Мейобентос зарослей макрофитов прибрежной зоны Новороссийской бухты
МИНИСТЕРСТВО
ОБРАЗОВАНИЯ И НАУКИ РОССИЙСКОЙ ФЕДЕРАЦИИ
Федеральное
государственное бюджетное образовательное учреждение высшего профессионального
образования
«КУБАНСКИЙ
ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ»
Кафедра
водных биоресурсов и аквакультуры
ВЫПУСКНАЯ
КВАЛИФИКАЦИОННАЯ РАБОТА
МЕЙОБЕНТОС
ЗАРОСЛЕЙ МАКРОФИТОВ ПРИБРЕЖНОЙ ЗОНЫ НОВОРОССИЙСКОЙ БУХТЫ
Работу выполнила С.А. Сержанова
Научный руководитель А.Н.
Пашков
Краснодар
2015
Реферат
Ключевые слова: МЕЙОБЕНТОС, ЗАРОСЛИ МАКРОФИТОВ,
БИОМАССА, ЧИСЛЕННОСТЬ, ЧЁРНОЕ МОРЕ, НОВОРОССИЙСКАЯ БУХТА, ТАКСОНОМИЧЕСКИЙ
СОСТАВ.
Целью данной работы являлось изучение
мейобентоса зарослей макрофитов прибрежной зоны Новороссийской бухты.
В ходе работы были описаны таксономический
состав, численность и биомасса мейобентоса в некоторых местах Новороссийской
бухты. Проведено сравнение мейобентоса по указанным показателям друг с другом.
Изучена динамика состояния мейобентоса прибрежной зоны Новороссийской бухты в
2014 г. в сравнении с данными предыдущих лет.
Содержание
Введение
. Аналитический
обзор
.1 Общая
характеристика мейобентоса
.2 Изучение
населения зарослей морских макрофитов
.3 История
развития и накопления знаний о мейобентосных организмах
.4 Распределение
мейобентоса
.5 Трофические
связи в сообществе мейобентоса
.6 Таксономический
состав мейобентоса. Основные группы мейобентосных организмов
. Физико-географическая
и гидрохимическая характеристика Новороссийской бухты
. Материал
и методы исследования
. Мейобентос
зарослей макрофитов прибрежной зоны Новороссийской бухты
.1 Таксономический
состав мейобентоса Новороссийской бухты
.2 Пространственная
изменчивость численности и биомассы мейобентоса Новороссийской бухты
.3 Временная
динамика численности и биомассы мейобентоса Новороссийской бухты
Заключение
Библиографический
список
Введение
Акватория Новороссийской бухты подвержена
сильному загрязнению. Наибольший ущерб экологии бухты наносят нефтяные выбросы,
поскольку Новороссийск является крупнейшим портом страны, да к тому же через
его терминалы на экспорт отправляется треть всего добываемого в России черного
золота. Помимо этого в гораздо большей степени акваторию загрязняют
промышленные и бытовые канализационные стоки самого города. Следствием этого
является обвальное снижение биомассы водорослей, морских животных, а также
гибель видов с узким коридором толерантности к интоксикации нефтью (таковых
видов в экосистемах Черного моря большинство) [Вершинин, 2003].
Термин «мейобентос» был введен в науку М.Мэр в
середине ΧΧ века для
обозначения совокупности микроскопических многоклеточных, выпадающих из
внимания исследователей морского бентоса при количественных учетах донной фауны
[Мокиевский, 2009].
Многолетние систематические наблюдения в
акваториях, наиболее подверженных прямому и косвенному воздействию различных
видов антропогенной деятельности, которая негативно сказывалась на
экологическом состоянии морских экосистем, установили, что видовое богатство,
плотность населения и показатели биомассы мейобентоса, а также соотношение
некоторых количественных характеристик может успешно применяться для оценки
экологического состояния бентали [Сергеева, Колесникова, 1996]. Ценность
использования качественных и количественных показателей мейобентоса заключается
также в возможности осуществления кратко- и долгосрочных прогнозов формирования
запасов не только кормового макрозообентоса, но и промысловых рыб придонного и
донного комплексов [Киселев, 1965].
Можно предположить, что в ближайшие годы
исследование морского мейобентоса и его отдельных групп будет оставаться одним
из наиболее актуальных направлений биоокеанологии, наблюдающийся в последние
годы «взрывной» рост числа исследований и публикаций заставляет ожидать
существенного приращения знаний об этом, все еще загадочном, компоненте
морского бентоса [Сергеева, Колесникова, 1996].
Целью данной работы является изучение
мейобентоса зарослей макрофитов прибрежной зоны Новороссийской бухты.
Для достижения этой цели были поставлены
следующие задачи:
. Определить таксономический состав
мейобентоса Новороссийской бухты;
. Сравнить таксономический состав
мейобентоса с разных станций отбора проб;
. Изучить структуру мейобентосного
сообщества в разных местах бухты;
. Изучить временную динамику
таксономического состава мейобентоса Новороссийской бухты по результатам
исследований 2003, 2008 и 2014 гг.;
. Изучить пространственную изменчивость
численности и биомассы мейобентоса Новороссийской бухты;
. Изучить временную динамику численности
и биомассы мейобентоса Новороссийской бухты.
1. Аналитический обзор
.1 Общая характеристика мейобентоса
Термин «мейобентос» был введен в науку М. Мэр в
1942 г. для обозначения совокупности микроскопических многоклеточных,
выпадающих из внимания исследователей морского бентоса при количественных
учетах донной фауны. Основанием для такого выделения послужили линейные размеры
организмов, а также их способность проходить при промывании зообентосных проб
через ячею 1×1 мм. Таксономические
исследования отдельных групп и видов мейобентосных организмов начались еще
раньше [Киселев, Славина, 1965]. Через 27 лет после выхода работы М. Ф. Мэр на
международной конференции в Тунисе учеными различных стран была принята ее
классификация [Воробьева, 1999].
Ни таксономический, ни размерный критерии не
позволяют провести однозначную границу меду макро- и мейобентосом. Таким
образом, можно более или менее формально определить мейобентос как совокупность
микроскопических организмов, образующих самостоятельный размерный,
таксономический и функциональный блок донной экосистемы [Мокиевский, 2012].
Критериями для выделения мейобентоса в качестве самостоятельной экологической
группы может служить таксономический состав, средний размер особей и набор
морфологических и физиологических адаптаций [Киселев 1965].
Выделение мейобентоса в качестве самостоятельной
экологической единицы носит, как и всякая классификация, сугубо утилитарный
характер: исследования мейобентоса требуют специфических технических приемов и
методов, специальной таксономической подготовки исследователей и иной, чем для
макробентоса, схемы планирования процесса сбора материала, исходя из специфики
пространственно-временных масштабов существования мейобентосных сообществ и
популяций [Гусева, Алемов, Кирюхина, 1998].
За последние десятилетия произошли заметные
изменения в направлениях исследований. К концу 1990-х гг. был накоплен
значительный материал по видовому составу и пространственной структуре
сообществ литорали и верхней сублиторали, выявлены основные закономерности
суточной и сезонной динамики, получены данные о питании различных групп
мейобентоса и сделаны первые попытки оценить роль мейобентоса в потоках энергии
через донные экосистемы [Мокиевский, 2009].В последние годы на первое место
вышли другие темы, вызывающие наибольший интерес исследователей:
.Изучение глубоководного мейобентоса в различных
аспектах - от описания таксономического состава основных групп до изучения
сезонной и многолетней динамики сообществ, исследования мелкомасштабной
пространственной структуры сообществ и роли различных факторов, как
биотических, так и абиотических, в ее поддержании.
.Исследование закономерностей видового
разнообразия в разных масштабах: от анализа многовидовых комплексов в пределах
квадратных метров и дециметров, до поиска закономерностей изменения видового
богатства на широтных градиентах в масштабе океанов.
.Составление обобщающих баз данных по различным
группам и регионам дает возможность перейти от частных исследований к более
широким обобщениям [Сергеева, Колесникова, 1996].
.2 Изучение населения зарослей морских
макрофитов
Впервые население зарослей макрофитов было
выделено в самостоятельные группировки в работе С. А. Зернова, посвященной
донным биоценозам Черного моря в 1913 г. Он выделил в самостоятельные биоценозы
население зарослей филлофоры и зоостеры, а население цистозиры включил в
биоценоз скал [Колесникова, Шадрин, 1981]. В 1927 г. В. А. Водяницкий обратил
внимание на высокую численность мелких организмов в зарослях цистозиры и
произвел количественный учет населения куста этих водорослей в Новороссийской
бухте. Он положил начало количественному изучению зарослей цистозиры
[Миловидова, 1969].
В работах Л. В. Арнольди и В. Н. Никитина о
населении цистозиры содержится сравнительно мало сведений, в то же время рыхлым
грунтам посвящены фундаментальные исследования. Л. В. Арнольди в 1949 г.,
описывая зообентос Каркинитского залива, приводит краткий список видов,
встречающихся в зарослях цистозиры [Афанасьев, Корпакова, 2008].
В 1952 г. И. В. Шаронов, описывая биоценоз скал
и каменистых россыпей приводит количественный данные и оп фауне цистозиры.
Однако, не выделяя сообщество зарослей цистозиры в самостоятельное, он считает
руководящим видом митилястер, «щетки» которого покрывают поверхность скал, а на
водорослях встречается только его молодь [Миловидова, 1969].
