Проведение сравнительного анализа генетической структуры городского и сельского населения Полесья, по генам, контролирующим группы крови АВО
Введение
В антропологии и других смежных науках проводятся обширные
исследования для решения актуальной проблемы взаимодействия человека и
биосферы, популяционно-генетической изменчивости человека в зависимости от
экологических и исторических факторов. С этой целью широко применяются
достижения в иммунологии, изосерологии, биохимии, генетике. Формирование
антропологических типов, генетико-демографических структур связано с адаптацией
человека к разнообразным условиям окружающей среды. Поскольку генетически
контролируемые гематологические системы АВО, МN, резус, Р, Льюис,
гаптоглобины и другие в онтогенезе не подвержены воздействию средовых факторов,
их использование при изучении эволюционных изменений и оказывается более
предпочтительным по сравнению с морфологическими признаками.
Изучением наследственности у человека занимается частная
генетика, называемая антропогенетикой, которая представляет собой важный раздел
и изучает:
1) генетическую детерминацию физиологических,
биохимических и морфологических свойств отдельных тканей и органов человека,
нервно-гуморальную координацию его психической (эмоциональной) и
интеллектуальной деятельности;
2) статистические закономерности распределения генных
частот в микропопуляциях;
3) методы защиты генотипа человека от поражения
различными факторами среды - химическими агентам в производстве, ионизирующей
радиацией, фармакологическими препаратами, космическими излучениями и др.;
4) генетическую обусловленность болезней, их передачу в
поколениях, проявление в онтогенезе, распространение в популяциях, возможность
медико-генетических консультаций по вопросам наследственных болезней,
географическое распространение и т.д.;
5) роль наследственности и среды в формировании
личности;
) молекулярные механизмы памяти, основанные на принципе
кодирования и передачи наследственной информации [5].
Исследованиями выявлен достаточно разнообразный генетический
полиморфизм человеческих популяций. Но не всегда удается выяснить, какими
причинами он обусловлен. В качестве основных причин выдвигаются адаптация
человека к условиям среды через генетическую изменчивость в поколениях,
стохастические процессы в случае изолированности, миграционные потоки
иноэтничного населения.
Все более детально разрабатывается математический аппарат,
достаточно эффективный при оценке исходных компонентов в смешанных группах, при
ретроспективном анализе хронологии формирования новых популяций посредством их
дивергенции и дифференциации в пределах генофонда. На основании выявленной
гетерогенности в распределении эритроцитарных групп крови все народы земного
шара разделены на десятки рас второго порядка. Правда, для этих целей
использовано не более двух-трех маркерных систем.
Анализ концентраций множества генов с обязательной оценкой
таксономических расстояний позволяет классифицировать популяции разных уровней
и генетико-антропологические типы. Поэтому данные обобщенной картографии
позволяют открыть центры происхождения генофонда этносов. Материалы
популяционной генетики находят все более широкое применение при решении этногенетических
вопросов, а также при выяснении миграционных потоков.
Исследование генофонда современного населения даст
возможность использовать его также в качестве биологического показателя,
необходимого медицинской науке при определении степени риска заболевания
отдельных индивидуумов рядом болезней.
Для успешного решения всех этих вопросов нужна новая
информация о генетической структуре, прежде всего коренного населения, а затем
и современных смешанных популяций. Несмотря на кажущуюся однородность народонаселения
Белоруссии, дифференцированный скрининг генетических маркеров предположительно
должен установить генетико-антропологические закономерности, которые сложились
в процессе адаптивной жизнедеятельности и репродуктивной преемственности
десятков и сотен прародительских поколений. Изучение индивидуальных
биологических свойств и популяционных генетических структур, таким образом,
поможет решить многие из выше обозначенных проблем, что свидетельствует об
актуальности данного исследования.
При исследовании репрезентативных выборок использовался
математический аппарат и вычислительная техника. С их помощью выполнено
таксономическое изучение населения и дан сравнительный анализ его биологической
изменчивости в процессе исторического становления и с учетом региональных
экологических и социально-демографических особенностей.
Целью нашей работы является: изучить целостную генетическую
систему современного населения, обладающую сложной пространственной и
динамичной временной структурой в виде специфического соотношения определенных
генотипов и аллелей; выявить вероятный гомеостаз в популяциях, их способность
сохранить свои особенности [6].
1. Обзор литературы
1.1
Человек как объект генетического исследования
Человек, являясь весьма высокоорганизованным биологическим и
социальным существом, нуждается в многостороннем изучении вообще и в
генетическом особенно. Однако, по мнению некоторых исследователей, существуют
определенные сложности и даже неблагоприятные моменты при анализе действия
генов у человека. Ими являются относительная продолжительность поколений,
значительная длительность дорепродуктивного периода, небольшое количество
потомков в семьях, относительно большое число хромосом, многочисленные
модификации наследственной изменчивости и некоторые другие причины, связанные с
морально-этическими моментами - абсолютной невозможностью проведения каких бы
то ни было экспериментов на человеке и народонаселении в целом.
Подробная изученность человека по большинству его
фенотипических особенностей на всем земном шаре, как в оптимальных, так и
экстремальных условиях пребывания позволяет глубоко исследовать его
потенциальные генетические возможности.
Все увеличивающаяся численности людей, обозримая длительность
существования человечества, возможности анализа семейных поколений или их
родословных представляют определенные преимущества при генетическом
исследовании человека по сравнению с животными.
Популяционная генетика человека в свою очередь изучает те
генетические процессы, которые происходят во времени и пространстве, в
популяциях человека, природу генетических различий и взаимосвязь между
отдельными группами людей. Наследственная изменчивость людей может проявиться
вследствие участившихся за последние годы миграций, на разных иерархических
уровнях, таких, как локальный, региональный, этнический, лингвистический и
другие [7].
Каждый из некогда существовавших или ныне живущих видов
представляет собой итог определенного цикла эволюционных преобразований на
популяционно-видовом уровне, закрепленный изначально в его генофонде. Последний
отличается двумя важными качествами. Во-первых, он содержит биологическую
информацию о том, как данному виду выжить и оставить потомство в определенных
условиях окружающей среды, а во-вторых, обладает способностью к частичному
изменению содержания заключенной в нем биологической информации. Последнее
является основой эволюционной и экологической пластичности вида, т.е.
возможности приспособиться к существованию в иных условиях, меняющихся в
историческом времени или от территории к территории [8].
Интерес к исследованиям популяций человека резко возрос после
установления генетических последствий радиоактивного и рентгеновского
облучений. Одновременно большое внимание стало уделяться изучению роли
генетических факторов в патогенезе различных заболеваний. Наряду с изучением
наследования нормальных признаков стали обращать внимание на наследование
патологических синдромов. Полиморфизм многих генетических систем человека,
генофонд человечества формируется под влиянием мутагенных факторов окружающей среды,
возможна определенная роль в этом и инфекционных болезней, прежде всего
болезней вирусной этиологии.
Популяция человека характеризуется высокой степенью
наследственного биохимического разнообразия.
Кроме анализа частот всевозможных генов, генотипов и
фенотипов, генетическая структура человеческих популяций изучается также путем
исследования систем браков. Географическая близость населения подразумевает его
родственные связи, но они вовсе необязательны. Различные системы браков могут
иметь определенные биологические последствия.
Сегодня эти и другие аспекты антропогенетики исследуются с
использованием популяционных и генодемографических методов [7].
1.2 Методы изучения генетики человека
Несмотря на указанные затруднения, генетика разработала
некоторые методы, которые позволяют шаг за шагом изучать наследственность и
наследование у человека. К методам, широко используемым при изучении генетики
человека, относятся генеалогический, популяционно-статистический, близнецовый,
метод дерматоглифики, цитогенетический, биохимический, методы генетики
соматических клеток, генодемографический метод [9].
Генеалогический метод
В основе этого метода лежит составление и анализ родословных.
Этот метод широко применяют с древних времен и до наших дней. Родословные
человека составлялись на протяжении многих столетий в отношении царствующих
семейств в Европе и Азии.
Как метод изучения генетики человека генеалогический метод
стали применять только с начала ХХ столетия, когда выяснилось, что анализ
родословных, в которых прослеживается передача из поколения в поколение
какого-то признака (заболевания), может заменить собой фактически неприменимый
в отношении человека гибридологический метод.
При составлении родословных исходным является человек -
пробанд, родословную которого изучают. Обычно это или больной, или носитель
определенного признака, наследование которого необходимо изучить. При
составлении родословных таблиц используют условные обозначения, предложенные Г.
Юстом в 1931 г. Поколения обозначают римскими цифрами, индивидов в данном
поколении - арабскими.
С помощью генеалогического метода может быть установлена
наследственная обусловленность изучаемого признака, а также тип его
наследования. При анализе родословных по нескольким признакам может быть
выявлен сцепленный характер их наследования, что используют при составлении
хромосомных карт. Этот метод позволяет изучать интенсивность мутационного
процесса, оценить экспрессивность и пенетрантность аллеля. Он широко
используется в медико-генетическом консультировании для прогнозирования
потомства. Однако необходимо отметить, что генеалогический анализ существенно
осложняется при малодетности семей [9].
Популяционно-статистический
метод
С помощью популяционно-статистического метода изучают
наследственные признаки в больших группах населения, в одном или нескольких
поколениях. Существенным моментом при использовании этого метода является
статистическая обработка получаемых данных. Этим методом можно рассчитать
частоту встречаемости в популяции различных аллелей гена и разных генотипов по
этим аллелям, выяснить распространение в ней различных наследственных
признаков, в том числе заболеваний. Он позволяет изучать мутационный процесс,
роль наследственности и среды в формировании фенотипического полиморфизма
человека по нормальным признакам, а также в возникновении болезней, особенно с
наследственной предрасположенность. Этот метод используют и для выяснения
значения генетических факторов в антропогенезе, в частности в расообразовании.
При статистической обработке материала, получаемого при
обследовании группы населения по интересующему исследователя признаку, основой
для выяснения генетической структуры популяции является закон генетического
равновесия Харди-Вайнберга. Он отражает закономерность, в соответствии с
которой при определенных условиях соотношение аллелей генов и генотипов в
генофонде популяции сохраняется неизменным в ряду поколений этой популяции. На
основании этого закона, имея данные о частоте встречаемости в популяции
рецессивного фенотипа, обладающего гомозиготным генотипом, можно рассчитать
частоту встречаемости указанного аллеля в генофонде данного поколения.
Распространив эти сведения на ближайшие поколения, можно предсказать частоту
появления в них людей с рецессивным признаком, а также гетерозиготных носителей
рецессивного аллеля.
Анализ частот встречаемости разных признаков в популяции в
случае их соответствия закону Харди-Вайнберга позволяет утверждать, что
признаки обусловлены разными аллелями одного гена [9].
Генодемографические
методы в антропологии
Среди наук, изучающих народонаселение, популяционная генетика
человека, пожалуй, самая молодая.
На протяжении эволюции структура популяций человека
неоднократно претерпевала изменения [10]. Элементарные эволюционные процессы
проявляются чаще всего в географически ограниченных сообществах людей. Такие
сообщества называют популяциями.
