Влияние ионов металлов на карбоангидразоподобную активность внешних водорастворимых белков PsbP и PsbQ фотосистемы 2
Министерство образования и науки
Российской Федерации
Тульский государственный университет
Кафедра БИОЛОГИИ
бакалаврская работа
Направление 06.03.01
Биохимия
Влияние ионов металлов на
карбоангидразоподобную активность внешних водорастворимых белков PsbP и PsbQ фотосистемы 2
Оглавление
. СПИСОК СОКРАЩЕНИЙ. ВВЕДЕНИЕ. ОБЗОР ЛИТЕРАТУРЫ
. Фотосинтез
.1 Роль фотосинтеза
.2 Строение и функционирование фотосинтетического аппарата
высших растений
.3 Фотосистема 2
Важную роль в обеспечении стабильности Mn4CaO5 - кластера
играет белок PsbO (марганец - стабилизирующий белок). В процесс образования
кислорода также принимают участие: ион Ca2+; один или два иона Cl-.
Предполагается, что в обеспечении этими ионами в процессе сборки
водоокисляющего комплекса, а также в их сохранении в нормально функционирующей
ФС-2 участвуют внешние водорастворимые белки ВОК PsbP и PsbQ.
.Карбоангидразная активность фотосистемы 2
.1 Карбоангидразная активность ФС-2 высших растений и её
свойства
.2 Носители карбоангидразной активности в ФС-2 высших
растений
. Роль ионов металлов в биохимических процессах высших
растений. ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЙ
. Выделение тилакоидов по методу Berthold, Babcock, Yocum,
1981 с модификациями Ford, Evans, 1983 (на основе оригинальной статьи)
. Выделение частиц ФС-2 из тилакоидов по методу schiller и
dau (2005 г)
. Измерение концентрации хлорофилла
.Измерение скорости фотосинтетического выделения кислорода
. Измерение фотоиндуцированных изменений выхода флуоресценции
хлорофилла
. Получение и очистка белков PsbP и PsbQ
. Приготовление диализных трубок
. Электрофорез
. Измерение концентрации белка
. Измерение карбоангидрзной активности. РЕЗУЛЬТАТЫ И
ОБСУЖДЕНИЕ
. Получение и характеристика препаратов ФС-2
.1 Определение концентрации хлорофилла
.2 Измерение скорости выделения кислорода
.3 Измерение флуоресценции ФС-2
.4 Электрофорез
. Получение и очистка белков РsbР и РsbQ
.1 NaCl обработка
.2 Диализ
.3 Разделение белков методом ионнообменной хромотографии
.4 Электрофорез
.5 Определение концентрации белка по методу Брэдфорд
.6 Измерение карбоангидразоподобной активности белков PsbP и
PsbQ. ЗАКЛЮЧЕНИЕ. ВЫВОДЫ. СПИСОК ЛИТЕРАТУРЫ
I. СПИСОК СОКРАЩЕНИЙ
АТФ- аденозинтрифосфат
БК- бикарбонат
ВОК - водоокисляющий комплекс
КА- карбоангидраза(ы)
КА-активность-- карбоангидразная активность
КП-активность- карбоангидразоподобная активность
НАДФ- никотинамидадениндинуклеотидфосфат
П680- первичный донор электрона в фотосистеме 2
ПААГ- полиакриламидный гель
РЦ- реакционный центр
ССК2- основной светособирающий комплекс переферической антенны
фотосистемы 2
Фео- феофитин
ФС-1- фотосистема 1
ФС-2- фотосистема 2
Хл- хлорофилл
ЭДТА- этилендиаминтетраацетат
ЭТЦ- электрон-транспортная цепьфрагменты тилакоидных мембран, обогащённые
фотосистемой 2 (выделенные согласно (Berthold et al., 1981))
CР43 -
белок антенны ядерного комплекса ФС2
CР47 -
белок антенны ядерного комплекса ФС2
D1 -
интегральный белок РЦ ФС2
D2 -
интегральный белок РЦ ФС2
DCBQ-
2,6-дихлор-п-бензохинон
HСО3-
- ион бикарбоната;- (N-морфолино)этансульфоновая кислотаCaO5-кластер - неорганическое ядро ВОК
ФС2
PsbP и
PsbQ- водорастворимые белки ВОК ФС2высших
растений и зеленых водорослей с молекулярными массами 23 кДа и 17 кДа,
соответственно-амино-2-гидроксиметил-пропан-1,3-диол
QA -
первичный хиноновый акцептор электрона в ФС2
QB -
вторичный хиноновый акцептор электрона в ФС
II .ВВЕДЕНИЕ
Фотосинтез - процесс образования богатых энергией органических веществ из
веществ энергетически бедных - углекислого газа и воды. Углекислый газ,
поступающий в клетку из окружающей среды, электроны в виде НАДФН и энергия в
виде АТФ, поступающие из цепи переноса электронов, ассоциированной с
тилакоидной мембраной обеспечивают синтез углеводов. В состав тилакоидной
мембраны входят три белковых комплекса - фотосистема 1, фотосистема 2 и
цитохромный b6f комплекс.
Благодаря фотосинтезу стала возможной жизнь на поверхности нашей планеты.
Кроме того, результатом деятельности фотосинтезирующих организмов являются
запасы топлива, к сожалению, исчерпаемые. Однако, знание механизмов этого процесса
поможет обрести человечеству источник неиссякаемой экологически чистой энергии.
Например, знание механизма фотосинтетического расщепления воды позволит
получать водород как топливо практически в неограниченном количестве.
Известно, что тилакоиды имеют как минимум два источника КА активности в
ФС-2. Один из источников - Mn-стабилизирующий
белок PsbO (молекулярная масса 33 кДа), другой
источник - ассоциирован с ФС-2, который можно выявить путём отмывки белка PsbO 1М раствором CaCl2. В ходе экспериментов было
установлено, что источники КА активности зависят от присутствия ионов металлов Ca2+, Mn2+. Возник интерес действию других металлов на КА активность,
например, входящих в КА или имеющих сходный радиус с теми металлами, для
которых уже показана активность, либо установить действие тяжелых металлов.
