Нуклеиновые кислоты, их строение и роль в биологических процессах
ГОСУДАРСТВЕННОЕ
БЮДЖЕТНОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО ПРОФЕССИОНАЛЬНОГО ОБРАЗОВАНИЯ
«КУБАНСКИЙ
ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ»
МИНИСТЕРСТВА
ЗДРАВООХРАНЕНИЯ
РОССИЙСКОЙ
ФЕДЕРАЦИИ
(ГБОУ
ВПО КубГМУ Минздрава России)
Реферат
На тему:
Нуклеиновые кислоты, их строение и роль в биологических процессах
Дисциплина:
Биология
Выполнила:
Оникиенко .А. П.
Студентка
1 курса, _ группы
Специальность:
060301 - «Фармация»
Преподаватель:
Голубцов В. И.
Краснодар
2014 год
Введение
Нуклеиновые кислоты как один из компонентов
живой материи были открыты в 1869 году швейцарским ученым Иоганом Мишером.
Однако бурное развитие химии и биохимии нуклеиновых кислот началось в конце
40-х - начале 50-х годов ХХ века, когда было установлено, что один из двух главных
типов нуклеиновых кислот - дезоксирибонуклеиновая кислота (ДНК) является
носителем наследственной информации.
На протяжении второй половины XX века
нуклеиновые кислоты были важнейшим объектом исследований химии и биологии.
Химики создали методы установления детальной химической структуры нуклеиновых
кислот, их искусственного синтеза, изучили их поведение при разных химических
воздействиях. Биохимики направили свои усилия на выяснение многочисленных
аспектов функционирования нуклеиновых кислот в живых организмах или выделенных
из них системах. Поскольку было выяснено, что строение молекул нуклеиновых
кислот специально приспособлено для выполнения некоторых основополагающих
биологических функций, область биохимии, изучающая поведение нуклеиновых кислот
в живых и модельных системах, обособилась в автономную область знания,
получившую название молекулярная биология.
Между тем изучение нуклеиновых кислот продолжает
оставаться одной из самых горячих точек на переднем крае современной науки.
. История открытия нуклеиновых кислот
Нуклеиновые кислоты - биологические полимерные
молекулы, хранящие всю информацию об отдельном живом организме, определяющие
его рост и развитие, а также наследственные признаки, передаваемые следующему
поколению. Нуклеиновые кислоты содержатся в ядрах клеток всех растительных и
животных организмов и выполняют важную биологическую роль.
Открытие нуклеиновых кислот как класса
органических веществ, связано с именем молодого швейцарского врача Фридриха
Мишера, работавшего в Тюбингенской физиолого-химической лаборатории, известной
в середине 19 века всему учёному миру. Изучая химический состав лейкоцитов
гноя, молодой учёный получил вещество с необычными для известных тогда
органических соединений свойствами. Мишер предположил, что это вещество
сдержится в ядре клетки. В то время ещё никто в биохимических лабораториях не
пытался выделить ядра или какие-либо другие субклеточные компоненты. Поэтому
Мишер предпринял более тщательные эксперименты по выделению клеточных ядер. В
конечном итоге, исследователь повторно выделил новое вещество. Оно не
разлагалось протеолитическими ферментами, то есть не являлось белком.
Отсутствие растворимости в горячем спирте указывало на то, что вещество не
являлось фосфолипидом. По-видимому, оно относилось к новому классу
биохимических соединений. Ввиду ядерного происхождения Мишер предложил для нег
название «нуклеин» (от лат. nucleus - ядро).
В дальнейшем Ф. Мишер использовал другой
материал для изучения состава ядер клеток. Он исследовал сперматозоиды - молоки
рейнского лосося. Исследования дали новые важные результаты: удалось разделить
нуклеин на белок (протамин), необходимый для стабилизации молекулы - полимера и
кислый остаток. Свободный от белка остаток нуклеина был назван в 1889 году
нуклеиновой кислотой.
Так были открыты нуклеиновые кислоты и новая
группа сложных белков - нуклеопротеины, содержащие в качестве простетической
группы нуклеиновые кислоты.
2. Структурно-функциональная организация
нуклеиновых кислот
Как известно, основная биологическая роль ДНК
сводится к хранению и передаче наследственной информации. Поэтому основное
требование, которое природа предъявила к структуре молекул ДНК, заключается в
стабильности ее структуры в физиологических условиях, обеспечивающей
сохранность генетической информации.
Подобно белкам, нуклеиновые кислоты -
биополимеры, а их функция заключается в хранении, реализации и передаче
генетической (наследственной) информации в живых организмах.
Существует два типа нуклеиновых
кислот - дезоксирибонуклеиновые (ДНК) и рибонуклеиновые (РНК). Мономерами в
нуклеиновых кислотах служат нуклеотиды. Каждый из них содержит азотистое
основание, пятиуглеродный сахар (дезоксирибоза - в ДНК, рибоза - в РНК) и
остаток фосфорной кислоты.
В ДНК входят четыре вида
нуклеотидов, отличающихся по азотистому основанию в их составе, - аденин (А),
гуанин (Г), цитозин (Ц) и тимин (Т). В молекуле РНК также имеется 4 вида
нуклеотидов с одним из азотистых оснований - аденином, гуанином, цитозином и
урацилом (У). Таким образом, ДНК и РНК различаются как по содержанию сахара в
нуклеотидах, так и по одному из азотистых оснований (табл. 1).
