Механизм действия ацетилхолина
Государственный
Университет Молдовы
Департамент
индустриальной и экологической химии
Факультет
химии и химических технологий
РЕФЕРАТ
На
тему: "Механизм действия ацетилхолина"
Выполнила студентка
Симион Диана ТРСМ-4
rus
Руководитель: T.
Cotelea
Кишинёв
г.
Холинэргические рецепторы (ацетилхолиновые
рецепторы) - трансмембранные рецепторы, лигандом которых является ацетилхолин.
Ацетилхолин служит нейротрансмиттером как в
пре-, так и в постганглионарных синапсах парасимпатической системы и в
преганглионарных симпатических синапсах, в ряде постганглионарных симпатических
синапсов, нервно-мышечных синапсах (соматическая нервная система), а также в
некоторых участках ЦНС. Нервные волокна, выделяющие ацетилхолин из своих
окончаний, называются холинергическими.
Синтез ацетилхолина происходит в цитоплазме
нервных окончаний; запасы его хранятся в виде пузырьков в пресинаптических
терминалях. Возникновение пресинаптического потенциала действия ведет к
высвобождению содержимого нескольких сотен пузырьков в синаптическую щель.
Ацетилхолин, выделяющийся из этих пузырьков, связывается со специфическими
рецепторами на постсинаптической мембране, что повышает ее проницаемость для
ионов натрия, калия и кальция и приводит к появлению возбуждающего постсинаптического
потенциала. Действие ацетилхолина ограничивается путем его гидролиза с помощью
фермента ацетилхолинэстеразы.
Специфические холинергические рецепторы с
фармакологичесой точки зрения разделяются на никотиновые (Н-рецепторы) и
мускариновые (М-рецепторы).
Ацетилхолиновый никотиновый рецептор является
одновременно и ионным каналом, т.е. относится к рецепторам-каналоформером,
тогда как ацетилхолиновый мускариновый рецепторотносится к классу серпентиновых
рецепторов, осуществляющих передачу сигнала через гетеротримерные G - белки.
Холинорецепторы вегетативных ганглиев и
внутренних органов различаются.
На постганглионарных нейронах и клетках
мозгового вещества надпочечников располагаются N-холинорецепторы
(чувствительные к никотину), а на внутренних органах - М-холинорецепторы
(чувствительные к алкалоиду мускарину). Первые блокируются ганглиоблокаторами,
вторые - атропином.
М-холинорецепторы подразделяются на несколько
подтипов:
М1-холинорецепторы располагаются в ЦНС и,
возможно, на нейронах парасимпатическихганглиев;
М2-холинорецепторы - на гладких и сердечной
мышцах и клетках железистого эпителия.
Селективным стимулятором М2-холинорецепторов
служит бетанехол. Пример селективного блокатора М1-холинорецепторов -
пирензепин. Этот препарат резко подавляет выработку HCl в желудке.
Стимуляция М2-холинорецепторов через Gi-белок
приводит к ингибированию аденилатциклазы, а стимуляция М2-холинорецепторов
через Gq-бeлок - к активации фосфолипазы С и образованию ИФ3 и ДАГ (рис. 70.5).
Стимуляция М3-холинорецепторов также приводит к
активации фосфолипазы С. Блокатором этих рецепторов служит атропин.
Методами молекулярной биологии были выявлены и
другие подтипы М-холинорецепторов, однако они пока недостаточно изучены.
Ацетилхолин (acetylcholine,
Ach) [лат. acetum - уксус, греч. chole - желчь и лат. -in(e) - суффикс,
обозначающий "подобный"] - уксусный эфир холина (см. Холин),
нейромедиатор, передающий нервное возбуждение через синаптическую щель в
парасимпатической нервной системе; синтезируется в тканях при участии
холинацетилазы, гидролизуется ферментом ацетилхолинэстеразой. А. обнаружен
также в составе некоторых растительных ядов. Впервые выделен из спорыньи в 1914
г. Г. Дейлом. За установление роли А. в передаче нервного импульса он совместно
с О. Леви получил Нобелевскую премию за 1936 г.
Ацетилхолин действует через холинергические
окончания нервов, концевые мионевральные пластинки и другие холинорецепторы.
Находясь в белково-липоидном комплексе (прекурссор), ацетилхолин освобождается
при электрическом и нервном возбуждении. Исследованиями Palay в 1956 г. с
помощью электронной микроскопии показано накопление капель жидкости в порах
синапса, часть из которых лопалась при прохождении нервного импульса. Полагают,
что секретируемая жидкость - ацетилхолин (теория пи-ноцитоза). Выделяясь в
холинергических субстанциях сердца, ацетилхолин воздействует на сопредельные
клеточные мембраны. Согласно современным взглядам, мебрана несет в покое
определенный электрический заряд, обусловленный перераспределением иона К.
