Аеробне дихання
Аеробне дихання
На відміну від анаеробних процесів, в
аеробних реакціях катаболізму піруват окиснюється до кінцевих продуктів - СО2
і Н2О, що супроводжується вивільненням значно більшої енергії.
У аеробів піруват перетворюється в
двовуглецеву сполуку - ацетил-КоА під дією піруватдегідрогеназного комплексу:
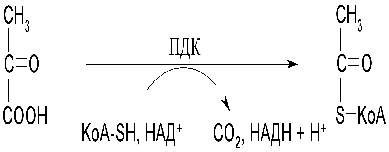
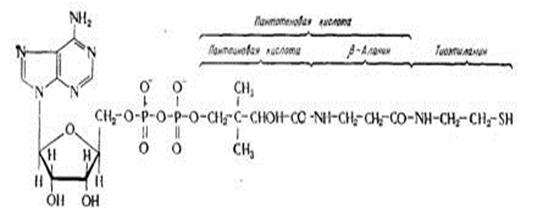
Кофермент А відноситься до групи мононуклеотидів, його будова:
Піруватдегідрогеназний комплекс локалізований в мітохондріях еукаріот
або в цитоплазмі прокаріот. Він досить складний, складається з 3-х різних
ферментів.
На наступній стадії клітинного дихання ацильні групи включаються в ЦТК
(цикл три карбонових кислот, цикл лимонної кислоти, цикл Кребса), в результаті якого
утворюється СО2 і атоми водню.
ЦТК в клітині має 2 функції:
1) Це досконалий клітинний
«котел», в якому здійснюється повне окиснння органічного субстрату і
відщеплення водню;
) ЦТК постачає клітині ряд
попередників для біосинтетичних процесів.
Енергетичним «паливом» для ЦТК
служать не тільки вуглеводи, а й жирні кислоти (після попередньої деградації до
ацетил-КоА), амінокислоти (після видалення аміногрупи).
В результаті утворюється 2 молекули
СО2
3 молекули НАДН2
молекула ФАДН2
Весь водень знаходиться на певних
переносниках, тепер потрібно його передати через інші переносники на
молекулярний кисень. 4 пари електронів від цих водневих атомів поступають в
дихальний ланцюг і забезпечують синтез АТФ у реакціях окисного фосфорилювання.
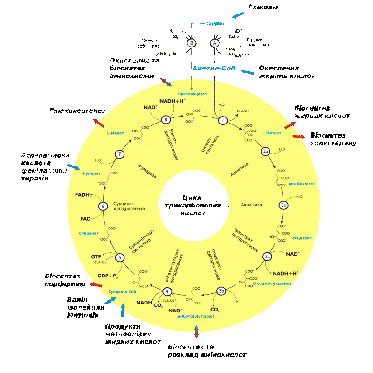
Рис.1 Цикл три карбонових кислот
Аконітаза перетворює лимонну кислоту в ізолимонну, яка не
відокремлюється від активного центра ферменту, α-кетоглутаратдегідрогеназа каталізує реакції,
аналогічні піруватдегідрогеназі, містить 3 ферменти та пов’язані з ними
кофактори: ТПФ, КоА, НАД, ФАД. Деякі інтермедіати ЦТК (α-кетоглутарат, сукцинат, оксалоацетат) можуть
бути використані як попередники амінокислот, що порушило би циклічність.
Існують спеціальні анаплеротичні реакції, які забезпечують поповнення пулу
попередників ЦТК (гліоксилатний шунт).
Дихальний ланцюг
В той час як більшість анаеробних організмів здатні синтезувати АТФ
тільки шляхом субстратного фосфорилювання, аероби здійснюють більш ефективну
регенерацію АТФ. Вони мають спеціальний апарат - дихальний ланцюг і
фермент АТФ-синтетазу.
Ці системи у прокаріот знаходяться в плазматичній мембрані, а у
еукаріот- на внутрішній мембрані мітохондрій. Електрони з відновлених
переносників (НАДН2, НАДФН2, ФАДН2) поступають
в дихальний ланцюг, де проходять через ряд етапів, опускаються поступово на
більш низькі енергетичні рівні і акцептуються сполуками - кінцевими акцепторами
електронів (О2 та інші). Перенос електронів призводить до суттєвої
зміни вільної енергії в системі. Ця вільна енергія переводиться у біологічну
форму, тобто АТФ. Компонентами дихального ланцюга є переносники білкової або
небілкової (хінони) природи з різноманітними простетичними групами, здатними
приєднувати і віддавати електрони або протони. Вони занурені у подвійний
ліпідний шар.
