Изучение влияния ментального и вербального биоэнергоинформационого воздействия на процессы онтогенеза у растений
Введение
Древние философы говорили: "Сначала было
слово". Другие заявляли, что первичным было дело. Но эти споры о
первичности не имеют никакого смысла, так как мыслеформа и мыслеобраз являются
категориями разных метрических пространств. Слово преобразует мыслеобраз в
мыслеформу и запускает мыслеформу в материализацию [1].
Язык общения между людьми - не просто
лингвистическое образование, сложившееся у данного этноса. Слово - ключ к
Информационным Полям. Поэтому энергетическому потенциалу вербального воздействия
отводит такое грандиозное значение - это всем известные молитвы, мантры,
заговоры, нейролингвистическое программирование сознания (НЛПи)... Простым
набором слов в фразе можно организовать абсолютно несанкционированный выход
через информационные поля в чужеродные эгрегориальные образования для
неконтролируемого запуска в материализацию деструктивных для нашей цивилизации
программ. Ведь мало кто из землян сможет понять, что случайно брошенная фраза
может привести к глобальной катастрофе через тысячу лет! [2].
Академик Казначеев В.П. пишет в своих трудах,
что человек- это уникальная биоэнергострукткра, отсюда вытекает невозможная
замена человека искусственным интеллектом. Возникает необходимость
органического слияния биосферы и человека на основе антиэнтропийных процессов
[4].
По словам В. М. Инюшина, в биологии уже сейчас
ощущается познавательный кризис, который обостряется в связи с проблемой
будущего человека как биологического вида на фоне экологических бедствий.
Всевозможные антропогенные факторы на фоне естественных всплесков солнечной
активности ведут к деградации психоэнергоструктуры человечества. Необходим
переворот в теоретическом и практическом мышлении. Особенно это актуально как
для биологии, так и для создания новых методов интенсификации сельскохозяйственного
производства и глобальных систем оздоровления человека, повышающих его
биоэнергетический потенциал [3].
Исходя из вышесказанного и основываясь на
работах В. М. Инюшина, В. П. Казначеева, П. П. Горяева было решено провести
эксперименты по интересующей нас проблематике.
Актуальность нашей работы определяется тем, что
вербальное влияние человека на полевоволновые структуры растений мало изучено,
хотя имеет большие перспективы в биологии и экологии для создания новых методов
интенсификации сельскохозяйственного производства и глобальных систем
оздоровления человека, повышающих его биологический потенциал.
Целью нашей научной работы является изучение
влияния вербального и ментального биоэнергоинформационного воздействия на рост
и развитие отдельных количественных признаков, а также биологической и
хозяйственной продуктивности перца горького, перца сладкого, баклажан,
картофеля.
Выбор объектов исследования не случаен, так как
данные культуры представляют большой интерес для сельскохозяйственного производства.
Для достижения цели работы были поставлены
следующие задачи:
. Изучить литературные источники по интересующей
нас проблематике, в частности, работы П. П. Горяева, В. М. Инюшина, Тихоплавов,
В. П. Казначеева и др.
. Составить текст-настрой для проведения
вербального и ментального биоэнергоинформационного воздействия.
. Разбить делянки и высадить перец горький,
перец сладкий баклажаны, картофель.
. Провести замеры отдельных количественных
признаков: длины стебля, длины и ширины листовой пластинки, величины
продольного и поперечного диаметра клубня, посчитать количество цветков, плодов
и количество стеблей в момент вегетации.
. Провести биометрическую обработку вариационных
рядов по вышеперечисленным признакам.
. Провести анализ результатов вербального и
ментального биоэнергоинформационного воздействия на рост и развитие отдельных
количественных признаков, а также на биологическую и хозяйственную
продуктивность перца горького, перца сладкого баклажан, картофеля.
вербальный ментальный биометрический
биоэнергоинформационный
1. Литературный обзор
Изучение влияния ментального и вербального
биоэнергоинформационого воздействия на процессы онтогенеза у растений
В настоящее время существуют несколько
направлений научных исследований, подтверждающих существование так называемых
"тонких структур" или "Тонкого Мира". Одной из первых
следует назвать концептуальный подход и обоснование самой возможности
существования "Тонкого Мира".
В своей работе "Неизбежность, реальность и
постижимость "Тонкого Мира". Владимира Никитовича Волченко, д.т.н.,
профессора МГТУ им. Н.Э.Баумана, президента Международного общественно научного
комитета "Экология человека и энергоинформатика" (цит. по Гаряеву),
приводит научно-философское обоснование существования Тонкого Мира на основе
духовной традиции и разнообразных феноменов информационно-энергетического
обмена на уровнях человека, Земли и Космоса. Сознание человека рассматривается
с более широких нежели материалистические позиций. Сознание трактуется как
энерго-информационная структура и в своих высших аспектах естественная часть
Космоса. Волченко вводит численные характеристики возможных границ миров в
системе координат "информативность - энергетичность". На основе
замеченной тенденции к повышению информационной эффективности систем
доказывается возможность постижимости "Тонкого Мира". Обсуждаются
свойства так называемых информационных безэнтропийных полей и пятого
фундаментального взаимодействия. Рассматриваются возможные модели
информационных полей (торсионная, бюонная, психоновая, семантическая,
реликтовых нейтрино, аксионовая, продольных электромагнитных полей и др.).
Автор утверждает, что метафизика должна стать наукой. Осуждается
технократический путь развития человеческой цивилизации и предлагается
синтетический подход. Подчёркивается значение морально-нравственной стороны в
эволюции [5].
Второе направление наиболее обширно. Его можно
обозначить как обоснование и доказательство объективного существования так
называемой "души" или энерго-информационной (полевой,
электромагнитной) сущности человека (сознания).
Одним из первых учёных, высказавших гипотезу о
полевой природе человеческого сознания был А. Г. Гурвич. В своей работе он
отмечает, что вся информация о строении потенциального организма содержится в
совокупном эмбриональном фотонном поле, излучаемом каждой хромосомой эмбриона.
А. Г. Гурвич утверждает, что такое интегральное поле создаёт волновой
биополевой каркас, план, по которому идёт строительство или самоорганизация
клеток в организм. В сущности предложена научная концепция биополя живых
существ. Также А. Г. Гурвич рассказывает о проявлениях митогенетического
излучения в опытах с умерщвлением креветок, когда чувствительная фотобумага
засвечивалась в результате излучения, генерируемого умирающими креветками при
их ошпаривании кипятком [6].
Работы Гурвича А. Г., Любищева А.А. и
Беклемишева В.Н. подтвердили идеи принципиальной возможности дистантной
трансляции и акцепции эпигенетических управляющих сигналов в форме особого вида
электромагнитного поля. Стало ясно, что "волновые гены" могут
существовать, в частности, как одна из форм явления возврата Ферми-Паста-Улама.
Это фундаментальное явление и легло в основу конструкции генератора
Ферми-Паста-Улама, приближенно моделирующего знаковые электродинамику и
акустику ДНК и потому способного "считывать" и ретранслировать
управляющие метаболизмом биосистем солитонные структуры с хромосомного
континуума биосистем-доноров и резонансно вводить их в геном
биосистем-акцепторов [7] .
Подтверждением предположения А.Г. Гурвича стала
работа доктора Х.С.Бурра (Йельский университет, США) [8]. Им был сконструирован
прибор, позволяющий регистрировать слабые электрические напряжения вблизи
живого объекта. После такого экспериментального подтверждения наличия некоего
энергетического поля доктор Бурр высказал гипотезу, по которой поле это
представляет собой как бы матрицу, исходный чертеж, формирующий структуру тела.
"Молекулы и клетки человеческого тела, - пишет он, - постоянно
перестраиваются, разрушаются и пополняются свежим материалом, поступающим из
пищи. Но благодаря контролю поля новые молекулы и клетки воспроизводятся по тем
же схемам, что и старые. Электромагнитную природу тонких тел живых существ
доказывал А.С. Пресман [7]. Он опровергает материалистическую точку зрения на
живой организм, как на систему биологического вещества. На основании
результатов своих многочисленных экспериментов Пресманом продемонстрировано
влияние слабых (в том числе естественных) электромагнитных полей на
биологические объекты. Эффект от воздействия электромагнитных полей определялся
не их интенсивностью (напряжённостью), а другими факторами, среди которых можно
назвать режим модуляции и направление вектора индукции. Автором высказывается
гипотеза о фундаментальной роли электромагнитного поля как носителя информации
в живой природе. При этом биологические эффекты воздействия поля зависят не
столько от величины их энергии, сколько от информационной насыщенности
воздействия.
В середине 1970-х годов советские учёные В.И.
Инюшин и В.Г. Адаменко сообщили о неких фантомных эффектах, наблюдавшихся у
повреждённых листьев растений при фотографировании их по методу Кирлиан. Сам
метод газоразрядной визуализации в высокочастотных высоковольтных электрических
полях (метод Кирлиан), был известен уже достаточно давно (с 1950-х годов), но
по стечению обстоятельств не привлёк к себе особого внимания
учёных-исследователей. Однако с этого момента дело начало развиваться по
другому. Учёные, получившие удивительные фотографии, на которых обрезанные
листья растений выглядели целыми, высказали предположение о существовании
невидимой глазу энергетической структуры биологических объектов, по которой
последние строят свою форму. Её назвали биополем [9].
Вокруг вопроса о том, чем же в действительности
являются изображения, проявляющиеся после удаления части листа растения,
разгорелась жаркая полемика. Лишь в 1979 году группа индийских исследователей
во главе с Ж.К. Чоудхари привела не только убедительные результаты, но и
подробную схему генератора, с помощью которого выявлялись фантомы удалённых частей
листа [10]. И хотя среди учёных, занимающейся кирлианографией, до сих пор нет
единства в этом вопросе, исследователями П.П. Гаряевым и его сотрудниками были
достигнуты большие успехи в части обоснования природы этого явления. В своей
работе они говорят, что, усовершенствовав установку для получения
фотоизображений методом газоразрядной визуализации (ГРВ), они добились
устойчивого получения различных фантомных эффектов, которые могут быть признаны
электрическим или полевым планом достройки части листа до целого. В
доказательство своего утверждения авторы приводят опыты, когда фантомные
эффекты были получены ими без применения метода ГРВ с использованием
специальных высокочувствительных фотопластинок для ядерных исследований. В
работе утверждается, что источниками фантомов могут быть только излучения
хромосомной ДНК в оставшейся неповреждённой части листа. Этот же механизм
голографической ассоциативной памяти, по мнению авторов, объясняет фантомные
боли после ампутации конечностей людей, а также сохранение памяти у людей в
случаях, когда часть мозга разрушена вследствие травмы или операции. Делается
попытка объяснить механизм записи и считывания голограмм - кодов с хромосом
организма. Обращается внимание на эффект "памяти" воды [11].
Исследовательской группе по физической химии из университета в Пенсильвании,
молекулы составляют в воде кластеры геометрически правильной формы в виде пяти
"тел Платона": тетраэдр а также исследователям из университета в
Беркли (Калифорния), удалось установить, что, гексаэдр, октаэдр, додекаэдр и
икосаэдр. С открытием "тел Платона" в структурах воды новейшие
научные исследования сомкнулись с тысячелетними древними знаниями. Вода
обладает единственной в своем роде космической резонансной способностью.
Поскольку "тела Платона" существуют во всей Вселенной, вода путем
резонирования с вибрационным образцом такой же формы может воспринимать
информацию и отдавать ее вновь.
Подобные скопления молекул воды являются ее
памятью, потому что она накапливает информацию в своей такой разнообразной
структуре. То, что образование кластеров не является продуктом случая, можно
увидеть по структурам кристаллов льда и снега, которые существуют в
бесконечных, но всегда правильных вариантах.
П.П. Гаряев, развивая идеи А.Г. Гурвича, в
другой своей работе доказывает, что синтез белка является результатом
генетического кодирования на волновом уровне. Генетическая память трактуется
П.П. Гаряевым и Е.А. Леоновой как солитонно-голографическая. Утверждается, что
хромосомы излучают свет и звук, что гены расщепляются на вещество и поле.
Методом спектроскопии корреляции фотонов в 1985 году П. П. Гаряеву удалось
зафиксировать необычные аномально долго затухающие звуковые колебания ДНК. Было
установлено, что ДНК обладает способностью синтезировать "незамолкающую
сложную мелодию с повторяющимися музыкальными фразами". Геном высших
организмов рассматривается авторами как солитонный биоголографический
компьютер, формирующий пространственно-временную структуру биосистем по
волновому образу-предшественнику. Таким образом, геном работает не только на
вещественном, но и на волновом (тонкоматериальном) уровне. Обращается внимание
на единство фрактальной структуры ДНК и человеческой речи. На основании этой
связи становится возможным воздействовать на ДНК посредством особых генераторов,
преобразующих речевые алгоритмы в солитонные модулированные поля. Далее Гаряеев
указывает что, по внешним данным, запись первичной генетической информации
происходит на квантово-волновом уровне в виде голограмм и текстов. Информация
поступает изнутри организма, но обусловлена космическими факторами. Гены
принимают ее и предают от клетки к клетке. Частным случаем
вакуумно-информационных процессов выступает феномен сознания, которое ни в коем
случае не локализует в одних лишь мозговых клетках, а проявляется в их
неразрывной взаимосвязи с другими объективными структурами окружающего мира, в
том числе и остающимися вне поля зрения современной науки.
В одной из своих последних работ Гаряев
рассматривает проблему расши- рения модели волнового генетического кодирования.
Он пишет что, исследования возглавляемой им группы доказывают, что синтез белка
- лишь один из примеров генетического кодирования на волновом, стратегическом
уровне и трактует генетическую память расширено как солитонно-голографическую,
свойственную генному био-компьютеру. Кроме того Горяев с сотрудниками в целях
более глубокого понимания волновой природы генома создали концепцию
когерентного физического излучения, голографии и солитоники фрактального
отображения структур ДНК и человеческой речи. Суть идей
Горяева-Березенина-Васильева "ГБВ модель" состоит в том, что геном
высших организмов рассматривается как солитонный биоголографический компьютер,
формирующий пространственно-временную структуру развивающихся эмбрионов по
каскадам реесторов волновых образцов-предшественников. При этов в качестве
носителей полевых генов выступает континуум волновых фронтов, задаваемый
мультиплексными геноголограммами, образуемыми гелевым жидкокристаллическим
хромосомным континуумом. Акт "считывания" информации осуществляют
сами же хромосомы, генерирующие лазерные свет и звук в широких диапазонах.
Близкую роль играют также солитоны ДНК - особый вид акустических и
электромагнитных полей, продуцируемых генетическим аппаратом самого организма и
способных к посредническим функциям по обмену стратегической регуляторной
информацией между клетками, тканями и органами биосистемы. Важно так же и то,
что квазиголографические решетки в том числе входящие в сотав колебательных
структур солитонов, являются лишь частным простейшим случаем кодово-образной
информации, зафиксированной в хромосомном континууме организма.
Мультиплетнокодовая ДНК, где аминокислотный код только малая часть, и
"эгоистическая" ДНК хромосомам анализируется в рамках ГБВ модели как
потенциальный стратегический информационный вектор всех клеток и тканей
организма, включая кору головного мозга. Геном работает не только на
вещественном на и на волновом, на "идеальном" (тонкоматериальном)
уровне. Это идеальная компонента, которую можно назвать супергенно-континуумом,
и является главной знаковой фигурой генома, обеспечивающей развитие и жизнь
человека, животных, растений, а также их программируемой естественное умирание.
Вместе с тем важно понять, что нет резкой и непреодолимой границы между
волновыми и материальными уровнями хромосом. Оба они образуются вещественными
матрицами, но гены дают материальные реплики в виде РНК и белков, а супергены
преобразуют падающие на них эндо-и экзогенные физические поля, формируя из них
пространственно-временные разметочные волновые структуры. Более того, гены
могут быть составной частью голографических решеток супергенов и регулировать
их полевую активность. И наконец, супергены могут формироваться как ДНК - РНК -
нуклеопротеид - лазерное поле промодулированное их текстами.[12].
