Биологические особенности кубанского рака
Биологические
особенности кубанского рака
Введение
Целью данной работы являлось изучение особенностей биологии кубанского
рака в Ахтарско-Гривенских лиманах.
В водоемах Северного Кавказа обитают пресноводные раки двух видов -
длиннопалый кубанский рак и широкопалый рак. Первый вид широко распространен и
имеет важное промысловое значение. Значимость длиннопалого кубанского рака
заключается в большей массе и большем количестве белка в массе тела [Черкашина,
1996].
Исследования биологии и плодовитости кубанского рака позволяют пополнить
сведения об этом промысловом виде, а так же позволяют дать оценку численности
популяции.
Основные задачи, поставленные для выполнения цели:
привести общую биологическую и экологическую характеристику кубанского длиннопалого
рака;
привести характеристику Ахтарско-Гривенских лиманов;
исследовать размерно-массовые показатели, возрастную и половую структуры
длиннопалого кубанского рака, выловленных в месте исследования;
описать плодовитость и дать оценку качества половых продуктов самок
длиннопалого кубанского рака;
провести исследования по питанию длиннопалого кубанского рака;
оценить экологическую обстановку в Ахтарско-Гривенских лиманах, как
среды, пригодной для обитания длиннопалого кубанского рака;
дать рекомендации по способам повышения численности популяции
длиннопалого кубанского рака в Ахтарско-Гривенских лиманах.
Исследования по биологическим особенностям и численности длиннопалого
кубанского рака проводятся практически ежегодно, но в основном осуществляются ФГУП
«АзНИИРХ» в водоемах Ростовской области, так как именно там сосредоточены
основные промысловые запасы (Веселовское, Пролетарское и Усть-Манычское
водохранилища), река Дон, включая водоемы поймы и бассейн реки Сал (реки Сал,
Кара-Сал, Джурак-Сал, Куберле, Акшибай) [Черкашина, 1981].
По оценкам последних исследований [Москул, 2005] численность кубанского
длиннопалого рака увеличилась, что говорит об улучшении экологической ситуации
в рыбохозяйственных водоемах Краснодарского края. Популяции рака были отмечены
в Варнавинском водохранилище и системе Ахтарско-Гривенских лиманов, однако до
сих пор главными проблемами, влияющими на численность популяции длиннопалого
кубанского рака, остаются браконьерский промысел и вылов молоди раков.
В настоящее время по ряду причин, промысел длиннопалого кубанского рака в
Краснодарском крае ведется лишь в системе Ахтарско-Гривенских лиманах:
Бойкиевский (994 шт./га), Пригиевский (752шт./га), Пимониевский (752 шт./га),
Ахтарский (694 шт./га) [Черкашина, 1990].
биологический рак лиман
1. Аналитический обзор
.1 Исследования об особенностях биологии кубанского рака
Центром происхождения раков семейства Astacidae, по мнению многих ученых,
является Понто-Каспийский бассейн [Мордухай-Болтовской, 1946]. Род Pontastacus имеет большой ареал, в котором
каждый из видов занимает ряд постоянных, порой удаленных друг от друга водоемов
с характерными условиями. Представители этого рода широко распространены в
бассейнах Черного и Азовского морей. Им свойственна значительная изменчивость,
что подтверждается образованием многих видов и рас [Бродский, 1967;
Старобогатов 1995; Скориков 1911]. Среди видов рода Pontastacus особое место занимает кубанский
длиннопалый рак (Pontastacus cubanicus Birst et Winog, 1934 syn. Astacus leptodactylus cubanicus).
Является промысловым видом, ежегодно его запасы оцениваются ФГУП
«АзНИИРХ». Утвержденные, согласно исследованиям биологии, ОДУ кубанского
длиннопалого рака достигали 33 т для водоемов Ростовской области и 3,37 т для
водоемов Краснодарского края (по официальным данным ОДУ ФГУП «АзНИИРХ»).
Исследования биологии раков для некоторых водоемов Кубани проводились в
разные периоды [Москул, 2005; Черкашина 1991, 1994, 1997]. Результаты
исследований показывали, что в целом, распространение популяции раков говорило
о благоприятной экологической обстановке в рыбохозяйственных водоемах, но
численность популяции все равно оставалась на невысоком уровне. Каких-либо
отклонений в биологии и развитии выявлено не было.
Широко распространен в водоемах Ростовской области, в меньшей степени - в
водоемах Краснодарского края (рисунок 1).
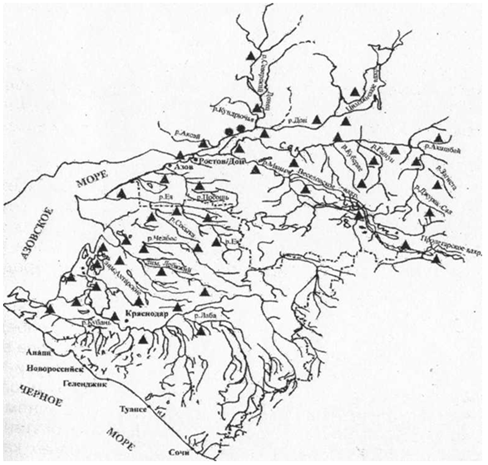
Рисунок 1 - Распространение кубанского рака в водоемах Краснодарского
края
биологический рак лиман
Основной задачей исследований прошлых лет была оценка численности и
условий обитания популяции длиннопалого кубанского рака (оценка качества воды,
кормовая база). Были сделаны выводы, о том, понтичный кубанский рак является
хорошим объектом для культивирования в искусственных условиях. Так же, были
получены следующие данные о биологии длиннопалого кубанского рака [Черкашина,
1990]:
отличительным признаком данного вида является ширина клешни,
пропорциональное соотношение отдельных частей тельца, большой размер;
мышцы содержат до 15% белка от сырого веса или 70-90% белка от сухого
остатка;
обладает быстрым ростом, высокими товарными и вкусовыми качествами;
средняя длина 12,5, для самцов - 12,7, для самок - 12,4 см;
средняя масса 59, для самцов - 63, для самок - 54 г;
средняя плодовитость самок - 290 икринок, число личинок - 150-2000 шт.
Было так же отмечено, что данные показатели могут изменяться в
зависимости от интенсивности промысла, флюктуации поколений и других
биотических и абиотических факторов [Александрова, 1994].
Результаты изучения генетической структуры [Бродский, 1974], а так же
биометрической обработки материалов по изменчивости ряда популяций кубанского
рака указывают на некоторую обособленность его от остальных форм. Кубанский рак
является типичной формой Азовского бассейна, а эстуарии Дона и Кубани - его
резервации. Этот вид имеет ареал в водоемах Нижнего Дона, Краснодарского края,
а его дунайская раса - в придунайских озерах. В последние годы исследования его
обнаружили в реке Ингулец - правом притоке Днепра, очевидно, что он был завезен
туда из Краснодарского края. Встречается на всех грунтах, кроме заиленных,
очень требовательны к кислородному режиму. Представители рода Pontastacus при
снижении его до 2,9 мг/л обычно выползают на берег, а при 0,37 мг/л - погибают.
Так, в Ахтарско-Гривенских лиманах (Малый Карпиевский, Сухой, Кривой, Орлиный,
Замарайков) в августе 1980 г. наблюдалась гибель кубанского рака в результате
резкого снижения содержания кислорода в воде до 1,25-1,85 мг/л, что явилось
следствием гниения водной растительности при высокой температуре [Черкашина,
1980]. Погибает этот вид при содержании кислорода в воде 2,23 мг/л (30 %
насыщения).
Всем ракам семейства Astacidae свойственно неравномерное заселение
экотопов, связанное с особенностями их жизненного цикла, кормовых организмов и
другими биотическими и абиотическими факторами [Бирштейн, 1940].
Сезонные перемещения скоплений раков при относительно стабильной
экологической ситуации не отмечены [Бирштейн, 1934]. При стабильной
экологической ситуации основные скопления раков приурочены к местам с высокой
биомассой 200-1000 г/м2 кормовых организмов, обычно зообентоса.
Так, в 2003-2004 гт. наиболее высокая их плотность была зарегистрирована
в западной части лимана (372 и 103 шт./га, соответственно), в 1992 г. - в
восточной (144 шт./га), в 1993 - в центральной (155 шт./га). В 2005 г. раки
распределились практически равномерно по всему водоему с плотностью 83 шт./га,
колеблясь от 98 (западная часть) до 73 шт./га (восточная), что связано, прежде
всего, с кормовой базой, и главным образом - с биомассой хирономид, которые
составляли 95,1 % от «мягкого» бентоса [Будников, 2005].