До найважливіших належать:
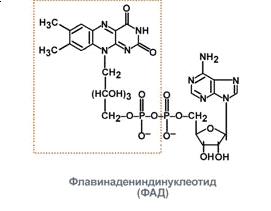
1. Флавопротеїни - ферменти, які
в якості простетичних груп мають ФМН або ФАД. Переносять Н+.
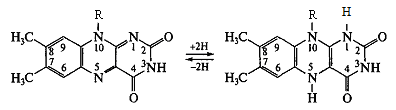
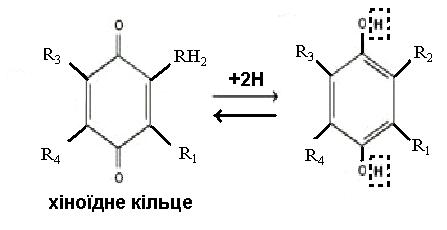
2. Залізосірковмісні білки (Fe-S-білки). Містять атоми Fe, зв’язані з неорганічною сульфідною
сіркою і сіркою цистеїну. Залізо змінює свою валентність Fe (II)→Fe (III). Переносять електрони.
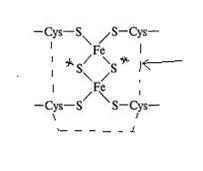
 -
неорганічна кислотолабільна сірка
-
неорганічна кислотолабільна сірка
3. Хінони. Переносять водень і
жиророзчинні сполуки. Убіхінон у грамнегативних, нафтохінон у грампозитивних.
4. Цитохроми - приймають участь
на заключному етапі ланцюга переносу електронів. Являють собою групу білків,
які містять залізопорфіринові простетичні групи (геми). Переносять тільки
електрони.
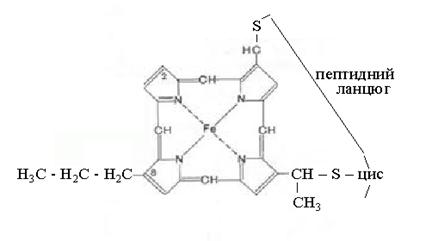
Fe2+  Fe3+ + e‾
Fe3+ + e‾
В мітохондріях еукаріот виявлено 5
цитохромів: b, c, c1, a, a3. Цитохроми забарвлені і
відрізняються спектрами поглинання і окисно-відновними потенціалами.
Найбільш поширеним та еволюційно
найдавнішим цитохромом, знайденим у всіх організмах з дихальним ланцюгом є
цитохром с. Кінцеві цитохроми (а+а3) передають електрони на
молекулярний кисень, являють собою власне цитохромоксидазу, в реакційному
центрі якої, крім двох гемів, містяться 2 атоми міді. Цитохромоксидаза
переносить на О2 4 електрона.
О2 + 4Fe2+ → 2O2 - + 4Fe3+
Якщо переноситься тільки 2 електрона,
то утворюється перекис водню, який потім руйнується каталазою або пероксидазою.
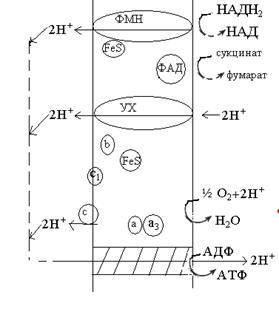
Дихальний ланцюг включає не менше 15
різноманітних переносників, розташованих у певній послідовності.

Компоненти дихального ланцюга мають свої значення окисно-відновних
потенціалів. Виявлені інгібітори, які специфічно діють на
певні ділянки дихального ланцюга (червоним на малюнку).
Топографія компонентів дихального ланцюга:
Основне призначення ЛПЕ - запасання енергії, яка звільнюється в процесі
електронного переносу шляхом трансформування її в хімічну енергію фосфатних зв’язків.
Протони при електронному транспорті переносяться через мембрану в
одному напрямі, і утворюють градієнт концентрації Н+ по обидві
сторони мембрани. Перенос протонів та електронів забезпечується розташуванням
переносників, а також тим фактом, що мембрана для протонів непроникна.
В мітохондріях на 3-х ділянках окиснювального ланцюга відбувається
виділення протонів у зовнішнє середовище (про це свідчить підкислення
середовища у дослідах на ізольованих мітохондріях). Відповідно 3 реакції ведуть
до утворення ∆µН+. Перша локалізована на початку дихального
ланцюга і пов’язана з функціонуванням НАД(Ф)·Н2-дегідрогенази.
