Морфологічні властивості мікроорганізмів
ЗМІСТ
ВСТУП
РОЗДІЛ І. Огляд літератури
1.1 Концепції виду в
бактеріології
1.2 Особливості визначення
систематичного положення мікроорганізмів
.3 Значення морфологічних
властивостей в систематиці мікроорганізмів
.4 Механізм ідентифікації
мікроорганізмів на основі морфологічних ознак
1.5 Обґрунтування обраного
напрямку роботи
РОЗДІЛ ІІ. Матеріали та методи
дослідження
2.1 Характеристика матеріалів
дослідження
2.2 Методи дослідження
2.3 Статистична обробка
результатів дослідження
РОЗДІЛ ІІІ Результати власних
досліджень
РОЗДІЛ IV. Аналіз результатів
досліджень
ВИСНОВКИ
ПРАКТИЧНІ РЕКОМЕНДАЦІЇ
СПИСОК ВИКОРИСТАНИХ ДЖЕРЕЛ
ДОДАТКИ
Додаток А
Додаток Б
Додаток В
Додаток Д
Додаток Е
Додаток Ж
Додаток З
Додаток И
ВСТУП
Актуальність теми. Залежно від
характеру трудової діяльності, стану здоров'я, індивідуального способу життя
час перебування у приміщенні коли-вається від 40 до 100%, а середній показник
становить - 75-80%. Вдихаючи повітря, ми контактуємо з великою кількістю
бактерій, при цьому 99,8% мікробів, які містяться в повітрі, затримуються в
дихальних шляхах [5]. Одні з них мирно співіснують, інші приносять користь,
треті - спричинюють захворювання. Відома також група умовно-патогенних
мікроорганізмів, які при порушення гігієнічних норм перетворюються на паразитів.
Тобто, повітряне середовище при значній зміні складу мікрофлори може стати
причиною різноманітних захворювань людини та джерелом забруднень харчових
продуктів [6]. Тому не випадково, об’єктом дослідження стала мікрофлора повітря
закритих приміщень.
Але визначення рівня бактеріального
забруднення потребує, перш за все, виявлення кількісного та якісного складу
мікроорганізмів [4]. Будь-які ознаки бактерії є важливими під час
ідентифікації. Проте деякі з них мають першочергове значення [22]. Вивчення
форми розмірів, взаємного розташування клітин, характеру розташування
джгутиків, наявності капсул та включень дозволяє скласти уявлення про
антигенні, патогенні та фізіолого - біохімічні, цитохімічні властивості
мікроорганізмів [9]. Таким чином, морфологія бактерій має велике таксономічне
значення, оскільки є важливим критерієм під час ідентифікації бактерій [17].
Метою дослідження є виявлення
взаємозв'язку між морфологічними властивостями та ідентифікацією сапрофітних
мікроорганізмів, що конкретизується у наступних поставлених завданнях:
1) виявити проблемні аспекти
тлумачення поняття “виду” в бактеріології;
) розглянути морфологічні
властивості мікроорганізмів;
) встановити зв'язок між
морфологічними властивостями та ідентифікацією бактерій;
) ідентифікувати бактерії
мікрофолори повітря;
) розглянути кількісні та якісні
закономірності формування мікрофлори повітря;
) порівняти результати досліджень із
санітарно-гігієнічними нормами.
Об'єкт
дослідження: морфологічні властивості мікроорганізмів.
Предметом дослідження є мікрофлора
повітря приміщень кафедри біології та методики її викладання
природничо-географічного факультету.
Методи дослідження: седиментаційний
метод Коха, фарбування за Грамом, статистичний; культивація мікроорганізмів
здійснювалася на щільному поживному середовищі, колонії рахували механічно.
Наукова новизна: здійснено аналіз
кількісного та якісного складу мікрофлори приміщень кафедри біології та
методики її викладання, обґрунтовано особливості ідентифікації мікроорганізмів
на основі морфологічних ознак.
Теоретичне значення: розглянуто
концепції виду в бактеріології та виділено проблеми, які виникають перед
мікробіологами при визначення виду мікроорганізмів як основної систематичної
категорії. Розширюються та поглиблюються наукові знання з морфологічних
властивостей мікроорганізмів, їх роль під час визначення роду бактерій,
виділено загальні закономірності механізму ідентифікації на основі морфології
мікроорганізмів.
Практична цінність полягає у тому,
що поширення знань з цього питання дозволить відповідальніше ставитися до свого
організму, адже допоможе усвідомити необхідність дотримання гігієнічних норм.
Обізнаність в санітарно-гігієнічному плані сприяє зменшенню загального числа
мікробів, з якими ми контактуємо щодня. Також, результати дослідження підтвердять
значення морфологічних властивостей в ідентифікації мікроорганізмів.
РОЗДІЛ І.
Огляд літератури
1.1 Концепції виду в бактеріології
В бактеріології вид як основна
систематична категорія залишається не визначеним. Взагалі, до одних найважчих завдань
перед якими поставлені нині мікробіологи можна з упевненістю віднести
визначення виду мікроорганізмів. Варто зазначити, що добитися єдиної думки
вчених у галузі таксономії в цілому та номенклатури бактерій зокрема - справа
надзвичайно складна.
По-перше, проблема полягає в тому,
що якщо рід бактерій в більшості випадків чітко окреслений, то найменший таксон
- вид - іноді дуже важко розрізнити не лише за морфологічними ознаками, але й
при врахуванні багатьох біохімічних властивостей та ретельному вивченні та
аналізі всіх особливостей штаму, який визначають [1]. Зрозуміло, що кожному
виду притаманна певна форма та розміри, але умови росту культури чинять значний
вплив на розміри бактеріальних клітин. Тому такий спрощений критерій, як
розмір, що застосовувався на зорі мікробіології, на даному етапі абсолютно
неприйнятний.
По-друге, труднощі, які виникають
при трактуванні поняття виду можна пояснити значною одноманітністю
морфологічних форм мікроорганізмів у порівнянні з еукаріотичними організмами
[16]. Адже якщо в основі визначення виду у рослин і тварин лежать чітко
виражені морфологічні та фізіологічні відмінності, зокрема неможливість
схрещуватись між представниками кількох популяцій, то у бактерій ці критерії
відсутні. Бактерії морфологічно не різноманітні, розмножуються нестатевим
шляхом, характеризуються наявністю плазмідної спадковості. Також, меншою мірою
у прокаріот проявляються цитологічні ознаки, виникають труднощі і при вивченні
їх філогенезу [5].
Розглянемо дану проблему на
конкретних прикладах. Бактерії можна поділити на основі їхньої здатності
викликати захворювання у людини, тварин, рослин. Формування особливих форм
патогенних та фітопатогенних бактерій відбувається лише за однією ознакою. Тому
не дивно, що з появою кожного нового визначника мікроорганізмів змінюється їх
групування. Мікроорганізми переміщаються з однієї таксономічної групи в іншу,
при цьому переміщають не тільки окремі види, але й роди, і навіть більш високі
систематичні одиниці. В одній і тій же групі організми розподіляють як за
морфологічними ознаками, так і за фізіологічними. А до однієї і тієї ж групи
можуть бути віднесені рухомі та нерухомі форми, як грампозитивні та
грамнегативні бактерії. Слід зауважити, що одна і та ж зовнішня морфологічна
ознака може мати різне значення. Наприклад, така суттєва для мікроорганізмів
ознака як взаєморозташування клітин зовсім не характерна для таких справ-жніх
бактерій як оцтовокислі бактерії, азотбактер та інші. Для мікобактерій
галуження нормальний формотворчий процес; для бактерій галуження - патологічний
явище, що спостерігається при дегенеративному переродженні [24].
Щоб підійти до визначення виду в
бактеріології, розглянемо історію розвитку цього поняття в біології взагалі.
Стихійне уявлення про види живих істот склалось ще в давні часи. З розвитком
біології як науки в додарвіністський період з'явилися дві основні концепції -
номіналістична і типологічна. Номіналісти вважають, що види реально не існують.
Типологічну концепцію можна вважати морфологічною, оскільки в даному разі
критерієм виду є морфологічна подібність. У 40-х роках ХХ ст. завдяки роботам
Ернста Майра сформувалась нова концепція виду - біологічна. Вона базується на
тому, що види складаються з популяцій, вони реальні, їм притаманна внутрішня
генетична інтегрованість. Згідно з біологічною концепцією вид - це група
організмів, здатних до схрещування, які утворюють природні популяції і які
репродуктивно ізольовані від інших груп [16].
До теперішнього часу відсутні єдині
принципи і підходи до об'єднання (або поділу) їх в різні таксономічні одиниці.
Слід зауважити, що в класифікації бактерій існує два основних принципи. Перший
з них - каталогізація форм, яка проводиться на основі однієї чи кількох ознак,
часто випадкових та не пов'язаних між собою. Другий принцип заснований на побудові
системи на основі філогенетичних даних, отриманих шляхом комплексного,
всебічного вивчення організмів. Дуже багато мікроорганізмів мають однакові
морфологічні ознаки, але розрізняються за будовою геномів, родинні зв'язки між
ними часто бувають неясними, а еволюція багатьох просто невідома. Більш того,
наріжне для кожної класифікації поняття виду бактерій до цього часу не має
чіткого визначення, а в ряді випадків справжня спорідненість між бактеріями
може виявитися спірною, оскільки вона лише відображає спільність походження від
одного далекого предка [11]. Серед мікробіологів поширена думка, що всі види
бактерій реально не існують. Це пов'язано з браком інформації про реальне
існування бактерій у природі, достатньо великою фізіологічною різноманітністю
ізолятів, які виділяють з природи [16].
Нині існує безліч тлумачень виду
бактерій. Розглянемо декілька визначень, які повною мірою розкривають суть
терміну. Для визначення виду бактерій цілком придатними є критерії, що
характеризують вид еукаріотів. Тобто, вид у бактерій - це група організмів, які
мають схожий набір і порядок генів на хромосомі та здатні (хоча б потенційно)
обмінюватися хромосомними генами і здійснювати їх гомологічну рекомбінацію з
великою частотою. Проте таке трактування виду довго залишалось не узаконеним
[16].
Як ще в минулому столітті зазначив
Красильников М. А. (1949), вид у бактерій - це сукупність організмів,
пов'язаних між собою родинними зв'язками. Специфіка тлумачення виду бактерій
Красильниковим, полягає у тому, що мікроорганізми повинні мати спільне
походження і на конкретному етапі еволюційного розвитку характеризуватися
сталими морфологічними ознаками. Також, Красильников стверджує, що вид бактерій
має бути у більший чи меншій мірі відокремленим еволюційним відбором та пристосованим
до певного середовища та зовнішніх умов існування [11].
Заварзін (1974) трактує поняття
таксону і виду бактерії так : “Таксон- група організмів, яка має задану ступінь
однорідності”. На думку дослідника, вид як основна таксономічна категорія
визначається на основі морфологічних, культуральних, фізіологічних,
біохімічних, антигенних та інших властивостей [16].
Дж. Стенлі та Н. Кріг у вступі до
дев'ятого видання Керівництва Бергі з систематики бактерій розглядають вид у
бактерій як колекцію штамів бактерій, які характеризуються сукупністю багатьох
спільних властивостей і відрізняються від інших штамів. Але вони зазначають, що
для чіткішого визначення виду може бути використаний рівень гомології ДНК -
показник генетичної спорідненості організму. Вид повинен включати штами з
рівнем подібності ДНК-ДНК 70% і вище [15].
На думку Герхардта (1984), в
бактеріології під видом розуміють типовий штам та всі інші штами, які вважають
достатньо подібними з типовим штамом, для того щоб належати до даного виду.
Типовий штам - це штам, який обрали в якості постійного зразка того, що
розуміють під даним видом, хоча він і не завжди є найбільш типовим
представником даного виду. Всі решта штамів, котрі можуть бути віднесеними до
даного виду, потрібно порівнювати з типовим штамом даного виду [7].
Вираз “достатньо подібними” з
наведеного вище визначення виду бактерій є джерелом більшості проблем в
класифікації бактерій. Проте очевидно: чим більше відомостей про якусь групу
бактерій, тим більш вірогідно що різноманітні дослідження допоможуть прийти до
однієї прийнятної схеми класифікації.
Використання молекулярно-біологічних
методів у мікробіології внесло певні корективи до уявлень мікробіологів щодо
суті виду бактерій. У 2002 р. відбулося засідання Спеціального міжнародного
комітету з систематики прокаріот. Комітетом процитовано визначення виду,
наведене Р. Розелло-Мора й Р. Аман (2001 р.) : “Вид - це категорія, яка обмежує
генотипово пов'язану групу штамів, що мають високий ступінь подібності за
незалежними ознаками, визначеними за певних стандартних умов” [16].
За сучасними даними, вид бактерії
розглядають як сукупність популяцій, що мають такі властивості: 1) спільне
походження; 2) пристосованість до певного середовища життя; 3) подібність
обміну речовин та характеру міжвидових відношень; 4) наявність подібного
генетичного апарату, морфологічних та фізіологічних ознак. Якщо при вивченні
виділених бактерій виявляють відхилення від типових видових властивостей, то
таку культуру розглядають як підвид [19].
Отже, існує декілька понять,
стосовно визначення виду. Дати одне чітке визначення виду бактерій важко у
зв'язку з біологічними особливостями цих мікроорганізмів. Види можуть бути
представлені багатьма підвидами, які можна розглядати як етап формування виду.
Але незалежно від різних варіацій тлумачення поняття виду, без сумніву, можна
виділити спільні риси даних визначень. По-перше, вид вважається найменшою
таксономічною одиницею, яка реально існує в природі. А всі інші таксономічні
категорії за винятком виду, штучно створені систематиками й описуються за
допомогою низки ознак. І, власне чи не найважливішими такими ознаками є саме
морфологічні. Також слід згадати і про цитологічні, фізіолого-біохімічні,
імунологічні тощо. По-друге, вид - являє собою продукт еволюції живої матерії, має
свою історію розвитку, формування і стабілізації в результаті пристосувань до
умов існування.
Існує також інфравидовий поділ, що
ґрунтується на відмінності за якимись незначними спадковими властивостями:
антигенними - серовар, морфологічними - морфовар, патогенністю - патовар [19].
Зрозуміло, що від ліннеївського
розуміння виду як стабільної одиниці до розуміння виду мікроорганізмів
сучасними біологами як найменшої реально існуючої категорії, яка розвивається,
пройшло багато часу. Вважають, що повне наукове обґрунтування виду у бактерій
можна буде дати тоді, коли буде вивчена їхня еволюція. Хоча в мікробіології
проблема виду вважається найбільш важкою, проте важливою, оскільки без
визначення виду, не можна побудувати класифікацію мікроорганізмів [7].
1.2 Особливості
визначення систематичного положення мікроорганізмів
Для визначення виду мікроорганізмів
спочатку систематизують основні його ознаки (морфологія, рухливість,
спороутворення, біохімічні та інші властивості), а потім за ними ідентифікують
мікроорганізм і знаходять за визначником його місце в класифікації бактерій.
Виявлення прокаріот, які морфологічно менш різноманітні ніж еукаріоти,
базується на використанні широкого спектру фенотипічних, а в багатьох випадках
генотипічних ознак. В даному випадку ідентифікація в більшій мірі, ніж
ідентифікація еукаріот, заснована на функціональних критеріях, тому, що
більшість бактерій можна виявити не за їхнім зовнішнім виглядом, а лише
визначивши, які процеси вони здатні здійснювати [19].
Саме тому, щоб
віднести мікрооганізми до тієї чи іншої таксономічної групи, необхідно
достатньою мірою володіти інформацією про їхні основні характеристики:
визначати форму мікроорганізмів, наявність джгутиків, капсул, здатність до
спороутворення, забарвлення за Грамом (див. Додаток А, Б, В, Д, Е.1) І не можна
обмежуватися будь-яким одним морфологічним критерієм при встановленні виду.
Лише за сукупністю ознак - морфологічних, культуральних, цитохімічних,
фізіолого-біохімічних можна охарактеризувати вид . Тобто, вид у бактерій визначається
сумою різноманітних ознак і властивостей [1].
1.3
Значення морфологічних властивостей в систематиці мікроорганізмів
Сукупність морфолого-культуральних
ознак були покладені в основу розподілу бактерій на групи різного рангу, тобто
вони стали критерієм класифікації бактерій.
Ще Антоні ван Левенгук описав всі
основні морфологічні форми бактерій - кокоподібні, паличкоподібні, вигнуті та
ін. [16].
Згідно з класифікацією К. Лемана та
Р. Неймана (1896), усі мікроорганізми за морфологією було поділено на три
родини: Coccaceae, Bacteriaceae, Spirilaceae. На основі морфологічних ознак
систему класифікації бактерій створили А. Клюйвер, К. Ван Ніль і Р. Стейнер
(1936, 1941) [19]. У міру нагромадження нових наукових даних про мікроорганізми
ця класифікація зазнала істотних змін і була значно доповнена, проте і в
зміненому вигляді вона не могла задовольнити мікробіологів, тому створювались
більш досконалі класифікації [22].
В основі сучасних уявлень про
морфологію бактерій лежать дослідження німецького фізіолога рослин Ф. Кона. Він
відніс бактерії до окремої групи організмів з характерними морфологічними
ознаками.
Морфологічні властивості бактерій є
одним із критеріїв класифікації бактерій за Д. Х. Берджі. Ще в 1923 р. він
склав перший міжнародний визначник бактерій. . А 1984 р. вийшло в світ нове
видання систематики бактерій Bergey's Manual of Systematic Bacteriology, у
підготовці якого брали участь 124 вчених із 14 країн світу. У ньому більш
детально розглянуто відомості про мікроорганізми [19].
За Берджі, усі бактерії поділені на
чотири категорії:
І. Грамнегативні бактерії, що мають
клітинну стінку;
ІІ. Грампозитивні бактерії, що мають
клітинну стінку;
ІІІ. Еубактерії, що позбавлені
клітинної стінки;
ІV. Архебактерії.
До першої категорії віднесено
прокаріоти зі складною (грамнегативного типу) будовою клітинної стінки. Дані
бактерії можуть бути сферичної або овальної форми, у вигляді прямих або
зігнутих паличок, спіралей, ниток. Деякі з них одягнені в чохол чи капсулу.
Міксобактерії можуть утворювати плодові тіла і міксоспори. Є рухливі
(“плавають”, “ковзають”) або нездатні до руху. Серед представників цієї
категорії є види патогенні для людини, тварин і рослин.
У другу категорію об'єднують
прокаріоти, що мають клітинну стінку грампозитивного типу. Фарбування за Грамом,
не завжди є позитивним. Клітини сферичної, паличкоподібної або ниткоподібної
форми. Палички і нитки здебільшого не галузяться, але у деяких організмів
спостерігається справжнє галуження. До цієї категорії відносять аспорогенні і
спороутворюючі бактерії, а також актиноміцети і споріднені організми
До третьої категорії відносять
прокаріоти, що позбавлені клітинної стінки. Їх називають мікоплазмами. Клітини
фарбуються за Грамом негативно. Дрібні форми проходять через бактеріальні
фільтри. Здебільшого - це нерухомі форми, хоча для деяких з них описаний
ковзний рух. У видів із спіралеподібною формою простежують обертовий і
поштовхоподібний рух. Серед мікоплазм трапляються патогенні форми. Останні
спричиняють хвороби у рослин, тварин, людини і здатні уражати культури тканин.
Архебактерії, які складають четверту
категорію, по-різному фарбуються за Грамом, оскільки клітинні стінки в них
сильно відрізняються. Форма клітин різноманітна: сферична, спіральна, пластична
і паличкоподібна. Трапляються поодинокі чи багатоклітинні форми у вигляді ниток
або агрегатів. Колір клітин може бути червоним, пурпуровим, рожевим,
оранжево-коричневим, жовтим, зеленим, темно-зеленим, сірим і білим [15].
Класифікація бактерій кожної
морфологічної групи наведена в додатку А.1.
Слід зауважити, що різноманітність
морфологічних властивостей бактерій, зокрема роду Clostridium показує, що
розміри і форма клітин не можуть бути достатніми критеріями для класифікації
прокаріот на рівні роду [ 22].
1.4
Механізм ідентифікації мікроорганізмів на основі морфологічних ознак
Вивчення морфології мікроорганізмів
лежить в основі мікроскопічно-го методу діагностики. Цього часом достатньо для
ідентифікації мікроорганізмів.
Ідентифікація - це порівняння
невідомих організмів з уже прокласифікованими бактеріями з метою встановлення
ідентичності чи найменування невідомих організмів [7].
У мікробіологічних дослідженнях, на
жаль, використовується порівняно невелика кількість стандартних методів і дуже
часто для ідентифікації пропонуються ознаки, виявлені специфічними методами.
Але ідентифікація повинна базуватись на простих, доступних для дослідника,
методах [16].
Як правило, виділений від хворого чи
із навколишнього середовища мікроорганізм не можна швидко і точно
ідентифікувати. З цією метою необхідно використовувати безліч фено- та
генотипічних ознак. При цьому велике значення приділяється дискримінаційній
силі тесту - тобто здатності виявляти відмінності між різними видами чи
варіантами бактерій (дискретним одиницям інформації). Наприклад, забарвлення за
Грамом, форма бактерій, спороутворення [22].
Морфологію мікроорганізмів вивчають
за допомогою світлової та електронної мікроскопії. У лабораторній практиці
частіше проводять мікроскопію фіксованих забарвлених мазків і рідше нативних
препаратів у вигляді стисненої чи висячої крапель. Такі препарати можна
фарбувати лише наднизькими концентраціями барвників - від 0,0001% до 0,001%
[7]. Живі препарати використовуються для вивчення розмірів, форми, структури,
рухливості, характеру розмноження, відношення клітин до різноманітних
подразників (хімічних, фізичних і т.д.). Дослідження у живому стані крупніших
мікроорганізмів дає змогу вивчати тонку структуру їх клітин краще , ніж у
забарвлених препаратах , а на зафіксованих і забарвлених препаратах вивчають
морфологію мікроорганізмів. Незабарвлені мікроорганізми, за винятком грибів,
погано видно у світловому мікроскопі через їх малу контрастність [17].
Також, виділяють загальні
закономірності механізму визначення розмірів, форми клітин, виявлення спор та
джгутиків, здатності до спороутворення, способу руху клітин мікроорганізмі (
див. Додаток Ж) .
1.5
Обґрунтування обраного напрямку роботи
бактеріологія
мікроорганізм ідентифікація морфологічний
У навколишньому повітрі завжди
міститься певна кількість мікроорганізмів.
Незважаючи на загальновідомий факт,
що мікрофлора повітря - вторинна за складом і досить бідна за видами
мікроорганізмів, а дефіцит вологи, поживних речовин та сонячної радіації
перешкоджає розмноженню бактерій, велике гігієнічне та епідеміологічне значення
має оцінка забрудненості повітря приміщень. Оскільки, збудники респіраторних
інфекцій поширюються повітряно-крапельними шляхами. Також, мікробіологічний
аерозоль може стати причиною алергічних захворювань, особливо за наявності в
повітрі цвільових грибів і актиноміцетів.
У зв'язку з цим, мікробіологічне
дослідження повітря має першочергове значення для контролю стану навколишнього
середовища замкнених приміщень: операційних, асептичних палат, боксів, аптечних
закладів, бактеріологічних лабораторій.
Ідентифікація мікроорганізмів на
основі їх морфологічних та тинкторіальних ознак дозволяє визначити роди
бактерій. Тобто, вивчення морфології мікроорганізмів лежить в основі
мікроскопічного методу діагностики. Цього часом достатньо для ідентифікації
мікроорганізмів. Оскільки, саме морфологія визначає багато властивостей
мікроорганізмів, включаючи і патогенні.
Зрозуміло, що морфологічні
властивості не можуть бути єдиними критерієм під час ідентифікації. Особливо
актуальним нині є питання механізму резистентності бактерій до антибіотиків.
Наприклад, відомі випадки, коли після фарбування за Грамом неможливо було
відрізнити роди стрептококів від стафілококів спираючись лише на морфологічні
ознаки. Проте, вивчення форми розмірів, взаємного розташування клітин, характеру
розташування джгутиків, наявності капсул та включень дозволяє скласти уявлення