Таблица 1. Компоненты
нуклеотидов ДНК и РНК
|
Нуклеиновая
кислота
|
Пятиуглеродный
сахар
|
Азотистые
основания
|
Остаток
фосфорной кислоты
|
|
ДНК
|
Дезоксирибоза
|
Аденин,
гуанин, цитозин, тимин
|
Остаток
фосфорной кислоты
|
|
РНК
|
Рибоза
|
Аденин,
гуанин, цитозин, урацил
|
Остаток
фосфорной кислоты
|
Молекулы ДНК и РНК существенно
различаются по своему строению и выполняемым функциям (табл. 2).
Таблица 2. «Сравнительная
характеристика ДНК и РНК»
|
Признаки
|
ДНК
|
РНК
|
|
Функции
|
1.
химическая основа хромосомного генетического материала (генов); 2. матрица
для синтеза ДНК; 3. матрица для синтеза РНК; 4. информация о структуре белка
|
1.
иРНК передает код наследственной информации о первичной структуре белка; 2.
рРНК входит в состав рибосом; 3. тРНК переносит аминокислоты к рибосомам; 4.
митохондриальная и пластидная ДНК входят в состав этих органоидов
|
|
Местонахождение
в клетке
|
ядро,
митохондрии, пластиды
|
ядро,
цитоплазма, рибосомы, митохондрии, пластиды
|
|
Строение
|
двойная
спираль: две комплементарные полинуклеотидные цепи
|
одинарная
полинуклеотидная цепь
|
|
Мономеры
|
дезоксирибонуклеотиды
|
рибонуклеотиды
|
|
Состав
нуклеотида
|
азотистое
основание (аденин, гуанин, тимин, цитозин), дезоксирибоза и остаток фосфорной
кислоты
|
азотистое
основание (аденин, гуанин, урацил, цитозин), рибоза и остаток фосфорной
кислоты
|
|
Типы
нуклеотидов
|
адениловый
(А), гуаниловый (Г), тимидиловый (Т), цитидиловый (Ц)
|
адениловый
(А), гуаниловый (Г), уридиловый (У), цитидиловый (Ц)
|
|
Свойства
|
способна
к репликации (самоудвоению), стабильна
|
не
способна к репликации, лабильна
|
2.1 Строение ДНК
ДНК (дезоксирибонуклеиновая
кислота) - нуклеиновая кислота, которая является основным компонентом хромосом
клеток эукариот и некоторых вирусов. ДНК часто называют «строительным
материалом» жизни, поскольку в ней хранится генетический код, являющийся
основой наследственности. Молекулярную структуру ДНК впервые установили Джеймс
Уотсон и Френсис Крик в 1953 г. Она состоит из двойной спирали, сложенной двумя
длинными лентами чередующихся молекул сахара (дезоксирибозы) и фосфатных групп,
связанных азотистыми основаниями. В целом молекула имеет форму, напоминающую
скрученную веревочную лестницу, перекладинами которой служат азотистые
основания - АДЕНИН (А), ЦИТОЗИН (С), ГУАНИН (G) и тимин (Т).
Молекула ДНК может включать
огромное количество нуклеотидов - от нескольких тысяч до сотен миллионов
(поистине гигантские молекулы ДНК удается «увидеть» с помощью электронного
микроскопа). В структурном отношении она представляет собой двойную спираль из
полинуклеотидных цепей (рис. 1), соединенных с помощью водородных связей между
азотистыми основаниями нуклеотидов. Благодаря этому полинуклеотидные цепи
прочно удерживаются одна возле другой.
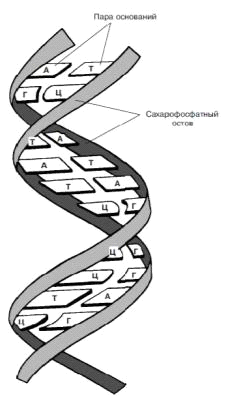
Две цепи обвиты одна вокруг
другой. Каждая цепь (изображенная в виде ленты) состоит из чередующихся
остатков сахара и фосфатных групп. Водородные связи между азотистыми
основаниями (А, Т, Г и Ц) удерживают две цепи вместе.
При исследовании различных ДНК
(у разных видов организмов) было установлено, что аденин одной цепи может
связываться лишь с тимином, а гуанин - только с цитозином другой.
Следовательно, порядок расположения нуклеотидов в одной цепи строго
соответствует порядку их расположения в другой. Этот феномен получил название
комплементарности(т. е. дополнения), а противоположные полинуклеотидные цепи
называются комплементарными. Именно этим обусловлено уникальное среди всех
неорганических и органических веществ свойство ДНК - способность к
самовоспроизведению или удвоению (рис. 2).
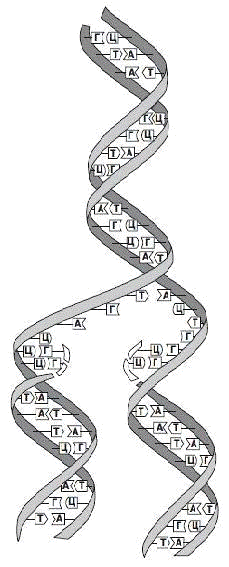
Рис. 2. Репликация ДНК
Двойная спираль
«расстегивается» по слабым водородным связям, соединяющим комплементарные
основания двух цепей. Каждая из старых цепей служит матрицей для образования
новой: нуклеотиды с комплементарными основаниями выстраиваются против старой
цепи и соединяются друг с другом.