Члены популяции должны занимать общую территорию и свободно
вступать в брак между собой. Но не общность территории, а родственные связи
играют важную роль в формировании популяций людей и их генофонда.
Генофонды популяций не остаются постоянными во времени.
Соотношение генотипов в популяциях, распространение одних генов и отсев других
происходят в процессе генетической и социальной среды обитания.
При экспедиционной работе антропологов наиболее
информативными являются исследования сельского населения, если необходимо
выявить издревне зафиксированные геногеографические структуры. Последующее
картографирование фенотипов и генов позволяет установить некоторые
закономерности в их распределении - последовательное нарастание или падение
концентрации на одних территориях, дисперсное распространение на других.
Так, наблюдается довольно плавное нарастание частоты
встречаемости гена В или гена N системы MN в пределах Евразии по мере движения с севера-запада
на юго-восток. Многие антропологи пишут о наличии группы В как о непременном
признаке монголоидной расы.
Факторы генетической динамики обуславливают огромное
индивидуальное разнообразие людей, однако это разнообразие ограничивается
пределами возможностей генофонда одной популяции или совокупной популяционной
системы. Генетические процессы в популяциях человека, будучи тесно
взаимосвязанными с демографическими процессами, в конечном счете, проявляются в
наследственном полиморфизме. Последний предстает в качестве основного источника
демографо-генетической информации.
Для изучения генетической структуры популяции и
морфофизиологической функции организма используются в качестве маркеров гены,
кодирующие биохимические, иммунологические, физиологические, морфологические
признаки. Проведенное в комплексе популяционное обследование позволяет получить
сведения не только о степени гомогенности или гетерогенности популяции по
собственно генетическим параметрам. Генетически важными являются и другие
источники информации.
В демографической структуре всего народонаселения
присутствуют различные иерархически соподчиненные уровни. Воспроизводство
генофондов осуществляется в популяциях, которые занимают нижний уровень
популяционной системы человечества.
В таком качестве в недалеком прошлом выступала деревня,
небольшие поселения городского типа, группы близко расположенных сел. Именно
они являются основным объектом демографо-генетических исследований.
Общество воздействует на генетический процесс чаще, чем
кажется на первый взгляд. Происходит это при выборе брачных партнеров. Степень
обладания идентичными генами, разнообразие генотипов в популяции тесно связано
с соотношением экзогамных и эндогамных браков, с увеличением числа последних не
всегда сокращается наследственный полиморфизм.
В другом случае изменение экологической, эпидемиологической,
социально-экономической ситуаций может повлечь за собой определенные потери.
Поэтому необходимо изучать закономерности наследственной передачи признаков
человека в каждой конкретной семье.
Семейная структура населения в отношении групп крови
представлена множеством генетически различимых типов. Частота каждого типа
определяется частотой аллельных генов в конкретной локальной популяции.
Так, семьи с типом иммуногенетической несовместимости
родителей, когда мать имеет группу О(I), а отец - A(II), в Западной Европе
встречается в два раза чаще, чем в Восточной Азии (20 и 10% соответственно).
Среди монголоидов резус-конфликт также исключается из-за фактического
отсутствия у них резус индивидуумов.
Таким образом, общность происхождения, кровнородственные
браки, другие социальные стороны семейной структуры населения играют важную
роль в формировании популяционного генофонда следующего поколения.
На генетической ситуации также сказываются и миграции
населения, расселения людей. Миграции населения являются ни чем иным, как
миграцией генов человека, которые изменяют адаптационные показатели генотипов
коренного населения.
Исследованиями установлено, что генетические процессы в
народонаселении оптимально отточены в ходе исторического развития коренного
населения разных континентов и человечества в целом.
Межпопуляционное генетическое разнообразие нивелируется,
когда миграция оказывается независимой от расстояния [7].
1.3 Антропологические исследования в БССР
По данным многолетних исследований белорусских антропологов и
медицинских генетиков на территории БССР в настоящее время локальные сельские
популяции белорусского населения сохраняются стабильными. По сведениям о
распределении групп крови и других наследственных признаков на территории
Белоруссии определено место белорусов на геногеографической карте Восточной
Европы. В восточнославянских группах по сравнению с западноевропейскими
популяциями концентрация гена В заметно возрастает. При этом одновременно
понижается содержание гена О. Увеличиваются также частоты генов М или N.
В свою очередь современное сельское население Белорусского
Полесья по эритроцитарным системам групп крови обнаруживает значительную
межгрупповую изменчивость.
Развитие обследованных сельских популяций БССР всегда
проходило в естественных условиях с постепенным увеличением генетического,
этносоциального и культурного разнообразия, с формированием относительно
сложной популяционной структуры. Ценность изучения сохранившихся сельских
популяций состоит в возможности определения тенденций генетического процесса в
условиях возрастающей урбанизации и усложняющейся экологической ситуации.
Полученные сведения об изменчивости генетических маркеров позволили исследовать
генетические процессы, а также проанализировать изменения показателей генного
разнообразия во времени.
Генетическая структура популяций перестраивается в поколениях
не только в соответствии с программой заданной наследственности, но и под
влиянием приспособительной изменчивости в меняющихся экологических условиях.
Например, в ряду трех последовательных поколений белорусов по направлению к
младшему снижается концентрация рецессивных гомозигот, а количество гетерозигот
индивидуумов по кодоминантным системам увеличивается.
Таким образом, сейчас нельзя утверждать, что наследственность
неизменна.
По мнению Ю.Г. Рычкова, генетическая структура быстро
растущих миллионных городов далеко не стабильна. Поэтому становится
закономерной необходимость расширения исследований, направленных на дальнейшее
изучение наследственности и изменчивости человека в быстро меняющихся
социальных и экологических условиях [7].
1.4
Геногеография
В отечественной же науке систематический разбор предмета и
задач геногеографии был выполнен еще в 20-е годы А.С. Серебровским. Ему
принадлежит и термин «геногеография» [11].
Отдельный человек рассматривается ею как носитель генов,
унаследованных от родителей в том неповторимом сочетании - генотипе, которым
задаются наследственные основы человеческой индивидуальности. Конкретные гены в
составе генотипа человека служат в популяционной генетике единицами наблюдения
и учета. Однако объект изучения - популяция, т.е. самостоятельная, относительно
обособленная часть населения, исторически сложившаяся на определенной территории
и самовоспроизводящаяся в границах своего ареала. Совокупность генов, которыми
обладает такая популяция, - её генофонд - неизбежно отличается от генофондов
других популяций. Причина этих отличий - в тех генетических процессах, которые
сопровождают самовоспроизводство популяции, её приспособление к условиям
окружающей среды, изменение этих условий в результате жизнедеятельности
популяции, взаимодействие с другими популяциями.
Генетические процессы в популяциях скрыты от глаз
наблюдателя, поскольку время, в котором они совершаются, измеряется
поколениями. В генофондах популяций человека и человечества в целом
генетические изменения происходят непрерывно. Происходят и потому, что
генетически различные поколения в популяции перекрываются во времени, и потому,
что одно и то же поколение в период своего нарождения и в период ухода из жизни
по генетическому составу не тождественно само себе. Причина - в связи
жизнеспособности людей с их генотипами. Лишь в редких случаях внутриутробной
гибели и ранней детской смертности эта связь проявляется зримо, в остальных она
осуществляется лишь в статистическом процессе по законам больших чисел.
Обладатели разных генотипов при прочих равных условиях имеют неравную
вероятность дожить до одного и того же возраста.
Ещё большие изменения в общем составе генофонда производит
неравная рождаемость, отличающая разные семьи одной популяции, городское
население от сельского, разные районы страны друг от друга. Так что
урбанизация, экономические условия, образ жизни, традиции имеют почти прямой
канал влияния на генофонд населения, вызывая неравный прирост разных частей
генофонда и тем самым, меняя его общие характеристики от поколения к поколению.
Изменения в составе генофонда как бы следует за всеми другими изменениями, в
том числе в социально-экономической и культурной сферах жизни населения.
От поколения к поколению происходят вызванные биологическими
и социально-культурными процессами генетические изменения, которые и
накапливаются в генофонде популяции. Вследствие этого генофонды различных
популяций, даже имеющих единое происхождение, под действием таких медленных, но
постоянных изменений генетически удаляются друг от друга. Генетические
изменения рано или поздно проявятся в таких свойствах населения, как
генетически обусловленные предрасположение или устойчивость к разным патогенным
воздействиям среды, структура заболеваемости, антропологическое своеобразие
населения.
Генетическое изучение населения очень трудоёмко, требует
больших затрат времени и средств, поэтому геногеографические данные
накапливаются довольно медленно. Но, если на основе этих, пусть весьма
неполных, данных, руководствуясь определенными генетически обоснованными
правилами, построить карту географического распределения генов, то о многих ещё
не изученных популяциях мы получим предварительную генетическую информацию,
носителем которой является геногеографическая карта.
Геногеография в том значении, в каком это понятие было
введено в науку в 1928 г. русским ученым А.С. Серебровским, есть ветвь
генетики, связывающая ряд генетических проблем с географией. Переход от
микромира генов в макромир природных и социальных ландшафтов нашей планеты
геногеография осуществляет, изучая распространение генов в земном пространстве.
На той или иной территории (и акватории) можно обнаружить длительно
сохраняющийся запас тех или иных генов, в котором Серебровский ещё в 20-е годы
призывал видеть такое же естественное богатство, как в запасах нефти, золота,
каменного угля и других невосполнимых природных ресурсов и обозначил этот
генетический запас термином «г е н о ф о н д». Так геногеография и генофонд
вошли в науку как тесно связанные понятия, при этом основной задачей
геногеографии стало изучение генофонда.
Советские генетики 20-30-х годов не только заложили основы
геногеографии, но и дали блестящие образцы геногеографических исследований,
выполненных в самом разном географическом масштабе - от геногеографии домашних
животных в долинах и ущельях Дагестана (А.С. Серебровский) до мировой
геногеографии культурных растений (Н.И. Вавилов). От географии генов домашних
животных и культурных растений, казалось бы, так далеко до обсуждаемой здесь
геногеографии народов. Но уже пионеры геногеографии продемонстрировали в своих
работах её потенциальные возможности в познании исторической географии популяций
человека - создателя этих пород и культур.
Сегодня, когда известны многие гены человека, работа по
изучению человеческого генофонда стала принципиально возможной. Развёртываются
работы по геногеографии населения: появились сводки по распределению различных
генов в населении мира, возник международный журнал «Gene Geography». Причина
современного пристального внимания к геногеографии состоит в том, что гены
человека лежат в основе его жизнеспособности, здоровья, адаптивных ресурсов
организма, его работоспособности, а в целом - трудовых и оборонных ресурсов
стран.
Решающее значение в географическом описании генофонда имеют
приемы картографирования генетических данных о населении.