Однако абсолютные уловы раков по сезонам меняются достаточно заметно, что
объясняется их физиологическим состоянием. Кубанский рак в летний период
активен только в течение месяца после линьки, то есть в середине июня-июля. В
этот период уловы раков практически мало отличаются от осенних, и скопления их
приурочены к тем же местам, что и осенью. Десятигодовые динамики выловов были
неравномерны и колебались (рисунок 2).
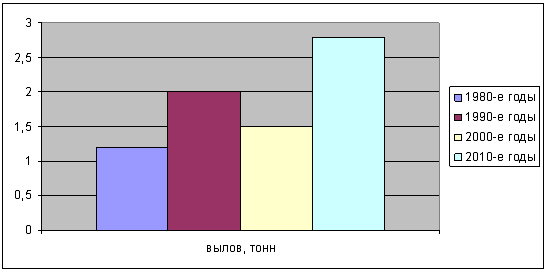
Рисунок 2 - Динамика вылова кубанского рака
Как видно из рисунка 2, максимальные размер выловов были достигнуты лишь
в 2010-е годы. В 2000-е годы наблюдалось снижение объемов вылова, это
объясняется снижением численности популяции кубанского рака из-за
неблагоприятной экологической обстановки в водоемах Краснодарского края
[Москул, 2005].
Исследования по изучению биологии, численности и запасов длиннопалого
рака так же проводили в 2002-2005 годах [Москул, 2005]. Для лова раков использовали
стандартные раколовки, ставные сети и 25-метровый мальковый невод. В раколовках
преобладали раки длиной 8-11 см, массой 40-85 г. В уловах ставными сегями
встречались особи длиной 6-9 см, массой 20-45 г. В неводных уловах, проведенных
в прибрежной части водохранилища, преобладали раки длиной 5-7 см, массой 15-35
г. Половозрелость кубанского рака наступает на третьем году жизни при длине 7-8
см. К концу второго года жизни при хороших условиях нагула у 75-80 % самок на
плеоподах была развивающаяся икра. Количество икры на плеоподах колебалась от
130 до 268 икринок. Спаривание раков начинается в конце сентября. Длина
половозрелых самок колеблется от 8 до 14 см, масса 35-110 г. Самки длиной 14 см
встречаются крайне редко. Наиболее активно принимают участие в размножении
самки длиной 9-11 см. Выклев личинок происходит в апреле - мае при температуре
воды 16-20°С. Средняя длина выклюнувшихся рачат - 8-9 мм, масса 20-25 мг.
Первая линька личинок происходит на 5-7-й день после вылупления, вторая - через
10-15 дней после первой. К осени длина сеголеток длиннопалого рака в
Варнавинском водохранилище колеблется от 3,8 до 4,7 см, масса от 3,5 до 5,1 г.
Молодь раков (1,1-1,9 см), перешедшая к самостоятельному образу жизни, питается
мелкими формами зоопланктона. По мере роста в рационе сеголетков доля дафний
уменьшается, а возрастает потребление личинок хирономид, бокоплавов, моллюсков,
а также личинок рыб. Хирономиды являются излюбленной пищей раков в Варнавинском
водохранилище и встречаются в желудках у всех возрастных групп. Популяция
длиннопалого кубанского рака представлена в Ахтарско-Гривенских лиманах всеми
размерными группами от 4 до 14 см. Преобладают средние возрастные группы длиной
тела от 7 до 9 см (50-70 %). Крупные раки длиной тела 12-14 см малочисленны (2-7
%). Неполовозрелые раки длиной до 7 см составляют от 28 до 43 %. Исследования
показали, что в 2005 году на 1 га площади нагула (2500 га) обитало 42,5 шт.
раков. Из них промысловых размеров, свыше 9,5 см - 26,3 шт./га. Общий запас
кубанского речного рака рассчитан на уровне 5,39 т (213,24 тыс.шт.).
Промысловый запас - 3,37 т (74,89 тыс.шт.). Увеличение численности раков может
быть достигнуто при улучшении общей экологической обстановки водоемов,
проведении в широких масштабах работ по культивированию и охране речных раков.
В целом, вид Pontastacus cubanicus является распространенным для
водоемов Краснодарского края, его биология изучалась на протяжении долгого
периода времени, накоплено достаточно сведений, которые возможно сравнить с
исследованиями настоящего времени и получить выводы об изменении в популяции
кубанского длиннопалого рака.
1.2 Исследования о состоянии Ахтарско-Гривенских лиманов
Согласно перечню водно-болотных угодий РФ место исследования относится к
группе лиманов между рекой Кубань и рекой Протокой и Ахтарско-Гривенской
системой лиманов Восточного Приазовья (рисунок 3). Эти группы представляют
собой единую систему, разделенную формально, поэтому их гидрологическая и
гидрохимическая характеристика дается совместно. Ахтарско-Гривенские лиманы
относятся к категории водно-болотных угодий регионального значения
Краснодарского края ЮФО. Общая площадь 173 тыс. га (группа лиманов между рекой
Кубань и рекой Протокой - 88 400 га; Ахтарско-Гривенская система лиманов: 84
600 га) [Лотышев, 2006].
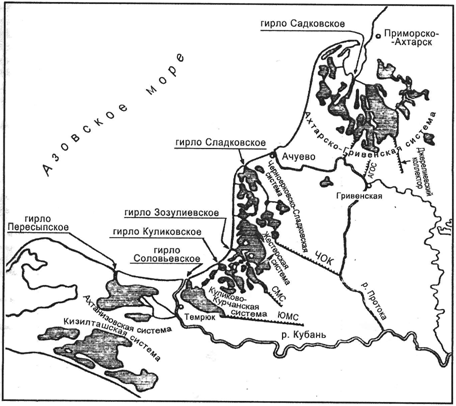
Рисунок 3 -
Место исследования: Ахтарско-Гривенские лиманы
Функции данной системы лиманов многочисленны
управление водно-болотными угодьями в целях оптимизации среды обитания
водоплавающих и околоводных птиц во все биологические периоды их годового
цикла;
охрана и рациональное использование обитающих водоплавающих птиц и других
животных;
поддержание оптимального или естественного гидрологического режима в
различные сезоны года, проведение мелиоративных и биотехнических мероприятий с
целью улучшения среды обитания водоплавающих и околоводных птиц, проведение
мероприятий по восстановлению нарушенного естественного гидрологического
режима;
сохранение высокого биологического разнообразия экосистем, высокой
продуктивности птиц и других животных, оптимального произрастания водной и
прибрежной растительности как кормовой базы и защитных условий для птиц и
других животных;
поддержание естественных и создание искусственных мест для гнездования,
кормежки и отдыха птиц.
Угодье находится в Краснодарском крае, на территории административных
районов: Приморско-Ахтарского, Славянского и Темрюкского и удалено на 3-56 км
от районных центров - городов Приморско-Ахтарска, Славянска-на-Кубани, Темрюка.
От краевого центра, г.Краснодара, удалено на 142 км. Угодье включает большую
часть современной дельты Кубани. Южная его граница идёт по берегу Курчанского
лимана, охватывая устье р. Кубань и выходит к Азовскому морю. Западная и
северо-западная морская граница пролегает вдоль по морю на удалении 500 м от
берега и выходит к середине лимана Ахтарский. По восточному побережью лимана
Ахтарский граница подходит к хутору Садки и, захватывая лиман Кирпильский,
уходит на юг. Восточная граница Рамсарского угодья идёт в основном по границе
рисовых систем с лиманами через пос. Слободка, Черноерковское, Свистельников,
Курганская [Воловик, 1996].
Климат
умеренно-континентальный с относительно мягкой зимой, среднемесячными
температурами в январе в пределах от -1 до -8 0С и жарким летом с температурами
в июле от +24 до +28 0С (рисунок 4).
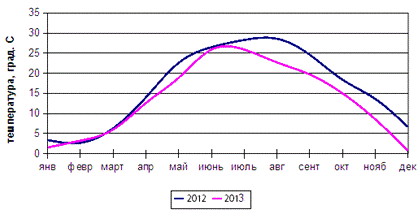
Рисунок 4 - Динамика температур 2012-2013 гг.
Наиболее крупные лиманы данной системы - Кирпильский и Ахтарский лиманы
[Макаров, 1996].
Ахтарский лиман - входит в одноименную группу Ахтарско-Гривенской системы
Кубанских лиманов. Площадь 65 кв. км, максимальная глубина 2 м. Ахтарский лиман
- типичная береговая лагуна, соединенная с морем широким проходом. Он
образовался несколько столетий назад в результате частичного отчленения
(Ачуевской косой) устья правого рукава Кубани от Азовского моря. Берега лимана
сформированы речными и морскими отложениями. На значительном протяжении они
низменны и заболочены. Лишь в районе Приморско-Ахтарска берегом лимана является
трехметровый обрыв коренной суши [Лотышев, 2006].