про антигенні, патогенні та фізіологічні властивості мікроорганізмів.
РОЗДІЛ ІІ.
Матеріали та методи дослідження
2.1 Характеристика матеріалів дослідження
Матеріалом дослідження є сапрофітна
мікрофлора повітря приміщень.
В атмосферному повітрі можуть
знаходитись десятки й сотні видів сапрофітних мікроорганізмів [14]. Проте,
якісний та кількісний склад мікрофлори повітря є випадковим [17].
Численні аналізи зразків
атмосферного повітря дали змогу ідентифікувати в ньому понад 1200 різних
мікроорганізмів. Серед них регулярно виявляють стафілококи, мікрококи, сарцини,
спороносні палички, актиноміцети, віруси [4]. Найпоширенішими є спорові
бактерії роду Bacillus
(B. subtilis, B. megaterium, B.
cereus), пігментовані сапрофітні бактерії роду Micrococcus, сарцини, дріжджі,
актиноміцети, плісневі гриби, тобто мікроорганізми стійкі до дії світла
(інсоляції) та висихання. З групи цвілевих грибів у повітрі є Mucor,
Asparagus, Pennicilium, Aspergillus, Risopus, а також дріжджові гриби -
Saccharomyces cereviseae, Candida, тощо [17].
У приміщеннях за наявності хворих
людей, тварин, у повітрі можуть міститися й патогенні види мікроорганізмів:
стафілококи, мікобактерії, бацили сибірки, бактерії туляремії, сальмонели,
збудники дифтерії, стрептококових, менінгококових і вірусних інфекцій [9].
Сапрофітні мікроорганізми, що є в
повітрі, можуть бути у трьох фазах бактеріального аерозолю - краплинній,
краплинно-ядерній і пиловій. Під аерозолем розуміють фізичну систему із дрібних
твердих або рідких часточок, що зависли в газовому середовищі.
2.2 Методи дослідження
В ході виконання курсової роботи
мною були використані наступні методи досліджень:
1. Седиментаційний метод Коха
Метод седиментації (Коха) базується
на осіданні мікроорганізмів, краплин вологи і пилинок під впливом сил тяжіння
на поверхню поживного середовища відкритої чашки Петрі.
Для аналізу були взяті проби повітря
з 6 приміщень, а саме: аудиторій № 21, № 18, коридорів на першому та другому
поверхах, живого куточку та туалету.
З цією метою, до початку занять та
після їх завершення було відкрито 12 чашок Петрі з МПА, які встановили на
горизонтальній поверхні в місці взяття проби. Експозиція чашок продовжувалася
15 хв. Потім посіви закритими, були поміщеними у термостат при 370 С на 24 год,
а потім залишеними на 48 год при кімнатній температурі [17].
2. Метод фарбування за Грамом в
модифікації Синьова
На зафіксований мазок досліджуваної
культури кладуть клаптик фільтрувального паперу і наносять на нього 2-3
краплини розчину карболового генціанвіолету. Фарбу витримують протягом 1-2
хвилин. Потім папірець знімають та діють на препарат двома краплями розчину
Люголя, і обробляють мазок етиловим спиртом протягом 30 сек. Після цього
препарат старанно промивають дистильованою водою, дофарбовано фуксином (1 хв),
знову промивають водою та висушують. Виготовлений препарат вивчала під
мікроскопом за допомогою імерсійної системи [4].
2.3 Статистична обробка результатів дослідження
Для кількісного обчислення
мікроорганізмів у повітрі проводились такі обчислення:
1) Визначення
площі поживного середовища з колоніями в чашці Петрі за формулою S = π ;
2) Перерахунок
кількості колоній, які утворилися на чашці, на площу 100 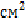 здійснювали
за наступною формулою:
здійснювали
за наступною формулою:
, де а - кількість колоній,
які утворились на чашці Петрі.
3) Відповідно
до даних В.Л. Омельського, на площу в 100 за 5 хв осідає стільки бактерій, скільки
їх міститься в 10 л повітря, тому перерахунок кількості мікроорганізмів в 1
повітря здійснювався за формулою:
, де а - кількості колоній,
які утворилися на чашці, на площу 100 [4].
РОЗДІЛ ІІІ.
Результати власних досліджень
Результати кількісного аналізу
Результати кількісного складу
мікрофлори приміщень кафедри біології та методики її викладання наведені в
таблиці 3.1. (обчислення наведені в Додатку З)
Кількісний склад мікрофлори
приміщень кафедри біології
Таблиця 3.1
|
Приміщення
|
Ауд. №21
|
Ауд. №18
|
Коридор на 2 -ому поверсі
|
Коридор на 1 - ому поверсі
|
Туалет
|
Живий куточок
|
|
Час забору проб
|
|
|
|
|
|
|
|
До занять
|
382
|
510
|
764
|
637
|
510
|
892
|
|
Після занять
|
1273
|
1146
|
1650
|
1415
|
4246
|
3057
|
Якісний аналіз мікрофлори повітря
Результати кількісного складу
мікрофлори приміщень кафедри біології та методики її викладання наведені в
таблиці 3.2.
Біорізноманіття мікрофлори приміщень
кафедри біології
Таблиця 3.2.
|
Мікро-організми
|
Коки
|
Палички
|
Незвична форма
|
|
|
Приміщення
|
Стрептококи
|
Монококи
|
Диплококи
|
Тетракоки
|
Сарцини
|
Бактерії
|
Диплобацили
|
Стрептобации
|
Клостридії
|
|
|
|
1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
|
|
Кор. 1-го поверху
|
+
|
-
|
+
|
-
|
+
|
+
|
-
|
+
|
-
|
+
|
|
|
Кор. 2-го поверху
|
+
|
+
|
-
|
-
|
+
|
+
|
+
|
-
|
+
|
+
|
|
Ауд. №21
|
+
|
+
|
-
|
-
|
+
|
-
|
-
|
-
|
+
|
-
|
|
Живий куточок
|
-
|
+
|
-
|
+
|
+
|
+
|
-
|
-
|
-
|
-
|
|
Туалет
|
+
|
+
|
+
|
+
|
+
|
+
|
+
|
-
|
-
|
+
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
РОЗДІЛ IV.
Аналіз результатів досліджень
Результати кількісного аналізу
Кількісний аналіз свідчить, що
найбільша концентрація мікроорганізмів наявна у туалеті (рис. 1), адже це
найбільш відвідуване місце, в якому не завжди дотримуються правил гігієни,
зберігається інвентар для прибирання. До приміщень з високою концентрацією
мікроорганізмів належить і живий куточок. Це обумовлено тим, що у повітрі
присутня велика чисельність мікрофлори організмів тварин, їх продукти
харчування містять епіфітну мікрофлору. Окрім вказаних факторів, на кількісний
склад мікрофлори повітря впливають сприятливі фізичні фактори. Зокрема,
відсутність прямих сонячних променів, сприятлива температура. Найменша
кількість мікроорганізмів виявлена в аудиторії №21, що може пояснюватись
регулярним провітрюванням, значною площею аудиторії та, можливо, найменшою
кількістю проведених пар в ній в день забору проб.
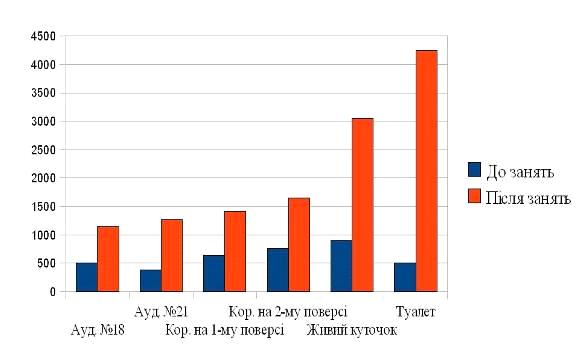
Рис. 1 Кількісний
розподіл мікроорганізмів до та після занять.
Привертає увагу, висока частка
грибів (40,65%) від загальної кількості колоній мікроорганізмів в повітрі
коридору на другому поверсі (див рис. Ж.12). Можна виділити декілька ймовірних
причин такого розподілу.
По - перше, велика чисельність
колоній грибів може бути обумовленою такою морфологічною ознакою як розміри
клітин. Адже маса міцелію грибів у порівнянні з іншими мікроорганізмами є
найбільшою, тому гриби швидше осідають. По - друге, гриби є індикаторами місць
з надлишковою вологістю. Так, показників вологості повітря для аудиторії № 21,
туалету, коридору на першому поверсі та кор. на другому поверсі складають
відповідно 51, 65, 71, та 74. Закономірним виявилось, що найбільша кількість
грибів була виявлена в приміщеннях коридорів. Таким чином, підтверджено
загальновідомий факт здатності грибів до розмноження в повітрі з надлишковою
вологістю та конденсованою вологістю на охолоджених поверхнях (стіни, скло,
відкоси вікон). Третьою ймовірною причиною високої частки грибів можуть
слугувати рослини, оскільки ґрунт є середовищем існування грибів.
Звертає увагу факт збільшення
кількості мікроорганізмів в повітрі аудиторій з кожним наступним поверхом, що
пояснюється зростанням температури в приміщенні. Значне зростання кількості
загального мікробного числа для мікрофлори повітря коридорів обумовлено тим, що
вони є місцем активного руху людей, які переносять значну кількість
мікроорганізмів на верхньому одязі.
Результати якісного аналізу
мікрофлори повітря
Закономірною виявилась переважання
пігментовані сапрофітні бактерії роду Micrococcus, сарцини, стрептококи,
бактерії роду Bacillus (рис. 2) . Це обумовлено тим, що пігменти захищають
мікроорганізми від надмірної сонячної інсоляції. Найменшим виявився вміст
диплобацил, стрептобацил (рис. 3).
Привертає увагу, незначне
біорізноманіття пігментоутворюючих мікроорганізмів живого куточку. В ході
ідентифікації тут не було виявлено стрептококів, диплококів, диплобацил,
стрептобацил (див. табл. 3.2., рис. 6), що є доказом поганої освітленості
даного приміщення.
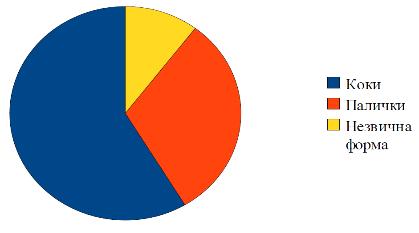
Рис. 2. Загальне співвідношення
мікроорганізмів: коки - 68 %; палички - 36 %; незвична форма - 12 %.
Рис. 3. Біорізноманітність
мікрофлори повітря у відсотках
Співвідношення сарцин, стрептококів,
монококів, диплококів та тетра-коків складає відповідно 29, 4 % : 23,5 % : 23,
5 % : 1,8 % : 11,8 % (рис. 4). Висока частка стрептококів є доказом
повітряно-крапельного забруднення повітря та індикатором місць довготривалого
перебування людей.
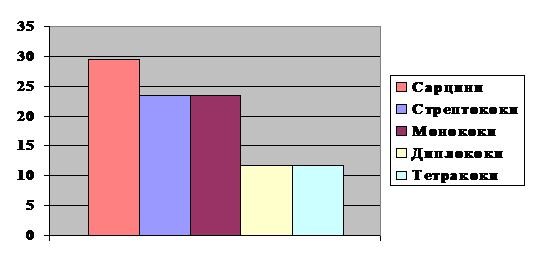
Рис. 4. Співвідношення коків
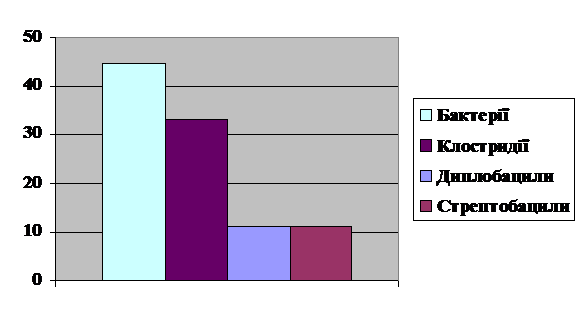
Рис. 5 Співвідношення паличок
Співвідношення бактерій, клостридій,
диплобацил, стрептобацил складає відповідно 44,5 % : 33, 3 % : 11, 1% : 11,1 %
(рис. 5). Присутність спороутворюючих паличок можна пояснити нерегулярним
провітрюванням та відсутністю вологого прибирання.
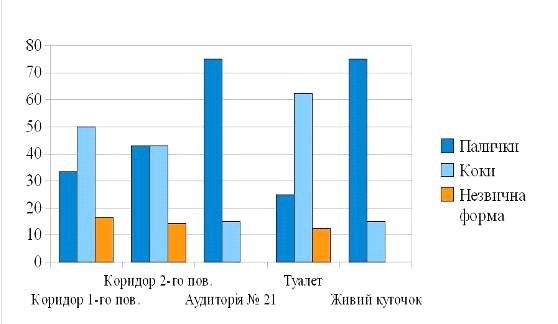
Рис. 6 .Біорізноманіття
мікрофлори приміщень кафедри біології
Таким чином, мікрофлора повітря є
вторинною та досить бідною за видовим складом. Насамперед, це пояснюється тим,
що на відміну від атмосферного повітря, у повітрі закритих приміщень не
відбувається процесів самоочищення.
Під час ідентифікації бактерій було
підтверджено загальновідомий факт, колій колоній не є ключовим при визначенні
видового складу мікрофлори (див. Додаток Е, рис. Е.2.1-Е.2.13). Морфологічні
ознаки їх клітин можуть бути однотипними. В ході проведеної роботи з
ідентифікації було встановлено, що більшість сапрофітних мікроорганізмів
мікрофлори повітря належить до грампозитивних бактерій.
ВИСНОВКИ
1. За
зростанням мікробного числа в 1 м3 повітря до занять досліджувані приміщення
можна розмістити у такому порядку: навчальні аудиторії ‹ туалет ‹ коридор І
поверху ‹ коридор ІI
поверху ‹ живий куточок, а після занять - навч.ауд ‹ коридор І поверху ‹
коридор ІІ пов. ‹ живий куточок ‹ туалет.
2. За
зростанням біорізноманітності мікроорганізмів в 1 м3 повітря досліджувані
приміщення можна розмістити у такому порядку: навч. аудиторія ‹ коридор І пов.
‹ коридор ІІ пов. ‹ живий куточок ‹ туалет.
3. Серед
мікрофлори повітря приміщень кафедри біології переважаючими є пігментовані
сапрофітні бактерії роду Micrococcus, сарцини. бактерії роду Bacillus .
Найменшим виявився вміст, клостридій та бактарій, мікроорганізмів неправильної
форми. Переважна більшість сапрофітних мікроорганізмів належить до
грампозитивних.
4. Кількість
мікроорганізмів в усіх досліджуваних приміщеннях за критерієм Шафіра
знаходиться в межах санітарних норм.
5. Труднощі
ідентифікації виду мікроорганізмів спричинені:
· меншою
різноманітністю морфологічних властивостей прокаріот у порівнянні з
еукаріотами;
· потрібною не
лише аналізу морфологічних властивостей, але й комплексу біохімічних,
детального всіх особливостей штаму;
· види можуть
бути представлені багатьма підвидами, які можна розглядати як певний етап
формування виду;
· здатністю
бактерій до плеоморфізму. Зокрема, резистентність бактерій під впливом
антибіотиків;
· впливом
умови росту культур на морфологічні властивості.
ПРАКТИЧНІ РЕКОМЕНДАЦІЇ
. Під час фарбування мікроорганізмів
необхідно зважати на те, що :
1.1 Краще фарбуються живі
фіксовані клітини, оскільки їхня клітинна стінка і цитоплазма лімітують
проникнення барвника;
1.2 Під час вітального
фарбування, з метою запобігання отримання артефактів, що з'являються в
результаті токсичної дії барвника на живі об'єкти потрібно використовувати
барвники у великому розведенні (1:10 000, 1:100 000);
.3 Лужні барвники (рідше
нейтральні) застосовують частіше, ніж
кислі, оскільки більшість бактерій
несуть на поверхні клітини негативний заряд, а в цитоплазмі переважають
речовини з кислими властивостями;
. 4 Під час фарбування
мікроорганізмів відповідно до методики Грама не рекомендується перетримувати
спирт;
. 5 Особливої уваги потребує
фарбування кислотостійких бактерій. У зв'язку з високим вмістом ліпідів (30-40
%) дана група бактерій погано фарбується звичайним розчином анілінових
барвників, тому необхідно застосовувати концентровані барвники за умови
прогрівання мазка, або флуорохроми (наприклпд, родамінон - аурамін);
.6 Під час дослідження культивованих
мікроорганізмів, характер росту колоній вказує на їх рухливість. Адже, рухливі
бактерії дають диференційний ріст по всій товщині поживного середовища.
.7 Молекулярний рух помилково може
бути прийнятим за рухливість мікроорганізмів. При цьому не рухливі бактерії
коливаються між двома близькими точками, ніби “танцьовуючи на місці”;
.8 Під час виявлення типу
розташування джгутиків варто звертати увагу на швидкість та характер руху
мікроорганізмів. Оскільки, швидкі рухи притаманні монотрихам та лофотрихам,
перетрихи та амфітрихи характеризуються менш енергійною та безладною
рухливістю.
. 9 Якість фарбування джгутиків
залежить від чистоти та знежирення скельця. Вік досліджуваної культури повинен
бути не старшим від 12-18 год.;
.10 За характером розташування
клітин можна судити про здатність бактерій до спороутворення. Адже, більшість
видів спороутворюючих бактерій утворюють ланцюжки. Тоді як, неспороутворюючі
бактерії ланцюжків не утворюють.
. 11 Якість забарвлення спор та
джгутиків можна підвищити, застосовуючи протрави (наприклад, солі танінової
кислоти).
. Під час ідентифікації
мікроорганізмів необхідно враховувати, що:
. 1 Колір колоній не може слугувати
ключовим критерієм, який визначає видовий спектр мікроорганізмів;
.2 Тинкторіальні властивості
досліджуваних культур частково вказують на патогенність. Адже, більшість
сапрофітних мікроорганізмів є грампозитивними, а серед грамнегативних
переважають патогенні;
.3 На результати фарбування за
Грамом суттєво впливають відхилення від умов культивування мікроорганізмів.
Так, старі культури грампозитивних можуть здатися грампозитивними. В кислому
середовищі всі мікроорганізми можуть бути сприйнятими за грамнегативних;
.4 Види бактерій можуть бути
представленими багатьма підвидами, які можна розглядати як етап формування
виду;
.5 Паличкоподібним бактеріям
найбільшою мірою характерний плеоморфізм;
3. До основних методів
зменшення загального мікробного числа належить:
3.1 Провітрювання:
3.2 Вологе
прибирання;
3.3 Підвищення
впливу сонячного випромінювання чи застосування за потреби ультрафіолетових
бактерицидних ламп;
3.4 Контроль
вологості повітря, яка має складати 30-50 %. Якщо вона вища за 50 % потрібні
висушувати повітря за допомогою електроприладів. Якщо нижча за 30 % -
зволожувати;
3.5 Збільшення
кількості кімнатних рослин , які не лише виділяють фітонциди, але й на своїх
листках затримують часточки пилу. Проте, якщо необхідно зменшити кількість
грибів в повітрі, то необхідно навпаки зменшувати кількість рослин, оскільки
грунт є середовищем розмноження грибів.
4. Адміністрації
факультету посилити увагу за контролем санітарного стану навчальних та
допоміжних приміщень протягом дня. Сприяти вирішенню питання щодо розмежування
аудиторій,де зберігається верхній одяг від навчальних аудиторій. Адже більшість
шкідливих мікроорганізмів залишаються на одязі та на взутті. Розробити
«Пам’ятку чистоти» та помістити її в загальнодоступному місці.
5. Кураторам
проводити виховні години, присвячені необхідності дотриманню правил особистої
гігієни студентами
6. Студраді
ініціювати проведення виховних заходів, присвячених питанням чистоти та гігієни
7. Студентам:
Провітрювати приміщення,принаймні 1 раз на добу. Але при можливості 4-6 разів
на добу. Слідкувати за особистою гігієною та чистотою свого одягу. Після
кожного дзвінка на перерву виходити в коридор.
СПИСОК ВИКОРИСТАНИХ ДЖЕРЕЛ
1. Блохина
И. Н. Систематика бактерий (с основами геносистематики) / Блохина И. Н,
Леванова Г. Ф., Антонов А. С. - Новгород: Изд-во Нижегород, 1992. - 169 с.
2. Борисов
Л. Б. Медицинская микробиология, вирусология, иммунология:підруч. [для студ.
вищ. навч. закл.] / Борисов Л. Б.. - М.: ООО Медицинское информационное
агентство, 2005. - 736 с.
3. Векірчик
К. М. Мікробіологія з основами вірусології: підручник [для студ. вищ. навч.
закл.] / К. М. Векірчик. - К.: Либідь, 2001. - 312 с.
4. Векірчик
К. М. Практикум з мікробіології: навч. Посібник/ К. М. Векірчик. - К.: Либідь,
2001. - 144 с.
5. Віннікова
О. І. Дидактичні матеріали з мікробіології та вірусології : методичний матеріал
/ О. І. Віннікова // Біологія. - 2008. - № 3 . - С. 2-11.
6. Віннікова
О. І. Практикум з мікробіології: [методичні рекомендації ] / О. І. Віннікова,
І. М Моргуль. - [2-ге вид.].- Х.: ХНУ імені В. Н. Каразіна, 2009. - 33 с.
7. Герхардт
Ф. Методы общей бактериологии; под ред. Ф. Герхардта.- Москва: Мир, 1983. -
1272 с.
8. Громов
Б. В. Строение бактерий / Б. В. Громов. - Ленинград : Изд-во Ленинградского
университета, 1985. - 192 с.
9. Гудзь
С. П. Мікробіологія: підруч. [для студ. вищ. навч. закл.] / Гудзь С. П., Гнатуш
С.О., Білінська. І. С. - Львів: Видавничий центр ЛНУ імені Франка, 2009. - 360
с.
10. Данилейченко
В. В. Мікробіологія з основами імунології / Данилейченко В. В., Федечко Й. М.,
Корнійчук О. П. - [2-ге вид.]. - К. : Медицина, 2009. - 392 с.
11. Жизнь
растенй В 6 т. / Гл. Ред. Ал. Федорова. - Т.1: Введение. Бактерии и
актиномицеты / Под. Ред. Н. А. Красильникова и А. А. Уранова. - М.:
Просвящение, 1974. - 487 с.
12. Люта
В.А. Основи мікробіології, вірусології та імунології: підруч. [для студ. вищ.
навч. закл.] / В. А. Люта, Г.І. Загорова. - Київ: Здоров"я, 2001.- 280 с.
13. Мікробіологія;
підр. [для студ. вищ. навч. закл.] / [Дикий І. Л., Холупяк І. Ю., Шевельова Н.
Ю. та інші]; за ред. І. Л. Дикого. - Х.: Вид-во НфаУ Оригінал, 2006. - 432 с .
14. Мікробіологія
та фізіологія харчування: посібник [для студ. вищ. навч. закл.] / [Маличіна В.
Д., Ракша-Слюсарева О.А., Рошова В. В. та ін.]. - К.: Кондор, 2009.- 242 с.
15. Определитель
бактерий Берджи: В 2-х т. [под ред. Дж. Хоулта, Н. Крига, П. Снита, Дж. Стейли,
С. Уилльямса].- Т. 1. пер. с англ. Г. А. Заварзіна. - [9-е изд.]. Москва: Мир,
1997. - 432 с.
16. Пирог
Т.П.Загальна мікробіологія / Пирог Т.П. - К.: Вид-во НУХТ, 2004. - 471 с .
17. Практична
мікробіологія / [Климнюк С.Л., Ситник I.O., Творко М.С., Широбоков В.П.]. -
Тернопіль: Укрмедкнига, 2004. - 440 с.
18. Практикум
по микробиологии: учеб. пособие [для студ. высш. учеб. Заведений] / [ Нетрусов
А. И.,. Егорова М. А, Захарчук Л. М. и др.]. под ред. А. И. Нетрусова. - М.:
Издательский центр “Академия”, 2005. - 608 с.
19. П'яткін
К.Д. Мікробіологія з вірусологією та імунологією: підруч. [для студ. вищ. навч.
закл.] / К.Д. Пяткін, Ю.С. Кривошеїн. - К. : Вища школа, 1992. - 431 с.
20. Сергійчук
М.Г. Мікробіологія: [методичні рекомендації до лабораторних робіт] / М.Г.
Сергійчук, Т.М. Фурзікова. - К.: Вид-во Міжнародного Соломонового університету.
- 66 с.
21. Сиротин
А.А Практикум по микробиологии / [сост. Сиротин А.А]. - Белгород: Изд-во БелГУ,
2007. - 80 с.
22. Современная
микробиология: В 2-х т./ Ленгелер Й., Древс Г., Шлегель Г. - т. 1: Прокариоты;
под. ред. Ленглера Й., Древс Г., Шлегель Г.; пер. с англ. Берга И.А.,
Ивановского Р.Н., Нетрусова А.И. и др..- Москва: Мир, 2005. - 656 с.
23. Теппер
Е. З. Практикум по микробиологии / Е. З. Теппер, В. К. Шильникова, Г. И.
Переверзева . - [2-е изд.]. - Москва : Колос, 1979 . - 216 с.
24. Титов
Л.П. Классификация, номенклатура и эволюция значимых для медицины бактерий //
Белорусский медицинский журнал. - Минск, 2011.
С.
224-227.
25. Федоров
М. В. Микробиология / М. В. Федоров; под. Ред. Е. А. Мишустина. - [7-е изд.]. -
М.: Изд-во сельськохозяйственной литературы, 1963. - 448 с.
26. Фробимер
М. Основы микробиологии / М. Фробимер; пер. С англ. В. А. Шорина. - Москва:
Мир, 1965. - 678 с.
Додаток А.1
Характеристика основних форм клітин
мікроорганізмів
Форма та розміри бактерій мають
велике таксономічне значення і є важливими критеріями для їх ідентифікації.
Науковці поділяють бактерії за
формою на чотири основні групи: сферична (коки), циліндрична (палички),
спіралеподібна (звивисті), ниткоподібна (нитчасті). [20] Між цими формами
існують чисельні проміжні, тому доволі часто важко визначити форму
бактеріальної клітини [21].
 Шароподібні форми
бактерій називають коками (від лат. kokkus - ягода, зерно) (рис. А.1.) .
Шароподібні форми
бактерій називають коками (від лат. kokkus - ягода, зерно) (рис. А.1.) .
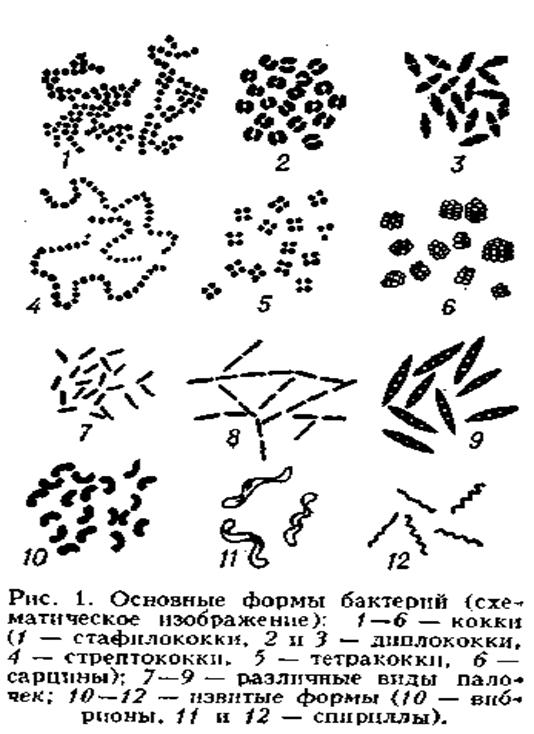
Мал. А.1 . Коки: 1 - стафілококи,
2 і 3 - диплококи, 4 - стрептококи, 5 - тетракоки, 6 - сарцини.
Кокоподібні бактерії мають вигляд
правильної кулі діаметром 1,0-1,5 мкм, а деякі бобо-, ланцето-, еліпсовидної
форми. Просторове розміщення клітин, що утворилися після поділу є сталою
ознакою в межах роду [9]. В залежності від кількості площин ділення та
розташування бактеріальних клітин після ділення коки поділяють на:
· мікрококи
(від лат. micro - маленький)
Діляться в одній або декількох
площинах, розташовуються поодиноко, або утворюють неоформлені скупчення.
Сапрофіти - типові мешканці води, ґрунту та повітря. Патогенних форм не
описано.
Представники: Micrococcus luteus, M.
roseus, M. lysodeikticus, M. varians.
· диплококи
(від лат. diplococcus - подвійний)
Діляться в одній площині, клітини
після поділу не розходяться, а об'єднуються попарно (Diplococcus muscosus, D.
magnus). Клітини можуть мати бобоподібну форму (Neisseria gonorrhoeae) або
ланцетоподібну форму.
Представники: Azotobacter
chroococcum, N. meningitidis.
· стрептококи (від
лат. streptos - ланцюг)
Виявлені Т. Більротом в 1874 р. І
через кілька років Пастером при дослідженні гнійних захворювань і сепсисі. Рід
Streptococcus включає багато видів, які різняться між собою за екологічними,
фізіологічними та біохімічними ознаками, а також патогенні для людини -
грампозитивні [2].
Діляться в одній площині, утворюють
ланцюжки різної довжини, оскільки клітини зберігають між собою зв'язок.
Представники: Streptococcus
pyogenes, S. faecalis, S. lactis, S. bovis.
· тетракоки (від гр.
tetra - чотири)
Діляться у двох
взаємоперпендикулярних площинах, утворюють тетради.
Представники: Deinococcus
proteolyticus, D. radiodurans, Stomatococcus mucilaginosus, Tetracoccus
anaerobicus, T. soyae.
· cарцини (від лат.
sarcina - зв’язка, пакет)
Діляться у трьох
взаємноперпендикулярних площинах і утворюють пакети із 8, 16, 32 і більше
клітин. Хвороботворних видів серед сарцин не встановлено.
Представник: S. ventriculi, S.
maxima.
· cтафілококи (від
лат. staphylos - грона винограду)
Відкриті Луї Пастером у 1880 році.
Рід Staphylococcus включає 19 видів, з них 3 види екологічно пов'язані з
організмом людини [5].
Діляться нерівномірно в декількох
площинах і утворюють скупчення у вигляді виноградних грон. Серед стафілококів
зустрічаються вільноживучі сапрофіти (на молочних, м‘ясних продуктах, у ґрунті,
повітрі, на поверхні рослин), а також патогенні для людини форми. Найчастіше є
збудниками гнійних запалень.
2. Найбільш чисельною та
різноманітною групою бактерій є паличкоподібні (циліндричні) форми бактерій
(рис. А.2). Довжина циліндричних бактерій коливається у досить широких межах,
від 0,8 мкм до 11 мкм, їх ширина є більш стабільним показником - 0,5-1,0 мкм.
Короткі палички (кокобактерії) мають розміри до 1 мкм (збудники коклюшу,
бруцельозу, тюляремії), довгі - понад 3 мкм (клостридії, кишкові палички та
інші.) [12].

