда подёнки. Изучение видового состава, распределения и количественного
развития донных организмов проводится различными методами и разными орудиями
лова. Для рек горного и предгорного типа с каменистым грунтом количественные
пробы бентоса могут быть отобраны:
а) с помощью бентометра;
б) путем сбора макробентоса с отдельных валунов.
Сбор и обработка проб зообентоса проводили по
общепринятым методикам.
Для количественного сбора бентоса использовали
бентометр конструкции Садовского, с помощью которого удобно работать в условиях
мелководных быстротекущих рек. Пробы, собранные дночерпателем, перед фиксацией,
отмучивали порциями в ведре или тазу. Для этого часть пробы помещали в ведро
или таз, наполняя его на 1/3 водой и вращательными движениями взмучивали.
Взмученную фракцию сливали в сачок-промывалку. Процесс отмучивания повторяли до
тех пор, пока вода, которой промывали пробу, не становится прозрачной.
Количественные пробы бентоса отбирали бентометром конструкции Садовского с
площадью покрытия дна 0,07 м², высота
50 см. Бентометр удобно использовать в условиях мелководных быстротекущих рек.
На каждой станции единовременно отбирали 2-3 проб.
Наиболее широко в практике гидробиологических
исследований используется бентометр В. Я. Леванидова [1976]. Прибор
представляет собой прямоугольный каркас, имеющий площадь захвата 0,3x0,42 м и
высоту 0,4 м. Три боковые грани каркаса затянуты планктонным газом N 15, а к
четвертой грани пришит мешок из планктонного газа N 23 длиной 1,5 м. Бентометр
плотно устанавливают на грунт до глубины не более 0,3 м. Оказавшиеся внутри
бентометра камни вынимают в ведро или таз, которые затем тщательно отмывают, а
имеющийся между камнями наполнитель взмучивают. Собранную в мешке фракцию
переносят в ведро с водой. Листья, палки и камни, попавшие в мешок, отмывают, а
полученный осадок промывают через сачок-промывалку. Сетки из планктонного газа
обеспечивают лучшую видимость дна при сборе материала, чем сделанные из
листового железа или стали.
Отбор качественных проб осуществляли с помощью
таких гидробиологических орудий как сачки, скребок, выбор которых зависит от
размера водотока, его глубины и скорости течения. Для более полного учета
видового состава водотоков применяли ручной сбор макробентоса с отдельных
валунов, личиночных шкурок.
Для установления качественного состава
макрозообентоса, а также в связи с большим количеством валунов в русле реки,
проводили смывы с камней. Грунт помещали в ведра и фиксировали 40 % формалином.
В лабораторных условиях организмы бентоса отделялись от грунта.
Затем личинки подёнок фиксировали 4 % раствором
формалина, проводили этикитирование.
Этикетку изготавливали из плотной бумаги или
матовой кальки. Надписи делали острым карандашом. На этикетке отражали
следующую информацию: название животного; место сбора (так, чтобы можно было
найти на карте); где именно был встречен объект (камень, коряга, ил,
поверхность воды); дата; имя и фамилия собравшего.
Этикетку помещали в банку вместе с материалом
или приклеивали скотчем снаружи.
Определение гидробионтов осуществляли с
использованием бинокуляра МБС-10 и микроскопа ЛОМО МИКМЕД-6.
Определение гидробионтов произведено
определителями [Хейсин, 1962; Определитель пресноводных …, 1997; Кантор, 2005;
Чертопруд, 2010].
Математическую обработку данных проводили с
использованием программных средств MS Excel 2000, Statistica 6.0.
4. Отряд Ephemeroptera
в исследуемых водоёмах
В процессе проведения исследований на территории
заказника нами были обследованы различные типы водоемов, которые располагались
в разных условиях (на полянах, в лесу), на различных высотах и экспозициях
горных склонов, с различающимися условиями - проточность, температура воды, тип
дна.
Первая стация - Большой водопад. Большой
университетский водопад расположен на ручье Горелая Балка, длина водопада более
20 м. Малооблачно, температура воздуха + 18 °С, температура воды + 5 °С.
Собрано 20 особей на 5 м2. Комментарии: дно каменистое, в месте
сбора глубина составила 0,45 м. В результате было обнаружено 35 представителей
данного отряда.
