Трансформация бактерий как основа генной инженерии и молекулярного клонирования
Министерство образования и науки РФ
Федеральное государственное
автономное образовательное учреждение высшего профессионального образования
ДАЛЬНЕВОСТОЧНЫЙ ФЕДЕРАЛЬНЫЙ
УНИВЕРСИТЕТ
РЕФЕРАТ
Трансформация бактерий как основа
генной инженерии и молекулярного клонирования
Выполнила студентка 3 курса
Группы Б8308а
Огнева З. В.
Проверила:
Владивосток 2013
Оглавление
Введение
. Генная инженерия. Молекулярное клонирование
. Вектор
.1 Плазмиды
.2 Бактериофаги
.3 Космиды
. Escherichia coli (E. coli) - основная модель
биотехнологических манипуляций
. Основные этапы трансформации E. Coli
. Практическое применение трансформированных микроорганизмов
Список литературы
Введение
Основой микробиологической, биосинтетической промышленности является
бактериальная клетка. Необходимые для промышленного производства клетки
подбираются по определённым признакам, самый главный из которых - способность
производить, синтезировать, при этом в максимально возможных количествах,
определённое соединение - аминокислоту или антибиотик, стероидный гормон или
органическую кислоту. Иногда надо иметь микроорганизм, способный, например,
использовать в качестве «пищи» нефть или сточные воды и перерабатывать их в
биомассу или даже вполне пригодный для кормовых добавок белок. Иногда нужны
организмы, способные развиваться при повышенных температурах или в присутствии
веществ, безусловно, смертельных для других видов микроорганизмов.
Задача получения таких промышленных штаммов очень важна, для их видоизменения
и отбора разработаны многочисленные приёмы активного воздействия на клетку - от
обработки сильнодействующими ядами, до радиоактивного облучения. Цель этих
приёмов одна - добиться изменения наследственного, генетического аппарата
клетки. Их результат - получение многочисленных микробов-мутантов, из сотен и
тысяч которых учёные потом стараются отобрать наиболее подходящие для той или
иной цели. Создание приёмов химического или радиационного мутагенеза было
выдающимся достижением биологии и широко применяется в современной
биотехнологии.
Но их возможности ограничиваются природой самих микроорганизмов. Они не
способны синтезировать ряд ценных веществ, которые накапливаются в растениях,
прежде всего в лекарственных и эфирномасличных. Не могут синтезировать
вещества, очень важные для жизнедеятельности животных и человека, ряд
ферментов, пептидные гормоны, иммунные белки, интерфероны да и многие более
просто устроенные соединения, которые синтезируются в организмах животных и
человека. Разумеется, возможности микроорганизмов далеко не исчерпаны. Из всего
изобилия микроорганизмов использованного наукой, и особенно промышленностью,
лишь ничтожная доля.
1. Генная
инженерия. Молекулярное клонирование
Молекулярное клонирование (molecular cloning
or gene cloning) - клонирование молекул ДНК (в том числе генов,
фрагментов генов, совокупностей генов, ДНК-последовательностей, не содержащих
гены), другими словами - наработка большого количества идентичных ДНК-молекул с
использованием живых организмов. Благодаря фундаментальным биологическим
открытиям XIX-XX-го веков, а именно: открытию клеточного строения тканей,
открытию структуры клеточного ядра, хромосом, ДНК, генов, - стало возможным то,
что ныне носит название молекулярного клонирования. Это технология клонирования
наименьших биологических объектов - молекул ДНК, их частей и даже отдельных
генов.
Генетическая инженерия (генная инженерия) - совокупность приёмов, методов и
технологий получения рекомбинантных РНК и ДНК, выделения генов из организма
(клеток), осуществления манипуляций с генами и введения их в другие организмы.
Генетическая инженерия не является наукой в широком смысле, но является
инструментом биотехнологии, используя методы таких биологических наук, как
молекулярная и клеточная биология, цитология, генетика, микробиология,
вирусология.
Процессы генной инженерии микроорганизмов и молекулярного клонирования
имеют одинаковое начало, но в итоге получается разный результат - в одном
случае наработка белка, а в другом - нуклеиновых кислот. Тем не менее, для этих
манипуляций необходимо трансформировать бактериальные клетки.