Рис. А.2. Різноманітність
паличок: 1 - палички, які спор не утворюють; 2-4 паличкоподібні спороутворюючі
бактерії бацилярного типу (2), клостридіального (3), плектридіального типу(4).
Крім розмірів у різних видів
паличкоподібних бактерій також звертають увагу на форму кінця клітини, вони
можуть бути загострені, овальні, різко обрізані або навіть дещо потовщеними
[9]. За діаметром розрізняють тонкі (мікобактерії туберкульозу) і товсті
(клостридії) палички, а за формою кінців - заокруглені (E. coli, шигели,
сальмонели), овоїдні (збудник чуми), обрубані (збудник сибірки), потовщені (рід
Corenebacterium), булавоподібні (збудник дифтерії), загострені (рід
Fusobacterium), а також мати яйцеподібну форму (Yersina pestis) [12].
На мікроскопічних препаратах також
звертають увагу на характер розташування клітин. Так, більшості видів
спороутворюючих бактерій властиво утворення ланцюжків, тоді як неспороутворюючі
бактерії ланцюжків не утворюють. Палички можуть розташовуватись поодиноко, попарно
у вигляді букв Х та V, або ієрогліфів (характерно для біфідобактерій).
Монобактерії розміщуються хаотично (більшість бактерій), диплобактерії, або
диплобацили, попарно, стрептобактерії, або стрептобацили - ланцюжком [12].
З морфологічної точки зору виділяють
два види паличок: спороутворюючі та аспорогенні. До спороутворюючих відносять
такі види бактерій: Amphibacillus, Bacillus, Clostridium, Desulfotomaculum,
Oscillospira, Sporohalobacter, Sporolactobacillus, Sporosarcina,
Syntrophospora, Sulfobacillus.
Хоча вважають, що спороутворюючі
бактерії є більш пристосованими до переживання несприятливих умов навколишнього
середовища, аспорогенні форми бактерій є більш розповсюдженою групою. Найбільш
відомими представниками серед неспороутворюючих паличок є : Esherichia coli,
Serratia marcescens - раніше мала назву Bacterium prodigiosum (чудесна
паличка). Серед неспороутворюючих паличок також досить відомими є бактерії, які
викликають такі розповсюджені захворювання, як дизентерія, черевний тиф,
сальмонельоз [20].
За аналогією із коками, залежно від
взаємного розташування у мазках, паличкоподібні мікроорганізми поділяються на
такі групи:
монобaктерії - при поділі
розташовуються поодиноко (Escherichia coli, Salmonella tiphy). До них належить
більшість бактерій;
Монобацили також розташовуються
поодиноко, але мають спори (Bacillus subtilus, Clostridium tetani);
диплобактерії розташовуються в
мазках попарно (Klebsiella pneumoniae, рід Zimomonas);
диплобацили - парне розташування
спорових мікроорганізмів;
стрептобактерії - безспорові
палички, які розташовані у вигляді ланцюжків (Haemophilus ducrey);
стрептобацили - спорові мікроби, які
розташовуються ланцюгом (Bacillus anthracis, B. Brevis) [9, 17].
Деякі палички утворюють незначні
розгалуження (представники родів Artrobacter, Corenebacterium, Nocardia,
Mycobacterium) [9].
Під час ідентифікації паличок
необхідно враховувати, що саме даній групі бактерій найбільшою мірою
притаманний поліморфізм [12].
. Третій групі бактерій притаманна
звивиста форма.