Всё разнообразие генов, использованных для построения
геногеографических карт, воплощено в картах с помощью единого, универсального
метода картирования. Благодаря этому в разнообразии построенных карт отражаются
исключительно свойства генов эволюционировать в пространстве в соответствии со
своей функциональной значимостью для жизнедеятельности человека и
распределяться в пространстве независимо друг от друга.
Картографическую работу такого объёма можно выполнить
однотипно и с полной воспроизводимостью только с помощью компьютера - от
расчета генетических характеристик в каждой точке пространства до вывода
изображения генокарт на дисплей и на печать, включая и все процедуры
оперирования картами при различных видах их анализа и обобщения. Для
математического обеспечения всего процесса построения карты создан оригинальный
пакет программ.
Геногеографические карты с определенной степенью точности
заполняют обширные пробелы в знаниях о генетике населения и могут служить
источником предварительной генетической информации. Такие карты представляют
прогноз, на который до получения прямой генетической информации из данного
района могут опираться специалисты в своих исследованиях. Важно отметить, что
эта информация лишена сугубо генетической специфики: язык этих картографических
образов доступен для представителей разных областей знания. Потому и проблемы,
поднимаемые геногеографией населения, являются не сугубо генетическими, а
междисциплинарными, и будучи изложены на наиболее универсальном научном языке,
позволяют объединить усилия как для решения комплексных проблем науки о
человеке, так и частных задач отдельных наук.
Геногеография имеет немалые возможности в разработке тех
фундаментальных и прикладных проблем народонаселения, которые лежат на стыке
биологии человека с другими областями знаний о человеке и его деятельности.
Важно, что геногеография не просто увеличивает объём научной информации о
населении за счет данных о генах человека, а упорядочивает и преобразует эту
информацию таким образом, что она приобретает значение для широкого круга
исследователей и практиков и может быть использована для решения самых
разноплановых задач в настоящем и будущем [12].
1.5
Открытие групп крови
Группа крови каждого человека наследуется от обоих родителей
посредствам передачи двух генов из трех возможных. Она не изменяется в течении
всей жизни. Сейчас среди всех человеческих рас и популящий известно более
двадцати генетических систем групп крови. Вот некоторые из них встречающиеся
наиболее часто: АВО, Rh, MNS, Льюис, Келл-Челлано, Даффи, Лютеран, каждая из
которых контролируется отдельным локусом [7]. К тому же для подавляющего
большинства этих локусов в настоящее время известны множественные аллели [13].
Изоантигены М и N были обнаружены в эритроцитах человека
Ландштейнером и Левиным в 1927 г. По многочисленным антропологическим данным
(сводки по АВО, MN и Rh), гены М и N, а также их генотипы ММ, MN и NN среди
людей на земном шаре распределены неравномерно. Антигены М и N находятся
в эритроцитах человека вместе или каждый в отдельности, но не могут
отсутствовать вовсе. Сформировавшись в раннем эмбриональном периоде, эти
антигены остаются постоянными и не подвергаются заметным количественным
изменениям в течение всей последующей жизни человека. Они передаются по
наследству как простые менделевские пары: у гомозиготных родителей все дети
имеют одинаковые признаки, а потомство гетерозиготных родителей неоднородно.
Причем наследуются групповые антигенные признаки М и N независимо от других
генетически обусловленных признаков, сочетаясь с ними во всевозможных
комбинациях в соответствии с законом вероятности. Кроме популяционной генетики
установленные правила наследования антигенов М и N широко используются
наряду с факторами ABO в судебно-медицинской практике и при
гомотрансплантации органов и тканей [6]. Для установления фенотипов М, МN и N использовали гетероиммунные
сыворотки анти-М и анти-N. Для определения брали свежую кровь и осуществляли его в
такой же последовательности, как это описано для системы АВО [14].
Система гаптоглобина относится к сывороточным системам крови.
Они не связаны с изосерологическими системами эритроцитов. Передаются по
наследству, не зависят от пола и возраста человека. Гаптоглобин относится к
сывороточным белкам. Его открыли в конце 30-х годов. Полоновски и Жейл. Позднее
Смитисом с помощью электрофореза сыворотки крови в крахмальном геле обнаружены
три типа Нр [15].
Считается, что система Льюис состоит из двух генов - Leа и Lеb, которые были
открыты соответственно в 1946 и 1948 гг. Мурентом и Андрезеном. Выявляются они
по степени выраженности реакции агглютинации между сывороткой и эритроцитами
[6]. Антигенов системы Льюис определяются по степени выраженности реакции
агглютинации между сывороткой и эритроцитами. Реакция агглютинации ставилась в
солевой среде при комнатной температуре. После инкубации в течение 45 минут
пробирки центрифугировали 2 минуты, а затем осадок рассматривали под
микроскопом [16]. Согласно данным Андрезена, признак Leа - носит
рецессивный характер. Многими исследователями установлена зависимость между
антигенами Le и способностью организма выделять групповые вещества АВН.
Ген секреции Se и ген Leb являются доминантными.
Законы наследственной передачи антигенных признаков системы Льюис до настоящего
времени остаются недостаточно изученными. Предполагается их связь с антигенами
системы АВО. Практически изоантигены данной системы используются в
судебно-медицинской экспертизе и реже в антропологии [6].
В 1937 и 1940 гг. Винером и Рейсом была открыта новая
изосерологическая система в организме человека. Это внесло существенный вклад в
антропологическую генетику и является важным фактором современной иммунологии.
Система оказалась весьма сложно организованной, детерминирована тремя парами
аллельных генов, которые в свою очередь контролируют наследование 8 гаплотипов.
В клинической практике до недавнего времени определялось только два
альтернативных признака по наличию или отсутствию в организме антигена Д. При
наличии этого антигена человек является резус-положительным, при его отсутствии
- резус-отрицательным [17].
Другие специфические обособленные антигены системы резус С,
Е, с и е, наличие которых удалось доказать при помощи изоиммунных человеческих
сывороток, также широко используются в антропологии, судебной медицине, клинике
[18]. Существует несколько методов определения антигенов резус в эритроцитах.
Систему резус мы определяли на чашках Петри в водяной бане при температуре +40оС
в течение 5 минут с гемагглютинирующей сывороткой антирезус. На основании
агглютинации делают вывод о том, является кровь резус-положительной или
резус-отрицательной [14].
Различные варианты антигенов резус встречаются у людей
неодинаково часто. В 1944 г. Фишер и Рейс доказали наличие внутренней
генетической связи между всеми антигенами резус. Ими же была предложена новая
номенклатура, которая в настоящее время широко распространена [18].
Антигены системы АВО наиболее хорошо изучены и были открыты
Ландштейнером в 1900 г. самыми первыми из групп крови [2]. Ландштейнер, проводя
свои долгие эксперименты, выявил не все четыре известные нам сегодня основные
группы крови, а три: 0 (I), A(II) и B(III). Четвертую группу крови, или AB(IV),
в 1907 году открыл чешский врач Якоб Янский и, на основании сделанного
Ландштейнером и им, предложил классификацию групп крови: I-IV. То, каким
антигеном будет обладать конкретный человек, зависит от трех генов(аллелей) A,
B и 0, причем они находятся в одном локусе двух гомологичных хромосом,
конкретно в длинном плече 9-й хромосомы (точная прописка 9q34.1-q34.2). И тут
надо пояснить: здесь мы имеем дело не с классической, то есть двухаллельной,
системой гена, а с системой множественных аллелей в данном случае трех. Трех в
принципе. Но всякий человек может обладать только двумя из трех возможных
аллелей данного локуса (по одному в каждой из гомологов 9-й хромосомы).
Группируя их попарно, теоретически получаем шесть возможных комбинаций
генотипов: AA, A0, BB, B0, AB, 00.
Генотипов шесть, но ведь групп крови системы AB0 всего четыре?
Правильно. Ибо аллели A и B доминантные, а аллель 0 рецессивный (его еще называют
«молчащим» геном), поэтому, например, при генотипе A0 группа крови
фенотипически будет второй A(II). А в случае генотипа AB? Кто кого подавляет? В данном случае никто и
никого. Это еще один генетический нюанс так называемые кодоминантные аллели:
каждый образует собственный антиген, что и имеет место при группе крови IV.
Иммунологически можно определить только фенотип группы крови
с первой по четвертую. А как распознать AB0-генотип человека, если
в этом возникает необходимость? Это делается с помощью анализа родословных.
Если известна группа крови потомка и его матери, то с определенной вероятностью
можно предположить, какой групой крови (и ее генотипом) может или не может
обладать отец ребенка. Наглядно это выглядит следующим образом:
Генотип родителей Возможные генотипы потомства
+ 00 00
00 + AA A0
+ A0 00, A0
+ BB B0
+ B0 00, B0+ AA AA+ BB AB+ B0 AB, A0, B0, 00+ AA
AA, A0+ A0 AA, A0, 00+ BB BB+ BB BB, B0+ B0 BB, B0, 00+ AB AB, AA, BB+ 00 A0,
B0+ AA AA, AB+ A0 AA, A0, AB, B0+ BB BB, AB+ B0 BB, B0, AB, A0
Таким образом, группы крови, во-первых, наследуются, и,
во-вторых, характер их наследования подчиняется строгим закономерностям
передачи простых (моногенных, или однолокусных) признаков из поколения в
поколение.
В основе того, что нас сейчас интересует, находится
гликопротеид с олигосахаридными цепями. Он универсален, первичен, то есть
образуется у любого человека, вне зависимости от его группы крови. А вот то,
каковой будет групповая принадлежность каждого из нас, напрямую связано с
терминальными последовательностями в олигосахаридных цепях гликопротеидов.
Если к такой основе присоединяется N-ацетил-D-галактозамин,
значит, в конечном счете возникает то, что названо A-специфичностью, или, на
уровне фенотипа, группой крови A(II); если терминальной последовательностью
окажется D-галактоза, то возникает B-специфичность, или группа крови B(III). Ну
а если к олигосахаридным цепям базисного гликопротеида не присоединится ни
первый, ни второй продукт, то тут мы имеем дело с ситуацией, названной
H-специфичностью то есть исходной, или с ситуацией предшественника; это и есть
группа крови 0 (I).
Аллели A и B определяют образование специфических
гликозилпереносящих ферментов (трансфераз), которые на конечных стадиях синтеза
катализируют присоединение группоспецифических молекул к концам углеводных
цепей соответственно либо N-ацетил-D-галактозамина, либо D-галактозы, либо
обеих этих молекул (генотип AB), либо, в случае если человек гомозиготен по
аллелю 0, никакого присоединения нет вовсе, поскольку эти трансферазы у него
наследственно отсутствуют.
Повторим: независимо от того, каким из аллелей локуса AB0
обладают разные люди, синтез макромолекул у них происходит одинаково вплоть до
стадии образования вещества с H-специфичностью (синтез этого антигена
определяет ген, расположенный в 19-й хромосоме). Ну а далее все зависит от
того, кто из нас какие транферазы получит от родителей. Точнее (то есть
изначально), какие гены системы AB0 [27].