Для получения генно-модифицированного организма, ДНК (обычно тем или иным
способом измененную) вводят в вектор (например, в бактериальную плазмиду или
геном бактериофага). Размножаясь, бактерии и фаги многократно увеличивают
количество введенной ДНК, в точности сохраняя её структуру, следовательно
увеличивают количество продуцируемых мРНК и белка. Чтобы затем выделить большое
количество продукта необходимо выделение и размножение бактериального или
фагового клона, содержащего необходимые молекулы ДНК. Для облегчения селекции
бактериальных клонов в плазмиды обычно вводят ген резистентности к антибиотику,
чаще всего ампициллину, в присутствии которого погибают все бактерии, не
имеющие клонируемой плазмиды. Такое клонирование необходимо для изучения
биологических молекул, их идентификации, решения вопросов клонирования тканей и
др.
Получение трансгенов стало возможным только после выделения
высокоспецифичных бактериальных ферментов, которые узнают определенные
последовательности оснований в двухцепочечной молекуле ДНК и расщепляют обе
цепи. Эти ферменты называются рестрицирующими эндонуклеазами типа II.
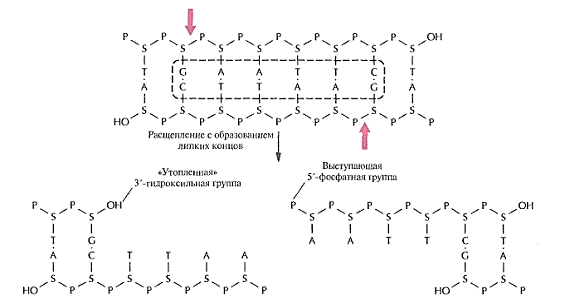
Одна из первых рестрицирующих эндонуклеаз типа II была выделена из
бактерии Escherichia coli и получила название EcoR. Этот фермент узнает
участок ДНК, содержащий специфическую палиндромную последовательность
(последовательность-перевертыш, идентичную в обеих цепях при прочтении в
направлении 5'-»3') из шести пар оснований и вносит разрыв между остатками
гуанина и аденина в каждой цепи. Разрывы в цепи ДНК располагаются наискось друг
от друга, в результате чего образуются одноцепочечные комплементарные концы с
«Хвостами» из четырех нуклеотидов в каждом (липкие концы).
Палиндромные последовательности, которые распознаются рестрицирующими
эндонуклеазами типа II и в которых происходит расщепление молекулы ДНК,
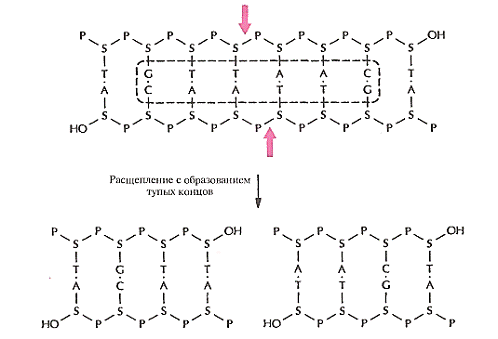
называются сайтами узнавания. Помимо рестриктаз, расщепляющих полинуклеотидную
цепь с образованием липких концов, существуют рестриктазы, которые вносят
разрывы в цепи строго друг против друга с образованием фрагментов ДНК с
«тупыми» концами.
Для осуществления модификаций в геноме микроорганизма недостаточно одних
только ферментов рестрикции. Во-первых, водородные связи между теми
четырьмя основаниями, которые образуют липкие концы, недостаточно прочны, чтобы
удержать два объединившихся фрагмента ДНК. Необходим какой-то инструмент для
устранения разрыва в сахарофосфатном остове молекулы. Таким инструментом
является ДНК-лигаза бактериофага Т4. Этот фермент катализирует образование
фосфодиэфирных связей между концами полинуклеотидных цепей, которые уже
удерживаются вместе благодаря спариванию липких концов. Кроме того, ДНК-лигаза Т4
«сшивает» тупые концы, которые сближаются друг с другом после того, как
объединяемые фрагменты связываются с ферментом. Во-вторых, объединение
разных молекул ДНК само по себе бесполезно, если вновь образованные комбинации
(рекомбинантные ДНК) не будут реплицироваться в клетке-хозяине. Таким образом,
если одна масть рекомбинантной молекулы ДНК несет нужный ген, который
предполагается клонировать, то другая должна содержать информацию, необходимую
для репликации в клетке рекомбинантной ДНК. Чтобы решить эту проблему,
используют клонирующие векторы. В-третьих, при рестрикции ДНК образуется
смесь разнообразных фрагментов, и после их легирования с векторной ДНК
образуется множество различных комбинаций. Необходимо уметь распознавать
тереципиентные клетки, которые содержат ДНК с нужной нуклеотидной
последовательностью.
