Изменение костных структур костей пальцев телят в постнатальном онтогенезе
МИНИСТЕРСТВО СЕЛЬСКОГО ХОЗЯЙСТВА
РОССИЙСКОЙ ФЕДЕРАЦИИ
ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ
ОБРАЗОВАТЕЛЬНОЕ
УЧРЕЖДЕНИЕ ВЫСШЕГО ПРОФЕССИОНАЛЬНОГО
ОБРАЗОВАНИЯ
«РЯЗАНСКИЙ ГОСУДАРСТВЕННЫЙ
АГРОТЕХНОЛОГИЧЕСКИЙ УНИВЕРСИТЕТ
ИМЕНИ П. А. КОСТЫЧЕВА» (ФГБОУ ВПО
РГАТУ)
Всероссийский конкурс на лучшую
научную работу среди студентов,
аспирантов и молодых ученых высших
учебных заведений
Министерства сельского хозяйства
Российской Федерации
по Центральному федеральному округу
ТЕМА: «ИЗМЕНЕНИЕ КОСТНЫХ СРУКТУР
КОСТЕЙ ПАЛЬЦЕВ ТЕЛЯТ В ПОСТНАТАЛЬНОМ ОНТОГЕНЕЗЕ»
СТУДЕНТА 13
ГРУППЫ 1 КУРСА ФАКУЛЬТЕТА ВЕТЕРИНАРНОЙ МЕДИЦИНЫ И БИОТЕХНОЛОГИИ
ХЛЕВТОВА
АЛЕКСАНДРА ДМИТРИЕВИЧА
Студент А. Д.
Хлевтов
Научный
руководитель
канд. вет.
наук, доцент В. В. Яшина
Ректор ФГБОУ
ВПО РГАТУ
д-р техн.
наук, профессор Н. В. Бышов
Рязань, 2014
Оглавление
ВВЕДЕНИЕ
1. ОБЗОР
ЛИТЕРАТУРЫ
2. МАТЕРИАЛЫ
И МЕТОДЫ ИССЛЕДОВАНИЙ
3. СОБСТВЕННЫЕ
ИССЛЕДОВАНИЯ
3.1 ИЗМЕНЕНИЕ МАКРОМИКРОСКОПИЧЕСКИХ СТРУКТУР КОСТЕЙ ПАЛЬЦЕВ
ГРУДНЫХ И ТАЗОВЫХ КОНЕЧНОСТЕЙ ТЕЛЯТ
.1.1 КОРОТКИЕ ТРУБЧАТЫЕ КОСТИ (ПУТОВЫЕ И ВЕНЕЧНЫЕ)
.1.2 КОПЫТЦЕВЫЕ КОСТИ
.2 МОРФОМЕТРИЧЕСКИЕ ИЗМЕНЕНИЯ КОСТЕЙ ПАЛЬЦЕВ ГРУДНЫХ И
ТАЗОВЫХ КОНЕЧНОСТЕЙ ТЕЛЯТ
.2.1 МАССЫ ПУТОВОЙ, ВЕНЕЧНОЙ, КОПЫТЦЕВОЙ КОСТЕЙ
.2.2 ДЛИНА ПУТОВЫХ И ВЕНЕЧНЫХ КОСТЕЙ
.2.3 ПЕРИМЕТР ДИАФИЗА ПУТОВЫХ И ВЕНЕЧНЫХ КОСТЕЙ
. ЗАКЛЮЧЕНИЕ
. ПРАКТИЧЕСКИЕ ПРЕДЛОЖЕНИЯ
СПИСОК ЛИТЕРАТУРЫ
ВВЕДЕНИЕ
кость скелет теленок конечность
Рост и формирование костей скелета животных в основе своей определяется
генетическим кодом, но вместе с тем постоянное влияние на них оказывают
многочисленные эндо - и экзогенные факторы (нейрогуморальные, алиментарные,
механические и т.д.), обеспечивая возможность адаптационных преобразований.
При рождении плод из водной среды плодных оболочек попадает в наземные
условия суши, в новое гравитационное поле. В этих условиях на новорожденного
действует большое количество факторов , «запускающих» врожденные реакции. Живой
организм, согласно биологической закономерности, в первые 10 -15 дней жизни
обладает высокой способностью воспринимать все раздражители, поступающие из
среды обитания и незамедлительно реализовывать все существующие в организме
генотипические возможности структурно-функциональной адаптации, обуславливающие
его дальнейшее развитие, выживаемость и сопротивляемость.
В сегодняшних условиях экосистемы животноводческих комплексов телята в
раннем постнатальном онтогенезе, а именно, с первых часов жизни до 20-ти суток
содержатся в клетках площадью 1- 1,25 мˆ2 при постоянстве параметров газового
состава воздуха, температурного и светового режимов, звуковых раздражителей и
ограничения реализации генетической потребности в движении. Это неизбежно
обуславливает изменение морфофункционального состояния всех без исключения
органов и систем.
В настоящее время накоплен большой фактический материал по костной
системе продуктивных животных, по которому можно сделать вывод, что наиболее
изучены и освещены структурно-функциональные особенности длинных трубчатых
костей (1). Сведения о костных органах пальца (путовая, венечная, копытцевая
кости), как органе, испытывающего в период локомоции действия, как статики
живой массы, так и противоударных сил при постановке конечности на почву, с
учетом местоположения фаланг в пальце, вида, возраста животного в научной
литературе малочисленны.
Нами проведены исследования, целью которых было выяснение изменений
костных структур костей пальцев телят в постнатальном онтогенезе.
Задачи исследования:
. Исследовать возрастные изменения костных структур пальцев телят на
макро-, макро-микроскопическом уровнях.
. Изучить морфометрические показатели костных органов пальцев грудных и
тазовых конечностей.
1. ОБЗОР ЛИТЕРАТУРЫ
Вся эволюция животных - доказательство единства организма и среды его
обитания и характеризуется колоссальными различиями формы тела, как формы аппарата
движения, формирующего в связи с разнообразием характера и степенью
двигательной активности, Видимо, поэтому движение явилось одним из условий
существования и совершенствования организма.