Бернштейн в 1924 г. предложил схему генетического контроля
этой системы: три аллельных гена, из них два кодоминантных аллеля А и В и один
рецессивный аллель О, которые при сочетании по два дают шесть генотипов: АА,
АО, ВВ, ВО, АВ и ОО, проявляющихся в четырех фенотипах: А, В, АВ и О.
Сейчас установлено, что антиген Н, который является
химическим предшественником антигенов А и В и конечным продуктом у людей с
группой крови О, контролируется отдельным локусом Н. При генотипах HH и Hh
образуется вещество Н, а при генотипе hh оно не образуется даже в том случае,
если эти гомозиготы являются носителями аллелей локуса АВО. Такие индивиды, у
которых не обнаружено антигенов АВН ни в эритроцитах, ни в секретах, впервые
наблюдались в нескольких семьях в Бомбее, их фенотип был назван «бомбейским». В
сыворотке крови лиц с таким фенотипом содержатся в высоком титре анти-А, анти-В
и анти-Н вещества.
А- и В-антигены имеют несколько вариантов. Антиген А
подразделяется на два основных варианта: А1 и А2, причем
антиген А2 имеет меньшую антигенную активность и проявляется слабо,
особенно в группе А2В. Гетерозиготы А1А2
серологически неотличимы от А1А1 и А1О. В
группе А открыты также антигены А3, А4, Аm, Ао,
Ах, Аz, Аg, Ае и Аend,
которые встречаются крайне редко. Есть также количественные варианты антигена,
например, промежуточный между А1 и А2, который
встречается у африканских народов, а также такой антиген, как Аhel,
который встречается с антигенами А1 и А2. Разновидностей
антигена В меньше и встречаются они реже: В2, В3, В4,
Вх; иногда при отсутствии в генотипе гена В, слабую В-подобную
реакцию дает антиген из E.coli. Некоторые разновидности антигенов А и В
контролируются, по-видимому, соответствующими мутантными аллелями, а другие,
вероятно, являются результатом действия гена-супрессора.
Описано два случая «химер», когда при рождении в течение всей
жизни у человека находятся эритроциты, реагирующие как с сывороткой анти - А, так
и с анти-О. Это явление объясняется тем, что у близнецов с развитыми
сосудистыми связями во время внутриутробной жизни эритроциты группы А поступают
в организм к близнецу с группой О, «приживаются» там и воспроизводятся в
течение всей жизни. Антиген Н содержится у всех лиц, в наибольшем количестве у
лиц с группой крови О, в наименьшем - с группой крови А1В.
В сыворотке крови не иммунизированных к антигенам системы АВО
людей содержатся так называемые нормальные агглютинины - антитела к
группоспецифическим веществам системы АВО. Они вырабатываются к отсутствующим у
данных лиц антигенам. Так, у индивидуума с группой крови 0 имеются анти-А и
анти-В антитела, у лиц с группой крови А и В-соответственно антитела анти-В- и
анти-А, при группе крови АВ эти антитела отсутствуют вовсе. Антитела
представляют собой 19s иммуноглобулины (IgG). Важными отличительными свойствами
этих IgG-агглютининов являются их способность проникать через плаценту и
большая агглютинационная сила. Нормальные антитела - полные IgM-антитела, а
иммунные антитела, как правило, являются неполными.
Антитела анти-Н иногда встречаются у людей с группами крови
В, А1, А1В. У большинства лиц, независимо от их групповой
принадлежности, анти-Н можно обнаружить в форме антител, реагирующих при
температуре от +4о до 0оС.
Происхождение нормальных агглютининов до сих пор не выяснено.
Некоторые исследователи полагают, что изоагглютинины образуются в результате
иммунизации антигенами, близкими по своей структуре к веществам А и В. Такие
антигены часто встречаются во многих продуктах питания и медикаментах. По
мнению других авторов, самостоятельное образование изоагглютининов у плода
генетически обусловлено.
Существуют данные в литературе о развитии антигенов в
онтогенезе. АВО-антигенная дифференцировка наблюдается в большинстве тканей
организма человека на самых ранних стадиях эмбриогенеза. Исследования,
проведенные М.А. Бронниковой и А.С. Гаркави, показали, что антигены А и Н
обнаруживаются в крови плода уже в возрасте 5 недель.
Известно несколько рецессивных аллелей, контролирующих на
различных этапах онтогенеза подавление биосинтеза этих антигенов. Однако ни
один из таких аллелей не получил достаточного распространения ни в одной из
обследованных популяций. Кроме того, АВО-локус функционирует практически в течение
всего онтогенеза, значит, потенциальные возможности регуляции его в раннем
онтогенезе не реализуются и поэтому можно предположить участие АВО-антигенов в
нормальном эмбриогенезе человека. Титр антигенов увеличивается с возрастом.
По данным М.А. Бронниковой и А.С. Гаркави, на ранних стадиях
онтогенеза у плода ни разу не был выявлен агглютинин, отсутствующий у матери.
При наличии в сыворотке у плода двух агглютининов - относительная выраженность
их находилась в соответствии с их выраженностью у матери. Указанные данные
свидетельствуют о том, что агглютинины проникают в кровь плода из крови матери.
По данным Фьюденберга и соавторов, иммуноглобулины IgM первыми из
иммуноглобулинов синтезируются в организме плода.
Раннее формирование антигенов АВО в организме плода в
сочетании с нормальным иммунологическим статусом женского организма
предопределяет возможность несовместимости по системе АВО между матерью и
плодом. Это приводит к конфликту, который может способствовать развитию у
младенца гемолитической болезни новорожденных, а также является причиной
недоношенности и спонтанных абортов. Явление несовместимости было открыто в
результате обследования 661 супружеской пары. Авторы обнаружили, что в браках 0
x A на 25% меньше детей с группой крови А, чем в реципрокных браках А х 0, и
объяснили это явление внутриутробной гибелью плодов с группой крови А. Конфликт
выражается в том, что антитела проникают из сыворотки крови матери в кровь
ребенка с группой крови А и вызывают агглютинацию его эритроцитов, что приводит
к гибели плода на ранних стадиях эмбриогенеза.
Современные данные (Головачев, 1983) свидетельствуют о двух
направлениях отбора при АВО-несовместимости, а не одного, как предполагалось
раньше. Один из векторов отбора направлен против несовместимости потомства, а
другой - против совместимого потомства. Направленность вектора зависит от
возраста матери и от количества родов.
Отсев на ранних стадиях развития плода не имеет клинических
проявлений, но на более поздних сроках беременности внутриутробная гибель плода
приводит к спонтанным выкидышам. Причем на частоту выкидышей в сроки
беременностей свыше 16 нед. АВО-несовместимость, по-видимому, не влияет. Отсев
на стадии зрелого плода преимущественно связан с гибелью от гемолитической
болезни новорожденных. АВО-гемолитическая болезнь новорожденных разной степени
тяжести встречается в 3-5% случаев, а ее тяжелые формы бывают редко. Различая
по степени выраженности болезни часто зависят от того, к какой возрастной
группе принадлежат матери новорожденных. Редкость тяжелых форм осложнений при
АВО-несовместимости объясняется существованием защитных механизмов. Одним из
таких механизмов является низкая активность А- и В-субстанций в эритроцитах
плода и новорожденного [11].
1.6
Полиморфизм гематологических признаков
Изучение внутрипопуляционного генетического полиморфизма
позволяет подойти к пониманию механизмов поддержания генетической структуры
популяций.
Существует несколько правил сравнения генетических систем
популяций в пространстве и времени: 1-сравнение популяций по генетическому
составу, например по группам крови, белковым фракциям, строению кожного
покрова, зубной системы и других известных к настоящему времени для человека
генетических признаков-маркеров; 2 - сравнение динамики генофонда больших более
стабильных популяций с аллелофондом малых популяций и выявление их генетической
структуры в зависимости от истории возникновения; 3 - исследование различий
между популяциями в зависимости от экологических особенностей и использованных
при обследовании признаков-маркеров [19].
Стабилизирующий отбор привел к закреплению в человеческих
популяциях сбалансированного наследственного полиморфизма.
Полиморфизм по многим локусам характерен для популяции
человека. Практически он приводит к тому, что (за исключением однояйцовых близнецов)
среди населения Земли невозможно найти двух генетически одинаковых людей[10].
Геногеографического исследования сельского населения
Белорусской ССР, описанное Микуличем, продолжалось с 1972 по 1984 г. Здесь
представлены данные о фенотипической изменчивости и генетической структуре 30
локальных популяций из 6 этнографических регионов. Основной акцент сделан на
изучение коренного населения белорусской национальности. Таким образом, было
сведено к минимуму давление миграций, влияние населения из других регионов
страны.
Сложившейся популяционной структуре коренного сельского
населения республики угрожают депопуляционные явления. Однако успели изучить
фенетический и генетический полиморфизм сельских популяций Белоруссии. Работ по
географической изменчивости наследственных изосерологических и гематологических
признаков на территории БССР ранее не проводилось. Основной задачей данного
исследования явилось изучение гетерогенности таких антропологически значимых
генетических систем, как ABO, - MN, Rh, P, Le, Hp, РТС, ЦС и др.
Весь материал был собран, систематизирован, обработан в
полевых и лабораторных условиях, проанализирован с помощью математической
статистики. При этом был выполнен последовательный анализ вначале распределения
фенотипов, а затем и частот генов. Результаты исследований приведены в
таблицах, а также на фено-и геногеографических картах, картах обобщенных
биологических расстояний. При исследовании особенностей географической
изменчивости каждого признака использованы шкалы с неодинаковым интервалом
частот в зависимости от их разброса между минимальными и максимальными
пределами и стандартными отклонениями межгрупповой изменчивости. Построение
карт осуществлено с помощью изолиний частот или обобщенных расстояний с учетом
локальной и этнографической принадлежности групп.
Число изученных территориальных групп практически по всем
генетическим системам одинаково и равняется 30, хотя объемы выборок не всегда
одинаковы, но достаточно представительны и статистически достоверны.
Система АВО. Скрининг по этой системе выполнен с помощью
гемагглютинирующих сывороток Белорусской республиканской станции переливания
крови. Каждый обследованный был определен как носитель одного из четырех
фенотипов. В системе АВО 3 гена ответственны за формирование 6 генотипов. 0 (I)
- группа крови является гомозиготной по гену r (О), AB(IV) - группа -
гетерозиготной по генам р(А) и q(B). Что касается двух других групп - A(II) и
B(III), то они могут оказаться или гомозиготными АА и ВВ или гетерозиготными АО
и ВО.
С учетом закономерности наследования этих генов на основании
закона Харди-Вайнберга выполнен статистический анализ. Последний показал, что
наблюдаемая частота фенотипов АВО повсеместно во всех локальных выборках
соответствует теоретически ожидаемым.
Анализ полученных результатов касается в первую очередь
амплитуды вариационных рядов, которая указывает на степень изменчивости каждого
из фенотипов среди 30 групп населения. АВ (IV) - группа крови, согласно
теоретическому ожиданию и по фактическим данным, в среднем по Белоруссии
равняется семи процентам с минимальным по сравнению с другими фенотипами
стандартным отклонением - 2,1. Минимальные частоты четвертой группы крови (3%)
обнаружены в Петриковской и Шкловской популяциях, максимальные (11%)-в
Малоритской и Верхнедвинской.
