Влияние натриево-сульфатного засоления на рост пшеницы
Введение
Засоленные почвы занимают около 30% всей суши.
По некоторым данным [Удовенко, 1978; Блэк, 1973] в бывшем СССР засолено около
10%, а в отдельных районах - до 90% всей посевной площади.
В последнее время кроме почв
первичного засоления, развивающихся естественным путем в силу природных
явлений, увеличивается и доля почв вторичного засоления, развивающихся не без
помощи человека. Масштабы вторичного засоления значительны - несколько десятков
тысяч гектаров в год. Есть серьезные опасения, что со временем вторичное
засоление может стать важным фактором, ограничивающим сельскохозяйственное
производство.
Борьба с засолением почв
приобрела народнохозяйственное значение, поэтому в последнее время вопросам
солеустойчивости растений уделяется огромное внимание [Третьяков, 2000].
Наряду с правильной агротехникой и
мелиоративными путями ослабления уровня отрицательного воздействия засоления
почвы, важное значение имеет задача повышения устойчивости самих растений к
стрессовым условиям [Жученко, 1988].
Целью работы являлось исследование влияние
натриево-сульфатного засоления почв на рост пшеницы
В задачи работы входило:
. исследование влияния натриево-сульфатного
засоления на рост корней и калеоптилей пщеницы.
. исследование влияния натриево-сульфатного
засоления на накопление биомассы пщеницы.
Глава 1. Литературный
обзор
.1 Засоление почв
За последние 50 лет уровень
Мирового океана поднялся на 10 см. Эта тенденция, по предсказаниям ученых,
будет продолжаться и дальше. Следствием этого является возрастающий дефицит
пресной воды, а доля засоленных вод увеличивается. Таким образом, к обширным
территориям в той или иной степени засоления будут прибавляться новые.
Однако кроме почв первичного
засоления, развивающихся естественным путем в силу природных явлений, увеличивается
и доля почв вторичного засоления, развивающихся не без помощи человека, чаще
всего при нерациональном орошении. Масштабы вторичного засоления значительны -
несколько десятков тысяч гектаров в год. Есть серьезные опасения, что со
временем вторичное засоление может стать важным фактором, ограничивающим
сельскохозяйственное производство.
Борьба с засолением почв
приобрела народнохозяйственное значение, поэтому в последнее время вопросам
солеустойчивости растений уделяется огромное внимание [Третьяков, 2000].
Засоление связано главным
образом с повышенным содержанием натрия в почве. В зависимости от
преимущественного накопления отдельных солей натрия засоление может быть
сульфатным, хлоридным, содовым или смешанным. Наиболее вредное влияние
оказывают ионы Na+ и С1-. Засоление приводит к созданию в почве низкого (резко
отрицательного) водного потенциала, поэтому поступление воды в растение сильно
затруднено [Якушкина, 1993].
Засоление почв может
происходить самыми различными способами. Одним из них является неумеренный,
бессистемный полив при отсутствии дренажа. Засоление почв имеет значительные
масштабы и представляет опасность для поливного земледелия.
Вторичное засоление ранее не
засоленных почв может происходить двояким путем. Во-первых, при неглубоком
залегании минерализованные грунтовые воды, поднимаясь по капиллярам почвы и
испаряясь, оставляют соли у поверхности. Во-вторых, при неумеренном поливе
происходит подъем грунтовых вод, заболачивание и засоление почвы солями,
растворенными в этих водах. Оросительные системы иногда при этом полностью
выходят из строя. Засоление может наблюдаться и при глубоком залегании
грунтовых вод, если в почвогрунте лежат уплотненные слои, служащие водоупором.
Образующаяся под этими слоями верховодка способствует капиллярному подъему и
выносу солей в пахотный слой.
Явление засоления почв имеет
широкое распространение и наблюдается во многих странах орошаемого земледелия
Азии, Африки, Америки. В результате вторичного засоления десятки миллионов
гектаров из некогда продуктивных земель превратились в малопродуктивные
засоленные почвы и соляные пустыни. Ежегодно в разных странах гибнут 200 - 300
тыс. га поливных земель от засоления и заболачивания.
Для предотвращения вторичного
засоления земель большое значение имеет дренаж, который необходим при близком
залегании минерализованных грунтовых вод. Применяют промывание засоленных почв.
Для полной ликвидации засоления рекомендуется строительство каналов в лотках и
бетонных ложах [Охрана природы, 1987].
1.2. Растения в
условиях стресса и механизмы адаптации
При любых экстремальных воздействиях в
растительном организме происходят изменения различных физиологических
параметров, что обусловлено взаимосвязанностью отдельных процессов в растении и
саморегулируемостью его метаболизма в целом.
