Биологическое действие лецитина:
Лецитин является основным питанием
всей нервной системы. Входит в состав оболочек нервных волокон. Обеспечивает
нормальный обмен фосфолипидов. При его нехватке появляется раздражительность,
нервные срывы, усталость.
Лецитин является важнейшим
строительным материалом для мозга. Рассеянный склероз, понижение памяти и
другие нарушения мозговой деятельности обычно сопровождаются пониженным
содержанием лецитина в организме человека. Нехватка лецитина у детей приводит к
рассеянному вниманию, низкой способности к обучению.
Лецитин снижает уровень холестерола
и концентрацию жирных кислот в крови, помогает очистить от холестероловых
бляшек стенки сосудов.
Лецитин улучшает функцию печени и почек,
препятствует образованию желчных камней.
Лецитин - помогает усвоению
жирорастворимых витаминов А, Д, Е и К, необходимых для питания всех клеток
организма. Лецитин помогает организму вырабатывать энергию. Его нехватка часто
наблюдается при больших физических нагрузках.
3. Хромопротеины.
Примеры белков этого класса, их биологическая роль
Хромопротеины состоят из простого
белка и связанного с ним окрашенного небелкового компонента, откуда и произошло
их название (от греч. chroma - краска). Среди хромопротеинов различают
гемопротеины (содержащие в качестве простетической группы железо),
магний-порфирины и флавопротеины (содержащие производные изоаллоксазина).
Хромопротеины наделены рядом
уникальных биологических функций: они участвуют в таких фундаментальных
процессах жизнедеятельности, как фотосинтез, дыхание клеток и целостного
организма, транспорт кислорода и углерода, окислительно-восстановительные
реакции, свето- и цветовосприятие и др.
Таким образом, хромопротеины играют
исключительно важную роль в процессах жизнедеятельности. Достаточно, например,
подавить дыхательную функцию гемоглобина путем введения оксида углерода или
утилизацию (потребление) кислорода в тканях путем введения синильной кислоты
или ее солей (цианидов), ингибирующих ферментные системы клеточного дыхания,
как моментально наступает смерть организма.
Хромопротеины являются непременными
и активными участниками аккумулирования солнечной энергии в зеленых растениях.
Хлорофилл (магний-порфирин) вместе с белком обеспечивает фотосинтетическую
активность растений, катализируя расщепление молекулы воды на водород и
кислород (с поглощением солнечной энергии); гемопротеины (железо-порфирины)
катализируют обратную реакцию - образование молекулы воды, связанное с
освобождением энергии.
Гемопротеины
К группе гемопротеинов относятся
гемоглобин и его производные, миоглобин, хлорофиллсодержащие белки и ферменты
(вся цитохромная система, каталаза и пероксидаза). Все они содержат в качестве
небелкового компонента структурно сходные железо (или магний) - порфирины, но
различные по составу и структуре белки, обеспечивая тем самым разнообразие их
биологических функций.
Гемоглобин в качестве белкового
компонента содержит глобин, а небелкового - гем. Видовые различия гемоглобина
обусловлены глобином, в то же время гем одинаков у всех видов гемоглобина.
Гемоглобину принадлежит уникальная
роль в транспорте кислорода от легких к тканям и углекислого газа от тканей к
легким.
В молекуле гемоглобина взрослого
человека, обозначаемого НbА (от англ. adult - взрослый), содержатся четыре
полипептидные цепи, которые вместе составляют белковую часть молекулы - глобин.
Из многообразия производных
гемоглобина, представляющих несомненный интерес для врача, следует прежде всего
указать на оксигемоглобин - НbО2 - соединение молекулярного
кислорода с гемоглобином. Кислород присоединяется к гему гемоглобина при помощи
координационных связей железа, причем валентность железа не меняется и железо
остается двухвалентным. Такой гемоглобин называют оксигенированным.
Помимо кислорода, гемоглобин легко
соединяется и с другими газами, в частности с СО, N0 и др. Так, при отравлении
оксидом углерода гемоглобин прочно с ним связывается с образованием
карбоксигемоглобина (НbСО). При этом из-за высокого сродства к СО гемоглобин
теряет способность связывать кислород и наступает смерть от удушья,
недостаточного снабжения тканей кислородом. Однако повышение парциального
давления кислорода во вдыхаемом воздухе приводит к частичному вытеснению СО из
связи с гемоглобином.