Другий генератор ∆µН+ визначається здатністю
убіхінона переносити водень. Останній локалізований в кінці дихального ланцюга
і пов’язаний з активністю цитохромоксидази.
Виведені назовні протони повертаються в середину бактеріальної клітини
(або в мітохондріальний матрикс еукаріот) за градієнтом концентрації через
спеціальні канали-пори в молекулі АТФ-синтетази, а вільна енергія, що при цьому
виділяється, використовується для синтезу АТФ.
Транспортування кожної пари електронів по ЛПЕ від НАДН2 до О2
супроводжується синтезом 3-х молекул АТФ у трьох точках спряження окиснення і
фосфорилювання. Всього ж при повному окисненні глюкози, враховуючи енергетичний
вихід гліколізу, окисненні пірувату та ЦТК утворюється 38 молекул АТФ.
Неповні окиснення
Більшість аеробних організмів окиснює
органічні речовини в процесі дихання до СО2 і Н2О.
Оскільки в молекулі СО2 досягається найвищий ступінь окиснення
вуглецю, то в такому випадку говорять про повне окиснення. При неповному
окисненні в якості продуктів обміну виділяються частково окиснені органічні
сполуки.
Кінцевими продуктами «неповних
окиснень» можуть бути кислоти: оцтова, глюконова, фумарова, лимонна, молочна та
ряд інших сполук. Оскільки ці продукти подібні до продуктів, що утворюються в
результаті бродіння, то неповні окиснення називають іноді «окисним бродінням»
або «аеробною ферментацією». Слова «бродіння» і «ферментація» в даному випадку
відображають швидше технологічний аспект.
Утворення оцтової кислоти
Оцет отримують із вина або
водно-спиртової суміші. Основна проблема - аерація. Розрізняють 3 основних
способи вирощування оцтовокислих бактерій:
поверхневий
фіксований (на носіях -
виноградний жом, букові стружки)
глибинний (посилена аерація)
Оцтовокислі бактерії - це
грамнегативні рухомі палички, облігатні аероби, належать до родів Acetobacter, Gluconobacter: A. aceti, A. xylinum, G.oxydans, G. melanogenum. Вони здатні окиснювати етанол,
гліцерол, глюкозу у відповідні кислоти (оцтову, гліцеринову, глюконову).
Вторинні спирти окиснюються ними до кетонів, наприклад, сорбіт (сорбітол) до
сорбози.
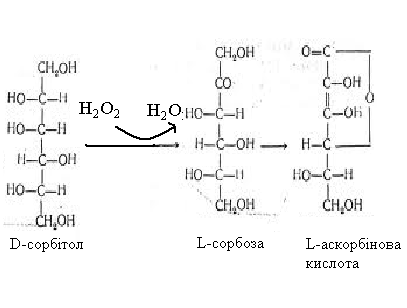
Таке окиснення є одним з етапів на шляху отримання аскорбінової кислоти
з глюкози. D-сорбітол можна отримати шляхом
електролітичного відновлення D-глюкози.
Інші органічні кислоти (глюконова, фумарова, лимонна, щавлева) можна
синтезувати із проміжних продуктів ЦТК грибами родів Mucor, Aspergillus, Rhizopus, а також дріжджами роду Candida.
Добре вивчений і налагоджений промисловий випуск лимонної кислоти
культурою гриба Aspergillus niger (lim Fe ↑вихід).
Щавлева кислота утворюється в результаті гідролізу щавлевооцтової
кислоти під дією оксалоацетатгідролази:
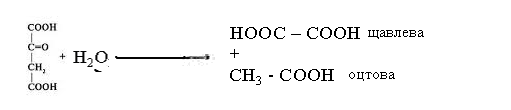
Її утворенню сприяє лужний рН середовища.
Глюконова кислота є продуктом ферментативного окиснення глюкози грибною
глюкозооксидазою. Глюконолактон, який при цьому утворюється, за допомогою
глюконолактонази (або не ферментативним шляхом) перетворюється в глюконову
кислоту.
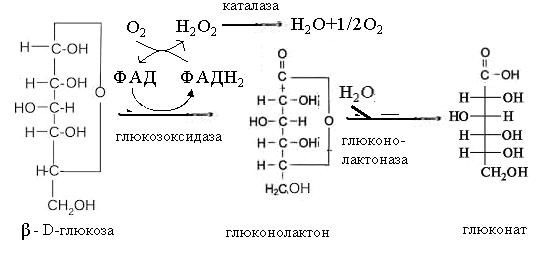
Відновлена глюкооксидаза переносить водень на кисень повітря з
утворенням перекису водню, який під дією каталази може розщеплюватись на воду і
кисень.