Кирпильский лиман - водоем в одноименной группе Ахтарско-Гривенской
системы Кубанских лиманов. Некоторыми авторами делится на две части: южную
(Большой Кирпильский лиман) и северную (Малый Кирпильский лиман). Площадь
акватории 61 кв. км, максимальная глубина 1,8 м. Прибрежная полоса густо
покрыта влаголюбивой растительностью. В настоящее время водоснабжение его
осуществляется через Джерелиевский главный коллектор и Гривенский
опреснительный канал. Кирпильский лиман - одно из рыбопромысловых угодий
Ахтарско-Гривенской системы [Лотышев, 2006].
Мелководность лиманов способствует быстрому прогреву и охлаждению вод под
воздействием температуры окружающей среды.
В зимний период зона лиманов находится под влиянием азиатского
антициклона и средиземноморских циклонов. Летом проявляется воздействие отрогов
субтропических областей повышенного давления, с выносом воздуха из Малой Азии.
В течение года преобладают ветра восточного и западного направлений.
Вода в Ахтарско-Гривенские лиманы поступает по Гривенскому шлюзу. В
среднемноголетнем разрезе величины атмосферных осадков составляют 45-50% от
величины испарения. Характерной особенностью лиманов является их мелководность,
обусловленная историческим развитием кубанской дельты, быстрая прогреваемость,
высокая кормность и благоприятно сложившийся водно-солевой режим.
Гидрохимический режим водоемов лиманов формируют два основных фактора:
температурный фон (который связан с изменениями температуры воздуха) и
гидрологический режим (количество, качество поступающей воды, сроки ее
поступления и связь с морем). Эти факторы влияют на общую минерализацию вод и
ее ионный состав.
Содержание хлора в нерестовый период во всех обследованных лиманах было в
пределах благоприятных величин, общая минерализация в пределах 0,7-2,1 г/л.
Наблюдалось уменьшение содержания в воде сульфатов и, на отдельных станциях,
увеличение ионов щелочных металлов.
Концентрация гидрокарбонатов колебалась в пределах206-269 мг/л. Ни в
одном лимане содержание гидрокарбонатных ионов не было ниже пороговой величины
(180 мг/л), что является важным положительным фактором для поддержания
стабильных величин активной реакции (рН) и газового режима.
Особенностью 2012-2013 года является увеличение концентрации ионов Са2+,
что благоприятно для роста и развития молоди рыб и раков. Отношение Са2+ к Mg2+
было максимальным в мае (2,13-2,30), с колебаниями от 0,28 до 1,02. В весенний
период рН воды лиманов оставался стабильным (8,0-8,5), что было на уровне
значений последних лет.
Основными формами неорганических соединений азота являются:N-NH4+;
N-NO2-, N-NO3-. Все соединения азота оказывают большое влияние на
продуцирование органического вещества в водоемах.
Из минеральных форм наибольшее биопродукционное значение имеет нитратный
и аммонийный азот. Доминирующим биогенным элементом из триады минерального
азота (N-NH4; N-NO2; N-NO3) весной были нитраты.
Средние значения содержания нитратов колебались от 0,009 до0,09 мг/л.
Концентрация ионов аммония не превышала ПДК (0,08 мг/л) во всех водоемах в
течение всего вегетационного периода. Содержание нитритов находилось на уровне
от 0,001 до 0,032 мг/л. Превышений ПДК (0,3 мг/л) зафиксировано не было.
Фосфор также является одним из важнейших элементов минерального питания.
Вместе с другими биогенными ингредиентами он оказывает существенное влияние на
биопродуктивность водоемов. В текущем году содержание минерального фосфора
колебалось от 0,018 до 0,16 мг/л.
Биомасса фитопланктона колеблется от 4,86 до 24,8 г/м3, составляя в
среднем за вегетационный сезон 13,9 г/м3 [Тевяшова, 1988, 1998].
Зоопланктон представлен тремя типичными планктонными группами организмов:
коловратками, веслоногими и ветвистоусыми рачками.
Наблюдения за сезонной динамикой зоопланктона, в период с апреля по
октябрь, показали, что он характеризуется резко выраженной периодичностью.
Биомасса зоопланктона весной (апрель - май) колеблется от 3,8 до 5,6
г/м3, летом происходит резкий спад (0,5-0,8 г/м3) и только к середине сентября
биомасса зоопланктона вновь начинает расти (1,2 - 1,6 г/м). Максимальное
развитие зоопланктона отмечается в мае (4,6 - 5,6 г/м3). Численность
зоопланктона колеблется от 12 до 320 тыс. экз./м3, а биомасса составляет 0,5 -
5,6 г/м3 , в среднем за вегетационный сезон - 2,3 г/м3.
В зообентосе отмечено наличие пяти групп организмов (личинки хирономид,
олигохеты, личинки жуков, мизиды, гаммариды). Наибольшее развитие достигают
личинки хирономид, биомасса которых колеблется от 0,4 до 3,5 г/м2.
Средневегетационная биомасса зообентоса составляет 1,89 г/м2.
Ихтиофауна представлена 26 видами рыб и кубанским раком. Промысловое
значение имеют сазан, лещ, карась, белый и пестрый толстолобики, белый амур,
плотва, судак, а также длиннопалый кубанский рак.
1.3 Промысловое и экологическое значение кубанского рака
Рак относится к нерыбным промысловым запасам.
В последние годы наблюдается увеличение промыслового запаса длиннопалого
кубанского рака, что свидетельствует о благоприятных экологических условиях
среды обитания и эффективной борьбе с браконьерским выловом немерного рака
(молоди). Однако, в ходе различных исследований [Черкашина, 1977; 1983, 1986]
отмечено, что по-прежнему в промысловых водоемах наблюдаются браконьерские
орудия лова.
Так же, вызывает беспокойство тот факт, что зарегулирование системы
Ахтарско-Гривенских лиманов ведет к обмелению и заиливанию водоемов, вследствие
чего возможно падение численности популяции [Черкашина, 1977; 1983, 1986].
По данным Москула Г.А. (2005) экологическая ситуация рыбохозяйственных
водоемов бассейна реки Кубани (водохранилища, лиманы, пруды руслового типа,
построенные на малых степных реках) в последние годы заметно улучшилась.
Промышленные, коммунальные предприятия и особенно сельское хозяйство,
включая рисоводство, стали меньше применять и, соответственно, меньше
сбрасывают загрязненные вещества, ядохимикаты, тяжелые металлы в
рыбохозяйственные водоемы. Это подтверждается тем, что практически во всех
водоемах стал обитать длиннопалый кубанский рак - Pontastacus cubanicus [Birst et Winoqr., 1934].
Если до 1992-1994 гг. кубанский рак встречался только в некоторых лиманах
и водоемах, расположенных на степных реках (Ея, Бейсуг, Челбас, Кирпили и их
притоках), то в настоящее время популяции кубанского длиннопалого рака
встречаются во всех приазовских лиманах, в водоемах, расположенных на степных
реках, а также в Краснодарском, Крюковском, Варнавинском, Шапсугском,
Шенджийском, Тахтамукайском и других малых водохранилищах бассейна реки Кубани.
В приазовских лиманах, водоемах, расположенных на степных реках ведется
промышленный лов рака [Москул, 2005].
2. Биологическая характеристика кубанского рака
.1 Систематическое положение
Длиннопалый кубанский рак, согласно современной систематике [Черкашина,
2007], занимает следующее положение:
Царство Животные - Animalia
Тип Членистоногие - Arthropoda
Подтип Ракообразные - Crustacea
Класс Высшие раки - Malacostraca
Отряд Десятиногие раки - Decapoda
Семейство Речные раки - Astacidae
Род Понтичные - Pontastacus
Вид Длиннопалый кубанский рак - Pontastacus cubanicus
2.2 Морфо-биологическая характеристика
Внешний вид длиннопалого кубанского рака представлен на рисунке 5.
Рисунок 5 - Кубанский длиннопалый рак (Pontastacus cubanicus)
Он, как и другие представители отряда десятиногих раков (Decapoda), относятся
к водным животным, дышат жабрами и имеют пять пар ходильных ног на головогруди.
Тело речных раков покрыто твердым хитиновым панцирем, пропитанным солями
кальция [Сущеня, 1972].
Среди других понтичных видов кубанский рак отличается по ширине клешни,
пропорциональному соотношению отдельных его частей. Мышцы его содержат до 15 %
белка от сырого веса или 70-90 % от сухого остатка. Средняя длина его
составляет 12,5 см, масса - 59 г, самок 12,4 и 54, самцов - 12,7 см и 63 г,
соответственно [Будников, 1952].