При отравлении оксидами азота, парами
нитробензола и другими соединениями часть гемоглобина окисляется в
метгемоглобин (НbОН), содержащий трехвалентное железо. Поскольку метгемоглобин
также теряет способность к переносу кислорода от легких к тканям, то и в
случаях метгемоглобинемии (вследствие отравления окислителями) в зависимости от
степени отравления может наступить смерть от недостатка кислорода.
Флавопротеины содержат прочно связанные с белком простетические группы,
представленные изоаллоксазиновыми производными - окисленными флавинмононуклеотидом
(ФМН) и флавинадениндинуклеотидом (ФАД). Флавопротеины входят в состав
оксидоредуктаз - ферментов, катализирующих оксилительно-восстановительные
реакции в клетке. Некоторые флавопротеины содержат ионы металлов. Типичными
представителями флавопротеинов, содержащих также негемовое железо, являются
ксантиноксидаза, альдегидоксидаза, СДГ, дигидрооротатдегидрогеназа,
ацил-КоА-дегидрогеназа и транспортирующий электроны флавопротеин. На долю двух
последних приходится до 80% митохондриальных флавопротеинов, выполняющих важную
роль в биоэнергетике клетки. Негемовое железо связывается с белковым
компонентом, отличающимся от гемсодержащих хромопротеинов. Железо ковалентно
связано с атомом серы остатка цистеина в белке. При кислотном гидролизе такого
белка освобождаются железо и Н2S. Несмотря на структурные отличия от
цитохромов, негемовые флавопротеины обладают аналогичной функцией в транспорте
электронов благодаря способности переходить из окисленного в восстановленное
состояние.
4. Витамин Е, биологическая
роль. Признаки гипо- и авитаминоза. Пищевые источники. Лекарственные препараты
витамина Е
Витамин Е по химической природе
является метильным производным токола.
аминокислота хромопротеин лецитин
пептид
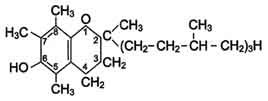
К витамину Е относят семейство 8
типов токоферолов. Витамин был выделен из зародышей пшеничных зерен.
Механизм действия. Витамин Е действует как антиоксидант, предотвращая образование в
клетках токсичных продуктов перекисного окисления липидов.
Роль в обмене веществ
Токоферол является природным
липофильным антиоксидантом, так как он способен инактивировать свободные
радикалы. Из-за гидрофобности антиоксидантные свойства витамина Е проявляются,
прежде всего, в липидном слое биомембран и таким образом предотвращается развитие
цепи перекисного окисления и липидов обеспечивается стабильность биологических
мембран. Токоферол отдает атом водорода свободному радикалу пероксида липида
(ROO*), восстанавливая его до гидропероксида (ROOH) и таким образом
останавливается развитие ПОЛ. Свободный радикал витамина Е, образовавшийся в
результате реакции, стабилен и не способен участвовать в развитии цепи.
Токоферол повышает биологическую активность витамина А, защищая от окисления
его ненасыщенную боковую цепь.
Гиповитаминоз Е.
Недостаточность витамина Е в отличие
от других витаминов не отличается специфическими клиническими проявлениями. Это
обусловлено тем, что при недостатке токоферола активируются
свободно-радикальные процессы, особенно в липидах биомембран, что проявляется
разнообразными нарушениями. Наиболее характерные признаки гиповитаминоза Е
наблюдается у недоношенных детей, когда поражаются биомембраны, клеток и,
прежде всего, эритроцитов, что приводит к развитию гемолитической анемии. Таким
образом, гиповитаминоз Е характеризуется патологией мембран, одним из признаков
которой является гемолиз.
Суточная потребность в витамине - 5
мг.
Источник витамина - растительные
масла, особенно оливковое, облепиховое масла, сардины, салат, семена злаков
(проросшие семена пшеницы), сливочное масло, яичный желток.