Костная система, как основная часть аппарата движения, благодаря своей
полифункциональности, привлекает к себе внимание исследователей различного
направления. Большое количество работ посвящено изучению кости, как органу, в
связи с освоением космического пространства и технического прогресса,
позволяющим уменьшить долю физического труда, тем самым ограничить действие
биомеханических нагрузок. Эти два фактора позволили выяснить значимость костных
органов и всей костной системы в жизнедеятельности организма.
Доказано, что в филогенезе скелет развивался как опорная система, и
совершенствование его шло в направлении повышения биомеханических свойств -
упругости, твердости с одновременной легкостью. Совершенствуясь в процессе
филогенеза, костные органы приобрели способность быстро перестраивать свои
структуры и реагировать на механические воздействия, возникающие при движении,
что в свою очередь привело к активному участию их в трофических и кроветворных
функциях и поставило эти функции в зависимость от движения животного. Таким
образом, костная система млекопитающих и птиц стала не только опорной
конструкцией, но и поставщиком полипотентных клеток гемопоэза и иммуногенеза,
участником обмена веществ (особенно минерального: Са, Р), источником
пъезоэлектричества, электролитического баланса организма. Лабильность костной
системы определяется ее постоянным разрушением и созиданием - ремоделяцией, что
приводит к обновлению в соответствии с запросами всего организма.
Следовательно, процессы морфогенеза кости и поддержание его
структурно-функционального гомеостаза обеспечиваются механизмами тканевого,
органного, системного и организменного уровней.
Изучая скелет на системном уровне, исследователи (8) отмечают, что рост и
формирование костей скелета животных в основе своей определяются генетическим
кодом, однако, вместе с тем, постоянное влияние на них оказывают и
многочисленные эндо- и экзогенные факторы (нейрогуморальные, алиментарные,
экологические, механические и др.). Авторами так же установлено, что
биомеханическая нагрузка, падающая на скелет во время локомоции (статика и
динамика), является основным фактором, обеспечивающим его морфофункциональное
состояние, каждой кости как органа. Во время движения силы, возникающие при
упругих деформациях в костях, определяют их строение, придают им
соответствующую величину и форму, приводят к тому, что структура костной ткани,
ее биофизические свойства проявляются в зависимости от расположения кости в
определенных звеньях скелета (3, 9).
В настоящее время уже ни у кого не вызывает сомнения тот факт, что каждая
кость скелета является самостоятельным органом, способным изменять форму и
внутреннюю структуру в связи с возрастом, видом, породой, полом, рационом
кормления и условиями функционирования локомоторного аппарата млекопитающих (5)
. Доказано, что в структуре каждой кости заложено два начала: статическое
(морфологическое), генетически обусловленное; динамическое (функциональное),
приобретенное на поздних антропогенезах. Условия же состояния статики и
динамики костных органов неизбежно изменяются в течение всей жизни индивидуума.
Ко всем этим условиям кость приспосабливается путем изменения внутренней
архитектоники, сочетая два разнонаправленных процесса - восстановление и
разрушение (6).
Основным фактором, вызывающим регенерацию и резорбцию кости являются две
формы биомеханической нагрузки, действующей на кость в процессе движения -
сжатие и растяжение. Костные поверхности, подвергающиеся непосредственному
давлению, растут на базе хряща, а все костные поверхности, подвергающиеся
влиянию растяжения, растут на основе соединительной ткани.
Костная ткань млекопитающих и птиц в зависимости от характера действия
биомеханической нагрузки формируется в двух направлениях: там, где кость
испытывает больше силы на излом, строится компактное вещество кости, там, где
действуют больше силы сжатия, формируется губчатое вещество (спонгиоза).
Под действием механической нагрузки в костях возникают упругие
деформации, силы и энергия которых оказывают влияние не только на кость, но и
на ее рецепторный аппарат (посылая эфферентные импульсы в мозг) и на условия
оттока и лимфы по сосудам (направляя кровь в сердце). Биомеханический фактор,
действующий на скелет, явился той необходимой причиной, тем стимулом, который
помимо основной филогенетически более древней опорно-двигательной функции
скелета обусловил в кости кроветворение, электролитические и трофические
функции, обеспечивающие гомеостаз организма. Влияние биомеханического фактора
на физиологическую перестройку костной ткани человека и животных в возрастном
аспекте характеризуется периодичностью процессов резорбции и костеобразования.
Криштофорова Б.В. (5), исследуя костную систему крупного рогатого скота и
кур в постнатальном онтогенезе при различной локомоции с учетом кормления и
содержания, установила четыре основных периода:1 -замедленного роста и
развития; 2- интенсивного роста и замедленного развития; 3- замедленного роста
и интенсивного развития; 4-4-постоянной и постепенной ремоделяции. Автор
отмечает, что действие гиподинамии изменяет последовательность стадийности
созревания костных структур и увеличивает одни и сокращает другие периоды роста
и развития костей.
Бамбуляк Н . Ф.(2) выделяет два периода роста и развития трубчатых костей
ремонтных свинок и указывает на тот общебиологический факт, что проявление
структурных изменений при различной степени локомоции больше в том периоде. В
котором происходит наиболее интенсивный рост и развитие органов, а значит и чем
моложе животное. Следовательно, различная степень двигательной активности в
динамике влияет на размеры, массу костей, физическую плотность, объем, а также
на процессы синтеза и резорбции кости, характеризующиеся временными величинами:
скоростью образования и созревания костной ткани, интенсивностью аппозиционного
и остеохондрального остеогенеза. Недостаточность в двигательной активности
обусловливает снижение массы и объема диафиза при неизменных плотности и длине
кости.