При этом сначала
комплементарные цепи молекул ДНК расходятся (под воздействием специального
фермента происходит разрушение связей между комплементарными нуклеотидами двух
цепей). Затем на каждой цепи начинается синтез новой («недостающей»)
комплементарной ей цепи за счет свободных нуклеотидов, всегда имеющихся в
большом количестве в клетке. В результате вместо одной («материнской») молекулы
ДНК образуются две («дочерние») новые, идентичные по структуре и составу друг
другу, а также исходной молекуле ДНК. Этот процесс всегда предшествует
клеточному делению и обеспечивает передачу наследственной информации от материнской
клетки дочерним и всем последующим поколениям.
Первичная структура ДНК.
Последовательность чередования нуклеотидов в
полинуклеотидной цепи ДНК составляет ее первичную структуру. Для обозначения
последовательности первичной структуры полинуклеотидной цепи используют
однобуквенные символы образующих ее нуклеотидов, которые идентичны символам
соответствующих азотистых оснований.
Определение первичной структуры ДНК - крайне
сложная и трудная задача, так как размеры молекул огромны, но при этом она
построена всего лишь из четырех типов нуклеотидов. Большие успехи в изучении
структуры ДНК были достигнуты в результате работ Э. Чаргаффа с сотрудниками,
которым впервые с помощью хроматографического метода удалось определить
нуклеотидный состав ДНК, выделенной из различных природных объектов. Оказалось,
что ДНК, выделенные из разных источников, существенно отличались по
количественному нуклеотидному составу (табл. 22), но во всех случаях
нуклеотидный состав молекул ДНК подчиняется универсальным закономерностям, которые
получили название правил Чаргаффа (табл. 3).
Таблица 3
|
ПРАВИЛА
ЧАРГАФФА
|
|
1
правило
|
2
правило
|
3
правило
|
4
правило
|
|
Суммарное
содержание пуриновых нуклеотидов равно суммарному содержанию пиримидиновых:
|
Содержание
тимина равно содержанию аденина:
|
Содержание
гуанина равно содержанию цитозина:
|
Количество
аденина и цитозина равно количеству гуанина и тимина:
|
|
А
+ Г = Ц + У + Т
|
А
= Т
|
Г
= Ц
|
А
+ Ц = Г + Т
|
*Такие соотношения не свойственны РНК
Вторичная структура ДНК.
Расшифровка вторичной структуры ДНК - одно из
крупнейших открытий молекулярной биологии, поскольку благодаря ему был раскрыт
механизм передачи наследственной информации в ряду поколений. В 1953 г.
американский биохимик Д. Уотсон и английский физик Ф. Крик на основании
большого числа экспериментальных данных (картины дифракции рентгеновских лучей
на нити ДНК) предложили модель структуры молекулы ДНК, существующей в водном
растворе. В основе модели Уотсона и Крика заложены следующие основные
положения:
. Молекулы ДНК построены из двух полинуклеотидных
цепей, ориентированных антипараллельно и по всей длине связанных друг с другом
водородными связями (причем в образовании водородных связей участвует каждый
мононуклеотид).
. Водородные связи между цепями образуются за
счет специфических взаимодействий остатка аденина одной цепи с остатком тимина
другой цепи (пара А···Т) и остатка гуанина одной цепи с остатком цитозина
другой цепи (пара Г···Ц).
. Первичная структура одной цепи молекулы ДНК в
составе двойной цепи комплементарна первичной структуре другой цепи. Это
положение легко понять на примере следующей схемы:
(5′ -> 3′)
А -Т - Т - Ц - Т - Ц - Г - T - Ц - Г - Г
T- A - A - Г
- A - Г
- Ц
- A - Г
- Ц
- Ц
(3′->5′)
Если в положении n (считая с 5′-конца)
первой цепи находится остаток аденина, то в положении n (считая с 3′-конца)
второй цепи находится комплементарный ему остаток тимина, а не другое азотистое
основание. Таким образом, зная первичную структуру одной цепи ДНК и используя
принцип комплементарности азотистых оснований, можно легко записать первичную
структуру другой цепи.
. Обе цепи закручены в спираль - двойная спираль
или дуплекс, которая имеет общую ось (рис. 32). При этом цепи могут быть
разделены только путем раскручивания (такие спирали называют
плектонемическими). Азотистые основания обращены внутрь спирали; их плоскости
перпендикулярны оси спирали и параллельны друг другу, образуя внутри спирали
стопку оснований. Между основаниями в этой стопке возникают гидрофобные, или
стэкинг- взаимодействия, которые, наряду с водородными связями, способствуют
стабилизации структуры двойной спирали в пространстве. Азотистые основания
упакованы очень плотно и не контактируют с окружающим водным раствором.
Третичная структура ДНК.
В частицах вирусов, клетках бактерий и высших
организмов, молекулы ДНК плотно «упакованы» и образуют довольно сложные
структуры. Например, в хромосоме E. coli содержится молекула ДНК длиной более 1
мм, хотя длина самой клетки не превышает 5 мкм. Вирусную ДНК можно отнести к
сравнительно мелким полимерным биомолекулам, но если ее вытянуть, то она
окажется во много раз длиннее, чем сам вирус. Для измерения длины молекул
нуклеиновых кислот в биохимии введена специальная единица длины, равная 1 000
пар нуклеотидов в случае двухцепочечных молекул нуклеиновых кислот - т.п.н. или
kb (от англ. kilobase - тысяча) или 1000 нуклеотидов в случае одноцепочечных
молекул - т.н. или kb. Так единица длины одноцепочечной ДНК величиной в 1 kb
имеет контурную длину 0,34 мкм и массу около 660 000. Выделенные из вирусных
частиц молекулы ДНК имеют либо линейную, либо кольцевую форму. Линейные
молекулы ДНК in vivo свертываются в плотный клубок. В таком состоянии они более
устойчивы к деградации. Например, кольцевая форма ДНК находится в фаге ϕΧ174.