Генетически обусловленный полиморфизм наиболее отчетливо
проявляется в изменчивости популяций по второй и первой группам крови:
стандартное отклонение увеличивается до 5,3 и 5,1% соответственно. Амплитуда
колебаний между максимальными и минимальными показателями достигает 25%.
Мозаичность изменчивости по третьей группе крови значительно менее выражена.
По данным Бойда, Моуранта, Бунака, на территории Европы по
факторам крови АВО прежде всего и некоторым другим гематологическим
системам выделяются шесть географических ареалов: северо-западноатлантический,
среднезападноевропейский, скандинавский, северобалкано-карпатский,
средиземноморский и восточноевропейский. Последний характеризуется большим
размахом в гематологической типологии населения, поскольку здесь в различные
исторические эпохи часто возникали условия для формирования смешанных
антропологических типов.
Приблизительно одинаковая распространенность первой и второй
групп крови (в пределах 23-49%) во всех обследованных популяциях коренного
сельского населения подтверждает принадлежность его к западному варианту
восточноевропейского ареала. Особо следует отметить их максимальные
концентрации по европейской шкале в некоторых локальных популяциях, таких, как
Лунинецкая, Смолевичская и Чашникская по О(I), Брагинская, Хойникская и
Волковысская по A (II). Большое распространение группы В (III) характерно для
азиатских народов. 5 из 30 малых популяций отличаются наибольшим показателем -
25%: Ельская, Слуцкая, Чериковская, Полоцкая и Верхнедвинская.
При картографировании изменчивости в группах по отдельным
фенотипам нельзя ожидать проявления какой-либо закономерности. Таковая может
быть в распределении самих материальных носителей наследственности - генов.
Однако, несмотря на подобное утверждение, минимальные концентрации фактора В (III) демонстрируют
центростремительную направленность, а максимальные - центробежную (приложение
А).
Попытаемся проследить подобный принцип фенотипической
изменчивости по другим признакам гематологической системы АВО. Популяциям
вдоль Белорусской гряды и в предполесских районах в целом присуща относительно
низкая концентрация фенотипа В(III), подобно тому, как это имеет место у
большинства центрально-европеоидного населения. Среднебелорусская, близкая к
модальной в пределах стандартного отклонения частота третьей группы крови при
изолинейном картографировании приблизительно повторяет линии изменчивости
минимальной и максимальной концентраций, всегда оставаясь в промежутке между
ними. Мы уделили так много внимания этому признаку потому, что он выступает
одним из индикаторов монголоидно-европеоидной изменчивости на Евразийском
континенте в пределах обозримой истории его заселения. Возможно, что
приблизительно трехлопастная конфигурация направлений основных изофенлиний
зафиксирована с момента начала консолидации трех известных племенных союзов в
единую белорусскую народность.
Таким образом, организованная структура фенотипической
изменчивости населения Белоруссии при почти нормальных кривых распределения (о
чем свидетельствуют коэффициенты асимметрии и эксцесса), скорее, говорит об его
автохтонном происхождении. Сказанное подтверждают и изофенлиний A (II). Причем концентрации
фенотипа A (II), который наиболее характерен для палеоевропеоидного населения,
максимальны в бассейнах Припяти, Немана и в верховьях Днепра (приложение Б).
При всей неоднородности распределения факторов АВО населению
Подвинья и Посожья свойственны высокая частота В(III) - фенотипа и низкая A (II). Такая зависимость в
становлении доминантных признаков в указанных регионах могла сложиться
исторически в процессе постепенного заселения всей территории республики.
Особенно четко эта закономерность прослеживается в географии
распределения первой группы крови (рецессивной) с тенденцией к выделению двух
очагов типообразования - северного и южного (приложение В).
На фоне всей Белоруссии средневзвешенная встречаемость гена
r(О) не превышает 60%-ного уровня. Вариабельность по этому фактору значительна:
местами достигает европейского максимума - 70%. Группы коренного населения,
локализованные приблизительно на одной трети центральной территории республики,
образуют зону повышенной концентрации рецессивного гена и самые низкие
концентрации доминантных генов системы АВО. От обнаруженного центра во всех
направлениях наблюдается уменьшение одних частот и увеличение других всегда
через промежуточные модальные концентрации.
Система резус Rh (CDE). За последние годы в мировой и
советской литературе появляются все новые сведения о распределении
многочисленных фенотипов и гаплотипов системы резус. Концентрация этих факторов
весьма не равномерна не только среди малых популяций, но и среди больших рас.
Так, преобладание резус-положительности характерно для монголоидной,
американоидной и негроидной рас. В Европе относительно часто встречается
резус-отрицательность - иногда до 30-40% в изолированных локальных популяциях.
Среди коренного населения Белоруссии размах изменчивости
достигает 15% при сравнительно небольшом стандартном отклонении (3,4%).
Асимметрия и эксцесс указывают на почти нормальную вариационную кривую
распределения. На географической карте максимальные частоты гена
резус-отрицательности и рецессивного генотипа - (более 20%) выявлены в
основном среди центральных выборок. Намечается наличие двух геногеографических
зон с разграничительной перетяжкой в направлении от Молодечно через Пуховичи на
Бобруйск.
Рассматривая распределение других аллеломорфных гаплотипов
системы резус в Белоруссии, можно отметить, что по частоте встречаемости после cde (39,2%) следует CDe (32,4%). Относительно
часто (более 10%) выявляются хромосомы cDE (12,6%) и cDe (11,4%). Из оставшихся
четырех хромосом только концентрация Cde превышает 1%-ный уровень
и достигает величины 3,7%. Сочетание CdE не выявлено нами вовсе,
так же как и другими исследователями в Советском Союзе и за рубежом.
Особенностью коренного сельского населения БССР является
также очень редкая (менее 1%) встречаемость cdE и СDЕ. Средняя частота аллеля
d(rh) у белорусов не превышает 40%, что соответствует центрально-европейским
концентрациям. Если факт нарастания доминантного гена D (Rh°) по направлению к
периферийным районам исследованной территории рассматривать как свидетельство
присутствия монголоидной примеси, то последняя присутствует как на западе, так
и на востоке республики. Поэтому, мы склонны такое влияние считать весьма
проблематичным, отдавая предпочтение автохтонному происхождению такой
наследственной изменчивости. [4].
1.7
Группы крови АВО и инфекционные заболевания
гематологический генетика полиморфизм кровь
Система АВО является системой сбалансированного полиморфизма.
В литературе приводятся многочисленные данные о разной резистентности фенотипов
различных систем эритроцитарных антигенов к инфекционным и неинфекционным
заболеваниям. Такую различную приспособленность лиц с различными фенотипами АВО
В.П. Эфроимсон считает результатом действия отбора мутантных генов. Существуют
гипотезы, объясняющие неравномерное распределение групп крови АВО на земном
шаре влиянием эпидемий таких инфекционных заболеваний, как чума, оспа и холера.
Было показано, что лица с группой крови О особо восприимчивы к холере и чуме, а
с группой А - к оспе. Разные реакции на инфекционные агенты автор связывает с
наличием у некоторых возбудителей инфекций антигенов, сходных с антигенами
крови у человека. Например, палочка чумы содержит антиген, подобный антигену О,
а вирус оспы - антигену А. Лица, имеющие соответствующие антигены, будут менее
устойчивы к указанным инфекциям, так как их организм не сразу «распознает»
антигены возбудителей как чужие и реагирует на них очень слабо. Действительно,
обращает на себя внимание низкая частота группы крови О у населения тех мест,
где свирепствовали эпидемии чумы: Индия, Северная Африка, Монголия, Турция.
Отмечена связь группы крови у детей в возрасте до 7 лет с
такими заболеваниями как паратиф, краснуха, скарлатина, коли-инфекция. Также
показано, что вирусным гепатитом чаще болеют люди с группой крови А, а менее
устойчивы к вирусу гриппа лица с группой О.У детей с группой крови А не
вырабатывается иммунитет против оспы даже при повторной вакцинации.
Статистически достоверно и показано многими авторами
преобладание у больных язвенной болезнью двенадцатиперстной кишки группы крови
О по сравнению с А. У больных язвой желудка с группой О выше уровень фруктозы в
моче и слюне. У детей с язвенной болезнью нет преобладания группы О. В случае
острого аппендицита отмечено, с одной стороны, повышение частоты группы А в
выборке больных, а с другой - у людей с группой В больше риск, что аппендицит
перейдет в перитонит, и чаще бывают нагноения при раневых повреждениях.
Раком желудка, поджелудочной железы, яичника, матки, слюнной
железы чаще болеют лица с группой крови А. Люди с группой крови В больше других
предрасположены к раку пищевода, полости рта, а с группой О - раку языка,
молочной железы, легких. Анализ распределения групп крови у больных с болезнями
опорно-двигательного аппарата показал понижение частоты фенотипа О и повышение
частоты АВ в группе больных остеохондрозом позвоночника; в группе больных
ревматизмом чаще встречается группа А; при поясничном остеохондрозе также
группа А.
Показана связь заболеваний дыхательных путей с группами
крови. У больных бронхиальной астмой достоверно увеличена частота группы О и
уменьшена - А, В и АВ; также повышена частота О у больных острой пневмонией.
Отмечено, что среди детей с врожденными пороками развития, а также умерших от
пневмонии, сепсиса, чаще встречаются группы А и В.
Риск заболеваний сердечно-сосудистой системы различен у людей
с разными группами крови: у больных атеросклерозом чаще встречается фенотип А и
реже - О; у людей с группой А повышена частота сердечно-сосудистых заболеваний
неревматического происхождения; наблюдается увеличение встречаемости группы А
по сравнению с В у больных с ишемической болезнью сердца; выявлена
отрицательная ассоциация группы О и положительная - группы А с инфарктом
миокарда, а также положительная связь А и отрицательная В с артериальной
гипертонией у мужчин.
По данным многих авторов, паралитическим полиомиелитом реже
болеют лица с группой B. У младенцев с фенотипом O дистрофия развивается реже,
чем при других группах крови. Отмечена тенденция к увеличению частоты группы А
у больных сахарным диабетом. В группе больных сифилисом соотношение фенотипов
B:O равно 1:5. У больных холециститом и желчнокаменной болезнью чаще встречается
группа А. Люди с группой А чаще болеют кожными заболеваниями.
Обширное исследование, предпринятое французскими учеными,
выявило существование ряда статистически достоверных корреляций определенных
фенотипов ABO со смертностью от таких болезней, как туберкулез, рак, диабет,
сердечно-сосудистые болезни, расстройства функций мозга, цирроз, грипп,
пневмония, бронхит, астма, а также от алкоголизма.
Корреляции между группами крови и определенными заболеваниями
могут занимать значительное место в процессах, поддерживающих полиморфизм.