Звивисті бактерії відрізняються
кількістю завитків (рис. А.3.). В залежності від кількості витків звивисті
форми бактерій поділяють на вібріони, спірили та спірохети [12]. Також деякі
вчені класифікують мікроорганізми звивистої форми на дві групи:форми, що не
гнуться (вібріони та спірили) і ті, що вигинаються (спірохети) [13].
Вібріони (від лат. vibrio -
вигинаюсь, або vibrare - тріпочучий) - вигнуті палички, які являють частину
(1/4) спіралі. Мають вигляд коми . Широко розповсюджені в морях та витоках
річок, на поверхні морських тварин та вмісті їх кишок. Деякі види виявлені
також у прісній воді. Біля 10 видів є патогенними для людини, декілька
викликають захворювання морських хребетних та безхребетних.

Рис. А.3. Звивисті клітини: 1 -
вібріони; 2 - спірили; 3 - спірохети.
Найбільш відомі збудники захворювань
людини - Vibrio cholerae, збудник холери,V. parahaemolyticus - збудник харчових
отруєнь, які викликані вживанням зараженої риби або молюсків, та V. vulnificans,
який викликає септицемію з високим відсотком летальності [10, 19].
Спірили (від лат. spiro - штопор) -
на відміну від вібріонів, їх клітини більш товсті, довгі та звивисті. Спірили
можуть мати від 1-2 завитків і нагадувати російську С, або англійську S, до
8-10 витків. Всі ці бактерій утворюють ригідну (шорстку) клітинну стінку. В
основному це сапрофіти, які мешкають у стоячих, забруднених водоймищах, а також
гниючих рештках рослинного та тваринного походження. Типовими представниками є
Spirillum pleomorphum, S. volutans, S. minus.
Спірохети - сильно звивисті палички,
які на думку Сергійчука М.Г., Фурзікової Т.М. (2001) мають більше 10 витків
[20]. Люта В. А. відносить спірохет до звивистих паличок, які мають більш ніж 3
завитки різної величини та щільності. Звивистість клітини зумовлена характером
руху бактерій [12].
До групи спірохет відносять 7 родів,
з яких 3 відіграють суттєву роль в інфекційній патології людини. Це роди
Treponema, Borrelia и Leptospira (рис. А.4.). У трепонем є 8-14 завитків І
порядку, однакових за формою та величиною. У борелій нараховується 4-8 великих
завитків неправильної форми, неоднакових за величиною. Ці мікроорганізми
найбільші серед патогеннихспірохет. Лептоспіри мають 12-18 завитків І порядку,
але вони дрібні й тісно прилягають один до одного, під звичайним світловим
мікроскопом практично не диференціюються [2]. На їхніх кінцях є вторинні
завитки, які надають їм S- або С- подібної форми і роблять їх схожими на гачки.
Вони є грамнегативними. Але фарбування за Грамом не використовують, бо
спірохети погано сприймають анілінові барвники [16].
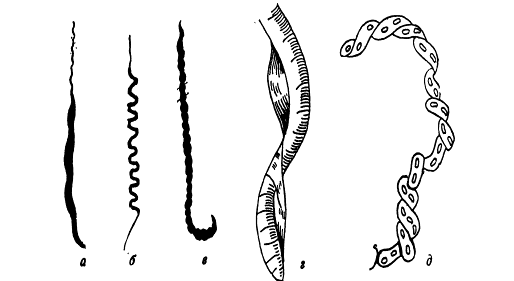
Рис. А.4. Морфологія різних видів
спірохет: а - борелія; б - трипонема, в - лептоспіра; г - спірохета; д -
кристиспіра.
Серед спірохет є вільноживучі форми,
які зустрічаються у прісних та солоних озерах, донних відкладеннях (Spirochaeta
plicatilis, S. isovalerica). Деякі спірохети є патогенними для людини
(Treponema pallidum - збудник сифілісу, Leptospira interrogans - збудник
інфекційної жовтухи, Borrelia duttoni, B. persica - збудник зворотного тифу)
[2].
4. В
окрему морфологічну групу можуть бути виділені мікобактерії (нитчасті). Прямі,
або зігнуті палички, іноді можуть галузитись, можливе утворення ниток, або
міцелієподібних утворень (рис. А.5.) Це переважно одноклітинні та
багатоклітинні мікроорганізми. Їхні нитки утворені багатьма клітинами,
з’єднаними за допомогою слизу, чохлів, піхв, плазмодесмів тощо. Нитчасті є
переважно водяними мікробами. Нитки трихомних бактерій можуть вільно плавати у
воді (Begiattoa alba, ціанобактерії родів Oscillatoria, Spirulina) або бути
прикріпленими до субстрату (Thiotrix nivea). [3] Але ці структури легко
розпадаються на палички або коки. Представники: Mycobacterium tuberculosis. M.
leprae [20].