Кольцевую ковалентно-связанную структуру имеют двухцепочечные ДНК бактерий,
вирусов, плазмид, митохондрий и др. Двухцепочечные кольцевые ДНК легко
переходят в суперспирализованное состояние, которое обеспечивает более плотную
упаковку громадной молекулы ДНК в малом объеме ядра или клетки (Рис. 3).
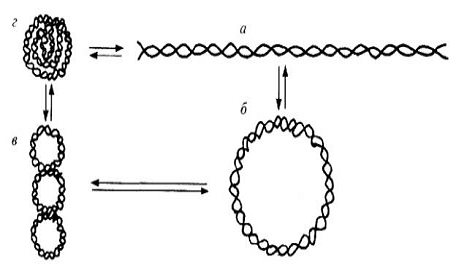
Рис. 3. Третичная структура молекул ДНК: а -
линейная; б - кольцевая; в - суперспирализованная (суперкольцевая); г -
компактный клубок
Длина молекул ДНК в среднем составляет около 3-5
см, а длина хромосом - всего несколько микрометров. Следовательно, степень
укладки ДНК в хромосомах благодаря дополнительному скручиванию нуклеосомной
нитки бус достигает нескольких тысяч. Высшие уровни укладки ДНК еще
недостаточно хорошо изучены.
.2 Строение РНК
РНК (рибонуклеиновая кислота) -
Нуклеиновая кислота, состоящая из рибонуклеотидов, участвует в процессах
реализации генетической информации. У некоторых вирусов в виде одно- или
двухцепочечных молекул может быть основным носителем наследственной информации.
В связи с тем, что молекулы РНК
имеют одноцепочечную структуру, их состав, в отличии от ДНК, не подчиняется
правилам Чаргаффа. Поэтому, как вторичная, так и третичная структуры РНК не
регулярны по строению.
Выделяют три вида РНК,
различающиеся по величине молекул и выполняемым функциям, - матричную (мРНК)
или информационную (иРНК), рибосомальную (рРНК) и транспортную (тРНК).
Три вида РНК
|
РНК
|
Число
нуклеотидов в молекуле
|
|
Матричные
(Информационные)
|
До
30 000
|
|
Рибосомальные
|
До
6000
|
|
Транспортные
|
Около
100
|
Матричная РНК
Открыта в 1961 году Жакобом и
Мано. Она составляет всего 2-3% от общего количества РНК клетки. Эта РНК не
имеет жесткой специфической структуры и ее полинуклеотидная цепь образует
изогнутые петли. В нерабочем состоянии м-РНК собрана в складки, свернута в
клубок, связана с белком; а во время функционирования цепь расправляется.
Матричные РНК синтезируются на ДНК в ядре. Процесс называется транскрипция
(списывание).
Роль мРНК - она несет
информацию об аминокислотной последовательности (т.е. о первичной структуре)
синтезируемого белка. Место каждой аминокислоты в молекуле белка закодировано
определенной последовательностью нуклеотидов в цепи м-РНК, т.е. в м-РНК имеются
«кодовые слова» для каждой аминокислоты - триплеты, или кодоны, или
генетические коды.
Свойства генетического кода. Генетическому
коду присущи:
) триплетность. Из 4-х
возможных мононуклеотидов м-РНК (УМФ, ГМФ, АМФ, ЦМФ) можно построить по
правилам перестановки 64 кодона. 61 кодон шифрует 20 аминокислот, а 3 кодона
(УАА, УАГ, УГА) не кодируют ни одной аминокислоты. Они играют роль
терминирующих (или «стоп-кодонов»), т.к. на них останавливается синтез п/п
цепи. Полный кодовый словарь представлен на таблице;
) неперекрещиваемость -
списывание информации идет только в одном направлении;
) непрерывность - код является
линейным, однонаправленным; не прерывается. Работает по принципу: одна
м-РНК-один белок
) универсальность, т.е. одна и
та же аминокислота у всех живых организмов кодируется одинаковыми кодами у всех
живых существ;
) вырожденность (избыточность).
Первые две буквы кодона определяют его специфичность, третья менее специфична.
Известно 20 аминокислот, а кодонов 61, следовательно, большинство аминокислот
кодируется несколькими кодонами (2-6).
Таким образом, мРНК принимает
непосредственное участие в биосинтезе белка. Основной постулат молекулярной
биологии, показывающий направление переноса генетической информации: ДНКàРНКàБелок.
Однако в 1974 году американские ученые Темин и Балтимор показали возможность
считывания информации и в обратном направлении с РНК на ДНК: ДНК↔РНКàбелок.
Этот процесс идет с участием фермента ревертазы. С его помощью можно
синтезировать участок ДНК по мРНК и перенести этот синтезированный ген в другие
объекты, что используется генной инженерией.
Рибосомальная РНК
На долю этого вида РНК
приходится более 80% от всей массы РНК клетки. Она входит в состав рибосом.
Рибосомы - это РНП, состоящие на 65% из р-РНК и на 35% из белка.
Полинуклеотидная цепь рРНК легко изгибается и укладывается вместе с белком в
компактные тельца.