Клетки растений реагируют сходными изменениями
на различные стрессы [Удовенко, 1978]. В основе такой неспецифической реакции
клетки лежит изменение конформационной подвижности белковых молекул.
Неспецифическая реакция растительного организма
проявляется и на других, более высоких уровнях организации биологических
систем.
Выделяют первичные изменения, обусловленные
непосредственным действием стресса на растение и вторичные, вызванные
первичными нарушениями метаболических функций [Зауралов, 1989].
К первичным относятся: изменения осморегуляции
клетки, биоэнергетических процессов, структурной целостности и состава мембран,
а также изменение структурного состояния и обусловленная им функциональная
активность ядерной ДНК [Строганов, 1962].
К вторичным относится большая часть
физиологических изменений в клетке и целом организме. Их общей чертой является
причинная зависимость от физиологических процессов, первично изменяющихся в
результате стресса. Функционально наиболее важное значение среди вторичных отклонений
имеет торможение белкового синтеза, увеличение концентрации фитогормонов,
подавляющих деление и растяжение клеток [Удовенко, 1979].
Существуют определенные закономерности в
изменении физиологических параметров под воздействием стресса, которые можно охарактеризовать
двумя типами кривых (рис. 1) [Жученко, 1987]. Обнаружено, что при стабильном и
длительном экстремальном воздействии уровень многих параметров меняется
экспоненциально и в дальнейшем сохраняется на отличном от оптимальной нормы
уровне (рис.1а). Другая же группа параметров изменяется по одновершинной
кривой, сначала существенно отклоняясь от нормы, а затем постепенно возвращаясь
к исходному уровню или близко к нему (рис.1б).

Рис. 1. Изменение физиологических параметров под
воздействием стресса: а - по логарифмическому, б - по одновершинному типу
По первому типу изменяются параметры,
характеризующие первичные нарушения и интегральные изменения. По второму типу в
экстремальных условиях меняются параметры вторичных отклонений. Они
характеризуют либо соотношение скоростных процессов, противоположно
направленных (например, синтез-гидролиз), или относительно независимых между
собой (накопление биомассы, поглощение минеральных ионов), либо состояние
постоянно обновляемых активно функционирующих биологических структур
(внутриклеточных мембран и органелл) [Жученко, 1987]. Предполагается, что
именно изменения второго типа реально отражают сущность саморегулируемости
метаболизма живой клетки и тем самым непосредственно характеризуют адаптацию
растительного организма к стрессовым условиям [Альтергот, 1982].
Важнейшими механизмами адаптации растений к
неблагоприятным факторам являются снижение скорости катаболических процессов и
более полная мобилизация потенциальных возможностей различных реакций,
компенсаторные изменения сопряженных процессов, а также включение шунтовых
механизмов [Удовенко, 1979].
Установлено, что в нормальных условиях
интенсивность различных процессов значительно ниже максимально возможного и при
неблагоприятных условиях может возрастать до такого уровня. Например, у
растений на засоленном фоне в течение некоторого времени скорость фотосинтеза
остается на том же уровне, что и на пресном [Белецкий, 1990].
Обнаружены некоторые компенсаторные изменения сопряженных
процессов. В частности, для компенсации дефицита энергетического баланса
организма у растений в условиях солевого стресса обычно усиливается
интенсивность дыхания при снижении его энергетической эффективности. У
слабоустойчивых растений, которые существенно угнетаются солями, такого
стимулирования дыхания не наблюдается или оно незначительно, и обнаружен резкий
дефицит их энергетического баланса.
Увеличение числа изоферментов в условиях
стресса, повышение активности одних ферментов при подавлении других,
катализирующих превращение тех же субстратов, активизация пентозофосфатного
пути дыхания при ослаблении гликолитического свидетельствует о включении
различных шунтовых механизмов при неблагоприятных для растения условиях
окружающей среды [Жученко, 1988].
Отмечено, что наиболее эффективная организменная
адаптация растений к длительно действующим неблагоприятным факторам
осуществляется через новообразование более устойчивых к стрессу клеток и
органов в процессе роста. То есть ростовые реакции, очевидно, являются
интегрирующими при определении адаптации растений к стрессовым условиям
[Кузнецов и др., 1991; Гамбарова, 1997].
Таким образом, адаптации растений к различным
неблагоприятным факторам окружающей среды являются неспецифическими и
определяются сложным комплексом процессов, координируемых системой
саморегуляции растений.
1.3.
Влияние солевого стресса на жизнедеятельность растений
О почвах говорят, что они засолены, если они
содержат избыток растворимых солей в корнеобитаемом слое [Блэк, 1973].