Сообщества беспозвоночных животных, обитающих в
зарослях макрофитов, обладают рядом общих (специфических) свойств, что
позволяет выделить их в группу «зарослевых сообществ» [Заика, Киселев,
Михайлова, 1992]. Приспособленность к существованию на макрофитах у
беспозвоночных животных определяется наличием специальных органов прикрепления.
Как правило, это железы, выделяющие слизистый секрет, которым животные либо
непосредственно прикрепляются к макрофитам (остракоды, нематоды, турбеллярии,
клещи и моллюски), либо склеивают из обрывков водорослей и детрита трубковидные
домики (бокоплавы и полихеты) [Маккавеева, 1966].
Нет ни одного вида беспозвоночных животных,
который был бы характерен только для одного макрофита.
В 1953 г. Маккавеева начала изучать макро- и
мейоэпифитон цистозиры в районе Севастополя на глубине до 1 м. Ею впервые были
проведены круглогодичные наблюдения, что позволило выявить сезонную динамику
численности и биомассы массовых видов и трофическую структуру этого сообщества
[Маккавеева, 1959].
Населению цистозиры у Западного побережья
Черного моря посвящено несколько работ румынских (Мюллер, Баческу, Бодеани,
Тиганус) и болгарских (Абаджиева) авторов [Макавеева 1966].
.3 История развития и накопления знаний о
мейобентосных организмах
макрофит мейобентос бухта биомасса
В середине XX
столетия трудно было предположить, что не изучен целый комплекс водных
организмов, представители которых есть практически во всех типах беспозвоночных
животных. По мере накопления сведений о них (богатому видовому разнообразию,
высокой плотности поселений, биологическим и экологическим особенностям) стало
ясно, что данная категория донных животных играет большую роль в жизни
природных водных экосистем [Заика, 1992]. В настоящее время раскрываются все
новые стороны значения мелких донных животных - мейобентоса - в жизни водоемов
[Воробьева, 1999].
Началом исследований мейофауны как
самостоятельной экологической группы принято считать работы Ремане в Кильской
бухте. Изучая фауну крупных песков литорали, Ремане впервые обнаружил, что
капилярные пространства между частицами песка представляют собой очень
своеобразный и богато населенный биотоп, обладающий рядом специфических черт,
требующих от его обитателей специальных адаптаций, сходных у представителей
разных таксономических групп [Мокиевский, 2012].
Для обозначения обитателей капилляров морского
песка - интерстициальных пространств, Николе в 1935 г. ввел термин
«интерстициальная фауна». Дальнейшие исследования морских и пресноводных
биотопов интерстициали позволило выявить основные морфологические адаптации
организмов:
мелкие размеры;
удлиненная (червеобразная) форма тела;
сильное развитие прикрепительных органов -
щетинок, присосок, крючков или желез, выделяющих клейкий секрет [Маккавеева,
1966].
Сходные морфологические черты проявляются в
самых разных таксономических группах. Так, удлиненная, червеобразная форма тела
характерна для интерстициальных представителей таких далеких таксономических
групп, как гарпактициды и турбеллярии [Маккавеева, 1966].
Позже Я. Тиль предложил термин «нанобентос» для
организмов размером 2-42 мкм, однако, этот термин получил распространение
только в глубоководных исследованиях и может быть встречен в относительно
небольшом числе публикаций [Киселев, 1965]. Для первых исследователей
удивительной и разнообразной микроскопической фауны внешне безжизненных
песчаных пляжей ее экологическая самостоятельность, непохожесть на известные
сообщества макробентоса, не вызывала сомнений. Неслучайно, первые же
исследователи этих биотопов предложили для их населения сразу несколько
специальных терминов [Киселев, 1985]. На протяжении следующих десятилетий
самостоятельность мейобентоса принималась большинством гидробиологов, но,
скорее, как удобный методический прием при исследовании бентоса. К обоснованию
мейобентоса как самостоятельной экологической категории Р. Варвик вернулся в
1984 г. Он сделал попытку сопоставить основные биологические характеристики
мейо- и макробентоса и подчеркнуть своеобразие каждой из этих групп
[Шереметьевский, 1991].
Этот анализ выявил с одной стороны,
существование заметных биологических различий между двумя группами
многоклеточных, но, с другой стороны, показал, что границы между ними
достаточно условны, и по каждому из критериев есть пересечение
признаков[Шереметьевский, 1991].
Взгляды разных исследователей на таксономический
объем мейобентоса сильно расходятся. Р. Хиггинс и Я. Тиль в число
рассматриваемых групп мейобентоса включают простейших - инфузорий и
фораминифер. Этой же точки зрения придерживается. Однако, при введении термина
«мейобентос» в 1942 г. М. Мэр четко и однозначно отделила его от микробентоса
(к которому относила инфузорий). Микробентос отличается от мейобентоса по всем
наиболее важным экологическим характеристикам, в том числе размерами особей и
временем оборота генераций, а также пространственными масштабами организации
сообществ [Миловидова, 1967].
Накопление знаний о морфологии, систематике и
распространении микроскопических многоклеточных позволяют сделать вывод о том,
что выделение мейобентоса, предложенное как технический прием, облегчающий
работу с бентосными пробами, имеет более глубокий биологический смысл. По
совокупности таксономических, морфологических и экологических характеристик
мейобентос представляет собой отдельный структурный блок донной экосистемы,
обладающий рядом уникальных черт [Киселев, 1985].
.4 Распределение мейобентоса
Работ, описывающих закономерности
пространственного размещения мейобентосных организмов, достаточно много - в
последние десятилетия это одно из самых популярных направлений в исследованиях.
Все работы можно разделить на две группы: пространственное распределение обилия
крупных таксономических групп в разных масштабах или более тонкое изучение
пространственной мозаики одного из крупных таксонов [Миловидова, 1967].
Трудности таксономической обработки материала - одна из основных причин, по
которой под изучением пространственной структуры сообществ понимается
исследование отдельных таксоценов [Киселев, 1985]. Наибольшее количество
исследований пространственного размещения мейобентосных организмов в разных
масштабах выполнено на литорали [Мокиевский, 2009]. На формирование
пространственной структуры влияют как биотические, таки абиотические факторы.
Наиболее благоприятные условия для существования большинства представителей
мейо- и макрозообентоса складываются в глубоководных участках бентали [Еремеев,
Гаевская, 2003].
Мейобентос по глубинам распределен неравномерно.
Как правило, наибольшие количественные характеристики приурочены к
псевдолиторали и верхней сублиторали, и прежде всего к зоне фиталя. Эта же зона
наиболее богата по таксономическому составу фауны [Колесникова, 1991].
.5 Трофические связи в сообществе мейобентоса
Несмотря на малые размеры, представители
мейобентоса обладают очень большой численностью. Высокая плотность поселений -
одна из важных их особенностей. Достигая численности свыше 5 ×
106 экз./м2,
они обладают высокой плодовитостью, в основном коротким жизненным циклом,
быстрым темпом роста, высокой интенсивностью метаболизма[Воробьева, 1999].
Организмы из состава мейобентоса играют важную роль в поддержании баланса
органических веществ в донных осадках, т. к. через них проходит значительная
доля всего энергопотока [Мокиевский, Малых, 2002].
На ухудшение условий существования мейобентос
отвечает сокращением одних групп, доминированием двух - трех групп и повышением
плотности организмов с минимальными размерами тела, обладающими, как правило,
короткими циклами развития [Мейобентос в санитарно-биологическом аспекте,
1995].
В процессе жизнедеятельности представители
мейобентоса, кроме детрита и диатомовых водорослей, потребляют значительное
количество бактериальной и инфузориальной продукции. Мейобентос также в
значительной мере существует за счет жизнедеятельности зоопланктона и
макрозообентоса [Маккавеева, 1967].
Представители микро-, мейо- и макрозообентоса
образуют сложные трофические связи, включающие питание не только мертвым
органическим веществом и продуктами жизнедеятельности гидробионтов из состава
других комплексов, но и живым органическим веществом.
Во всех сообществах мейобентосных животных
непрерывно осуществляется сложный механизм перестроек, обусловленный сезонными
изменениями в количественных и качественных характеристиках представителей
самого мейобентоса, их пищевых компонентов, макрозообентоса и ихтиофауны
[Колесникова, Мазгумян, 2002].
.6 Таксономический состав мейобентоса. Основные
группы мейобентосных организмов
В состав мейобентоса входят представители
практических всех известных типов беспозвоночных, но относительное число
микроскопических видов в разных таксонах различно.
Некоторые типы представлены исключительно видами
мейобентосных размеров, в большинстве же типов известно лишь небольшое число
мелких видов.
Степень изученности биологии различных
мейобентосных групп варьирует в широких пределах.
Наиболее подробно изучены самые массовые группы
- нематоды, гарпактициды, остракоды.
Большое число исследований посвящено таксономии,
строению и образу жизни этих организмов. Сведения по биологии большинства
редких и малочисленных групп скудны и отрывочны.
Для некоторых таксонов известен лишь факт
нахождения микроскопических представителей, сопровождающийся иногда кратким
очерком анатомии и морфологии [Воробьева, 1999]. Основные виды представителей
мейобентоса представлены в таблице 1.