Вторая стация - родник на территории биостанции.
Родник на биостанции «Камышанова Поляна». Безоблачно, температура воздуха +18 °C,
температура воды + 4 °С. Комментарии: поймано 50 особей с 5 м2.
Водоём освещён частично, вода проточная, дно каменистое, встречаются крупные
камни, вода прозрачная, глубина в месте сбора 15 см. Были найдены представители
данного отряда, в количестве 31 особь.
Третья стация - Малый водопад. Расположен на р.
Мезмайка. Координаты: 44°10'813"N
и 40°03'28"Е. Высота 1100 метров. Безоблачно, температура воздуха + 16 °С,
температура воды + 8 °С. Комментарии: дно каменистое, наблюдаются валуны,
течение достаточно быстрое. Сбор ручной. Был собран материал в количестве 34
особи.
Четвертая стация - Лесное озеро №1. Водоём
заболочен, заросший растительностью. Глубина незначительная. Температура воды +
9 °C. Температура
воздуха в момент забора пробы + 20 °C.
Представители отряда Ephemeroptera
не обнаружены.
Пятая стация - река Горелая Балка, правый
приток. Очень полноводный приток, уровень воды непостоянный. Из-за дождей
становится полноводной. Температура воды + 5 °C,
температура воздуха в момент забора пробы + 16 °C.
Дно каменистое. Вода слегка мутная. Координаты: высота 1082 м, 44°11'047"N
и 40°02'395"Е. Было обнаружено 44 особи.
Шестая стация - река Горелая балка, левый рукав
левого притока. Река Горелая балка - левый приток реки Мезмайки, представляет
собой небольшой ручеек. Дно - каменистое. Температура воды + 5 °C.
Температура воздуха в момент забора пробы + 19 °C.
В результате исследований было найдено 34 представителя отряда подёнок.
Седьмая точка - река Горелая Балка, правый рукав
левого притока. Пасмурно. Температура воды + 5 °C
, температура воздуха в момент забора пробы + 16 °C.
Дно - каменистое. Вода прозрачная. В результате исследований было найдено 32
представителей отряда подёнок.
Восьмая точка - река Горелая балка, левый
приток. Река Горелая балка - левый приток реки Мезмайки. Дно - каменистое.
Температура воды + 5 °C.
Температура воздуха в момент забора пробы + 17 °C.
В результате исследований было найдено 38 представителей отряда поденок.
Девятая точка - родниковый водоем возле Базов.
Водоем полностью освещён. По берегам растут ситник лягушачий, рогоз. Дно
илистое. Температура воды + 12 °C,
температура воздуха в момент забора пробы + 16 °C.
Максимальная глубина 100 см. Метод отбора - сачок. В результате проведённых
исследований, материал не обнаружен, т.к. экологические условия не характерны
для представителей данного отряда.
Десятая точка - Лесное озеро №2. Координаты:
44°11'363"N и
40°02'023"Е. Малооблачно температура воздуха + 14 °С, температура воды +
12 °С, материал не обнаружен. Комментарии: озеро практически не проточное, вода
на момент сбора была мутной, сбор осуществляли с помощью гидробиологического
сачка. В результате проведённых исследований, материал не обнаружен, т.к.
экологические условия не характерны для представителей данного отряда.
Одиннадцатая точка - ручей, выходящий из Лесного
озера №2. Водоём освещён частично. Грунт - каменистый. Вода прозрачная. Глубина
в месте сбора до 20 см. Температура воды + 5 °C,
температура воздуха в момент забора пробы + 16 °C.
Были обнаружены данные представители отряда в количестве 32 особей.
Двенадцатая стация - мочак по дороге на водопад
Малый. Мочак слабо освещен. Вода стоячая, загрязненная. Дно - илистое. В
результате проведённых исследований, материал не обнаружен, т.к. экологические
условия не характерны для представителей данного отряда.
Тринадцатая стация - мочак возле Лесного озера.
Вода стоячая, загрязненная. Дно - илистое. Мочак окружен растительностью.
Температура воды + 12 °C.
В результате проведённых исследований, материал не обнаружен, т.к.
экологические условия не характерны для представителей данного отряда.
Четырнадцатая стация - мочак возле биостанции.