Показания к применению препаратов
витамина Е
Применение препаратов витамина Е
показано:
) недоношенным детям, при
искусственном вскармливании;
) при проведении сеансов
кислородотерапии под избыточным давлением для профилактики токсического
действия кислорода;
) при комплексном лечении различных
анемий, особенно связанных с разрушением эритроцитов;
) при лечении различных гипотрофий и
дистрофий миокарда;
) для стимуляции синтеза антител и
неспецифических факторов резистентности к инфекции;
) при лечении некоторых форм
бесплодия и при патологии беременности (способствует нормальному развитию и
функции плаценты).
Витамин Е применяют также при
профилактике и лечении рахита (усиливает лечебное действие витаминов Д и С) и
для предотвращения осложнений при передозировке витаминов Д и А.
Препараты витамина Е
Витрум витамин Е. Мощный природный антиоксидант. Нейтрализует свободные радикалы,
превращая их в безвредные вещества, которые выводятся из организма. Замедляет
процессы старения, обладает омолаживающим зффектом; предупреждает развитие
атеросклероза, снижает риск развития ишемической болезни сердца,
инсулинозависимость, усиливает иммунитет, снижает утомляемость, уменьшает
влияние канцерогенных веществ, восстанавливает репродуктивные функции, повышает
мужскую потенцию, снимает усталость. Является прекрасным средством для лечения
ожогов. Витрум Витамин Е выпускается в капсулах.
Доппельгерц витамин Е
форте. Растительный препарат витамина Е,
полученный из молодых ростков злаков. Применяется при гиповитаминозах, в период
выздоровления после заболеваний, протекавших с лихорадочным синдромом, при
высоких физических нагрузках, в пожилом возрасте, при заболеваниях связочного
аппарата и мышц. Эффективен при климактерических вегетативных нарушениях, дегенеративных
и пролиферативных изменениях суставов и связочного аппарата позвоночника и
крупных суставов.
Супер формула
антиоксидантов. Эта специальная
суперформула содержит повышенные дозы важнейших антиоксидантов (витамина А - 20
000 МЕ, витамина Е - 200 МЕ, витамина С - 200 мг) В сочетании с минералами
(селен - 100 мг, цинк - 12 мг, кальций - 15 мг) и комплексом продления жизни.
Также добавлены концентраты моркови, апельсинов и миндаля. Препарат выпускается
в капсулах, которые рекомендуется применять по 2 в день.
Гипервитаминоз Е
Гипервитаминоз Е. Длительный прием витамина Е в дозах от 100 до 800 мг не вызывает
побочных реакций. Возможны проявления токсического действия витамина Е при
парентеральном введении больших доз (креатинурия, потенцирование коагулопатии
при недостаточности витамина К и ухудшение заживления ран), возможно также
снижение эффективности фагоцитоза, повышение риска септических осложнений.
5. Что такое ингибиторы?
Охарактеризуйте ингибиторы непрямого действия
Действие ферментов можно полностью
или частично подавить (ингибировать) определенными химическими веществами
(ингибиторами). По характеру действия ингибиторы могут быть обратимыми и
необратимыми. В основе этого деления лежит прочность соединения ингибитора с
ферментом.
Обычно для установления того, каким
именно ингибитором является исследуемое вещество, реакционную смесь, содержащую
все компоненты ферментативной реакции, подвергают диализу или проводят
гельхроматографирование с целью отделить фермент от других компонентов. В результате
этого сравнительно непрочная связь обратимого ингибитора с ферментом
разрывается и активность фермента полностью восстанавливается. Если
взаимодействие фермента шло с необратимым ингибитором, то фермент-ингибиторный
комплекс обычно не распадается и активность фермента не восстанавливается.
Проявление ингибирующего действия на иммобилизованные ферменты существенно
отличается от их действия в гомогенных системах.
Другой способ деления ингибиторов
основывается на характере места их связывания. Одни из них связываются с
ферментом в активном центре, а другие - в удаленном от активного центра месте.