По словам Горяева, обычный шум в эфире не
разрушает генетический аппарат. Он ничего не понимает в таком
"радиогудении" и поэтому не реагирует на него. Но достаточно тихонько
"шепнуть" одно слово, как аппарат на него живо откликнется. От
громкости радиозвука мало что зависит. Весь секрет в смысловом содержании. Ведь
ядра клеток обмениваются информацией с помощью кодовых слов. Если они
образуются из внешних волн и содержат негативный смысл, клетки испытывают
электромагнитный шок. Они начинают вырабатывать особые белки, которые вызывают
тяжелые расстройства и даже смерть. - Еще один, казалось бы, парадоксальный
вопрос. С древнейших времен монахи и отшельники проводили жизнь в молитве -
непрестанно славили Бога. Долгие десятилетия это считалось чуть ли не мракобесием.
Исследования в области волновой генетики показали, что все наоборот: монахи
поступают мудро - благодатными песнопениями создают в организме такой волновой
режим, который защищает их от грязи в эфире. Слова молитвы включают
наследственные программы оздоровления души и тела. Дело не столько в физическом
действии акустических волн, сколько в смысле слов.
Исследователи провели эксперименты с зернами
пшеницы и ячменя, которые получили дозы от 2 до 10 тысяч рентген. Частицы
высоких энергий разрывали хромосомы - казалось зерна уже мертвые. Но ученые
обрабатывали их особыми электромагнитными волнами, на которые были наложены
акустические колебания человеческой речи. И не простой, а ритмичной, обращенной
к высокому идеалу - проще говоря, молитвами. В результате в зернах
образовывались волновые "каркасы" здоровых хромосом, и на них
восстанавливались разорванные хромосомы. Генетический аппарат клеток чудесным
образом избавлялся от повреждений. Молитва - сильнее радиации.
У Литтлом (1965) принципиально доказывается
возможность сверхпроводимости в биологических молекулах. Следовательно, энергия
может переноситься в живых системах практически без потерь. Литтл доказывает,
что в органической молекуле, состоящей из длинной цепочке атомов, происходит
свободное перемещение электронов вдоль её оси. Вся эта, определённым образом
организованная система, является заряженными частицами в условиях организма
четвёртым состоянием живого вещества или биологической плазмой - материальной
основой биологического поля [3].
В своей книге "Тайны русского народа"
В. Дёмин приводит цитату английского биолога Лайэлла Уотсона: "Каждое тело
имеет биоплазменного двойника, который существует на менее физическом уровне,
принимает приблизительно те же формы, что и тело, и имеет некоторое отношение к
контролю и организации жизненных функций. Его нелегко измерить, но его
существование вытекает из практики иглоукалывания и может обнаруживаться с
помощью специальной техники, состоящей из высокочастотной аппаратуры. Он не
исчезает в момент клинической смерти". Глубинное информационное поле
Вселенной кодирует и хранит в виде голограмм любую информацию, исходящею от
живых и неживых структур. С незапамятных времен многими великими умами
утверждалось, что в любой точке Мироздания содержится информация обо всех
событиях и сущностях Вселенной. Информация не хранится пассивно, а отбирается и
передается в необходимых дозах, в необходимое время и в необходимом
направлении. Процессы эти невозможны без непрерывной энергетической подпитки и
информационного круговорота, в ходе которого возникают устойчивые смысловые
структуры, сохраняемые и передаваемые от одних носителей (живых и неживых)к
другим. Сказанное относится и к слову. Перефразирую афоризм средневекового
индийского мудреца Бхатрихари:"Бесконечный, вечный Брахман (космическое
всеединство) - это сущность слова, которое неуничтожимо", можно сполной
уверенностью утверждать: "Бесконечное, вечная Вселенная непрерывна
порождает и накапливает разнокачественную информацию (включая слово). Поэтому
эта информация неуничтожима и вечна как сама Вселенная, как весь бесконечный
Космос. Включая слово".
В указанном смысле устное Слово - это
направленное волевым усилием акустическое выражение внутренней энергии
индивида, приводящее в движение механизм раскодирования информации на различных
физических уровнях, включая глубинный, - пока во многом неизвестный и
неисследованный. Являясь объективной аккустико-энергосмысловой структурой,
Слово непосредственно замыкается на информационный банк (поле) Вселенной и
репродуцирует заложенное в нем знание.
Слово как звуковая речь - всегда представляет
собой акустические колебания молекул, но одновременно - и волновые колебания
образующих их атомов, элементарных частиц и соответствующим образом
закодированные поля. При этом слово не только кодирует низшие формы движения
материи, образуя и постоянно обогащая информационное поле, но так же черпает из
этого поля энергию и передает ее в случае необходимости обратно к источнику,
производящему Слово, т.е. к человеку. Так, гневное слово возбуждает того,
против кого оно направлено, не через одно лишь осмысление, но и через
возникаемую энергию отрицательного эмоционального состояния, которое возбуждает
информационное поле и распространяет его вокруг себя. Тоже - со смехом. Можно
не знать языка, выражающего гнев или радость, но плакать и смеяться наравне со
всеми.
В определенной мере положительные и
отрицательные эмоции могут возникать под непосредственным воздействием
окружающего энергоинформационного поля. Резкие выкрики при выпаде и ударе
восточных единоборствах, русское "гых-х-х", при рубке дров
концентрирует энергию в соответствующем направлении и сообщает человеку
дополнительную силу именно за счет подпитки, почерпнутой из общего энергополя
Вселенной, и за счет включения соответствующих каналов под влиянием его
информационной составляющей.
Аналогичный эффект дают воинственные крики на
поле брани (вроде русского "Ура") повышая энергетический потенциал
сражающихся людей. Крик от боли или страха также мобилизует энергетические
ресурсы организма, противодействуя факторам, вызывающим боль.
Взаимодействие между живым организмом и
принадлежащим ему биополем, с одной стороны, и информационно-энергетическим
фоном Вселенной, с другой, может принимать самые причудливые формы. Так
кровавое жертвоприношение в прошлом имело, судя по всему, колоссальный
психофизический эффект: на месте жертвоприношения возбуждалось и менялось
информационное поле, а энергия перераспределялась в пользу приносящих жертву
(отсюда столь массовое и повсеместная приверженность к подобным кровавым
спектаклям). Тоже, видимо, происходит и во время поединков,: побежденный теряет
свой энергетический потенциал и частично передает его победителю. В особенности
это касается смертельной схватки, когда один из соперников гибнет.
Давно замечено и эффективно используется так же
и психофизическое воздействие слова, музыки и песнопений, концентрируемых под
сводами Храмов всех без исключения религиозных культов. Здесь действует
одновременно и чисто внешняя сторона (купол-стены) и целенаправленное
воздействие энергоинформационного поля, приводящее к таким психологически
нетривиальным следствиям, как молитвенный экстаз, благодать, очищение
(катарсис), успокоение и такого типа.
Те прозрения, откровения, голоса и ведения,
которые случаются во время молитвы (независимо от того, какую религию
исповедует верующий), дают некоторое представление о формах возможных контактов
с энергоинформационным полем. Их универсальность и абсолютная несвязанность с
конкретным содержание религиозного культа - лучшее доказательство доступности
для любого человека параллельно существующего мира. Сказанное в одинаковой
степени относится к индивидуальным, и к коллективным ритуальным актам в виде
хоровых песнопений, всенощных бдений, массовых молитв или танцев. Однако
коллективное действо, вне всякого сомнения, усиливает эффективность контактов
между впадающими в экстаз больших или малых групп людей, с одной стороны, и
возбуждаемым ими параллельным миром, с другой.
Таким образом, не приходится сомневаться, что
существует некое общее непрерывное и всепронизывающая материальная среда,
именуема "физическим вакуумом" и вполне объясняемая при помощи
концепции взаимодействия и взаимопроникновения Макро- и Микрокосма человека. В
результате разности потенциалов движения и энергии в "физическом
вакууме" возникают устойчивые структуры, обеспечивающие все многообразие
живого и неживого во Вселенной. Здесь же кроется разгадка
информационно-генетических закономерностей. [13].
Необычный подход применил в своих исследованиях
Г.П. Крохалёв. В своей работе он доказывает реальность психической энергии, как
энергии сознания человека, обращает внимание на психотронику как на область
знания, изучающую проявления энергий психических актов человека Г. П. Королёв
утверждает, что экспериментально установил, что при зрительных галлюцинациях
происходит обратная передача зрительной информации от центра зрительного
анализатора к периферии с электромагнитным излучением из сетчатки глаз в
пространство зрительных образов в виде плоскостных или объёмных
(голографических) изображений. Также была проверена область слуховых
галлюцинаций, которые с помощью специальной методики фиксировались на магнитной
плёнке обычного магнитофона, в слуховых галлюцинациях происходит обратная
передача слуховой информации от центра слухового анализатора к периферии с
звукоизлучением Кортиева органа [14].
Подобную работу провёл доктор
физико-математических наук Анатолий Серобабин [15]. Он занимался
фотографированием мысли людей как голографического изображения, проецируемого
из глаз экстрасенсов. В 1996 году учёный направил заявку на открытие "Об
экспериментальном обнаружении физических полей, вызванных концентрацией
мысли".
Но наибольшего успеха в области регистрации
энергетического воздействия мысли добился Роберт Джан - руководитель
лаборатории по изучению аномальных явлений Пристонского университета (США)
(Цит. по Гаряеву). Он провёл большое количество опытов по воздействию
операторов на генераторы случайных чисел, которые управляли движением
механического робота. Было экспериментально доказано, что под воздействием
операторов движение робота из случайного становилось более упорядоченным и шло
в одном из определённых направлений.
В России подобные опыты производятся под
руководством Юрия Попова - профессора Московского инженерно-физического
института. В центре студенческих инициатив был сконструирован свой
чувствительный прибор, регистрирующий мысленное воздействие. При этом, по
мнению Ю. Попова, для влияния на прибор не обязательно находиться в
непосредственно близости от него. Достаточно лишь мысленно представить его
[16].
По мере развития биофизики, как науки о
физических процессах в биологическом пространстве, появляется новое направление
- биофизика индивидуальности. Каждый человек имеет свою уникальную
биофизическую структуру, т.е. биофизическую индивидуальность. В России, в
городе Уфе, в Башкирском университете это направление развивается доктором
психологии, профессором Г. А. Аминьевым [17].
Любой психический процесс - эмоции, мысли и т.д.
- сопровождается изменением энергетического пространства вокруг личности. При
этом изменяется структура пространства. Пространство, энергия и время
взаимосвязаны. А. Бергсон первый высказывался по теории биологического времени,
считая, что психическое время отличается от физического времени. С точки зрения
биофизики индивидуальности или основ психоэнергетики необходимо учитывать
временную структуру личности (речь идёт о познании особенностей психического
времени). Психика имеет биоэнергетическое содержание, но ее состояние зависит
от экзогенной гелиогеофизической среды [18].
Нельзя думать, что психика - это изолированная
система, не реагирующая на внешние воздействия. Взаимодействие психики с
экзогенной средой - космическими излучениями, магнитными бурями на Солнце,
электромагнитными полями, количеством зарядов в атмосфере Земли, холодной
геоплазмой Земли - являются важными составляющими в единстве: человек - Земля -
Космос. В работе А.Л. Чижевского "Физические факторы исторического
процесса" показана взаимосвязь солнечной активности с психическими факторами
исторического процесса на Земле. Выброс солнечной плазмы, пятна на Солнце
вызывают психические изменения у огромных масс людей на планете Земля.
Психоэнергетический статус отдельного человека и масс людей в значительной
степени зависит от космических и геофизических флуктуаций [19].
Космическое воздействие обусловлено, прежде
всего, действием Солнца, фазами Луны, процессами, происходящими в Солнечной
Системе и Галактике. Такие факторы влияют на большие массы людей в целом и на
отдельные личности, в частности. Все возмущения, которые происходят в Земле,
отражаются, прежде всего, через геоплазму Земли на биоплазме мозга человека.
Так, человек, живущий в какой-то геоаномальной зоне, может чувствовать приливы
психической энергии или, наоборот, депрессию, которая может приобрести окраску
психосоматического заболевания. Можно полагать, что одной из причин многих
болезней является действие аномалии геоплазмы на человека в течение длительного
периода его жизни. Флуктуации или возмущения в геоплазме постепенно разрушают
биоэнергетику и психоэнергетику человека. Причем характеристики геоплазмы, а,
значит, и биоплазмы, изменяются в зависимости от фаз активности Солнца [16].
Любое цивилизованное общество строилось на
энергетической пирамиде, основание которой представляет энтропийная энергия
естественной и техногенной природы. Далее - биологическая энергия живой
природы, плодами которой мы пользуемся. Она занимает большую часть пирамиды.
Вершину пирамиды представляет высокоорганизованная форма энергии - пси-энергия.
Флуктуация энтропийного энергетического основания приведёт к сдвигам во всей
пирамиде. Техногенизация общества приводит к падению психоэнергетического
потенциала и его деградации. Увеличивается энтропийный базис, понижается высота
вершины пирамиды. Кроме того, А.Л. Чижевский указывал на глобальность влияния
экзогенных факторов солнечной активности на гео-, био- и пси-энергетику.
Всплески солнечной активности расшатывают всю пирамиду, приводят к
деградационным процессам, нарушают безопасность психосоциальной сферы. Происходят
глубочайшие возмущения в биосфере и ноосфере (сфере разума) [20].
Многие филососфы, психологи считают, что
психические процессы лишены материальной основы, определяются только через
понятия, образы и на этом строили познание психики человека. Однако, уже З.
Фрейд понимал, что без энергии понять психологические процессы и нарушения в
психике на атомно-молекулярном уровне невозможно, и искал основу новой
парадигмы. Существующее механистическое мировоззрение не могло объяснить всех
проявлений, которые происходят в пси-мире человека (Цит. по Гаряеву).
Необходимость разработки совершенно новых
понятий в биологии и физике возникла в связи с необходимостью объяснения
феномена биологического целого, который впервые в отечественной литературе был
сформулирован А.Г. Гурвичем на строгой экспериментальной и единственно научной
эпигенетической основе. А.Г. Гурвич впервые сформулировал положение о
"факторе целого" для клеточной ориентации на основании движения
эпителиальных клеток при развитии головного мозга позвоночных (зародышей
акулы). Он сформулировал следующую теорему: "При формообразовании клетки
ориентируются в своем движении так, как будто они притягиваются некоторой
силовой поверхностью, совпадающей с границами окончательной поверхности".
Эта силовая поверхность динамически преформированной морфы получила название
"предморфа" [21].
Предморфа представлена структурированным
физическим вакуумом, виртуальной биоплазмой. Вакуум имеет свое время,
пространство и наполнен волнами и торсионными полями. Первоначально А.Г. Гурвич
отмечал, что источник поля не связан с каким-либо материальным носителем.
Однако, принцип поля, даже в его геометрическом
воплощении, оказался весьма полезным. Позднее, в 20-е годы, Гурвич полагал, что
каждая хромосома является носителем поля, длинная ось которого совпадает с ее
осью. Поле вне хромосом рождается ими и управляет развитием. ДНК содержит
стратегические пространственно-временные коды, именно поэтому она не
элиминируется естественным отбором. Клеточное деление - митоз - сопровождается актами
сверхслабого излучения, которое имеет физиологическое значение.
Митогенетическое излучение, как полагают последователи школы А.Г. Гурвича,
является одним из факторов, регулирующих физиологические и морфологические
процессы, как на клеточном, так и органном уровнях [22]. Так, Р. Шелдрейком
(1981) выдвинута концепция о морфогенетических полях. Предполагается наличие
специфических голограмм, ответственных за характер, форму и организацию
биосистем на всех уровня сложности. Они упорядочивают системы, связанные с
ними, влияя на события, которые, с энергетической точки зрения, кажутся
недетерменированными или вероятностными. Делается вывод, что если
морфогенетические поля ответственны за организацию и форму материальных систем,
то они сами должны обладать характерными структурами. Следовательно, каждый
организм, в том числе и человек, находится под влиянием морфогенетических
следов, оставленных в физическом вакууме. Таким образом, генетическая программа
находится не в зиготе, а в физическом вакууме в виде волновой структуры
(биоголограммы) [23].