Дно - илистое. Вода не проточная, загрязненная. Высота 1240 метров. В
результате проведённых исследований, материал не обнаружен, т.к. экологические
условия не характерны для представителей данного отряда.
Пятнадцатая стация - ручей на Шанхае.
Малооблачно, температура воздуха + 18 °С, температура воды + 7 °С. Комментарии:
ручей не постоянный, во время отсутствия дождей пересыхает, дно
каменисто-илистое. Протяженность ручья 250-300 метров. Средняя ширина ручья 40
см. Было отмечено 3 категории участков: в дистальной части - заболоченный
участок, затем проточная часть и пересыхающий участок. Были исследованы все три
категории ручья. Сбор проводили ручной, в результате было обнаружено 13 особей.
Шестнадцатая стация - река Курджипс. Река берёт
своё начало в районе горного массива Лагонаки, является левым приток реки
Белой. Это горная, быстротечная, крутопадающая река, протекающая по северному
склону Главного хребта Большого Кавказа. Температура воздуха в момент забора
пробы + 20 °C, температура воды
+ 7 °C. В результате
исследований было найдено 36 представителей отряда подёнок.
Семнадцатая стация - ручей на правобережье реки
Курджипс. Первые правобережные скальные стены поросли лесом. Вода проточная.
Дно - каменистое. Температура воздуха в момент забора пробы + 20 °C.
Температура воды + 7 °C.
Было найдено 34 представителей отряда подёнки.
Восемнадцатая стация - верховья реки
Курджипс. Это горная, быстротечная река, протекающая по северному склону
Главного хребта Большого Кавказа
<#"869654.files/image004.gif">
Рисунок 4 - Река Курджипс в месте сбора
материала

Рисунок 5 - Левый приток р. Горелая Балка в
месте сбора материала
Семейство Heptageniidaе семидневные подёнки
представлено родами: Ecdyonurus, Heptagenia. Распространение Heptagenia
fuscogrisea (Retzius,
1783) центральная Англия, Шотландия, Украина, Кавказ; Heptagenia
flava Rostock
1878 Австрия, Чехия, Дания, Франция, Нидерланды, Румыния; Heptagenia
sulphurea (Müller,
1776) Австрия, Чехия, Дания, Франция, Нидерланды, Румыния, Люксембург,
Норвегия, Словения, Великобритания; Ecdyonurus
venosus (Fabricius,
1775) Австрия, Чехия, Дания, Франция, Нидерланды, Румыния, Италия, Люксембург,
Норвегия, Словения, Великобритания, Ирландия. Семейство Baetidae
представлен родом Baetis, видом Baetis rhodani
(Pictet, 1845)
распространен Австрия, Чехия, Дания, Франция, Нидерланды, Румыния, Италия,
Люксембург, Норвегия, Словения, Великобритания, Ирландия, Бельгия, Португалия,
Россия - Кавказ.
Семейство Ephemerellidae представлен одним родом
Ephemerella, в Московской области Ephemerella
ignita (Poda,
1761), Распространение Австрия, Чехия, Канада, Великобритания, Кавказ.
Три из шести видов принадлежат к роду Heptagenia,
и по одному виду относятся к родам Ecdyonurus,
Baetis и Ephemerella.
В сравнении с фауной подёнок всего Кавказа, насчитывающей 59 видов, выявленный
нами видовой список составляет 10 % от фауны Кавказа. Всего в мировой фауне
более 1500 видов. Основные показатели водоёмов, в которых велись исследования,
и абсолютная численность собранных личинок подёнок, показаны в таблице 2.
Для сравнения данные по численности подёнок в
каждом из исследованных нами водоемов были сопоставлены и графически изображены
на рисунке 6 в виде гистограммы.
Таблица 2 - Абсолютная
численность подёнок в водоёмах заказника «Камышанова Поляна»
|
Место
сбора материала
|
Основные
параметры водоема: проточность, освещенность
|
Вид
и его численность, экз.
|
|
|
Heptagenia fuscogrisea
|
Ecdonurus venosus
|
Heptagenia flaga
|
Heptagenia sulfura
|
Baetis rhodani
|
Ephemerella ignita
|
|
1.Большой
водопад
|
проточный,
освещенный
|
13
|
--
|
--
|
--
|
--
|
--
|
|
2.