Таблица 1 - Основные представители мейобентоса
(Е. Б. Макавеева [1967])
|
Отряд
|
Виды
|
|
Coetenterata
|
Agtaopheniapluma Plumutanalincoi
Lucemaria campanula
|
|
Polychaeta
|
Fabriciasabelta Grubealimbata
Harmothoe reticulate Nereiszonata Platynereisdumerilii Spirorbismolitaris
Spirorbispusilta
|
|
Loricata
|
Acanthohitonafascicularis
Lepidochitonacinerea
|
|
Gastropoda
|
Gibbulaadriatica Nana donovani
Nana neritea Rissoasplendida Rissoamembranacea
|
|
Bivalvia
|
Mytilasterlieatus
Mytilusgallaprovincialis
|
|
Cirripedia
|
Balanusimprovisus
|
|
Cumacea
|
Cumacea sp.
|
|
Tanaidacea
|
Lepcocheliasavignyi
|
|
Isopoda
|
Idoteaostroumovi Sphaeromaserratum
Synisomacapito
|
|
Amphipoda
|
Apherusabispinosa
Biancolinacuniculus Hyaledolffusi Hyaleperieri Hyalepontica
Stenothoemonoculoides
|
|
Decapoda
|
Decapod asp. Decapodazoea
Hippolitelongirostris
|
|
Chironomida
|
Chironomidae larvae
|
|
Bryozoa
|
Lepraliapallasiana
|
|
Tunicata
|
Diplosomalisterianum
|
По суммарному числу видов мейобентос ненамного
уступает макроскопическим беспозвоночным. Приблизительный расчет на основании
списков видов для морей показывает, что число известных видов мейобентоса
составляет около 25% от видового разнообразия бентоса. Доля мейобентоса от
общего числа видов свободноживущих беспозвоночных в наиболее хорошо изученных
морях (Белое, Баренцево) равна 16-20 %, но падает до 5-6 % в морях Сибири, что
связано со слабой изученностью мейофауны. Локальное разнообразие мейофауны
может быть сопоставимо и даже превышать разнообразие макробентосных видов
[Мокиевский, 2009].
Чутко реагируя на изменения условий обитания,
мейобентос в последние десятилетия также претерпел изменения в своем составе,
численности и биомассе. Вероятно, эти перемены должны были отразиться на
биологии связанных с мейобентосом организмов, в том числе и на естественном
воспроизводстве донных рыб [Заика, Киселев, Михайлова, 1992].
Обилие и фаунистическое разнообразие эпифитона
больше всего зависит от вида водоросли, на которой обитают животные в биотопе.
Основными зарослеобразующими макрофитами Новороссийской бухты являются
Cystoseira barbata и Cystoseira crinita. Таксономический и количественный сосав
мейобентоса определялся в зарослях Cystoseirabarbata [Колесникова, 1981].
Заросли цистозиры, произрастая на прибрежной
полосе скал и валунов, занимают значительные площади шельфа северо-восточной
части Черного моря. Они служат субстратом, пищей и убежищем для многочисленных
беспозвоночных, в том числе мейобентоса [Миловидова1969]. Известно, что обилие
и фаунистическое разнообразие мейоэпифитона больше всего зависят от вида
водоросли, на которой обитают животные, а также от степени прибойности и
интенсивности водообмена в биотопе. Поэтому зарослевые биоценозы открытого
прибрежья, по сравнению с защищенными участками моря, имеют ряд особенностей.
На макрофитах, растущих в открытых незащищенных от волнового воздействия местах
прибрежья, показатели численности мейобентоса могут быть ниже на порядок, чем
на тех же видах водорослей защищенных бухт [Ремизова, 2008].
Весной в мейобентосе обнаружено 16
таксономических групп, среди которых к эумейобентосу относятся: Turbellaria,
Nematoda, Harpacticoida, Ostracoda, Foraminifera, Aearina к
псевдомейобентосу-Polychaeta, Oligochaeta, Cumacea, Anisopoda, Isopoda,
Amphipoda, Loricata, MytilidaeBivalvia, Gastropoda, Chironomidae (Insecta)
[Ремизова, 2008].
Мейобентос Черного моря, как объект изучения,
представляет особый интерес. Во-первых, это связано с тем, что фауна моря имеет
различное происхождение, отражающее геологическое прошлое водоема [Гусева,
1995]. В составе бентических форм (включая мейобентос) встречаются понтические
(каспийские) реликты, виды бореально-атлантического происхождения,
средиземноморские иммигранты и, наконец, экзотические виды, случайно завезенные
судами из других районов Мирового океана [Зайцев, Александров, Миничева, 2006].
Во-вторых, в силу ряда природных и антропогенных причин (высокая изоляция от
Мирового океана, обширная водосборная площадь, замедленное вертикальное
перемешивание водных масс, индустриализация прибрежной зоны, эвтрофирование и
т. д.) в Черном море произошли глубокие биотопические изменения, отразившиеся
на их обитателях. В последние годы этот аспект привлек к себе внимание не
только местных специалистов, но и международных организаций, включая структуры
ООН [Александров, 2004].
2. Физико-географическая и гидрохимическая
характеристика Новороссийской бухты
Для Черного моря, особенно Кавказского
побережья, характерна небольшая изрезанность берегов.
Новороссийская бухта образовалась во время
вторжения моря в сушу при разломе и опускании одного из прибрежных горных
хребтов. Она представляет собой естественный постоянный водоём, заполненный
морской водой [Тильба, Мнацеканов, Крутолапов, 2006]. Его западные берега не
образуют поднятий, и протянувшаяся вдоль береговой полосы Суджукская коса
сформировала небольшую лагуну с мелководными участками. Восточные берега бухты
представляют собой высокие каменно-земляные обрывы с узкой пляжной полосой.
Внутренняя часть бухты обрамлена набережными, к которым подступают городские
кварталы. Угодье является крупным морским портом, снабжено выдающимися далеко в
море молами, пристанями [Маккавеева, 1979].
Новороссийская бухта, известная также под
названием Суджукской (по названию турецкой крепости Суджук-Кале) и Цемесской
(по названию впадающей в нее реки Цемесс), является самой обширной и
глубоководной на Кавказском побережье. Карта-схема Новороссийской бухты
представлена на рисунке 1. Длина бухты 15 км, ширина у входа 9 км, в средней
части 4,6 км. [Харитонова, 1981]. Береговая полоса почти не изрезана. Восточный
и западный берега значительно отличаются друг от друга. Восточный берег ограничен
сплошным хребтом гор (хребет Мархот), круто спускающимся к бухте отвесными
утесами. Этот берег разделен мысами Шесхарис и Пинай, примерно, на три равные
части. Западный берег представляет собой невысокую, полого спускающуюся к морю
холмистую местность [Маккавеева, 1979].
Рельеф дна бухты ровный, с преобладающими
глубинами 21-27 м. У берега глубина резко уменьшается.
Расположение грунтов типичное для Черного моря.
Вдоль берегов идет полоса скал, расширяющаяся по мере продвижения выхода из
бухты на глубине от 5 до 25 м. На глубинах 5-15 м. и 19-25 м. лежит слой
крупного ракушечника или песка [Харитонова, 1981].
По гидрологическому режиму Новороссийская бухта
делится на три района: порт, средняя часть бухты и район выхода из бухты
[Харитонова, 1981].
Порт - отгороженная молами вершина бухты. Для
него характерна мелководность (10-12 м.) и значительная удаленность от вод
открытого моря, это является одной из причин сильной загрязненности портовой
части [Макавеева, 1979].
Средняя часть располагается от моловых
сооружений до мыса Пенай. Максимальная глубина 25 м. Для той части бухты
характерно широкое сообщение вод с открытым морем. Загрязнение городскими
стоками здесь меньше, но все же значительное [Харитонова, 1981].
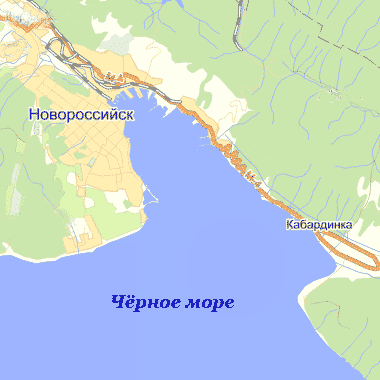
Рисунок 1 - Карта-схема Новороссийской бухты
Часть бухты, расположенная у выхода, имеет
глубину до 30- 35 м. Гидрологический режим определяется гидрологическим режимом
прибрежной полосы северо-восточной части Черного моря, отличный от условий
открытого моря [Тильба, Мнацеканов, Крутолапов, 2006].
К концу ҲҲ столетия экологическое
состояние Черного моря учеными и специалистами было определено как кризисное.
Это не могло не вызвать обеспокоенность мирового сообщества и проживающих на
его берегах народов.
К основным экологическим проблемам Черного моря
на данный момент относят:
. эвтрофикация (цветение воды). Из-за попадания
в прибрежные акватории остатков химических удобрений и плохо очищенных сточных
вод;
. загрязнение воды нефтепродуктами и другими
токсическими веществами;
. биологическое загрязнение чужеродными видами
(рапана, гребневик);
. уменьшение биологического разнообразия,
выраженное в снижении численности или полном исчезновении черноморских видов
флоры и фауны;
. сокращение площади природных ландшафтов,
лесных и особо охраняемых территорий в прибрежных зонах;
. накопление твердых бытовых отходов [Долинский,
Патлатюк,2013].
Площадь поверхности Черного моря в шесть раз
меньше площади его водосбора. По этой причине, состояние Черного моря сильно
зависит от хозяйственной деятельности на суше, как прибрежных, так и других
государств его бассейна. На качество прибрежных вод Черного моря влияют
береговые источники антропогенного загрязнения и в первую очередь большие
населенные пункты на побережье [Глумов, Кочеткова,1966]. Вследствие
деятельности объектов коммунального хозяйства населенных пунктов в морские воды
поступает значительная часть загрязняющих веществ. В последнее время в связи с
усилением природоохранной деятельности, снижения объемов сбросов и улучшением
качества очистки сточных вод в регионе наметилась четкая тенденция по снижению
поступления загрязняющих веществ в море. К сожалению, мер, которые
предпринимают черноморские государства недостаточно [Вершинин, 2003].