Раки обладают превосходным зрением и обонянием. Они различают запахи на
значительном расстоянии, как в воде, так и на суше. Завидев какой-нибудь
предмет красного цвета, брошенный человеком на дно реки, они устремляются к
нему со всех сторон, принимая его за кусок мяса. Тухлую рыбу, речные раки
обнаруживают с более далекого расстояния, чем свежую, которую обнаруживают по
цвету [Воропаев, 1990].
2.3 Размножение
Средняя физиологическая плодовитость самок - 290 икринок, число личинок -
150-200 шт, по другим данным абсолютная плодовитость самок длиннопалого рака,
составляет 276 икринок, рабочая - 193 икринки. Эти показатели могут изменяться
в зависимости от интенсивности промысла, флюктуации поколений и других
абиотических и биотических факторов [Иванов, 1955].
С наступлением весны, происходит размножение речных раков. Самец
несколько крупнее чем самка того же возраста, половые отверстия находятся в
разных местах: у самца - у основания пятой пары ног, у самки - у основания
третьей пары ног. Ниже ног, у речных раков находятся брюшные ножки
(ложноножки), у самца первые две пары их развиты наиболее сильно и направлены к
голове, у самки ложноножки или совершенно отсутствуют, или имеют вид мягких
придатков. Плес у самца относительно уже, чем у самок [Куделина, 1931].
Самцы и самки сравнительно легко различаются (рисунок 6).

Рисунок 6 - Самец и самка кубанского длиннопалого рака
Половозрелость кубанского рака в естественных условиях наступает на
третьем-четвертом году жизни. Спаривание у него зарегистрировано весной (третья
декада марта - первая декада апреля) при температуре воды 8-12 °С. Самки после
спаривания по прошествии 10-20 дней, а иногда и сразу приступают к яйцекладке.
У кубанского рака, как и у всех весеннеспаривающихся особей, доэмбриональная
диапауза длится 6 месяцев (от завершения оогенеза и сперматогенеза до
спаривания) [Черкашина, 1982]. Вылупление личинок происходит при температуре
воды 21-24 °С. Переход их на самостоятельное питание - через 7-10 дней после
вылупления [Куренков, 1951].
Процесс спаривания речных раков происходит следующим образом: самец
преследует самку, и, схватив ее ногами, прижимается к нижней стороне самки
своей нижней стороной. Потом, через половые отверстия переливает сперму во
внутренние половые органы самки. Таким образом, оплодотворение икры у речных
раков происходит внутри тела. При спаривании, самка сильно сопротивляется,
старается вырваться от самца, и, если самец оказывается слабее, уходит от него.
Оплодотворенная самка, из-за рьяного сопротивления, уходит от самца очень
потрепанной, поэтому иногда гибнет, и икра пропадает. Она скрывается в норе и
выходит из нее только днем, когда самцы отдыхают в убежищах. Самец подряд может
оплодотворить 2-3 самки, поэтому он сильно истощается и бывает настолько
голоден, что не может удержаться и съедает последнюю самку [Мацкявичене, 1986].
Через 3-4 недели после спаривания, самки приступают к икрометанию,
выпуская икру через половое отверстие, которая тут же приклеивается под плесом
к ложноножкам и остается там до вылупления личинок. Это самый тяжелый период в
жизни самки. Икра требует непрестанного промывания водой, обогащенной
кислородом, поэтому самка беспрестанно гонит воду под плесом, подгибая и
разгибая конец хвоста [Мирошниченко, 1966].
В спокойной воде, особенно когда самка сидит в норе, вода застаивается,
обедняется кислородом и обогащается продуктами обмена веществ, из-за этого икра
погибает. Самка постоянно промывает икру, очищает ее от грязи, нитчатых
водорослей и плесени. При этом, часть икринок отрывается от плеса, падает на
дно и погибает [Шпет, 1967].
Как бы ни была плодовита самка, вылупляются не более 60 личинок. Они, в
зависимости от погоды и места обитания, вылупляются в начале или середине лета
(июнь, июль). Внешне личинки мало отличаются от взрослых раков, за исключением
размеров. Развитие личинок и сравнение с размерами взрослых особей представлено
в таблице 1.
Таблица 1 - Стадия развития речных раков в естественных условиях
|
Стадия развития
|
Продолжительность развития
|
Размер личинки, мм
|
Масса, г
|
|
1 стадия
|
1-7 день
|
1,5-2 мм
|
-
|
|
2 стадия
|
5-8 день
|
8,7 мм
|
0,147 г
|
|
3 стадия
|
9-14 день
|
12 мм
|
0,346 г
|
|
Сеголетка
|
90 дней
|
30 мм
|
8-19 г
|
|
Двухлетка
|
2 года
|
60 мм
|
32 г
|
|
Половозрелые
|
3 года
|
67 мм
|
Вылупление и развитие речного рака, происходит следующим образом: рачок
вылупляется из икры, разрывая яйцевую оболочку вдоль нижней части тела зародыша
движением брюшка и конечностей, вылупившаяся личинка повисает на так называемой
«гиалиновой нити», и через 2-3 суток эта нить обрывается, но личинка
ухватывается за стебелек или оболочку икринки клешнями, которые сильно
заострены и имеют на концах загнутые крючочки. В таком положении, личинки
пребывают от 1 до 4 дней (в зависимости от температуры воды), питаясь желтком
из желточного мешка, который находится под спинным щитком головогруди. Панцирь
личинки 1 стадии мягкий, поэтому тело и масса ее увеличиваются. На этой стадии
личинки еще не похожи на взрослого рака [Нефедов, 1974].стадия развития личинки
начинается после ее первой линьки, которая наступает на пятый день после
вылупления. Желточный мешок к этому времени исчезает, головогрудь удлиняется,
панцирь становится тверже, чем у личинок первой стадии, расгрум выпрямляется,
личинки поедают яйцевую оболочку. На расширившемся тельсоне, появляются
веерообразно расположенные щетинки [Семенов, 2000].
Длина однодневных личинок достигает 1-1,5 мм. Первое время, они остаются
прикрепленными под плесом у самки. Через неделю или две, начинают плавать возле
самки, но при малейшей опасности быстро прячутся под плес. Только в возрасте
1,5-2 месяцев, личинки покидают навсегда самку и ведут самостоятельный образ
жизни [Сущеня, 1975].
Растут личинки медленно, и к осени едва достигают 2,5-3 см длины. К концу
второго года жизни, молодые раки вырастают до 6 см, прибавляя каждый год по 1
см. В 10-летнем возрасте они достигают 9-10 см. Личинки становятся очень
подвижными. Нередко, в поисках пищи уходят далеко от самки, но в случае
опасности прячутся под ее брюшком (плесом) [Сущеня, 1975]. После второй линьки,
личинки переходят в III стадию, и метаморфоз завершается. Личинка приобретает
внешний вид взрослого рака, ведет самостоятельный образ жизни и окончательно
покидает самку.
Личинки III стадии растут до полного затвердевания панциря (размер 1,2
см, масса 34,6 мг). На сроки и количество линек очень влияет температура воды.
В Ростовской области молодь растет в основном 2,5-3,5 месяца. За этот период,
происходит 6-9 линек, так как в прудах температура воды обычно выше, чем в
естественных водоемах. К концу сезона, личинки III стадии переходят в стадию
сеголеток и достигают 5-6 см длины и массы около 6 г (иногда 7,8 и 14 г) в
искусственных водоемах, и, соответственно, - 3 см и 8-10 г в реках [Черкашина,
1977, 1980].
Двухлетние раки, за теплый сезон линяют в прудах 8-9 раз, и достигают
промысловой длины 10 см, массы 32 г, а некоторые даже максимального размера
-12,3 см и 70,5 г веса [Тамкявичене, 1986].
Молодь, выращенная в реках и озерах, достигает промысловых размеров на
третье или четвертое лето [Федотов, 1993].
Выживаемость сеголеток в прудах, при хорошей кормовой базе за
вегетативный период значительно больше (85-90%), чем в естественных водоемах
(10-15%).
Высокий темп роста и выживаемость молоди речных раков, объясняются
хорошими кормовыми и температурными условиями, которые они находят в
искусственных водоемах, в то время как в реках молодь не получает даже
минимального рациона, покрывающего расходы энергии на поиски пищи и обмен
веществ в организме [Цукерзиз, 1963, 1970].
Половозрелость речных раков в естественных условиях, наступает на третьем
году жизни, при минимальном размере самок 6,7 см.
Сроки спаривания зависят от условий в водоеме и температуры воды, что
обычно наблюдается, например, в Краснодарском крае. В марте-апреле при
температуре воды 8-12сС; личинки из икры вылупляются при температуре воды
21-24°С во второй половине мая - первой половине июня.
К самостоятельному обитанию личинки в этом регионе переходят через 10-14
дней после вылупления [Черкашина, 1972, 1989].