Рибосома состоит из 2-х субъединиц
- большой и малой (соотношение их 2,5:1). В рибосоме различают 2 участка - А
(аминокислотный, или участок узнавания) и Р - пептидный, здесь присоединяется
п/п цепь. Эти центры расположены на контактирующих поверхностях обеих
субъединиц. Рибосомы могут свободно перемещаться в клетке, что дает возможность
синтезировать белки в клетке там, где это необходимо. Рибосомы мало специфичны
и могут считывать информацию с чужеродных мРНК, вместе с мРНК рибосомы образуют
матрицу.
Роль рРНК - обуславливает
количество синтезируемого белка.
Транспортная РНК
Этот вид изучен лучше всего,
составляет 10% всей клеточной РНК. Содержится в цитоплазме, молекулярная масса
небольшая (20тыс.Da) состоит из 70-80 нуклеотидов.
Основная роль - транспорт и
установка аминокислот на комплиментарном кодоне мРНК.
· акцепторный стебель
с последовательностью нуклеотидов АЦЦ, к нему присоединяется аминокислота.
· участок для
присоединения к рибосоме;
· антикодон -
участок, комплиментарный кодону мРНК, который кодирует аминокислоту,
присоединенную к данной тРНК .
Особенностью первичной
структуры тРНК является то, что они содержат минорные, или модифицированные
основания (7-метилгуанин, гипоксантин, дигидроурацил, псевдоурацил,
4-тиоурацил), которые способны к неклассическому спариванию. Это ускоряет
белковый синтез. Таким образом, тРНК «метит» аминокислоту, придавая ей
специфичность и способствует установлению аминокислоты на определенный участок
мРНК.
генетический нуклеиновый кислота
рибосомальный
3. Физико-химические свойства нуклеиновых
кислот
Уникальные биохимические и физико-химическими
свойства нуклеиновых кислот определяются высокой молекулярной массой,
особенностями химического состава и структурной организации на различных
уровнях надмолекулярного строения. Среди характерных физико-химических свойств
нуклеиновых кислот (и их растворов) следует выделить самые главные:
кислотно-основные свойства, хелатирующую способность, способность к
денатурации; оптические, коллоидные, осмотические свойства и высокую вязкость
растворов. Кроме того, нуклеиновые кислоты в среде живой клетки могут
находиться в жидкокристаллическом состоянии, что является крайне важным при
описании их биохимических свойств. При растворении в воде нуклеиновые кислоты
набухают и образуют вязкие коллоидного типа растворы. Растворимость нуклеиновых
кислот главным образом определяется гидрофильностью фосфатных групп, а также
особенностями надмолекулярной упаковки.
.1 Кислотно-основные свойства
В физиологических условиях среды (рН ~ 7,4)
фосфатные группы в молекулах ДНК и РНК полностью ионизированы, поэтому в
условиях внутренней среды живых организмов нуклеиновые кислоты существуют в
форме полианионов, т.е. несут множество отрицательных зарядов. Фосфатные группы
нуклеиновых кислот сильно полярны и характеризуются значениями рКI < 2.
Поверхность нуклеиновых кислот в целом несет отрицательный заряд, т.к. внешние
фосфатные группы полностью ионизированы уже при рН = 4 (рН внутриклеточной
среды близко к нейтральному значению). Именно поэтому нуклеиновые кислоты
склонны к взаимодействию с полиаминами, у которых между атомами азота
содержатся две или три метиленовые группы. Большой интерес вызывает
кислотно-основное взаимодействие нуклеиновых кислот с белками, в результате
которого, собственно и формируются сложные белки - нуклеопротеины. Особенно
эффективными оказываются взаимодействия нуклеиновых кислот с белками, имеющими
оснóвный
характер (рΙ > 8), т.к. в
биосредах суммарный заряд этих белков принимает положительные значения. Как уже
отмечалось ранее, ДНК образует комплексы с гистонами, входящими в состав хромосом.
Гистоны электростатически взаимодействуют с отрицательно заряженными фосфатными
группами, расположенными по периферии двойной спирали ДНК, и образуют
достаточно прочный комплекс, в котором ДНК оказывается стабилизированой
дополнительно. Вследствие изменения ионного состава среды, в которой находится
данный комплекс, может происходить дестабилизация ДНК, чем и определяется
регуляторная роль гистонов в функционировании генома.
Надо отметить, что кислотно-основные свойства
нуклеиновых кислот обусловлены не только наличием в них фосфатных групп, но и
присутствием236 азотистых оснований. Кислотно-основные свойства
гетероциклических оснований влияют главным образом на состояние и прочность
водородных связей и стэкинг-взаимодействий, стабилизирующих вторичную структуру
ДНК.
3.2 Хелатирующая способность
В водных растворах нуклеиновые кислоты проявляют
свойства активных полидентатных лигандов. Полидентатность нуклеиновых кислот
обусловлена наличием ионизированных фосфатных групп и полярных групп азотистых оснований
(карбонильных, имино- и др.), способных к образованию координационных связей с
катионами металлов. С помощью ионизированных фосфатных групп нуклеиновые
кислоты хелатируют катионы щелочных и щелочноземельных металлов, причем с
катионами щелочных металлов нуклеиновые кислоты образуют лабильные, а с
катионами щелочноземельных металлов (Mg2+,Ca2+) - более прочные комплексы. За
счет взаимодействия с полярными группами азотистых оснований нуклеиновые
кислоты образуют достаточно стабильные комплексы с катионами d-металлов.
.3 Денатурация
Все внешние факторы, которые приводят к
ослаблению или нарушению водородных связей или стэкинг-взаимодействий, вызывают
денатурацию нуклеиновых кислот. При этом происходит нарушение вторичной и
третичной структуры нуклеиновых кислот, но сохраняется первичная структура
молекул (рис. 4).