2. Вектор
Вектор (в генетике) - молекула нуклеиновой кислоты, чаще всего ДНК,
используемая в генетической инженерии для передачи генетического материала
другой клетке.
Существующие векторы:
• плазмиды
• вектор на основе бактериофага
• векторы на основе вирусов эукариотических организмов
.1
Плазмиды
Многие свойства бактерий, интересные с точки зрения биотехнологии,
кодируются плазмидами. Плазмиды-это кольцевые молекулы ДНК, которые стабильно передаются
потомству бактериальных клеток независимо от хромосомной ДНК. В генетической
инженерии плазмиды используются для клонирования нужных генов.
Плазмиды различаются по молекулярной массе, так самые мелкие плазмиды
кодируют один-два белка среднего размера, тогда как более крупные - 300 или
более белков. Крупные плазмиды могут кодировать множество ферментов,
необходимых для работы целой последовательности биохимических реакций, например
для превращения толуола в катехол.
В бактериальных клетках плазмиды существуют в виде кольцевых
двухцепочечных ДНК, которые, кроме того, находятся в сверхспиральном состоянии.
Некоторые из плазмид могут существовать лишь в клетках одного или двух близких
видов. К числу плазмид с широким кругом хозяев относятся RP4, RK2 и сходные с ними плазмиды, несущие маркер устойчивости к
лекарственным препаратам (R-плазмиды),
которые принадлежат Р-группе несовместимости (члены одной и той же группы
несовместимости не могут сосуществовать в бактериальной клетке). Эти плазмиды
были перенесены во многие виды грамотрицательных бактерий; создается
впечатление, что их хозяевами могут быть все штаммы грамотрицательных бактерий.
Таким образом, R-плазмиды,
относящиеся к группе Р, особенно подходят для генетических экспериментов с
грамотрицательными бактериями, используемыми в промышленности. При помощи этих
плазмид мы можем осуществлять перенос хромосомных генов между неродственными
видами. Небольшие по размеру производные плазмидRP4 и RK2
используются как векторы для клонирования молекул ДНК, которая затем может быть
перенесена в клетки самых разных видов.
Если плазмиды не могут стабильно сосуществовать в одной клетке в условиях
отсутствия селективного давления, их называют несовместимыми. Несовместимость
плазмид обусловливается подавлением репликации одной из них и (или)
блокированием распределения дочерних молекул ДНК по клеткам перед их делением.
Эти оба механизма действуют независимо друг от друга.
Несовместимость, вызванная подавлением репликации, наблюдается у плазмид
как со строгим, так и с ослабленным контролем репликации. Она обусловлена
существованием у плазмид генетического механизма поддержания числа
плазмидных копий на определенном уровне, который приводит к тому, что в
клетке только одна из двух плазмид (резидентная или с меньшей
вероятностью вошедшая) сохраняет способность к удвоению.
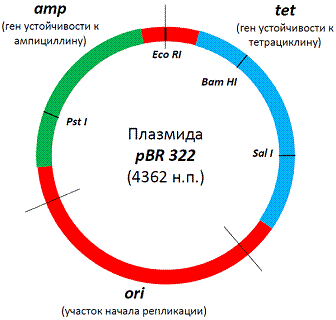
Структура
знаменитой плазмиды PBR322 <#"698719.files/image004.gif">
Однако при недостатке питательных веществ или иных неблагоприятных
обстоятельствах интегрированная фаговая ДНК высвобождается, и запускается
литический цикл развития. Размер ДНК фага λ составляет примерно 50т. п. н.,
причем значительная ее часть (около 20 т. и. н.) несущественна для размножения
фага и отвечает за его встраивание в хозяйскую ДНК. В связи с этим возникла
идея, что ее можно заменить фрагментом другой ДНК эквивалентного размера.
Образующаяся рекомбинантная молекула будет реплицироваться в клетке как ДНК
«рекомбинантного» фага λ, «вставшего» на литический путь развития.
Чтобы понять, как функционирует векторная система на основе фага λ, необходимо рассмотреть молекулярные
аспекты литического цикла развития. Инфекционная фаговая частица имеет головку,
в которой заключена плотно упакованная ДНК длиной примерно 50 т. п. н., и
отросток с отходящими от него тонкими белковыми нитями (фибриллами). Сборка
головки отростка и упаковка ДНК четко скоординированы. ДНК фага λ
- это линейная двух
цепочечная молекула длиной 50 т. п. н. с одноцепочечными 5'-«хвостами» из 12
нуклеотидов. Их называют липкими (cos) концами, поскольку они взаимно комплементарны и могут спариваться друг
с другом. После того как фаговая ДНК проходит через отросток и попадает в Е.
coli, cos-концы соединяются с образованием кольцевой молекулы.