2.4 Питание
Из растений наибольшую роль в питании европейских видов раков играют
высшие водные и околоводные растения, богатые известью: роголистник, элодея,
некоторые рдесты и харовые виды. Частота встречаемости харовых в желудках
длиннопалого рака - 14-70%. Элодея в желудках встречается 17,7% по весу
содержимого. Для речных раков такие качества харовых растений, как жесткость и,
возможно, выделяемые ими фитонциды, имеют меньшее значение, чем для мелких
видов ракообразных. Строение ротового аппарата позволяет речным ракам
использовать наряду с мягкими растениями и жесткие. Они очень охотно поедают
стебли и корневища тростника, камыша, клубнекамыша и осоки, причем речные раки
поедают все части этих растений. Их суточный рацион составляет примерно 2,5% от
живого веса рака [Фан-Хыу-Дык, 1970].
Врагами рака в природе являются хищные рыбы (щука, окунь, отчасти судак и
сом), голенастые птицы, водяные крысы и лисы [Цихон-Луконина, 1968].
Речные раки в пищевой цепи служат не только объектом питания для хищных
животных, но и сами поедают животную пищу: мелких улиток, водяных червей,
личинок разнообразных насекомых, особенно ручейников, головастиков и очень
редко небольших рыб. Впрочем, вполне здоровые рыбки успевают увернуться от
этого охотника. Пищевой спектр речного рака меняется в зависимости от его
возраста. Личинки, например длиннопалого рака, поедают до 70-80% животной пищи.
Сразу после перехода к самостоятельному образу жизни сеголетки длиною 1,2-2 мм
питаются дафниями (59%), хирономидами (25%). По мере роста, доля дафний в
рационе уменьшается до 5%, а в возрасте двух лет, дафний совсем нет.
Молодь всех разномерных групп потребляет хирономид (24-25%). Сеголетки,
при достижении длины 2 см, начинают питаться насекомыми (18- 45%) и их
личинками. В частности, личинками ручейников, поденок, веснянок и др. По мере
роста речных раков, возрастает потребление бокоплавов, с 5% у сеголетков, до
63% у молоди длиною 8-10 см. Моллюски появляются в пище сеголетков, когда их
длина достигает 3 см, а рыбы - при длине сеголетков 4 см.
Пищевой спектр молоди кубанского рака в водоемах меняется так же по мере
роста. Сразу после перехода к самостоятельному образу жизни молодь длиною
1,2-2,0 см питается дафниями (59 %), хирономидами (25 % от веса пищевого
комка). По мере роста сеголетков доля дафний в рационе уменьшается до 5 %, а
двухлетки ими не питаются. Молодь всех размерных групп потребляет хирономид
(24-25 %) [Черкашина, 1982].
Сеголетки при достижении 2 см начинают питаться насекомыми (18-49 %) и их
личинками, в частности, личинками ручейников, поденок, веснянок и др. По мере
роста возрастает потребление бокоплавов с 5 % у сеголетков до 63 у молоди
длиною 8-10 см. Половозрелые раки питаются рыбными остатками (62 %), которые
начинают появляться в желудках молоди длиною 3 см и моллюсками (16 %)
[Черкашина, 1977].
2.5 Экология обитания
Раки требовательны к содержанию кислорода в воде и при падении его до
2,9-1,2 мл/л обычно выползают на берег, и возвращаются в водоем при
восстановлении кислородного режима. При 0,37 мл/л они погибают. Оптимальное
содержание кислорода в воде для раков - 6-7 мл/л.
В естественных условиях обитают в тихой проточной воде, заселяя реки,
протоки, каналы и крупные ручьи, а также озера и проточные пруды с тенистыми
берегами, прячутся в норах, под корягами или корнями больших деревьев, растущих
на берегах водоемов. Дышат они, сложно устроенными жабрами, но в сильно влажной
атмосфере могут некоторое время дышать также воздухом. Речные раки очень
требовательны к качеству воды, и при ее загрязнении или цветении покидают такой
водоем и переселяются в тот, где находят благоприятные условия [Черкашина,
1983, 1985].
Ухудшение качества воды в водоеме, в зимнее время ведет к полной гибели
популяции речных раков. В воде, бедной минеральными веществами, панцирь у раков
становится мягким. На темпы роста молоди раков отрицательно влияет накопление
продуктов обмена. При концентрации аммонийного азота 3,5 мг/л рост раков
останавливается, а при 10,0 мг/л наступает их гибель (Шяштокас, Цукерзис,
1972). Кубанский рак является эвритермным видом и переносит колебания
температуры от 4 до 32 °С. Оптимальная температура для роста - 22-24 °С. Длиннопалые
раки, в отличие от широкопалых, активны весь год, поэтому возможен их зимний
промысел. Они чаще живут на мягких, глинистых почвах и прячутся в норы, вырытые
в обрывах рек, активны ночью. Днем они затаиваются в различных убежищах. В
теплое время года, убежища находятся близко от поверхности воды, а зимой
устраиваются на такой глубине, чтобы нора не закрылась льдом, так как ледяной
плен приводит к их гибели. С наступлением сумерек рак выходит из убежища на
охоту в поисках пищи. Обычно он передвигается по дну водоема и очень редко
плавает, Иногда, в поисках пищи выходит на берег, почуяв запах падали, которую
он очень любит. В очень редких случаях взрослого рака можно встретить на суше
днем.
2.6 Биологические особенности развития
Чрезвычайно ответственным периодом в жизни раков является линька.
Линька речных раков происходит один-два раза в год у взрослых особей, а у
молоди каждый раз при росте и развитии. Сроки наступления линьки различны, и
зависят от местности, в которой обитают раки. Рост молоди происходит сразу
после линьки и до затвердения нового панциря. Взрослые самцы кубанского рака
линяют два раза; самки, участвующие в размножении - один раз. Первая линька
самцов происходит при температуре воды 19-22 °С (конец мая - начало июня),
вторая линька самцов и первая самок - в августе. Молодь раков линяет в
естественных условиях 5-6 раз. Весь сезонный ход размножения и линьки раков
контролируется температурой воды.
Убыль популяции раков при стабильной экологической ситуации в водоемах
тесно связана с размножением и ростом особей, которые ее регулируют. Каждый вид
раков, как и любого животного, характеризуется смертностью, которая колеблется
в пределах определенной амплитуды. От старости погибает очень небольшой процент
раков, так как их вылавливают раньше.
Для каждого вида раков специфична не только общая величина смертности, но
и ее распределение по отдельным возрастным группам и этапам развития.
Наибольшая гибель раков приходится на периоды икрометания, раннего развития
эмбриогенеза и перехода молоди от самок на самостоятельное питание.
Определяющими причинами гибели икры при икрометании и на стадии раннего
онтогенеза являются абиотические факторы, особенно температура воды. С
переходом молоди раков на самостоятельное питание действие этих факторов обычно
снижается и на первое место выступает обеспеченность ее пищей, а также гибель
от хищников. К концу сезона выживаемость сеголеток раков, перешедших на
самостоятельное питание, составляет около 4-5 %.
Гибель половозрелой части популяции кубанского рака при стабильной
экологической ситуации составляет 2,5 %, что совпадает с литературными данными
[Бродский, 1981].
Таким образом, при стабильной экологической ситуации в водоемах основными
причинами гибели раков на ранних этапах онтогенеза являются в основном абиотические
факторы, при переходе молоди к самостоятельному образу жизни. Гибель
промысловых особей характеризуется определенной, более или менее стабильной
величиной.
Интенсивное загрязнением водоемов токсическими веществами приводит к
нарушению экологии раков и значительной гибели - до 32,7 %, а, следовательно, -
к резкому сокращению их численности, биомассы и нарушению структуры популяции.
3. Материалы и методы исследования
Учет численности популяций кубанского рака в водоемах системы
Ахтарско-Гривенских лиманов осуществлялся методом прямого учета с
использованием пассивных орудий лова - стандартных раколовок, а также с
применением мальковой волокуши, снабженной фальшподборой (рисунок 7).
Коэффициенты уловистости раколовок (определенные АзНИИРХ) составляют - 0,71,
мальковой волокуши - 0,13. Учитывая, что одна раколовка облавливает раков в
среднем с площади 100 м2, расчет удельной численности раков производится по
формуле [Методы…, 2005]: для раколовок: А = 1,4a*100 (1)
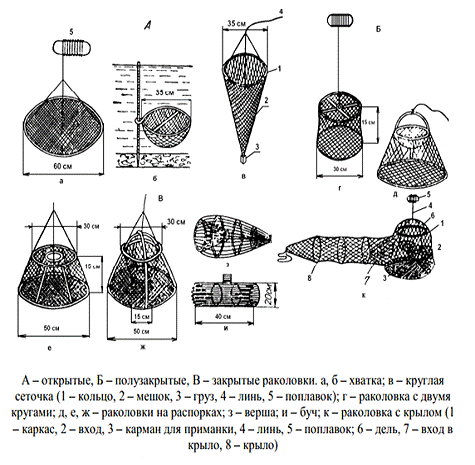
Рисунок 7 - Орудия ловли раков
Для определения размерно-массовых характеристик и оценки численности
кубанского рака в лиманах, учетными орудиями (раколовки, и мальковая волокуша)
выловлено 179 экземпляров взрослого рака и 96 экземпляров молоди рака.