Харченко Л.Г. (11), учитывая функциональную дифференциацию между
конечностями, указывает, что масса путовой и венечной костей грудной конечности
крупного рогатого скота, независимо от уровня двигательной активности,
закономерно выше, чем тазовой, исключая копытцевую кость. Автор отмечает, что
интенсивность роста костей акроподия в постнатальном онтогенезе незначительна
по сравнению с проксимальными звеньями конечностей. В постнатальном онтогенезе
закономерно уменьшается в костной ткани органические и увеличиваются
минеральные компоненты, обусловливающие плотность и характеризующие ее
реактивность и устойчивость к патологическим процессам. Плотность костной ткани
и кости в целом изменяется не только с возрастом животного, но и с положением
кости в скелете, а также участка в одной и той же кости. Плотность каждой кости
изменяется в зависимости от падающей на нее биомеханической нагрузки. Снижение
нагрузки на костную систему вызывает нарушение белкового и минерального (Са и
Р) обменов, что приводит к уменьшению плотности костей. Доказано, что
повышенная двигательная активность способствует увеличению плотности в костях,
обусловливая их прочность. Линейные параметры костей конечностей млекопитающих
и человека в онтогенезе также обусловлены величиной и характером функциональных
нагрузок. Мельник К.П., Клыков В.И. (8)констатируют, что продольные размеры
костей тазовых конечностей всегда больше грудных. Однако, влияние динамических
и статических нагрузок на изменение линейных размеров костей по данным
исследователей носит противоречивый характер. Усиленная функция мышц в
результате активации движения стимулирует продольный рост трубчатых костей,
Интенсивность этого роста повышается при умеренно увеличенных динамических и
физических нагрузках, чрезмерные же нагрузки вызывают замедление роста костей.
Повышенная статическая нагрузка на конечности вызывает увеличение, в основном,
поперечных размеров скелетных элементов.
В онтогенезе каждая кость конечности имеет различный градиент роста.
Дистальные звенья конечностей перестают расти раньше. Исаенков Е.А. (4),изучая
линейный рост скелета тазовой конечности в онтогенезе романовских овец,
отмечает, что в утробном развитии каждая кость конечности растет в длину тем
интенсивнее, чем дальше она расположена от тазовых костей, за исключением 3-ей
фаланги пальцев. После рождения наименьшей интенсивностью роста отличается
плюсневая кость. А по направлению вверх и вниз от нее интенсивность роста
костей увеличивается. В результате неравномерного роста отдельных костей
конечностей с возрастом изменяется относительная длина их в пределах
конечности. Автор указывает, что кости стопы более быстрыми темпами
приближаются к дефинитивной длине, составляющие у новорожденных 60-75% от их
длины взрослых животных, тогда как вышерасположенные кости - 43-53%. Самой
первой костью, достигшей к 12-ти месячному возрасту окончательной величины,
является 1-ая фаланга; к 15 - месячному возрасту 2-ая фаланга, бедренная,
плюсневая и заплюсневые; к 18 - месячному - 3 -я фаланга.
В процессе роста и развития трубчатых костей происходит увеличение
размера поперечника диафиза. Толщина диафизарных стенок зависит от
интенсивности костеобразовательных процессов, происходящих с наружной стороны
кости и от активности резорбции со стороны костно-мозгового участка диафиза.
Изменение этих параметров связано с возрастом, полом животных, активностью
движения. Доказано, что новорожденные животные, испытывающие наименьшую
биомеханическую нагрузку живой массы, имеют костно-мозговой участок диафиза
больше в длинных трубчатых костях дистальных звеньев, чем проксимальных, Он
достигает у крупного рогатого скота 50% длины кости, а у птиц (цыплят) менее
40%. В костях пальцев имеется костно-мозговой участок диафиза, хотя и
незначительный.
Упоминая о мере биологической надежности структур растущей кости в
процессе адаптации к физическим нагрузкам, Криштофорова Б.В. (6) указывает, что
она может происходить при расширении диафиза и костно-мозгового участка
диафиза, стабильном или даже истончающемся компактном веществе; сужении диафиза
и его костно-мозгового участка при отчетливом утолщении компактного вещества.
Автор доказывает, что биомеханические свойства кости, как устойчивой
механической конструкции, в первом случае более надежны. Снижение локомоции
также обусловливает «убыль» костной ткани и увеличение костно-мозгового участка
диафиза. Однако, действие же длительных интенсивных физических нагрузок
длительного характера также приводит к уменьшению толщины компактного слоя длинных
трубчатых костей. В условиях постепенно возрастающих динамических нагрузок
кортикальный слой диафиза становится широким. А губчатое вещество занимает в
кости большее пространство.
Высокая чувствительность костной ткани к деформациям, возникающим при движении,
обусловливает адаптационную перестройку внутренней структуры кости, как органа,
т.е. перестройку губчатого и компактного веществ кости.
Компактное вещество, формируя стенку кости, распределяется по ее длине
неодинаково в различных скелетных звеньях конечностей. Оно характеризуется
консервативностью к приспособительной перестройке, по сравнению с губчатым
веществом, выполняющим функцию выравнивания напряжений, действующих на
кортикальную субстанцию кости. Скорость перестройки спонгиозы в 8-10 раз больше,
чем компактного вещества. Характеризуя развитие компактного вещества в
скелетных элементах конечностей, исследователи отмечают постепенное утолщение
стенок плечевой кости от проксимального конца к дистальному, в костях
предплечья и пясти в средней трети и проксимальной половине (стопо - и
пальцеходящие) или в средней трети (фалангоходящие). Для элементов тазовой
конечности характерно утолщение в средней трети и начале проксимальной
(бедренная кость и кости плюсны) или средней и начале дистальной трети
(большеберцовая кость ) (8).
Слесаренко Н. А.(10), изучая кости акроподия крупного рогатого скота,
сообщает о наибольшей толщине компакты в области средней частей 1- ой и 2 - ой
фаланг и среднего участка дорсальной поверхности 3 -ей фаланги Автор указывает,
что увеличение компакты в дистальном направлении происходит независимо от
условий содержания и кормления. Имеются сведения. Что у кроликов, находившихся
в условиях свободного содержания, проксимальная фаланга первого пальца
толстостенная. Стенки проксимальных и средних фаланг других пальцев лучше
развиты в дистальной трети таким образом, что костно-мозговой участок диафиза
конусовидно сужен к дистальному концу. Дистальные фаланги также относительно
тонкостенны. Ограничение подвижности проявляется также в истончении и резорбции
губчатого вещества.
Структура губчатого вещества трубчатых костей конечностей отличается от
компактного более разряженным строением, неоднородностью и зависит от места
расположения в кости, положения кости в звене конечности возраста животного и
степени его локомоции. Оно представлено костными трабекулами, пластинами,
балками с очень разнообразным расположением в соответствии с направлением
основных линий действия силы сжатия и растяжения ткани.