Они могут связывать и блокировать функциональную группу молекулы фермента,
необходимую для проявления его активности. При этом они необратимо, часто
ковалентно, связываются с ферментом или фермент - субстратным комплексом и
необратимо изменяют нативную конформацию. Это, в частности, объясняет действие
Hg2+, Pb2+, соединений мышьяка. Ингибиторы такого рода
могут быть полезны при изучении природы ферментативного катализа. Например,
диизопропилфторфосфат ингибирует ферменты, имеющие серин в активном центре.
Таким ферментом является ацетилхолинэстераза, катализирующая следующую реакцию:
|
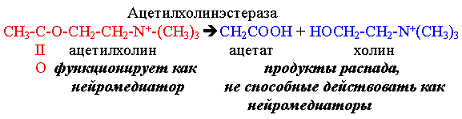
|
Реакция происходит каждый раз после
проведения нервного импульса, прежде чем второй импульс будет передан через
синапс. Диизопропилфторфосфат - одно из отравляющих веществ
нервно-паралитического действия, так как приводит к утрате способности нейронов
проводить нервные импульсы.
|
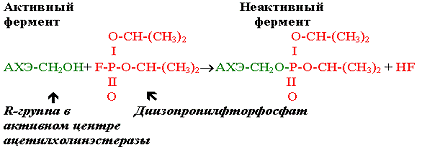 Действие диизопропилфторфосфата на фермент
Действие диизопропилфторфосфата на фермент
|
Действие необратимого ингибитора
обычно возрастает с увеличением времени реагирования, поэтому оно может
характеризоваться константой скорости взаимодействия фермента с ингибитором:
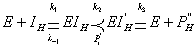
Кора надпочечников человека
синтезирует 3 основных класса стероидных гормонов, которые обладают широким
спектром физиологических функций. Они включают глюкокортикоиды,
минералокортикоиды и адренальные андрогены. Эти гормоны образуются в различных
слоях надпочечников из холестерола липопротеинов низкой плотности или ацетил
коэнзима А, или эфиров холестерина из внутриклеточных депо.

Основным и наиболее активным
глюкокортикоидом, образующимся в организме человека, является гидрокортизон
(кортизол), другие, менее активные, представлены кортизоном, кортикостероном,
11-дезоксикортизолом, 11-дегидрокортикостероном.
Механизм действия глюкокортикоидов
на молекулярном уровне до конца не выяснен. Считают, что действие
глюкокортикоидов на клетки-мишени осуществляется, главным образом, на уровне
регуляции транскрипции генов. Оно опосредуется взаимодействием глюкокортикоидов
со специфическими глюкокортикоидными внутриклеточными рецепторами
(альфа-изоформа). Эти ядерные рецепторы способны связываться с ДНК и относятся
к семейству лиганд-чувствительных регуляторов транскрипции. Рецепторы
глюкокортикоидов обнаружены практически во всех клетках.
После проникновения через мембрану
внутрь клетки глюкокортикоиды связываются с рецепторами, что приводит к
активации комплекса. При этом олигомерный белковый комплекс диссоциирует -
отсоединяются белки теплового шока (Hsp90 и Hsp70) и иммунофилин. В результате
этого рецепторный белок, входящий в комплекс в виде мономера, приобретает
способность димеризоваться. Вслед за этим образовавшиеся комплексы
«глюкокортикоид + рецептор» транспортируются в ядро, где взаимодействуют с участками
ДНК, расположенными в промоторном фрагменте стероид-отвечающего гена - т.н.
глюкокортикоид-отвечающими элементами (glucocorticoid response element, GRE) и
регулируют (активируют или подавляют) процесс транскрипции определенных генов
(геномный эффект). Это приводит к стимуляции или супрессии образования м-РНК и
изменению синтеза различных регуляторных белков и ферментов, опосредующих
клеточные эффекты.
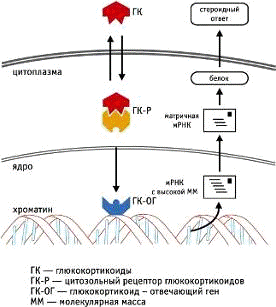
Рис. 1. Молекулярный механизм
действия глюкокортикоидов:
ГК - глюкокортикоид; ГК-R -
цитозольный рецептор глюкокортикоидов; ГК-ОГ - глюкокортикоид-отвечающий ген;
ММ - молекулярная масса
Кроме того, недавно открыт еще один
механизм действия глюкокортикоидов, связанный с влиянием на транскрипционную
активацию цитоплазматического ингибитора NF-kB - IkBa.