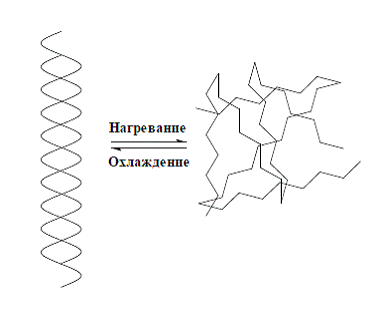
Рис. 4 Изменение вторичной структуры ДНК в
процессе денатурации
Факторы, вызывающие денатурацию нуклеиновых
кислот абсолютно те же, что и факторы, приводящие к денатурации белков, но
интенсивность денатурирующего действия конкретного фактора в случае нуклеиновых
кислот может быть иной. При денатурации ДНК ее двойная спираль полностью или
частично разделяется (раскручивается) на составляющие цепи.
Контроль за процессом денатурации ДНК можно
осуществлять с использованием ряда методов. Чаще всего измеряют поглощение в
УФ-области спектра. Интенсивность поглощения света пуриновыми и пиримидиновыми
основаниями зависит от того, присутствует ли оно в свободном состоянии или
входит в состав полинуклеотидов. При образовании двухспиральной структуры ДНК
интенсивность поглощение света каждым основанием уменьшается (гипохромный
эффект). Разрушение двухцепочечной спирали ДНК при денатурации вызывает
увеличение интенсивности поглощения УФ-света пуриновыми и пиримидиновыми
основаниями (гиперхромный эффект). Если относительное поглощение при 260 нм
отложить по оси ординат, а температуру (как фактор денатурации) - по оси
абсцисс, то получается S-образная кривая (рис. 5).
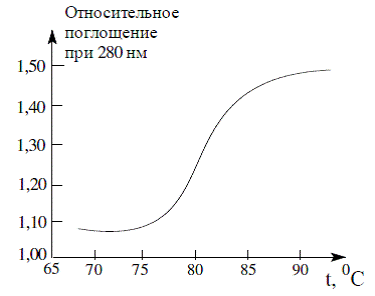
Рис. 5 График денатурации (плавления) ДНК
Она показывает, что при нагревании ДНК ведет
себя подобно кристаллам: двухцепочечная молекула «расплетается» на составляющие
ее цепи в пределах небольшого температурного интервала. Поэтому денатурацию ДНК
нередко называют плавлением, а температуру, при которой ДНК денатурирована на
50%, - температурой плавления - Тпл.
Нужно отметить, что Тпл зависит от
относительного содержания пар гуанин - цитозин и аденин - тимин, поскольку первая
пара более прочная. S-образные профили плавления показывают, что денатурация
ДНК - процесс кооперативный, т.е. каждое предшествующее изменение повышает
вероятность последующего. Так, если в определенном месте спирали происходит
нарушение водородных связей, то это приводит к нарушению
стэкинг-взаимодействий, что облегчает разрыв последующих водородных связей и т.
д.
3.4 Молекулярная гибридизация
Так как при денатурации нуклеиновых кислот их
первичная структура сохраняется, то данный процесс может иметь обратимый
характер. На способности нуклеиновых кислот восстанавливать свою нативную
конформацию после денатурации (ренативация) основан чрезвычайно важный метод
определения степени гомологичности, или родственности нуклеиновых кислот. Этот
метод называют молекулярной гибридизацией. Сущность этого метода сводится к
следующему: сначала смешиваются растворы ДНК, выделенных их организмов разного
вида (например, лягушки и кролика); затем эти растворы нагревают (для
денатурации ДНК), а потом охлаждают. При этом возникают двухспиральные
структуры разного состава: наряду с двухспиральными молекулами, идентичными
исходным молекулам ДНК, могут образовываться гибридные молекулы, содержащие
одну полинуклеотидную цепь из ДНК лягушки, а другую - из ДНК кролика. Такие гибридные
молекулы не совершенны: спирализованные участки в них чередуются с
неспирализованными, в которых полинуклеотидные цепи не комплементарны друг
другу. Изучение гибридизации ДНК - ДНК позволило биологам сделать вывод, что
первичная структура ДНК характеризуется видовой специфичностью. Сходным образом
может происходить гибридизация ДНК - РНК: в этом случае гибридная молекула
содержит одну дезоксирибонуклеотидную цепь и одну рибонуклеотидную.
При гибридизации ДНК и РНК, выделенных их одного
и того же организма, образуются совершенные гибриды. Иначе говоря, вся РНК
организма комплементарна ДНК того же организма. Это означает, что все
соображения относительно видовой специфичности ДНК в равной степени применимы и
к РНК.
3.5 Жидкокристаллическое
состояние нуклеиновых кислот
Для нуклеиновых кислот характерна определенная
ориентационно-пространственная организация нуклеотидов, каждый из которых
анизотропен, причем, при образовании комплементарной пары азотистых оснований в
полинуклеотидной цепи анизотропные свойства усиливаются. Следовательно, при
описании свойств двойной спирали ДНК роль анизотропии становится весьма
существенным фактором. Для крупных молекул ДНК, молекулярная масса которых
достигает 109 Да, вполне естественно, что в растворе отдельные фрагменты
молекулы могут находится в жидкокристаллическом состоянии. Число таких
фрагментов и их ориентация в пространстве сильно влияют на состояние ДНК в
клетке и на ее биологические функции. Кроме того, различные
жидкокристаллические состояния могут формироваться в системах полинуклеотиды -
вода или нуклеопротеины - вода. В таких системах могут происходить
множественные переходы из одного жидкокристаллического состояния в другое,
которые будут изменять биологические функции системы в целом под действием направленного
поля или самопроизвольно. В настоящее время установлено, что
жидкокристаллические состояния нуклеиновых кислот или их молекулярных
комплексов с белками играют важную роль в процессах передачи наследственной
информации и биосинтеза новых нуклеиновых кислот и белков на молекулярном
уровне.