Одна из самых серьезных экологических проблем
Черного моря заключается в том, что впадающие в него реки несут с собой не
только пестициды или тяжелые металлы, но и азот и фосфор, в больших количествах
смывающиеся с полей.
Основным международным документом, регулирующим
вопросы охраны Черного моря, является Конвенция о защите Черного моря от
загрязнения, подписанная шестью черноморскими странами - Болгарией, Грузией,
Россией, Румынией, Турцией и Украиной в 1992 году в Бухаресте [Долинский,
Патлатюк, 2004].
3. Материал и методы исследования
Материал для данной работы был собран в период с
23.06.2014 г. по 20.04.2014 г. в Новороссийской бухте на базе Новороссийского
учебного и научно-исследовательского морского биологического центра Кубанского
государственного университета.
Пробы мейобентоса отбирались в акватории
Новороссийской бухты в трех различных точках: «Яхтклуб», «Рыбзавод», «поселок
Алексино». При выборе станций для отбора проб основным критерием была
непосредственная близость или удаленность от акватории порта. Станция «Яхтклуб»
находится ближе всего к акватории порта, за ней идет станция «Рыбзавод»,
«поселок Алексино» находится в открытой части бухты.
Пробы отбирались два раза (01.07.2014 и
09.07.2014) в урезовой зоне (0,5м).
С каждой точки было взято по шесть проб. Станции
отбора проб показаны на рисунке 2.
Сбор и обработку материала проводили по
общепринятым методикам.
При сборе материала таллом водорослей мы
накрывали мешком из мельничного газа № 49, подрезали подошву, затем затягивали
отверстие мешка и выносили его на поверхность.
Каждый таллом водорослей нами этикетировался и
помещался в полиэтиленовый мешок, и сутки пробы выдерживались без фиксации для
более полного отделения животных от водорослей. Пробы промывали пресной
проточной водой в большой емкости.
Смыв процеживали через систему сит (верхнее сито
- макрозообентосное, с ячеей 1 мм, нижнее, с ячеей 0,1 мм - мейозообентосное),
после чего пробы фиксировали 4% раствором формалина [Маккавеева, 1979].
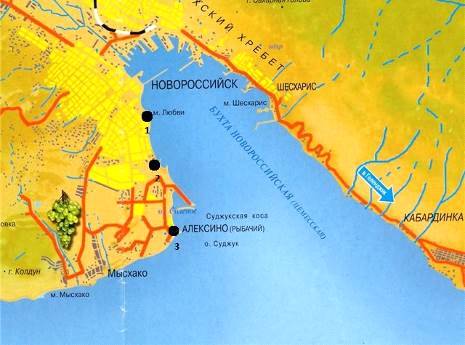
- Яхтклуб; 2 - Рыбзавод; 3 - поселок Алексино
Рисунок 2 - Станции отбора проб
Водоросли идентифицировались нами до вида,
определялась их биомасса с целью последующего пересчета мейофауны на 1 кг массы
водоросли. Для пересчета численности мейобентоса на 1 м2 дна мы
использовали данные по общему проективному покрытию.
Разбор проб проводился в камере Богорова под
бинокуляромМБС-10 при увеличении 8×2.
Произвели подсчет основных групп мейобентоса [Маккавеева, 1979]. При расчете
биомассы мейобентоса используются данные по средним весам, полученные для
отдельных групп мейобентоса. Для расчета биомассы отдельных крупных
мейобентосных организмов используются номограммы Численко [Численко, 1968].
Для идентификации водорослей был использован
определитель зеленых, бурых и красных водорослей южных морей СССР [Зинова,
1967]. Для идентификации организмов, обнаруженных в пробах, был использован
определитель фауны Черного и Азовского морей [Баранова, Бэческу, Голиков,
1986]. Также в работе было проведено сравнение сходства таксономического
состава на разных станциях отбора проб. Для этого использовался коэффициент
Серенсена, формула для расчета данного коэффициента выглядит таким образом
(формула 1):
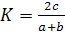 , (1)
, (1)
где а - число видов в одном
сообществе, - число видов в другом сообществе,
с - число видов, общих для 2
сообществ.
4. Мейобентос зарослей макрофитов прибрежной
зоны Новороссийской бухты
.1 Таксономический состав мейобентоса
Новороссийской бухты
Пробы мейобентоса отбирались в акватории
Новороссийской бухты в трех различных точках, таких как Яхтклуб, Рыбзавод,
поселок Алексино дважды (01.07.2014 и 09.07.2014) в урезовой зоне (0,5м). При
выборе станций для отбора проб основным критерием была непосредственная
близость или удаленность от акватории порта. Станция Яхтклуб находится ближе
всего к акватории порта, за ней идет станция Рыбзавод, поселок Алексино
находится в открытой части бухты.
В результате исследований в составе летнего
мейобентоса удалось зарегистрировать представителей 14 таксономических групп
(таблица 2).
Таблица 2 - Сравнение таксономического состава
мейобентоса на разных станциях
|
Таксономическая
группа
|
Станция
|
|
Яхтклуб
|
Рыбзавод
|
П.
Алексино
|
|
Эумейобентос
|
|
Turbellaria
|
+
|
+
|
+
|
|
Nematoda
|
+
|
+
|
+
|
|
Harpacticoida
|
+
|
+
|
+
|
|
Ostracoda
|
+
|
+
|
+
|
|
Acarina
|
+
|
+
|
+
|
|
Foraminifera
|
+
|
-
|
+
|
|
Псевдомейбентос
|
|
Polychaeta
|
+
|
+
|
+
|
|
Anisopoda
|
+
|
-
|
+
|
|
Isopoda
|
+
|
+
|
+
|
|
Amphipoda
|
+
|
+
|
+
|
|
Bivalvia
|
+
|
+
|
+
|
|
Gastropoda
|
+
|
+
|
-
|
|
Olygochaeta
|
+
|
+
|
+
|
|
Chironomidae
|
+
|
+
|
+
|
|
Итого
|
14
|
12
|
13
|
Из таблицы мы видим, что состав мейобентоса по
количеству таксономических групп в разных районах бухты различается
незначительно.
Так, на станции Яхтклуб, которая расположена
ближе всего к акватории порта зарегистрировано 14 таксономических групп, таких
как Turbellaria, Nematoda, Harpacticoida, Ostracoda, Acarina, Foraminifera,
Polychaeta, Anisopoda,
Isopoda, Amphipoda,
Bivalvia, Gastropoda,
Olygochaeta, Chironomidae. На
станции Рыбзавод, что находится в средней части бухты зарегистрировано 12
таксономических групп, таких как Turbellaria, Nematoda, Harpacticoida,
Ostracoda, Acarina, Polychaeta, Isopoda, Amphipoda, Bivalvia, Gastropoda,
Olygochaeta, Chironomidae. А на станции п. Алексино, которая находится в
открытой части бухты, зарегистрировано 13 таксономических групп, таких как Turbellaria,
Nematoda, Harpacticoida, Ostracoda, Acarina, Foraminifera, Polychaeta,
Anisopoda, Isopoda, Amphipoda, Bivalvia, Olygochaeta, Chironomidae. На
основании данных таблицы 2 можно подсчитать коэффициент сходства
таксономического состава мейобентоса зарослей макрофитов. Он рассчитывается по
формуле Серенсена. Этот коэффициент является наиболее универсальным при оценках
сходства двух или более совокупностей данных. Полученные данные по сходству
таксономического состава мейобентоса представлены в таблице 3.
Таблица 3 - Коэффициент сходства
таксономического состава мейобентоса
|
Сравниваемые
станции
|
Коэффициент
Серенсена
|
|
Яхтклуб
- Рыбзавод
|
0,92
|
|
Яхтклуб
- п. Алексино
|
0,96
|
|
Рыбзавод
- п. Алексино
|
0,88
|
Чем ближе значение коэффициента к единице, тем
выше сходство таксономического состава на разных точках отбора проб. Из таблицы
мы видим, что наибольшим сходством обладают такие точки, как Яхтклуб и п.
Алексино, которые находятся в наибольшем удалении друг от друга. Наименьшим
сходством обладают Рыбзавод и п. Алексино. Такие результаты скорее всего
связаны с тем, что из-за стоков ливневых и бытовых отходов в районе Рыбзавода
зеленые водоросли (кладофора и ульва) в прибрежной зоне развиваются сильнее чем
бурые, в районе Яхтклуба таких стоков нет, а в районе п. Алексино трубы с
городскими сточными водами отведены достаточно глубоко в море и на этих
станциях в прибрежной зоне преобладают бурые водороссли.
На следующем этапе работы было проведено
сравнение таксономического состава мейобентоса за 2003, 2008 и 2014 гг.
(таблица 4).