Известно, что современная генетика не монет
ответить на вопросы, связанные с пространственным становлением живых объектов,
пользуясь только биохимическим и молекулярным подходом. В настоящее время
назрела необходимость по новому взглянуть на процессы морфогенеза. В новом
подходе недостаточно останавливаться только на генетическом программировании
пространственного развития живых существ. Ряд исследователей: (Р. Кастлер, X.
Равен), подсчитав количество информации в зиготе и в развивающемся организме,
пришли к выводу, что количество информации в сформированном организме
возрастает на несколько порядков, по сравнению с той, которая была вначале
развития. В сложившихся условиях не обойтись без теории пространственных информационных
матриц, которые принимают непосредственное участие как в онтогенезе, так и в
историческом развитии -филогенезе. Концепция пространственных информационных
матриц, складывающих общее информационное поле, гласит:
Информационное поле образовано пространственными
биоматрицами, представляющими собой фрактальные информационные структуры,
непосредственно связанные со спиралями ДНК и спирализованными хромосомами,
которые выходят на следующий организационный уровень [14].
Пространственный анализ показывает, что
фрактальные структуры и спирали могут контактировать друг с другом и служить
переходным мостом, где информация биоматриц непосредственно реализуется
материально в процессе морфогенеза, определяя дифференциальную активность генов
в ДНК.
Биологические поля не могут быть общими и
одинаково организованными, как это мы находим в неживой природе. Существуют
электрическое, магнитное, гравитационное поля, которые имеют одинаковую
организацию во всех живых и неживых субстанциях. Биологическое поле
индивидуально для каждого организма, и поэтому ему дано название
"информационное поле", элементом которого могут быть биоматрицы[ 23]
.
В ходе рассмотрения разнообразных концепций
доказывается, что морфогенетическое поле связано с генетическим аппаратом и
представляет собой сложный паттерн, включающий все известные и неизвестные
поля. Это слабоэнергетическое, но высокоинформативное комплексное поле
индивидуально и его следует называть информационным. Развитие морфогенетической
системы продолжается до тех пор, пока не сгладится пространственное
несоответствие между развивающееся структурой и информационным полем или
информационной биоматрицей. Данные подтверждаются экспериментами на ряде
гидробионтов [4]. Биоматрицы используются живыми организмами, как в филогенезе,
так и в онтогенезе.
Одним из разделов, в проблеме митогенеза
излучения, является вопрос об излучении нервной системы. Оно характерно для
всех отделов нервной системы и присуще любым физиологическим состояниям. А.Г.
Гурвич обозначил контуры биоголографической концепции, в которой состояние
клеточного субстрата детерминируется или, другими словами, подчиняется
пространственным директивам доминирующего фактора - биополя. Введение научного
понятия биополя в биологию имеет огромное, революционное значение для понимания
особенностей живого состояния. Следует вспомнить, что академик В.И. Вернадский
подчеркивал необычную организованность живого вещества по сравнению с неживым.
При изучении живого вещества мы уже имеем дело с гетерогенным пространством.
Живой организм представлен в пространстве веществом и полем. Живой организм
является многомерным "конденсированным" пространством, конфигурация
которого в значительной мере обусловлена действием полей микрокосмоса. В отрыве
от физических полей биополе рассматривать нельзя [6].
Нельзя не отметить фундаментальные разработки
профессора Н.И. Кобозева, который показал, что живая система обладает
отрицательной энтропией, и для нее не существует полной процедуры
воспроизведения. Сама многомерность биологического пространства неизбежно
приводит к постулату о невоспроизводимости биоструктур. В этом уникальность
пространственно-энергетического состояния биоплазмы - одно из проявлений
живого. Уникальность биоэнергетической структуры живого, и, прежде всего,
человека, накладывает принципиальный запрет - "табу" на создание
искусственного живого интеллекта, искусственной пищи, воспроизведение жизни из
неорганических компонентов (цит. по Гаряеву) [5]. Основные положения Н.И.
Кобозева следующие:
• Мозг как биохимическая, молекулярная, клеточная
и нейрофизиологическая система не способен обеспечивать процесс мышления.
• Для обеспечения процесса мышления необходимо
наличие антиэнтропийных структур, обеспечивающих подавление теплового движения
высокомолекулярных компонентов мозга.
• Вещественный вакуум является необходимым
атрибутом жизни.
• Обогащение биологических систем вещественным
вакуумом повышает их жизнестойкость, а обеднение приводит к достаточно
серьезным заболеваниям.
• Одним из путей поступления вещественного
вакуума в организм человека является пища, важнейшим показателем которой
является концентрация в ней вещественного вакуума.
• Изучение свойств вещественного вакуума - один
из возможных ключевых подходов к пониманию общих свойств материи.
Психика, как антиэнтропийная структура, обуславливает
четкость взаимодействия биоголограмм - формирует их проявление, регулирует
уровень разнообразия и накопления. Организатором процессов "синтеза
узоров" является психика, как верхний этаж всей биоэнергетической основы.
Тонкая инфраструктура психики своими корнями уходит в глубинные структуры
организованной материи. Следовательно, антиэнтропийную компоненту нужно искать
в таких биоэнергетических структурах. Хотелось бы подчеркнуть, что без
признания биоплазмы как матрицы для поддержания устойчивых когерентных волновых
структур невозможно понять феномен жизни.
Биоплазма - организованная плазма, пятое
состояние вещества. В целом, в живой клетке все плазменные структуры, куда
входят и виртуальные частицы, образуют единый биоплазменный ансамбль клетки,
который является целостной системой, гомеостаз которой тесно связан с
устойчивостью атомно-молекулярных компонентов (вода, органические молекулы и
т.д.). В клетке можно выделить три вида биоплазмы:
. Соматическую, локализованную в мембранах.
. Гермобиоплазму, заполняющую преимущественно
ядерные структуры.
. Пси-биоплазму, интегрирующую клетку как целое
и обеспечивающую межклеточные коммуникации в тканях и органах.
Соматическая биоплазма состоит из лёгких частиц
(электронов, экситонов и т.д.). Обуславливает многообразие соматических
процессов - двигательную активность, мышление. Соматическая биоплазма менее
энергоёмкая, более лабильная, с анизотропными свойствами с доминированием
кинетических форм энергии.
Биоплазма гермы - тяжёлая организованная плазма,
которая представлена протонами, ионами, частицами с большой массой. Биоплазма
гермы наиболее энергоёмкая, с большим запасом потенциальной энергии, с меньшей
анизотропией. Создаёт энергетическую основу для размножения, с очень высокой
стабильностью и устойчивостью. В ней находит отражение наследственный ключ для
входа в физические вакуумные структуры.
Пси-биоплазма - биоплазма мозга и других
структур, обладающих псибиоголограммами, имеющие антиэнтропийные вакуумные
структуры [9].
Условно, в целом организме можно выделить единое
"биоплазменное тело", которое имеет микро- и макроструктуру. В нем
представлены три вида биоплазмы: соматическая, герма- и пси-. У человека все
три структуры гармонично развиты. Энергетические потенциалы гермоплазмы и
соматоплазмы в онтогенезе меняются за счет основного биоэнергетического
процесса. Сущность основного процесса: "В многоклеточном организме
происходит увеличение энергетического потенциала одних клеток за счет
уменьшения потенциала у других". Можно предположить, что при деградационных
процессах происходит симметричное разделение энергетического потока. Часть
энергии соматической биоплазмы рассеивается в виде тепла, обесценивается
согласно второму началу термодинамики, другая часть перебрасывается в
гермобиоплазму, повышая ее биоэнергетический потенциал [24].
Разница потенциалов между внутренней и внешней
стороной клеточной оболочки является причиной возникновения токов активности у
нервов. Измерение электрического потенциала мозга нашло своё применение в
электроэнцефалографической диагностике [25].
Открытие так называемого "пика" в
активных точках окончательно решило вопрос о существовании электромагнитных
полей в живых системах.
Электромагнитное излучение мозга изучается
сейчас довольно активно в связи с биологической телекоммуникацией. Нормально
работающий человеческий мозг представляет собой станцию электромагнитного
излучения силой в 2*10-10 Вт в широкополосном спектре с длиной волны
от сантиметров до тысячи метров.
Через биоплазму осуществляется миграция энергии,
её перераспределение согласно пространственной матрице и проводящим путям.
Биоплазма трансформирует энергию из одного вида в другой и является основной
структурой, обуславливающей раздражаемость. Стабильность параметров биоплазмы
находится в тесной зависимости от электрического состояния окружающей среды
[26].
Таким образом, микроструктура биоплазмы имеет
место уже на субмолекулярном уровне. В живой клетке биоплазмой заполняются
мембранные системы. В этих нитевидных структурах, проявляющих
жидко-кристаллические свойства, находится лабильная электронно-дырочная плазма,
она и обуславливает высокую функциональную активность биологических мембран. В
клеточном ядре сосредоточены большие плотности электронно-протонной плазмы
[25].
Мозг человека - вершина структурности
соматогермобиоплазмы. Биоплазма в мозгу человека имеет наибольшую плотность
(1038 электронов в 1 см3). Она насыщена суперлёгкими частицами, которые
подавляют энтропийные явления. Мы полностью согласны с Н.И. Кобозевым в том,
что жизнь на всех своих уровнях содержит антиэнтропийную компоненту, у мозга
человека уровень антиэнтропии наиболее высокий.
Мыслительная продукция мозга невозможна на
атомно-молекулярном уровне, и даже биохимического материала, безусловно,
недостаточно для создания мыслящего мозга. Участие элементарных частиц
расширяет возможности мозга как антиэнтропийного генератора [27].
Термодинамическое рассмотрение процессов
информации и мышления приводит Н.И. Кобзева к необходимости допущения
существования устойчивого комплекса элементарных частиц. Мы считаем, что этот
комплекс и есть биоплазма. Соматическая биоплазма имеет очень сложную
структуру, наличие затухающих волновых структур. И здесь имеет место
биоголограммное состояние поля биоплазмы. Биоплазма, как организованная
структура, является еще и излучающей системой, она генерирует композиционное -
упорядоченное поле со сложной конфигурацией - биополе [28].
Мысль возникает из "пустоты", из
структурированного вакуума, накопленного поколениями предков [29].
Первые следы биоплазмы в эксперименте были
обнаружены с помощью эффекта Кирлиан. Создание новой, биоголографической,
системы понимания жизни дает основу для научно-технической революции, создает
предпосылки для осуществления действенной природоохранительной стратегии,
разрешения проблемы укрепления здоровья человека, открывает новые перспективы в
развитии экопсихологической безопасности общества [30].
Основной биотермодинамический процесс, открытый
Э. Бауэром еще в 1935 году, отчетливо проявляется в динамике
психоэнергетической структуры общества.
Принято считать, что нервный импульс прост, что
он является всего лишь волной деполяризации, и поэтому шифровка информации
здесь происходит только по частотному механизму, но частотные модуляции
недостаточны для кодирования. Исследование А.А. Березина показало, что нервные
импульсы - это все те же солитоны (электромагнитные волны) в рамках явления
возврата Ферми-Паста-Улама , при этом такие солитоны отображают в своей
структуре РНК-"тексты", а следовательно, и ДНК. В развитии этой идеи
выдвинуто предположение о знаковой смысловой связи солитонов на хромосомном
уровне и солитонов нервных импульсов. Это свидетельствует в пользу связи
сознания и его отображения - слова с основной информационной фигурой биосистем
- молекулами ДНК с их новыми, неизвестными ранее, типами памяти. Солитоны ДНК
способны после "прочитывания" ими генотекстов покидать пределы ДНК в
форме особых электромагнитных полей с тем, чтобы узнавать нужные участки других
молекул ДНК и формировать целостные новые "тексты", необходимые в
данный момент для выполнения биофункций, в том числе и путем передачи
информации в нервные импульсы [7].
Используя эти теоретические разработки, удалось
экспериментально доказать возможность свертки генетической информации в форме
солитонных волновых пакетов. Такие волновые пакеты с искусственно введенной в
них биоинформацией, продуцируемые соответствующими Ферми-Паста-Улама-
радиоэлектронными устройствами, способны входить в резонансный информационный
контакт с генетическим аппаратом животных, растений и, вероятно, человека с
последующим резким и направляемым изменением их обмена веществ. Вводя
определенные кодовые вербальные команды через генератор Ферми-Паста-Улама в
генетический аппарат радиационно поврежденных семян растений, удалось
достоверно уменьшить число хромосомных аберраций, то есть фактически
блокировать поражающее действие рентгеновского излучения. Более того,
оказалось,что возможна превентивная защита генома растений от жесткого
рентгеновского излучения с помощью адекватных волновых команд [31].
Существует и другая семантическая ниша знаковых
процессов в наследственном аппарате высших биосистем, связанная с его
квази-речевыми характеристиками, а также с генетической атрибутикой
словообразований в естественных человеческих языках. Ранее получены
доказательства, что развитие языков и человеческой речи подчиняются законам
формальной генетики. По сути, "тексты" ДНК (квазиречь) и письменность
людей, их разговор (истинная речь) выполняют одинаковые управленческие,
регуляторные функции, но в разных фрактально-сцепленных масштабированиях. ДНК
генетически функционирует на клеточно-тканевом уровне, а человеческая речь, как
макрогенетическая структура, используется на уровне общественного
суперорганизма. Выяснилось, что ДНК и человеческая речь (тексты) обладают
стратегически близкой фрактальной структурой в геометрическом смысле. Вероятно,
это каким-то образом коррелирует с фрактальной структурой солитонного
акустического и электромагнитного Ферми-Паста-Улама - поля, генерируемого
хромосомным аппаратом высших биосистем. Возможно, по этой причине и удалось
зарегистрировать управленческие эффекты на геномах растений, вызываемые с
помощью Ферми-Паста-Улама - трансформированной человеческой речи, которая
резонансно взаимодействует с хромосомной ДНК in
vivo [32].
Многие ученые заговорили о возможности создания
биокомпьютера на генетических структурах - молекулах ДНК. Ясно, что при его
разработке необходимо условие реализации возможностей ДНК, которая должна
находиться в привычной среде - в водном растворе или и в жидкокристаллическом
состоянии. Прежде всего, необходимо начать практическое использование новых
типов памяти геноструктур и для этого пытаться конструировать ячейки памяти,
работающие на явлении Ферми-Паста-Улама - резонансов и на способности
записывать голограммы. Такая память будет на многие порядки по объёму и
быстродействию превосходить память существующих магнитных, оптических дисков и
голографических систем [11].
Недостатки современных ортодоксальных теорий о
строении мира привели к появлению все новых и новых гипотез. Многие из них не
выдерживали проверкии временем и забывались. Другие вновь появлялись на
страницах различных изданий. Необходимость существования скорости передачи
информации в Мироздании, если не мгновенной, то по крайней мере многократно
превышающей скорость света, привела к появлению гипотез о существовании
микролептонных, тахионных, торсионных полей. В этих гипотезах делалась попытка
отыскания новых дальнодействующих полей, так как получаемая информация о
Вселенной, существующей только в электромагнитном спектре, оказывалась
недостаточной и не позволяла объяснить многие, если не большинство, из
наблюдаемых фактов [31].
В 1913 году французский математик Эли Картан
указал на возможность существования торсионных полей, возникающих вокруг
всякого вращающегося тела. А в 1993 году Г.И. Шипов в книге "Теория
физического вакуума" обосновал это теоретически. Если гипотетические
гравитационные поля порождаются массой, электромагнитные - зарядом, то
торсионные поля формирует классический спин, представляющий собой квантовый
аналог углового момента вращения. Константа спинторсионных взаимодействий,
служащая показателем их силы, оценивалась величиной не более 10-66,
что первоначально не привлекало к себе повышенного внимания ученых из-за
чрезвычайной слабости этих полей. К примеру, константа электромагнитных
взаимодействий составляет 10-2. Со временем ученые пришли к выводу,
что для динамического кручения, когда создается волновое торс ионное
взаимодействие, снимается ограничение на величину константы. При этом, если константа
становится очень большой, торсионные проявления становятся визуализированными.
Это позволило разработать новые торсионные технологии в области энергетики,
транспорта, связи, коммуникаций и т.д. [29].