Выяснение механизма действия ионов тяжелых металлов на ФС-2 является актуальной
проблемой, важной для развития сельского хозяйства и фундаментальных
представлений о механизме функционирования ФС-2. Так как, по имеющимся в
настоящий момент данным, ионы металлов могут влиять на КП-активность белков ВОК
ФС-2, необходимо изучить этот вопрос более детально. Поэтому целью нашей работы
явилось изучение влияния ионов металлов на карбоангидразоподобную активность
внешних водорастворимых белков PsbP и
PsbQ фотосистемы 2. Для достижения данной
цели мы выделили несколько задач:
) Выделить фотохимически активные фрагменты тилакоидных мембран
хлоропластов, обогащенных ФС-2 из листьев гороха (PisumsativumL. cv. Moscowski 559);
) Выделить из них белки водоокисляющего комплекса - PsbPи PsbQ;
) Установить влияние ионов двухвалентных металлов на
карбоангидразоподобную активность белков PsbPи PsbQ;
III.ОБЗОР ЛИТЕРАТУРЫ
1. Фотосинтез
.1 Роль фотосинтеза
Фотосинтез
- это окислительно-восстановительный процесс, протекающий в несколько этапов, в
котором происходит восстановление диоксида углерода до углеводов и окисление
воды до кислорода.. (<#"868134.files/image001.gif">
Рис. 1. Строение молекулы хлорофилла;
Хлорофилл a: R1= CH3, R2= С2Н5 ,
Хлорофилл b: R1= CH3, R2 = CH=CH2
Хлорофиллd: R1 = СООСН3, R2= CH=CH2
Растения имеют характерную зелёную окраску из-за того, что хлорофиллы
отражают зеленый, а поглощают красный и сине-фиолетовый свет. Молекулы
хлорофилла в мембранах входят в состав пигмент-белково-липидных комплексов,
названных фотосистемами. У растений и сине - зеленых водорослей имеются ФС-1 и
ФС-2, у фотосинтезирующих бактерий - ФС-1. Только ФС-2 имеет способность
отбирать электроны у воды.
Световая
фаза фотосинтеза осуществляется при участии двух фотосистем - ФС-1 и ФС-2,
содержащих пигменты и различные белки. Они вместе с цитохромным комплексом
являются основными компонентами фотосинтетической цепи переноса электронов.
Электрон-транспортная цепь организована в мембране в виде трансмембранных
белковых комплексов (ФС-1, ФС-2 и цитохромный b6f комплекс
<#"868134.files/image002.gif">
Рис. 2. Схематическое изображение белковых комплексов, включённых в
реакции фотоиндуцированного переноса электронов и протонов в тилакоидной
мембране фотосинтетических оксигенных организмов.
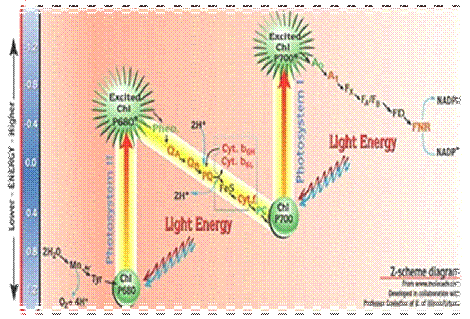
Рис. 3. Z-схема фотосинтеза
-схема, или схема нециклического транспорта
электронов, получила свое название из-за сходства с буквой Z. Её принцип
впервые был предложен в 1960 г. Р. Хиллом и Ф. Бендаллом и в 1961 г. был
подтвержден работами Л. Дюйзенса.
Конечным этапом световых стадий фотосинтеза является
восстановление NADP+, которое осуществляет ФС-1. При возбуждении П700 в
реакционном центре ФС-1 энергией, эквивалентной 1 кванту длинноволнового
красного света, электрон захватывается мономерной формой хлорофилла a (A1;
акцептор электрона) и затем последовательно передается переносчикам электронов
А2 и Ав (железосерные белки FeS), ферредоксину (водорастворимый FeS-белок) и
ферредоксин - NADP-оксидоредуктазе с FAD в качестве кофактора. Наконец, редуктаза
восстанавливает NADP+. Именно в ФС-2 происходит фотоиндуцированное окисление
воды. П700 является сильным окислителем. Донором электронов для него является
ФС-2. Электроны от ФС-2 переходят на ФС-1 посредствам мобильных переносчиков и
цитохромом b6f-комплексов.Исходным донором электронов для ФС-2 является
вода.Энергия, освобождающаяся при движении электронов от П680 до П700,
используется для синтеза АТФ из AДФ и неорганического фосфата
(фотофосфорилирование). (Холл Д., Рао К., 1983)
Фотохимический реакционный центр ФС-2П680 поглощает
энергию, эквивалентную 1 кванту коротковолнового красного света, и переходит в
синглетное возбужденное состояние, отдает электрон феофетину (Фео). От Фео
электроны, теряя энергию, последовательно передаются на пластохиноны - QA и QB,
в пул липидорастворимых молекул пластохинона (PQ), переносящих через липидную
фазу мембраны электроны и протоны в цитохромный b6f комплекс, который затем
восстанавливает Cu-содержащий белок пластоцианин (Пц). Электроны с Пц переходят
на вакантное место в П700.
Вакантное место («дырка») в П680 заполняется двумя
электроном из содержащего Мn -
неорганического ядра водоокисляющего комплекса. Этот комплекс связывает воду и
восстанавливается за счет её электронов. Для осуществления этой реакции в
белковом комплексе необходимы Мn и
С1-, а также Са2+. Таким образом, основным источником электронов для
восстановления NADP+ является
вода, разложение которой происходит в ФС-2.
1.3 Фотосистема 2
ФС-2 (Рис. 4) содержит более 20 различных белков и пигмент-белковых
комплексов, ее состав зависит от условий окружающей среды. Полипептиды ФС-2
выполняют ряд важных функций: часть поглощают энергию света и передают энергию
возбуждения, участвуют в разделении зарядов и окислении воды, стабилизируют
структуру димеров ФС-2, участвуют в сборке мономера и образовании димера,
связывают и ориентируют молекулы хлорофилла а и b, а также каротиноиды. В ФС-2 условно выделяют части,
выполняющих разные функции: 1) реакционный центр , 2) светособирающий комплекс,
3) водоокисляющий комплекс.
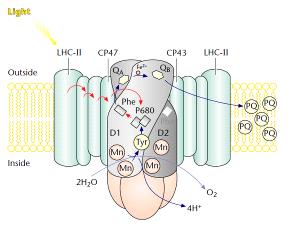
Рис. 4. Схематическое изображение фотосистемы 2 в тилакоидной мембране.
D1 иD2 белкиреакционногоцентра (РЦ). (Mn) - марганцевый кластер, участвующийв
фотоокислении воды; P680 (П680) - специальная пара молекул хлорофилла
реакционного центра; Phe (Фео) - феофитин; QA - связанный пластохинон; QB -
пластохинон, подвижный переносчик электронов;Tyr - аминокислотный остаток
тирозина-161 белка D1 (Yz); PQ - пул подвижных молекул пластохинона. Антенные
комплексы: LHC-II (ССК-2) - периферийный светособирающий комплекс ФС-2; CP47 и
CP43- хлорофилл-белковые комплексы внутренней антенны, имеющие соответственно
молекулярные массы 47 и 43 кДа.