В трубчатых костях млекопитающих выделяют мелкоячеистую зону губчатого
вещества, занимающую эпифизарные субхондральные зоны, среднеячеистую и
крупноячеистую - в проксимальных и дистальных участках диафиза
Развитие спонгиозы по данным Мельника К.П. , Клыкова И.В.(8) положительно
коррелирует со степенью полифункциональности локомоторного аппарата животных.
Авторы отмечают, что повышение функциональной специализации грудной конечности
(локомоция, рытье, хватательные движения у норных и полуводных форм грызунов)
усиливает развитие спонгиозы, которая заполняет (частично или полностью) весь
костно-мозговой участок диафиза лучевой кости. Однако, узкая локомоторная
специализация конечностей, ограничение подвижности, повышение доли статических
нагрузок в локомоторном цикле ограничивают развитие губчатого вещества,
вызывают очаговую резорбцию и частичную деструкцию балок. Эти явления, хотя и
прогрессируют с возрастом, протекают на фоне компенсаторных изменений.
Резорбция, прежде всего, происходит тех костных балок, которые меньше всего
испытывают механическое напряжение, отмечается стирание границ между костными
пластинами, которые становятся расплывчатыми, утрачивая структуру кости. Тем
самым подчеркивается значимость характера и направления биомеханической
нагрузки в определении количественного содержания и архитектоники костных
структур в кости. Одновременно было доказано, что костные пластинки в трубчатых
костях распределяются соответственно направлениям максимальной тяги и давления,
отвечая законам статики и динамики.
Формирование кости, ее рост, изменения в постнатальном онтогенезе под
влиянием функции совершается во всех ее участках, где имеются клетки в
состоянии активности. Такая активность может проявляться в костных клетках со
стороны надкостницы, эндоста или суставных и метафизарных хрящей. Вот почему
возможен рост кости в толщину - за счет надкостницы и эндоста (аппозиционный
рост) и в длину за счет зоны роста, а также за счет клеток остеонов
(мезостальный рост). В разных зонах растущей кости остеопластический процесс
существенно отличается по своей интенсивности, которая зависит от влияния
каких-либо факторов и условий, определяющих общее течение процессов
морфогенеза. Одним из раздражающих факторов являются различного рода
биомеханические нагрузки, (статические, динамические). Тяга мышц активизирует
клетки надкостницы и компактного вещества, что способствует как мезостальному
так и аппозиционному росту кости. Важным побуждающим фактором роста является не
только тракция мышц, но и растяжение надкостницы при сгибании костей под
тяжестью тела, В этом случае клетки надкостницы также активизируются, что
стимулирует рост кости в толщину. Однако постоянное давление в одном участке
тормозит рост кости в длину. Известно, что активная поверхность 1 - го г
костной ткани составляет около 200 м², а всей костной ткани примерно 2 км², чем и объясняются быстро
развивающиеся адаптационные преобразования в костной системе животных при смене
режимов двигательной активности. Рост диафиза длинной трубчатой кости в длину
осуществляется за счет активной деятельности метафизарного хряща. Функционально
он обеспечивает ориентацию новообразованных трабекул, участвует в передаче и
распределении силовых нагрузок в растущей кости. Рост эпифизов происходит за
счет клеток суставного хряща. Толщина суставных и метафизарных хрящей различна
(от 0,5 мм до 4-5 мм) и соответствует испытываемому давлению. При подвижности в
суставе в хрящ сохраняет жизнеспособность. Важную роль в функционировании
хрящей играет подлежащая к нему субхондральная кость. Функциональное единство
кости и хряща определяет темп, характер развития и функционирования основных
звеньев скелета конечностей, его адаптационные возможности при изменении
условий существования организма.
Исследования Криштофоровой Б.В.(7) показывают, что величина и структура
метафизарных хрящей и их субхондральных костей изменяются с возрастом, степенью
двигательной активности и положением кости в звене конечности животных.
Известно, что кости стило-, зейгоподия - биэпифизарные, кости метаподия -
дистально моноэпифизарные, кости акроподия (путовая, венечная ) -проксимально
моноэпифизарные. Кость третьей фаланги (копытцевая кость) парнокопытных
животных характеризуется наличием суставного хряща для сочленения с дистальным
концом кости второй фаланги и надкостницы, покрывающей оставшуюся поверхность
кости.
Сроки синостоза эпифизов и диафизов трубчатых костей конечностей животных
являются одним из признаков физиологической зрелости организма. Причем
биомеханическая нагрузка прямым образом влияет на процессы синостозирования
трубчатых костей. Существует зависимость между сроками синостозирования костей
и объемом движения в суставах. Синостозирование костей, выполняющих большую
статическую нагрузку в сочетании с достаточным объемом движений, начинается
раньше, чем в менее нагруженных. Процесс синостозирования сопровождается
структурными изменениями в метафизарном хряще, эпиметафизарной и диметафизарной
субхондральных костях.
Таким образом, анализ литературы показывает, что у продуктивных животных
больше всего освещены структурно-функциональные особенности длинных трубчатых
костей во взаимосвязи с условиями содержания и кормления. Сведения о структуре
костных органов пальцев (путовая, венечная, копытцевая) малочисленны.
2. Материал и методы исследований
Работа выполнена на кафедре анатомии и физиологии сельскохозяйственных
животных Рязанского государственного агротехнологического университета им. П.А.
Костычева.
Проводили морфологические исследования путовых, венечных, копытцевых
костей (по n=32) пальцев грудных и тазовых
конечностей телят черно-пестрой породы: новорожденные, 30 - суточные,
выращиваемых согласно технологии, принятой в условиях животноводческого
хозяйства «Стенькино» Рязанского р-на, Рязанской области. Отобранный материал
фиксировали в растворе (3-,5-,10%) нейтрального формалина. Использовали
комплекс морфологических методик. Отпрепарированные элементы (кости) пальцев
взвешивали, линейные промеры коротких трубчатых костей (рис. 1,2) производили
при помощи линейки с ценой деления в 1 мм. Рентгенографию провели по
общепринятым методам. На органном - целые кости, интроорганном уровне - шлифы
костей. Рентгенограммы изучали при помощи негатоскопа и лупы.