В теории торсионных полей присутствует ряд удивительных
свойств, в корне отличающихся от общепринятых научных догм. Энергия и импульс
торсионного поля равны нулю, как и потенциальная энергия спин-торсионного
взаимодействия. Торс ионное поле переносит информацию без переноса энергии. Это
было экспериментально доказано украинскими учеными В.П. Майбородой и И.И.
Тарасюком при воздействии торсионным генератором на кристалл типа
кадмий-ртуть-теллур. При этом наблюдалось изменение магнитных свойств на
величину, требующую в миллион раз больше энергетических затрат, чем было
затрачено на работу торсионного генератора [4].
В отличие от гравитационных и электромагнитных
полей, характеризуемых центральной симметрией, торсионные поля спинирующих
объектов обладают осевой симметрией. Здесь не работает закон обратных квадратов,
поэтому интенсивность торсионного поля не зависит от удаленности от источника
поля и обладает исключительной проникающей способностью в любых природных
средах. В качестве квантов торсионного поля - тордионов, - выступают
низкоэнергетические реликтовые нейтрино. Торсионные поля, схожие по своей
природе c гравитационными, невозможно экранировать. Если гравитация при
моделировании интерпретируется как спиновая продольная поляризация, то
торсионные поля - как поперечная поляризация физического вакуума [29].
Групповая скорость торсионных волн составляет не
менее 109с (с - скорость света). Сверхсветовые скорости присутствуют в теории
гравитации Ньютона и составляют основу концепции гипотетических сверхсветовых
частиц - тахионов, позволяющих в свою очередь объяснить явления
сверхпроводимости и сверхтекучести [7].
Ещё одно уникальное свойство - это взаимное
притяжение одноименных и отталкивание разноименных торсионных зарядов.
Торсионное поле, обладающее "памятью", содержит 24 независимые
компоненты и разлагается на три самостоятельные части. Эти три части поля и
образуют некую общность, называемую торсионным полем [8].
Перспективы торсионных технологий спровоцировали
новый виток техногенного развития, в том числе и в области психотронных
технологий воздействия на подсознание человека, а также на биологические поля
всех живых организмов. К сожалению, трудно предугадать, какие плоды принесут
данные открытия человечеству.
2. Материал и методика исследования
.1 Биоморфологическая характеристика вида
баклажан Solanum
meongena (Сорт
"Донской 14")
По данным "Справочника овощных и бахчевых
культур произрастающих в Казахстане" (Шабыкин Л.В., Колоколова Л.С., 1980
г.) баклажан относится к семейству Паслёновых. В нашей стране - это однолетнее
растение. Плод баклажана - ягода самой разновидной формы. Длина плодов
колеблется от 5 до 20 см, окраска в технической спелости - белая, фиолетовая,
тёмно-фиолетовая, зелёная, а в семенной спелости плоды светлеют, окраска
становится буро-жёлтой или серо-зелёной. Плоды в технической спелости в
зависимости от сорта весят от 50 до 1300 гр. Окраска мякоти белая или кремовая.
Семена светло-жёлтые, чечевицеобразной формы с гладкой, блестящей кожицей.
Основная масса корней баклажана расположена в верхнем горизонте почвы на
глубине до 30 - 40 см. Баклажан - светолюбивое, влаголюбивое и теплолюбивое
растение.
Сорт баклажана "Донской 14" -
среднепоздний. Листья зелёные, слабовыемчатые. Цветок розово-фиолетовый. Плоды
удлинённо-грушевидной формы, средний вес плода около 300 гр. Куст сомкнутый,
высотой 60 - 75 см. Урожайность высокая. Районируется этот сорт во всех
областях нашей республики.
Полезные свойства баклажан Solanum
meongena
В плодах баклажанов содержится около 0,2 %
кислот, 0,1 - 0,4 % жира, 1,2 % белковых веществ, до 1,2 % клетчатки, 0,5 %
минеральных солей и в небольших количествах каротин. Горьковатый вкус
баклажанов обусловливается наличием в плодах соланина (0,004-0,009 % сухого
вещества).
Баклажаны широко используются в кулинарии.
Ценность баклажанов состоит в том, что они способствуют выведению из организма
холестерина, а, следовательно, и предупреждению атеросклероза. Их рекомендуют
употреблять пожилым людям, больным сердечно-сосудистыми заболеваниями, при
отёках и подагре.
.2 Биоморфологическая характеристика вида Перец
сладкий Capsicum
annum L.
(Сорт "Ротунда" )
Перец относится к семейству Паслёновые и
происходит из центральной тропической Америки. В нашей стране его выращивают
как однолетник. Стебель травянистый, но с возрастом древеснеет у основания, что
обеспечивает прямостоячую форму куста. Высота стебля 20 - 125 см, в зависимости
от сорта. Плод перца- многосемянная ягода с толщиной стенок от 1 до 9 мм, самой
разнообразной формы. Вес плодов от 50 до 350 гр. Окраска плодов в технической
спелости зелёная, салатная, почти белая или кремовая, а в биологической,-
жёлтая, оранжевая, красная. Период от всходов до начала сборов плодов 90 - 110
дней. Перец сладкий очень светолюбив, требователен к теплу и влаге.
Сорт "Ротунда" среднеспелый. Высота
стебля 50 - 75 см. Листья цельнокрайные, гладкие. Цветки белые, кремовые. Плоды
округло-сплющенные, слаборебристые. Мякоть нежная, очень толстая, сладкая.
Окраска плодов от жёлто-зелёной до красной. Средний вес плода 270 гр.
Полезные свойства Перца сладкого Capsicum
annum L.
В свежих плодах перца сладкого технической
спелости содержится 1 -1,5 % белка, до 1 % жира, 0,5 - 0,6 % минеральных
веществ, 1,4 % клетчатки, от 0,5 % до 4,5 мг % провитамина А, 0,06 мг %
витамина В1, 0,06 мг % витамина В2 и около 1,0 мг %
витамина РР. Суточная норма витамина С для взрослого человека может быть
обеспечена 25 гр, а витамина А - 45 гр плодов перца. Алкалоида капсаицина
содержится до 0,01 %, Эфирных масел, - 0,1 - 1,25 % от сухого вещества.
2.3 Биоморфологическая характеристика вида Перец
горький Capsicum
annum L.
(Сорт "Астраханский 147")
Перец горький также является представителем
семейства Паслёновые. Выращивается как однолетник. Стебель прямостоячий,
травянистый, одревесневший у основания. Высота стебля 20 - 125 см, в
зависимости от сорта. Плоды - стручки, ложная многосемянная ягода, состоящая из
мякоти, семяносцев и семян. Толщина мякоти от 1 до 5мм. Вес плода от 5 до 50
гр, в зависимости от сорта. Окраска зелёная, красная- зависит от сорта и
спелости.
Сорт "Астраханский 147" скороспелый,
от восходов до начала покраснения проходит 105 - 110 дней. Цветки белые. Плоды
красные, конусовидные, серповидно-изогнутые, гладкие, длиной 8 см, диаметром
2-3 см, весом 20 - 40 гр., очень острые. Сорт засухоустойчивый.
Полезные свойства Перца горького Capsicum
annum L.
В плодах перца горького содержится от 15 до 32 %
сухого вещества, сахаров примерно то же количество, что и в сладком перце, до
400 мг % Витамина С, до 5 мг % провитамина А, эфирные масла ( 0,7 - 2,5 %),
алкалоид капсаицин (0,02 - 1 %) Используется горький перец как приправа в
кулинарии, при солении, мариновании и консервировании. Применим в народной
медицине в качестве настоек, компрессов.
.4 Биоморфологическая характеристика вида
картофеля
Картофель сорт голландский. По длине
вегетационного периода сорта картофеля делится на ранние, среднеранние,
среднепоздние и поздние. Ранние сорта вегетируют 60-70 дней, среднеранние
70-80, среднеспелые 90-100, среднепоздние 110-130, поздние 150-180 дней.
Сорт голландский имеет, высокий прямостоячий
куст. Стебли толстые, с точечной пигментацией в нижней части. Листья
среднерассеченные. Цветки белые с красно- фиолетовой прожилкой. Цветение
обильное.
Сорт среднеспелый, высоко урожайный, устойчивый,
обычным расам фитофторы не поражается. Вкусовые качества высокие, крахмалистость
12,7-17,8%, легкость хорошая. Клубни крупные, белые, овальные, с неглубокими
глазками. В засушливые годы подвержен значительному вырождению. Районирован в
Алма-атинской и Северо-казахстанской областях.
.5 Ход эксперимента
Тексты для вербального и ментального
биоэнергоинформационного воздействия проводились по методике Г.Н. Сытина
(1993г). Основой данной методики являются следующие постулаты:
В текстах должны быть конкретные установки,-
оптимистичные для положительного эксперимента и пессимистичные для
отрицательного.
При воздействии необходим эмоциональный
компонент.
Тексты нужно произносить внятно, чётко,
уверенно, в состоянии сосредоточения на объекте воздействия.
• Текст положительного вербально и ментально
биоэнергоинформационного воздействия:
Я использую все свои духовные силы для того,
чтобы вы - баклажаны (перец горький, перец сладкий, картофель), выросли
большими, красивыми, полезными. Поток энергии света, вливаясь в вас, делает ваш
организм, каждую вашу клеточку всё более здоровыми, всё более крепкими. Весь
ваш организм постоянно, непрерывно поддерживает полную боевую готовность к
преодолению всех вредных влияний внешней среды. Не смотря ни на что, вы
продолжаете расти, здороветь и крепнуть. С каждым днём вы становитесь всё
крепче, ваше биополе становится мощнее. Весь ваш организм живёт и дышит легко,
свободно, полноценной жизнью. Я твёрдо верю, что всё сказанное мною обязательно
исполнится и вы, баклажаны (перец горький, перец сладкий, картофель), вырастите
большими, здоровыми, полезными.
• Текст отрицательного вербально и ментально
биоэнергоинформационного воздействия:
Я использую все свои духовные силы для того,
чтобы вы - баклажаны (перец горький, перец сладкий, картофель), прекратили свой
рост и развитие. Вселенная не поможет вам своей энергией, вы вырастете
маленькими и больными. Вы никому не нужны в этом мире, вы - бесполезные
создания. Природа не станет поддерживать вашу жизнь своими минеральными
элементами и энергией, вы не получите необходимых вам веществ для роста и
развития. Болезни измучают ваши организмы, ваши плоды будут маленькими и
чахлыми, а ваши семена не дадут проростков. Я твёрдо верю, что всё сказанное
мною обязательно исполнится и вы, баклажаны (перец горький, перец сладкий,
картофель), не сможете преодолеть вредных воздействий окружающей среды.
Для выполнения поставленных задач перечисленные
растения были рассажены в пять делянок по 25 растений каждого вида для трёх
экспериментов: положительного, отрицательного и контроля. Растения различных
экспериментов были пространственно разделены. Над положительно-
экспериментальными растениями с интервалом в один день читалась положительная
установка, над отрицательно-экспериментальными, - отрицательная. Контрольные
растения не подвергались воздействию вовсе. Схема высадки и интенсивности
воздействия приведена в Таблице 1.
Таблица 1 Сроки высадки и схема информационных
воздействий
|
Название
растения
|
Виды
воздействия
|
Сроки
высадки
|
Интенсивность
воздействия
|
|
Баклажаны
|
Положительное
ментально- биоэнергоинформационное воздействие
|
11.05.09г.
|
15
раз по 10 мин, с интервалом 1 день
|
|
Контроль
|
11.05.09г.
|
Не
проводилось
|
|
Отрицательное
ментально- биоэнергоинформационное воздействие
|
11.05.09г.
|
15
раз по 10 мин, с интервалом 1 день
|
|
Перец
сладкий
|
Положительное
ментально- биоэнергоинформационное воздействие
|
12.05.09г.
|
15
раз по 10 мин, с интервалом 1 день
|
|
Контроль
|
12.05.09г.
|
Не
проводилось
|
|
Отрицательное
ментально- биоэнергоинформационное воздействие
|
12.05.09г.
|
15
раз по 10 мин, с интервалом 1 день
|
|
Перец
горький
|
Положительное
ментально- биоэнергоинформационное воздействие
|
12.05.09г
|
15
раз по 10 мин, с интервалом 1 день
|
|
Контроль
|
12.05.09г
|
Не
проводилось
|
|
Отрицательное
ментально- биоэнергоинформационное воздействие
|
12.05.09г
|
15
раз по 10 мин, с интервалом 1 день
|
|
картофель
|
Положительное
вербально- биоэнергоинформационное воздействие
|
12.05.09г.
|
15раз
по 10 мин, с интервалом 1 день
|
|
Контроль
|
12.05.09г.
|
Не
проводилось
|
|
Отрицательное
вербально- биоэнергоинформационное воздействие
|
12.05.09г.
|
15
раз по 10 мин, с интервалом 1 день
|
Следует указать что грядки на которых
высаживались растения которые в последствии подвергались вербальному и
ментальному биоэнергоинформационому процессу, были пространственно разобщены,
расстояния между ними было не менее 25м.
Схема проведения замеров дана в Таблице 2.
Таблица 2 Схема
проведения замеров
|
Название
растения
|
Делянки
|
Замеры
вегетативных структур
|
Замеры
генеративных структур
|
|
Баклажаны
|
Положительное
ментально- биоэнергоинформационное воздействие
|
30.06.09г
30.07.09г 30.08.09г
|
18.06.09г.
|
|
Контроль
|
30.06.09г
30.07.09г 30.08.09г
|
18.06.09г.
|
|
Отрицательное
ментально- биоэнергоинформационное воздействие
|
30.06.09г
30.07.09г 30.08.09г
|
18.06.09г.
|
|
Перец
сладкий
|
Положительное
ментально- биоэнергоинформационное воздействие
|
30.06.09г
30.07.09г 30.08.09г
|
23.06.09г.
|
|
Контроль
|
30.06.09г
30.07.09г 30.08.09г
|
23.06.09г.
|
|
Отрицательное
ментально- биоэнергоинформационное воздействие
|
30.06.09г
30.07.09г 30.08.09г
|
23.06.09г.
|
|
Перец
горький
|
Положительное
ментально- биоэнергоинформационное воздействие
|
30.06.09г
30.07.09г 30.08.09г
|
23.06.09г.
|
|
Контроль
|
30.06.09г
30.07.09г 30.08.09г
|
23.06.09г.
|
|
Отрицательное
ментально- биоэнергоинформационное воздействие
|
30.06.09г
30.07.09г 30.08.09г
|
23.06.09г.
|
|
картофель
|
Положительное
вербально- биоэнергоинформационное воздействие
|
30.06.09г
30.07.09г 30.08.09г
|
18.06.09г.
|
|
Контроль
|
30.06.09г
30.07.09г 30.08.09г
|
18.06.09г.
|
|
Отрицательное
вербально- биоэнергоинформационное воздействие
|
30.06.09
30.07.09г 30.08.09г
|
18.06.09г.
|
.6 Методика обработки результатов
Биометрическая обработка вариационных рядов
произведена по общепризнанным методикам Удольской, 1976 и Сазоновой, 1986.
Для характеристики изменчивости количественного
признака определялись следующие показатели:
1. Средняя арифметическая, указывающая на
наиболее характерную для совокупности величину признака, определялась по
формуле:
X=A
+∑ af/n
где А - условное среднее,
а - отклонение вариаций от условной
средней
/ а = Х - А /,
f - частота
встречаемости признака,
n - число растений в
выборке,
λ - величина классового
промежутка.
. Среднее квадратическое отклонение находилось
по формуле
G=√∑af(∑(af)/n)n-1
. Ошибка средней арифметической определялась по
формуле
M=g/√n
. Коэффициент вариации определялся по формуле
C(V%)=(g/x)100%
. Критерий достоверности различий вычислялся по
формуле
Td=X-X/√m=m
Для характеристики уровня изменчивости признаков
использована классификация Мамаева, 1975, по которой можно распределить
изученные признаки по соответствующим уровням изменчивости:
Таблица 3
|
Уровень
изменчивости
|
Коэффициент
вариации (%)
|
|
Очень
низкий
|
До
7
|
|
Низкий
|
7
- 12
|
|
Средний
|
13
- 20
|
|
Высокий
|
21
- 40
|
|
Очень
высокий
|
Более
40
|
Биологическая продуктивность (биомасса,
созданная за весь период вегетации) и хозяйственная продуктивность (биомасса
полезной хозяйственной части урожая) определена по следующей методике:
. Для определения продуктивности берётся 10
растений;
. Растения выкапываются, разделяются на стебли с
корнями и листьями и плоды;
. Взвешивается сырая биомасса отдельно растений,
отдельно плодов.