Попередником ітаконової кислоти є цис-аконітова кислота. Під час
декарбоксилювання подвійний зв’язок переміщується з положення 2,3 у положення
3,4:
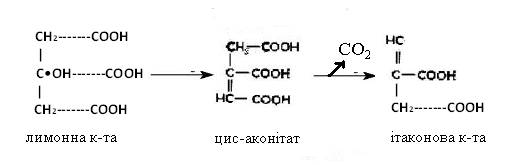
Окиснення одновуглецевих сполук
Серед великої різноманітності
мікроорганізмів, які отримують енергію в процесі аеробного дихання, особливу
фізіологічну групу складають метилотрофи, здатні окиснювати одновуглецеві
субстрати (метан, метанол, форміат, метиламіни). Вони не здатні використовувати
вуглеводи з довгим ланцюгом.
До метилотрофних мікроорганізмів
належать:
) бактерії: архебактерії
фотосинтезуючі бактерії
) дріжджі: Pichia, Candida,
Hansenula, Rhodotorula, Torulopsis
) міцеліальні гриби: Gliocladium, Paecilomyces,
Penicillum, Trichoderma.
Прокаріотичні метилотрофи поділяють
на 2 фізіологічні групи:
облігатні: використовують
лише С1-субстрати, як правило, метан
факультативні: ростуть на
середовищах із іншими субстратами, але не на метані
Реакції засвоєння метану протікають
через такі послідовні стадії:
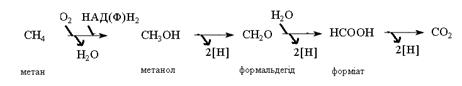
На першому етапі відбувається окиснення метану СН4 до
метилового спирту СН3ОН ферментом метанмонооксидазою, причому
цей процес пов’язаний з використанням лише одного атома кисню, тоді
як другий відновлюється до води.
Метанол СН3ОН до формальдегіду НСОН бактерії окиснюють за
допомогою метанолдегідрогенази. Електрони від цього окиснення
направляються в ЛПЕ для синтезу АТФ.
Синтез клітинних сполук (асиміляція) здійснюється із формальдегіду -
високо реакційної та токсичної сполуки. Саме тому ферменти його утворення та
асиміляції локалізовані поза цитоплазмою. Бактеріальна
метанолдегідрогеназа прикріплена ззовні до периплазматичної мембрани. У
дріжджів ферменти катаболізму метанолу локалізовані у спеціальних органелах-
пероксисомах.
Метилотрофні бактерії реалізують 3 циклічні шляхи асиміляції
одновуглецевих субстратів: сериновий, рибулозомонофосфатний та
рибулозодифосфатний:
1) з формальдегіду і гліцину
утворюється серин → фосфоенолпіруват
) формальдегід +
рибулозо-5-фосфат → гексулозодифосфат
Біолюмінесценція
Серед бактерій поширене явище
світіння - біолюмінесценція. Світіння є окиснювальним процесом, який
зумовлюється ферментом люциферазою. Більшість люмінесцентних бактерій належить
до родів Vibrio, Photobacterium, Alteromonas, Beneckia. Вони переважно є галофільними
грамнегативними факультативними анаеробами. За типом живлення -
хемоорганогетеротрофи. Бувають різної форми - коки, палички, прямі і зігнуті.
Більшу частину бактерій виділено з морської води. Вони також зустрічаються у
тілі і м’язах дрібних ракоподібних
планктонних організмів. У деяких риб є спеціальні органи для
фотобактерій-симбіонтів.
Світіння відбувається лише у
присутності кисню, пов’язане з окисними реакціями, які супроводжуються не
синтезом АТФ, а виділенням (емісією) синьо-зеленого світла (470-500 нм).
Бактеріальна люцифераза складається з
двох різних субодиниць - α і β, α-субодиниця виконує
каталітичну функцію, β-субодиниця - регуляторну.
Джерелом енергії для світіння є
реакція перетворення альдегіду з довгим вуглецевим ланцюгом до відповідної
жирної кислоти за участю відновленого флавінмононуклеотиду.