Родник на территории биостанции
|
проточный,
слабосвещенный
|
11
|
--
|
--
|
--
|
--
|
--
|
|
3.
Малый водопад
|
проточный,
слабосвещенный
|
15
|
--
|
--
|
--
|
--
|
10
|
|
4.
Лесное озеро№1
|
слабопроточный,
слабосвещенный
|
--
|
--
|
--
|
--
|
--
|
--
|
|
5.
Горелая Балка, правый приток
|
проточный,
слабосвещенный
|
21
|
12
|
--
|
--
|
--
|
10
|
|
6.
Горелая балка, левый рукав левого притока
|
проточный,
слабосвещенный
|
10
|
--
|
--
|
--
|
--
|
--
|
|
7.
Горелая Балка, правый рукав левого притока
|
проточный,
слабосвещенный
|
15
|
--
|
--
|
--
|
--
|
14
|
|
8.
Горелая балка, левый приток
|
проточный,
слабосвещенный
|
--
|
--
|
--
|
--
|
--
|
--
|
|
9.
родниковый водоем возле Базов
|
стоячий,
слабосвещенный
|
--
|
--
|
--
|
--
|
--
|
--
|
|
10.
Лесное озеро№2
|
стоячий,
освещенный
|
--
|
--
|
--
|
--
|
--
|
--
|
|
11.
Ручей, выходящий из Лесного озера
|
проточный,
слабосвещенный
|
12
|
--
|
--
|
--
|
--
|
-
|
|
12.
Мочак по дороге на водопад Малый
|
стоячий,
слабосвещенный
|
--
|
--
|
--
|
--
|
--
|
--
|
|
13.
Мочак возле Лесного озера
|
стоячий,
слабосвещенный
|
--
|
--
|
--
|
--
|
--
|
-
|
|
14.
Мочак возле биостанции
|
стоячий,
освещенный
|
--
|
--
|
--
|
--
|
--
|
--
|
|
15.
Ручей на Шанхае.
|
слабопроточный,
слабосвещенный
|
--
|
--
|
--
|
--
|
--
|
--
|
|
16.
Река Курджипс
|
проточный,
освещенный
|
18
|
--
|
--
|
--
|
--
|
--
|
|
17.
Ручей на правобережье реки Курджипс
|
проточный,
освещенный
|
13
|
--
|
--
|
--
|
--
|
12
|
|
18.
Верховья реки Курджипс
|
проточный,
освещенный
|
23
|
--
|
--
|
--
|
--
|
--
|
|
19.
Ручей щели Инструкторская
|
проточный,
освещенный
|
--
|
--
|
--
|
--
|
--
|
--
|
|
20.
Верховья реки Белая
|
проточный,
освещенный
|
15
|
--
|
--
|
--
|
--
|
--
|
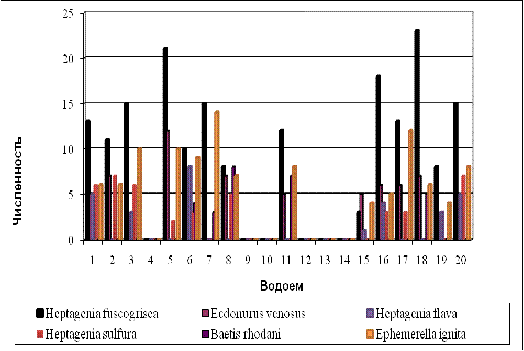
Рисунок 6 - Показатели относительной численности
различных видов подёнок в исследованных водоемах
Максимальной численностью
характеризуются виды Heptagenia
fuscogrisea, Ephemerella
ignita, Ecdyonurus
venosus - они встречаются
в большинстве водоемов: Большой и Малый водопады; реки Мезмай, Курджипс,
Горелая Балка с притоками, Белая.
В таких водоемах, как мочак по
дороге на водопад Малый, мочак возле Лесного озера, мочак возле биостанции, а
так же в родниковом водоеме возле Базов, Лесное озеро №2 поденки отсутствуют
полностью, все они характеризуются стоячей водой, илистым дном, заросшими
травой берегами, температура воды в данных водоёмах в среднем составляет +
10-12 °С, данные параметры в совокупности обеспечивают неблагоприятные условия
для жизни представителей.