Однако Штерн и В.П. Эфроимсон считают, что так как большинство болезней
неинфекционной природы поражают людей в пострепродуктивном возрасте, то они
незначительно влияют на относительное число аллелей в последующих поколениях. По
мнению Рида, связи между группами крови и неинфекционными болезнями,
по-видимому, свидетельствуют о том, что гены групп крови потенциально
подвержены действию естественного отбора. На подверженность групп крови АВО
естественному отбору указывает изменение их частот в экстремальных условиях
среды (например, в высокогорье), по сравнению с нормальными.
В.В. Бунак не считает окончательно установленной связь
кровяных групп с различными нормальными или аномальными свойствами организма,
кроме несовместимости матери и плода. В различных популяциях и возрастных
группах одной популяции знак ассоциации с данным заболеванием может меняться.
Система групп крови АВО подвержена возрастной динамике.
Показано, что до старческого возраста чаще доживают люди с группой крови О по
сравнению с А, так как первые меньше предрасположены к наиболее опасным и
распространенным в настоящее время болезням. Эта гипотеза была подтверждена
также при исследовании выборки геронтов в Москве.
В некоторых работах прослеживается неравномерное распределение
групп крови между мужчинами и женщинами одного возраста. В Дели группы крови А
и О чаще встречаются у женщин, а В-у мужчин. Однако, по данным других авторов,
в населении Индии частота групп крови А и О выше у мужчин.
Было показано, что многоплодие достоверно чаще имеет место в
браках A х A.
Система АВО, несомненно, является одной их основных систем
совместимости у человека, т.е. играет важную роль при переливании крови и
пересадке тканей и органов, а ее гены обладают выраженным плейотропным эффектом
в предрасположенности к некоторым заболеваниям [11].
Инфекционные заболевания оказывают специфическое воздействие
на иммунный ответ организма. Если группы крови АВО влияют на иммунный ответ, то
отбор, возникающий из-за дифференциальной восприимчивости к инфекционным
заболеваниям, может привести к дифференциальной детской и юношеской смертности
[20].
1.8 Система «человек-среда»
Рассматривая отношение геногеографии к экологии
народонаселения, будем исходить из понимания предмета экологии человека как изучения
связей и взаимодействий человека с окружающим миром. Изучается ли адаптация
человека к условиям природной среды и среды, создаваемой деятельностью самого
человека, или воздействие урбанизации на человека, изучаются ли
хозяйственно-культурные и этнокультурные ландшафты как человеческие экосистемы,
или условия сохранения равновесия в системе «человек - среда», изучаются ли
демографические последствия экономического развития, научно-технического
прогресса и преобразований в обществе, - тем самым открываются и познаются все
новые многообразные связи и взаимодействия человека с окружающим его миром
[12].
Охрана наследственности человека в условиях резко меняющейся
среды - привлекает сегодня всеобщее внимание [21]. В ноосфере, в отличие от
биосферы, такое многообразие связей не ведет к состоянию равновесия, поскольку
воздействия, испытываемые человеком со стороны изменяемой им окружающей среды,
изменяют и его самого. Поэтому в центре всего круга проблем экологии человека
стоят такие связи и взаимодействия в системе «человек - мир», которые вызывают
изменения в самом человеке. Среди этих изменений могут быть и такие, что имеют
долгосрочные последствия для человека, то есть, изменения генетического
характера. На присутствие генетического контекста в экологии человека указывают
те фундаментальные проблемы нормальной генетики человека, исследование которых
лежит явно на пересечении генетики с экологией. Одна из них - наследственный
полиморфизм человека, а другая - человеческий генофонд. Наследственный
полиморфизм людей - это первоисточник индивидуального и личностного
многообразия, без которого немыслимо существование и процветание человеческого
общества. Одновременно наследственный полиморфизм - свидетельство
предшествующей эволюции человека и условие ее продолжения в соответствии с
требованиями окружающей среды. Эта генетическая проблема, безусловно,
пересекается с проблемами экологии человека, поскольку генетический полиморфизм
есть особо реактивное состояние наследственности человека, чувствительное к
малейшим изменениям в окружающей среде, будь то природной, или социальной.
Острота этой чувствительности связана именно с многообразием людских генотипов.
Если считать предметом экологии человека его связи с окружающим миром, то не
будет преувеличением сказать, что генетическое многообразие человека
конвариантно многообразию этих связей, посредством которых возникает и
регулируется.
Геногеография, в свою очередь, целиком основывается на
информации о наследственном полиморфизме населения, создает картографические
модели распространения и распределения этой информации в населении, исследует
свойства этих моделей и выявляет географические закономерности эволюции
наследственного полиморфизма под действием факторов природной и социальной
среды. Таким образом, в контексте наследственного полиморфизма связь между
геногеографией и экологией населения достаточно очевидна.
Проблема генофонда в экологическом ключе имеет свое особое
звучание как проблема главного жизненного ресурса экосистемы «человек - среда».
Если другие энергетические ресурсы могут быть в разной мере доступны, в разной
мере исчерпаны, в разной степени эффективны, то, по-видимому, должны быть
эквиваленты этих различий в свойствах и состоянии генофонда как ресурса жизни
экосистемы. В полной мере это предстоит выявить именно с помощью геногеографии,
но крайние состояния могут быть уяснены и дедуктивным путем.
Из генофонда черпаются уникальные генные сочетания -
человеческие генотипы. В него же гены и возвращаются, если генотип (точнее его
обладатель) выдержал все испытания, которым подвергла его среда обитания, и
оставил потомству свои гены. Число и исход этих испытаний, кроме случайностей,
определяются числом и характером связей в системе «человек - среда». Далеко не
все гены, почерпнутые из генофонда при формировании человеческого поколения,
возвращаются в том же числе и составе по завершении поколением своего цикла.
Число это может быть и больше (рост популяции) и меньше исходного (вымирание),
но даже при его постоянстве - состав генов в генофонде изменяется, и изменение
это есть реакция генофонда на состояние окружающей человека среды.
Одно из крайних состояний человеческого генофонда мыслимо
лишь теоретически: полная фиксация по всем генам и, следовательно, полная
идентичность и гомозиготность всех генотипов на всем ареале. Нежизнеспособность
такой популяции и такого общества очевидна. Возможно именно поэтому такое
состояние генофонда (достижимое при разведении лабораторных животных) ни разу
не наблюдалось в популяциях человека. Сложнее обстоит с противоположным состоянием
- максимального разнообразия по каждому из генов в составе генофонда, - но, все
же одно следствие очевидно, если считать, что видовое единство человечества
покоится на фундаменте мономорфных генов. Геногеографическому исследованию
доступны все промежуточные состояния генофонда как жизненного ресурса, и именно
геногеография может обнаружить распределение этих состояний в географическом
пространстве.
География состояний генофонда помимо факторов природной среды
в еще большей мере определяется факторами социальной среды. В качестве
передаточного механизма действует демографический механизм воспроизводства
народонаселения. Тип демографического развития народонаселения является
результирующим всех взаимодействий в экосистеме «человек - среда». Он различен
не только для разных исторических формаций человеческого общества, но и для
разных исторических культур.
Хотя экологический аспект геногеографии народонаселения
трудно отделить от исторического, все же, можно, следуя сложившемуся в
этнологии различению историко-культурных и хозяйственно-культурных типов
населения, считать, что культура хозяйствования в большей мере отражает тип
отношения общества к природной среде. Соответственно, исторически не
родственные группы населения могут стать однотипны в хозяйственно-культурном
плане, что имело место и в древности (типы охотников и рыболовов,
скотоводов-номадов, земледельцев и т.д.), а в еще большей мере при
распространении современной цивилизации. Хозяйственно-культурная типология
систематизирует географическое разнообразие населения по признакам способа
освоения человеком природной среды и характеризует не биологическую, а именно
хозяйственно - культурную адаптацию населения к условиям среды. Но и такая
адаптация может иметь генетические последствия, правда, не в виде
распространения или исчезновения тех или иных генов, а в виде изменений
состояния генофонда. Разнообразие хозяйственно-культурных типов населения - это
разнообразие имевших место в истории способов взаимодействия человека со средой
обитания. Вклад этого разнообразия в дифференциацию генофонда по его состояниям
наиболее полно можно выявить и охватить именно средствами геногеографии.
Урбанизация - классическая экологическая проблема, в которой
присутствует генетический контекст. У процесса урбанизации множество
парадоксальных генетических последствий, которые обнаруживаются уже при первых,
пока еще единичных, исследованиях. Весь объем и характер влияний урбанизации,
на состояние генофонда населения еще предстоит выяснить. С другой стороны
география именно городских поселений является разработанной областью
экономической и социальной географии. Геногеография способна соединить эти два
подхода и тем самым проследить влияние центров урбанизации на распределение
состояний генофонда в географическом пространстве.
Оперируя с наиболее поддающейся математической формализации
генетической информацией о населении, находящемся в той или иной экологической
ситуации, геногеография тем самым создает своими - генокартографическими -
средствами легко воспринимаемые образы состояния и процессов в экосистеме [12].
Законы природы подчеркивают настоятельную необходимость
перестать загрязнять окружающую среду, вырубать леса, истощать почвы или
промышлять диких животных в размерах превышающих их естественную способность к
самовосстановлению [21].
2. Объект и методика исследования
.1 Методы исследования
Определение групп крови проводилось на базе лаборатории
ГОИКБ.
Строгое постоянство наследуемости групповых признаков АВО
позволило вычислить эмпирическое и теоретическое распределение фенотипов и их
аллелей по формулам Бернштейна[19]. Достоверность статистических показателей
оценивалась по критериям Стьюдента [22]. Установлено, что все выборки находятся
в генетически оптимальном равновесии в соответствии с законом Харди-Вайнберга
[6].
Групповую принадлежность определяли с помощью реакции
агглютинации при помощи стандартных гемагглютинирующих сывороток.
Систему группы крови АВ0 составляют два групповых
эритроцитарных агглютиногена (А и В) и два соответствующих антитела - агглютинины
плазмы альфа (анти-А) и бета (анти-В). Различные сочетания антигенов и антител
образуют 4 группы:
Группа 0 (I) - на эритроцитах отсутствуют групповые
агглютиногены, в плазме присутствуют агглютинины альфа и бета; группа А(II) -
эритроциты содержат только агглютиноген А, в плазме присутствует агглютинин
бета; группа В(III) - эритроциты содержат только агглютиноген В, в плазме
содержится агглютинин альфа; группа АВ(IV) - на эритроцитах присутствуют
антигены А и В, плазма агглютининов не содержит[23].
Определение группы крови проводится при температуре +15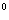 - +25С на маркированной плоскости (тарелке) белого цвета, где
записывается фамилия лица, у которого определяется кровь, согласно международной
классификации маркируются точки нанесения сыворотки. Используются стандартные
сыворотки двух серий групп О(I), А(II), В(III). После их нанесения получается
шесть капель, расположенных в два ряда в следующем порядке: О(I), А(II),
В(III). Каждая сыворотка наносится своей пипеткой. Исследуемая кровь углами
предметного стекла или стеклянными палочками вносится в сыворотку в количестве
в десять раз меньшем и тщательно перемешивается. Можно перемешивать кровь с
сывороткой одним и тем же углом предметного стекла (или стеклянной палочкой),
но в этом случае после каждого перемешивания необходимо промывать стекло в воде
и насухо вытирать. За ходом реакции, в общей сложности, наблюдают не менее 5-ти
минут, периодически покачивая тарелку. В капли, где к третьей минуте наблюдения
произошла агглютинация, вносят по капле физиологического раствора и наблюдают
еще две минуты. Добавление физиологического раствора не разрушает истинной
агглютинации и позволяет отдифференцировать ее от псевдоагглютинации,
обусловленной самопроизвольным склеиванием эритроцитов в монетные столбики.