Таблица 4 - Сравнение таксономического состава
мейобентоса за 2003, 2008 и 2014 гг.
|
Таксономическая
группа
|
Год
|
|
2003
(фондовые данные НУНМБЦ)
|
2008
(фондовые данные НУНМБЦ)
|
2014
(наши данные)
|
|
Эумейобентос
|
|
Turbellaria
|
+
|
+
|
+
|
|
Nematoda
|
+
|
+
|
+
|
|
Kinorhyncha
|
-
|
+
|
-
|
|
Harpacticoida
|
+
|
+
|
+
|
|
Ostracoda
|
+
|
+
|
+
|
|
Foraminifera
|
-
|
+
|
+
|
|
Acarina
|
+
|
+
|
+
|
|
Псевдомейобентос
|
|
Polychaeta
|
+
|
+
|
+
|
|
Cirripedia
|
-
|
+
|
-
|
|
Anisopoda
|
+
|
+
|
+
|
|
Cumacea
|
-
|
+
|
-
|
|
Isopoda
|
+
|
+
|
+
|
|
Amphipoda
|
+
|
+
|
+
|
|
Bivalvia
|
+
|
+
|
+
|
|
Gastropoda
|
+
|
+
|
+
|
|
Olygochaeta
|
-
|
+
|
+
|
|
Chironomidae
|
+
|
+
|
+
|
|
Итого
|
12
|
17
|
14
|
Пробы в 2003, 2008 и 2014 гг. отбирались на
одних и тех же станциях: Яхтклуб, Рыбзавод, п. Алексино. Из таблицы видно, что
наибольшее количество таксономических групп было зарегистрировано в 2008 г. 17
таксономических групп, таких как Turbellaria, Nematoda, Kinorhyncha,
Harpacticoida, Ostracoda, Foraminifera, Acarina, Polychaeta, Cirripedia,
Anisopoda, Cumacea,
Isopoda,Amphipoda,
Bivalvia, Gastropoda,
Oligochaeta, Chironomidae. В 2003 году было зарегистрировано наименьшее
количество таксономических групп - 12, таких как Turbellaria, Nematoda,
Harpacticoida, Ostracoda, Acarina, Polychaeta, Anisopoda, Isopoda, Amphipoda,
Bivalvia, Gastropoda. В 2014 г. было зарегистрировано 14 таксономических групп,
таких как Turbellaria, Nematoda, Harpacticoida, Ostracoda, Foraminifera,
Acarina, Polychaeta, Anisopoda, Isopoda, Amphipoda, Bivalvia, Gastropoda,
Oligochaeta, Chironomidae.
Наиболее богатым таксономический состав был в
2008 г., он составил 17 таксономичекских групп, в 2014 г. обнаруженный
таксономический состав был беднее и составил 14 таксономических, в пробах 2014
г. не обнаружено таких групп, как Kinorhyncha, Cirripedia и Cumacea. В 2003 г.
таксономический состав мейобентоса был наиболее беден, он составил всего 12 таксономических
групп, в данных пробах не было обнаружено таких групп организмов, как
Kinorhyncha, Foraminifera, Cirripedia, Cumacea, Oligochaeta.
4.2 Пространственная изменчивость численности и
биомассы мейобентоса Новороссийской бухты
На рисунке 3 приведены сведения по общей
численности мейобентоса на трех станциях сбора проб: Яхтклуб, Рыбзавод и п.
Алексино.
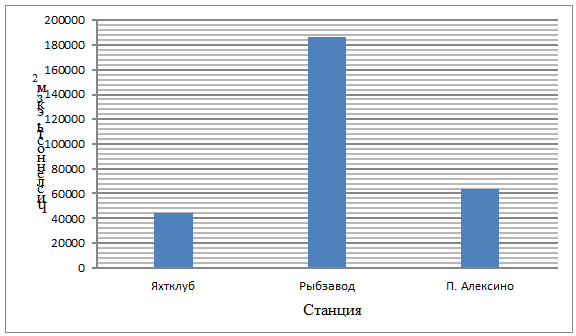
Рисунок 3 - Сравнение численности
мейобентоса с разных станций сбора проб
Из диаграммы мы видим, что на станции
Яхтклуб, которая находится ближе всего к акватории порта - самой загрязненной
части бухты, - общая численность организмов мейобентоса самая низкая из всех
станций, она составляет 43795 экземпляров/м2. На станции п.
Алескино, которая относится к открытой части бухты, численность выше и
составляет 63513 экземпляров/м2. Самая высокая численность
организмов наблюдалась на станции Рыбзавод, она приблизительно в три - четыре
раза выше, чем на двух других станциях и составила 186069 экземпляров/м2.
Это может быть связано с большим содержанием органики из-за выхода в этом
районе в море городских ливневых и бытовых сточных вод, что позволяет в большом
количестве развиваться фитопланктону, входящему в состав пищевого рациона
мейобентосных организмов.
На следующем этапе исследования были
проведены сравнения численности организмов, принадлежащих к разным
таксономическим группам, в пробах, отобранных на разных станциях. Численность
рассчитывалась в процентах (таблица 4) и в экземплярах/м2 (таблица
5).
Таблица 4 - Сравнение относительной
численности таксономических групп с разных станций отбора проб В процентах
|
Таксономическая
группа
|
Яхтклуб
|
Рыбзавод
|
П.
Алексино
|
|
Эумейобентос
|
|
Turbellaria
|
24,8
|
7,2
|
20,8
|
1,9
|
5,5
|
8,4
|
|
Harpacticoida
|
27,7
|
54,1
|
20,1
|
|
Ostracoda
|
3,0
|
0,8
|
17,0
|
|
Foraminifera
|
<0,1
|
<0,1
|
<0,1
|
|
Acarina
|
21,0
|
15,2
|
6,2
|
|
Псевдомейобентос
|
|
Polychaeta
|
12,7
|
11,8
|
12,1
|
|
Anisopoda
|
1,0
|
<0,1
|
0,3
|
|
Isopoda
|
1,3
|
<0,1
|
1,0
|
|
Amphipoda
|
2,9
|
0,8
|
3,4
|
|
Bivalvia
|
1,5
|
<0,1
|
0,9
|
|
Gastropoda
|
1,8
|
1,2
|
0,9
|
|
Olygochaeta
|
<0,1
|
<0,1
|
<0,1
|
|
Chironomidae
|
0,4
|
3,4
|
8,9
|
|
Итого
|
100
|
100
|
100
|
В процентном соотношении на станции Яхтклуб
доминировали Harpacticoida (27,7 %), Turbellaria (24,8 %) и Acarina (21,0 %).
Polychaeta(12,7 %) занимают субдоминантное положение. Остальные таксономические
группы, такие как Nematoda, Ostracoda, Foraminifera, Anisopoda, Isopoda,
Amphipoda, Bivalvia, Gastropoda, Oligochaeta, Chironomidae составляли гораздо
меньший процент. На станции Рыбзавод доминантное положение в процентном
соотношении занимали Harpacticoida (54,1 %), субдоминантами были Acarina (15,2
%) и Polychaeta (11,8 %). Относительная численность остальных таксономических
групп, таких как Turbellaria, Nematoda, Ostracoda, Foraminifera, Anisopoda,
Isopoda, Amphipoda, Bivalvia, Gastropoda, Oligochaeta, Chironomidae была
значительно ниже.
На станции п. Алексино в процентном соотношении
доминировали Turbellaria (20,8 %) и Harpacticoida (20,1 %), субдоминантное
положение занимали Ostracoda (17,0 %) и Polychaeta (12,1 %). Остальные группы,
такие как Nematoda, Foraminifera, Acarina, Anisopoda, Isopoda, Amphipoda,
Bivalvia, Gastropoda, Olygochaeta, Chironomidae составляли гораздо меньший
процент.
Таблица 5 - Сравнение численности мейобентоса
цистозиры с разных станций сбора проб В экз./м2
|
Таксономическая
группа
|
Численность
|
|
Яхтклуб
|
Рыбзавод
|
П.
Алексино
|
|
Эумейобентос
|
|
Turbellaria
|
10867
|
13491
|
13190
|
|
Nematoda
|
829
|
10482
|
5357
|
|
Harpacticoida
|
12130
|
100914
|
12773
|
|
Foraminifera
|
21
|
-
|
13
|
|
Ostracoda
|
1299
|
1430
|
10791
|
|
Acarina
|
921
|
28741
|
3950
|
|
Псевдомейобентос
|
|
Polychaeta
|
5543
|
22129
|
7646
|
|
Anisopoda
|
432
|
131
|
606
|
|
Isopoda
|
569
|
11
|
206
|
|
Amphipoda
|
1272
|
1750
|
2082
|
|
Bivalvia
|
669
|
15
|
593
|
|
Olygochaeta
|
2
|
443
|
559
|
|
Gastropoda
|
778
|
21
|
33
|
|
Chironomidae
|
190
|
6511
|
5663
|
|
Итого
|
43795
|
186069
|
63513
|
Из данной таблицы мы видим, что Turbellaria были
одной из самых многочисленных групп на всех станциях. Их численность составила
на станции Яхтклуб 10867 экземпляров/м2, на станции Рыбзавод 13491
экземпляров/м2, а на станции п. Алексино 13190 экземпляров/м2.
Nematoda были наиболее многочисленны на станции Рыбзавод, их численность
составила 10482экземпляров/м2,, на станции п. Алексино численность
нематод была примерно в 2 раза ниже, чем на Рыбзаводе и составила
5357экземпляров/м2, а на станции п. Алексино их численность была
очень маленькой по сравнению с двумя другими станциями и составила
829экземпляров/м2.
Численность Harpacticoida была очень высокой на
станции Рыбзавод и составила 100914экземпляров/м2, что в несколько
раз выше, чем на Яхтклубе (12130 экземпляров/м2) и в п. Алексино
(12773 экземпляров/м2).