Главными катионными компонентами растворимых
солей в засоленных почвах являются натрий, кальций, магний, а главными
анионными компонентами - сульфаты, хлориды и карбонаты. Второстепенными
компонентами могут быть калий, бикарбонаты, нитраты [Строганов, 1962]. Чаще
всего засоление представлено ионами натрия в сочетании с ионами хлора, сульфата
или карбоната. Почвы с карбонатно-натриевым засолением практически непригодны
для произрастания большинства растений. Поэтому наиболее подробно изучается
влияние на растения хлорно - и сульфатно-натриевого засоления [Удовенко, 1978].
Уровень накопления иона соли в растительном
организме при данной концентрации его в среде обусловливается совокупностью
двух определяющих факторов: сорбционной емкости и растворяющей способности (или
коэффициента растворимости по отношению к воде) цитоплазмы [Блэк, 1973;
Строганов, 1962]. Ионы хлора аккумулируются в основном в надземных органах,
прежде всего в молодых листьях. Натрий также в большей степени аккумулируется в
надземных органах, особенно в стеблях. Концентрации сульфат - иона оказываются
наиболее высокими в корнях [Удовенко, 1977].
Неодинаково распределение ионов при засолении у
сортов с разной степенью солеустойчивости: более устойчивые формы локализуют
соли преимущественно в корнях, представляющих для них своеобразный барьер, а у
неустойчивых основная масса ионов накапливается в надземных органах.
При засолении претерпевают изменения
разнообразные физиологические параметры, что обусловлено взаимосвязанностью
физиологических функций в самокоординируемой системе растительного организма.
Эти изменения имеют динамический характер, отражающий последовательно идущие в
клетках фазы раздражения, повреждения и адаптации организма к засолению
[Альтергот, 1982]. Четкое представление о характере этих изменений необходимо
для разработки эффективных способов повышения солеустойчивости растений.
Физиологические процессы солеустойчивости
растений лежат в пределах осмотического давления 0,3-1,8 мПа, что соответствует
концентрации хлорида натрия 0,42-2,52% [Елисеева, 1987].
Засоление оказывает серьезное ингибиторное
влияние прежде всего на рост растений. В связи с этим выделяют ионные,
осмотические и токсичные эффекты воздействия солей на растение [Строганов,
1962; Жученко, 1988].
При засолении у растений повышается осмотический
потенциал клеточного сока, вследствие накопления в цитоплазме гидрофильных,
осмотически активных ионов солей [Белянская, Исханов, Шамина, 1991]. Наряду с
изменением уровня осмотического потенциала при засолении повышается и проницаемость
протоплазмы клеток. Установлено, что при этом выделяются кальций и белки.
Одновалентные катионы солей разрушают связь кальция со структурными и
растворимыми белками клетки, обуславливая этим изменение свойств цитоплазмы.
Добавление кальция в среду восстанавливает проницаемость протоплазмы клеток
[Калинин, 1980]. Также постоянно наблюдаемым эффектом воздействия солей на
клетки является отхождение протоплазмы от клеточных стенок. При этом происходит
разобщение плазмодесм и нарушение межклеточных связей.
Помимо осмотического действия, обуславливающего
и обезвоживание, прямое влияние солей заключается в сдвиге ионного равновесия,
которое изменяет все внутриклеточные процессы, а также неблагоприятно действует
на протоплазму, органоиды и биополимеры клетки.
В условиях засоления у большинства
культивируемых видов растений изменяется интенсивность дыхания, снижается
интенсивность фотосинтеза [Белецкий, 1990], синтеза белков и нуклеиновых кислот
[Филатова, Мельникова, 1987], фосфорного обмена [Строганов, 1962], поглощения
элементов минерального питания и их метаболического использования в надземных
органах [Хориков, 1990; Климашевский, 1991], а также доступность воды для
растения (состояние «физиологической засухи»), что в целом приводит к заметному
снижению роста [Удовенко, 1977].
Одной из причин торможения ростовых процессов у
растений при засолении является также заметное изменение содержания и
соотношения эндогенных стимуляторов и ингибиторов роста [Подлиток, 1987;
Рейтарова, Черников, 1991].
К одной из главных причин повреждения растений в
условиях засоления Б.П.Строганов [1962], относит нарушение процессов азотного
обмена. При этом в частности, замедляется синтез белков, усиливается гидролиз
запасных белков, что приводит к появлению токсичных диаминов (таких, как
путресцин и кадаверин), вызывающих появление некрозов у растений [Филатова,
Мельникова, 1987; Жученко, 1988].
Согласно Жученко [1980], при солевом стрессе
возможны повреждения мембран клеток как следствие изменения липидного
комплекса, денатурации и агрегации белков, нарушения обменных процессов,
возникновения осмотического стресса и стресса, обусловленного «голоданием»
растений. Считается, что эти нарушения носят вторичный, косвенный характер.