1.3.1 Реакционный центр
Реакционный центр обеспечивает фотосинтетическое разделениезарядов -
первый этап преобразования солнечной энергии в энергию химических связей. Он
содержит интегральные белки D1 (PsbA) и D2 (PsbD),
каждый из которых образован пятью трансмембранными
α-спиралями,
расположенными в мембране пррактически параллельно друг другу (Barberetal., 1987). В состав РЦ ФС-2 также
входит третий интегральный белок - цитохром b559,количество молекул которого на
один РЦ точно не установлено (предполагается 1-2 молекулы).
На белках D1 и D2 расположены кофакторы переноса
электрона в ФС-2 - фотоактивная А-цепь: TyrZ, П680, ChlD1,
PheoD1, QA и QB. Энергия света,
поглощенная пигментами светособирающего комплекса ФС-2, переносится к
первичному донору электрона P680 - специализированному димеру хлорофиллов а
(Renger, 1992).Разделение зарядов в реакционном центре происходит за счет
перехода молекулы P680в возбужденное
синглетное состояние, за счет энергии поглощенного кванта света. В первой
стадии разделения зарядов принимает участие молекула хлорофилла a (ChlD1), которая располагается между П680 и PheoD1 (рис. 5.). Энергия возбуждения
возможно делокализуется между четырьмя молекулами хлорофилла(специальная пара PD1PD2 и вспомогательными ChlD1 и ChlD2). (Barber, 2002)
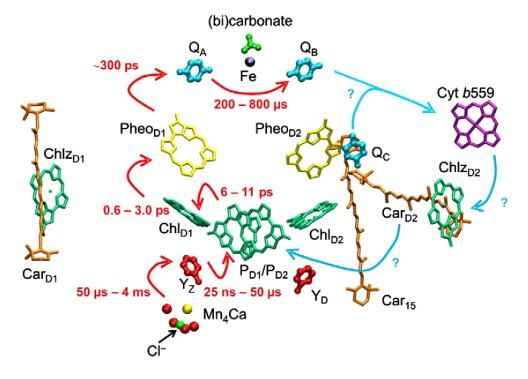
Рис. 5. Схема расположения кофакторов переноса электрона в ФС-2 (Guskovetal., 2009). Красные стрелки - этапы
переноса электронов с указанием времени переноса электрона. Голубые - возможные
пути переноса электрона (Muh et al., 2012).
РЦ входит в состав более крупного белкового комплекса, называемого «ядерным»
В его состав у высших растений, кроме белков D1, D2,
цитохром b-559 входят белкиCP43, CP47, PsbO;
могут входить белки PsbP, PsbQ.
Структура ядерного комплекса высших растений крайне близка к структуре
цианобактерий: для цианобактерии Thermosynechococcus elongatus показано наличие
35 молекул хлорофилла а, 2 молекул феофитина, 3-х пластохинонов и 12 β-каротинов в расчёте на один мономер,
а также марганцевого кластера и других кофакторов. Для «ядерных» комплексов из
высших растений (шпината) показано наличие от 35 до 80 молекул хлорофилла,
одной молекулы QA и марганцевого
кластера в расчете на один РЦ.
1.3.2 Светособирающий комплекс (ССК)
Светособирающий комплекс необходим для эффекктивного улавливания квантов
света с большой площади и направленной передачи энергии света в виде энергии
возбужденного состояния хлорофилла в реакционный центр ФС-2. Белки ССК
выполняют следующие функции:
определяют специфичное связывание, ориентацию и распределение молекул
пигментов;
служат посредниками во взаимодействии с другими белковыми компонентами в
структурной организации антенных систем, обеспечивая перенос энергии
возбуждения и регуляцию этого процесса
ССК условно разделяют на внешнюю и внутреннюю антенны.
Внутреннюю антенну ФС-2 составляют два белка - CP43 и CP47, которые
расположены по обе стороны от D1 и D2 (белков РЦ ФС-2). СР43 (с ним
скоординировано 13 молекул хлорофилла) находится со стороны белка D1, а СР47 (содержит 16 молекул
хлорофилла) - со стороны белка D2.К
внутренней антенне акже относят белки CP29, CP26, CP24. Внешняя антенна представленна белками ССК-2 разного
состава.
Эти комплексы формируют внешнюю антенну с высоким соотношением пигмент:
белок. Регулирование светособирающей функции происходит за счёт обратимого,
находящегося под окислительно-восстановительным контролем фосфорилирования
белков ССК-2 (Lhcb1 и Lhcb2), и происходящая в результате латеральная миграция
ССК-2 от ФС-2 к фотосистеме 1 (ФС-1) приводит к перераспределению энергии
возбуждения между ФС-2 и ФС-1. (Шитов, 2013)
1.3.3 Водоокисляющий комплекс (ВОК)
Для окисления воды в растениях и цианобактериях требуются мембранные
белки РЦ и внутренней антенны: СР47, СР43, D1, D2, α и β субъединицы цитохрома b-559. Фотоокисление воды происходит в
части ФС-2, обращенной в просвет тилакоида.
В
активном центре водоокисляющий комплекс содержит четыре иона марганца и один
ион кальция, которыесвязаны между собой при помощи μ-оксо мостиков (Mn4CaO5-кластер),
активный в окислительно-восстановительном отношении тирозин-161 (TyrZ)
и другие аминокислотные остатки, которые вместе функционируют как каркас. Mn4CaO5
- кластер ФС-2 высших растений окружён полипептидными цепями интегральных
белков РЦ и периферических белков водоокисляющего комплекса (PsbO,
PsbP, PsbQ). Эти полипептидные цепи стабилизируют структуру Mn4CaO5-кластера,
создавая условия для фотоокисления воды. Для образования одной молекулы
кислорода
<#"868134.files/image006.gif">
Рис. 6. График фотосинтетического выделения кислорода ФС-2. Стрелкой
вверх показан момент включения света. Стрелка вниз - момент выключения света.
Вычисление скорости выделения О2 за 1 час по формуле: V(О2) = ΔU*K*60мин/СХл, где ΔU- значение изменения вольтажа за 1
минуту, K- калибровочный коэффициент(мкМО2/мл*мин*В),
СХл- концентрация хлорофилла(мг/мл). Значение калибровочного
коэффициентавычисляется по специальному калибровочному графику (рис.7). Его
значение равно665, 79 мкМО2/ мл*мин*В. За тем по графику (рис.6) нашли
изменение напряжения (В) за 1 минуту после включения света (начальная скорость
на прямолинейном участке).
Скорость выделения кислорода составила 344,7 ± 12 мкМО2/ ч *мг/мл
Для этого метода получения BBYмаксимальное
значение для выделения кислорода показано порядка 600 мкМО2/ч*мг/мл, а среднее
значение колеблется в пределах от 300 до 450 мкМО2/ч*мг/мл. Полученные нами
данные свидетельствуют о достаточно хорошей функциональной активности ФС-2.