3. Результаты исследований
.1 Анализ результатов направленного
биоэнергоинформационно ментального воздействия на параметры количественной
характеристики и амплитуды изменчивости отдельных признаков баклажан
Анализ фенотипической
изменчивости длины стебля
Таблица 4 Показатели количественной
характеристики и амплитуды изменчивости длины стебля при положительном
воздействии (см )
|
№
п/п
|
Замер
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
1
|
30.06.09
|
31,54
+ 0,33
|
34,80
- 28,60
|
1,66
|
5,26
|
Очень
низкий
|
|
2
|
30.07.09
|
41,52
+ 0,24
|
43,50
- 38,70
|
1,22
|
2,94
|
Очень
низкий
|
|
3
|
30.08.09
|
47,76
+ 0,28
|
50,20
- 44,80
|
1,40
|
2,93
|
Очень
низкий
|
Таблица 5 Показатели количественной
характеристики и амплитуды изменчивости длины стебля при отрицательном
воздействии (см )
|
№
п/пЗамерХ + mlim Xmax - XminGVУровень
изменчивости
|
|
|
|
|
|
|
|
1
|
30.06.09
|
29,69
+ 0,24
|
31,60
- 27,20
|
1,09
|
3,67
|
Очень
низкий
|
|
2
|
30.07.09
|
37,93
+ 0,30
|
41,60
- 35,20
|
1,44
|
3,80
|
Очень
низкий
|
|
3
|
30.08.09
|
43,65
+ 0,32
|
47,60
- 40,60
|
1,60
|
3,66
|
Очень
низкий
|
Таблица 6 Показатели количественной характеристики
и амплитуды изменчивости длины стебля у растений, не подвергавшихся воздействию
(см)
|
№
п/пЗамерХ + mlim Xmax - XminGVУровень
изменчивости
|
|
|
|
|
|
|
|
1
|
30.06.09
|
30,09
+ 0,28
|
32,50
- 27,30
|
1,39
|
4,62
|
Очень
низкий
|
|
2
|
30.07.09
|
39,48
+ 0,37
|
42,80
- 35,60
|
1,86
|
4,71
|
Очень
низкий
|
|
3
|
30.08.09
|
45,50
+ 0,30
|
48,80
- 42,40
|
1,50
|
3,30
|
Очень
низкий
|
Как видно из таблиц 4 - 6 в процессе онтогенеза
рост стебля и при отрицательном и при положительном воздействии, так же как и у
растений, не подвергавшихся воздействию, увеличился в среднем на 25 - 30%. Но
уже со второго замера видно, что отрицательное воздействие замедляет рост
стебля по сравнению, как с растениями положительного эксперимента, так и с
контрольными. Дальнейший анализ таблиц 4 -6 показал, что уровень изменчивости,
определяемый по числовому значению коэффициента вариации, определён нами как
очень низкий. Как известно из работ учёных, изучающих физиологию и биохимию
растений (Р. Ленинджер, 1976) низкий уровень изменчивости количественных
признаков появляется в том случае, если растения произрастают в благоприятных
условиях.
Таблица 7 Критерии достоверности различий длины
стебля у растений экспериментальных и контрольной делянок
|
Сравниваемые
делянки
|
td при измерении
|
|
Положит.
эксперимент - контроль
|
5,51
|
|
Отрицат.
эксперимент - контроль
|
4,21
|
|
Положит.
Экспер. - отрицат. экспер.
|
9,56
|
Р = 95 %. Таким образом, статистические расчёты
показывают достоверность различий длины стебля между экспериментальными и
контрольными растениями. Особенное значение имеет тот факт, что выявлена
достоверность различий длины стебля положительно обработанных растений и
контрольных.
Анализ фенотипической изменчивости ширины
листовой пластинки
Таблица 8 Показатели количественной
характеристики и амплитуды изменчивости ширины листовой пластинки при
положительном воздействии
Замер
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
1
|
30.06.09
|
5,63
+ 0,15
|
7,10
- 4,20
|
0,76
|
13,50
|
Средний
|
|
2
|
30.07.09
|
7,58
+ 0,16
|
9,20
- 5,80
|
0,79
|
10,42
|
Низкий
|
|
3
|
30.08.09
|
8,90
+ 0,14
|
10,30
- 7,40
|
0,69
|
7,75
|
Низкий
|
Таблица 9 Показатели количественной
характеристики и амплитуды изменчивости ширины листовой пластинки при
отрицательном воздействии
|
№
|
Замер
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
1
|
30.06.09
|
5,30
+ 0,15
|
6,70
- 3,80
|
0,73
|
13,77
|
Средний
|
|
2
|
30.07.09
|
6,92
+ 0,18
|
8,70
- 5,30
|
0,89
|
12,86
|
Средний
|
|
3
|
30.08.09
|
8,25
+ 0,14
|
9,60
- 6,70
|
0,69
|
8,36
|
Низкий
|
Таблица 10 Показатели количественной
характеристики и амплитуды изменчивости ширины листовой пластинки у растений,
не подвергавшихся воздействию (см)
|
№ЗамерХ
+ mlim
Xmax - XminGVУровень изменчивости
|
|
|
|
|
|
|
|
1
|
30.06.09
|
4,45
+ 0,20
|
6,70
- 2,80
|
0,98
|
22,02
|
Высокий
|
|
2
|
30.07.09
|
7,02
+ 0,20
|
8,70
- 4,80
|
0,99
|
14,10
|
Средний
|
|
3
|
30.08.09
|
8,01
+ 0,18
|
9,60
- 6,20
|
0,92
|
11,49
|
Низкий
|
Проведённые нами исследования по изучению
влияния ментально биоэнергоинформационного воздействия на количественную
характеристику показал, что ширина листовой пластинки от первого замера к
третьему увеличивается в среднем на 20 - 30%, как у экспериментальных, так и у
контрольных растений. Как видно из таблиц 8 - 10 уровень изменчивости
колеблется от среднего до низкого у экспериментальных групп растений, и от
высокого к низкому - у контрольных, что свидетельствует о снижении уровня
изменчивости посредством ментально биоэнергоинформационного воздействия.
Таблица 11 Критерии достоверности различий
ширины листовой пластинки у растений экспериментальных и контрольной делянок
|
Сравниваемые
делянки
|
td при измерении
|
|
Положит.
эксперимент - контроль
|
1,32
|
|
Отрицат.
эксперимент - контроль
|
2,75
|
|
Положит.
экспер. - отрицат. экспер.
|
3,25
|
Р = 95 %
Определённый нами критерий достоверности
различий количественных характеристик растений показал достоверность различий
ширины листовой пластинки между экспериментальными и контрольными растениями.
Анализ фенотипической изменчивости длины
листовой пластинки
Таблица 12 Показатели количественной
характеристики и амплитуды изменчивости длины листовой пластинки при
положительном воздействии
|
№
|
Замер
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
1
|
30.06.09
|
11,91
+ 0,18
|
13,60
- 10,20
|
0,89
|
7,47
|
Низкий
|
|
2
|
30.07.09
|
14,06
+ 0,16
|
15,70
- 12,30
|
0,81
|
5,76
|
Очень
низкий
|
|
3
|
30.08.09
|
15,60
+ 0,16
|
16,90
- 14,00
|
0,79
|
5,01
|
Очень
низкий
|
|
|
|
|
|
|
|
|
|
Таблица 13 Показатели количественной
характеристики и амплитуды изменчивости длины листовой пластинки при
отрицательном воздействии
|
№
|
Замер
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
1
|
30.06.09
|
11,45
+ 0,14
|
12,80
- 9,90
|
0,69
|
6,03
|
Очень
низкий
|
|
2
|
30.07.09
|
13,80
+ 0,14
|
15,20
- 12,30
|
0,69
|
5,00
|
Очень
низкий
|
|
3
|
30.08.09
|
15,04
+ 0,13
|
16,30
- 13,90
|
0,64
|
4,26
|
Очень
низкий
|
Таблица 14 Показатели количественной
характеристики и амплитуды изменчивости длины листовой пластинки у растений, не
подвергавшихся воздействию (см)
|
№ЗамерХ
+ mlim
Xmax - XminGVУровень изменчивости
|
|
|
|
|
|
|
|
1
|
30.06.09
|
11,79
+ 0,18
|
13,50
- 10,10
|
0,89
|
7,55
|
Низкий
|
|
2
|
30.07.09
|
13,80
+ 0,16
|
15,50
- 12,10
|
0,81
|
5,87
|
Очень
низкий
|
|
3
|
30.08.09
|
15,18
+ 0,12
|
16,40
- 14,00
|
0,61
|
4,02
|
Очень
низкий
|
Сравнительное изучение длины листовой пластинки
показало, что как у экспериментальных, так и у контрольных растений в процессе
онтогенеза происходит статистически выявляемое увеличение количественной
характеристики данного признака.
Исходя из данных Таблиц 12 - 14 видно, что длина
листовой пластинки, как у экспериментальных, так и у контрольных растений по
уровню изменчивости варьирует от низкого к очень низкому, т. е. можно сказать,
что ментальное биоэнергоиформационное воздействие оказывает заметное влияние на
уровень изменчивости данного признака.
Таблица 15 Критерии достоверности различий длины
листовой пластинки у растений экспериментальных и контрольной делянок
|
Сравниваемые
делянки
|
td при измерении
|
|
Положит.
эксперимент - контроль
|
2,10
|
|
Отрицат.
эксперимент - контроль
|
0,78
|
|
Положит.
экспер. - отрицат. экспер.
|
2,67
|
Р = 95 %
Данные таблицы 15 свидетельствуют о том, что
ментальное биоэнергоинформационное воздействие водой оказывает статистически
достоверное влияние на рост листовой пластинки, особенно это наблюдается при
сравнении положительно и отрицательно ментально биоэнергоинформационно обработанных
растений.
Анализ фенотипической изменчивости
количественной характеристики числа цветков и плодов
Таблица 16 Показатели количественной
характеристики и амплитуды изменчивости количества цветков (см)
|
№
|
Признак
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
1
|
"
+ "
|
12,36
+ 0,35
|
15
- 9
|
1,75
|
14,16
|
Средний
|
|
2
|
"
- "
|
7,16
+ 0,32
|
10
- 4
|
1,60
|
22,35
|
Высокий
|
|
3
|
Контроль
|
9,08
+ 0,36
|
13
- 6
|
1,78
|
19,60
|
Средний
|
" + " - растения, обработанные
положительно биоэнергоинформационно ментальным воздействием
"-" - растения, обработанные
отрицательно биоэнергоинформационно ментальным воздействием
Наши исследования показали, что количество
цветков у растений, подвергшихся положительному ментально
биоэнергоинформационному воздействию почти на 15% больше, чем у контрольных, а
по сравнению с растениями, подвергшихся отрицательному ментально
биоэнергоинформационному воздействию водой, это число составляет 50%.
Также из таблицы 16 видно, что отрицательное
ментально биоэнергоинформационное воздействие водой вызывает увеличение уровня
изменчивости.
Таблица 17 Критерии достоверности различий
количества цветков у растений экспериментальных и контрольной делянок
|
Сравниваемые
делянки
|
td при измерении
|
|
Положит.
эксперимент - контроль
|
6,56
|
|
Отрицат.
эксперимент - контроль
|
4,00
|
|
Положит.
Экспер. - отрицат. экспер.
|
10,40
|
Р = 95 %
Данные по определению критерия достоверности
(Таблица 17) свидетельствуют, что различия по числу цветков между
экспериментальными и контрольными растениями достоверны.
Это указывает на то , что ментальное биоэнергоинфрмационное
воздействие обладает способностью увеличения закладки и дальнейшего в процессе
онтогенетического развития роста почечных структур цветкам, которые позже
станут лепестками, тычинками и пестикам.
Таблица 18 Показатели количественной характеристики
и амплитуды изменчивости количества плодов (см)
|
№
п/п
|
Признак
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
1
|
"
+ "
|
6,36
+ 0,30
|
9
- 4
|
1,50
|
23,58
|
Высокий
|
|
2
|
"
- "
|
4,52
+ 0,24
|
7
- 2
|
1,19
|
26,32
|
Высокий
|
|
3
|
Контроль
|
5,40
+ 0,28
|
8
- 3
|
1,38
|
25,55
|
Высокий
|
" + " - растения, обработанные
положительно биоэнергоинформационно ментальным воздействием
" - " - растения, обработанные
отрицательно биоэнергоинформационно ментальным воздействием
Анализ таблицы 18 показывает, что
положительно-экспериментальные растения почти на 15% превышают показатели
контрольных, а по сравнению с отрицательно-экспериментальными растениями этот
процент удваивается. Уровень изменчивости отрицательно экспериментальных
растений выше, чем у положительно-экспериментальных и контрольных.
Таблица 19 Критерии достоверности различий
количества плодов у растений экспериментальных и контрольной делянок
|
Сравниваемые
делянки
|
td при измерении
|
|
Положит.
эксперимент - контроль
|
2,34
|
|
Отрицат.
эксперимент - контроль
|
2,26
|
|
Положит.
экспер. - отрицат. экспер.
|
4,72
|
Р = 95 %
Статистические расчёты таблицы 19 показывают
достоверность различий количества плодов между экспериментальными и
контрольными растениями.
3.2 Показатели количественной характеристики и
амплитуды изменчивости отдельных признаков перца горького
Анализ фенотипической изменчивости длины стебля
Таблица 20 Показатели количественной
характеристики и амплитуды изменчивости длины стебля при положительном
воздействии (см)
|
№
|
Замер
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
1
|
30.06.09
|
33,89
+ 0,32
|
37,30
- 30,90
|
1,62
|
4,78
|
Очень
низкий
|
|
2
|
30.07.09
|
43,68
+ 0,38
|
47,20
- 39,80
|
1,90
|
4,35
|
Очень
низкий
|
|
3
|
30.08.09
|
49,58
+ 0,38
|
53,10
- 45,70
|
1,91
|
3,85
|
Очень
низкий
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Таблица 21 Показатели количественной
характеристики и амплитуды изменчивости длины стебля при отрицательном
воздействии (см)
|
№
|
Замер
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
1
|
30.06.09
|
30,86
+ 0,36
|
34,60
- 27,70
|
1,82
|
5,90
|
Очень
низкий
|
|
2
|
30.07.09
|
39,17
+ 0,43
|
43,30
- 34,90
|
2,16
|
5,51
|
Очень
низкий
|
|
5
|
30.08.09
|
40,95
+ 0,43
|
45,90
- 37,50
|
2,14
|
5,23
|
Очень
низкий
|
Таблица 22 Показатели количественной
характеристики и амплитуды изменчивости длины стебля у растений, не
подвергавшихся воздействию (см)
|
№ЗамерХ
+ mlim
Xmax - XminGVУровень изменчивости
|
|
|
|
|
|
|
|
1
|
30.06.09
|
34,21
+ 0,34
|
37,10
- 30,70
|
1,69
|
4,94
|
Очень
низкий
|
|
2
|
30.07.09
|
41,78
+ 0,38
|
45,30
- 37,90
|
1,90
|
4,55
|
|
3
|
30.08.09
|
45,66
+ 0,38
|
49,30
- 41,90
|
1,91
|
4,18
|
Очень
низкий
|
Сравнивая пределы вариабельности длины стебля мы
обнаружили, что количественная характеристика данного признака от первого
замера к третьему увеличивается в среднем на 25 - 30% у всех групп растений.
Дальнейший анализ таблиц 20 - 22 показывает, что
уровень изменчивости по числовому значению коэффициента вариации определён нами
как очень низкий, следовательно, ментально биоэнергоинформационное воздействие
практически не влияет на уровень изменчивости.
Таблица 23 Критерии достоверности различий длины
стебля у растений экспериментальных и контрольной делянок
|
Сравниваемые
делянки
|
td при измерении
|
|
Положит.