Численность Foraminifera была незначительной на
станциях Яхтклуб (21 экземпляров/м2) и п. Алексино (13 экземпляров/м2),
а на станции Рыбзавод данная группа организмов не обнаружена. были наиболее
многочисленны на станции п. Алексино, там их численность составила 10791экземпляров/м2,
на станциях Яхтклуб и Рыбзавод данных организмов было значительно меньше, там
их численность составила 1299и 1430экземпляров/м2соответственно.были
наиболее многочисленны на станции рыбзавод, их численность составляла
28741экземпляров/м2, на станциях Яхтклуб и п. Алексино их
численность по сравнению с Рыбзаводом была значительно ниже и составила 921 и
3950 экземпляров/м2 соответственно. были наиболее многочисленны на
Рыбзаводе - 22129 экземпляров/м2, на Яхтклубе их численность
составила 5543 экземпляров/м2, в п. Алексино - 7646 экземпляров/м2.
Численность Anisopoda на станции Яхтклуб -
432экземпляров/м2, на станции Рыбзавод - 131 экземпляр/м2,
на станции п. Алексино - 606 экземпляров/м2. : на Яхтклубе -
569экземпляров/м2,на Рыбзаводе их численность была незначительной -
11 экземпляров/м2, в п. Алексино - 206экземпляров/м2.
Численность Amphipodaна станциях Яхтклуб,
Рыбзавод и п. Алексино была довольно высокой и составила 1272, 1750 и
2082экземпляров/м2 соответственно. Численность Bivalvia: на Яхтклубе
- 669экземпляров/м2, на Рыбзаводе - 15экземпляров/м2, в
п. Алексино - 593экземпляров/м2.
Численность Olygochaeta на станции Яхтклуб была
незначительной и составила 2 экземпляра/м2, на станциях Рыбзавод и
п. Алексино их численность была значительно выше и составила 443 и
559экземпляров/м2 соответственно. Gastropoda наиболее
многочисленными были на станции Яхтклуб - 778экземпляров/м2, на
станции Рыбзавод - 21экземпляр/м2, на станции п. Алексино - 33
экземпляра/м2. имели наименьшую численность на станции Яхтклуб - 190
экземпляров/м2, на станциях рыбзавод и п. Алексино численность
данной таксономической группы была значительно выше и составила 6511 и
5663экземпляров/м2 соответственно.
Ниже приведены результаты исследования биомассы
мейобентоса цистозиры: график, наглядно показывающий, пространственные
изменения биомассы мейобентоса (рисунок 4).
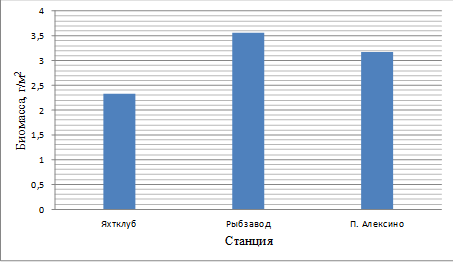
Рисунок 4 - Сравнение биомассы мейобентоса с
разных станций сбора проб
Из рисунка мы видим, что наибольшая биомасса
мейобентоса была зарегистрирована на станции Рыбзавод, она составила 3,564 г/м2,
на станции п. Алексино биомасса была незначительно меньше чем на предыдущей
станции и составила 3,177 г/м2. На станции Яхтклуб биомасса была
самой низкой - 2,339 г/м2. Это связано с более высокой численностью
организмов на Рыбзаводе и в п. Алексино, по сравнению с Яхтклубом.
Таблица 6 - Сравнение относительной биомассы
мейобентоса цистозиры с разных станций сбора проб В процентах
|
Таксономическая
группа
|
Биомасса
|
|
Яхтклуб
|
Рыбзавод
|
П.
Алексино
|
|
Эумейобентос
|
|
Turbellaria
|
27,9
|
22,7
|
24,9
|
|
Nematoda
|
0,0
|
0,1
|
0,0
|
|
Harpacticoida
|
3,1
|
17,1
|
2,5
|
|
Foraminifera
|
<0,1
|
0,0
|
<0,1
|
|
Ostracoda
|
2,2
|
1,6
|
13,6
|
|
Acarina
|
7,9
|
16,1
|
2,5
|
|
Псевдомейобентос
|
|
Polychaeta
|
4,7
|
12,5
|
4,8
|
|
Anisopoda
|
5,6
|
1,1
|
2,0
|
|
Isopoda
|
14,5
|
0,0
|
11,5
|
|
Amphipoda
|
21,8
|
19,6
|
26,3
|
|
Bivalvia
|
2,0
|
0,0
|
1,3
|
|
Olygochaeta
|
0,0
|
0,0
|
0,0
|
|
Gastropoda
|
10,0
|
3,7
|
5,2
|
|
Chironomidae
|
0,3
|
5,5
|
5,4
|
|
Итого
|
100
|
100
|
100
|
Из таблицы мы видим, что на станции Яхтклуб
домининтное положение в процентном соотношении по биомассе занимали Turbellaria
(27,9 %) и Amphipoda (21,8 %), субдоминантами были Isopoda (14,5 %) и
Gastropoda (10,0 %), остальные группы организмов составляли значительно меньший
процент. На станции Рыбзавод доминантное положение занимали те же организмы что
и на предыдущей станции: Turbellaria (22,7 %) и Amphipoda (19,6 %),
субдоминантами были Harpacticoida (17,1 %), Acarina (16,1 %) и Polychaeta (12,5
%), остальные таксономические группы составили гораздо меньший процент.
На станции п. Алексино доминантное положение
также как и на двух других станциях занимали Amphipoda (26,3 %) и Turbellaria
(24,9 %), субдоминантами были Ostracoda (13,6 %) и Isopoda (11,5 %), остальные
таксономические группы составили гораздо меньший процент.
Таблица 7 - Сравнение биомассы мейобентоса
цистозиры с разных станций сбора проб В г/м2
|
Таксономическая
группа
|
Биомасса
|
|
Яхтклуб
|
Рыбзавод
|
П.
Алексино
|
|
Эумейобентос
|
|
Turbellaria
|
0,652
|
0,809
|
0,791
|
|
Nematoda
|
0,000
|
0,004
|
0,002
|
|
Harpacticoida
|
0,073
|
0,605
|
0,077
|
|
Foraminifera
|
<0,001
|
0,000
|
<0,001
|
|
Ostracoda
|
0,052
|
0,057
|
0,432
|
|
Acarina
|
0,184
|
0,575
|
0,079
|
|
Псевдомейобентос
|
|
Polychaeta
|
0,111
|
0,443
|
0,153
|
|
Anisopoda
|
0,130
|
0,039
|
0,062
|
|
Isopoda
|
0,342
|
0,001
|
0,364
|
|
Amphipoda
|
0,509
|
0,700
|
0,833
|
|
Bivalvia
|
0,047
|
0,001
|
0,041
|
|
Olygochaeta
|
<0,001
|
0,001
|
0,001
|
|
Gastropoda
|
0,234
|
0,133
|
0,164
|
|
Chironomidae
|
0,006
|
0,195
|
0,170
|
|
Итого
|
2,339
|
3,564
|
3,177
|
Из таблицы мы видим, что биомасса таких
организмов как Nematoda, Foraminifera и Olygochaeta на всех станциях была
ничтожно мала, в то время как другие организмы, такие как Turbellaria,
Amphipoda и Polychaeta на всех станциях имели довольно большую биомассу.
Биомасса остальных групп организмов на разных станциях сильно варьировала. Так,
биомасса Harpacticoida на станции Рыбзавод была самой высокой и составила 0,605
г/м2, на станциях Яхтклуб и п. Алексино биомасса данных организмов
была значительно ниже и составила 0,073 и 0,077г/м2 соответственно.
Ostracoda на станции Рыбзавод имени наибольшую биомассу - 0,432г/м2,
на станции Яхтклуб она составила 0,052г/м2, на станции п. Алексино -
0,057г/м2. Биомасса Acarina на Яхтклубе была 0,184г/м2,
на Рыбзаводе - 0,575 г/м2, в п. Алексино - 0,079 г/м2.Биомасса
Anisopoda наибольшей была на Яхтклубе - 0,130г/м2, на Рыбзаводе она
составила 0,039г/м2, в п. Алексино - 0,062г/м2. Isopoda
на станциях Яхтклуб и п. Алексино имели биомассу 0,342 и 0,364 г/м2
соответственно, а на станции Рыбзавод биомасса этих организмов была
незначительной и составила всего 0,001г/м2. Биомасса Bivalvia на
всех трех станциях была низкой: Яхтклуб - 0,047 г/м2, Рыбзавод -
0,001г/м2, п. Алексино - 0,041 г/м2. Gastropoda
наибольшую биомассу имели на станции Яхтклуб - 0,234г/м2, на
станциях Рыбзавод их биомасса различалась незначительно и составляла 0,133 и
0,164г/м2 соответственно. Биомасса Chironomidae на станциях Рыбзавод
и п. Алексино была почти одинаковой и составляла 0,195 и 0,170 г/м2
соответственно, на станции Яхтклуб биомасса данных организмов была
незначительной - 0,006г/м2.
.3 Временная динамика численности и биомассы
мейобентоса Новороссийской бухты
На следующем этапе исследования было проведено
сравнение численности (таблица 8) и биомассы (таблица 9) мейобентоса зарослей
макрофитов за 2003, 2008 и 2014 гг.