Рис. А.5. Нитчасті бактерії: 1 -
Beggiatoa; 2 - Thiothrix; 3 - Saptospira; 4- Simonsiella; 5 - Caryophanon; 6 -
ціанобактерії роду Microcoleus; 7- Leptotrix; 8 - Sphaerotilus; 9 - Сrenothrix.
Отже, мікроорганізми здатні
формувати асоціації різноманітного рівня складності - трихоми, агрегати
правильної чи неправльної форми, а також стабільні чи нестабільні “спільноти” з
іншими видами мікроорганізмів чи тканинами еукаріотів [22].
Крім перелічених основних форм
прокаріотів, виявлені бактерії, які мають незвичну форму. Бактерії незвичної
форми морфологічно різноманітні: тороїдальні (Microcyclus major, M. aquaticus),
тубероїдні, червоподібні, нагадують правильну шестикутну зірку (рід Stella),
пласкі квадратові пластинки, гантелі, черв'якоподібної форми. Архебактерії роду
Haloarcula мають форму плоских неправильних трикутників і прямокутників. Вони
беруть участь у процесах біодегратації природніх органічних сполук [3].
Деякі одноклітинні бактерії
утворюють різноманітні вирости. Ці вирости можуть бути у вигляді простек, або
стеблинок (рис. А.6.) .