Однако ряд эффектов глюкокортикоидов
(например быстрое ингибирование глюкокортикоидами секреции АКТГ) развиваются
очень быстро и не могут быть объяснены экспрессией генов (т.н. внегеномные
эффекты глюкокортикоидов). Такие свойства могут быть опосредованы
нетранскрипторными механизмами, либо взаимодействием с обнаруженными в
некоторых клетках рецепторами глюкокортикоидов на плазматической мембране.
Полагают также, что эффекты глюкокортикоидов могут реализовываться на разных
уровнях в зависимости от дозы. Например, при низких концентрациях
глюкокортикоидов (>10-12 моль/л) проявляются геномные эффекты (для их
развития требуется более 30 мин), при высоких - внегеномные.
Глюкортикоиды вызывают множество
эффектов, т.к. оказывают влияние на большинство клеток организма. Они обладают
противовоспалительным, десенсибилизирующим, противоаллергическим и
иммунодепрессивным действием, противошоковыми и антитоксическими свойствами.
Глюкокортикоиды влияют на альтеративную и экссудативную фазы воспаления,
препятствуют распространению воспалительного процесса.
ГКС играют важную роль в регуляции
основных метаболических процессов. Существует дозозависимый эффект их влияния
на углеводный обмен. Они стимулируют печеночный глюконеогенез и гликогенолиз,
влияют на синтез других гормонов вовлеченных в глюконеогенез (глюкагон,
адреналин) и ингибируют утилизацию глюкозы, индуцируя периферическую
инсулинрезистентность. Одним из основных проявлений этого процесса является
рост гликемии.
ГКС оказывают влияние на жировой
метаболизма через активацию липолиза и ингибирование захвата глюкозы
адипозоцитами. Влияние их на белковый обмен проявляется в подавлении синтеза
белка и активации протеолиза в мышцах. Освобожденные при этом аминокислоты
служат субстратом для глюконеогенеза.
Основными минералокортикоидами,
секретируемыми корой надпочечников, являются альдостерон и
11-дезоксикортикостерон (ДОК). Альдостерон и другие минералокортикоиды влияют
на транспорт ионов в эпителиальных клетках, действуя на уровне Ка+-К+-АТФазы.
Основной их эффект заключается в поддержании нормальных концентраций Na+
и К+, а также объема внеклеточной жидкости. Проникая через клеточную
мембрану, они взаимодействуют с минералокортикоидными рецепторами цитозоля.
Активный стероид-рецепторный комплекс перемещается в ядро клетки, где
модулирует транскрипцию ряда генов, приводя к изменению синтеза определенных
РНК и соответствующих белков.
Минералокортикоиды являются жизненно
важными гормонами, гибель организма после удаления надпочечников можно
предотвратить, вводя гормоны извне. Минералокортикоиды усиливают воспаление и
реакции иммунной системы. Избыточная их продукция ведет к задержке в организме
натрия и воды, отекам и повышению артериального давления, потере калия и
водородных ионов, вследствие чего возникают нарушения возбудимости нервной
системы и миокарда. Недостаток альдостерона у человека сопровождается
уменьшением объема крови, гиперкалиемией, гипотензией, угнетением возбудимости
нервной системы.
7. Что такое
антиоксиданты? Механизм действия каталазы и пироксидазы
Антиоксиданты (антиокислители) -
соединения, способные тормозить, уменьшать интенсивность свободнорадикального
окисления, нейтрализовывать свободные радикалы путем обмена своего атома
водорода (в большинстве случаев) на кислород свободных радикалов. Антиоксиданты
могут быть природного (биоантиоксиданты) и синтетического происхождения.
Вещества этой группы имеют подвижный атом водорода и поэтому реагируют со
свободными радикалами, а также катализаторами свободнорадикального окисления и,
прежде всего, с ионами металлов переменной валентности. Подвижность атома
водорода обусловлена нестойкой связью с атомами углерода (С-Н) или серы (S-Н).