Рис. 7. Калибровочнаякривая для расчета скорости выделения кислорода
1.3 Измерение флуоресценции ФС-2
Ещё одним из наиболее достоверных методов определения функциональной
активности ФС-2 является метод измерения флуоресценции. Пигменты
фотосинтетического аппарата поглощают энергию света, часть высвечивается
обратно в виде флуоресценции или рассеивается в виде тепла. При комнатной
температуре в спектре флуоресценции наблюдается преимущественно полоса при 685
нм, принадлежащая антенному хлорофиллу а ФС-2. (Кизимирко Ю. В., 2006)
Молекула П680, входящая в состав реакционного центра ФС-2, в возбужденном
состоянии является первичным донором электрона для акцептора Qa. Реакционный
центр находится в открытом состоянии, когда молекула П680 окислена, а хиноновый
акцептор Qa восстановлен. После появления первичной пары П680+Фео- происходит
восстановление П680+ от вторичных доноров. В результате РЦ оказывается
состоянии П680Qa, называемое закрытым (Goh, Schreiber, Hedrich et al., 1999;
Renger, 1992). Эти состояния характеризуются разным выходом флуоресценции (в
закрытом состоянии флуоресценция максимальна, а в открытом - минимальна).
Разность между максимальным и минимальным выходом флуоресценции
называется переменной флуоресценцией. Было установлено, что под действием света
происходит восстановление первичного акцептора QA и выход флуоресценции
возрастает от начального уровня (Fo) до максимального уровня (Fm), т.е.
возникает так называемая переменная флуоресценция (Fv = Fm - Fo).Эта величина
пропорциональна той части энергии света, которая используется в фотохимических
реакциях фотосинтеза при открытых реакционных центрах ФС-2 и теряется в виде
флуоресценции и тепла при закрытых реакционных центрах. Для расчета данных,
полученных в ходе измерения следует брать одно из важнейших соотношений Fv/Fm,
т.е. отношение интенсивностей переменной и максимальной флуоресценции. (Ланкин
А.В. с соавт., 2014) Это отношение равно квантовому выходу использования
энергии света открытыми РЦ ФС-2. По изменению этой величины можно судить о
потенциальной эффективности первичных процессов фотосинтеза в ФС-2. Этот
параметр флуоресценции, как показатель состояния и эффективности
функционирования фотосинтетического аппарата, широко используется в
фундаментальных и прикладных исследованиях. (Antalandal., 1998), (G. H. Krause, E. Weis, 1991).
Измерение флуоресценции проводилось для препаратов изначально
адаптированных к темноте. Расчеты были проведены исходя из данных графика
флуоресценции (рис.8).
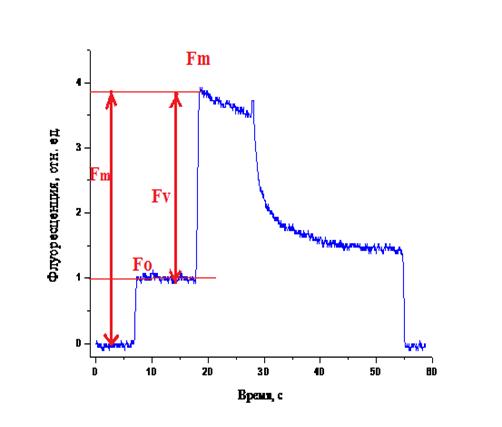
Рис. 8. График флуоресценции ФС-2.
Fm-
максимальный квантовый выход флуоресценции;
Fv-
зачение переменной флуоресценции;
Fo-
начальный уровень флуоресценции;
Отношение интенсивностей переменной и максимальной флуоресценции
составило 0,744. Из других источников известно, что максимальный квантовый
выход у листа растения составлял 0,83.(Лукаткин А. С. с соавт., 2009). Таким
образом, полученные нами BBY
частицы обладают довольно высоким показателем максимального квантового выхода
фотохимии ФС-2и этот показатель, в совокупности с данными о фотосинтетическом
выделении кислорода этим препаратом, свидетельствует о хорошей функциональной
активности образца, достаточной для дальнейшей процедуры выделения белков PsbP
и PsbQ.
1.4 Электрофорез
Ещё одним методом, позволяющем судить о целостности препаратов является
денатурирующий электрофорез. Электрофорез проводили в полиакриламидном геле для
идентификации ФС-2. (рис.9). Результаты электрофореза свидетельствуют о наличии
в препаратах ФС-2 всех необходимых нам для дальнейшей работы белков.
Исходя из экспериментальных данных о фотохимической активности и
электрофореза можно судить о функциональной активности субмембранных препаратов
ФС-2 и её целостности, позволяющей проводить с этими препаратами дальнейшие
обработки.
2. Получение и очистка белковРsbР и РsbQ
2.1 NaCl обработка
Субмембранные препараты ФС-2 обрабатывали 1М раствором NaCl для экстракции гидрофильных белков PsbP (18 кДа) и PsbQ (24 кДа) при температуре +4оС при комнатном освещении
в течение 30 минут. ссылка на статью
2.2 Диализ
Обессоливание белков проводилось методом диализа в присутствии 20 мМ HEPES-KOH-буфера (рН 7,5), содержащего 10 мМ NaCl и 2 мМ ЭДТА. ЭДТА использовалось в
качестве комлексообразователя, то есть препараты таким образом были избавлены
от металлов.
2.3 Разделение белков методом ионнообменной хромотографии
Ионообменная хроматография позволяет разделять
молекулы, основываясь на ионных взаимодействиях. Чаще всего ионообменная
хроматография является первым шагом к дальнейшей очистке белков.
Для разделения белков использовалась анионная
ионообменная хроматография. Она характеризуется неподвижной фазой с
положительно заряженными функциональными группами и задержкой отрицательно
заряженных анионов.
Изначально система хроматографа (ActaFPLCc колонкойMonoQ) промывается этиловым спиртом и деионизованной водой.
Разделение смеси белков PsbQ и
PsbP проводилось в присутствии буфера HEPES (рН 7.5). Cкорость нанесения белка на колонку выбиралась в зависимости
от давления и варьировала от 0,2 до 0,5 мл/мин.
После нанесения белковой смеси на колонку наблюдалось возрастание
показателя поглощения в ультрафиолетовой области, что свидетельствовало о
выходе белка из колонки(рис.10). Длина волны при поглощении в УФ области
составляет 280 нм. Первым из колонки выходил белок PsbQ (он был собран во фракцию, обозначенную номером 1).
Его выход был связан с тем, что он имеет изoэлектрическую точку (рI) равную 8.3, а белок PsbР - 6.5. Общий заряд белка определяется величиной рН.