На раннем этапе литического цикла в результате репликации кольцевой молекулы
ДНК образуется линейная молекула, состоящая из нескольких сегментов длиной 50
т. п. н.
Каждый из таких сегментов упаковывается в белковую головку, к последней
присоединяется уже собранный отросток и образуется новая фаговая частица. При
упаковке молекулы ДНК длиной менее 38 т. п. н. получается неинфекционная
фаговая частица, а фрагменты длиной более 52 т. п. н. не умещаются в головку.
Сегменты длиной 50 т. п. н. в линейной молекуле ДНК разделены cos-сайтами, и именно по этим сайтам
разрезается молекула, когда очередной сегмент заполняет головку. Разрезание
осуществляет фермент, находящийся у входа в головку.
В результате исследований по изучению сборки фага λ была разработана система упаковки
молекул ДНК in vitro с образованием инфекционных фаговых
частиц. Смешав в пробирке очищенные пустые головки, фаговую ДНК и собранные
отростки, можно получить инфекционные фаговые частицы.
Задача исследователя состоит втом, чтобы заменить этот центральный
участок нуклеотидной последовательностью длиной примерно 20 т. п. н. нужной
ДНК. ДНК, предназначенную для клонирования, тоже расщепляют на фрагменты
размером от 15 до21 т. п. н. Оба препарата - фаговую и чужеродную ДНК -
объединяют и добавляют ДНК - лигазу. Фрагменты ДНК длиной 50 т. п. н.
упаковываются в головки, к ним присоединяются отростки и образуются
инфекционные фаговые частицы. Фрагменты большего (>52 т. л. н.) или меньшего
(<38 т. п. н.) размера упаковываться не могут.
2.3
Космиды
Гибрид плазмиды и фага. По сути дела, это просто плазмида, в которую
добавлены сайты для связывания с белками оболочки фага (они называются
cos-сайтами, и именно благодаря им космиды получили свое название). Белковая
оболочка делает космиду стабильнее, благодаря чему в нее можно загружать более
длинные вставки.
Векторы, называемые космидами, могут включать до 40 т. п. н. чужеродной
ДНК и при этом активно амилифипицироваться в Е. coli как плазмиды. Космиды объединяют в
себе свойства плазмидных векторов и векторов на основе фага λ.
Например, широко
применяемая космида pLFR-5
(приблизительно 6 т. п. н.) имеет два cos-сайта фага λ, разделенных сайтом рестрикции, полилинкер с шестью
уникальными сайтами рестрикции, точку начала репликации ДНК (ori) и ген устойчивости к тетрациклину
(Тс). Эта космида может интегрировать чужеродную ДНК длиной до 40 т. п. н.
Препараты ДНК смешивают и лигируют. Те продукты легирования, которые содержат
вставку длиной 40 т. п. н., имеют суммарный размер, близкий к 50 т. п. н., и,
следовательно, могут упаковываться in vitro в головки
фага λ.
Реассоциировавшие
молекулы pLFR-5, не содержащие вставок, упакованы
не будут. После сборки фаговых частиц инфицируют ими Е. coli.
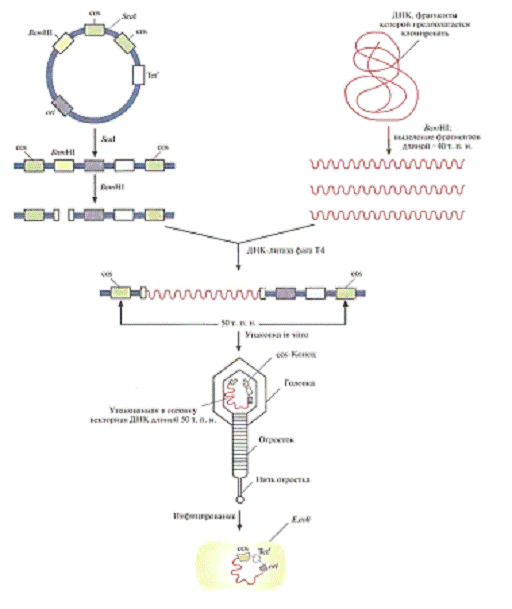
Оказавшись в бактериальной клетке, линейная молекула со вставкой
замыкается в кольцо благодаря спариванию cos-сайтов. В такой стабильной конфигурации она может
долгое время существовать в клетке и реплицироваться как гибридная плазмида,
поскольку содержит все необходимые для этого элементы. Кроме того, ген
устойчивости к тетрациклину обеспечивает рост колоний, несущих данную космиду.