Концентрация калия в покое много выше внутри клетки, нежели снаружи. Для
натрия, наоборот, концентрация снаружи клетки велика, а внутри - мала.
Концентрация ионов натрия внутри клетки остается постоянной благодаря активному
удалению его из клетки во время процесса, называемого "натриевым
насосом". Калий же проникает на поверхность клетки, оставляя более
массивный анион внутри ее, поэтому наружная поверхность клетки получает избыток
положительных зарядов, внутренняя - отрицательных. Чем больше катионов калия
выйдет из клетки, тем выше оказывается заряд ее мембраны, и наоборот - при
замедлении выхода калия потенциал мембраны снижается. Прямые измерения
потенциала покоя показали, что он равен в миокарде желудочков и предсердий
приблизительно 90 мв, в синусовом узле 70 мв. Если по какой-либо причине
потенциал мембраны снизится до 50 мв, резко меняются свойства мембраны и она
пропускает внутрь клетки значительное количество ионов натрия. Тогда внутри
клетки превалируют положительные ионы и мембранный потенциал меняет свой знак.
Перезарядка (деполяризация) мембраны вызывает электрический потенциал действия.
После сокращения восстанавливаются концентрации калия и натрия, свойственные
состоянию покоя (реполяризация).
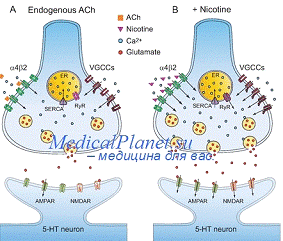
Установлено, что холинергические
(парасимпатомиметические, парасимпатотропные, трофотропные) реакции возникают
при действии ацетилхолина (или других соединений холина) на холинорецепторы,
субклеточные образования, клетки, ткани, органы или организм в целом. Помимо
своего основного (холинергического) действия, ацетилхолин вызывает освобождение
калия, связанного белками, повышает или снижает проницаемость биологических
мембран, принимает участие в регуляции избирательной проницаемости эритроцитов,
изменяет активность отдельных дыхательных ферментов, влияет на активность
катепсинов, на обновляемость фосфатной группы в фосфолипидах, на метаболизм
макроэр-гических фосфорных соединений, повышает устойчивость отдельных тканей и
организма в целом к гипоксии. Коштоянц высказал предположение, что, осуществляя
медиаторное действие, ацетилхолин вступает в круг тканевых биохимических
превращений.
Нормальный механизм автоматизма в сердце основан
на спонтанном уменьшении потенциала синусового узла до -50 мв (генераторный
потенциал). Это происходит в синусовом узле посредством особого метаболического
процесса, основанного на снижении проницаемости мебраны для калия. Ацетилхолин,
напротив, специфически увеличивает проницаемость для К мембраны синусового
узла, тем самым повышая выход К и препятствуя развитию генераторного
потенциала. Поэтому частота сердечных сокращений падает. Если же концентрацию
ацетилхолина увеличить еще более, то генераторный потенциал развивается
настолько медленно, что мембраны синусового узла теряют способность развивать
потенциал действия (аккомодация мембраны). Наступает остановка сердца.
Повышение проницаемости для калия под влиянием ацетилхолина обусловливает более
быстрый процесс восстановления потенциала покоя мембраны (реполяризацию).
Введенный ацетилхолин разносится кровью не всегда равномерно. Поэтому в
предсердии этот процесс ускоренной реполяризации также может идти неравномерно,
что при сохранившемся возбуждении синусового узла проявляется как трепетание и
мерцание предсердий. Желудочки сердца, лишенные холинергических окончаний,
остаются нечувствительными к ацетилхолину. Активация центров автоматизма II
порядка (пучка Гиса) связана со свойством волокон Пуркинье развивать спонтанную
деполяризацию так же, как это происходит в синусовом узле.