Таблица 3 - Средняя численность
подёнок в исследуемых водоёмах
|
Вид
|
Число
наблюдений
|
Среднее
с ошибкой, экз.
|
Минимум/максимум,
экз.
|
|
Ecdyonurus venosus
|
20
|
3,00±0,824
|
0,00/12,00
|
|
Heptagenia fuscogrisea
|
20
|
9,25±1,700
|
0,00/23,00
|
|
Heptagenia flava
|
20
|
1,45±0,531
|
0,00/8,00
|
|
Heptagenia sulphurea
|
20
|
2,25±0,656
|
0,00/8,00
|
|
Baetis rhodani
|
20
|
1,20±0,501
|
0,00/7,00
|
|
Ephemerella ignita
|
20
|
5,45±0,985
|
0,00/14,00
|
В соответствии с рисунком 6, по
принципу отсутствия подёнок, точки сбора материала 4, 9, 10, 12-15 - это
объединение включило водоемы одного типа. Все они являются стоячими или слабо
проточными - мочаками или лесными озерами. В этих водоемах поденки не
встречены. Высокая степенью сходства поденок в различных притоков рек Горелая
Балка и Курджипс, которые объединяются на расстоянии объединения 5 км. Более значительные
различия имеют проточные водотоки родников и ручьев заказника, а также
примыкающая р. Белая.
Отличается фауна поденок
Большого водопада, которая имеет сходство с притоками р. Горелая Балка, так как
Большой водопад расположен на р. Горелая Балка после слияния всех ее притоков.
Отсутствие материала в мочаках
свидетельствует о том, что экологические условия не соответствуют, условиям
необходимым для существования поденок, отсутствие достаточного количества
кислорода, проточность, низкой температуры.
Проанализировав выше сказанное,
можно сказать о том, что на территории Апшеронского и Майкопского района
подёнки в большом количестве предпочитают водоёмы: проточные реки с быстрым
течением и каменистым дном. Значительно меньше видовое разнообразие характеризуют
загрязненные водоемы. Подёнки не были встречены в водоёмах: слабопроточных и
стоячих, илистым дном.
Так же проводили сбор материала
в водоемах Отрадненского района, нами было исследовано 7 точек, различающиеся
друг от друга по типу дна, количеству воды в русле, по питанию водоемов, по
течению. Нами было собрано 171 особь из трех семейств.
Первая стация р. Тегинь в районе станице
Малотегинской температура воздуха + 25оС, температура воды + 10 оС
погода ясная, дно каменистое, вода прозрачная поймано 10 особей из отряда
подёнки.
Вторая стация р. Уруп в районе
хутора Ильич. Малооблачно, температура воздуха + 23 оС, температура
воды + 10 оС, дно каменистое, поймано 15 особей, так как глубина
реки в месте сбора составила 38 см, что достаточно глубоко для представителей
данного отряда.
Третья стация р. Аргош в районе
станицы Передовая температура воздуха + 25оС, температура воды + 8 оС
дно каменистое поймано 18 особей.
Четвертая стация Картамук
температура воздуха + 21 оС воды + 7 оС малооблачно, дно
каменистое, встречено 43 особи.
Пятая стация Озеро, облачно
температура воздуха + 20 оС, температура воды + 8,5оС,
дно илистое особей не обнаружено, а в выходящем из него Ручье №1 на расстоянии
80 м встречено 35 особей температура воды + 8 оС
Шестая стация Ручей №2 в ст.
Надежная, облачно, температура воздуха + 18оС, вода + 7 оС,
дно каменистое встречено 27 особи.
Седьмая стация р. Кува в районе
хутора Ильич температура воздуха + 22оС, температура воды + 7,5 оС,
малооблачно, дно каменистое, встречено 23 особи.
Всего нами было встречено 171
особь, из 3 семейств: Ephemerella ignita (Poda, 1761), Baetis
rhodani (Pictet,
1845), Baetis
digitatus (Бенгтссон, 1912),
Epeorus caucasicus
(Tshernova, 1938).
Численность по видовому составу по приведена в таблице 4.