Рисунок 1 - Длина костей: а) путовой, б) венечной
Рисунок 2 - Периметр диафиза костей: а) путовой, б) венечной
3. Собственные исследования
.1 Изменения макро-микроскопических структур
костей пальцев грудных и тазовых конечностей телят
Аналогичность биомеханической функции при поступательном движении (опора
и тракция сухожилий мышц пальцевых сгибателей и разгибателей) стала одной из
причин общности морфологии костей акроподия.
3.1.1 Короткие трубчатые кости (путовые и
венечные)
На обзорных рентгенограммах костей (путовых и венечных) пальцев грудных и
тазовых конечностей прослеживается определенная структура, характерная для
коротких трубчатых костей новорожденных телят. В монофизарных путовых и
венечных костях четко различимы проксимальный и эпифиз и диафиз, отграниченные
друг от друга линией зоны роста: метафизарным хрящом, эпифизарной и
диаметафизарной субхондральными костями. В коротких трубчатых костях поперечник
эпифизов несколько больше по сравнению со средним участком диафиза, что
особенно проявляется в путовых костях. Диафизы путовых костей вытянуты более
продольно, чем в венечных. Губчатое вещество проксимальных и дистальных
участков диафизов этих костей мелкоячеистой, а компактное сетчатой структуры
(рис. 3,4).
Компактный слой диафиза путовых костей тонкий, лишь в дистальной части
несколько утолщается. В венечных костях компактный слой сетчатой структуры. В
зоне костно-мозгового участка диафиза путовых костей губчатое вещество
разреженное и содержит тонкие единичные трабекулы, ориентированные параллельно
длинику кости. В венечных он заполнен среднеячеистым губчатым веществом.
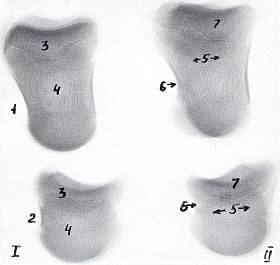
Рисунок 3 - Рентгенограммы костей пальцев грудной конечности теленка.
Новорожденный. Ι - 3-ий и ΙΙ - 4-ый пальцы.
- путовая и 2 - венечная кости, 3 - эпифиз, 4 - диафиз, 5 - губчатое и 6
- компактное вещества, 7 - зона роста
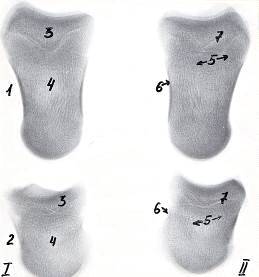
Рисунок 4 - Рентгенограммы костей пальцев тазовой конечности теленка.
Новорожденный. Ι - 3-ий и ΙΙ - 4-ый пальцы.
- путовая и 2 - венечная кости, 3 - эпифиз, 4 - диафиз, 5 - губчатое и 6
- компактное вещества, 7 - зона роста
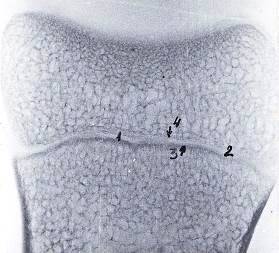
Рисунок 5 - Рентгенограмма шлифа проксимального эпифиза и диафиза путовой
кости 3-го пальца грудной конечности теленка. Новорожденный
- метафизарный хрящ, 2 - очаг синостоза, 3 - диаметафизарная
субхондральная кость, 4 - эпиметафизарная субхондральная кость
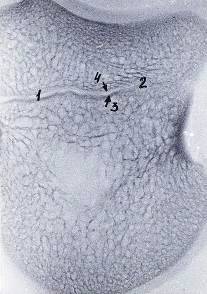
Рисунок 6 - Рентгенограмма шлифа венечной кости 3-го пальца грудной
конечности теленка. Новорожденный.
- метафизарный хрящ, 2 - очаг синостоза, 3 - диаметафизарная
субхондральная кость, 4 - эпиметафизарная субхондральная кость
На макро-микрорентгенограммах шлифов (рис. 5,6) под субхондральной костью
суставного хряща эпифизов отмечается некоторое утолщение костных трабекул,
которые располагаются параллельно суставной поверхности. Субхондральная кость
суставных хрящей без видимых границ переходит в примитивное губчатое вещество
проксимальных эпифизов и дистальных участков диафиза.
В зоне роста, в толще метафизарного хряща путовых и венечных костей на
рентгенограммах и их шлифов выявляются очаги синостоза, как в центральной части,
так и ближе к периосту. В участках разрушения метафизарного хряща,
эпиметафизарной, диметафизарной субхондральной костей наблюдается
бесструктурное губчатое вещество, состоящее из обломков костных трабекул,
пониженной рентгенплотности.
У телят 30-ти суточного возраста выявляются структурные отличия костных
структур коротких трубчатых костей пальцев по отношению к новорожденным.
На рентгенограммах костей телят 30-суточного возраста, наряду с
резорбцией губчатого вещества в диафизе коротких трубчатых костей 3-го и 4-го
пальцев, отмечается и некоторое утолщение компактного вещества. В большей мере
это проявляется на плантарных поверхностях путовых костей тазовых конечностей
(рис.7,8) . По строению компактный слой сетчатой (верхняя 1/3 стенки диафиза) и
пластинчатой (дистальные 2/3 стенки диафиза) структуры.
В венечных костях 3-го и 4-го пальцев компактное вещество нечеткое,
слоистой структуры, с различной рентгенплотностью. Периостальные его
поверхности ровные, а эндостальные - неровные с наличием костных трабекул,
отходящих под разным углом. На рентгенограммах шлифов коротких трубчатых костей
губчатое вещество проксимального и дистального участков диафиза мелкоячеистой
структуры.
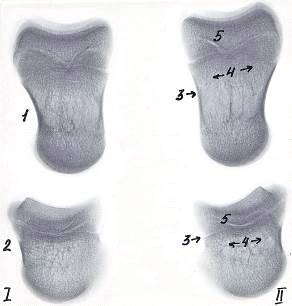
Рисунок 7 - Рентгенограммы костей пальцев грудной конечности теленка.