Немедиаторное действие ацетилхолина в целостном
организме представляет один из наименее изученных и наиболее спорных разделов
гуморально-гормональной регуляции функций. Установлено, что холинергические (
парасимпатомиметические, парасимпатотропные, трофо-тропные) реакции возникают
при действии ацетилхолина (или других соединений холина) на холинорецепторы,
субклеточные образования, клетки, ткани, органы или организм в целом. Помимо
своего основного (холинергического) действия, ацетилхолин вызывает освобождение
калия, связанного белками, повышает или снижает проницаемость биологических
мембран, принимает участие в регуляции избирательной проницаемости эритроцитов,
изменяет активность отдельных дыхательных ферментов, влияет на активность
катепсинов, на обновляемость фосфатной группы в фосфолипидах, на метаболизм
макроэргических фосфорных соединений, повышает устойчивость отдельных тканей и
организма в целом к гипоксии. Коштоянц высказал предположение, что, осуществляя
медиаторное действие, ацетилхолин вступает в круг тканевых биохимических
превращений. [1] А торможение действия ацетилхолина в какой-то мере
функционально эквивалентно увеличению концентрации дофамина. [5]
Биохимический эффект
ацетилхолина заключается в том, что его присоединение к рецептору открывает
канал для прохождения ионов Na и К через мембрану клетки, что ведет к
деполяризации мембраны. Блокирование действия ацетилхолина чревато серьезными
проблемами, вплоть до смертельного исхода. Именно в этом заключается биохимическое
действие нейротоксинов. Ниже показаны структуры двух наиболее сильных
нейро-токсинов - хистрионикотоксина и хлорида D-тубокурарина. Как и
ацетил-холин, молекула D-тубокурарина содержит аммониевые фрагменты. Она
блокирует место присоединения ацетилхолина к рецептору, исключает передачу
нервного сигнала, предотвращает перенос ионов через мембрану. Создается
ситуация, называемая параличом живой системы. [9]
Влияние ацетилхолина на сердце.
Холинергические механизмы. На наружной мембране
кардиомиоцитов представлены, в основном, мускаринчувствительные (М-)
холинорецепторы. Доказано наличие в миокарде и никотинчувствительных (N-)
холинорецепторов, однако их значение в парасимпатических влияниях на сердце
менее ясно. Плотность мускариновых рецепторов в миокарде зависит от
концентрации мускариновых агонистов в тканевой жидкости. Возбуждение
мускариновых рецепторов тормозит активность пейсмекерных клеток синусного узла
и в то же время увеличивает возбудимость предсердных кардиомиоцитов. Эти два
процесса могут привести к возникновению предсердных экстрасистол в случае
повышения тонуса блуждающего нерва, например ночью во время сна. Таким образом,
возбуждение М-холинорецепторов вызывает снижение частоты и силы сокращений
предсердий, но повышает их возбудимость.
Ацетилхолин угнетает проводимость в
атриовентрикулярном узле. Это связано с тем, что под влиянием ацетилхолина
возникает гиперполяризация клеток атриовентрикулярного узла вследствие усиления
выходящего калиевого тока. Таким образом, возбуждение мускариновых холинорецепторов
оказывает противоположное, по сравнению с активацией B-адренорецепторов,
действие на сердце. При этом снижается частота сердечных сокращений, угнетается
проводимость и сократимость миокарда, а также потребление миокардом кислорода.
Возбудимость предсердий в ответ на применение ацетилхолина возрастает, тогда
как возбудимость желудочков, напротив, уменьшается.
Ацетилхолин относится к числу самых важных
нейромедиаторов мозга. Самая выдающаяся роль ацетилхолина реализуется в
нейромышечной передаче, где он является возбуждающим трансмиттером. Известно,
что ацетилхолин может оказывать как возбуждающее, так и ингибирующее действие.
Это зависит от природы ионного канала, который он регулирует при взаимодействии
с соответствующим рецептором.
Нейротрансмиттер ацетилхолин высвобождается из
везикул в пресинаптических нервных терминалях и связывается как с никотиновыми
рецепторами, так и мускариновыми рецепторами на поверхности клетки. Эти два
типа ацетихолиновых рецепторов значительно отличается как по структуре, так и
по функциям.
Ацетилхолин - уксуснокислый эфир холина,
является медиатором в нервно-мышечных соединениях, в пресинаптических
окончаниях мотонейронов на клетках Реншоу, в симпатическом отделе вегетативной
нервной системы - во всех ганглионарных синапсах, в синапсах мозгового вещества
надпочечников и в постганглионарных синапсах потовых желез; в парасимпатическом
отделе вегетативной нервной системы - также в синапсах всех ганглиев и в
постганглионарных синапсах эффекторных органов . В ЦНС ацетилхолин обнаружен во
фракциях многих отделов мозга, иногда в значительных количествах, однако
центральных холинэргических синапсов обнаружить не удалось.
Ацетилхолин синтезируется в нервных окончаниях
из холина, который поступает туда с помощью неизвестного пока транспортного
механизма. Половина поступившего холина образуется в результате гидролиза ранее
высвободившегося ацетилхолина, а остальная часть, по-видимому, поступает из
плазмы крови . Фермент холин-ацетилтрансфераза образуется в соме нейрона и
примерно за 10 дней транспортируется по аксону к пресинаптическим нервным
окончаниям. Механизм поступления синтезированного ацетилхолина в синаптические
пузырьки пока неизвестен.