Если значение pH ниже изоэлектрической точки - заряд белковой молекулы является
положительным, при pH выше - отрицательный. Таким образом при рН 7,5, белок РsbQ (заряженный положительно) выходит из
колонки, а PsbР (заряженный отрицательно)
задерживается на ней. Белок PsbР
элюировали в градиенте NaCl
от 0 до 500 мМ (время элюации 58 минут, выход белка наблюдался при 165-260 мМ NaCl). По мере элюции выходящий раствор
собирали в различные фракции. Исходя из данных поглощения в ультрафиолетовой
области, белки предположительно должны содержаться во фракциях 1, 6, 7 и 11.
Эти фракции взяли для анализа на содержание белков. Для этого использовали
электрофорез в денатурирующих условиях. Так как ряд фракций содержал высокую
концентрацию NaCl, которая мешает проведению
электрофореза, белки предварительно осадили с помощью трихлоруксусной кислоты
(надосадочную жидкость с высоким содержанием соли слили), а затем перевели
осадок в буфер для нанесения образцов для электрофореза.
2.4 Электрофорез
Электрофорез проводился в 12,5% ПААГ (рис.9). Растворы для нанесения на
гель готовились по табл. 4. Ячейки геля заполнялись в соответствии со схемой
(табл. 5).
Табл. 4. Растворы для нанесения на гель
|
Анализируемый компонент
|
Количество компонента, мкл
|
Буфер для нанесения, мкл
|
Меркаптоэтанол
|
Деионизованная вода
|
Буфер для нанесения
|
20
|
|
|
|
|
2
|
Маркеры RainBow
|
5
|
|
|
|
|
3
|
BBY-препараты ФС-2
|
14
|
33
|
5
|
48
|
|
4
|
ФР 6
|
10
|
33
|
5
|
52
|
|
5
|
ФР7
|
15
|
33
|
5
|
47
|
|
6
|
ФР 11
|
10
|
33
|
5
|
52
|
|
7
|
ФР1.14
|
62
|
33
|
5
|
|
|
8
|
Фр1.19
|
62
|
33
|
5
|
|
|
9
|
ФС-2 после NaCl обработки
|
28
|
33
|
5
|
34
|
|
10
|
Маркеры PageRuler
|
5
|
|
|
|
Экспериментов по разделению белков было два, и нам удалось собрать
фракцию 1 дважды. В таблицах отражены обе фракции под номерами 1.14 и 1.19
(номера после точки соответствуют дате выделения).
Табл. 5.Схема заполнения геля
|
Буфер для нанесения, мкл
|
Маркеры RainBow, мкл
|
BBY-препа-раты ФС-2, мкл
|
ФР 6, мкл
|
ФР7,мкл
|
ФР 11, мкл
|
ФР1.14, мкл
|
ФР1.19, мкл
|
ФС-2 после NaClобработки,
мкл
|
Маркеры PageRuler, мкл
|
|
20
|
5
|
10
|
10
|
10
|
10
|
20
|
20
|
10
|
5
Рис. 9. Электрофорез в ПААГ
) Буфер для нанесения (контроль)
) МаркерыAmershamECLRainbow
- FullRange (Рис.11)концентрация белка
приблизительно 2 мг / мл, спектр определения молекулярной массы 12-225 кДа.
) BBY - препараты ФС-2 (описание в разделе
"Получение и характеристика препаратов ФС-2")
) ФР6 - Обнаружен белок PsbP с
молекулярной массой 23 кДа
) ФР7 - Белка не обнаружено
) ФР11 - примеси белков
,8) ФР 1.14 и 1.19 - Обнаружен белок PsbQ (17 кДа) c примесями других белков.
) Препараты ФС-2 после NaCl-обработки
) МаркерыFermentasPagerulerUnstainedProteinLadder (рис. 11)Спектр определения белка от 10 кДа до 200
кДa
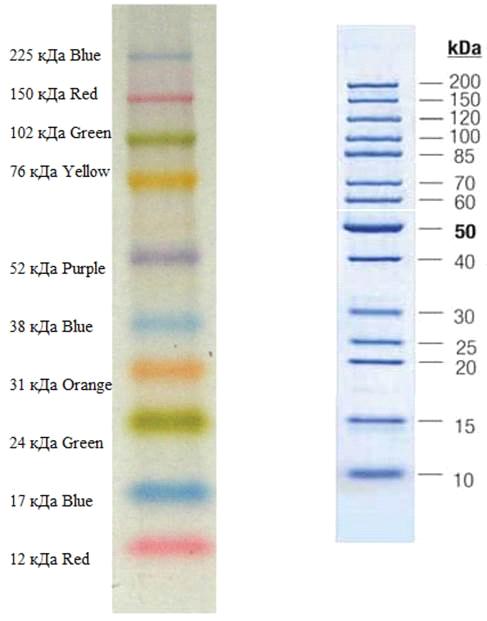
Рис.
11. Маркеры Amersham ECL Rainbow -
Full Range (слева), маркерыPageRuler Unstained Protein Ladder (справа)
Из результатов электрофореза видно (рис. 9; полосы геля 3 и 9), что нам
удалось полностью удалить белки PsbP и PsbQ из препаратов ФС-2 с помощью
обработки NaCl. Кроме того, нам удалось отделить
белки PsbP и PsbQ друг от друга при помощи ионообменной хроматографии (рис. 9
полосы геля 4, 7 и 8). Таким образом, полученные фракции белков можно
использовать для исследования их карбоангидразоподобной активности. К
сожалению, фракции изолированных белков PsbP и PsbQ содержат примеси: во
фракции белка PsbP по-видимому присутствует белок PsbS; а фракция белка PsbQ содержит некоторые белки ССК-2.Для
дальнейшей идентификации примесных белков необходимы другие методы и подходы,
но это не входило в цели данной работы. Кроме того, известно, что белки ССК-2
не обладают карбоангидразной активностью (Шитов с соавт., 2009) и,
следовательно, не могут помешать исследованию карбоангидразоподобной активности
белка PsbQ.
Итак, нам удалось разделить белки PsbP и PsbQ и мы могли производить на
них по отдельности дальнейшие исследования.
2.5 Определение концентрации белка по методу Брэдфорд
Для дальнейшей работы с белковыми фракциями было необходимо знать
концентрацию белка в них. Для её определения был выбран метод Брэдфорд.
Концентрация белка определялась исходя из данных по его оптической
плотности(табл.6) через уравнение графика калибровки (рис.12). Расчеты
произведенные по уравнению калибровочной кривой представлены в табл. 7.