Таблица 4 - Абсолютная
численность по видовому составу подёнок рек Отрадненского района
|
Место
сбора материала
|
Основные
параметры водоема
|
Вид
и его численность, экз.
|
|
|
Ephemerella
ignita
|
Baetis
rhodani
|
Baetis
digitatus
|
Epeorus
caucasicus
|
|
1.
Тегинь
|
проточный,
освещенный
|
--
|
--
|
--
|
--
|
|
2.
Уруп
|
проточный,
освещенный
|
--
|
--
|
--
|
-
|
|
3.
Аргош
|
проточный,
освещенный
|
--
|
--
|
--
|
--
|
|
4.
Картамук
|
проточный,
освещенный
|
18
|
10
|
--
|
--
|
|
5.
Озеро
|
слабопроточный,
слабосвещенный
|
--
|
--
|
--
|
--
|
|
6.
Ручей №1
|
проточный,
слабосвещенный
|
12
|
12
|
--
|
--
|
|
7.
Ручей №2
|
проточный,
слабосвещенный
|
--
|
--
|
--
|
--
|
|
8.
Кува
|
проточный,
освещенный
|
--
|
--
|
--
|
--
|
Вид Baetis digitatus
встречается на территориях Германия, Австрия, Великобритания для Кавказа не
отмечен; вид Epeorus
caucasicus распространен на
территории Кавказа, данный вид является эндемиком. Epeorus
caucasicus, отмечен только на
Кавказе.
Было обнаружено 4 вида из трех
семейств, два вида из семейства Baetidae
Baetis rhodani, Baetis digitatus; и по одному виду из семейств Ephemerellidae
Ephemerella ignita и Еpeoridae
Epeorus caucasicus. В сравнени с фауной «Камышановой Поляны» имеются различия
по двум видам Baetis digitatus. Epeorus caucasicus. Средняя численность подёнок
приведена в таблице 5.
Таблица 5 - Средняя численность
подёнок в исследуемых водоемах Отрадненского района
|
Вид
|
Число
набл.
|
Среднее
с ошибкой, экз.
|
Минимум/максимум,
экз.
|
|
Ecdyonurus venosus
|
8
|
8,25±1,704
|
0,00/18,00
|
|
Baetis
rhodani
|
8
|
5,81±0,942
|
0,00/12,00
|
|
Baetis
digitatus
|
8
|
4,13±0,640
|
0,00/8,00
|
|
Epeorus caucasicus
|
8
|
3,21±0,780
|
0,00/7,00
|
Наиболее встречаемым оказался
вид Ephemerella ignita так как представители вида имеют широкий ареал
распространения, переносят температуру воды плюс 6-10 оС, наименее
встречаемым был вид Epeorus caucasicus, так как благоприятными условия для
существования вида необходимо температура воды + 6 оС, тип дна
галька, большое количество кислорода.
В таблице 6 представлены
количество семейств, родов и видов встреченных в исследуемых водоёмах
Апшеронского, Майкопского и Отрадненского районов.
Таблица 6 - Количество
семейств, родов, видов в исследуемых водоёмах
|
Отряд
|
Семейство
|
Род
|
Вид
|
|
Ephemeroptera
|
4
|
5
|
8
|
Наиболее многочисленное
семейством по количеству видов это Heptageniidaе четыре из восьми видов
принадлежат к данному семейству. По два вида из семейства Baetidae,
и по одному виду из семейств Ephemerellidae
и Epeoridae.
ЗАКЛЮЧЕНИЕ
В 2013 г. нами было проведено изучение видового
состава и особенностей экологии подёнок в различных водоемах Лагонакского
нагорья, а так же в некоторых водоемах Отрадненского района. Для изучения были
использованы общепринятые методики проведения сборов личинок, ведения
наблюдений, этикетирования, коллекционирования и обработки количественных
данных. Результаты сформулированы в следующие выводы:
. Видовой состав подёнок в исследуемых водоёмах
Апшеронского и Майкопского района насчитывает 6 видов, из трех семейств, и трех
родов, что составляет 10 % от общего числа видов Северо-Западного Кавказа.
2. В исследуемых водоемах Апшеронского и
Майкопского района преобладает вид Heptagenia
fuscogrisea численность
которого составляет 9,25 ± 1,700. Минимальной средней численностью
характеризуется Baetis rhodani - составляет 1,20 ± 0,501 особей.