По-видимому, лишь небольшая часть (15-20%)
запаса ацетилхолина, который хранится в пузырьках, составляет фракцию
немедленно доступного медиатора, готовую к высвобождению - спонтанно или под
влиянием потенциала действия .
Депонированная фракция может мобилизоваться
только после некоторой задержки. Это подтверждается, во-первых, тем, что вновь
синтезированный ацетилхолин высвобождается примерно вдвое быстрее, чем ранее
присутствовавший, во-вторых, при нефизиологически высоких частотах стимуляции
количество ацетилхолина, высвобождаемое в ответ на один импульс, падает до
такого уровня, при котором количество ацетилхолина, высвобождаемое в течение
каждой минуты, остается постоянным. После блокады поглощения холина
гемихолинием из нервных окончаний высвобождается не весь ацетилхолин.
Следовательно, должна быть третья, стационарная фракция, которая, возможно, не
заключена в синаптические пузырьки. Видимо, между этими тремя фракциями может
происходить обмен. Гистологические коррелянты этих фракций еще не выяснены, но
предполагают, что пузырьки, расположенные около синаптической щели, составляют
фракцию немедленно доступного медиатора, тогда как остальные пузырьки
соответствуют депонированной фракции или ее части.
Ацетилхолин служит медиатором всех
преганглионарных нейронов, постганглионарных парасимпатических нейронов,
постганглионарных симпатических нейронов, иннервирующих мерокриновые потовые
железы, и соматических нервов . Он образуется в нервных окончаниях из
ацетил-КоA и холина под действием холинацетилтрансферазы . В свою очередь,
холин активно захватывается пресинаптическими окончаниями из внеклеточной
жидкости . В нервных окончаниях ацетилхолин хранится в синаптических пузырьках
и высвобождается в ответ на поступление потенциала действия и вход
двухвалентных ионов кальция. Ацетилхолин относится к числу самых важных
нейромедиаторов мозга.
Если концевая пластинка
подвергается действию ацетилхолина в течение нескольких сотен миллисекунд, то
мембрана, деполяризованная вначале, постепенно реполяризуется, несмотря на
постоянное присутствие ацетилхолина, то есть постсинаптические рецепторы
инактивируются. Причины и механизм этого процесса пока не изучены.
Обычно действие ацетилхолина на
постсинаптическую мембрану продолжается всего 1-2 мс, потому что часть
ацетилхолина диффундирует из области концевой пластинки, а часть гидролизуется
ферментом ацетилхолинэстеразой (т.е. расщепляется на неэффективные компоненты
холин и уксусную кислоту). Ацетилхолинэстераза в больших количествах имеется в
концевой пластинке (так называемая специфическая или истинная холинэстераза),
однако холинэстеразы имеются также в эритроцитах (также специфические) и в
плазме крови (неспецифические, т.е. расщепляют и другие эфиры холина). Поэтому
ацетилхолин, который диффундирует из области концевой пластинки в окружающее
межклеточное пространство и поступает в кровоток, тоже расщепляется на холин и
уксусную кислоту. Большая часть холина из крови снова поступает в
пресинаптические окончания.
Действие ацетилхолина на
постсинаптическую мембрану постганглионарных нейронов может быть воспроизведено
никотином, а на эффекторные органы - мускарином (токсин мухомора). В связи с
этим возникла гипотеза о наличие двух типов макромолекулярных рецепторов
ацетилхолина, и его действие на эти рецепторы называется никотиноподобным или
мускариноподобным. Никотоноподобное действие блокируется основаниями, а
мускариноподобное - атропином.
Вещества, действующие на клетки
эффекторных органов так же, как холинэргические постганглионарные
парасимпатические нейроны, называются парасимпатомиметическими, а вещества,
ослабляющие действие ацетилхолина - парасимпатолитическими.
Список литературы
холинергический
рецептор ацетилхолин нейрон
1. Харкевич Д.А. Фармакология. М.:
ГЭОТАР-МЕД, 2004
. Зеймаль Э.В., Шелковников С.А. -
Мускариновые холинорецепторы
. Сергеев П.В., Галенко-Ярошевский
П.А., Шимановский Н.Л., Очерки биохимической фармакологии, М., 1996.
. Хуго Ф. Нейрохимия, М,
"Мир", 1990 г.
. Сергеев П.В., Шимановский Н.Л.,
В.И. Петров, Рецепторы, Москва - Волгоград, 1999 г.