(ФМН) • Н2 + О2 + R - COH → ФМН + Н2O + R - COOH + hυ
Нітратне дихання
Для багатьох прокаріотів кінцевим
акцептором електронів при анаеробному диханні є нітрати. Ці бактерії
називаються денітрифікуючими, а нітратне дихання - дисиміляційною
нітратредукцією або денітрифікацією.
Денітрифікуючі бактерії -
хемоорганогетеротрофи. В анаеробних умовах вони здійснюють окиснення органічних
субстратів звичайним каталітичним шляхом (гліколіз, ЦТК - в аеробів, бродіння -
в анаеробів), при цьому Н+ та електрони поступають на дихальний
ланцюг.
Як кінцевий акцептор водню ці
бактерії використовують нітрати, які відновлюються через нітрити до
молекулярного азоту:
NO3- → NO2-
→ N2 (або N2O)
Нітратредуктаза А нітритредуктаза
Процес нітратного дихання каталізують
2 мембранзв’язні ферменти - нітратредуктаза
А і нітритредуктаза. Обидва індукуються лише в анаеробних умовах
(для більшості денітрифікаторів потрібна наявність NO3- в середовищі, для деяких не
потрібна). Серед денітрифікуючих бактерій є такі, що використовують як кінцевий
акцептор водню не лише нітрати, але й нітрити та закис азоту.
До денітрифікаторів відносять:
Pseudomonas aeruginosa
P. fluorescens
Paracoccus denitrificanslicheniformisdenitrificans
Розвиток денітрифікуючих бактерій у
ґрунті приводить до пониження вмісту доступних для рослин форм азоту. Оранка
ґрунту сприяє створенню аеробних умов і пригніченню денітрифікації
(репресія синтезу нітрит- і нітратредуктази). Проте це єдиний процес, в якому
зв’язаний N перетворюється в молекулярний N2. Крім дисиміляційної нітрат редукції
бактерії використовують нітрат для синтезу азотовмісних компонентів клітини.
Цьому передує відновлення нітрату до амоніаку.
NO3- → NO2- → HNO → NH2OH → NH3
Процес отримав назву асиміляційна
нітратредукція і здійснюється бактеріями як в аеробних, так і в
анаеробних умовах. Відновлення нітрату при асиміляційній нітратредукції
каталізує фермент нітратредуктаза В, яка знаходиться в цитоплазмі та індукується
тільки тоді, коли єдиним джерелом азоту є нітрат. Завершується відновний процес
за участю нітритредуктази.
Деякі ентеробактерії (enteron - кишечник) (роди Enterobacter, Escherichia) здатні відновлювати нітрат тільки
до нітриту, і тільки ця ланка спряжена з переносом електронів і одержанням
енергії. Нітрит далі може відновлюватись до NH3+ в реакціях асиміляційної
нітратредукції з виділенням в середовище NH4+. Процес одержав назву амоніфікація
нітрату.
Споживання багатих нітратами
продуктів або води приводить до відновлення їх мікрофлорою кишечника. При цьому
нітрит, що потрапив у кров, спричиняє утворення метгемоглобіну. В
результаті еритроцити втрачають здатність транспортувати О2.
Сульфатне дихання
Сульфатне дихання (дисиміляційна
сульфатредукція) здійснюється у бактерій виключно в анаеробних умовах
SO42- + 8[H+]
→ H2S + 2H2O + 2OH-
або
4H2 + SO42- → S2- + 4H2O
Цей процес спряжений з утворенням АТФ
в реакціях окисного фосфорилювання на ЛПЕ.
Багато мікроорганізмів здатні
утилізувати сульфати в асиміляційних процесах, зокрема синтезах сірковмісних
метаболітів. Відновлення сульфатів до сульфідів здійснюється в реакціях асиміляційної
сульфатредукції. Обидва процеси - дисиміляційна і асиміляційна
сульфатредукція - проходять через стадію утворення сульфіту.
Desulfotomaculum, Desulfovibrio, Desulfosarcina, які заселяють різні екологічні ніші
(грунти, водойми, рубець жуйних). Сульфатредукуючі бактерії є облігатними анаеробами.
Донорами водню для них служать прості низькомолекулярні сполуки, які утворилися
при анаеробному розкладі біомаси, в основному целюлози: лактат, ацетат,
пропіонат, бутират, форміат, етанол, вищі жирні кислоти, молекулярний водень.
Асиміляційна сульфатредукція призводить до утворення H2S. Різниця: 1) більш поширена; 2)
відновлена сірка йде на біосинтетичні процеси; 3) перший етап подібний, при
асиміляційній сульфатредукції йде ще одна реакція активації активованого (АТФ)
сульфату.