4. Биологические функции нуклеиновых кислот
Нуклеиновые кислоты, прежде всего ДНК, являются
материальными носителями наследственной информации и определяют видовую
специфичность организма, сложившуюся в ходе биологической эволюции. Важно
уяснить, что именно азотистые основания - пуриновые или пиримидиновые, являются
носителями наследственной (генетической) информации, подобно тому, как боковые
заместители аминокислот определяют химические и функциональные свойства
аминокислоты. Сочетания трех рядом стоящих нуклеотидов в цепи ДНК называются
триплетами оснований или кодонами. Все кодоны ДНК составляют генетический код.
Молекула ДНК организованна в клетке в структурные единицы - гены. Гены, в свою
очередь, локализованы в хромосомах, которые находятся в ядре животных или
растительных клеток. Именно ген содержит информацию, определяющую
фенотипический признак организма: цвет глаз и волос, рост, пол и др. Все
количество ДНК, содержащееся в клетке, называется геномом. Наука, объектом
исследования которой является совокупность всей наследственной информации
организма, т.е. наука о геноме, называется геномикой и в последнее время
претерпевает бурное развитие.
Количество генов в геноме человека ранее
оценивалось в 60-80 тысяч. По последним данным, их меньше - 25-35 тысяч. Причем
большинство генов из генома «молчит» (у человека примерно 4/5 от общего
количества генов не активны), т.е. не кодирует молекулярные процессы, жизненно
необходимые для клетки и организма в целом. Генетическая информация передается
от родительской клетки к дочерней путем репликации - точного воспроизведения
ДНК in vivo. Генетическая информация, заложенная в ДНК, в процессе транскрипции
«переписывается» в полинуклеотидную последовательность мРНК. Матричная РНК, в
свою очередь, взаимодействует с соответствующими специфическими аминоацил-тРНК,
в результате чего происходит последовательное присоединение аминокислот.
Перевод генетической информации из РНК в специфическую аминокислотную
последовательность белка называется трансляцией.
Схематично процесс передачи генетической
информации можно представить следующим образом
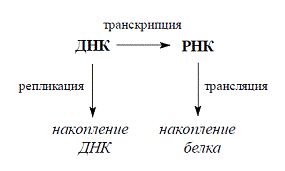
4.1 Процесс
репликации
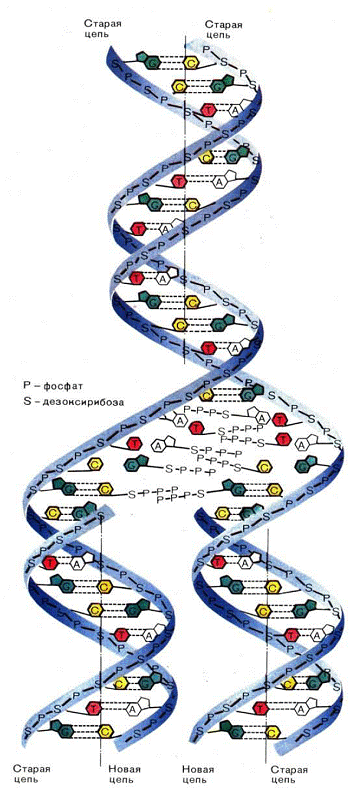
Рис. 6 - Репликация ДНК
Репликация (от позднелат. replicatio -
повторение) - самовоспроизведение нуклеиновых кислот (обычно ДНК, у некоторых
вирусов РНК), обеспечивающее точное копирование генетической информации и передачу
ее от поколения к поколению. При репликации ДНК нуклеотидная последовательность
копируется (целиком или частично) в виде комплементарной последовательности
дезоксирибонуклеотидов.
В процессе репликации двойная спираль ДНК,
состоящая из двух комплементарных полинуклеотидных цепей, раскручивается на
отдельные цепи и одновременно начинается синтез новых полинуклеотидных цепей;
при этом исходные цепи ДНК играют роль матриц. Новая цепь, синтезирующаяся на
каждой из исходных цепей, идентична др. исходной цепи. Когда процесс
завершается, образуются две идентичные двойные спирали, каждая из которых
состоит из одной старой (исходной) и одной новой цепи (рис. 6). Таким образом,
от одного поколения к другому передается только одна из двух цепей,
составляющих исходную молекулу ДНК, так называемый полуконсервативный механизм
репликации.
4.2 Процесс транскрипции
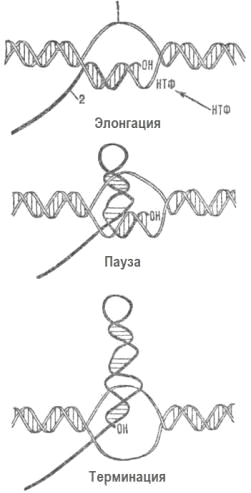
Транскрипция (от лат. transcriptio -
переписывание), биосинтез РНК на матрице ДНК; первая стадия реализации генетической
информации, в ходе которой нуклеотидная последовательность ДНК считывается в
виде нуклеотидной последовательности РНК. В основе этого процесса лежит принцип
комплементарного спаривания пуриновых и пиримидиновых оснований.