Табл. 6.Данные оптической плотности (λ = 595 нм) растворов разных концентраций
бычьего сывороточного альбумина и исследуемых фракций белков.
|
Анализируемый компонент
|
Количество компонента в
анализируемом растворе, мкл
|
Оптическая плотность (А)
|
|
NaCl
|
150
|
0,0039
|
|
БСА (0.5 мг/мл)
|
5
|
0,457
|
|
7,5
|
0,619
|
|
10
|
0,786
|
|
12,5
|
0,831
|
|
15
|
0,906
|
|
17,5
|
0,979
|
|
20
|
0,923
|
|
ФР 6
|
5
|
0,282
|
|
|
0,274
|
|
ФР 1 (14.05)
|
5
|
0,054
|
|
|
0,053
|
|
ФР 1 (19.05)
|
5
|
0,149
|
|
|
0,134
|
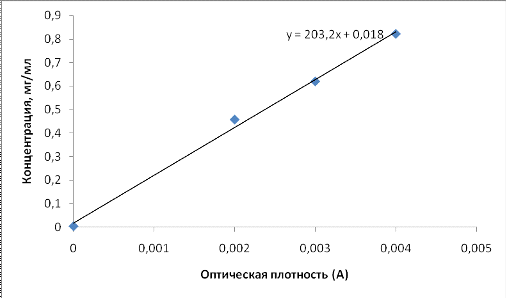
Рис.12. Калибровочная кривая
Табл.7.Конечная концентрация белка во фракциях
|
ФР 6
|
ФР 1 (14.05)
|
ФР1 (19.05)
|
|
Концентрация, мг/мл
|
0,3205
|
0, 0774
|
0,1402
|
2.6 Измерение карбоангидразоподобной активности белков PsbP и PsbQ
Как было сказано выше, КА-активность внешних белков ВОК (PsbP, PsbOи PsbQ)
не подавлялась ингибиторами карбоангидраз и поэтому активность обозначили как
карбоангидразоподобную (КП-активность).
Исходя из данных концентрации белка, полученных при помощи метода
Брэдфорд рассчитываем количество образца для измерения карбоангидразной
активности (табл. 8 и 9).
Измерение КП-активности для белка РsbР проводим по схеме описанной в табл.8, для белка РsbQ - в табл. 9. Также измеряем
КА-активность для ФС-2, чтобы сравнить её с активностью белков. Результаты
измерения КП-активности отражены на рис.13.
Табл. 8. Схема измерения КП-активности белка PsbP
|
Вероналовый буфер, мкл
|
HEPES (рН 7.5), мкл
|
PsbP, мкл
|
Раствор ионов металла (200
мкМ), мкл
|
Деионизованная вода,
насыщенная СО2, мкл
|
|
1
|
1472,8
|
|
|
750
|
|
2
|
1472,8
|
|
27,2
|
|
|
|
3
|
1450,3
|
|
27,2
|
22,5 (MnSO4)
|
|
|
4
|
1450,3
|
|
27,2
|
22,5 (ZnSO4)
|
|
|
6
|
1472,8
|
27,2
|
|
|
|
|
7
|
1450,3
|
|
27,2
|
22,5 (MgSO4)
|
|
|
8
|
1450,3
|
|
27,2
|
22,5 (CaCl2)
|
|
|
9
|
1472,8
|
27,2
|
|
|
|
|
10
|
1450,3
|
|
27,2
|
22,5 (CuSO4)
|
|
Табл. 9. Схема измерения КП-активности белка PsbQ
|
Вероналовый буфер, мкл
|
HEPES (рН 7.5), мкл
|
PsbQ, мкл
|
Раствор ионов металла (200
мкМ), мкл
|
Деионизованная вода,
насыщенная СО2, мкл
|
|
1
|
1447,69
|
52,31
|
|
|
750
|
|
2
|
1447,69
|
|
52,31
|
|
|
|
3
|
1425,19
|
|
52,31
|
22,5 (MnSO4)
|
|
|
4
|
1425,19
|
|
52,31
|
22,5 (ZnSO4)
|
|
|
6
|
1447,69
|
52,31
|
|
|
|
|
7
|
1425,19
|
|
52,31
|
22,5 (MgSO4)
|
|
|
8
|
1425,19
|
|
52,31
|
22,5 (CaCl2)
|
|
|
9
|
1447,69
|
52,31
|
|
|
|
|
10
|
1425,19
|
|
52,31
|
22,5 (CuSO4)
|
|
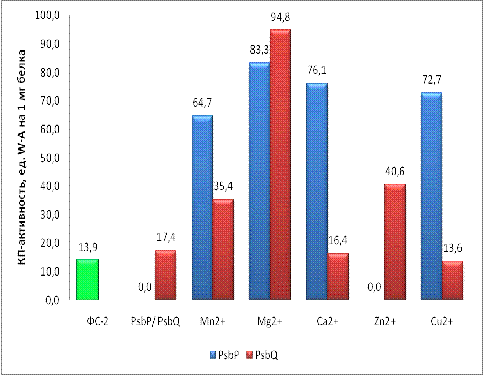
Ранее было показано, что КП-активность белка PsbP увеличивалась в присутствии Mn2+ (Шитов с соавт.,
2009). Однако, в этой работе белок PsbP содержал Mn в своём составе
и осталось неясным, будет ли обладать КП-активностью этот белок, если удалить
ионы Mn из белка. В нашей работе был получен
образец белка PsbP, не содержащего ионов металлов в
своём составе (для этого проводился диализ в присутствии ЭДТА). Нами было
выяснено, что обработанный ЭДТА белок не обладал КП-активностью, но добавление
ионов Mn2+ вызывало появление КП-активности этого белка. Как было показано
ранее, также ведёт себя белок PsbO (Шитов с соавт., 2009). Ионы Ca2+ и Mg2+ и
даже Cu2+ ещё более значительно увеличивали КП-активность белка PsbP,
максимальное значение зафиксировано в опыте с ионами Mg2+ (83,29). Эти данные
свидетельствуют о возможном сродстве белка PsbP к ионам двухвалентных металлов.
Это предположение подтверждается результатамими, опубликованными ранее в
иностранных научных статьях. Пока непонятно, насколько специфично действие
определённых металлов на КП-активность PsbP и как связано это явление с механизмом
функционирования и сборки ФС-2.Все эти вопросы требуют дальнейшего
исследования.
В присутствии ионов Zn2+ КП-активность PsbP не проявлялась. Пока
непонятно, с чем может быть связан этот результат. Возможно, играет какую-то
роль неспособность ионов цинка проявлять степень окисления больше двух (как
Mn), или этот ион имеет размеры, которые не подходят для мест связывания на
белковой молекуле. Для решения этоих вопросов требуются дополнительные
исследования.