суток. Ι - 3 - й и ΙΙ - 4-й пальцы
- путовая кость, 2 - венечная кость, 3 - компактное вещество,
- губчатое вещество, 5 - зона роста
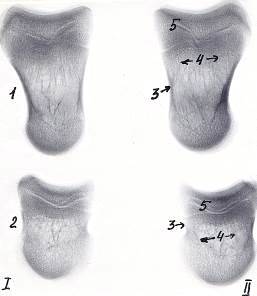
Рисунок 8 - Рентгенограммы костей пальцев тазовой конечности теленка. 30
суток. Ι
- 3-ий и ΙΙ - 4-ый пальцы
- путовая кость, 2 - венечная кость, 3 - эпифиз, 4 - диафиз,
- губчатое вещество, 6 - компактное вещество, 7 - зона роста
В центральной части диафиза костные балки спонгиозы истончены,
фрагментированы. Более наглядно это выражено в путовых костях пальцев грудных и
тазовых конечностей, где архитектоника спонгиозы изменяется с образованием
обширных пространств, заполненных костным мозгом, с редкими, косо-продольно
расположенными костными трабекулами (рис.9). В венечных костях 3-го и 4-го
пальцев телят также отмечается разряжение спонгиозы в середине диафиза (10).
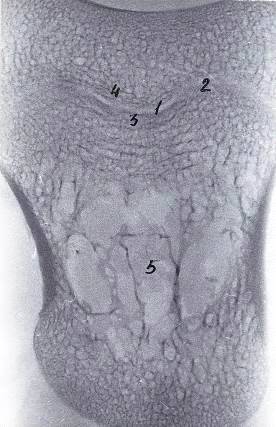
Рисунок 9 - Рентгенограмма шлифа путовой кости 3-го пальца тазовой
конечности теленка. 30 суток
- метафизарный хрящ, 2 - очаг синостоза, 3 - диаметафизарная
субхондральная кость, 4 - эпиметафизарная субхондральная кость, 5 -
костно-мозговой участок.
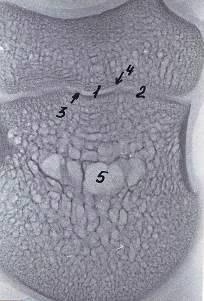
Рисунок 10 - Рентгенограмма шлифа венечной кости 3-го пальца тазовой
конечности теленка. 30 суток
- метафизарный хрящ, 2 - очаг синостоза, 3 - диаметафизарная
субхондральная кость, 4 - эпиметафизарная субхондральная кость, 5 -
костно-мозговой участок
На рентгенограммах шлифов путовых и вечных костей в зонах роста
метафизарного хряща отмечается наличие участков синостоза. Метафизарный хрящ
имеет вид прерывистой извилистой линии. В местах его разрушения формируется
мелкоячеистое губчатое вещество.
3.1.2 Копытцевые кости
Копытцевая кость по своей форме отличается от коротких трубчатых костей
пальцев. На ренрентгенограммах имеет вид треугольника с основанием по линии
опоры (рис.11).
А
Б
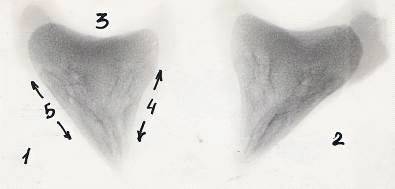
Рисунок 11 - Рентгенограммы копытцевых костей теленка. Новорожденный.
А - грудная конечность, Б - тазовая конечность
- 3-й палец, 2 - 4-й палец, 3 - суставная поверхность, 4 - дорсальная
поверхность, 5 - подошвенная поверхность
Зона роста в ней представлена суставным хрящом и его субхондральной
костью. В копытцевой кости отмечается периостальное костеобразование,
обусловливающее образование компактного слоя. Дорсальная и подошвенная стенки
кости выявляются в виде слабо выраженных костных структур: сетчатой (со стороны
подошвы), слоистой (со стороны дорсальной стенки) (рис.12). От их внутренних
поверхностей отходят под острым углом трабекулы, анастомозирующие друг с другом
и ориентированные параллельно соответствующим стенкам кости. Губчатое вещество
располагается в виде треугольника: от суставной поверхности к центру кости.
Субхондральная кость суставного хряща копытцевой кости пальцев без видимых
границ переходит в примитивное губчатое вещество. Костные трабекулы формируют
костные каналы для прохождения сосудов и нервов.
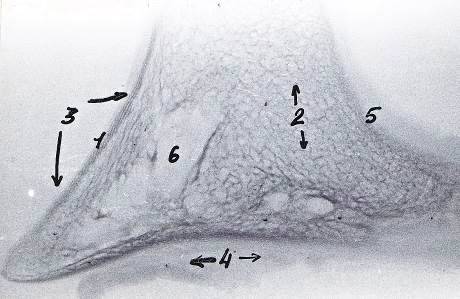
Рисунок 12 - Рентгенограмма шлифов копытцевой кости 3-го пальца грудной
конечности теленка. Новорожденный
- компактное вещество, 2 - губчатое вещество, 3 - дорсальная стенка, 4 -
подошвенная стенка, 5 - субхондральная кость суставного хряща, 6 - костные
каналы сосудов
У телят 30-суточного возраста в копытцевых костях на рентгенограммах
отмечается ориентация костных трабекул губчатого вещества под субхондральной
костью. Контур дорсальной стенки едва различим и имеет рентгеноплотность ниже,
чем подошвенная (рис.13). Слоистое строение компактного слоя подошвы занимает
первые 2/3 части всей ее длины. На дорсальной стенке компактное вещество
сетчатой структуры и пониженной рентгенплотности в нижней части кости. Губчатое
вещество в центре кости разряжено, без четкой ориентации костных трабекул
(рис.14).