Космиды имеют большое преимущество по сравнению с плазмидами: в них можно
встраивать более протяженные фрагменты, а это означает, что для создания
геномной библиотеки нужно меньшее число клонов и потребуется меньше времени на
их скрининг.
3.
Escherichia coli (E. coli) -
основная модель биотехнологических манипуляций
Кишечная палочка (лат. Escherichia coli, E. coli, по имени
Теодора Эшериха) - грамотрицательная палочковидная бактерия, широко встречается
в нижней части кишечника теплокровных организмов.E. coli - грамотрицательная
бактерия, факультативный анаэроб, не образует эндоспор. Клетки палочковидные,
со слегка закруглёнными концами, размером 0,4-0,8 х 1-3 мкм, объём клетки
составляет около 0,6-0,7 μm³. Штаммы, имеющие жгутики, способны
передвигаться. Жгутики расположены перитрихально. Протопласт E. coli
одет в муреиновый мешок, прилегающий к внешней мембране. E. coli
относится к микроорганизмам, не обладающим физиологической компетентностью к
поглощению экзогенной ДНК.
Кишечная палочка может жить на разных субстратах. В анаэробных условиях E.
coli образует в качестве продукта жизнедеятельности лактат, сукцинат,
этанол, ацетат и углекислый газ. Часто при этом образуется молекулярный
водород, который мешает образованию указанных выше метаболитов, поэтому E.
coli часто сосуществует с микроорганизмами, потребляющими водород -
например, с метаногенами или бактериями, восстанавливающими сульфат.
Оптимальный рост достигается культурами E. coli при
температуре 37 °C, некоторые штаммы могут делиться при температурах до 49 °C.
Рост может стимулироваться аэробным или анаэробным дыханием, различными парами
окислителей и восстановителей, в том числе, окислением пирувата, формиата,
водорода, аминокислот, а также восстановлением кислорода, нитрата,
диметилсульфоксида и триметиламин N-оксида.
E. coli играет важную роль в современной промышленной микробиологии и биологической
инженерии. Работа Стенли Нормана Коэна и Герберта Бойера на E. coli, с
использованием плазмид и эндонуклеаз рестрикции для создания рекомбинантной
ДНК, находится у истоков современной биотехнологии. Усовершенствование методов
получения сферопластов E. coli и их трансфекции позволили достичь
достаточно высокой эффективности трансформации молекулами ДНК различных фагов.
Кишечную палочку считают универсальным организмом для синтеза
чужеродных белков. В
E. coli исследователи вводят гены при помощи плазмид, что позволяет
осуществлять биосинтез белков для промышленной ферментации. Также разработаны
системы для синтеза в E. coli рекомбинантных белков. Одним из первых
примеров использования технологии рекомбинантных ДНК является синтез аналога
инсулина человека. Модифицированные E. coli используют при разработке
вакцин, синтеза иммобилизованных ферментов и решения других задач. Однако, в
организмеE. coli невозможно получать некоторые крупные белковые
комплексы, содержащие дисульфидные связи, в частности, белки, для проявления
биологической активности которых требуется посттрансляционная модификация.
Чужеродные гены клонируют в так называемых челночных векторах. Эти
вектора с одинаковым успехом реплицируются в клетках нескольких хозяев, в
данном случае, в клетках E. coli. Векторы были получены комбинацией in
vitro фрагментов этих плазмид.
Для конструирования рекомбинантной ДНК, содержащей в своем составе ген,
который должен экспрессироваться, придерживаются следующей стратегии.
Синтезируют к ДНК или из клонотеки выделяют клетки, несущие фрагмент генома с
нужным геном, и клонируют их в соответствующем векторе. Фрагменты геномной ДНК
подвергают модификации - удаляют из них некодирующие области и участки соседних
генов. Часто для проведения этой операции необходимо секвенирование данного
фрагмента ДНК. Затем конструируются промежуточные рекомбинантные ДНК, в которых
ген помещается под контроль бактериальных регуляторных элементов (промотор,
оператор, точка связывания с рибосомами). Эти регуляторные элементы выделяют из
гибридных плазмид, сконструированных специально как источники регуляторных
элементов. Полученная конструкция встраивается в подходящий вектор, например,
pBR 322, и ген экспрессируется в бактериальной клетке.