эксперимент - контроль
|
7,26
|
|
Отрицат.
эксперимент - контроль
|
8,26
|
|
Положит.
экспер. - отрицат. экспер.
|
15,14
|
Р = 95 %
Статистические расчёты таблицы 23 показывают
достоверность различий длины стебля между экспериментальными и контрольными
растениями.
Анализ фенотипической изменчивости ширины
листовой пластинки
Таблица 24 Показатели количественной
характеристики и амплитуды изменчивости ширины листовой пластинки при
положительном воздействии
|
№
|
Замер
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
1
|
30.06.09
|
3,96
+ 0,13
|
5,10
- 2,70
|
0,63
|
15,91
|
Средний
|
|
2
|
30.07.09
|
4,83
+ 0,11
|
5,70
- 3,80
|
0,55
|
11,39
|
Низкий
|
|
3
|
30.08.09
|
5,64
+ 0,14
|
6,90
- 4,50
|
0,69
|
12,32
|
Низкий
|
Таблица 25 Показатели количественной
характеристики и амплитуды изменчивости ширины листовой пластинки при
отрицательном воздействии
|
№ЗамерХ
+ mlim
Xmax - XminGVУровень изменчивости
|
|
|
|
|
|
|
|
1
|
30.06.09
|
2,72
+ 0,13
|
3,90
- 1,50
|
0,65
|
23,90
|
Высокий
|
|
2
|
30.07.09
|
3,75
+ 0,12
|
4,70
- 2,80
|
0,51
|
13,60
|
Средний
|
|
3
|
30.08.09
|
4,58
+ 0,15
|
6,10
- 3,20
|
0,77
|
16,81
|
Средний
|
Таблица 26 Показатели количественной
характеристики и амплитуды изменчивости ширины листовой пластинки у растений,
не подвергавшихся воздействию (см)
|
№ЗамерХ
+ mlim
Xmax - XminGVУровень изменчивости
|
|
|
|
|
|
|
|
1
|
30.06.09
|
3,15
+ 0,10
|
4,10
- 2,20
|
0,51
|
16,19
|
Средний
|
|
2
|
30.07.09
|
4,38
+ 0,12
|
5,50
- 3,10
|
0,61
|
13,93
|
Средний
|
|
3
|
30.08.09
|
4,87
+ 0,15
|
6,20
- 3,30
|
0,76
|
15,61
|
Средний
|
Проведённые нами исследования по изучению
влияния биоэнергоинформационного воздействия на количественную характеристику
показал, что ширина листовой пластинки от первого замера к третьему
увеличивается в среднем на 15 - 25%, как у экспериментальных, так и у
контрольных растений. Как видно из таблиц 24 - 26 уровень изменчивости
колеблется от высокого до низкого, что свидетельствует о снижении уровня
изменчивости посредством биоэнергоинформационного воздействия.
Таблица 27 Критерии достоверности различий
ширины листовой пластинки у растений экспериментальных и контрольной делянок
|
Сравниваемые
делянки
|
td при измерении
|
|
Положит.
эксперимент - контроль
|
3,67
|
|
Отрицат.
эксперимент - контроль
|
1,38
|
|
Положит.
экспер. - отрицат. экспер.
|
5,05
|
Р = 95 %
Результаты сравнения ширины листовой пластинки
(Таблица 27) показывают статистически достоверные различия по данному признаку
между экспериментальными и контрольными растениями.
Анализ фенотипической изменчивости длины
листовой пластинки
Таблица 28 Показатели количественной
характеристики и амплитуды изменчивости длины листовой пластинки при
положительном воздействии
|
№
|
Замер
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
1
|
30.06.09
|
5,44
+ 0,16
|
6,70
- 3,80
|
0,78
|
14,34
|
Средний
|
|
2
|
30.07.09
|
6,50
+ 0, 14
|
8,10
- 5,20
|
0,72
|
11,08
|
Низкий
|
|
3
|
30.08.09
|
8,01
+ 0,18
|
9,60
- 6,20
|
0,92
|
11,49
|
Низкий
|
Таблица 29 Показатели количественной
характеристики и амплитуды изменчивости длины листовой пластинки при
отрицательном воздействии
|
№
|
Замер
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
1
|
30.06.09
|
4,23
+ 0,15
|
5,70
- 2,80
|
0,76
|
17,97
|
Средний
|
|
2
|
30.07.09
|
5,73
+ 0,18
|
7,60
- 4,20
|
0,91
|
15,88
|
Средний
|
|
3
|
30.08.09
|
7,53
+ 0,18
|
9,20
- 5,80
|
0,91
|
12,08
|
Низкий
|
Таблица 30 Показатели количественной
характеристики и амплитуды изменчивости длины листовой пластинки у растений, не
подвергавшихся воздействию (см)
|
№ЗамерХ
+ mlim
Xmax - XminGVУровень изменчивости
|
|
|
|
|
|
|
|
1
|
30.06.09
|
4,90
+ 0,14
|
6,20
- 3,30
|
0,73
|
14,90
|
Средний
|
|
2
|
30.07.09
|
6,58
+ 0,17
|
8,20
- 4,80
|
0,89
|
13,53
|
Средний
|
|
3
|
30.08.09
|
7,62
+ 0,17
|
9,40
- 6,00
|
0,89
|
11,68
|
Низкий
|
Проведённые нами исследования показали, что в
процессе онтогенеза ширина листовой пластинки у растений положительного
эксперимента и контрольных увеличилась в среднем на 21 - 30. Из данных Таблиц
28 - 30 видно, что изучаемый признак, как у экспериментальных, так и у
контрольной групп растений уровень изменчивости уменьшается от среднего к
низкому, что доказывает наличие ментального биоэнергоинформационного
воздействия.
Таблица 31 Критерии достоверности различий длины
листовой пластинки у растений экспериментальных и контрольной делянок
|
Сравниваемые
делянки
|
td при измерении
|
|
Положит.
эксперимент - контроль
|
1,12
|
|
Отрицат.
эксперимент - контроль
|
0,36
|
|
Положит.
экспер. - отрицат. экспер.
|
1,42
|
Р = 95%
Статистические расчёты, приведенные в таблице 31
показывают, что различия по длине листовой пластинки между экспериментальными и
контрольными группами растений не достоверны.
Анализ фенотипической изменчивости
количественной характеристики числа цветков и плодов
Таблица 32 Показатели количественной
характеристики и амплитуды изменчивости количества цветков (см)
|
№
|
Признак
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
1
|
"
+ "
|
20,24
+ 0,32
|
23
- 17
|
1,62
|
8,00
|
Низкий
|
|
2
|
"
- "
|
14,40
+ 0,39
|
19
- 12
|
1,94
|
13,47
|
Средний
|
|
3
|
Контроль
|
15,68
+ 0,48
|
21
- 13
|
2,38
|
15,18
|
Средний
|
" + " - растения, обработанные
положительно биоэнергоинформационно ментальным воздействием
" - " - растения, обработанные
отрицательно биоэнергоинформационно ментальным воздействием
Согласно данным таблицы 32, количество цветков у
растений подвергшихся положительному воздействию на 30% превышает контрольный
показатель, а по сравнению с растениями, подвергшимися отрицательному
воздействию, более чем на 40%.
Ментальное биоэнергоинформационно положительное
и отрицательное воздействие вызвало понижение уровня изменчивости.
Таблица 33 Критерии достоверности различий
количества цветков у растений экспериментальных и контрольной делянок
|
Сравниваемые
делянки
|
td при измерении
|
|
Положит.
эксперимент - контроль
|
7,86
|
|
Отрицат.
эксперимент - контроль
|
2,06
|
|
Положит.
экспер. - отрицат. экспер.
|
11,68
|
Р = 95 %
В таблице 33 приведены данные по определению
критерия достоверности, которые свидетельствуют, что между
отрицательно-экспериментальными и контрольными растениями, а также между
положительно-экспериментальными и отрицательно-экспериментальными растениями
существуют достоверные различия.
Таблица 34 Показатели количественной
характеристики и амплитуды изменчивости количества плодов (см)
|
№
п/п
|
Признак
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
1
|
"
+ "
|
9,48
+ 0,28
|
12
- 7
|
1,38
|
14,56
|
Средний
|
|
2
|
"
- "
|
6,52
+ 0,26
|
9
- 4
|
1,32
|
20,25
|
Высокий
|
|
3
|
Контроль
|
7,32
+ 0,24
|
10
- 5
|
1,22
|
16,67
|
Средний
|
" + " - растения, обработанные
положительно биоэнергоинформационно ментальным воздействием
" - " - растения, обработанные
отрицательно биоэнергоинформационно ментальным воздействием
По результатам исследований, приведенным в
таблице 34, можно сделать выводы, что число цветков у растений, подвергавшихся
положительному ментально биоэнергоинформационному воздействию, на 20% выше, чем
у контрольных, и на 33% выше, чем у отрицательно-экспериментальных групп
растений.
Уровень изменчивости данного признака
показывает, что у положительно-экспериментальных и контрольных групп растений
он средний, а у отрицательно-экспериментальных - высокий, что позволяет
говорить о способности положительного ментально биоэнергоинформационного
воздействия снижать уровень изменчивости.
Таблица 35 Критерии достоверности различий
количества плодов у растений экспериментальных и контрольной делянок
|
Сравниваемые
делянки
|
td при измерении
|
|
Положит.
эксперимент - контроль
|
5,84
|
|
Отрицат.
эксперимент - контроль
|
2,29
|
|
Положит.
экспер. - отрицат. экспер.
|
7,79
|
Р = 95 %
Статистические расчёты таблицы 35 показывают
достоверность различий количества плодов между экспериментальными и
контрольными растениями.
.3 Показатели количественной характеристики и
амплитуды изменчивости отдельных признаков перца сладкого
Анализ фенотипической изменчивости длины стебля
Таблица 36 Показатели количественной
характеристики и амплитуды изменчивости длины стебля при положительном
воздействии (см)
|
№
п/п
|
Замер
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
1
|
30.06.09
|
37,14
+ 0,32
|
39,80
- 33,90
|
1,60
|
4,31
|
Очень
низкий
|
|
2
|
30.07.09
|
48,50
+ 0,34
|
51,70
- 45,30
|
1,72
|
3,55
|
Очень
низкий
|
|
3
|
30.08.09
|
50,69
+ 0,30
|
53,30
- 47,90
|
1,49
|
2,94
|
Очень
низкий
|
Таблица 37 Показатели количественной
характеристики и амплитуды изменчивости длины стебля при отрицательном
воздействии (см)
|
№
|
Замер
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
1
|
30.06.09
|
35,77
+ 0,28
|
38,60
- 33,10
|
1,40
|
3,91
|
Очень
низкий
|
|
2
|
30.07.09
|
46,40
+ 0,33
|
43,50
- 49,40
|
1,66
|
3,58
|
Очень
низкий
|
|
3
|
30.08.09
|
48,31
+ 0,30
|
51,40
- 45,50
|
1,50
|
3,10
|
Таблица 38 Показатели количественной
характеристики и амплитуды изменчивости длины стебля у растений, не
подвергавшихся воздействию (см)
|
№ЗамерХ
+ mlim
Xmax - XminGVУровень изменчивости
|
|
|
|
|
|
|
|
1
|
30.06.09
|
36,76
+ 0,25
|
39,00
- 34,60
|
1,23
|
3,35
|
Очень
низкий
|
|
2
|
30.07.09
|
47,50
+ 0,34
|
50,60
- 44,20
|
1,72
|
3,62
|
Очень
низкий
|
|
3
|
30.08.09
|
49,56
+ 0,33
|
52,60
- 46,70
|
1,64
|
3,31
|
Очень
низкий
|
В результате наших исследований мы получили
следующие данные (таблица 36 - 38), которые говорят, что на более ранних
стадиях онтогенеза рост стебля, как у экспериментальных, так и у контрольных,
т.е. не подвергавшихся воздействию растений, увеличился в среднем на 30%. Но
уже второй замер позволил выявить, что отрицательное воздействие замедляет рост
стебля по сравнению и с положительно-экспериментальной и с контрольной группами
растений.
Дальнейший анализ данных таблиц 36 - 38 показал,
что уровень изменчивости изучаемого признака, как у контрольных, так и у
экспериментальных растений очень низкий, следовательно, ментально
биоэнергоинформационное воздействие на уровень изменчивости ощутимого влияния
не оказывает
Таблица 39 Критерии достоверности различий длины
стебля у растений экспериментальных и контрольных делянок
|
Сравниваемые
делянки
|
td при измерении
|
|
Положит.
эксперимент - контроль
|
2,51
|
|
Отрицат.
эксперимент - контроль
|
2,78
|
|
Положит.
экспер. - отрицат. экспер.
|
5,67
|
Р = 95 %
Определённый нами критерий достоверности
различий количественных характеристик экспериментальных и контрольных растений
(таблица 39) показывает достоверность различий по данному признаку.
Анализ фенотипической изменчивости ширины
листовой пластинки
Таблица 40 Показатели количественной
характеристики и амплитуды изменчивости ширины листовой пластинки при положительном
воздействии
|
№
|
Замер
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
1
|
30.06.09
|
4,42
+ 0,11
|
5,50
- 3,10
|
0,56
|
12,70
|
Низкий
|
|
2
|
30.07.09
|
5,13
+ 0,15
|
6,70
- 3,80
|
0,76
|
14,81
|
Средний
|
|
3
|
30.08.09
|
7,10
+ 0,16
|
8,80
- 5,40
|
0,81
|
11,41
|
Низкий
|
Таблица 41 Показатели количественной
характеристики и амплитуды изменчивости ширины листовой пластинки при
отрицательном воздействии
|
№ЗамерХ
+ mlim
Xmax - XminGVУровень изменчивости
|
|
|
|
|
|
|
|
1
|
30.06.09
|
3,50
+ 0,14
|
4,80
- 1,90
|
0,69
|
19,71
|
Средний
|
|
2
|
30.07.09
|
5,15
+ 0,14
|
6,60
- 3,70
|
0,69
|
13,40
|
Средний
|
|
3
|
30.08.09
|
5,83
+ 0,18
|
7,50
- 4,10
|
0,89
|
15,27
|
Низкий
|
Таблица 42 Показатели количественной
характеристики и амплитуды изменчивости ширины листовой пластинки у растений,
не подвергавшихся воздействию (см)
|
№ЗамерХ
+ mlim
Xmax - XminGVУровень изменчивости
|
|
|
|
|
|
|
|
1
|
30.06.09
|
3,82
+ 0,17
|
5,40
- 2,00
|
0,87
|
22,87
|
Высокий
|
|
2
|
30.07.09
|
4,29
+ 0,17
|
6,30
- 2,90
|
0,87
|
20,28
|
Средний
|
|
3
|
30.08.09
|
5,86
+ 0,13
|
7,00
- 4,60
|
0,63
|
10,75
|
Низкий
|
Как видно из Таблиц 40 - 42 исследование
количественной характеристики ширины листовой пластинки показало, что, в
процессе онтогенеза данный признак, при последнем замере, у
отрицательно-экспериментальных и у контрольных растений практически не
отличается, а у положительно-экспериментальных растений превышает их показатель
более чем на 20%, что говорит о способности положительного ментально
биоэнергоинформационного воздействия влиять на ростовые процессы.
Таблица 43 Критерии достоверности различий
ширины листовой пластинки у растений экспериментальных и контрольной делянок
|
Сравниваемые
делянки
|
td при измерении
|
|
Положит.
эксперимент - контроль
|
7,75
|
|
Отрицат.
эксперимент - контроль
|
0,14
|
|
Положит.
экспер. - отрицат. экспер.
|
5,29
|
Р = 95 %
Статистические расчеты по определению критерия
достоверности, приведённые в Таблице 43, показывают, что между
положительно-экспериментальными и контрольными, а так же
положительно-экспериментальными и отрицательно-экспериментальными группами
растений существует достоверность различий.