А
Б
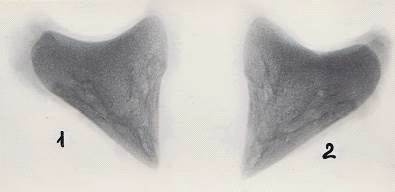
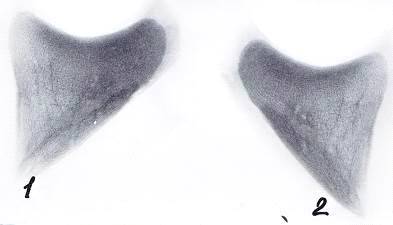
Рисунок 13 - Рентгенограммы копытцевых костей теленка. 30 суток
А - грудная конечность, Б - тазовая конечность
- 3-й палец, 2 - 4-й палец, 3 - суставная поверхность, 4 - дорсальная
поверхность, 5 - подошвенная поверхность.
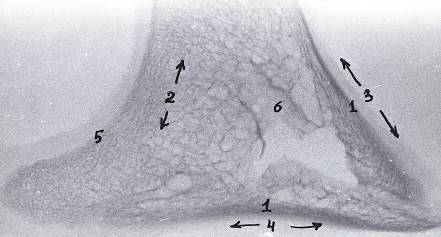
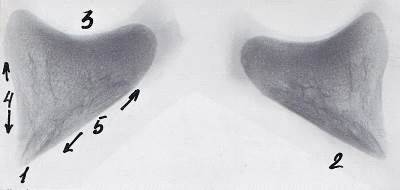
Рисунок 14 - Рентгенограмма шлифов копытцевой кости 3-го пальца грудной
конечности теленка. 30 суток
- компактное вещество, 2 - губчатое вещество, 3 - дорсальная стенка, 4 -
подошвенная стенка, 5 - субхондральная кость суставного хряща, 6 - костные
каналы сосудов
.2 Морфометрические изменения
костей пальцев грудных и тазовых конечностей телят
Адаптационная перестройка внутренней архитектоники костных органов пальцев
сопряжена с изменением их органной структуры. Интенсивность изменений роста и
развития фаланг пальцев грудных и тазовых конечностей телят проявляется
остеометрическими характеристиками.
.2.1 Массы путовой, венечной,
копытцевой костей
Таблица 1
Изменения абсолютной массы костей пальцев телят, (г)
|
возраст
|
конечность
|
палец
|
путовая
|
венечная
|
копытцевая
|
|
новорожденный
|
грудная
|
3
|
19,68 ± 0,74
|
11,98 ± 0,39
|
10,57 ± 0,37
|
|
|
4
|
19,05 ± 0,84
|
11,56 ± 0,47
|
9,48 ± 0,29
|
|
тазовая
|
3
|
20,13± 0,77
|
11,96 ± 0,44
|
9,93 ± 0,26
|
|
|
4
|
19,64 ± 0,86
|
11,89 ± 0,51
|
9,90 ± 0,36
|
|
30 суток
|
грудная
|
3
|
19,64 ± 0,48
|
12,02 ± 0,49
|
11,23 ± 0,30
|
|
|
4
|
18,73 ± 0,62
|
11,92 ± 0,43
|
9,89 ± 0,29
|
|
тазовая
|
3
|
18,76 ± 0,76
|
11,93 ± 0,25
|
9,79 ± 0,05
|
3.2.2 Длина путовых и венечных костей
Длина коротких трубчатых костей пальцев телят преобладает в путовых
костях и тазовых конечностях (табл.2).У новорожденного теленка длина путовых
костей на тазовых конечностях на 6,75%(на 3-ем пальце) и на 4,75%(на 4-ом
пальце) больше, чем на грудных. К 30-суточному возрасту телят отмечается
уменьшение длины путовых и венечных костей по сравнению с новорожденными. Также
наблюдается уменьшение длинниковых промеров коротких трубчатых костей с большей
интенсивностью на тазовых конечностях.
Таблица 2
Изменения длины коротких трубчатых костей пальца телят, (см)
|
возраст
|
конечность
|
палец
|
путовая
|
венечная
|
|
новорожденный
|
грудная
|
3
|
4,59 ± 0,09
|
3,31 ± 0,06
|
|
|
4
|
4,63 ± 0,07
|
3,31 ± 0,08
|
|
тазовая
|
3
|
4,90 ± 0,11
|
3,31 ± 0,05
|
|
|
4
|
4,85 ± 0,11
|
3,32 ± 0,06
|
|
30 суток
|
грудная
|
3
|
4,56 ± 0,01
|
|
|
4
|
4,56 ± 0,01
|
3,14 ± 0,05
|
|
тазовая
|
3
|
4,70 ± 0,04
|
3,20 ± 0,07
|
|
|
4
|
4,67 ± 0,03
|
3,27 ± 0,04
|
3.2.3 Периметр диафиза коротких трубчатых костей
Периметры диафизов (в самой узкой его части) коротких трубчатых костей
телят изменяются по следующей закономерности: чем меньше длина кости, тем
больше величина периметра (табл.3). У новорожденных телят периметр диафиза
костей 1-ой и 2-ой фаланг пальцев колеблется незначительно (в путовых -
6,43-6,68 см, венечных - 6,53-7,09см) при наибольшем периметре диафиза венечной
кости 3-го пальца грудной конечности (7,09см).
К 30-ти суточному возрасту телят происходит (по отношению к предыдущему
возрасту) уменьшение периметров диафиза путовых и венечных костей и в большей
степени в костях 3-го пальца тазовой конечности (путовая 7,73%, венечная
7,63%).
Исследования показывают, что активизация функциональной перестройки в
этом возрасте отражается на морфометрических показателях костных органов телят.
Уменьшение морфометрических параметров в коротких трубчатых костях пальца
происходит, возможно, из-за действия на них статистики увеличивающейся массы
тела и быстрого разрушения утробных структур.