Однако удобнее встраивать ген в специальный вектор для экспрессии,
который уже содержит регуляторные элементы, обеспечивающие активную экспрессию
после введения рекомбинантной плазмиды в бактериальную клетку. К таким
эффективным регуляторным участкам относится, например, сильный промотор гена
бэта-лактамазы (ген устойчивости к пенициллину, входящий в состав плазмиды pBR
322). Ряд генов, в том числе и ген инсулина, встраивали в сайт рестрикции Pst
I, который расположен в структурной части гена. Промотор этого гена
обеспечивает эффективную транскрипцию, которая продолжается до тех пор, пока
РНК-полимераза не дойдет до сигнала терминации встроенного гена.
С другой стороны, если фрагмент чужеродной ДНК встраивается в один из
генов устойчивости, то последний инактивируется. Следовательно, успешное
встраивание фрагмента чужеродной ДНК в один из этих генов легко детектировать
по исчезновению у бактерий устойчивости к данному антибиотику.
4.
Основные этапы трансформации E. Coli
молекулярное клонирование кишечная палочка
1. Амплификация интересующего участка гена с помощью полимеразной цепной
реакции (ПЦР)- экспериментальный метод молекулярной биологии, позволяющий
добиться значительного увеличения малых концентраций определённых фрагментов
нуклеиновой кислоты (ДНК) в биологическом материале (пробе).
Метод основан на многократном избирательном копировании определённого
участка ДНК при помощи ферментов в искусственных условиях (in vitro).
При этом происходит копирование только того участка, который удовлетворяет
заданным условиям, и только в том случае, если он присутствует в исследуемом
образце. В отличие от амплификации ДНК в живых организмах, (репликации), с
помощью ПЦР амплифицируются относительно короткие участки ДНК.

Для проведения ПЦР в простейшем случае требуются следующие
компоненты:
· ДНК-матрица, содержащая тот участок ДНК, который требуется
амплифицировать.
· Два праймера, комплементарные противоположным концам разных
цепей требуемого фрагмента ДНК.
· Термостабильная ДНК-полимераза - фермент, который
катализирует реакцию полимеризации ДНК. Полимераза для использования в ПЦР
должна сохранять активность при высокой температуре длительное время, поэтому
используют ферменты, выделенные из термофилов - Thermusaquaticus
(Taq-полимераза), Pyrococcusfuriosus (Pfu-полимераза), Pyrococcuswoesei
(Pwo-полимераза) и другие.
· Дезоксирибонуклеозидтрифосфаты (dATP, dGTP, dCTP, dTTP).
· Ионы Mg2+, необходимые для работы полимеразы.
· Буферный раствор, обеспечивающий необходимые условия реакции
- рН, ионную силу раствора.
2. Выделение ПЦР-продукта из геля
Электрофорез ДНК - это аналитический метод, применяемый для разделения
фрагментов ДНК по размеру (длине) и форме (в случае, если ДНК образует
вторичные структуры, например шпильки). Силы электрического поля,
прикладываемого к образцам, заставляют фрагменты ДНК мигрировать через гель.
Сахарофосфатный остов молекул ДНК заряжен отрицательно и поэтому цепи ДНК
двигаются от катода, заряженного отрицательно, к положительному аноду. Более
длинные молекулы мигрируют медленнее, так как задерживаются в геле, более
короткие молекулы двигаются быстрее.
К образцам обычно добавляют низкомолекулярный кислый краситель (например,
динитрофенол, бромфеноловый синий), чтобы визуализировать ход электрофореза в
процессе. Краситель также необходим для того, чтобы определить, когда стоит
остановить процесс.
После разделения (иногда краситель вносят в расплавленную агарозу)
фрагменты ДНК разной длины визуализируют при помощи флюоресцентных красителей,
специфично взаимодействующих с ДНК, например, агарозные гели обычно красят бромистым
этидием, который интеркалирует между азотистыми основаниями дуплекса и
флюоресцирует в УФ-лучах.
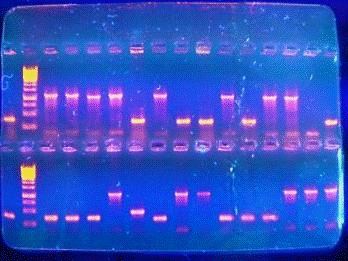
Определение размеров производят путем сравнения коммерчески доступных
фрагментов ДНК (DNA ladder, «линейка»), содержащий линейные фрагменты ДНК
известной длины.
Выделение продуктов из геля осуществляется, сначала механическим
выделением необходимого участка агарозного геля, в котором заключена ДНК, а
затем с помощью специальных наборов («ЕВРОГЕН», «Glassmilk», и т.д.) проводится экстракция ДНК.