Таблица 8 - Динамика численности мейобентоса
Новороссийской бухты В экз./м2
|
Таксономическая
группа
|
Численность
|
|
2003
г. (фондовые данные НУНМБЦ)
|
2008
г. (фондовые дан-ныеНУНМБЦ)
|
2014
г. (наши данные)
|
|
Эумейобентос
|
|
Turbellaria
|
602
|
69
|
12516
|
|
Nematoda
|
1741
|
4611
|
5556
|
|
Kinorhyncha
|
0
|
33
|
0
|
|
Harpacticoida
|
4768
|
3883
|
41939
|
|
Foraminifera
|
0
|
40
|
11
|
|
Ostracoda
|
653
|
654
|
4506
|
|
Acarina
|
2323
|
6833
|
11204
|
|
Псевдомейобентос
|
|
Polychaeta
|
26
|
316
|
11774
|
|
Cirripedia
|
0
|
39
|
0
|
|
Anisopoda
|
396
|
703
|
390
|
|
Cumacea
|
0
|
20
|
0
|
|
Isopoda
|
314
|
110
|
263
|
|
Amphipoda
|
4173
|
6267
|
1701
|
|
Bivalvia
|
558
|
5549
|
1277
|
|
Olygochaeta
|
0
|
2
|
334
|
|
Gastropoda
|
26
|
82
|
277
|
|
Chironomidae
|
106
|
4121
|
|
Итого
|
17709
|
29317
|
95869
|
Из таблицы мы видим, что численность организмов
от 2003 к 2014 г. имеет тенденцию к увеличению. Так, общая численность
организмов в 2003 г. составила 17709 экземпляров/м2, в 2008 - 29317
экземпляров/м2, а к 2014 г. численность организмов значительно
возросла и составила 95869 экземпляров/м2.
Особенно существенную вспышку численности дали
такие организмы как Turbellaria, Harpacticoida, Acarina, Ostracoda, Polychaeta,
Olygochaeta. Такие организмы как Kinorhyncha, Foraminifera, Cirripedia,
Cumacea, Isopoda в данный период имели низкую численность, без заметных вспышек
и спадов.
Организмы, которые дали вспышку численности
имеют высокую устойчивость к органическим загрязнениям, эти загрязнения обеспечивают
увеличение кормовой базы, что благотворно влияет на развитие данных организмов.
Те организмы, численность которых осталась неизменной или снизилась, имеют
меньшую устойчивость к загрязнениям, в том числе органическим.
В период с 2003 по 2014 г. доминантные и
субдоминантные таксономические группы не изменились.
Таблица 9 - Временная динамика биомассы
мейобентоса Новороссийской бухты В г/м2
|
Таксономическая
группа
|
Биомасса
|
|
2003
г. (фондовые данные НУНМБЦ)
|
2008
г. (фондовые данные НУНМБЦ)
|
2014
г. (наши данные)
|
|
Эумейобентос
|
|
Turbellaria
|
0,036
|
0,004
|
0,751
|
|
Nematoda
|
0,001
|
0,002
|
0,006
|
|
Kinorhyncha
|
0,000
|
<0,001
|
0,000
|
|
Harpacticoida
|
0,028
|
0,023
|
0,252
|
|
Foraminifera
|
0,000
|
0,001
|
<0,001
|
|
Ostracoda
|
0,026
|
0,026
|
0,180
|
|
Acarina
|
0,058
|
0,137
|
0,279
|
|
Псевдомейобентос
|
|
Polychaeta
|
<0,001
|
0,006
|
0,236
|
|
Cirripedia
|
0,000
|
0,004
|
0,000
|
|
Anisopoda
|
0,077
|
0,211
|
0,077
|
|
Cumacea
|
0,000
|
0,002
|
0,000
|
|
Isopoda
|
0,282
|
0,066
|
0,236
|
|
Amphipoda
|
1,670
|
2,507
|
0,681
|
|
Bivalvia
|
0,013
|
0,388
|
0,030
|
|
Olygochaeta
|
0,000
|
<0,001
|
<0,001
|
|
Gastropoda
|
0,017
|
0,025
|
0,177
|
|
Chironomidae
|
0,064
|
0,003
|
0,124
|
|
Итого
|
2,255
|
3,405
|
3,029
|
Из таблицы мы видим, что наибольшей биомассы
мейобентосные организмы достигали летом 2008 г. (3,405 г/м2), в 2014
г. биомасса мейоентоса была немного ниже (3,029 г/м2). В 2003 г.
биомасса была наиболее низкой и составила 2,255 г/м2.
Анализируя две последние таблицы: временная
динамика численности и временная динамика биомассы мейобентоса Новороссийской
бухты, мы видим, что в 2014 г. численность организмов была значительно выше,
чем в 2008 и 2003 гг. но биомасса животных отличается незначительно.
Численность организмов в 2003 г. была в 5,4 раз ниже чем в 2014 г., а биомасса
была ниже всего в 1,3 раза выше. Это связано с тем, что довольно большой
численности в 2003 и 2008 гг. достигали крупные организмы с большой массой
тела, такие как Amphipoda, а в 2014 г. доминантное положение заняли мелкие
организмы такие как Harpacticoida.
Данные по относительной численности (таблица 10)
и биомассе (таблица 11) за 2003, 2008 и 2014 гг. позволяют проследить изменения
в структуре сообщества мейобентоса с 2003 по 2014 гг.
Таблица 10 - Временная динамика относительной
численности мейобентоса Новороссийской бухты В процентах
|
Таксономическая
группа
|
Численность
|
|
2003
г. (фондовые данные НУНМБЦ)
|
2008
г. (фондовые данные НУНМБЦ)
|
2014
г. (наши данные)
|
|
Эумейобентос
|
|
Turbellaria
|
3,4
|
0,2
|
13,1
|
|
Nematoda
|
10,2
|
15,7
|
5,8
|
|
Kinorhyncha
|
0,0
|
0,1
|
0,0
|
|
Harpacticoida
|
26,8
|
13,2
|
43,7
|
|
Foraminifera
|
0,0
|
0,1
|
<0,1
|
|
Ostracoda
|
3,7
|
2,3
|
4,7
|
|
Acarina
|
13,1
|
23,3
|
11,7
|
|
Псевдомейобентос
|
|
Polychaeta
|
0,1
|
1,1
|
12,3
|
|
Cirripedia
|
0,0
|
0,1
|
0,0
|
|
Anisopoda
|
2,1
|
2,4
|
0,4
|
|
Cumacea
|
0,0
|
0,1
|
0,0
|
|
Isopoda
|
1,7
|
0,4
|
0,3
|
|
Amphipoda
|
23,6
|
21,4
|
1,8
|
|
Bivalvia
|
3,2
|
18,9
|
1,3
|
|
Olygochaeta
|
0,0
|
<0,1
|
0,3
|
|
Gastropoda
|
0,1
|
0,3
|
0,3
|
|
Chironomidae
|
12,0
|
0,4
|
4,3
|
|
Итого
|
100,0
|
100,0
|
100,0
|
Из таблицы мы видим, что в 2003 г. доминантное
положение в сообществе мейобентоса занимали Harpacticoida (устойчивы к
органическим загрязнениям) и Amphipoda (чувствительны к органическим
загрязнениям). В 2008 г. из состава доминантов выпали Harpacticoida, их
заменили Acarina, также хорошо переносящие органику. В 2014 г. чувствительные к
органическим загрязнениям Amphipoda резко снизили численность и выпали из
состава доминантов. Доминирующее положение заняли Harpacticoida.
Таблица 11 - Временная динамика относительной
биомассы мейобентоса Новороссийской бухты В процентах
|
Таксономическая
группа
|
Биомасса
|
|
2003
г.
|
2008
г.
|
2014
г.
|
|
Эумейобентос
|
|
Turbellaria
|
1,5
|
<0,1
|
24,9
|
|
Nematoda
|
<0,1
|
<0,1
|
0,3
|
|
Kinorhyncha
|
0,0
|
<0,1
|
0,0
|
|
Harpacticoida
|
1,2
|
0,8
|
8,4
|
|
Foraminifera
|
0,0
|
<0,1
|
<0,1
|
|
Ostracoda
|
1,1
|
0,8
|
5,9
|
|
Acarina
|
2,6
|
4,1
|
9,3
|
|
Псевдомейобентос
|
|
Polychaeta
|
<0,1
|
<0,1
|
7,9
|
|
Cirripedia
|
0,0
|
<0,1
|
0,0
|
|
Anisopoda
|
3,2
|
6,3
|
2,5
|
|
Cumacea
|
0,0
|
<0,1
|
0,0
|
|
Isopoda
|
12,4
|
2,0
|
7,5
|
|
Amphipoda
|
74,1
|
73,7
|
21,3
|
|
Bivalvia
|
0,5
|
11,5
|
1,0
|
|
Olygochaeta
|
0,0
|
<0,1
|
<0,1
|
|
Gastropoda
|
0,7
|
0,8
|
5,8
|
|
Chironomidae
|
2,7
|
<0,1
|
4,2
|
|
Итого
|
100
|
100
|
100
|
Из данной таблицы мы видим, что относительная
биомасса таких организмов как Turbellaria, Harpacticoida, Ostracoda, Acarina,
Polychaeta, Gastropoda, Chironomidae в период с 2003 по 2014 гг. значительно
увеличилась. В то время как Nematoda,
Kinorhyncha, Foraminifera,
Cirripedia, Anisopoda,
Cumacea, Olygochaeta
имели почти неизменную численность на протяжении данного периода времени.
Isopoda и Amphipoda значительно снизили свою относительную биомассу. В 2008 г.