Анализ фенотипической изменчивости длины листовой
пластинки
Таблица 44 Показатели количественной
характеристики и амплитуды изменчивости длины листовой пластинки при
положительном воздействии
|
№
|
Замер
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
1
|
30.06.09
|
7,54
+ 0,22
|
9,60
- 5,20
|
1,09
|
14,46
|
Средний
|
|
2
|
30.07.09
|
9,82
+ 0,18
|
11,60
- 8,20
|
0,89
|
9,06
|
Низкий
|
|
3
|
30.08.09
|
11,46
+ 0,16
|
13,10
-9,70
|
0,81
|
7,07
|
Низкий
|
Таблица 45 Показатели количественной
характеристики и амплитуды изменчивости длины листовой пластинки при
отрицательном воздействии
|
№ЗамерХ
+ mlim
Xmax - XminGVУровень изменчивости
|
|
|
|
|
|
|
|
1
|
30.06.09
|
6,67
+ 0,19
|
8,60
- 4,70
|
0,97
|
14,54
|
Средний
|
|
2
|
30.07.09
|
8,38
+ 0,17
|
10,00
- 6,60
|
0,84
|
10,02
|
Низкий
|
|
3
|
30.08.09
|
10,39
+ 0,21
|
12,10
- 8,20
|
1,06
|
10,20
|
Низкий
|
Таблица 46 Показатели количественной характеристики
и амплитуды изменчивости длины листовой пластинки у растений, не подвергавшихся
воздействию (см)
|
№ЗамерХ
+ mlim
Xmax - XminGVУровень изменчивости
|
|
|
|
|
|
|
|
1
|
30.06.09
|
7,08
+ 0,22
|
9,10
- 5,20
|
0,98
|
13,84
|
Средний
|
|
2
|
30.07.09
|
8,94
+ 0,18
|
10,70
- 7,20
|
0,81
|
9,06
|
Низкий
|
|
3
|
30.08.09
|
10,66
+ 0,22
|
12,60
- 8,70
|
0,98
|
9,19
|
Низкий
|
Сравнительное изучение длины листовой пластинки
(Таблицы 44-46) показало, что как у экспериментальных, так и у контрольных
растений в процессе онтогенеза происходит статистически выявляемое увеличение
количественной характеристики данного признака, в среднем эта цифра составляет
25 - 30 %. Исходя из табличных данных видно, что длина листовой пластинки, как
у экспериментальных, так и у контрольных растений, по уровню изменчивости, варьирует
от среднего к низкому, т. е. можно сказать, что ментальное
биоэнергоинформационное воздействие оказывает заметное влияние на уровень
изменчивости данного признака.
Таблица 47 Критерии достоверности различий длины
листовой пластинки у растений экспериментальных и контрольной делянок
|
Сравниваемые
делянки
|
td при измерении
|
|
Положит.
эксперимент - контроль
|
3,87
|
|
Отрицат.
эксперимент - контроль
|
0,67
|
|
Положит.
экспер. - отрицат. экспер.
|
4,12
|
Р = 95 %
Так же нами был определён критерий достоверности
различий количественных характеристик экспериментальных и контрольных растений.
Как свидетельствуют данные Таблицы 47, различия по числу плодов между ментально
отрицательно-экспериментальными и контрольными группами растений не достоверны.
Анализ фенотипической изменчивости количественной характеристики числа цветков
и плодов
Таблица 48 Показатели количественной
характеристики и амплитуды изменчивости количества цветков (см)
|
№
|
Признак
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
1
|
"
+ "
|
18,44
+ 0,40
|
22
- 14
|
2,02
|
10,95
|
Низкий
|
|
2
|
"
- "
|
12,76+
0,49
|
17
- 8
|
2,44
|
19,12
|
Средний
|
|
3
|
Контроль
|
15,60
+ 0,44
|
19
- 12
|
2,22
|
14,23
|
Средний
|
" + " - растения, обработанные
положительно биоэнергоинформационно ментальным воздействием
" - " - растения, обработанные
отрицательно биоэнергоинформационно ментальным воздействием
Статистические расчеты числа цветков в Таблице
48 показывают, что число цветков у растений, подвергавшихся ментально
положительному биоэнергоинформационному воздействию, на 15% выше чем у
контрольных, а по сравнению с растениями, подвергшимися отрицательному
ментально-биоэнергоинформационному воздействию, это число в два раза больше.
Таблица 49 Критерии достоверности различий
количества цветков у растений экспериментальных и контрольной делянок
|
Сравниваемые
делянки
|
td при измерении
|
|
Положит.
эксперимент - контроль
|
4,81
|
|
Отрицат.
эксперимент - контроль
|
4,30
|
|
Положит.
экспер. - отрицат. экспер.
|
9,02
|
Р = 95 %
Как видно из Таблицы 49, статистические расчеты
показывают достоверность различий количества цветков между экспериментальными и
контрольными группами растений. Особенное значение имеет тот факт, что выявлена
достоверность различий количества цветков у экспериментальных растений.
Таблица 50 Показатели количественной
характеристики и амплитуды изменчивости количества плодов (см)
|
№
п/п
|
Признак
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
1
|
"
+ "
|
8,56
+ 0,26
|
11
- 6
|
1,32
|
15,42
|
Средний
|
|
2
|
"
- "
|
5,44
+ 0,26
|
8
- 3
|
1,32
|
24,32
|
Высокий
|
|
3
|
Контроль
|
6,64
+ 0,26
|
9
- 4
|
1,32
|
19,88
|
Средний
|
" + " - растения, обработанные положительно
биоэнергоинформационно ментальным воздействием
" - " - растения, обработанные
отрицательно биоэнергоинформационно ментальным воздействием
Анализ Таблицы 50 показывает, что
положительно-экспериментальные растения более чем на 25% превышают показатели
контрольных, а по сравнению с отрицательно-экспериментальными растениями этот
процент удваивается. Уровень изменчивости положительно-экспериментальных и
контрольных растений ниже, чем у отрицательно-экспериментальных, что
свидетельствует о возможности снижать уровень изменчивости посредством
положительного ментально-биоэнергоинформационного воздействия.
Таблица 51 Критерии достоверности различий
количества плодов у растений экспериментальных и контрольной делянок
|
Сравниваемые
делянки
|
td при измерении
|
|
Положит.
эксперимент - контроль
|
5,19
|
|
Отрицат.
эксперимент - контроль
|
3,24
|
|
Положит.
экспер. - отрицат. экспер.
|
8,43
|
Р = 95%
Как показывает Таблица 51, различия количества
плодов между экспериментальными и контрольными группами растений достоверны.
Важно отметить факт достоверности различий между положительно- и
отрицательно-экспериментальными группами растений.
.4 Анализ результатов направленного
информационно- вербального на параметры количественной характеристики и
амплитуды изменчивости отдельных признаков у картофеля
Анализ фенотипической изменчивости величины
продольного диаметра клубня
Проведенные нами исследования показали что
информационно- вербальное положительное так и информационно- вербальное
отрицательное воздействия уменьшает X
max - X
min величину или
амплитуду изменчивости продольного диаметра клубней испытуемых растений,
(таблица 52).
При этом обнаруживаются, что уровень
изменчивости у растений подвергшихся положительному биоэнергоинформационому
воздействию низким или очень низким, тогда как у контрольных растений уровень
изменчивости также низкий( I
и III замеры), а во II
замере мы обнаружили, что уровень изменчивости средний. Эти данные говорят о
том, что количество клубней должно быть больше.
Таблица 52 Показатели количественной характеристики
продольного диаметра клубней
|
Делянки
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
I замер
|
|
|
|
|
|
|
Эксперимент
положительный
|
27 + 0,9
|
29-25
|
1,8
|
6,6
|
Очень
низкий
|
|
Контроль
|
23 + 1,3
|
28-18
|
2,7
|
11,7
|
Низкий
|
|
Эксперимент
отрицательный
|
20 + 0,9
|
22-18
|
1,8
|
9
|
Низкий
|
|
II замер
|
|
|
|
|
|
|
Эксперимент
положительный
|
27,2
+
0,7
|
29-26
|
1,5
|
5,5
|
Очень
низкий
|
|
Контроль
|
22,2
+
1,5
|
25-18
|
3,08
|
13,8
|
Средний
|
|
Эксперимент
отрицательный
|
20,3 + 0,7
|
19-2
|
1,2
|
5,9
|
Очень
низкий
|
|
III замер
|
|
|
|
|
|
|
Эксперимент
положительный
|
27,2
+ 1,1
|
2,3
|
8,4
|
Низкий
|
|
Контроль
|
25
+ 1,2
|
27-23
|
2
|
8
|
Низкий
|
|
Эксперимент
отрицательный
|
19,7
+ 0,7
|
21-18
|
5
|
7,6
|
Низкий
|
Следует отметить что между растениями, которые
подвергались информационно- вербальному воздействию и растениями ему не
подвергавшиеся выявлена статистически достоверная разница, эти данные
представлены в таблицы 53.
Таблица 53 Критерии достоверности различий
продольного диаметра клубня экспериментально положительной и контрольной группы
|
Сравниваемые
делянки
|
td при измерениях
|
|
Эксперимент
положительный- контроль
|
I
|
II
|
III
|
|
3,6
|
3,1
|
1,4
|
Р=95
Анализ таблицы 53 показывает, что при I
и II замерах
продольного диаметра клубня выявлены статистические достоверные различия между
растениями, подвергавшимися информационно- вербально положительному воздействию
и не подвергавшиеся ему. Это позволяет сделать вывод, о том, что информационно-
вербально положительное воздействие стимулирует увеличение продольного диаметра
клубня.
Таблица 54 Критерии достоверности различий
продольного диаметра клубня экспериментально отрицательной и контрольной группы
|
Сравниваемые
делянки
|
Td при измерениях
|
|
Эксперимент
отрицательный- контроль
|
I
|
II
|
III
|
|
1,8
|
1,1
|
4
|
Р=95%
Анализ таблицы 54 показывает, что при I
и II замерах
продольного диаметра клубня не выявлены статистически достоверные различия
между растениями, подвергавшиеся информационно- вербально отрицательному
воздействию. Статистически достоверные различия между растениями выявлены
только при III замере
продольного диаметра клубня. Это позволяет сделать вывод о том, что
информационно- вербально отрицательное воздействие не значительно стимулирует
уменьшение продольного диаметра клубня.
Анализ фенотипической изменчивости величины
поперечного диаметра клубня
Изучение поперечного диаметра клубня показало,
что информационно- вербальное воздействие в значительной степени не влияет на X
max - X
min величину или
амплитуду изменчивости, так как у экспериментально положительных растений:
I замер 4 см, II
замер 2 см, III замер 5см.
У экспериментально отрицательных растений:
I замер 1см, II
замер 2см, III замер 1см.
У контрольных растений:
I замер 5см, II
замер 5см, III замер 2см,
(таблица 60)
Таблица 55 Показатели количественной
характеристики продольного диаметра клубней
|
Делянки
|
Х
+ m
|
lim Xmax - Xmin
|
G
|
V
|
Уровень
изменчивости
|
|
|
I замер
|
|
|
|
|
|
|
|
Эксперимент положительный
|
23,7
+
0,9
|
25-21
|
1,8
|
7,6
|
Низкий
|
|
Контроль
|
19,5
+ 1,1
|
21-16
|
2,3
|
11,7
|
Низкий
|
|
Эксперимент
отрицательный
|
17,5+
0,25
|
18-17
|
0,5
|
2,8
|
Очень
низкий
|
|
II замер
|
|
|
|
|
|
|
Эксперимент положительный
|
23,7 + 0,5
|
25-23
|
1
|
4,2
|
Очень
низкий
|
|
Контроль
|
20,5 + 1,3
|
23-18
|
2,6
|
12,8
|
Средний
|
|
Эксперимент
отрицательный
|
18,6
+ 1,2
|
21-17
|
2,09
|
11,2
|
Низкий
|
|
III замер
|
|
|
|
|
|
|
Эксперимент
положительный
|
24,2
+ 1,1
|
26-21
|
2,2
|
9,09
|
Низкий
|
|
Контроль
|
21,3
+ 1,5
|
22-20
|
2,6
|
2,2
|
Очень
низкий
|
|
Эксперимент
отрицательный
|
19,5
+ 0,25
|
18-17
|
0,5
|
2,8
|
Очень
низкий
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Следует отметить, что между растениями, которые
подвергались положительному воздействию и растениями ему не подвергавшиеся
выявлена статистически достоверная разница, эти данные представлены в таблицы
55.
Таблица 56 Критерии достоверности различий
поперечного диаметра клубня экспериментально положительной и контрольной группы
|
Сравниваемые
делянки
|
td при измерениях
|
|
Эксперимент
отрицательный- контроль
|
I
|
II
|
III
|
|
3
|
3,2
|
1,6
|
Р=95%
Анализ таблицы 56 показывает, что при I
и II замерах
поперечного диаметра клубня выявлены статистические достоверные различия между
растениями, подвергавшимися информационно- вербально положительному воздействию
и не подвергавшиеся ему. Это позволяет сделать вывод, о том, что информационно-
вербально положительное воздействие стимулирует увеличение поперечного диаметра
клубня.
Таблица 57 Критерии достоверности различий
поперечного диаметра клубня экспериментально отрицательной и контрольной
группы.
|
Сравниваемые
делянки
|
td при измерениях
|
|
Эксперимент
отрицательный- контроль
|
I
|
II
|
III
|
|
1,8
|
0,9
|
2,5
|
Р=95%
Анализ таблицы 57 показывает, что при I
и II замерах
поперечного диаметра клубня не выявлены статистически достоверные различия
между растениями, подвергавшиеся информационно- вербально отрицательному
воздействию. Статистически достоверные различия между растениями выявлены
только при III замере
поперечного диаметра клубня. Это позволяет сделать вывод о том, что
информационно- вербально отрицательное воздействие не значительно стимулирует
уменьшение поперечного диаметра клубня.
Анализ показателей изменчивости количества
стеблей, в момент вегетаций
Таблица 58 Показатели изменчивости количество
стеблей, в момент вегетаций
|
Сравниваемые
делянки
|
Измерение
в см.
|
|
I
|
II
|
III
|
|
Эксперимент
положительный
|
5
|
6
|
6
|
|
Контроль
|
4
|
4
|
5
|
|
Эксперимент
отрицательный
|
3
|
4
|
4
|
Анализ данных таблиц 58 показывает, что
информационно-вербально положительное воздействие стимулирует увеличение
количество стеблей в момент вегетаций, и незначительно влияет информационно-
вербально отрицательное воздействие на уменьшение количество стеблей в момент
вегетаций (таблица 58).
Анализ показателей изменчивости хозяйственной
продуктивности
Таблица 59 Показатели изменчивости хозяйственной
продуктивности
|
Сравниваемые
делянки
|
Измерение
в гр.
|
|
I
|
II
|
III
|
|
Эксперимент
положительный
|
980
|
920
|
940
|
|
Контроль
|
600
|
640
|
520
|
|
Эксперимент
отрицательный
|
540
|
500
|
520
|
Так как у экспериментально положительной группы
растений вес гнезда при I
замере равен 980гр , при II
замере 920гр , при III
замере 940гр, анализ 59 таблицы показывает, что информационно- вербально
положительное воздействие стимулирует увеличения хозяйственной продуктивности.
Тогда как у контрольной группы растений вес гнезда составляет при I
замере 600гр, при II
замере 640гр, III замере
520гр. Также мы можем говорить о том, что информационно- вербально
отрицательное воздействие уменьшает хозяйственную продуктивность, так как у
экспериментально отрицательной группы растений вес гнезда при I
замере равен 540гр, при II
замере 500гр, при III
замере 520гр.
3.5 Биологическая и хозяйственная продуктивность
изученных видов, подвергшихся положительно и отрицательно ментально
биоэнергоинформационному воздействию
Анализ влияния ментально
биоэнергоинформационного воздействия на биологическую и хозяйственную
продуктивность баклажан
Для определения биологической и хозяйственной
продуктивности нами был взвешен вес десяти растений, произросших на площади
равной 1,5 м2, биологическая их продуктивность представлена в кг/м2.
Таблица 60 Результаты изучения биологической и
хозяйственной продуктивности баклажан (кг)
|
№
п/п
|
Воздействие
|
Биологическая
продуктивность
|
Хозяйственная
продуктивность
|
|
1
|
"
+ "
|
4,50
|
3,60
|
|
2
|
"
_ "
|
2,96
|
2,36
|
|
3
|
"
0 "
|
4,34
|
3,50
|
" + " - растения, обработанные
положительно биоэнергоинформационно вербальным воздействием
" - " - растения, обработанные
отрицательно биоэнергоинформационно вербальным воздействием
" 0 " - растения, не подвергавшиеся
воздействию.