В результате взаимодействия возникают малоактивные радикалы самого
антиоксиданта (они не способны к продолжению цепи), гидроперекиси разлагаются
без диссоциации на активные радикалы (под действием серосодержащих соединений),
образуются комплексоны с металлами переменной валентности. Образующиеся
свободные радикалы антиоксидантов малоактивны и выводятся из организма в виде
молекулярных соединений - продуктов взаимодействия с другими антиоксидантами
(токоферолами, хинонами, витаминами группы К, серосодержащими соединениями).
Ряд антиоксидантов не обрывает, а замедляет продолжение цепи, т.е. обладает
пролонгирующим действием. Несмотря на малую активность радикалов
антиоксидантов, их накопление в клетках нежелательно.
Ферментные антиоксиданты
катализируют реакции, в которых активные формы кислорода и некоторые другие
окислители восстанавливаются до стабильных и нетоксичных продуктов.
Перекись водорода разрушается двумя
классами родственных ферментов, катализирующих ее двухэлектронное
восстановление до Н2О
Н2О2 →
2Н2О + О2
и использующих в качестве донора
электронов Н2О2 в случае каталазы или различные
органические соединения в случае пероксидазы. При низком содержании Н2О2
органические пероксиды преимущественно катализируются пероксидазой. Однако, при
высоких концентрациях Н2О2 работают каталазы.
8. Какое количество АТФ
будет образовываться при окислении 3 молекул ацетил-КоА в цикле Кребса путем
субстратного фосфорилирования? Где протекают реакции этого цикла, и каково его
значение для организма?
В матриксе митохондрий ацетил-КоА
постепенно окисляется до углекислого газа с одновременным отщеплением дегидрогеназами
атомов Н, которые в виде НАДН и ФАДН2 становятся субстратами
(донорами электронов) в реакции дыхательной цепи.
Цикл Кребса протекает в матриксе
митохондрий и является участком аэробного окисления ацетил-КоА по схеме:
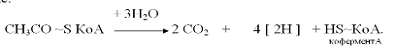
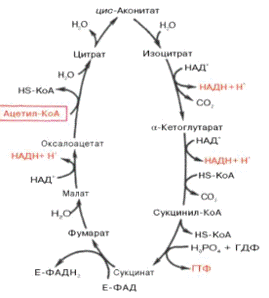
В ходе окисления б-кетоглутарата
синтезируется одна молекула ГТФ (субстратное фосфорилирование), что равносильно
одной молекуле АТФ.
Баланс энергии при окислении
ацетил-КоА в цикле Кребса:
СН3ЇСО~КоА→ 6 НАД×Н2 + 2 ФАД×Н + 2 АТФ.
Следовательно, в ЦТК при окислении 2
молекул ацетил-КоА образуется 2 АТФ в результате субстратного фосфорилирования.
Тогда при окислении 3 молекул ацетил-КоА будет образовываться 3 молекулы ГТФ,
что равносильно образованию 3 молекул АТФ.
Цикл трикарбоновых кислот занимает
важное место в процессе обмена веществ. При окислении ацетилКоА в нем
образуется ряд промежуточных продуктов, которые приводят к синтезу других
важных соединений: щавелево-уксусная и б-кетоглутаровая кислоты, подвергаясь
восстановительному аминированию, образуют аспарагиновую и глутаминовую кислоты,
сукцинилКоА идет на синтез порфиринов. В цикле Кребса осуществляется
взаимосвязь между обменом углеводов, органических кислот, жиров, аминокислот и
белков в клетках живых организмов. Таким образом, ЦТК - это амфиболический путь
метаболизма. Функции его связаны не только с катаболическими, но и с
анаболическими процессами, для которых он поставляет вещества -
предшественники.
9. Напишите цепь митохондриального
окисления для лактата и глицеролфосфата. Укажите участки выделения АТФ
Глицеролфосфат является субстратом
окислительного фосфорилирования, протекающего в матриксе митохондрий.