Таблица 3
Изменения периметра диафиза коротких трубчатых костей пальца телят, (см)
|
возраст
|
конечность
|
палец
|
путовая
|
венечная
|
|
новорожденный
|
грудная
|
3
|
6,68 ± 0,12
|
7,09 ± 0,16
|
|
|
4
|
6,51 ± 0,11
|
6,79 ± 0,16
|
|
тазовая
|
3
|
6,43 ± 0,10
|
6,7 ± 0,10
|
|
|
4
|
6,63 ± 0,22
|
6,53 ± 0,13
|
|
30 суток
|
грудная
|
3
|
6,27 ± 0,05
|
6,83 ± 0,08
|
|
|
4
|
6,18 ± 0,08
|
6,75 ± 0,09
|
|
тазовая
|
3
|
5,97 ± 0,09
|
6,30 ± 0,01
|
|
|
4
|
6,17 ± 0,07
|
6,60 ± 0,06
|
4. ЗАКЛЮЧЕНИЕ
Исследования костных органов пальцев грудных и тазовых конечностей телят,
выращиваемых по технологии, принятой в хозяйстве, показывают, что их
морфофункциональный статус изменяется в зависимости от возраста. Характер их
структурной адаптации зависит от активности движения и местоположения фаланг в
пальце грудных тазовых конечностей животных. Вместе с тем, структурные
изменения копытцевой кости обусловлены, по-видимому, ее положением в роговом
копытце. Адаптогенез костных органов пальцев на разных этапах роста и развития
имеет свои особенности, что проявляется в их интраорганной структуре и
морфологических показателях.
Для новорожденных животных характерно наличие утробных структур в костных
органах пальцев грудных и тазовых конечностей. Утробные структуры определяются
наличием синостоза проксимального эпифиза и диафиза в коротких трубчатых
костях. Компактный слой костей сетчатой структуры с минимальной развитостью.
Губчатое вещество примитивное. Костно-мозговые участки диафиза в путовых и
венечных костях на обзорных рентгенограммах практически неразличимы. На
макро-микрорентгенограммах шлифов этих же костей в участках разрушения
метафизарного хряща обнаруживается бесструктурное губчатое вещество.
Мы думаем, что утробный синостоз, отмечаемый в путовых и венечных костях
в зоне метафизарного хряща, носит временной характер, так как в дальнейшем
происходит его замещение и образование структур зон роста костей. Особенность
структуры копытцевой кости определяется энхондральным и эндесмальным
остеогенезом.
Костные органы пальцев новорожденных телят характеризуются определенными
морфометрическими параметрами: масса уменьшается в дистальном направлении
пальца, длина превалирует в путовых костях грудных и тазовых конечностей,
периметры в венечных.
У 30-суточных телят отмечается функциональная перестройка костных органов
пальцев. Отмечается резорбция костных структур, проявляясь «убылью» костной
ткани, уменьшением морфометрических параметров и больше в путовых и венечных
костях как грудных, так и тазовых конечностей. Видимо, действие на них статики
увеличивающейся массы обусловливает более быстрое разрушение утробных структур.
На обзорных и шлифах рентгенограмм путовых и венечных костей отмечаем в
зонах роста метафизарного хряща наличие участков синостоза со структурой
мелкоячеистого губчатого вещества.
Таким образом, в своих исследованиях мы показали, что костные органы
пальцев телят изменяются в соответствии с ростом, развитием и структурным
адаптогенезом всего организма. Возможно, ограничение в движении с первых дней
жизни телят (клеточное содержание) обусловливает вначале задержку разрушения
утробных структур. В последующем утробные структуры так интенсивно разрушаются,
что созидание новых функциональных структур запаздывает и проявляется в «убыли»
костной массы. Такой адаптационный процесс можно рассматривать как задержку
функциональной перестройки.
5. ПРАКТИЧЕСКИЕ ПРЕДЛОЖЕНИЯ
При выращивании телят в условиях промышленных комплексов предлагается при
разработке технологии содержания учитывать их генетическую потребность в
движении с первого дня жизни.
СПИСОК ЛИТЕРАТУРЫ
1. Акаевский,
А. И. Анатомия домашних животных / А. И. Акаевский. - М.: Колос, 1971. - 581 с.
. Бамбуляк,
Н. Ф. Морфология и минеральный состав некоторых трубчатых костей свиньи при
различной двигательной активности: автореф. дис. .. .канд. вет. наук / Н. Ф.
Бамбуляк. - Киев, 1992. - 22 с.
. Гаврилин,
П. Н. Морфофункциональные особенности костной и иммунной систем телочек
новорожденного и молочного периодов при различной двигательной активности:
автореф. дис…. канд. вет. наук / П. Н. Гаврилин - Киев, 1992. - 20 с.
. Исаенков,
Е. А. Линейный рост скелета тазовой конечности в онтогенезе романовских овец /
Е. А. Исаенков // Влияние антропогенных факторов на структурные преобразования
органов, тканей, клеток человека и животных: Материалы 2-ой Всероссийской конф.
- Саратов, 1993, Ч. 2. - С. 23.
. Криштофорова,
Б. В. Морфофункциональный адаптогенез, периодизация роста и развития
млекопитающих и птиц различной локомоции / Б. В. Криштофорова, П. Н. Гаврилин
// Влияние антропогенных факторов на структурные преобразования органов,
тканей, клеток человека и животных: Материалы 2-ой Всероссийской конф. -
Саратов, 1993, Ч. 2. - С. 27.
. Криштофорова,
Б. В. Рост и развитие костномозгового участка диафиза трубчатых костей
конечности животных при различной двигательной активности / Б. В. Криштофорова
// Сб. науч. тр. Моск. вет. академии. - М., 1987. - С. 30 - 38.
. Криштофорова,
Б. В. Строение, рост и развитие суставных и метафизарных хрящей трубчатых
костей конечностей животных при различной двигательной активности / Б. В.
Криштофорова // Сб. науч. тр. Моск. вет. академии. - М., 1987. - С. 18 - 30.
. Мельник, К.
П. Локомоторный аппарат млекопитающих / К. П. Мельник, В. И. Клыков. - Киев:
Наукова думка, 1991. - 20 с.
. Слесаренко,
Н. А. Морфология костей 3-го пальца крупного рогатого скота при различных
условиях содержания и кормления / Н. А. Слесаренко // Сб. научн. тр. Московск.
вет. академии. - М., 1978. - Т.100. - С. 69-70.
. Харченко,
Л. Г. Морфометрические характеристики костей акроподия крупного рогатого скота
разной породности в зависимости от уровня двигательной активности / Л. Г.
Харченко / / Межвуз. сб. научн. трудов. - М., 1988. - С. 23-26.