3. В водоёмах Апшеронского и Майкопского
районах подёнки в большом количестве предпочитают водоёмы: проточные реки с
быстрым течением и каменистым дном. В водоемах с не проточной или
слабопроточной водой, илистым дном, средней температурой воды + 10 °С подёнки
родов: Heptagenia,
Ecdyonurus, Baetis
и Ephemerella не встречаются,
так как являются олигосапробами.
. Видовой состав подёнок водоёмов
Отрадненского района насчитывает 4 вида, из трех семейств и трех родов.
Наиболее встречаемым оказался вид Ephemerella ignita так как представители вида
имеют широкий ареал распространения, переносят температуру воды плюс 6-10 оС,
наименее встречаемым был вид Epeorus caucasicus, так как благоприятными условия
для существования вида необходимо температура воды + 6 оС, тип дна
галька, большое количество кислорода.
БИБЛИОГРАФИЧЕСКИЙ СПИСОК
1. Байкова
О. Я. К познанию поденок бассейн Амура // Изв. ТИНРО. СПб., 1970. С. 207-232.
. Байкова
О. Я. Новые и малоизвестные виды поденок (Ephemeroptera) из бассейна Амура //
Энтомологическое обозрение. 1972. Т. 49, Вып. 1. С. 146-155.
. Бродский
К. A. К познанию Ephemeroptera Южной Сибири // Энтомологическое обозрение.
1976. Т. 24, Вып. 2. С. 31-40.
. Борисов
В. И. Реки Кубани. Краснодар, 1976. 38с.
. Голубков
С. М., Тиунова Т. М. Влияние содержания кислорода в воде на интенсивность
дыхания реофильных личинок подёнок // Тез. Докл. XI Всесоюзн. Симпоз.
Биологические проблемы Севера. 1986. Вып. 4. С. 83-84.
. Жадин
М. В. Фауна рек и водохранилищ. М. 1940. 259с.
. Жильцова
Л. А. Отряд Plecoptera. Веснянки // Определитель насекомых Европейской части
СССР. Изд-во АН СССР. 1964. Т. 1. С. 177-200.
8. Жильцова
Л. А. Новые и малоизвестные виды веснянок (Plecoptera, Nemouridae, Capnidae,
Leuctridae) фауны СССР // Энтомологическое обозрение. Т. 68, Вып.З. 1989. С.
582-586.
9. Жильцова
Л. А., Черчесова С. К. Описание личинок Perla pallida // Энтомологическое
обозрение. 2003. Т. 2, Вып. 85. С. 23-326.
. Заика
В. В. Особенности формирования фауны реофильных амфибионтных насекомых горных
территорий Тувы // Животный мир горных территорий. М., 2009. С. 46-50.
. Канонников
А. М. Природа Кубани и Причерноморья. Краснодар, 1984. 75с.
. Касымов
А. Г. Биология бокоплава Pontogammarus robustoides // Зоологический журнал.
1972.Т. 8, Вып. 35. С. 356-357.
. Клюге
Н. Ю. Систематика и филогения подсемейства Heptegeniinae (Ephemeroptera,
Heptageniidae). М., 1985. 14с.
. Клюге
Н. Ю. Новый вид поденок (Ephemeroptera, Heptageniidae) из Закавказья.
Зоологический журнал. 1987. Т. 5, Вып. 66. С. 775-777.
. Клюге
H. Ю. Ревизия родов сем. Heptageniidae
(Ephemeroptera) //
Энтомологическое обозрение. 1993. Т. 7, Вып. 12. С. 39-54.
. Клюге
H. Ю. Каталог
типовых экземпляров коллекции Зоологического Института РАН. Подёнок (Ephemeroptera).
СПб., 1995. 52с.
. Клюге
H. Ю. Определитель пресноводных беспозвоночных России и сопредельных
территорий: в 6 т. Т. 3: Паукообразные и Низшие насекомые. СПб., 1997. 439 с.
. Кустарева
Л. А. Подёнки рода Iron (Ephemeroptera, Heptageniidae) фауны Средней Азии //
Энтомологическое обозрение. 1984. Т. 3, Вып. 47. С. 14-25.