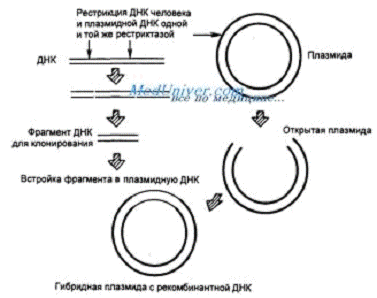
3. Лигирование ампликона в вектор (обычно плазмида)- образование
фосфодиэфирной связи между двумя основаниями одной цепи ДНК, разделенных
разрывом, с помощью фермента ДНК-лигазы. ДНК-лигазы абсолютно необходимы в
процессах репарации ДНК и в процессах репликации - при удвоении цепи ДНК.
Лигирование применяют в молекулярной биологии для соединения каких-либо молекул
ДНК.
Сначала проводится рестрикция образца ДНК и плазмиды одной и той же рестриктазой
(по липким или тупым концам), затем посредством водородных связей липкие концы
плазмиды и образца ДНК «захлопываются» но остается разрыв на векторе, поскольку
нуклеотиды сами не способны образовывать фосфодиэфирные связи. Для этого
добавляется фермент ДНК-лизага, которая образует фосфодиэфирные связи в новой
гибридной молекуле ДНК. Теперь мы имеем готовый вектор.
4. Трансформация полученным вектором E. Coli
Трансформация - то перенос вектора в клетки специального штамма E.
coli
Трансформация состоит из двух частей это получение компетентных клеток с
ночного высева бактерий с суспензионную среду и непосредственно трансформация.
Между этими двумя этапами часто используют недлительную заморозку в жидком
азоте или холодильнике на -80, что улучшает производительность. Получение
компетентных клеток проходит в несколько этапов, суть которых заключается
сначала в отделении клеток от среды LB, а затем в помещении в раствор хлорида кальция с 10мМ TrisHcl и 60% раствор глицерина. На всех
этапах используют центрифугирование на низких оборотах(4600об/мин).
Непосредственно трансформация осуществляется действием на клетки теплового
шока, после добавления раствора плазмиды, смесь охлаждают до +4 градусов, а
затем на 2 минуты помещают на водяную баню на +42 градуса. В это время клетка
«захватывает» плазмиды, и после добавления среды LB полученную смесь инкубируют в термостате около часа на +37
градусов. После этого происходит растирание клеток на твердую среду, в которую
заранее был добавлен антибиотик.
5. Отбор клонов (бактерии, содержащие вставку в плазмиде искомой
последовательности)
Отбор клонов осуществляется реакцией ПЦР на исследуемые вставки. Детекция
амплифицированного ПЦР-продукта осуществляется так же с помощью электрофореза.
При наличии необходимой вставки, ПЦР продукты используются в дальнейшем при
реакции секвенирования.
6. Секвенирование - определение последовательности ДНК исследуемого
ПЦР-продукта.
5.
Практическое применение трансформированных микроорганизмов
Важной составной частью биотехнологии является генетическая инженерия.
Родившись в начале 70-х годов, она добилась сегодня больших успехов. Методы
генной инженерии преобразуют клетки бактерий, дрожжей и млекопитающих в
"фабрики" для масштабного производства любого белка. Это дает возможность
детально анализировать структуру и функции белков и использовать их в качестве
лекарственных средств.
В настоящее время кишечная палочка (E. coli) стала поставщиком
таких важных гормонов как инсулин и соматотропин. Ранее инсулин
получали из клеток поджелудочной железы животных, поэтому стоимость его была
очень высока. Для получения 100 г кристаллического инсулина требуется 800-1000
кг поджелудочной железы, а одна железа коровы весит 200 - 250 грамм. Это делало
инсулин дорогим и труднодоступным для широкого круга диабетиков.
В 1978 году исследователи из компании "Genentec" впервые
получили инсулин в специально сконструированном штамме кишечной палочки.
Инсулин состоит из двух полипептидных цепей А и В длиной 20 и 30 аминокислот.
При соединении их дисульфидными связями образуется нативный двухцепочечный
инсулин. Было показано, что он не содержит белков E. coli, эндотоксинов и
других примесей, не дает побочных эффектов, как инсулин животных, а по
биологической активности от него не отличается. Впоследствии в клетках E. coli
был осуществлен синтез проинсулина, для чего на матрице РНК с помощью обратной
транскриптазы синтезировали ее ДНК-копию. После очистки полученного проинсулина
его расщепили и получили нативный инсулин, при этом этапы экстракции и
выделения гормона были сведены к минимуму. Из 1000 литров культуральной
жидкости можно получать до 200 граммов гормона, что эквивалентно количеству
инсулина, выделяемого из 1600 кг поджелудочной железы свиньи или коровы.