Учет сеголеток (4-6 см) проводился раколовками и мальковым тралом с ячеей
6,5мм. При проведении сравнительного анализа уловистости пассивных и активных
орудий лова отмечена более высокая уловистость мальковых тралов на грунтах с
редкой водной растительностью, и, соответственно, раколовок в густой водной
растительности.
В промысловых орудиях лова с ячеей 16мм доля молоди раков (4-6см) не
превышала 3,7%, раколовки с ячеей 18мм практически полностью исключали прилов
молоди. В многозаходных раколовках «корейского производства» доля прилова
молоди была самой высокой - 10,4-19%, вместе с молодью рака в эти раколовки
попало большое количество молоди судака. В Гривенском лимане молодь судака была
в 7 раколовках из 10 с максимальной численностью в одной раколовке 18 экз.
Одной из мер снижения прилова молоди раков и рыбы может быть введение
ограничения минимальной ячеи в раколовках - 18мм.
Сбор материалов по питанию проводился согласно инструкции Брейтбурга А.М.
(1961).
4. Биологические особенности кубанского рака
В Ахтарско-Гривенских лиманах ракопродуктивные водоемы составляют 13,9
тыс.га (по оценке АзНИИРХ, 2007).
В 2013 г., в связи с благоприятным гидролого-гидрохимическим режимом и
отсутствием массовых заморов, несмотря на положительную температурную аномалию
в летний период, был высокий урожай раков. Условия обитания и состояние
популяции кубанского рака в лиманах после зимовки весной 2014 г. были
относительно благополучными, массовых заболеваний и гибели раков не
наблюдалось. Встречаемость яловых самок во всех водоемах не превышала 5 %, а
ржавопятнистого заболевания - 4,6%, (лиманы Дончиков и Пригибский).
Условия обитания кубанского рака в лиманах 2013-2014 гг. были
относительно благополучными. Максимальные концентрации молоди рака длиной
43-52мм отмечены в прибрежной части лиманов. Уловы сеголеток в
Ахтарско-Гривенской группе лиманов (лиман Гривенский) находились на уровне -
0,04 - 0,17 экз./м², доля прилова годовиков находилась в пределах 16 -
32%. Воспроизводственный потенциал раков в лиманах в последние годы находится
на достаточно высоком уровне и должен обеспечить хорошее пополнение промысловых
запасов в ближайшие годы.
4.1 Размерно-массовая характеристика
Размерно-массовый состав части популяции длиннопалого кубанского рака,
был представлен особями размерной группы от 6,5 до 13,0 см. Доминирующая группа
(54,2%) была представлена особями 8,6-11,0 см. Массовая часть была представлена
особями от 8,2 до 88,5 г (рисунок 8, таблица 2).
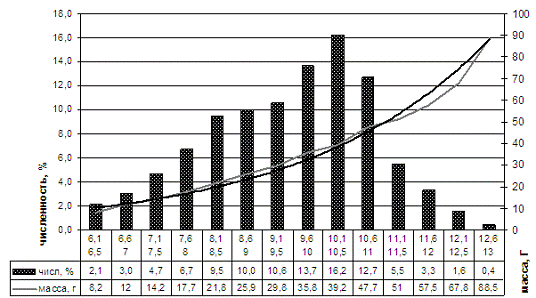
Рисунок 8 - Размерно-массовая характеристика части популяции длиннопалого
кубанского рака
Таблица 2 - Размерный состав и масса раков по группам
|
Группа
|
Масса, г
|
|
Средняя
|
Минимальная
|
Максимальная
|
|
Группа пополнения (6,5 -
8,5 см)
|
16,9
|
12,1
|
21
|
|
1-я промысловая группа (8,6
- 12,0 см)
|
41,0
|
25,8
|
57,5
|
|
2-я промысловая группа
(12,1 - 13,0 см)
|
72,9
|
70,5
|
88,5
|
4.2 Возрастная и половая структуры
В 2014 г, в среднем по лиману, молодь (6,5-8,5см) составляет 28 %, раки 1
промысловой группы (8,6-12,0 см) - 70%, раки 2 промысловой группы (12,0-13,0
см) - 2% (рисунок 9)
Соотношение полов в общей структуре популяции характеризуется небольшим
преобладанием самцов (1,1-1,6:1), что в пределах нормы и обеспечивает
нормальные воспроизводственные показатели.

Рисунок 9 - Доли разновозрастных групп в общей части популяции
Средняя рабочая плодовитость самок промысловых размеров (9,1-12,0 см)
была на уровне показателей предыдущих лет и составляла 148-265 икринок, у более
крупных особей - 264-292 икринки, а у самок из группы пополнения - 93-212
икринок (таблица 3).
Таблица 3 - Показатели плодовитости и полового соотношения у части
популяции длиннопалого кубанского рака
|
Молодь, (6,5 - 8,5 см)
|
1 пром. группа, (8,6 - 12,0
см)
|
2 промысловая группа, (12,1
- 13,0 см)
|
Средняя
|
|
Плод. шт. икринок
|
93-212
|
214-240
|
264-292
|
198-242
|
|
Соотношение самцов и самок,
%
|
|
Самцы
|
62,3
|
69,6
|
53,4
|
61,8
|
|
Самки
|
37,7
|
30,4
|
46,6
|
38,2
|
|
Соот. самцов и самок
|
1,7:1
|
2,3:1
|
1,2:1
|
1,6:1
|
|
|
|
|
|
|
Икринки имели ровную и упругую структуру, чёткий контур
4.4 Исследования по питанию
Таблица 4 - Частота встречаемости различных пищевых компонентов в
желудках длиннопалого кубанского рака.
|
Количество вскрытых особей,
шт.
|
168
|
|
Количество желудков с
пищей, шт.
|
144
|
|
Зеленые нитчатки
|
41
|
|
Роголистник
|
82
|
|
Уруть
|
69
|
|
Тростник
|
112
|
|
Камыш
|
2
|
|
Кувшинки
|
4
|
|
Рдест
|
11
|
|
Неопределенные остатки
растений
|
72
|
|
Кладоцеры
|
7
|
|
Остраподы
|
6
|
|
Мизиды
|
9
|
|
Ручейник
|
8
|
|
Хирономиды
|
12
|
|
Прочие личинки насекомых
|
14
|
|
Моллюски
|
8
|
|
Остатки панцирей речных
рачков
|
1
|
|
Рыба
|
6
|
|
Мышечная ткань (не опр.)
|
2
Таблица 5 - Процентное соотношение пищевых компонентов
|
Общий объем пищевого комка,
%
|
100
|
|
Зеленые нитчатки
|
0,5
|
|
Роголистник
|
18,8
|
|
Уруть
|
14,4
|
|
Тростник
|
20,8
|
|
Камыш
|
8,6
|
|
Кувшинки
|
0,2
|
|
Рдест
|
1,8
|
|
Осока
|
11,6
|
|
Фоктуалис
|
0,7
|
|
Неопределенные остатки
растений
|
12,2
|
|
Кладоцеры
|
0,1
|
|
Остраподы
|
0,1
|
|
Мизиды
|
0,6
|
|
Ручейник
|
1,2
|
|
Хирономиды
|
0,4
|
|
Прочие лич. нас.
|
0,5
|
|
Моллюски
|
4,3
|
|
Остатки панцерей речных
рачков
|
0,1
|
|
Рыба
|
1,8
|
|
Мышечная ткань (не опр.)
|
1,4
|
|
Процент растительности в
желудках
|
89,6
|
|
Процент животной пищи
|
10,4
|
Зеленые нитчатки встречались в значительном количественном разнообразии.
Ни в одном случае водоросли не составляли значительной доли в общем содержании
желудков. Весьма значительный процент различных групп зеленых водорослей из
обрастаний.
Индексы наполнения желудков составляли от 3,8 до 1,3% к весу тела.
Отмечено уменьшение индекса с уменьшениями размеров тела.
Процент соотношения животной и растительной пищи был различен. У молоди,
как и утверждается в литературных данных, растительная пища, особенно
растения-обрастатели, составляет основу рациона, тогда как у взрослых особей
наблюдается изменения рациона в сторону животной пищи.