Из данных Таблицы 60 видно, что биологическая и
хозяйственная продуктивность растений, получивших положительное вербальное
биоэнергоинформационое воздействие больше, чем у растений, получивших
отрицательное вербальное биоэнергоинформационое воздействие, так же
биологическая продуктивность растений получивших положительное вербальное
биоэнергоинформационое воздействие, больше чем у контрольных растений, не
получивших не кого воздействия .Так, биологическая продуктивность растений,
подвергшихся вербальному биоэнергоинформационому положительному воздействию на
4 % больше, а у растений подвергшихся вербальному биоэнергоинформационому
отрицательному воздействию на 32 % меньше, чем у контрольных. Прирост массы
хозяйственной продуктивности составил: у растений подвергшихся вербальному
биоэнергоинформационому воздействию на 3 % больше, чем у растений, подвергшихся
вербальному биоэнергоинформационому отрицательному воздействию на 32 % меньше,
чем у контрольной.
Более наглядно эти данные проиллюстрированы на
Рисунках 1, 2.
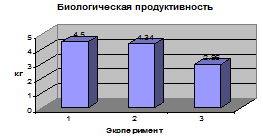
Рисунок 1. Биологическая продуктивность баклажан
- положительное ментально
биоэнергоинформационное воздействие;
- воздействие не оказывалось;
- отрицательное ментально
биоэнергоинформационное воздействие.
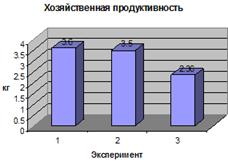
Рисунок 2. Хозяйственная продуктивность баклажан
- положительное ментально
биоэнергоинформационное воздействие;
- воздействие не оказывалось;
- отрицательное ментально
биоэнергоинформационное воздействие.
Анализ влияния ментально
биоэнергоинформационного воздействия на биологическую и хозяйственную
продуктивность перца горького
При определении биологической и хозяйственной
продуктивности экспериментальных, то есть подвергшихся ментальному
биоэнергоинформационому воздействию, и контрольных растений перца горького,
нами были получены следующие результаты (Таблица 61, Рисунки 3,4)
Таблица 61 Результаты изучения биологической и
хозяйственной продуктивности перца горького (кг)
|
№
п/пВоздействиеБиологическая продуктивностьХозяйственная продуктивность
|
|
|
|
|
1
|
"
+ "
|
1,14
|
0,48
|
|
2
|
"
_ "
|
1,06
|
0,40
|
|
3
|
"
0 "
|
1,10
|
0,46
|
"+" - растения, обработанные
положительно биоэнергоинформационно ментальным воздействием
" - " - растения, обработанные
отрицательно биоэнергоинформационно ментальным воздействием
" 0 " - растения, не подвергавшиеся
воздействию.
Из данных Таблицы 61 видно, что биологическая и
хозяйственная продуктивность положительно-экспериментальных растений больше,
чем у отрицательно-экспериментальной и контрольной групп растений. Так,
биологическая продуктивность растений, подвергшихся положительному воздействию
на 4 % больше, а у отрицательно-экспериментальных на 3 % меньше, чем у
контрольных. Прирост массы хозяйственной продуктивности составил: у
положительно-экспериментальной группы растений на 5 % больше, а у
отрицательно-экспериментальной - на 13 % меньше, чем у контрольной. Разницу в
показателях наглядно демонстрируют Рисунки 3, 4.
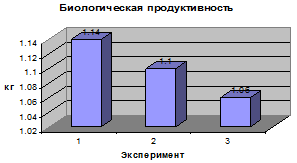
Рисунок 3. Биологическая продуктивность перца
горького
- положительное ментально
биоэнергоинформационное воздействие;
- воздействие не оказывалось;
- отрицательное ментально
биоэнергоинформационное воздействие.
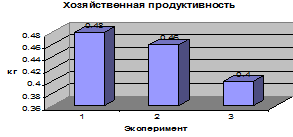
Рисунок 4. Хозяйственная продуктивность перца
горького
- положительное ментально
биоэнергоинформационное воздействие;
- воздействие не оказывалось;
- отрицательное ментально
биоэнергоинформационное воздействие.
Анализ влияния ментально
биоэнергоинформационного воздействия на биологическую и хозяйственную
продуктивность перца сладкого
Исследования биологической и хозяйственной
продуктивности экспериментальных и контрольных растений показали следующие
результаты (Таблица 62, Рисунки 5, 6).
Таблица 62 Результаты изучения биологической и
хозяйственной продуктивности перца сладкого (кг)
|
№
п/пВоздействиеБиологическая продуктивностьХозяйственная продуктивность
|
|
|
|
|
1
|
"
+ "
|
6,96
|
4,80
|
|
2
|
"
_ "
|
6,24
|
4,34
|
|
3
|
"
0 "
|
6,56
|
4,56
|
" + " - растения, обработанные
положительно биоэнергоинформационно ментальным воздействием
" - " - растения, обработанные
отрицательно биоэнергоинформационно ментальным воздействием
" 0 " - растения, не подвергавшиеся
воздействию.
Из данных Таблицы 62 следует, что биологическая
и хозяйственная продуктивность положительно-экспериментальных растений больше,
чем у отрицательно-экспериментальной и контрольной групп растений. Так,
биологическая продуктивность растений, подвергшихся положительному воздействию
на 6 % больше, а у отрицательно-экспериментальных на 5 % меньше, чем у
контрольных. Прирост массы хозяйственной продуктивности составил: у
положительно-экспериментальной группы растений на 5 % больше, а у отрицательно-экспериментальной
- на 5 % меньше, чем у контрольной. Разницу данных показателей наглядно
демонстрируют "Рисунки 5, 6.
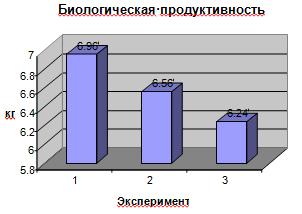
Рисунок 5.Биологическая продуктивность перца
сладкого
- положительное ментально биоэнергоинформационное
воздействие;
- воздействие не оказывалось;
- отрицательное ментально
биоэнергоинформационное воздействие.
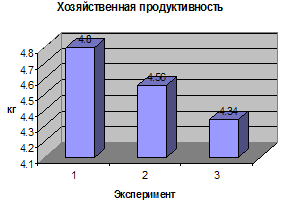
Рисунок 6. Хозяйственная продуктивность перца
сладкого
- положительное ментально биоэнергоинформационное
воздействие;
- воздействие не оказывалось;
- отрицательное ментально
биоэнергоинформационное воздействие
Заключение
Таким образом проведенные нами изучение местных
источников раскрывающею проблемы биоэнергоинформационных взаимодействий в
разных системах показало, что пионерами в области этих исследований Гурвич
А.Г., который экспериментально доказал, что проросшие семена оказывают
дистанционное влияние на другие семена побуждая их так же к прорастанию. Это
влияние он объяснил способностью проросших семян излучать слабые лучи. В более
поздних работах Гурвич А.Г. выявил существования на дегратационное излучение,
которое создаётся любым живым объектом и по физической сути своей представляет
так же поток фотонов в слабом ультрофиолетовомспектре.
П.П. Гаряев, развивая идеи А.Г. Гурвича, в
другой своей работе доказывает, что синтез белка является результатом
генетического кодирования на волновом уровне. Генетическая память трактуется
П.П. Гаряевым и Е.А. Леоновой как солитонно-голографическая. Утверждается, что
хромосомы излучают свет и звук, что гены расщепляются на вещество и поле.
Методом спектроскопии корреляции фотонов в 1985 году П. П. Гаряеву удалось
зафиксировать необычные аномально долго затухающие звуковые колебания ДНК. Было
установлено, что ДНК обладает способностью синтезировать "незамолкающую
сложную мелодию с повторяющимися музыкальными фразами". Геном высших
организмов рассматривается авторами как солитонный биоголографический
компьютер, формирующий пространственно-временную структуру биосистем по
волновому образу-предшественнику. Таким образом, геном работает не только на
вещественном, но и на волновом (тонкоматериальном) уровне. Обращается внимание
на единство фрактальной структуры ДНК и человеческой речи. На основании этой
связи становится возможным воздействовать на ДНК посредством особых
генераторов, преобразующих речевые алгоритмы в солитонные модулированные поля .
Проведенные нами исследования вербального и
ментального биоэнергоинформационного воздействия человека на ростовые процессы
у растений показало, что клубни картофеля выросшие при получении положительной
текстовой информации (в количестве 3 раза в неделю) биометрически достоверна
отличаются от контрольных. При чем эти показатели достигаются в результате
увеличения объема и веса клубней получивших положительную информацию. В то же
время растения картофеля получившие вербальную отрицательную информацию
оказались достоверно меньше по объему и весу как положительно образованных так
и контрольных клубней.
Рассада баклажан, перца сладкого и перца
горького выросшая при получении положительной текстовой информации (в
количестве 3 раза в неделю) биометрически достоверна отличаются от контрольных.
При чем эти показатели достигаются в результате увеличения длины стебля, длины
и ширины листовой пластинки, числа цветков и плодов получивших положительную
информацию. В то же время растения баклажан перца сладкого и перца горького
получившие ментальную информацию оказались достоверно меньше по длине стебля,
длине и ширине листовой пластинки, количеству числа цветков и плодов как
положительно образованных так и контрольных растений.
Выводы:
Результаты наших исследований выделяют ряд
фактов, позволяющих говорить об эффективности вербального и ментальное
биоэнергоинформационно воздействия на рост и развитие растений.
Положительное ментальное биоэнергоинформационное
воздействие вызывает статистически достоверное увеличение по всем исследуемым
признакам: длине стебля, длине и ширине листовой пластинки, количеству цветков
и плодов.
Отрицательное ментальное биоэнергоинформационное
воздействие вызывает статистически достоверное уменьшение длины стебля, длины и
ширины листовой пластинки, количества цветков и плодов.
Положительное ментальное биоэнергоинформационное
воздействие снижают уровень изменчивости ширины и длины листовой пластинки, а
так же числа цветков.
Отрицательное ментальное биоэнергоинформационное
воздействие в ряде случаев повышает уровень изменчивости числа цветков и
плодов.
Положительное вербальное биоэнергоинформационное
воздействие повышает количественную характеристику продольного и поперечного
диаметра клубней.
Отрицательное вербальное биоэнергоинформационое
воздействие снижает количественную характеристику продольного и поперечного
диаметра клубней.
Положительное вербальное и ментальное биоэнергоинформационное
воздействие повышает показатели биологической и хозяйственной продуктивность
растений, а отрицательное воздействие, - понижает.
Таким образом, наши исследования подтверждает
полученные ранее данные экспериментов, проведенных В.М. Инюшиным, Ж. Калиевой и
др. Вербальное и ментальное биоэнергоинформационно воздействие на ростовые
процессы растений может быть использовано как альтернативный,
экологически-чистый метод интенсификации сельскохозяйственного производства, а
также для разработки глобальных систем оздоровления человека, повышающих его
биологический потенциал.
Список использованной литературы
1. Тихоплав В.Ю., Тихоплав Т.С.
"Время Бога". Санкт - Петербург, ИД "ВЕСЬ", 2003.
. Тихоплав В.Ю., Тихоплав Т.С.
"Жизнь на прокат". Санкт - Петербург, ИД "ВЕСЬ", 2003.
. Инюшин В. М., Ильясов Г.У.,
Непомнящих И.А. "Биоэнергетические структуры - теория и практика",
Алма-Ата, "Казахстан",1992.
. Казначеев В. П., Михайлова Л. П.
"Биоинформационная функция естественных электромагнитных полей",
Новосибирск, "Наука",1985.
. Гаряев П.П. "Тупик
генетического кода",1997, Интернет: www.bank
referatov.ru,
С. 2 - 5.
. Гурвич А. Г "Теория
биологического поля", Москва, изд-во "Наука",1944.
. Гаряев П. П., Внучкова В. А.,
Шелепина Г. А., Комиссаров Г. Г. "Вербально-семантические модуляции
резонансов Ферми-Паста-Улама как методология вхождения в командно-образный
строй генома" 1994, Интернет: www.bank
referatov.ru,
С. 17 - 28.
. Гаряев П. П., Маслов М. Ю.,
Решетняк С. А., Щеглов В. А. "Взаимодействие электромагнитного изучения с
информационными биомакромолекулами" // Краткие сообщения по физике.
Физический Институт РАН, 1996, №1 - 2.
. Инюшин В. М, Чекуров П. Р.
"Биостимуляция лучем лазера и биоплазма", Алма-Ата, "Кітап",
1975.
. Коротков К.Г., Чумак А.В.
"Влияние сознания на материальный мир - концепции и эксперимент. От
эффекта Кирлиан к биоэлектрографии", СПб, "Ольга", 1998.
. Гаряев П. П., Македонский С. Н.,
Леонова Е. А. "Биокомпьютер на генетических молекулах как реальность"
// Информационные технологии, №5, Москва, 1997.
. Гаряев П. П. "Волновой
геном", Москва, "Наука",1994.
. Дёмин В. Н. "Тайны русского
народа" // Жур. "Мир на изнанку", Москва "Вече", 2000.
. Гаряев П. П., Чудин В. И., Березин
А. А., Ялакас М. С. "Хромосомный Биокомпьютер" // "Врач",
1991, №4, С. 30 - 33.
. Серобабин А. И. "Что такое
биополе", Санкт-Питербург, "Питер", 1997.
. Казначеев В.П., Спирин Е.А.
"Космопланетарный феномен человека", Новосибирск, "Наука",
Сибирское отделение, 1991.
. Серобабин А. И., Серобабин В. А.
"Познай своё биополе", Москва, 1997.
. Полонников Р.И. "Феномен
информации и информационного взаимодействия", СПб. Санкт-Питербург.
Институт информатики, 2001.
. Чижевский А. Л. "Земное эхо
солнечных бурь", Москва, 1991.
. Мартынов А.М. "Исповедимый
путь", Москва. 1990.
. Гурвич А. Г. "Избранные
труды", Москва, 1977.
. Гурвич А. Г.
"Митогенетическое излучение", Москва, 1932.
. Инюшин В. М. "Биоплазма и
холодная плазма Земли", Алма-Ата, "Казахстан",1997.
. Вейник А. И. "Термодинамика
реальных процессов", Минск, 1999.
. Маковски Е. М. "Природа и
структура живой материи", Бухарест, 1976.
. Сатпрем Р. "Разум
клеток", СПб. "Мирра",1995.
. Карпенко М. Е. "Разумная
Вселенная", Москва, 1992.
. Лупичев Н.Л. "Гомеопатия и
энергоинформатика", Москва, ТОО РИФ "Рой",1994,С.1-2
. Калиева Ж. "Изучение слабого
физического /полевого/ воздействия на пшеницу. В кн.: Биоплазма - феномен
жизни: материалы симпозиума", под ред. Инюшина В. М., Алматы, 1997.
. Шипов Г.И. "Теория
физического вакуума. Теория, эксперименты и технология", 2-е изд., испр. и
доп., Москва, "Наука", 1997.
. Коротков К.Г. "Разработка
научных основ и практическая реализация биотехнических
измерительно-вычислительных систем анализа газоразрядного свечения,
индуцированного объектами биологической природы", СПб. 1999.
. Дёмин В. Н. "Тайны
Вселенной" Москва "Вече", 2005.
. Тихоплав В.Ю., Тихоплав Т.С.
"Великий переход". Санкт - Петербург, ИД "ВЕСЬ", 2004.
. . Сазонова О.Н. "Темы
курсовых работ по генетике с основами селекции" - Москва,
"Просвещение", 1981.
. Тихоплав В.Ю., Тихоплав Т.С.
"Гармония Хаоса или Фрактальная реальность". Санкт - Петербург, ИД
"ВЕСЬ", 2003.
. Удольская Н.Л. "Введение в
биометрию", Алма-Ата "Кітап"-1976.