19. Леванидов В. Я. Биомасса и структура
донных биоценозов малых водотоков Чукотского полуострова // Пресноводная фауна
Чукотского полуострова. Владивосток, 1976. С. 104-122.
20. Леванидова
И. М. Амфибиотические насекомые горных областей Дальнего Востока СССР. СПб.,
1982. 214 с.
. Мамаев
В. И., Медведев Л. Н., Правдин Ф. Н. Определитель насекомых Европейской части
СССР: в 5 т. Т. 2. М., 1976. 303 с.
. Мартынов
A. B. Ручейники. Определители по фауне СССР. Т. 3. СПб., 1934. 343 с.
. Макаров
К. В. Личинка пещерной жужелицы Jeannelius birsteini // Энтомологическое
обозревание. 2000. Т. 4, Вып. 47. С. 836-845.
. Новикова
E. A. Подёнки семейства Baetidae (Ephemeroptera). Краснодар, 1987. 16 с.
. Панов
В. Д. Ледники бассейна реки Терек, 1971. 287 с.
. Синиченкова
Н. Д. Личинки подёнок палеарктичуских видов рода Rhithrogena (Ephemeroptera,
Heptageniidae) // Вестник МГУ. 1973. Т.5, Вып. 39. С. 9-17.
. Синиченкова
Н. Д. Подёнки рода Iron (Ephemeroptera, Heptageniidae) фауны Кавказа //
Энтомологическое обозрение. 1976. Т. 5, Вып. 43. С. 853-862.
. Синиченкова
Н. Д. Новый вид подёнок фауны Кавказа // Энтомологическое обозревание. 1979. Т.
2, Вып. 34. С. 7-9.
. Синиченкова
Н. Д. Подёнки рода Rhithrogena фауны Кавказа // Энтомологическое обозрение.
1979. Т. 5, Вып. 23. С. 811-820.
. Цалохин
С. Я. Определитель пресноводных беспозвоночных России и сопредельных
территорий. М., 2001. 345 с.
. Чернова
О. А. К познанию подёнок (Ephemeroptera) Восточного Закавказья. Тр. Зоол. ин-та
Азерб. фил. АН СССР. 1938. Т. 4, Вып. 32. С. 55-64.
. Чернова
О. А. Фауна подёнок Европейского Севера СССР // Зоологический журнал. 1941. Т.
7, Вып. 23. С. 213-236.
. Чернова
О. А. Географическое распространение подёнок (Ephemeroptera) и особенности
фауны бассейна Амура // Энтомологическое обозрение. 1958. Т. 3, Вып. 49. С.
64-84.
. Чернова
О. А. Определитель насекомых европейской части СССР: в 5 т. Т. 1: Низшие,
древнекрылые, с неполным превращением. М., 1964. С. 110.
. Хаберман
X. М. Материалы по фауне подёнок (Ephemeroptera) Эстонской ССР // 1952. Т. 2,
Вып. 23. С. 214-225.
. Хатухов
А. М., Якимов А. В. О фауне диплозоид (Diplozoidae) бассейна Терека //
Материалы докладов IX Международной конференции «Биологическое разнообразие
Кавказа». Махачкала, 2007. С. 249-250.
. Шиолошвили
М. Н. Бентос р. Малка // Вестник МАНЭБ. 2008. Т. 13, Вып. 3. С. 84-87.
38. Kluge
N. Ju. Pterothorax structure of mayflies (Ephemeroptera) and its use in
systematics. Bull // Soc. Entomol. France 1994. Vol. 99 (1). P. 41-61.
. Kluge
N. Ju. New subgenera of Holarctic mayflies (Ephemeroptera: Heptageniidae,
Leptophlebiidae, Ephemerellidae) // Zoosyst. Rossica. 2009. Vol. 34, P.
233-235.
. Zwick
P. Plecoptera aus Anatolien und benachbarten Gebieten // Mitt. Schweiz.
Entomol. Ges. Bd. 44, Hf. 3-4. 1971. S. 233-264.
41. Zimmermann
W. Rhilhrogena leberdensis n. sp. eine neue Heptageniidae (Ephemeroptera) aus
dem Einzugsgebiet der Teberda (UdSSR, Westkaukasus). Entomol. Nachr. 1981. Bd.
27. S. 17-27.