Мал. А.6. Бактерії, які здатні
утворювати вирости: 1- Caulobacter; 2- Hyphomicrobium; 3 - Ancalomicrobium; 4 -
Gallionella.
Кількість простек може бути різною,
від однієї до декількох. До того ж, простеки можуть роздвоюватись. Кількість,
розмір, розташування простек є диференційною ознакою для визначення роду.
Представники: Ancalomicro-bium adetum, Asticcacaulis excenticus.
Стеблинка - неклітинний виріст, не
містить цитоплазми і не оточений клітинною стінкою. Клітини кокоподібні,
овальні, грушоподібні [16]. Не мають пептидоглікану. Утворюють розетки або
нитки. Представник: Planctomyces bekefii [20].
Слід зазначити, що більшість
бактерій характеризується постійністю форми завдяки клітинній стінці. Але є
бактерії для яких, характерним є плеомофізм.
Люта В. А. (2001) тлумачить поняття
плеоморфізму так: “Плеоморфізм - це здатність змінювати форму під дією різних
факторів (антибіотиків, дезінфекційних розчинів, умов культивування)” [12].
Також, під плеоморфізмом розуміють різноманітність форм і розмірів
мікроорганізмів залежно від віку та стадії розвитку, що зумовлюється
індивідуальною мінливістю [19].
Ще в 1850 р. було опубліковано дані
про зміну морфологічних форм в циклі розмноження іржастих та сажкових грибів.
Ця публікація поклала початок дискусії про плеоморфізм та мономорфізм бактерій.
Деякі дослідники вважали, що одна і та ж бактерія може існувати в різноманітних
формах (плеоморфізм), обумовлюючи при цьому хвороби і утворюючи різні продукти
метаболізму, залежно від умов росту. Прибічниками даних тверджень були такі
авторитетні вчені як Т. Більрот, К. Негелі, Є. Хеліер, Дж. Лістер, Р. Ланкастер
і Цонф. Інші вчені, такі як Пастер, Кон, Кох і О. Брефельд, доводили важливість
вивчення чистих культур і, таким чином, підкреслювали важливість експериментів.
Отже, визнання мономорфізму, тобто постійності видів бактерій, потребувало
багато часу. Твердження про постійність бактерійних видів звісно викликало
сумніви, адже дослідники спостерігали мінливість і мутації навіть в чистих культурах
[22].
Плеоморофізм властивий таким видам
бактерій як: Corynebacterium diphtheriae, Arthrobacter globiformis,
нокардіобактерії, в яких в циклі розвитку спостерігається зміна форми клітин:
кок-паличка-кок. До плеоморфних бактерій також належать мікоплазми
(Acholeplasma laidlawii, Mycoplasma mycoides , L- форми, клітини яких не мають
постійної форми [20].
У процесі росту форма клітин може
суттєво змінюватись від коків до паличок (і навпаки), неправильних форм,
розгалужених ниток [9]. Мікроорганізми дуже пластичні і легко змінюються під
впливом факторів середовища (антибіотики хімічні реагенти, підвищена або
понижена температура, концентрація солей, кислотність, продукти метаболізму,
дезинфікуючі агенти, лікарські засоби, інгібітори організмі тощо). В результаті
можуть виникати незвичайні за формою і розміром клітини (інволюційні форми):
дуже збільшені, роздуті, кулясті, колбоподібні або ниткоподібні. Вони
ревертують до вихідних форм, коли дія факторів, що індукували їхнє утворення,
припиняється [9, 19].
Такі зміни морфології пов'язані з
порушенням або синтезу клітинної стінки бактерій, або механізму регуляції їх
клітинного поділу.
Необхідно зауважити, що бактерії
характеризуються високим ступенем поліморфізму (індивідуальна множинність
форми, що не передається спадково) особливо під час культивування на
синтетичних поживних середовищах
Властивість мікроорганізмів
змінюватись під впливом різних факторів навколишнього середовища враховують у
лабораторній діагностиці інфекційних захворювань, при виготовленні інфекційних
препаратів, що застосовуються з інфекційною та лікувальною метою [13, 19].
Під час ідентифікації бактерій
необхідно враховувати, що при частковому розчиненні клітинної стінки під
впливом лізоциму (Salton, 1952) або інших факторів паличкоподібні (переважно
грамнегативні) клітини перетворюються у сферичні тіла, що дістали назву
сферопласти. Також існують і протопласти, які утворюються в результаті
пригнічення синтезу клітинної стінки, що веде до порушення координації між
ростом і поділом клітини; найчастіше це буває під впливом пеніциліну.
Морфологічно вони являють собою крупні кулясті і ниткоподібні плазматичні
структури. До протопластів відносять L-форми бактерій. Розрізняють стабільні і
нестабільні L-форми. Стабільні за своїм властивостями дуже подібні до мікоплазм
[19].
Отже, мікроорганізми у порівнянні з
еукаріотами характеризується значною одноманітністю морфологічних форм.
Додаток
А.2.
Класифікація мікроорганізмів за
Бергі
У дев'ятому виданні
Визначника бактерій Бергі всі виявлені організми, віднесені в царство
Prokaryotae, розділені на 35 груп.
Група 1. Спірохети.
Включає порядок Spirochaetales. Тонкі спіралевидні одноклітинні форми, що
володіють своєрідною морфологією й способом руху. Довжина клітин коливається
від 5 до 250 мкм. Схильні до утвору аномальних форм (гранул, цист). Клітини
складаються із протоплазмового циліндра, аксіальної нитки й зовнішньої
оболонки. Оболонка тонка й еластична, що й забезпечує спірохетам своєрідний
спосіб пересування. Грамнегативні.
Група 2. Аеробні,
рухливі спіралеподібні або вигнуті грамнегативні бактерії. Прокаріоти, що
входять у цю групу, мають тверду клітинну стінку, так що клітина свою форму не
змінює. Рух здійснюється за допомогою одного або безлічі полярно розташованих
джгутиків.
Група 3. Нерухливі
грамнегативні вигнуті бактерії. Включає сімейство Spiromonaceae, що поєднує
облігатно аеробні форми з характерною клітинною морфологією: від прямих паличок
до кілець, не повністю або повністю закручених; при цьому в культурі одночасно
можуть бути присутнім клітини різної форми.
Група 4.
Грамнегативні аеробні палички й коки. Група представлена восьма родинами. До
родини Pseudomonadaceae відносяться поодинокі прямі або злегка вигнуті рухливі
палички. Рух здійснюється за допомогою полярно розташованих джгутиків. Типові
представники родини об'єднані в рід Pseudomonas.
Група 5.
Факультативно анаеробні грамнегативні палички. Поєднує 3 родини: зокрема
Enterobacteriaсеае. До складу першої родини належать рухливі або нерухливі
неспороутворюючі палички. Деякі види утворюють капсули.
Група 6. Анаеробні
грамнегативні прямі, вигнуті або спіралевидні палички. Це палички правильної
форми або схильні до плеоморфізму, неспороутворюючі, нерухливі або рухливі.
Група 7. Бактерії,
що характеризуються дисиміляційним відновленням сірки або сульфату. У складі
групи еубактерій з різною морфологією й наступними однаковими властивостями:
грамнегативні строгі анаероби.
Група 8. Анаеробні
грамнегативні коки. Представлені однією родиною Veillonellaceae. Коки, як
правило, з'єднані попарно, але можуть утворювати ланцюжки або скупчення клітин.
Група 9. Рикетсії й
хламідії. До складу групи включено два порядки: Rickettsiales і Chlamydiales.
Перший поєднує бактерії, що характеризуються в більшості випадків сукупністю
наступних ознак: плеоморфні, нерухливі, грамнегативні, з типовими для
еубактерій клітинними стінками, що розмножуються розподілом всередині
клітин-хазяїв. Серед рикетсій є рухливі види, що фарбуються позитивно по Граму.
Група 10.
Мікоплазми. До них відносяться форми, у яких відсутня клітинна стінка.
Таксономічне значення цієї ознаки дозволила всі прокаріоти, що не мають
клітинної стінки, виділити в групу, привласнивши їй ранг відділу. Для них
характерний яскраво виражений поліморфізм. У культурі одного виду можна
одночасно виявити великі кулясті тіла, дрібні зерна, клітини еліпсовидної,
дископодібної, паличкоподібної й ниткоподібної форми. Останні можуть
галузитись, утворюючи структури, подібні міцеліальним.
Група 11.
Ендосимбіонти. У цю групу виділені прокаріоти - ендосимбіонти найпростіших,
комах, грибів і безхребетних. Для більшості представників ендосимбіоз
облігатний і їх не вдалося культивувати в лабораторії в чистій культурі.
Група 12.
Грампозитивні коки. До складу групи входять представники 15 родів, що значно
різняться філогенетично й фенотипічно.
Бактерії,
поєднувані в сімейство Micrococcaceae - коки, що діляться більш ніж в одній
площині, схильні не розходитися після поділу й тому утворюючі скупчення
сферичної або неправильної форми.
Група 13.
Грампозитивні палички й коки, що утворюють ендоспори. У складі групи
представники 6 родів. Два з них (Bacillus і Clostridium) найбільш численні й
цікаві. Рід Bacillus поєднує рухливі паличкоподібні клітини, розміри яких
коливаються в досить широких межах. Джгутики розташовані перитрихіально.
Фарбування по Граму по-різному: позитивно або позитивно тільки в молодій
культурі. До складу роду Clostridium входять палички, що відрізняються від
попереднього роду формою спороутворення й облігатно анаеробним способом
існування.
Група 14.
Грампозитивні палички правильної форми. Група - конгломерат, що складається з 7
родів, об'єднаних декількома загальними морфологічними й фізіологічними
ознаками: клітини паличкоподібної форми (від кокоподібних до подовжених, від
поодиноких до здатних утворювати ланцюжки).
Група 15.
Грампозитивні палички неправильної форми. Група різноманітна. Більшість -
грампозитивні палички неправильної форми, що ростуть у присутності повітря й не
утворюють ендоспор, але є в групі бактерії, що мають форму коків або паличок
правильної форми, що зафарбовуються негативно по Граму, що і є строгими
анаеробами.
Так, до роду
Corynebacterium відносяться форми, схильні до морфологічної мінливості. Крім
коротких паличок у культурі можна виявити кокоподібні форми, клітини, що мають
булавоподібні випинання, слабкорозгалуджені форми. Для представників цього роду
характерний утворення фігур, що складаються із розташованих під кутом або, що
примикають друг до друга дочірніх клітин.
Група 16. Ковзаючі
бактерії, які утворюють плодові тіла. Джгутики відсутні. Не здатні до ковзання
по твердій поверхні. За нестачі поживних речовин клітини здатні агрегуються з
утворенням плодових тіл, що формуються з видозміненого слизу та клітин, часто
яскраво забарвлені і видимі неозброєним оком. Їх плодові тіла варіюють за
будовою від простих грудочок до складних структур.
Група 17.
Грампозитивні коки. Не здатні утворювати спори. Серед яких виділяють:
· аеробні
коки, які можуть розташовуватись попарно, тетрадами та утворювати скупчення;
· факультативні
анаеробні чи мікроаерофільні коки, які розташовуються попарно, утворюють
ланцюги, кластери, тетради;
· строго
анаеробні коки, які зустрічаються в парах, тетрадах чи пакетах кубічної форми.
Група 18.
Грампозитивні коки і палички, які утворюють ендоспори. Серед коків лише один рід
ж рухомим (в тетрадах чи кубічних пакетах). Частіше виявляють рухомі палички.
Група 19.
Грампозитивні палички правильної форми, які не утворюють спор. Паличковидні
клітини (від коковидних до видовжених паличок чи ниток), Незабарвлені (лише для
одного роду характерна світло-жовта пігментація).
Група 20.
Грампозитивні палички не правильної форми, які не утворюють спор. У деяких
моєуть бути присутні булавовидні форми, ниткоподібні елементи чи одночасно
палички або ниткоподібні елементи і коковидні форми. Деяким видам характерний
цикл розвитку палички/коки. Один з родів включає в себе організми від
грамнегативних до грам варіативних.
Група 21.
Мікобакетрії. Аерлбні, нерухомі, спор не утворюють. Мають паличковидну форму,
характеризуються кислотостійкістю. За Грамом фарбуються слабко. Іноді утворюють
нитки, що галузяться. Повітряний міцелій не утворюють.
Група 22-29
Актиноміцети. Грампозитивні бактерії. Утворюють нитки, що галузяться чи гіфи у
вигляді міцелія, який має бути стабільним чи розпадатися на паличковидні чи
коковидні елементи. Деякі види мають джгутики.
Група 31.
Метаногени. Суворо анаеробні. Здатні утворювати метан як кінцевий продукт
метаболізму. Можуть давати синьо-зелену флуоресценсію при опроміненню світлом з
довжиною хвилі 420 нм.
Група 32. Сульфадредукуючі
археї. Суворо анаеробні, здатні утворювати із сульфату в процесі дисиміляційної
сульфатредукції. При мікроскопії в УФ виявляють синьо-зелену флуоресценсію.
Група 33.
Галорбактерії. Грамнегативні чи грампозитивні бактерії. Клітини паличковидні, форма
від правильної до вираженої неправильної.
Група 34.
Архебактерії, що позбавлені клітинної стінки. Мають коковидні клітини,
позбавлені клітинної стінки.
Група 35. Екстремальні
термофіли і гіпертермофіли, метаболізуючі [15].
Додаток Б
Розміри клітин мікроорганізмів
Розміри як і форма мікроорганізмів
мають визначене таксономічне значення і є важливим критерієм при їх
ідентифікації, оскільки це відносно стабільні ознаки у суворо визначених умовах
культивування на штучних поживних середовищах [2].
Розміри бактеріальних клітин сильно
варіюють (табл. Б.1.). Зазвичай вони занадто малі для того, щоб їх можна було
виявити неозброєним оком. Але є виключення - бактерія Epulopiscium fishelsoni,
розмір якої в мільйон раз перевищують об'єм, порівняно з рештою бактерій [22].
Діаметр сферичних бактерій становить
від 0,2 до 0,25 мкм. Найменшими є мікоплазми (0,15 мкм). Цей розмір є
теоретичною межею клітинного рівня організації життя, у якому в клітині ще може
бути мінімум молекул білка (1200) і мінімум ферментативних реакцій, необхідних
для підтримання клітинної структури. До того ж, розміри клітин мікроорганізмів
суттєво можуть суттєво змінюватись під впливом чинників зовнішнього середовища.
Під дією кислот і лугів, зміні температур, через накопичення продуктів обміну у
середовищі існування та інших факторів, клітини зазнають змін [14].
Таблиця Б.1
Розміри деяких бактерій
|
Найменування бактерій
|
Довжина клітини, мкм
|
Діаметр клітини, мкм
|
|
Молочнокислий стрептокок
|
0,8-1,2
|
0,5-0,8
|
|
Вершковий стрептокок
|
0,6-0,7
|
0,6-0,7
|
|
Паличка болгарська
|
4,0-0,5
|
0,6-1,0
|
|
Ацидофільна паличка
|
1,5-6,0
|
0,6-0,9
|
|
Паличка протея
|
0,6-4,0
|
0,4-0,5
|
|
Кишкова паличка
|
1,2-0,3
|
0,5-0,8
|
|
Паличка бруцельозу
|
0,5-2,0
|
0,2-0,5
|
З наведених даних випливає, що
розміри бактерій коливаються від 0,2 до 10 мкм (більшість з них має розміри від
о,5-0,8 мкм до 2-3 мкм) [12]. Так, паличкоподібні бактерії мають товщину
0,5-1,0 мкм, довжину від 1-2 до 10 мкм. Нитчасті форми можуть досягати
макроскопічних розмірів (1 мм) їх можна побачити неозброєним оком. Довжина
спірохет коливається від 1-3 до 100-500 мкм. Більшість патогенних мають розміри
0,2 - 10 мкм [13]. Серед сапрофітів трапляються, велетенські бактерії, що
досягають 55-125 мкм. Нижній розмір одноклітинних бактерій визначається
простором, необхідних для упаковки апарату, який забезпечує незалежне існування
клітини, верхній - оптимальним співвідношенням між поверхнею клітини та об'ємом
[13].
Додаток В
Здатність до утворення спор та
розташування спор в бактеріальній клітині
Кон вперше спостерігав спори сінної
палички (1877), названої пізніше Bacillus subtilis. Описані Р. Кохом Bacillus
antrhracis підтвердило значення ендоспор як диференціальної ознаки для
класифікації бацил [22].
На думку Борисова (2005), спори
бактерій можна розглядати як форму збереження спадкової інформації клітини
бактерій в несприятливих умовах зовнішнього середовища [2]. Здатністю до
утворення ендоспор володіє порівняно невелике число як патогенних, так і
непатогенних бактерій. Ця властивість виявлена у представників 15 родів, які
відрізняються морфологічною і фізико-хімічною різноманітністю. Серед них є
паличкоподібні, сферичні, спіральні, нитчасті форми. Усі вони мають клітинну
стінку, характерну для грампозитивних бактерій, хоча деякі з них не фарбуються
за Грамом [9]. До першої групи відносять бактерії родів Bacillus, Clostridium,
а до непатогенних - сапрофітні представники згаданих родів і деякі коки. В
цілому, до спороутворюючих відносять такі види бактерій: Amphibacillus,
Bacillus, Clostridium, Desulfotomaculum, Oscillospira, Sporohalobacter,
Sporolactobacillus, Sporosarcina, Syntrophospora, Sulfobacillus [13].
При мікроскопічному дослідженні
спори можна побачити завдяки високому заломленню світла. Спори містять усю суху
речовину материнської клітини, хоча займають у 10 разів менший об'єм.
Спори являють собою округлі, овальні
чи еліпсовидні утворення. Бакеріальні ендоспори - це особливий тип спочиваючих
клітин грампозитивних бактерій, які мають специфічні структури: багатошарові
білкові покриви, зовнішню та внутрішню мембрани, кортекс, іноді екзоспоріум.
Спори різних видів мають різну
форму, розміри, характер поверхні та локалізацію у клітині (рис. А.7.). Якщо
діаметр спори не перевищує діаметр клітини, в якій утворюється спора, то
клітина належить до бацилярного типу. У тілі бацил і клостридій спори можуть
розташовуватись: центрально (збудник сибірки), субтермінально (збудники
ботулізму, анаеробної інфекції та ін.), термінально (збудник правця). Якщо ж
діаметр перевищує, то в залежності від розташування спори (в центрі чи на кінці
клітини) - клостридіальний чи плектридіальний. В бацилярній клітині спора може
розташовуватись як в центрі клітини (центральне розташування) і на кінці
клітини (термінальне), ближче до одного з кінців (субтермінальне), і при цьому
клітина не змінює своєї форми. Такі клітини називають бацилами (Baccilus).
Спори можуть займати клостридіальне положення в клітині. В даному випадку при
формуванні спори клітина змінює форму, набуваючи вигляду човника чи веретена.
Такі клітини називають клостридіями (Clostridium, від грец. Closter - веретено)
[9]. В клостридій діаметр спор перевищує діаметр бактеріальної клітини.
Плектридіальне розташування характеризується тим, що спора локалізуються
термінально, у місці її розміщення клітина розширюється і набуває форми
ракетки. Клостридіальний і плектридіальний типи розміщення спор властиві в
основному видам роду Clostridium і нерідко одночасно зустрічаються в культурі
одного виду При субтермінальній локалізації спор такі особини стають
веретеноподібними. До них належать клостридії молочнокислого бродіння (C.
Butyricum) [13]. У клостридій правця діаметр спори перевищує її ширину, але
розміщується спора на кінці клітини, в результаті чого мікроорганізм за формою
нагадує барабанну паличку. До того ж, в анаеробних бактерій на спорах
утворюються різноманітні за формою відростки: фібрилярні, стрічкоподібні,
булавовидні тощо. Вони формуються перед закладанням кортексу з білків або
полісахаридів. Їхня функція остаточно не з'ясована [16].