Белок PsbQ, лишённый металлов, в отличии от белка PsbP, обладал
КП-активностью, которая была сравнима с КА-активностью ФС-2. В присутствии Mn2+
и Ca2+ его КП-активность увеличивалась примерно в 3 раза. Резкое увеличение
КП-активности наблюдалось с ионами Zn2+(94.77). Присутствие ионов Mg2+ и Сu2+
не приводило к увеличению КП-активности. Эти данные свидетельствуют о большем
сродстве PsbQ к ионам Mn2+, Ca2+ и особенно Zn2+. Подтверждением этого сродства
могут быть данные о важности белков PsbPи PsbQдля сохранения ионов Caи Cl в составе ВОК ФС-2. Косвенным подтверждением сродства Zn к PsbQ может являться тот факт, что в ФС-2 ранее
обнаруживали присутствие ионов Zn2+ (Шитов с соавт., 2009).Однако, пока
непонятна роль этого связывания в функционировании ФС-2. Необходимо отметить,
что ионы цинка могут связываться не только с PsbQ, но и с другими белками, которые входят в состав
выделенной нами фракции белка PsbQ.Для
подтверждения связывания ионов Zn (а
также Caи Mn), необходимо дальше очистить эту фракцию с применением
других физико-химических методов (например, катионнообменной хроматографии) с
целью получения белка PsbQв
чистом виде и проведения аналогичных экспериментов с КП-активностью.
Так как ионы Zn2+ и Ca2+ обладали наибольшим влиянием на КП-активность,
необходимо было определить зависимость активности от концентрации этих ионов
(рис.14).
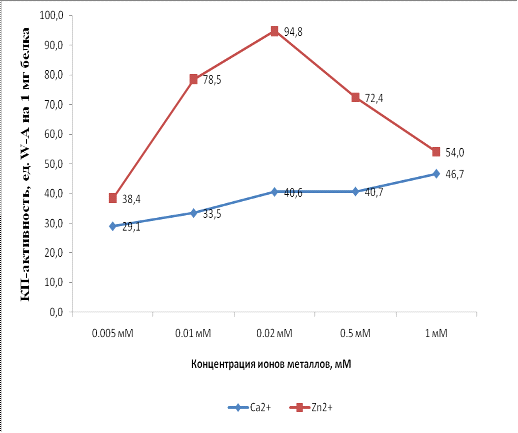
Из графиков видно, что зависимости КП-активности PsbQ от концентраций
разных ионовзначительно отличаются. На графике концентрационной зависимости для
ионов Zn2+ прослеживается пик активности (94,8 ед. W-A/мг белка при 200 мкМ
Zn), а в присутствии высоких концентраций (1мМ) КП-активность уменьшается. Ионы
Ca2+ действуют иначе, активность линейно возрастает по мере увеличения
концентрации иона. По-видимому, это связано с различной природой взаимодействий
этих ионов с белком. Причины различий в действии этих металлов пока не ясны и
этот вопрос требует дальнейшего изучения.Наличие оптимума концентрации
двухвалентного металла характерно для металлоферментов (в том числе и
карбоангидраз) и это может быть либо признаком того, что белок PsbQявляется карбоангидразой нового
класса, либо признаком присутствия посторонней карбоангидразы во фракции белка PsbQ.В любом случае, эта работа ещё не
завершена и поставленные нами вопросы требуют дальнейшего исследования.
VI.
ЗАКЛЮЧЕНИЕ
В настоящее время наличие карбоангидразной активности в ФС-2 высших
растений (горох, шпинат, пшеница, кукуруза) показано многими исследователями и
не должно вызывать сомнений. Но пока неясно, какова природа носителя
карбоангидразной активности в ФС-2. Им может быть один (или несколько) из
известных белков ФС-2, обладающий неизвестной ранее функцией (карбоангидразной
активностью), или неизвестный белок.Принимая во внимание значимость
карбоангидразной активности для функционирования донорной стороны ФС-2, особое
внимание было уделено исследованию карбоангидразной активности внешних
водорастворимых белков водоокисляющего комплекса. В данной работе впервые был
проведен анализ действия ионов двухвалентных металлов на карбоангидразоподобную
активность белков РsbРи РsbQпо отдельности.
Было выяснено, что белок PsbPне
обладал КП-активностью пока к нему не добавляли ионы двухвалентных металлов.
Особенно эффективным активатором КП-активности оказался ион Mg2+, несколько менее эффективными -
ионы Ca, Cu, Mn. Вопрос о специфичности
ионов металлов в активации КП-активности PsbP пока остаётся открытым.
Было показано, что очищенный от ионов металлов белок PsbQобладал КП-активностью, которая
сильно возрастала в присутствии ионов Zn2+, в присутствии ионов Caи Mn КП-активность увеличивалась с в два
раза меньшей эффективностью. Также было выявлено, что Zn и Ca
по-разному влияют на увеличение КП-активности, что может свидетельствовать о
разной природе взаимодействий ионов этих металлов с белком PsbQ. Природу этих взаимодействий необходимо
более тщательно исследовать, поскольку это важно для понимания закономерностей
функционирования фотосистемы 2. В силу актуальности этой тематики, эта работа
требует продолжения.
Роль обнаруженных носителейкарбоангидразной активности, находящихся в люменальной
части ФС-2, в непосредственной близости к ВОК, может быть важна для
фотосинтетического окисления воды, подобно функциональной активности
карбоангидразы cah3, обнаруженной
ранее в составе «ядерного» комплекса ФС-2 клеток C. reinhardtii и
необходимой (наряду с анионом бикарбоната) для формирования, стабилизации и
функционирования Mn2+-содержащего
водоокисляющего комплекса (Villarejoetal., 2002;Shutovaetal., 2008).
VII. ВЫВОДЫ
. Фрагменты тилакоидных мембран хлоропластов, обогащенных фотосистемой 2,
выделенные из листьев гороха, обладали хорошей фотохимической активностью и по
своему белковому составу соответствовали препаратам фотосистемы 2, описанным
ранее в научной литературе.
. Выбранные нами методы и подходы позволили выделить изолированные фракции
белков PsbPи PsbQс сохранением их КП-активности.
.Выявлено, что изолированный и очищенный от металлов белок PsbP не
обладал карбоангидразоподобной активностью. Эта активность проявлялась только в
присутствии ионов двухвалентных металлов, таких как Mn2+, Ca2+, Mg2+, Cu2+. Ионы Zn2+не
вызывали увеличения КП-активности.
. Белок РsbQобладал
КП-активностью без добавления ионов двухвалентных металлов. Однако, его
активность значительно возрастала в присутствии ионов Zn2+, Ca2+ и
Mn2+.Было выяснено, что ионы Zn2+и Ca2+ по-разному воздействуют на карбоангидразоподобную
активность белка PsbQ.
VIII.
СПИСОК ЛИТЕРАТУРЫ
Antal
T. K., Venediktov P. S., Konev Yu. N., Matorin D. N., Hapter R., and Rubin A.
B. (1998) Assessment of Vertical Profiles of Phytoplankton Photosynthetic
Activity by the Fluorescence MethodD.I. (1949) Copper Enzymes in Isolated
Chloroplasts.Polyphenoloxidase in Beta vulgaris.Plant Physiology 24, 1-15.K.