Восстановленный флавопротеин
(фермент-ФАДН2) вводит на уровне KoQ приобретенные им электроны в
цепь биологического окисления и сопряженного с ним окислительного
фосфорилирования, а диоксиаце-тонфосфат выходит из митохондрий в цитоплазму и
может вновь взаимодействовать с цитоплазматическим НАДН + Н+.
Лактат образуется из пирувата в
качестве конечного продукта анаэробного гликолиза. Пируват + NADH+ЛДГ H+
= Лактат + NAD.
При большой потребности в АТФ и
недостаточном поступлении кислорода пируват за счет анаэробного гликолиза
восстанавливается в молочную кислоту (лактат), которая диффундирует в
кровь
Глюкоза + 2АДФ + 2Фн = 2
лактат + 2АТФ + 2H2O.
Образование молочной кислоты
является тупиковой ветвью метаболизма, но не конечным продуктом обмена веществ.
Под действием лактатдегидрогеназы молочная кислота окисляется снова, образуя
пируват, который участвует в дальнейших превращениях. В норме получившийся
лактат переводится в пируват, который превращается в ацил-кофермент А
(ацил-КоА), который, в свою очередь, переносится через митохондриальную
мембрану и в митохондриальном матриксе превращается в ацетил-КоА. Ацетил-КоА
вступает в цикл Кребса, в процессе которого происходит накопление протонов
водорода в виде соединений НАД·Н и ФАД·Н2.
Далее, на внутреннем слое митохондриальной мембраны через так называемую
дыхательную цепь с участием энергии накопленных протонов происходит синтез
молекул АТФ.
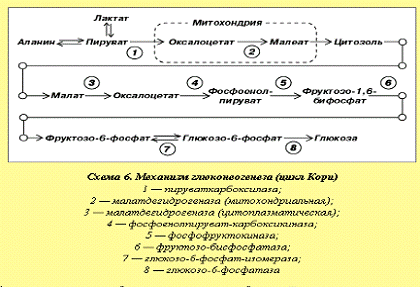
Образование глюкозы из лактата
осуществляется в печени (цикл Кори). При этом из лактата образуется пируват и
далее Г-6-Ф, который превращается затем в гликоген или глюкозу в зависимости от
состояния обмена веществ в организме (схема 6). Лактат может превращаться в
глюкозу и в волокнах белых мышц, но при этом необходимы высокая концентрация
лактата и высокое соотношение концентраций АТФ и АДФ. Глюконеогенез в
незначительной степени происходит и в почках при более низкой концентрации
лактата. Образование глюкозы из лактата - энергоемкий процесс, требующий
достаточного количества АТФ. В печени соотношение АТФ и АДФ составляет около
10, в других тканях оно ниже. В цикле Кори из лактата образуется пируват, далее
- Г-6-Ф, который превращается в глюкозу, поступающую в кровоток и
преобразующуюся в мышцах в гликоген.
10. Переваривание
сахарозы, лактозы и мальтозы в ЖКТ. Всасывание образовавшихся продуктов
Усвоение пищевых олиго- и
полисахаридов начинается с их гидролитического (под действием воды) расщепления
в процессе пищеварения до моносахаридов.
Гидролитическое расщепление
углеводов в процессе пищеварения происходит под действием ферментов гликозидаз,
расщепляющих 1-4 и 1-6 гликозидные связи в молекулах сложных углеводов. Простые
углеводы пищеварению не подвергаются, может только происходить брожение
некоторой части их в толстом кишечнике под действием ферментов микроорганизмов.
К гликозидазам относятся амилаза слюны,
поджелудочного и кишечного соков, мальтаза слюны и кишечного сока, конечная
декстриназа, сахараза и лактаза кишечного сока. Гликозидазы активны в
слабощелочной среде и угнетаются в кислой среде, за исключением амилазы слюны,
которая катализирует гидролиз полисахаридов в слабокислой среде и теряет
активность при увеличении кислотности.
В ротовой полости начинается
пищеварение крахмала под воздействием амилазы слюны, которая расщепляет 1-4
гликозидные связи между остатками глюкозы внутри молекул амилозы и
амилопектина. При этом образуются дектстрины и мальтоза. В слюне содержится в
небольших количествах и мальтаза, гидролизующая мальтозу до глюкозы. Другие
дисахариды во рту не расщепляются.