В целом, доли растительной и животной пищи составили 89,6 и 10,4
соответственно (рисунок 10).
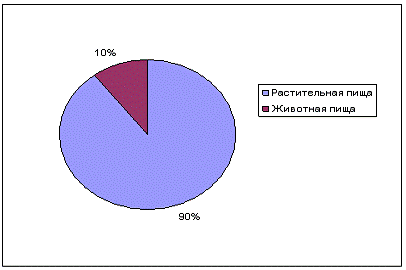
Рисунок 10 - Процентное соотношение животной и растительной пищи в
рационе части популяции длиннопалого кубанского рака
4.5 Связь состояния популяции и места исследования
В целом, состояние популяции длиннопалого кубанского рака в
Ахтарско-Гривенских лиманах, не вызывает опасения. Однако, восстановление
популяции длиннопалого кубанского рака произошло сравнительно недавно и любое
нарушение равновесия экологических факторов среды обитания способно значительно
подорвать численность данной популяции. Любое загрязнение, связанное с
сельскохозяйственной деятельностью, а так же изменение водного режима лиманов
способно значительно сократить численность кубанского длиннопалого рака. Из-за
изменения глубины лиманов (проведение обводных каналов) уже наблюдается сильное
зарастание и обмеление западных участков лиманов. И, как следствие, уловы, как
молоди, так и взрослых особей там составляют небольшое число, по сравнению с
другими участками.
Заключение
Целью работы было выявить и описать особенности биологии кубанского
длиннопалого рака в Ахтарско-Гривенских лиманах, провести исследования по
размерно-массовым характеристикам, возрастной и половой структурам, дать оценку
плодовитости, а так же описать качество и состояние половых продуктов у самок,
провести анализ питания длиннопалого кубанского рака.
В процессе работы были получены различные данные исследования,
перечисленные ниже:
размерно-массовый состав части популяции длиннопалого кубанского рака,
был представлен особями размерной группы от 6,5 до 13,0 см. Доминирующая группа
(54,2%) была представлена особями 8,6-11,0 см. Массовая часть была представлена
особями от 8,2 до 88,5 г. Полученные данные практически совпадают с данными
предыдущих исследований, что говорит о стабильном состоянии популяции
длиннопалого кубанского рака;
- было выделено три возрастные группы, выборка проводилась исходя из
размера особи;
в среднем по лиману в 2014 г., молодь (6,5-8,5см) составляла 28 %, раки 1
промысловой группы (8,6-12,0 см) - 70%, раки 2 промысловой группы (12,0-13,0
см) - 2%;
- соотношение полов в общей структуре популяции характеризовалось
небольшим преобладанием самцов (1,1-1,6:1);
средняя рабочая плодовитость самок промысловых размеров (9,1-12,0 см)
была на уровне показателей предыдущих лет и составляла 148-265 икринок, у более
крупных особей - 264-292 икринки, а у самок из группы пополнения - 93-212
икринок;
икра имела четкую структуру, без посторонних включений, была плотно
прикреплена к плеоподам, имела равномерную окраску, без каких-либо отклонений;
спектр питания длиннопалого кубанского рака был разнообразен, всего в
рационе отмечено 18 различных пищевых остатков, отмечено изменение в рационе,
происходящее по мере роста рака;
в целом, доли растительной и животной пищи в рационе длиннопалого
кубанского рака составили 89,6% и 10,4% соответственно;
индексы наполнения желудков составляли от 3,8 до 1,3% к весу тела;
экологическое состояние Ахтарско-Гривенских лиманов в настоящее
благоприятно для развития длиннопалого кубанского рака, однако зарег. стока и
проведение обводных каналов может существенно сократить численность данного
вида из-за обмеления водоемов и их заиливания;
повышение численности популяции длиннопалого кубанского рака возможно при
соблюдении следующих рекомендаций:
. Осуществление мелиоративных процессов в местах обитания длиннопалого
кубанского рака, очистка водоемов от ила;
. Повышение эффективности борьбы с браконьерским ловом молоди раков,
контроль за поставленными раколовушками, извлечение браконьерских орудий лова,
особенно в прибрежной зоне, где наблюдается повышенное скопление молоди рака,
соблюдение сезонных циклов при промысловой добыче длиннопалого кубанского рака;
. Проведение мероприятий, направленных на искусственное пополнение
популяции длиннопалого кубанского рака (искусственное воспроизводство раков с
последующим выпуском в естественные водоемы);
. Контроль за качеством воды в системе Ахтарско-Гривенских лиманов,
особенно важно соблюдение ПДК с хлорорганическими, азотными и фосфорными
соединениями.
. Недопуск попадания в водоемы стоков с сельскохозяйственных угодий,
рисовых чеков и продуктов нефтепереработки.
Библиографический список
1.
Александрова Е.М. Промышленное культивирование речных раков//Рыбоводство и
рыболовство. - 1994. - Вып.№4. - с. 27-28.
. Бирштейн
Я.А. Высшие раки. Жизнь пресных вод. - 1940. - 110с.
. Бирштейн
Я.А. Пресноводные десятиногие СССР и их географическое
распространение//Зоологический журнал. - 1934. - Вып.№1.- с. 121-130.
. Брейтбург
А.М. Физиология питания. - М.: 1961.- 630 с.
. Бродский
С.Я. Речные раки Советского союза.//Распространение длиннопалого рака//Вестник
зоологии.-1974.- с. 51-58.
. Бродский
С.Я. Речные раки и использование их запасов//Рыбное хозяйство.-1967.- с. 26-27.
. Будников
К.Н. Рак, его разведение и промысел. - М., 2005.
. Будников
К.Н., Третьяков Ф.Ф. Речные раки и их промысел.-М., 1952.- 95 с.
. Вернадский
В.И. Биосфера. - М.: Мысль, 1967. - 376 с.
. Воловик
СП., Макаров Э.В., Семенов А.Д. Азовское море: возможен ли выход из
экологического кризиса // Осн. пробл. рыбн. хоз-ва и охраны рыбохоз. водоемов
Азовского басе: Сб. научн. тр. АзНИИРХ. - Ростов-на-Дону, 1996. - С. 115-125.
. Воропаев
Н.В., Полосьянц Т.Ю., Новосельцев Г.Е., Понкратов О.В. Междунар. симпоз. по
разведению и выращ. раков // Рыбное хоз-во, сер. «Рыбохоз использ. внутр. водоемов».
- ВНИиПК экономики, информации и автоматизир. систем управления рыбн. хоз-ва,
1990. - С. 3-9.
. Гаевская
Н.С. Роль высших водных растений в питании животных пресных водоемов. - Мм
1966. - 328 с.
. Гексли Т.
Рак. - М., 1800. - 272 с.
. Дилов X.
Микроводоросли. - София: Изд-во на академия на научи, 1985. - 194 с.
. Зинова А.Д.
Определитель зеленых, бурых и красных водорослей Южных морей СССР. - М.-Л.:
Наука, 1967. -281 с.
. Золотницкий
Н.Ф. О расселении пресноводных раков на земном шаре // Дневник отд. русск.
об-ва акклим. животных и растений. - 1900.-Вып. 1.-С. 72-83.
. Иванов А.В.
Промысловые водные беспозвоночные. - М.,1955.
. Инструкция
по сбору и обработке материала для исследований питания раков в естественных
условиях. - М.: Изд-во ВНИРО, 1971.-С. 1-76.
. Куделина
Е.Н. Биология и промысел азово-донского и азово-кубанского рака. - Рукопись
ВНИРО, 1931.
. Куренков
И.И. Питание речного рака // Тр. Моск. техн. ин-та рыбной промышленности и
хоз-ва им. Микояна. - 1951. -Вып. 4. - С. 82-90.
. Лотышев И.
П. География Кубани. Энциклопедический словарь. Майкоп, 2006. - 138 с.
. Макаров
Э.В., Семенов А.Д. Экологические аспекты проблемы развития рыбного хозяйства в
Азовском бассейне // Осн. пробл. рыбн. хоз-ва и охраны рыбохоз. водоемов
Азовского басе: Сб. научн. тр. АзНИИРХа. - Ростов-на-Дону, 1996. - С. 6-20
. Мацкявичене
Г.И. Влияние питания на уровень обмена веществ у молоди длиннопалого рака //
Тез. докл. IV Всес. конф. по промысловым беспозвоночным. Часть II. -
Севастополь, 1986. -С. 113-114.
. Мирошниченко
А.З. К вопросу о питании и росте молоди длиннопалого и широкопалого раков //
Гидробиол. ж. - 1966. -№3.-С. 56-59.
. Москул
Г.А., Москул Н.Г. Современное состояние популяции длиннопалого кубанского рака
в некоторых водоемах Краснодарского края//ВНИРО.- 2005. с. 182-183.