Соматотропин - гормон роста человека, секретируемый гипофизом. Недостаток
этого гормона приводит к гипофизарной карликовости. Если вводить соматотропин в
дозах 10 мг на кг веса три раза в неделю, то за год ребенок, страдающий от его
недостатка, может подрасти на 6 см. Ранее его получали из трупного материала,
из одного трупа: 4 - 6 мг соматотропина в пересчете на конечный
фармацевтический препарат. Таким образом, доступные количества гормона были
ограничены, кроме того, гормон, получаемый этим способом, был неоднороден и мог
содержать медленно развивающиеся вирусы. Компания "Genentec" в 1980
году разработала технологию производства соматотропина с помощью бактерий,
который был лишен перечисленных недостатков. В 1982 году гормон роста человека
был получен в культуре E. coli и животных клеток в институте Пастера во
Франции, а с 1984 года начато промышленное производство инсулина и в СССР. При
производстве интерферона используют как E. coli, S. cerevisae
(дрожжи), так и культуру фибробластов или трансформированных лейкоцитов.
Аналогичными методами получают также безопасные и дешевые вакцины.
На технологии рекомбинантных ДНК основано получение высокоспецифичных
ДНК-зондов, с помощью которых изучают экспрессию генов в тканях,
локализацию генов в хромосомах, выявляют гены, обладающие родственными
функциями. ДНК-зонды также используются в диагностике различных заболеваний.
Технология рекомбинантных ДНК сделала возможным нетрадиционный подход
"белок-ген", получивший название "обратная генетика".
При таком подходе из клетки выделяют белок, клонируют ген этого белка, модифицируют
его, создавая мутантный ген, кодирующий измененную форму белка. Полученный ген
вводят в клетку. Если он экспрессируется, несущая его клетка и ее потомки будут
синтезировать измененный белок. Таким образом можно исправлять дефектные гены и
лечить наследственные заболевания.
У дрожжей Saccharomycescerevisiae с помощью промотора
алкогольдегидрогеназы удалось добиться прямой транскрипции в дрожжах гена
инсулина человека, а в опытах с использованием промотора фосфоглицераткиназы
получен высокий уровень экспрессии генов интерферонов.
С помощью бактерий были получены с высоким выходом некоторые белки -
продукты генов животных и их вирусов. Так, были созданы штаммы Е. coli,
у которых 20% всего клеточного белка составляли коровый антиген вируса гепатита
В, гормон роста человека или главный капсидный антиген вируса ящура. У одного
из сконструированных штаммов В. sublilis последний составлял около 1%
синтезируемого этой бактерией белка.
Новые разработки в области генетической теории и технологии послужили
основой для создания целого арсенала методов получения новых штаммов и путей
модернизации технологических процессов. Хорошо освоенные мутагенез и отбор
будут использоваться и далее, но специфичность этого метода станет гораздо выше
благодаря введению мутаций в клонированные гены in vitro.
Усовершенствования оборудования и способов контроля за работой ферментеров
существенно облегчат постоянный отбор, нацеленный на улучшение количественных
характеристик организма-продуцента.
С помощью методов генетической инженерии можно будет получать продукты,
которые не могли ранее применяться из-за того, что выделять их из тканей
животных было сложно и дорого. К числу их относятся интерфероны. В клиниках
сейчас очень широко изучается эффективность синтезируемых бактериями интерферонов
при лечении вирусных заболеваний и разных форм рака.
Сейчас, даже трудно предсказать все возможности, которые будут
реализованы в ближайшие несколько десятков лет.
Список
литературы
1. Глик Б.,
Пастернак Дж. Молекулярная биотехнология. Принципы и применение. Пер. с англ. -
М.: Мир, 2002. - 589 с, ил.
2. Рыбчин
В. Н. Основы генетической инженерии. 2-е изд., перс роб идол.: Учебник для
вузов. СПб.: Изд-во СП61ТУ, 2002. 522 с.
. Сартакова
О.Ю. Основы микробиологии и биотехнологии. Часть 1: Уч. Пособие. / Алтайский
государственный технический университет им. И.И.Ползунова - Барнаул. Изд-во
АГТУ, 2001. - 64 с.
. http://ru.wikipedia.org
. http://www.biotechnolog.ru