Shanker (2008) Trace elements: Nutritional benefits, environmental
contamination and health; 21 Modeofaction and toxicity of trace elements,Edited
by M.N.V. PrasadJ. (2002)P680: what is it and where is it?,
Bioelectrochemistry. Jan;55(1-2):135-8.J., Chapman D.J. and Telfer A. (1987)
Characterization of a PSII reaction center isolated from the chloroplasts of
Pisum sativum. FEBS Lett. v.220, p.67-73.
Basics
G. H. Krause E. Weis (1991) Сhlorophyll
fluorescence and photosynthesisM. M. (1976) A Rapid and Sensitive Method for
the Quantitation of Microgram Quantities of Protein Utilizing the Principle of
Protein-Dye Binding
Dai
X., Yu Y., Zhang R., Yu X., He P. and Xu C. (2001) Relationship among
Photosystem II carbonic anhydrase, extrinsic polypeptides and manganese
cluster. Chinese Science Bulletin46, 406-408.A., Kern J., Gabdulkhakov A.,
Broser M., Zouni A., Saenger W. (2009) Cyanobacterial photosystem II at 2.9 A
resolution: role of quinones, lipids, channels and chloride. Nat. Struct. Mol.
Biol. 16, 334-342.D., Gho Y.S., Suh M. and Kang C. (2002) Highly Sensitive and
Fast Protein Detection with Coomassie Brilliant Blue in Sodium Dodecyl
Sulfate-Polyacrylamide Gel Electrophoresis.Bulletin of the Korean Chemical
Society23, 1511-1512.M.S., Ignatova L.K., Rudenko N.N., Ivanov B.N. and Klimov
V.V. (2004) Photosystem II associated carbonic anhydrase activity in higher
plants is situated in core complex. FEBS Letters577, 305-308.V.V.,
Allakhverdiev S.I., Shuvalov V.A. and Krasnovsky A.A. (1982) Effect of
extraction and re-addition of manganese on light reactions of photosystem-II
preparations.FEBS Letters148, 307-312.U.K. (1970) Cleavage of structural
proteins during the assembly of the head of bacteriophage T4.Nature227,
680-685.Y.K. and Stemler A.J. (2007) Differing responses of the two forms of
photosystem II carbonic anhydrase to chloride, cations, and pH.Biochimica et
Biophysica Acta (BBA) - Bioenergetics 1767, 633-638.Y.K., Theg S.M. and Stemler
A.J. (2005) Carbonic anhydrase activity of the photosystem II OEC33 protein
from pea.Plant and Cell Physiology 46, 1944-1953.I.L., Badger M.R., Wydrzynski
T. and Hillier W. (2007) A quantitative assessment of the carbonic anhydrase
activity in photosystem II.Biochimica et Biophysica Acta (BBA) - Bioenergetics
1767, 639-647.J.V., Ma Y., Frey W.D., Fusilier K.A., Pham T.T., Simms T.A.,
DiMario R.J., Yang J.and Mukherjee B. (2011) The carbonic anhydrase isoforms of
Chlamydomonas reinhardtii: intracellular location, expression, and
physiological roles. Photosynthesis Research109, 133-149.O.V., Razguljayeva
A.Y., Shutova T.V., Khristin M.S., Ivanov B.N. and Klimov V.V. (1999) Carbonic
anhydrase activity of different Photosystem II preparations. In: Garab G. (ed.)
Photosynthesis: Mechanism and Effects, Vol. 2, pp. 1201-1204. Kluver Academic
Publishers, Dordrecht.G. (1992) Energy transfer and trapping in photosystem II.
-In: Topics in photosynthesis, the photosystems: structure, functions and
molecular biology. (ed.: Barber J.), Elsevier, Amsterdam, 45-99.F., Glockner
C., Hellmich J., Zouni A., (2012) Light-induced quinone reduction in
photosystem II. Biochimica et Biophysica Acta 1817, 44-65.N.N., Ignatova L.K.
and Ivanov B.N. (2007) Multiple sources of carbonic anhydrase activity in pea
thylakoids: soluble and membrane-bound forms. Photosynthesis Research 91,
81-89.T., Nikitina J., Deikus G., Andersson B., Klimov V. and Samuelsson G.
(2005) Structural dynamics of the manganese-stabilizing protein-effect of pH,
calcium, and manganese. Biochemistry44, 15182-15192.A. (1986) Carbonic
anhydrase associated with thylakoids and Photosystem II particles from maize.
Biochimica et Biophysica Acta (BBA) - Bioenergetics850, 97-107.A., Shutova T.,
Moskvin O., Forssen M., Klimov V.V. and Samuelsson G. (2002) A photosystem
II-associated carbonic anhydrase regulates the efficiency of photosynthetic
oxygen evolution. EMBO Journal21, 1930-1938.K.M. and Anderson N.G. (1948)
Electrometric and colorimetric determination of carbonic anhydrase.The Journal
of Biological Chemistry176, 147-154.
Гольцев В.Н.,
Каладжи М.Х., Кузманова М.А., Аллахвердиев С.И.(2014). Переменная и замедленная
флуоресценция хлорофилла a - теоретические основы и практическое приложение в
исследовании растений . Ижеск-Москва: Институт компьютерных исследований.
Казимирко Ю.
В. Разработка флуорометрических методов оценки состояния фотосинтетического
аппарата для биоиндикации среды: диссертация ... кандидата биологических наук :
03.00.02, 03.00.16. - Москва, 2006. - 117 с.
Ланкин А.В.,
Креславский В.Д., Худякова А.Ю., Жармухамедов С.К., Аллахвердиев С.И. (2014)
Влияние нафталина на фотохимическую активность фотосистемы 2
Лукаткин А.
С., Ревин В. В., Башмаков Д. И., Кренделева Т. Е., Антал Т. К., Рубин А. Б.
(2009), Экологическая оценка состояния древесных растений г. Cаранска по флуоресценции хлорофилла
Малиновский
В. И. (2004) Физиология растений, Владивосток: ДВГУ
Остерман, Л.
(1981). Методы исследования белков и нуклеиновых кислот: электрофорез и
ультрацентрифугирование. Москва: Наука.
Полевой В. В.
(1989) Физиология растений, Москва: Высшая школа
Холл Д., Рао
К. (1983) Фотосинтез, Москва: Мир
Шитов, А. В.
Исследование карбоангидразной активности фотосистемы 2 гороха:диссертация ...
кандидата биологических наук:03. 01. 04- Пущино, 2013. - 116 с.
Якушкина И.
И. (2004). Физиология растений, Москва: Владос
Яныкин, Д. В.
Фотопоглощение молекулярного кислорода на донорной стороне фотосистемы 2 в
субхлоропластных мембранных препаратах с разрушенным водоокисляющим комплексом:
диссертация ... кандидата биологических наук:03. 01. 04- Пущино, 2013. - 133с.
Похожие работы на - Влияние ионов металлов на карбоангидразоподобную активность внешних водорастворимых белков PsbP и PsbQ фотосистемы 2
|