.
Мордухай-Болтовской Ф.Д. К вопросу о происхождении каспийской фауны в
Азово-Черноморском бассейне // Зоолог, ж. - 1946. - 25. - Вып. 5. - С. 411-426.
. Нефедов
В.Н. Размножение длиннопалого рака Astacus leptodactylus Esch// Тр. Волгоградского
отд. ГосНИОРХ. - 1974. - Т. 8. - С. 68-82.
. Никоноров
А.М. и др. Биомониторинг тяжелых металлов в пресноводных экосистемах. - П.:
Гидрометеоиздат, 1985. -210 с.
. Павлютин
А.П. К методике определения усвояемости пищи у водных животных // Зоолог, ж. -
1970. - Т. 49. - №2. - С. 48-52.
. Румянцев
В.Д. Изучение биологии, состояния запасов и мест концентрации речного рака. -
Рукопись. КаспНИРХ / Архив ВНИРО.- 1968. - 30 с.
. Семенов
А.Д., Короткова Л.И., Сапожникова Е.В., Короленко Е.О. Современное состояние
пестицидного загрязнения водных объектов Азовского бассейна // Осн. пробл.
рыбн. хоз-ва и охраны рыбохоз. водоемов Азово-Черноморского басе: Сб. научн.
тр. АзНИИРХа. - Ростов-на-Дону, 2000. - С. 301-306.
. Скориков
А.С. Новый вид речного рака с Кавказа // Изв. Кавказок, музея. - 1911. - Т. 5.
- Вып. 4. - С. 328-340.
.
Старобогатов Я.М. Определитель пресноводных беспозвоночных России и
сопредельных территорий. - Т. 2. Ракообразные. - СПб., 1995. - С. 177-183.
. Сущеня Л.М.
Интенсивность дыхания ракообразных. - Киев: Наукова думка, 1972. - С. 1-194.
. Сущеня Л.М.
Количественные закономерности питания ракообразных. - Минск: Наука и техника,
1975. - 205 с.
. Тамкявичене
Е.А. Рост и развитие молоди аборигенных и акклиматизированного видов речных
раков // Тез. докл. IV Всес. конф. по промысл, беспозвоночным. - Севастополь,
1986. -С. 120-121.
. Тевяшова
О.Е. Влияние антропогенного воздействия на зоопланктон водоемов Азово-Донского
бассейна // Сб. научн. тр. АзНИИРХ. - Ростов-на-Дону, 1998. - С. 134-143.
. Тевяшова
О.Е. Влияние одноклеточных водорослей на рост и выживаемость личинок раков //
Рекомендации по подращиванию личинок раков: Отчет о НИР // АзНИИРХ; Рук.
Черкашина Н.Я. - Ростов-на-Дону, 1988. - С. 15-20.
.
Фан-Хыу-Дык. Материалы к изучению питания раков из рода Astacus // Ученые
записки Азерб. ун-та / Сер. биол.-Баку, 1970.-Вып. 3.-С. 183-186.
. Федотов
В.П. Разведение раков. - СПб: Биосвязь, 1993. - 101 с.
.
Цихон-Луконина Е.А. Об усвояемости пищи донными ракообразными Азовского моря и
методика ее определения // Океанология. - 1968. - Т. VIII. - Вып. 3. - 487 с.
. Цукерзис
Я.М. Биология длиннопалого рака. - Вильнюс: Милятис, 1970.-204 с.
. Цукерзис
Я.М. Зависимость между газообменом и весом тела у широкопалого и длиннопалого
раков // Тр. АН ЛитССР. -1967. -Сер. В.-Т. 3(44).-С. 87-90.
. Черкашина
Н.Я., Тевяшова О.Е., Карпенко В.Н., Новикова Е.С. Состояние популяций раков
рода Astacus в водоемах Азово-Донского бассейна в условиях антропогенного
воздействия // Сб. научн. тр. АзНИИРХ. - Ростов-на-Дону, 1996. -С. 206-211.
. Черкашина
Н.Я. Биотехника выращивания длиннопалого рака в прудах // Информ. листок
Ростовского ЦНТИ № 419-81. - Ростов-на-Дону: РЦНТИ, 1981. -4 с.
. Черкашина
Н.Я. Влияние качества самок длиннопалого кубанского рака на потомство //
Экология. - 1979. - №4. - С. 76-81.
. Черкашина
Н.Я. Временная инструкция по выращиванию длиннопалого рака в прудах. - М.:
ВНИРО, 1982. - С. 1-24.
. Черкашина
Н.Я. Интенсивность дыхания раков рода Astacus из Азовского бассейна // Зоолог,
ж. - 1982. - Т. XI. - №1 -С. 451-454.
. Черкашина
Н.Я. К биологии молоди длиннопалого рака Astacus leptodactylus cubanicus //
Использование промысловых беспозвоночных на пищевые, кормовые и технические
цели: Тез. докл. Всес. конф. - М., 1977. - С. 109-110.
. Черкашина
Н.Я. Определение возраста раков графическим методом «вероятностной бумаги» //
Экспресс-информация «Рыбохоз. использов. внутренних водоемов». - 1973. -Вып.
12.-С. 1-4.
. Черкашина
Н.Я. Основные направления увеличения продукции длиннопалого кубанского рака //
Тез. докл. Y Всесоюз. конф. по промысловым беспозвоночным. - М., 1990. - С.
72-73.
. Черкашина
Н.Я. Питание раков семейства Astacidae в Азовском бассейне // Гидробиол. ж. -
1985. - Т. XXX. - №6. - С. 34-39.
. Черкашина
Н.Я. Раки из рода Astacus в Азовском бассейне // Тр. ВНИРО. - 1980. - С. 78-91.
. Черкашина
Н.Я. Раки рода Astacus и перспективы их культивирования. - Ростов-на-Дону:
Изд-во «ТОР», 1994. - 143 с.
. Черкашина
Н.Я. Рост и питание молоди длиннопалого рака Astacus leptodactylus (Decapoda,
Astacidae) // Зоолог, ж. - 1977. -Т. 56. - Вып. 5. - С. 704-708.
. Черкашина
Н.Я., Буртыка Б.Б. Речные раки // Кн. «Ресурсы живой фауны». Часть 1. -
Ростов-на-Дону: Изд-во РГУ, 1980. -С. 62-67. '
. Черкашина
Н.Я., Карнаушенко И.В. Доэмбриональная диапауза у длиннопалого кубанского рака (Astacus
leptodactylus cu-banicus Bir. et Win.) // Ж. общ. биологии. - 1982. - Т. XIII. -
№5. - С. 687-689.
. Черкашина
Н.Я., Ковалевский В.Н., Шукашева СИ. Запасы и промысел длиннопалого рака в
водоемах Азовского бассейна // Тез. докл. обл. научн. конф. по итогам работы
АзНИИРХ в XI пятилетке. - Ростов-на-Дону: АзНИИРХ, 1986. -С. 196-199.
. Черкашина
Н.Я., Ковалевский В.Н., Шукашева СИ. Речные раки Азовского бассейна // Тез.
докл. обл. научн. конф. по итогам работы АзНИИРХ за 25 лет. - Ростов-на-Дону,
1983. -С. 114-116.
. Черкашина
Н.Я., Стась Л.Р., Карпенко В.Н. К антропогенному воздействию на длиннопалого
кубанского рака (Astscus leptodactylus cubanicus) // Тез. докл. V Всесоюз.
конф. по промысловым беспозвоночным. - М., 1990. - С. 70-72.
. Черкашина
Н.Я., Тевяшова О.Е., Карпенко В.М., Новикова Е.С. Состояние популяций раков
рода Astacus в водоемах Азовского бассейна (Ростовская область) в условиях
антропогенного воздействия: Отчет о НИР / АзНИИРХ. - Ростов-на-Дону, 1991.-70
с.
. Черкашина Н.Я.,
Тевяшова О.Е., Карпенко В.Н., Беляева В.А., Глушко Е.Ю., Новикова Е.С. Раки
рода Astacus в водоемах Азовского бассейна и рекомендации по сохранению и
увеличению их численности // Сб. «Проблемы охраны, рационального использования
и воспроизводства речных раков». - М.: Мединар, 1997. - С. 96-104.
. Черкашина
Н.Я., Шуховцов В.В. Сезонная динамика химического состава и пищевой ценности
длиннопалого рака // Использование промысловых беспозвоночных на пищевые,
кормовые и технические цели: Тез. докл. Всес. конф. - М., 1977. -С. 104-106.
. Шпет Г.И.,
Бродский С.Я. Речные раки и их продуктивность // Гидробиол. ж. - 1967. - №1. -
С. 73-77.
Похожие работы на - Биологические особенности кубанского рака
|