Результат оценивается по наличию агглютинации. Эритроциты группы О(I) не
содержат антигенов и в реакцию агглютинации не вступают. Эритроциты группы
А(II) не дают агглютинации только с сывороткой своей группы, эритроциты В(III)
также не дают агглютинации только с сывороткой своей группы. Если во всех
сыворотках произошла агглютинация, то для исключения неспецифической
панагглютинации и подтверждения, что это эритроциты IV группы крови, следует дополнительно
исследовать их стандартной сывороткой группы АВ (IV). Для этого используется
одна сыворотка, исследование проводится с соблюдением всех выше указанных
условий. Сыворотка группы АВ(IV) не содержит никаких антител, а поэтому не
должна давать агглютинации. При определении групп крови следует обратить особое
внимание на сроки чтения результата - они не должны быть менее 5-ти минут. Так
разновидность второй группы крови А(II) дает агглютинации, как правило, раньше,
чем обычно - к исходу 3-4-ой минуты (таблица 1).
- +25С на маркированной плоскости (тарелке) белого цвета, где
записывается фамилия лица, у которого определяется кровь, согласно международной
классификации маркируются точки нанесения сыворотки. Используются стандартные
сыворотки двух серий групп О(I), А(II), В(III). После их нанесения получается
шесть капель, расположенных в два ряда в следующем порядке: О(I), А(II),
В(III). Каждая сыворотка наносится своей пипеткой. Исследуемая кровь углами
предметного стекла или стеклянными палочками вносится в сыворотку в количестве
в десять раз меньшем и тщательно перемешивается. Можно перемешивать кровь с
сывороткой одним и тем же углом предметного стекла (или стеклянной палочкой),
но в этом случае после каждого перемешивания необходимо промывать стекло в воде
и насухо вытирать. За ходом реакции, в общей сложности, наблюдают не менее 5-ти
минут, периодически покачивая тарелку. В капли, где к третьей минуте наблюдения
произошла агглютинация, вносят по капле физиологического раствора и наблюдают
еще две минуты. Добавление физиологического раствора не разрушает истинной
агглютинации и позволяет отдифференцировать ее от псевдоагглютинации,
обусловленной самопроизвольным склеиванием эритроцитов в монетные столбики.
Результат оценивается по наличию агглютинации. Эритроциты группы О(I) не
содержат антигенов и в реакцию агглютинации не вступают. Эритроциты группы
А(II) не дают агглютинации только с сывороткой своей группы, эритроциты В(III)
также не дают агглютинации только с сывороткой своей группы. Если во всех
сыворотках произошла агглютинация, то для исключения неспецифической
панагглютинации и подтверждения, что это эритроциты IV группы крови, следует дополнительно
исследовать их стандартной сывороткой группы АВ (IV). Для этого используется
одна сыворотка, исследование проводится с соблюдением всех выше указанных
условий. Сыворотка группы АВ(IV) не содержит никаких антител, а поэтому не
должна давать агглютинации. При определении групп крови следует обратить особое
внимание на сроки чтения результата - они не должны быть менее 5-ти минут. Так
разновидность второй группы крови А(II) дает агглютинации, как правило, раньше,
чем обычно - к исходу 3-4-ой минуты (таблица 1).
Таблица 1 - Возникновение агглютинации эритроцитов (+) при
смешивании крови со стандартными сыворотками
|
Группы крови
|
Антигены
эритроцитов
|
Антитела
сыворотки
|
Реакция
эритроцитов на стандартную сыворотку крови группы
|
|
|
|
О
|
А
|
В
|
АВ
|
|
0 (I)
|
О
|
анти-А анти-В
|
-
|
+
|
+
|
+
|
|
А(II)
|
А
|
анти-В
|
-
|
-
|
+
|
+
|
|
В(III)
|
В
|
анти-А
|
-
|
+
|
-
|
+
|
|
AB(IV)
|
АВ
|
-
|
-
|
-
|
-
|
-
|
Оценка результатов определения групп крови при помощи
стандартных гемагглютинирующих сывороток. Знаком плюс (+) обозначено наличие
агглютинации, знаком минус (-) - её отсутствие [24].
Активность реакции агглютинации, ее наличие или отсутствие
регистрировались в протоколах обследования (рисунок1) [11].
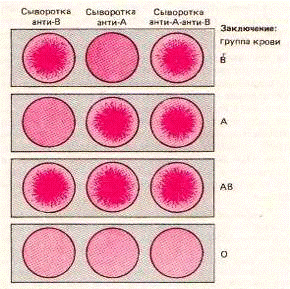
Рис. 1 Реакция агглютинации
В ходе исследования проведены вычисления аллельных и генных
частот определяющих генетическую структуру для генотипически неоднородных групп
крови II и III. Оценка частот (p, q, r) генов А, В, 0 проводилась по известным
формулам Ф. Бернштейна (Mourant Е.А., 1976; Ли, 1978) следующим образом.
Находим предварительные оценки частот генов 0, А и В:
r' (O) = 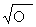 ,
,
p’ (A) = 1-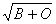 ,
,
q (B) = 1- 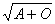 .
.
где 0', A' и B' - частоты фенотипов, т.е. отношение числа лиц
с определенным фенотипом к объему выборки.
Когда сумма предварительных частот генов не равна 1, вводим
поправку
= 1 - (r» + p» + q»).
Далее вычисляем окончательные (уточненные) оценки частот
генов:
r = (r’ + 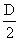 ) (1 + ),
) (1 + ),
p = p’(1 + ),
q = q’(1 + ),+ p + q = 1.
Эти формулы справедливы для оценки частот генов групп крови АВ0.
На основе данных изосерологических исследований и формул
Бернштейна, используя математический аппарат и вычислительную технику, дан
сравнительный анализ генетической структуры населения Полесья по генам,
контролирующим группы крови АВО[4].
3. Результаты исследования и их обсуждение
Все обследованные являются коренными жителями Белоруссии (в III поколениях), это было
выяснено с помощью генеалогического метода, метода анкетирования и посемейного
опроса. Объем выборки составил 79 человека, проживающие в г. Гомеле.
Распределение групп крови по фенотипам отражено в таблице 2.
Таблица 2 - Фенотипическое распределение групп крови среди
белорусов в III поколении г. Гомеля
|
N
|
Население г.
Гомеля
|
Год рождения
|
Место рождения
|
Группа крови
|
|
1
|
Прокопцова О.А.
|
1982
|
Гомель
|
A(II)
|
|
2
|
Блоцкая И.М.
|
1982
|
Гомель
|
O
(I)
|
|
3
|
Ракова А.В.
|
1982
|
Гомель
|
B
(III)
|
|
4
|
Васьковцова
А.В.
|
1982
|
Гомель
|
B
(III)
|
|
5
|
Шпаньков В.В.
|
1975
|
Гомель
|
B
(III)
|
|
6
|
Яроцкий В.И.
|
1975
|
Гомельск.обл.,
д. Моклищи
|
AB(IV)
|
|
7
|
Олейник А.А.
|
1970
|
Гомель
|
O (I)
|
|
8
|
Воробей Ю.А.
|
1981
|
Гомель
|
O (I)
|
|
9
|
Борисов А.Г.
|
2001
|
Гомель
|
B (III)
|
|
10
|
Гагарина М.А.
|
2002
|
Гомель
|
A(II)
|
|
11
|
Субботина М.А.
|
1981
|
Гомель
|
AB(IV)
|
|
12
|
Марков А.В.
|
2000
|
Гомель
|
AB(IV)
|
|
13
|
Дерюжков И.С.
|
2001
|
Гомель
|
B
(III)
|
|
14
|
Бондарев М.С.
|
2000
|
Гомель
|
B
(III)
|
|
15
|
Кожемякина Е.О.
|
1975
|
Гомель
|
B
(III)
|
|
16
|
Веренчикова
В.Г.
|
1968
|
-
|
O
(I)
|
|
17
|
Савостьянова
Л.Н.
|
1983
|
Гомель
|
O
(I)
|
|
18
|
Семченко Д.В.
|
1977
|
Гомель
|
A(II)
|
|
19
|
Ткач А.В.
|
1975
|
Гомель
|
AB(IV)
|
|
20
|
Синева В.П.
|
1971
|
Гомель
|
O
(I)
|
|
21
|
Корнеев Р.В.
|
1994
|
Гомель
|
O
(I)
|
|
22
|
Коробущенко
С.А.
|
1967
|
Гомель
|
A(II)
|
|
23
|
Горбачева Т.В.
|
1977
|
Рогочев
|
O
(I)
|
|
24
|
Бондаренко Т.Я.
|
1963
|
Добрушск. р-н,
д. Логуны
|
|
25
|
Перекрестова
Е.С.
|
1977
|
Гомель
|
A(II)
|
|
26
|
Саприко Е.Л.
|
1979
|
Гомель
|
A(II)
|
|
27
|
Грачева О.М.
|
2002
|
Гомель
|
A(II)
|
|
28
|
Ковалева Н.А.
|
1977
|
Гомель
|
B
(III)
|
|
29
|
Волоткович Т.А.
|
1980
|
Гомель
|
B
(III)
|
|
30
|
Громыко П.С.
|
1982
|
Гомель
|
A(II)
|
|
31
|
Кирсанова Т.Б.
|
1976
|
Гомель
|
O
(I)
|
|
32
|
Петкевич Р.В.
|
2001
|
Гомель
|
A(II)
|
|
33
|
Ковалев В.А.
|
2001
|
Гомель
|
A(II)
|
|
34
|
Громыко Л.А.
|
2001
|
Гомель
|
O
(I)
|
|
35
|
Чекан О.Н.
|
1978
|
д. Юровичи
|
A(II)
|
|
36
|
Бондаренко В.А.
|
2001
|
Гомель
|
O (I)
|
|
37
|
Чернявский Е.Е.
|
2001
|
Гомель
|
A(II)
|
|
38
|
Лукашевич К.В.
|
2000
|
Гомель
|
A(II)
|
|
39
|
Дубровская О.С.
|
1986
|
Гомель
|
В(III)
|
|
40
|
Томазова В.В.
|
1975
|
Гомель
|
A(II)
|
|
41
|
Шаклеина Н.А.
|
1983
|
Гомель
|
A(II)
|
|
42
|
Тимашкова С.С.
|
1981
|
Гомель
|
O
(I)
|
|
43
|
Ядченко Н.И.
|
1982
|
Гомель
|
A(II)
|
|
44
|
Мельникова И.И.