Большая часть молекул полисахаридов
не успевает гидролизоваться во рту. Смесь крупных молекул амилозы и
амилопектина с более мелкими - декстринами - мальтозой, глюкозой, поступает в
желудок. Сильно кислая среда желудочного сока угнетает ферменты слюны, поэтому
дальнейшие превращения углеводов происходят в кишечнике, сок которого содержит
бикарбонаты, нейтрализующие соляную кислоту желудочного сока.
Амилазы поджелудочного и кишечного
соков более активны, чем амилаза слюны. В кишечном соке содержится также
конечная декстриназа, гидролизующая 1-6 связи в молекулах амилопектина и
декстринов. Эти ферменты завершают расщепление полисахаридов до мальтозы. В
слизистой оболочке кишечника вырабатываются также ферменты, способные
гидролизовать дисахариды: мальтаза, лактаза, сахараза. Под воздействием
мальтазы мальтоза расщепляется на две глюкозы, сахароза под воздействием
сахаразы - на глюкозу и фруктозу, лактаза расщепляет лактозу на глюкозу и
галактозу.
Под действием ферментов
микроорганизмов продукты распада сложных углеводов могут подвергаться брожению,
в результате чего образуются органические кислоты, СО2, СН4
и Н2. Схема превращений углеводов в пищеварительной системе:
Образовавшиеся в результате
гидролиза углеводов моносахариды по своей структуре одинаковы у всех живых
организмов. Среди продуктов пищеварения преобладает глюкоза (60%), она же
является главным моносахаридом, циркулирующим в крови. В кишечной стенке
фруктоза и галактоза частично превращаются в глюкозу, так что содержание ее в
крови, оттекающей от кишечника, больше, чем в его полости.
Образовавшиеся в результате гидролиза
углеводов моносахариды по своей структуре одинаковы у всех живых организмов.
Среди продуктов пищеварения преобладает глюкоза (60%), она же является главным
моносахаридом, циркулирующим в крови. В кишечной стенке фруктоза и галактоза
частично превращаются в глюкозу, так что содержание ее в крови, оттекающей от
кишечника, больше, чем в его полости.
Всасывание моносахаридов - активный
физиологический процесс, протекающий с затратой энергии. Ее обеспечивают
окислительные процессы, происходящие в клетках кишечной стенки. Моносахариды
получают энергию, взаимодействуя с молекулой АТФ в реакциях, продуктами которых
являются фосфорные эфиры моносахаридов. При переходе из кишечной стенки в кровь
фосфорные эфиры расщепляются фосфатазами, и в кровоток поступают свободные
моносахариды. Поступление их из крови в клетки различных органов также
сопровождается их фосфорилированием.
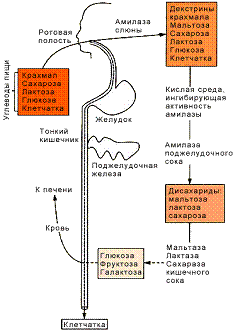
Рис. 2. Переваривание углеводов в
ЖКТ
Литература
1. Филиппович Ю.Б.
Основы биохимии. - М.: Высшая школа. - 1985.
2. Кнорре Д.Г.,
Мызина С.Д. Биологическая химия. - М.: Высшая школа. - 1996.
. Биохимия.
Сборник задач и упражнений /Под.ред. Кучеренко Н.Е., Бабенюк Ю.Д., Васильев
А.Н. и др.-К.: Выща шк. - 1988.
. Филиппович
Ю.Б., Севастьянова Г.А., Щеголева Л.И. Упражнения и задания по биологической
химии. - М.: Просвещение. - 1986.
. Ленинджер А.
Биохимия. - М.: Мир. - 1999.
. Березов Т.Т.,
Коровкин Б.Ф. Биологическая химия.-М.: Медицина. - 1998.
. Р. Марри, Д.
Греннер, П. Мейес, В. Родуэлл Биохимия человека. - М.: Мир. - Т. 1,2. - 1993.
. Степанов В.М.
Молекулярная биология. Структура и функции белков. - М.: Высшая школа. - 1996.