Транскрипция осуществляется с участием фермента
РНК-полимеразы, использующей в качестве субстратов рибонуклеозидтрифосфаты.
Кроме того, в транскрипции участвует большое число вспомогательных белков,
регулирующих работу РНК-полимеразы.
Транскрипция происходит на участках ДНК, которые
называют единицами транскрипции или трапскриптонами.
В начале и конце транскрилтона расположены
специфические нуклеотидные последовательности - промотор и терминатор.
Существование множества транскриптонов обеспечивает возможность независимого
считывания разных генов, их индивидуального включения и выключения. У животных,
растений и др. эукариот в состав транскриптона, как правило, входит один ген.
Транскриптоны бактерий обычно называют оперонами; множество из них содержат по
несколько генов, обычно функционально связанных (например, кодирующих несколько
ферментов, участвующих в синтезе той или иной аминокислоты).
Процесс синтеза РНК можно разделить на четыре
основные стадии: 1) связывание РНК-полимеразы с промотором, 2) начало синтеза
цепи РНК (инициация), 3) рост цепи РНК (элонгация), 4) завершение синтеза цепи
РНК (терминация).
4.3 Процесс трансляции
Трансляция (от лат. translatio-передача) -
программируемый генами процесс синтеза белка. Посредством трансляции
осуществляется реализация генетической информации нуклеиновых кислот. По
современным представлениям, исходный ген в виде ДНК непосредственно
транслироваться не может. Для трансляции он должен быть сперва
транскрибирован-переписан в форме молекул матричных рибонуклеиновых кислот
(мРНК) и именно последняя связывается с рибосомой и транслируется. В ходе
трансляции последовательность нуклеотидов мРНК определяет последовательность
остатков аминокислот в синтезируемом полипептиде.Происходит как бы перевод с
языка нуклеотидной последовательности гена на язык аминокислотной
последовательности белка. Считается, что аминокислотная последовательность
синтезируемого на рибосоме полипептида однозначно задает пространственную
структуру белка через процесс сворачивания (фолдинга), идущего одновременно с
трансляцией (котрансляционное сворачивание) или по ее завершении
(посттрансляционное сворачивание). Кроме того, в формировании конечной
структуры функционального белка могут принимать участие процессы ко- и
посттрансляционных модификаций. Трансляция каждой молекулы мРНК рибосомой
разделяется на три четкие последовательные стадии - инициацию, элонгацию
полипептида (собственно трансляция) и терминацию (Рис. 7).
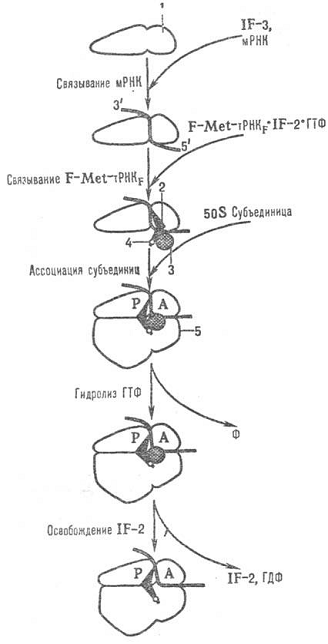
Рис. 7 - Схема последовательности событий в
процессе инициации трансляции у прокариот: 1-малая субъединица рибосомы;
2-инициаторная тРНК; 3-фактор инициации IF-2; 4-N-формилметионил (сокр. F-Met);
5-большая субъединица рибосомы; P и А-тРНК-связывающие участки рибосомы,
F-фосфат
Заключение
Несколько десятилетий тому назад биологическая
роль нуклеиновых кислот была совершенно неясна, в настоящее же время
установлено их первостепенное значение в живой природе.
Значение нуклеиновых кислот очень велико.
Особенности их химического строения обеспечивают возможность переноса в цитоплазму
и передачи по наследству дочерним клеткам информации о структуре белковых
молекул, которые синтезируются в каждой клетке. Белки обусловливают большинство
свойств и признаков клеток. Из этого следует, что стабильность структуры
нуклеиновых кислот - важнейшее условие нормальной жизнедеятельности клеток и
организма в целом. Любые изменения строения нуклеиновых кислот влекут за собой
изменения структуры клеток или активности физиологических процессов в них,
влияя, таким образом, на жизнеспособность.
Исследование нуклеиновых кислот и
нуклеопротеинов позволяет понимать механизмы возникновения инфекционных
заболеваний, поскольку по химической природе функциональная часть любого вируса
есть не что иное, как нуклеопротеин. Поэтому борьба с многочисленными вирусными
заболеваниями невозможна без глубокого познания строения и свойств нуклеиновых
кислот. И, наконец, сами мономерные звенья нуклеиновых кислот - нуклеотиды
играют важную самостоятельную роль в метаболизме: некоторые из них -
коферменты, другие - аккумуляторы энергии в клетке, третьи - циклические
нуклеотиды - регуляторы обмена веществ.
Используемая литература
1.
М. С. Гиляров, А. А. Бабаев, Г. Г. Винберг, Г. А. Заварзин и др. «Биологический
энциклопедический словарь»;
2. В. Зенгер «Принципы структурной организации
нуклеиновых кислот» М., 1987;
. Н. С. Курбатова, Е. А. Козлова "Конспект
лекций по общей биологии";
. З. А. Шабарова, А. А.Богданов, «Химия
нуклеиновых кислот и их компонентов», 1978;
. А. П. Горкин «Биология. Современная
иллюстрированная энциклопедия», М.: Росмэн, 2006;
6.
Д. Г. Кнорре «Биохимия нуклеиновых кислот», 1996