|
1982
|
Гомель
|
O
(I)
|
|
45
|
Никонович С.Н.
|
1977
|
Гомель
|
B
(III)
|
|
46
|
Тарасова О.Л.
|
1973
|
Гомельск.р-н,
д. Пионер
|
B(III)
|
|
47
|
Васильева С.А.
|
1967
|
д. Ченки
|
А(II)
|
|
48
|
Шаклеина Н.А.
|
1983
|
Гомель
|
А (II)
|
|
49
|
Кочкин В.М.
|
1947
|
Гомель
|
О(I)
|
|
50
|
Бардина С.И.
|
1973
|
Гомель
|
О(I)
|
|
51
|
Климович С.А
|
1983
|
Бобруйск
|
А(II)
|
|
52
|
Бондарь И.В.
|
1982
|
Солигорск
|
АВ(IV)
|
|
53
|
Помозова В.В.
|
1976
|
г/п Костюковка
|
А(II)
|
|
54
|
Клобук В.В.
|
1985
|
г. Гомель
|
А(II)
|
|
55
|
Никитенко А.Г.
|
1971
|
г. Гомель
|
А(II)
|
|
56
|
Никитина Н.С.
|
1982
|
г. Гомель
|
В(III)
|
|
57
|
Ермашневич О.И.
|
1979
|
г. Гомель
|
В(III)
|
|
58
|
Стеланович В.М.
|
1962
|
г. Гомель
|
В(III)
|
|
59
|
Ковалева Е.П.
|
1983
|
г. Гомель
|
В(III)
|
|
60
|
Сенчилова И.Н.
|
1974
|
Гомельск.р-н.
|
О(I)
|
|
61
|
Сатырева И.В.
|
1982
|
г. Гомель
|
О(I)
|
|
62
|
Валюшенко И.Г.
|
1982
|
г. Гомель
|
А(II)
|
|
63
|
Хомиченко Н.П.
|
1983
|
г. Гомель
|
О(I)
|
|
64
|
Шпанькова Д.А.
|
2002
|
г. Гомель
|
А(II)
|
|
65
|
Ремова А.С.
|
1981
|
г. Гомель
|
О(I)
|
|
66
|
Рыдзевская В.В.
|
2003
|
г. Гомель
|
О(I)
|
|
67
|
Зайцева Е.В.
|
1980
|
г. Гомель
|
А(II)
|
|
68
|
Синицына Т.Г.
|
1983
|
г. Гомель
|
А(II)
|
|
69
|
Павленок Д.А.
|
2002
|
г. Гомель
|
О(I)
|
|
70
|
Никитенко Д.А.
|
2003
|
г. Гомель
|
О(I)
|
|
71
|
Ещенко Е.Н.
|
1983
|
г. Гомель
|
А(II)
|
|
72
|
Полекова М.В.
|
1979
|
г. Гомель
|
О(I)
|
|
73
|
Гончарова Н.В.
|
1982
|
г. Гомель
|
В(III)
|
|
74
|
Якубович С.Г.
|
1982
|
г. Гомель
|
В(III)
|
|
75
|
Таранова С.А.
|
1967
|
г. Гомель
|
АВ(IV)
|
|
76
|
Жигалова Ж.М.
|
1973
|
г. Гомель
|
А(II)
|
|
77
|
Ревуцкая М.А.
|
1978
|
г. Гомель
|
А(II)
|
|
78
|
Озолынч Г.И.
|
2003
|
г. Гомель
|
А(II)
|
|
79
|
Верочинская
О.В.
|
1980
|
г. Гомель
|
А(II)
|
|
80
|
Рачковская О.В.
|
1982
|
г. Гомель
|
А(II)
|
|
81
|
Марченко Н.И.
|
1975
|
г. Гомель
|
А(II)
|
|
82
|
Cуднеко С.В.
|
1976
|
г. Гомель
|
О(I)
|
|
83
|
Денисенко Л.А.
|
1969
|
г. Гомель
|
В(III)
|
|
84
|
Ежикова Н.Ю.
|
1989
|
г. Гомель
|
О(I)
|
|
85
|
Сковородников
Ю.А.
|
1989
|
г. Гомель
|
О(I)
|
|
86
|
Пенязькова Е.М.
|
1973
|
г. Гомель
|
В(III)
|
|
87
|
Медведева Е.А.
|
1967
|
г. Гомель
|
А(II)
|
|
|
|
|
|
|
Для оценки частот фенотипов и генов в популяциях человека
используются принятые в популяционной генетике методы (Ниль, Шелл, 1958;
Mourant et al., 1976; Ли, 1978).
На основании полученных данных была рассчитана частота
аллелей, ответственных за группы крови АВО, с использованием математического
аппарата и вычислительной техники (таблица 3).
Таблица 3 - Распределение частот генов АВО среди белорусов в III поколении г. Гомеля
|
Популяция
|
N
|
O(I)
|
A(II)
|
B(III)
|
AB(IV)
|
r
|
p
|
q
|
|
население г.
Гомеля
|
87
|
26
|
34
|
20
|
7
|
0,55
±0,05
|
0,28 ±0,05
|
0,11 ±0,04
|
По литературным данным, распределение частоты аллеля О в
популяциях коренного населения Белоруссии составляет 50-55%, аллеля А 20-30% и
аллеля В 10-15% [25].
При сравнении данных нашего исследования с литературными, по
частоте аллеля О, А, В наблюдается соответствие, что совпадает с данными из
литературных источников (Фогель, Мотульски, 1990; Айала, Кайгер, 1988) [21,26].
Наглядно распределение частот генов выглядит следующим
образом (рисунок 2).
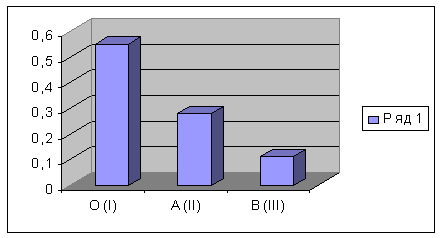
Рис. 2 Распределение частот генов
При анализе генетической структуры городского населения с
сельским, проведенное Микуличем, наблюдается следующее распределение частот
генов (таблица 4).
Таблица 4 - Количество фенотипов и частота генов АВО
сельского населения
|
Популяция
|
N
|
O(I)
|
A(II)
|
B(III)
|
AB(IV)
|
r
|
q
|
|
Сельское
население
|
92
|
38
|
30
|
19
|
5
|
0,52 ±0,04
|
0,260 ±0,03
|
0,140 ±0,02
|
Заключение
Строгое постоянство наследуемости групповых признаков АВО
позволило вычислить эмпирическое и теоретическое распределение фенотипов и их
аллелей по формулам Бернштейна. Достоверность статистических показателей
оценивалась по критериям Стьюдента [24]. Установлено, что все выборки находятся
в генетически оптимальном равновесии в соответствии с законом Харди-Вайнберга
[6].
В результете проведенных исследований было установлено, что
частота аллеля О составила 0,55, частота аллеля А - 0,28, частота аллеля В -
0,11. Данные частоты гомологичных вариантов генов, ответственных за определение
групп крови, отражают генетическую структуру городского населения под
воздействием объективных факторов, на протяжении определенного исторического
периода.
Популяционно-генетическое обследование коренных жителей
Белоруссии с использованием изосерологических тестов позволяет получить
информацию о частоте соответствующих аллелей, изучить геногеографию их
распределения, выявить взаимозависимость с другими биологическими,
историческими и общественными явлениями [4].
Список использованных источников
гематологический генетика полиморфизм кровь
1. Bernstein F. // Z. induct. Abstammungs -
und Vererbungs - lehre. 1930. №56. S. 223.
2. Hirschfeld L., Hirschfeld H. // Lancet.
1919. №2. P. 33.
. Landsteiner K. // Wien Klin. Wschr.
1901. №14. P. 19.
. Becrman L.A. Contribution to the
Physical Anthropology and Population Genetics of Sweden. Lund, 1959.
5. М.Е. Лобашев. Генетика. Изд. ЛГУ, 1967. - 753
с.
6. Микулич А.И. Геногеография сельского
населения Белоруссии. - Мн.: Наука и техника, 1989. - 182 с.
7. Микулич А.И. Наша генетическая память:
Современные аспекты антропогенетики. - Мн.: Наука и техника, 1987. - 72 с.
. Биология. В 2 кн. Кн.2: Учеб.для медиц.
спец. вузов/ В.Н. Ярыгин, В.И. Васильева, И.Н. Волков, В.В. Синельщикова; Под
ред. В.Н. Ярыгина. - 3-е изд., стер. - М.:Высш. Шк., 2000. -352 с.: ил.
9. Биология. В 2 кн. Кн.1: Учеб.для медиц.
спец. вузов/ В.Н. Ярыгин, В.И. Васильева, И.Н. Волков, В.В. Синельщикова; Под
ред. В.Н. Ярыгина. - 3-е изд., стер. - М.:Высш. Шк., 2000. - 448 с.: ил.
. Общая генетика: Учеб. для студ. биол.
спец. ун-тов (С.И. Алиханян, А.П. Акифьев, Л.С. Чернин - М.: Высш. Шк., 1985. -
448 с., ил.
11. Генофонд и геногеография народонаселения
/ Под ред. Ю.Г. Рычкова: Том 1. Генофонд населения России и сопредельных стран.
СПб.: Наука, 2000. 611 с.
12. Генофонд и геногеография народонаселения
/ Под ред. Ю.Г. Рычкова: Том 2. Геногеографический атлас населения России и сопредельных
стран. СПб.: Наука, (в печати).
13. Маккьюсик В. Генетика человека. М., 1967.
14. Landsteiner K., Levine P. // Prok.
Soc. Exp. Viol.
(N.Y.). 1927. №24. P. 600-602.
. Спицын В.А. Биохимический полиморфизм человека.
М.,
1985.
. Levine P. // Hum.biology. 1958. №14. P.
28.
. Wiener A. // Amer. J. Hum. G enet. 1952.
Vol. 4, №3. P. 271.
. Race R. // Nature. 1944. №153. P. 771
19. Яблоков А.В. Популяционная биология. М.,
1987.
21. Фогель Ф., Мотульски А.
Генетика человека: В 3-х т. Т. 2: Пер. с англ. - М.: Мир, 1990. - 378 с., ил.
22. Ракицкий Л.Ф. Биологическая статистика. -
Минск: Вышэйшая школа, 1973. - 318 с.
23. Энциклопедия клинических лабораторных тестов
под ред. Н.У. Тица. Издательство «Лабинформ» - М. - 1997 - 942 с.
. Руководство по медицине в 2-х томах. Гл.
редактор Р. Беркоу. - М - «Мир» - 1997. - т 1 - 1045с; т II - 872 c.
. Атухов Ю.П. Генетические процессы в популяциях.
Москва «Наука» 1989.
. Айала Ф., Кайгер Дж. Современная генетика: В
3-х т. Т.3. Пер. с англ. - М.: Мир, 1988. - 335., ил.
27. Источник: А.А. Травин: «Химия
и жизнь», 2003, №1