Рис. А.7 Схематичне зображення
типових форм спор та їхня локалізація у клітинах бактерій: 1- термінально
розміщена спора з білковим включеннями; 3 - термінальна спора булавоподібна; 4
- центральна спора клостридієподібна; 5 - кругла термінальна спора; 6 -
латеральна спора.
Спороутворення - один із
найскладніших процесів диференціації бактеріальної клітини. Вважається, що цей
процес принципово однотиповий у всіх видів, які мають ендоспори. У кожній
бактеріальній клітині формується тільки одна ендоспора. Найкраще процес
спороутворення досліджений у представників родів Bacillus та Clostridium,
зокрема Bacilus subtilis, Streptomyces та Myxococcus xanthus.
Виділяють сім етапів спороутворення
(рис. А.8.). Споруляція починається після завершення логарифмічної фази росту,
відрахунок стадій спороутворення - після останнього клітинного поділу.
Рис. А.8. Схематичне зображення
формування ендоспори спороутворюючими бактеріями: 1- нуклеоїд; 2 - цитоплазма;
3 - ЦПМ; 4 - клітинна стінка; 5 - спорова перегородка; 6 -зовнішня мембрана
спори; 7 - внутрішня мембрана спори; 8 - кортекс; 9 - покриви спори; I
-вегетативна клітина; II - на одному з полюсів клітини відбувається ущільнення
цитоплазми; III - формування септи; IV -мембрана більшої клітини та її
цитоплазма оточує меншу клітину; V- утворюється проспора; VI - утворюється
кортекс; VII - завершується дозрівання спор; VIII -лізис материнської клітини
та звільнення спори; IX -вільна зріла спора; X -проростання спори.
На початковій стадії клітина містить
один або декілька нуклеоїдів.
На першій стадії у клітині
з'являється поздовжнє хроматинове тіло (осьовий тяж), з'єднане через мезосому з
цитоплазматичною мембраною. Ця стадія спороутворення є оборотною, і в разі
перенесення клітин у повноцінне поживне середовище відновлюється поділ
нуклеоїда і клітини.
На другій стадії осьовий тяж
ділиться на дві частини: велику - нуклеоїд спорангія (спороутворююча клітина)
та малу - нуклеоїд майбутньої спори. Малий нуклеоїд локалізується на полюсі
клітини. Цей поділ супроводжується появою перегородки, що утворюється за
рахунок інвагінації цитоплазматичної мембрани. Проте після появи перегородки не
утворюється поперечна стінка, як це простежується в разі клітинного поділу.
Замість цього мембрана більшої частини клітини (спорангія) та її цитоплазма
оточують меншу частину клітини. Отже, утворюється проспора, оточена двома
частинами мембрани - зовнішньою та внутрішньою. На цьому завершується третя
стадія, після якої процес спороутворення стає необоротним. Наступна стадія
характеризується утворенням специфічної оболонки спори - кортексу. Він
формується між мембранами проспори з пептидоглікану певної структури.
На п'ятій стадії спору оточують
білкові оболонки, що становлять від 30 до 60% її ваги. Спори на цій стадії є
оптично активними та стійкими до дії різних факторів середовища.
У багатьох бактерій ззовні білкових
оболонок формується екзоспоріум, який складається з ліпідів та білків і виконує
захисну функцію.
На шостій стадії завершується
дозрівання спор. Воно супроводжується певними хімічними перебудовами, а саме:
синтезом дипіколінової кислоти. Спори на цій стадії стають термостійкими.
На завершальній стадії відбувається
лізис спорангія та звільнення спори. Утворені спори містять незначну кількість
води ( до 15% від маси спори) і перебувають у стані анабіозу.
Спори не є обов'язковою стадією
життєвого циклу бактерій. Утворення спор простежується лише тоді, коли не
вистачає поживних речовин, або ж в разі надмірного нагромадження окремих
продуктів метаболізму [16].
За спритливих умов спори проростають.
Спочатку вони поглинають зі середовища значну кількість води, набрякають. Потім
відбуваються глибокі фізіологічні зміни, активуються ферментні системи,
інтенсифікується дихання. Ці процеси супроводжуються виділенням зі спори
амінокислот, дипіколінової кислоти, іонів кальцію, мономерів пептидоглікану (до
30 % маси спори). У цей час спори втрачають термостійкість. У проростаючій
спорі починається синтез РНК та білків, а через одну-дві години після початку
цього процесу - синтез ДНК. Руйнуються кортекс, оболонки спори, і ростова
трубка виходить назовні на кінці або збоку спори (полярне та екватооріальне
проростання) [9].
Здатність організмів до
спороутворення використовується в їх систематиці [19].
Додаток Д
Здатність до руху і характер
розташуванням джгутиків
Джгутики вперше описав Кон в 1872
році, а Р. Кох (1877) та Ф. Лефлер (1877) розробили методики їх фарбування
[22].
Кількість та розташування джгутиків
у різних бактерій неоднакова, окрім того вони притаманні не всім бактеріям.
Проте розташування джгутиків у певних видів є стабільним і часто
використовується як диференційна ознака [2]. Особливо важливим даний критерій
для виявлення паличкоподібних грамнегативних бактерій [9]. До того ж, кількість
джгутиків, їхня локалізації, діаметр, амплітуда спіралі генетично детерміновані
і є таксономічною ознакою для певних груп бактерій [18].
За кількістю та розміщенням
джгутиків мікроорганізми (рис. Г.1.) поділяють на:
· монотрихи:
Клітина має один джгутик, локалізований на одному з полюсів клітини (холерний
вібріон).
· Розташування
декількох (пучка) джгутиків на одному з полюсів клітини отримало назву
лофотрихального (паличка синьо-зеленого молока, Alcalignes faecalis). Пирог
Т.П. (2004), під лофотрихами тлумачить бактерії з монополярно-політрихальним
розміщенням джгутиків (Pseudomonas, Chromatium).
· Бактерії,
які містять один або декілька джгутиків на обох полюсах клітини, називають
амфітрихами (Spirillum volutants). Дані види бактерій мають біполярно -
політрихальне розміщення джгутиків.
· Якщо ж
джгутики розташовані по всій поверхні бактеріальної клітини, то такий тип
джгутикування називають перитрихальним (сальмонели черевного тифу, паратифів А
і В, ешерихії).

Рис. Г.1. Типи розташування
джгутиків у бактерій: 1- монотрихальне; 2 - лофотрихальне; 3 - латеральне; 4 -
амфітрихальне; 5 - перетрихальне; 6 - “зміщене” полярно-перетрихальне.
Наведена класифікація є умовною. За
допомогою електронної мікроскопії виявлено, що в деяких монотрихів джгутик
міститься не на кінці тіла, а в місці переходу бокової поверхні до полюса, а
амфітрихи - це дві клітини монотрихів, що не повністю поділилися, клітини із
джгутиками на дистальних кінцях [12]. Доведено, що бактерії, які раніше
вважалися монотрихами мають по кілька джгутиків. Самостійне існування
амфітрихів заперечується [13].
Найбільш поширеними є два типи
розташування джгутиків: полярний та перетрихальний; полярні джгутики слугують
для руху в рідких середовищах, а латеральні характерні для видів, середовищем
існування яких є волога тверда поверхня [22].
Рух бактерій вперше спостерігав
Левенгук в 1863 році. А Дж. Будер (1915) і П. Метцнер (1920) виявили
властивість джгутиків обертатись [13]. Саме розміщення джгутиків визначає
характер руху бактерій. Моно- і лофотрихи рухаються прямолінійно, перетрихи під
час прямолінійного руху безладно пересуватися [22]. В цілому, тип руху залежить
від кількості джгутиків, віку і властивостей культури, температури і наявності
хімічних речовин та інших факторів. Найбільшу рухливість мають монотрихи (60
мкм/с) [9].
Джгутики являють собою спірально
закручені нитки. У різних видів бактерій вони різняться за товщиною (12-18 нм),
довжиною до 20 мкм, а також за довжиною та амплітудою витка.
Хоч наявність джгутиків і є видовою
ознакою, однак вони не завжди життєво необхідні, є безджгутикові варіанти
рухливих видів бактерій [19].
У різних видів бактерій є ворсинки
(рис), які набагато коротші і тонші від джгутиків [19].
Поверхня деяких бактерій вкрита
великою кількістю (від 10 до кількох тисяч) довгих таких прямих ниток завтовшки
3-25 нм та завдовжки до 12 нм, які називаються фімбріями. Фімбрії зустрічаються
у бактерій, які мають і не мають джгутиків [16].
Пілі є поверхневими структурами
переважно грамнегативних бактерій. У E. coli вони мають форму порожнистих
циліндрів. Розрізняють загальні (ворсинки першого типу) та статеві пілі
(ворсинки другого типу). Загальні пілі розміщені по всій поверхні клітини, їхня
кількість становить від 50 до 400, а довжина до 1,5 мкм. Ці пілі зумовлюють
адгезивні властивості бактерій.[13] Статеві пілі, або пілі типу F, були
виявлені у клітин-донорів E. coli К 12, у штамів, що містять статевий фактор F
(F, Hfr) F пілі можуть бути тільки по одній або по дві на клітину. Також,
ворсинки другого типу характерні для нейсерій, гемофільних видів, стрептококів
групи А, коринебактерій та ін. [9].
У деяких паличкоподібних форм
поверхня клітин покрита виростами, шипами [13].
Додаток Е.1
Фарбування за Грамом
Фарбування мікроорганізмів -
складний фізико-хімічний процес, у механізмі якого значну роль відіграють явища
адсорбції, капілярності, хімічної спорідненості між барвником і об'єктом
фарбування [20].
Данський вчений Х. Грам у 1884 р.
Запропонував специфічний метод фарбування бактерій, згідно з яким фіксовані
клітини бактерій обробляють розчином барвника кристалічного фіолетового та
йодом. Фарбування за Грамом є важливою діагностичною ознакою бактерій..Слід
зауважити, що характер забарвлення прокаріот за Грамом залежить від віку
культури, факторів зовнішнього середовища тощо [9].
Донедавна розглядалися різні теорії,
що пояснюють диференційне забарвлення бактерій за Грамом (наприклад, хімічна,
мембранна, ізоелектрична). Згідно сучасних уявлень, після проникнення у клітини
розчинна хлорна форма генціан-віолету переходить у нерозчинну йодну форму і
випадає у осад, при цьому забарвлюється цитоплазма клітини. При обробці
препарату розчинником (етиловий спирт, ацетон) із цитоплазматичної мембрани
екстрагуються ліпіди, це призводить до підвищення її пористості. Таким чином,
мембрана не є перешкодою для вимивання комплексу генціан-віолет-йод. Але
зазвичай клітина має муреїновий шар (різної товщини та пористості у різних
бактерій), який характеризується високою стійкістю до органічних розчинників.
Багатошаровий малопористий муреїновий шар перешкоджає вимиванню барвника -
клітини забарвлюються у синьо-фіолетовий колір (грампозитивно), а при наявності
моношару муреїну із крупними порами, клітини забарвлюються за Грамом -
негативно [18].
Отже, фарбування мікроорганізмів має
велике диференціальне значення оскільки дає змогу встановити морфологічні і
тинкторіальні властивості мікроорганізмів, тобто їхнє відношення до барвників.
Додаток Е.2
Результати фарбування за Грамом
Рис. Е.2.1. Мікрофлора повітря
аудиторії № 21
Рис. Е.2.2. Мікрофлора повітря
аудиторії № 21
Рис. Е.2.3. Мікрофлора повітря
аудиторії № 21
Рис. Е.2.4. Мікрофлора повітря
аудиторії № 21
Рис. Е.2.5. Мікрофлора повітря
аудиторії № 2