по сравнению с 2003 и 2014 гг. имели большую биомассу такие таксономические
группы как Anisopoda, Bivalvia.
По биомассе доминантное положение в 2003 и 2008
гг. занимали Amphipoda (более 70 % биомассы сообщества), в настоящее время
(2014 г.) их биомасса значительно снизилась и немного превысила 21 %. В 2014 г.
наряду с Amphipoda довольно высокой биомассы достигли Turbellaria (24,9 %).
Такие спады и подъемы в биомассе могут быть
связаны с тем что, пробы отбирались в разное время и большей или меньшей
степенью загрязненности морских вод.
Заключение
По результатам выполненных исследований можно
сделать следующие выводы:
. Мейобентос зарослей макрофитов
прибрежной зоны Новороссийской бухты в летний период 2014 г. был представлен 14
таксономическими группами.
. Таксономический состав мейобентоса в
разных участках бухты отличался незначительно. Максимальное разнообразие этой
группы зообентоса (14 таксонов) было характерно для станции «Рыбзавод»,
минимальное (12 таксонов) - для станции «Яхтклуб».
. Разнообразие мейобентоса Новороссийской
бухты изменяется по годам. В 2003 г. было зарегистрировано 12, в 2008 - 17, в
2014 - 14 таксономических групп организмов.
. Численность и биомасса организмов
мейобентоса на разных станциях отбора проб значительно отличалась. Наименьшей
(43795 экземпляров/м2 и 2,339 г/м2) они были
на станции «Яхтклуб», расположенной наиболее близко к акватории порта. На
станции «Рыбзавод», где близко к берегу выходят ливневые и канализационные
городские стоки, численность и биомасса организмов мейобентоса была наибольшей
(186069 экземпляров/м2 и 3,564 г/м2). На станции «п.
Алексино» (открытая часть бухты) численность и биомасса организмов составила
63513 экземпляров/м2 и 3,177 г/м2.
. Структура меобентосных сообществ по
численности характеризовалась преобладанием на всех трех участках Harpacticoida
(от 20,1 % в районе п. Алексино до 54,1 % на станции «Рыбзавод»). Также в
состав доминантов по численности входили Turbellaria (на станциях «Яхтклуб» и
«п. Алексино») и Acarina (на станциях «Яхтклуб» и «Рыбзавод»).
. По относительной биомассе доминантное
положение на всех трех участках занимали Turbellaria (от 22,7 % на станции
«Рыбзавод» до 27,9 % на станции «Яхтклуб») и Amphipoda (от 19,6 % на станции
«Рыбзавод» до 26,3 % на станции «п. Алексино»). Также в состав доминантов по
биомассе входили: на станции «Рыбзавод» - Harpacticoida (17,1 %), и Acarinа
(16,1 %), на станциях «Яхтклуб» и «п. Алексино» - Isopoda
.
. Численность организмов в 2014 г. по
сравнению с 2003 г. увеличилась в 5,4 раз, в то время как биомасса всего в 1,3
раз. За указанный период произошло резкое увеличение численности и биомассы
групп зообентоса устойчивых к органическим загрязнениям, таких как
Harpacticoida, Acarina, Polychaeta.
Библиографический список
1. Александров
Б. Г. Проблема переноса водных организмов судами и некоторые подходы к оценке
риска новых инвазий // Морской экологический журнал. 2004. Вып. 1. С. 5-17.
.
Афанасьев Д.Ф., КорпаковаИ.Г. Макрофитобентосроссийского Азово-Черноморья.
Ростов н/Д., 2008. 291 с.
.
Баранова З. И., Бэческу М., Голиков А. Н. Определитель фауны Черного и
Азовского морей. Том 2. 1986. 569 с.
.
Вершинин А. Н. Жизнь Черного моря. М., 2003. 45 с.
.
Воробьева Л. В. Мейобентос украинского шельфа Черного и Азовского морей. Киев,
1999. 300 с.
.
Гусева Е. В. Мейобентос бухты Казачья // Морская санитарная гидробиология /
Редактор О.Г. Миронов. Севастополь, 1995. 4-12 с.
.
Гусева Е.В., АлемовС.В., Кирюхина Л.Н. Мейобентос севастопольских бухт по
данным 1994-1997 годов // Экология моря. 1998. Вып. 47. С. 7-12.
.
Долинский С. К., Патлатюк Е. Г. Экологическое состояние Черного моря.
Научно-практическая конференция 2004.
.
Многолетние изменения зообентоса Черного моря / В. Е. Заика [и др.]. Киев,
1992. 248 с.
.
Зайцев Ю. П., Александров Б. Г., Миничева Г. Г. Северо-западная часть Черного
моря: биология и экология. Киев, 2006. 701 с.
.
Зинова А. Д. Определитель зеленых, бурых и красных водорослей южных морей
СССР.М.-Л., 1967. 243 с.
.
Киселева М. И. Видовой состав и количественное развитие различных размерных
группировок бентоса в некоторых сообществах Черного моря // Экология моря.
1985. Вып. 212. С. 15-19.
.
Киселев М. И. Качественный состав и количественное распределение мейобентоса у
западного побережья Крыма // Бентос. Киев, 1965. С. 48-56.
.
Киселева М. И., Славина О. Я. Качественный состав и количественное
распределение макро- и мейобентоса у северного побережья Кавказа // Бентос.
Киев, 1965. С. 62-81.
.
Колесникова Е. А. МейобентосфиталиЧерного моря // Экология моря. 1991. Вып. 39.
76-81 с.
.
Колесникова Е. А., Мазгумян С. А. Сезонная динамика мейобентоса
зарослейцистозиры, в бухте Круглая // Проблемы устойчивого развития приморских
городов. Труды первой международной конференции: Приморские города и поселки
Черного и Азовского морей: совместно к устойчивому развитию, март 2001, ред. Н.
В. Шадрин. Севастополь 2002. - С.192-203.
.
Колесникова Е.А. Состав, обилие и миграции гарпактицид в некоторых донных
сообществах Черного моря: автореф. диссертация доктора биол. наук. Севастополь,
1981. 18с.
.
Колесникова Е. А., Шадрин Н. В. Влияние веса таллома цистозиры на плотность
популяций мейобентосных животных // Экология морских организмов. Киев, 1981. С.
83-93.
.
Маккавеева Е. Б. Беспозвоночные зарослей макрофитов Черного моря. Киев, 1979.
228 с.
.
Маккавеева Е. Б. Беспозвоночные зарослей макрофитов Черного моря. Киев, 1966.
227 с.
.
Маккавеева Е. Б. Биоценоз Cestoseira barbata прибрежного участка Черного моря
// Труды Севастопольской биологической станции. Том XII, 1959. С. 168-191.
.
Маккавеева Е. Б. Об экологии и количественном распределении паукообразных в
северо-восточной части Черного моря // Гидробиологический журнал. 1966. Том II,
№5. С. 30-33.
.
Маккеева Е. Б. Роль макрофитов и экологических факторов в формировании видового
состава зарослевых сообществ // республиканский межведомственный сборник. Серия
«Биология моря». Киев. 1967. С. 52-62.
.
Мейобентос в санитарно-биологическом аспекте / О.Г. Миронов [и др.] // Микромир
в морских санитарно-биологических исследованиях. Севастополь, 1995. С. 44-57.
.
Миловидова Н.Ю. Зообентос бухт северо-восточной части Черного моря. Автореф.
диссертация доктора биол. наук. Ростов-н/Д., 1967. 18с.
.
Миловидова Н.Ю. Количественная характеристика биоценоза цистозиры
северо-восточной части Черного моря // Морские подводные исследования. М.,
1969. С. 78-88.
.
Многолетние изменения зообентоса Черного моря / отв. ред. В.Е. Заика. Киев,
1992. 248 с.
.
Мокиевский В. О. Экология морского мейобентоса: количественное распределение,
таксономическое разнообразие и организация сообществ. М., 2012. 105 с.
.
Мокиевский В.О. Экология морскогомейобентоса. М., 2009. 286 с.
.
Мокиевский В.О., Малых Е.П. О методах количественного учета мейобентоса //
Океанология. Т. 2. 2002. С. 249-253.
.
Ремизова Н. П. Мейобентос зарослей цистозиры прибрежной зоны Северо-Восточной
части Черного моря. Новороссийск, 2008.С.
.
Сергеева Н.Г., Колесникова Е.А. Результаты изучения мейобентоса Черного моря //
Экология моря. 1996. Вып. 45. С. 54-62.
.
Современное состояние биоразнообразия прибрежных вод Крыма (черноморский
сектор) / под ред. В.Н. Еремеева, А.В. Гаевской. Севастополь, 2003. 511 с.
.
Техногенное загрязнение и процессы естественного самоочищения Прикавказской
зоны Черного моря / под ред. И. Ф. Глумова, М. В. Кочеткова. М., 1996. 502 с.
.
Тильба П. А., Мнацеканов Р. А., Крутолапов В. А. Водно-болотные угодья
Северного Кавказа. М., 2006. 245 с.
.
Харитонова О. С. Зообентос рыхлых грунтов бухт Северо-Восточной части Черного
Моря. Краснодар, 1981. С. 11-13.
.
Численко Л. Л. Номограммы для определения веса водных организмов по размеру и
форме тела. Л., 1968. 195 с.
.
Шереметевский А. М. К вопросу о компенсации развития макробентоса мейобентосом
на примере мидиевых банок Белого моря // Экология моря. 1991. Вып. 39. С.
89-92.