Як акцептор водню деякі бактерії
можуть використовувати елементарну сірку, відновлюючи її до H2S у процесах окиснення органічних
субстратів (сірчане дихання).
В наш час добре вивчений Desulfuromonas acetoxidans. Цю бактерію, яка пересувається за
допомогою латерально розташованого джгутика, можна виділити з морської води і
отримати нагромаджувальну культуру на середовищі, що містить мінеральні солі,
ацетат і сірчаний цвіт (порошкоподібну сірку) при дотриманні анаеробних умов.
Сульфатредукувальні бактерії поширені
в прибережних мулах, промислових стоках, багатих сульфатами. Саме за рахунок їх
життєдіяльності тут нагромаджується сірководень. Його вміст різко зростає при
підвищенні концентрації органічних речовин у стічних водах.
Сульфатредукувальні бактерії є
причиною нагромадження значної кількості сірководню у Чорному морі.
За участю сульфатредукувальних
бактерій утворені поклади сірки біогенного походження (Роздольське на
Львівщині; Техас, Луізиана, Мексика). Анаеробна корозія заліза та будівельних
матеріалів, що містять сульфати, також результат їх життєдіяльності. Отримати
нагромаджувальні культури суьфатредукувальних бактерій можна, якщо додати в
середовище лактат, сульфат, залізний цвях (скріпку) для створення низького
окисно-відновного потенціалу (в результаті катодної поляризації, анаеробних
умов), (Е0’= -200 мB).
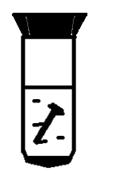
Рис.2 Отримання нагромаджу вальної культури сульфатредукувальних
бактерій. Інокулянт - сірководневий мул
Карбонатне дихання
В клітинах деяких анаеробних бактерій
кінцевим акцептором водню є СО2. До них належать метаногенні
бактерії (Archaeobacteria).
Морфологічно вони є паличкоподібні (Methanobacterium), коки (Methanococcus), сарцини (Methanosarcina), спірили (Methanospirillum), неправильної форми (Methanococсeoides). Вони мешкають (зустрічаються) на
площах, зайнятих тундрою і болотами (метан - болотний газ), рисові поля, осади
на дні ставків і озер; лимани, марши, естуарії; відстійники очисних споруд,
шлунки (рубці) жуйних тварин (більше ніж 109 куо/мл).
Метанутворюючі бактерії становлять останню
ланку анаеробного харчового ланцюга. В ньому приймають участь:
) бактерії, які зброджують
целюлозу до сукцинату, пропіонату, бутирату, лактату, ацетату, спиртів, СО2,
Н2;
) ацетогенні бактерії, які
зброджують вищеназвані продукти до ацетату, форміату, СО2, Н2;
) метаногени, що
використовують ці розчини в якості субстратів.
4Н2 + СО2 →
СН4 + 2Н2О
За типом живлення метаногени є
переважно гетеротрофами, хоча описані види, які використовують СО2 як
єдине джерело вуглецю.
В різних місцях утворюється метан,
відбувається також утворення оцтової кислоти. Підкислення середовища у
відстійниках пов’язане з діяльністю бактерій, які перетворюють СО2 і
Н2 в оцтову кислоту згідно рівнянню:
Н2 + 2СО2 →
СН3 - СООН + 2Н2О
Ацетогенні бактерії - це анаеробні
хемолітоавтотрофи; грампозитивні палички
Clostridium aceticum.
thermoaceticumwoodi.
дихання біолюмінесценція
окиснення нітратний
Висновки
1. Мікроорганізми здатні
здійснювати окиснення органічних сполук в анаеробних та аеробних умовах.
. Найбільш оптимальний спосіб
окиснення - цикл Кребса. Електрони із сполук метаболізму поступають в ланцюг
переносу електронів, що дозволяє клітині запасати енергію у вигляді АТФ.
. Мікроорганізми здатні
здійснювати неповне окиснення органічних сполук, в результаті чого утворюються
оцтова, глюконова, фумарова, лимонна, молочна, ітаконова кислоти.
. Прикладом неповного
окиснення є люмінесценція.
. У випадку якщо кінцевим
акцептором електронів є не кисень, а інші органічні сполуки, то розрізняють
сульфатне, карбонатне, нітратне дихання.
Список літератури
1. Prescott L. M., Harley J. P., Klein D. A.
Microbiology. Third edition. - Dubuque: WCB, 1996