Рис. Е.2.6. Мікрофлора повітря
аудиторії № 21
Рис. Е.2.7 Мікрофлора
повітря аудиторії № 21
Рис. Е.2.8 Мікрофлора повітря
аудиторії № 21

Рис. Е.2.9. Мікрофлора
повітря аудиторії № 21
Рис. Е.2.10. Мікрофлора повітря
аудиторії № 21
Рис. Е.2.11 Мікрофлора повітря
аудиторії № 21
Рис. Е.2.12. Мікрофлора повітря
туалету
Рис. Е.2.13 Мікрофлора
повітря туалету
Мал. Е.2.14 Мікрофлора
повітря туалету
Додаток Ж
Механізм визначення розмірів клітин
мікроорганізмів
Клітини мікроорганізмів виміряють
під мікроскопом за допомогою окулярної лінійки - мікрометра чи окулярного
гвинтового мікрометра. Для вимірювання краще використовувати живі, а не
фіксовані клітини, так як фіксація і забарвлення клітин призводить до деяких
змін справжніх розмірів. Якщо клітини здатні до руху, препарат необхідно трохи
підігріти або ж до краплини досліджуваної суспензії додають 1 мл 0,1 % - ного
водного розчину агару. Розміри клітин визначають у мікрометрах (мкм) [18].
Механізм визначення форми клітини
Форму клітин виявляють, як правило,
на препаратах “роздавлена крапля” в світловому чи фазово-контрастному
мікроскопі. Для визначення форми клітини дрібних паличкоподібних бактерій
таких, як Serratia marcescens, готують препарат фіксованих клітин і фарбують їх
простим способом. Клітини бактерій, які мають малі розміри і мають вирости
(роди Stella, Caulobacter, Prosthecomicrobium) необхідно досліджувати в темному
полі. Розташування клітин в колонії актиноміцетів і міцеліальних грибів
вивчають методом відбитків [18].
Механізм виявлення спор
мікроорганізмів
Спори характеризуються значною
заломлюваністю світла. Вони мають такий самий показник заломлювання світла як і
зневоднений білок. Це вказує на те, що в спорах велика кількість багатого
білком матеріалу зосереджена в малому об'ємі [16]. Спора містить майже всю суху
речовину материнської клітини, але займає в 10 разів менший об'єм. При
бактеріологічному дослідження у незабарвленому стані мають вигляд блискучих
зерен; вони погано забарвлюються [19].
У сумнівних випадках виявити в
клітинах істинні ендоспори можна за допомогою спеціального фарбування.
Оболонка мікроорганізмів стійка до
дії більшості хімічних речовин, тому погано фарбується [16]. Спори важко
взаємодіють з барвниками, тому за простих методів фарбування вони залишаються
безбарвними на тлі забарвлених вегетативних тіл бактерій [23] .
Для зафарбовування спор важливою
умовою є спеціальне оброблення мазка, що забезпечує розм'якшення оболонки. Його
протравлюють кислотою, потім фарбують концентрованим розчином барвника при
високій температурі. Наприклад, карболовим розчином фуксину. При цьому, як
правило, оболонка спори міцно зв'язують барвник і не знебарвлюються, навіть при
обробці етанолом чи 1М оцтовою кислотою (в умовах, коли решта клітини стає
безбарвною) [21, 23].
Механізм виявлення джгутиків
Виявлення джгутиків є однією з
найважчих процедур у бактеріологічній техніці. Це пов'язано з тим, що джгутики
можна дуже легко пошкодити при виготовленні мікропрепарату. Розглянути джгутик
(пучок джгутиків) у світловому мікроскопі чи в умовах фазового контрасту
вдається тільки у небагатьох представників родів Bdellovibrio, Thiospirrilum,
Chromatium. Легко виявити джгутики шляхом нанесення на них барвників або
металу, а також за допомогою електронного мікроскопа [9].
Наявність джгутиків виявляють
визначенням рухливості бактерій, мікроскопіюванням у темному полі,
застосуванням особливих методів обробки їх протравами, адбсорбції на поверхні
клітин бактерій різних барвників, а також за допомогою електронної мікроскопії.
Останній дав змогу виявити спіралеподібну форму і гвинтоподібну будову
джгутиків [19].
Скло для мікропрепаратів повинно
бути абсолютно чистим і знежиреним. Зазвичай, використовують нові покривні
скельця. Дуже важливо, щоб вік досліджуваної культури був не старшим від 12-18
год. Якщо скло абсолютно чисте, то краплина з мікроорганізмами набуде
правильної округлої форми, легко пошириться і швидко висохне на повітрі без
підігріву. Препарати перед фарбуванням обов'язково фіксують протравою, яка
посилить інтенсивність забарвлення.
Прикладом методик виявлення
джгутиків є метод Леффлера. В даному випадку джгутики фарбуються у рожево-червоний
колір. При використанні метода Шнека джгутики набувають синього забарвлення
[21].
Особливості механізму виявлення
спороутворення, способу руху клітин мікроорганізмів
Процеси спороутворення, вивчення
характеру руху клітини проводять на живих клітинах мікроорганізмів,
використовуючи “роздавлену” та “висячу краплю”, також спори виявляють шляхом
диференційного фарбування цитоплазми і спор. Висяча крапля широко
використовується для визначення проростання спор у вегетативних форм [21]. Слід
зазначити, що у практичних лабораторіях вивчення мікроорганізмів у живому стані
використовують для дослідження їхньої рухливості як непрямого підтвердження
наявності в них джгутиків [ 23].
Рухливість бактерій вивчають
методами висячої чи роздавленої краплі. Визначення рухливості використовують у
лабораторній практиці для ідентифікації холерних вібріонів, бактерій черевного
тифу, паратифів та ін.
Досить часто рухливість
мікроорганізмів можна виявити неозброєним оком. Для цього досліджувану культуру
сіють уколом у стовпчик напіврідкого живильного середовища. Посів інкубують у
термостаті протягом 18-20 годин. Якщо бактерії не мають джгутиків, їх ріст
(інтенсивне помутніння) буде тільки впродовж лінії уколу. Рухливі бактерії
дають дифузний ріст по всій товщині живильного середовища.
Якщо для встановлення характеру руху
клітини використовують “висячу краплю”, то в імерсійному об'єктиві можна
спостерігати наступне: рухливі бактерії проходять з однаковою швидкістю значну
відстань, часом через усе поле зору, роблячи кругові і ґвинтові рухи. Найбільш
швидкі і прямолінійні рухи роблять монотрихи і лофотрихи. Перитрихам і
амфітрихам властива менш енергійна і безладна рухливість.
Варто зазначити, що початківець може
помилково прийняти молекулярний рух за справжній. При цьому нерухливі бактерії
коливаються між двома близькими точками, ніби “танцюють” на місці.
Для збільшення контрастності
досліджуваних об'єктів використовують малотоксичні й майже нешкідливі барвники:
метиленовий і толуоїдний синій, конго і нейтральний червоний, акридиновий
оранжевий, янус зелений [17].
Однак ці методи мають свої недоліки.
У живих бактерій, що активно рухаються, важко виявити деталі структури. При
такому дослідженні можна мати лише загальне уявлення про їх морфологію.
Отже, у випадку виявлення у
мікроорганізмів здатності до утворення спор необхідно звернути увагу, тип
спороутворення, на розташування спор в клітині, форму вільних спор та визначити
їхні розміри [18].
Слід зауважити, що вік культури,
склад середовища і умови культивування суттєво впливають на морфологію
мікроорганізмів.
Додаток З
Обрахунки кількісного складу
мікрофлори повітря
1. Результати посівів
мікроорганізмів взятих з повітря аудиторії № 21:
· До початку
занять ( температура складала)
На площі чашки Петрі (78,5 )
за 48 год утворилося 3 колонії. Відповідно на площі 100 колоній буде більше:
,5 - 3 колонії,
- х колоній,
Перерахунок кількості колоній
на площу 100 доцільно проводити тому, що за приблизними даними В. Л.
Омелянського на цю площу за площу за 5 хв осідають усі мікроби та їхні спори,
які містяться в 10 л повітря. Цей показник дає можливість перерахувати
кількість мікроорганізмів у повітря досліджуваного приміщення:
колонії - 10 л
х - 1000 л,
Отже, в повітря аудиторії №
21 міститься 382 мікробних тілець та їхніх спор.
Після завершення занять (
температура складала)
На площі чашки Петрі (78,5 )
за 48 год. утворилося 10 колоній. Відповідно на площі 100 колоній буде більше:
,5 - 10 колоній,
- х колоній,
Перерахунок кількості колоній
на площу 100 :
, 73 - 10 л
х - 1000 л,
Отже, в повітря аудиторії №
21 міститься 1273 мікробних тілець та їхніх спор.
Результати посівів
мікроорганізмів взяті з аудиторії № 18:
· До початку
занять ( температура складала)
На площі чашки Петрі (78,5 )
за 48 год. утворилося 4 колонії. Відповідно на площі 100 колоній буде більше:
,5 - 4 колонії,
- х колоній,
Перерахунок кількості колоній
на площу 100 :
,1 - 10 л
х - 1000 л,
Отже, в повітря аудиторії №
18 міститься 510 мікробних тілець та їхніх спор.
Результати посівів
мікроорганізмів взяті з коридору на другому поверсі :
· До початку
занять ( температура складала)
На площі чашки Петрі (78,5 )
за 48 год. утворилося 6 колоній. Відповідно на площі 100 колоній буде більше:
,5 - 6 колоній,
- х колоній,
Перерахунок кількості колоній
на площу 100 :
, 64 - 10 л
х - 1000 л,
Отже, в повітря коридору на
першому поверсі міститься 510 мікробних тілець та їхніх спор.
Після завершення занять (
температура складала)
На площі чашки Петрі (7,065 )
за 48 год. утворилося _ колоній. Відповідно на площі 100 колоній буде більше:
,065 - 2 колоній,
- х колоній,
Перерахунок кількості колоній
на площу 100 :
, 26 - 10 л
х - 1000 л,
Отже, в повітря туалету
міститься 4246 мікробних тілець та їхніх спор.
Результати посівів
мікроорганізмів взяті з коридору на першому поверсі:
· До початку
занять ( температура складала)
На площі чашки Петрі (78,5 )
за 48 год. утворилося 5колоній. Відповідно на площі 100 колоній буде більше:
,5 - 5 колоній,
- х колоній,
Перерахунок кількості колоній
на площу 100 :
, 37 - 10 л
х - 1000 л,
Отже, в повітря коридору на
першому поверсі міститься 637 мікробних тілець та їхніх спор.
Після завершення занять (
температура складала)
На площі чашки Петрі (7, 65 )
за 48 год. утворилося 1 колонія. Відповідно на площі 100 колоній буде більше:
,065 - 1 колонія,
- х колоній,
Перерахунок кількості колоній
на площу 100 :
, 15 - 10 л
х - 1000 л,
Отже, в повітря коридору
першого поверху після занять міститься 1415 мікробних тілець та їхніх спор.
Результати посівів
мікроорганізмів взяті з повітря живого куточку:
· Після завершення
занять температура складала).
На площі чашки Петрі (7,065 )
за 48 год. утворилося 2 колонії. Відповідно на площі 100 колоній буде більше:
,065 - 2 колоній,
- х колоній,
Перерахунок кількості колоній
на площу 100 :
, 3 - 10 л
х - 1000 л,
Отже, в повітря живого
куточку 2830 мікробних тілець та їхніх спор.
Додаток
И

Рис. И.1. Виготовлення поживного
середовища

Рис. И.2. Виготовлення поживного
середовища

Рис. И.3. Виготовлення поживного
середовища

Рис. И.4. Виготовлення поживного
середовища

Рис. И.5. Виготовлення поживного
середовища

Рис. И.6. Виготовлення поживного
середовища

Рис. И.7. Виготовлення
тимчасових мікропрепаратів

Рис. И.8. Виготовлення
тимчасових мікропрепаратів

Рис. И.9. Результати культивування
мікроорганізмів

Рис. И.10. Результати культивування
мікроорганізмів аудиторії № 21 (до занять)
Рис. И.11. Результати культивування
мікроорганізмів аудиторії №21 (після занять)

Рис. И. 12. Результати
культивування мікроорганізмів кор. другого поверху (до занять).