Эффект торможения процессов роста и развития
растений при засолении проявляется, начиная с прорастания, наклевывания семян,
роста проростка и сохраняется в течение всего периода вегетации.
Торможение синтетических процессов, в первую
очередь белка и нуклеиновых кислот, и понижение энергообеспеченности организма
ведет не только к резкому сокращению ростовых процессов, но и создает у
растений диспропорцию синтеза и гидролиза. Если уровень засоления субстрата
достигает летального порога или превышает его, то размеры нарушений постоянно
возрастают, прогрессивно увеличивается диспропорция между продуктами синтеза и
распада в клетках и организм погибает. Однако, когда степень засоления ниже,
растения выживают, но при этом интенсивность синтетических реакций в них
стабилизируется на новом, пониженном уровне.
Таким образом, приспособление к засолению (в том
числе и к многим другим экстремальным воздействиям, как считают многие
исследователи) в целом выражается в переходе всех основных реакций метаболизма
на новый, менее интенсивный уровень [Удовенко, 1975]. За счет этого, с одной
стороны, восстанавливается нормальное равновесие между интенсивностью
метаболических процессов, что обеспечивает дальнейшее существование организма,
как сложной и скоординированной биологической системы. С другой стороны,
понижается чувствительность организма к отклоняющимся от оптимальной нормы
условиям среды, что объясняет и причину появления у растений так называемой
сопряженной солеустойчивости, и сущность явления закаливания.
1.4 Солеустойчивость,
основные механизмы защиты, методы оценки
По устойчивости к засолению среди видового и
экотипического разнообразия наиболее часто выделяют две группы растений:
галофиты и гликофиты [Удовенко, 1977; Блэк, 1973]. Среди культурных растений
практически все виды являются гликофитами [Калинин, 1980].
При оценке солеустойчивости исследователи
различают два аспекта - биологический и агрономический [Удовенко, 1977;
Строганов, 1962]. С биологической точки зрения под солеустойчивостью вида или
сорта понимают тот предел засоления, при котором растения еще способны
полностью завершить онтогенетический цикл развития и воспроизвести всхожие
семена. Выше этого предела растения гибнут раньше завершения цикла онтогенеза
или образуют невсхожие семена с недоразвитым зародышем [Жученко, 1987]. При
агрономическом подходе к солеустойчивости растений обращают внимание насколько
данный сорт или вид снижает свою продуктивность в конкретных условиях засоления
(при определенном его уровне) по сравнению с его урожайностью без засоления
[Удовенко, Гончарова, 1982].
Известно, что ионы соли находятся в протоплазме
в двух различных состояниях: адсорбционно - связанном белками и другими
полимерными молекулами и свободно-растворенном в жидкой фазе цитоплазмы.
Очевидно, что при одном и том же уровне содержания иона в клетке эффект воздействия
его на метаболизм будет различным по величине при разном соотношении количеств
иона в этих состояниях. Именно этим, считает Г. В. Удовенко [1977],
определяется уровень устойчивости растений к избытку засоляющих ионов.
Большинство авторов в настоящее время выделяют
три вероятных способа защиты от солевого стресса:
. «Осмотическая защита» - поддержание тургора
клетки активацией биосинтеза осмопротекторов, балансирование внутриклеточного
осмотического давления аккумуляцией осмотически активных веществ, активация
ферментов с защитными свойствами [Белянская, Исханов, Шамина, 1991; Жученко,
1987; Блэк, 1973];
. «Вакуолярная защита» - предотвращение
токсичности действия ионов путем компартментализации в метаболически инертных
структурах [Удовенко, 1977; Дридзе, Майсурян, 1993]
. «Мембранная защита» - изменения в структуре
клеточной мембраны, препятствующие проникновению токсичных ионов в клетку или
обеспечивающие их выброс за пределы клетки с помощью «протонной помпы»
[Строганов, 1962; Климашевский, 1991].
Только сочетание всех трех вариантов защиты от
стрессовой ситуации позволяет клетке проявить максимальную резистентность к
воздействию соли [Дридзе, Майсурян, 1993].
Свойство солеустойчивости на организменном
уровне может обеспечиваться за счет накопления концентрации солей в тканях и
частях растения, не несущих большой метаболической нагрузки и выполняющих
осмотические функции без значительного воздействия на ферментативную
активность. В условиях высокого содержания хлорида натрия в субстрате некоторые
виды растений освобождаются от избытка ионов с помощью железистых клеток
[Жученко, 1987]. Растения других видов экскретируют излишнее количество солей
через специальные выделительные клетки, расположенные на верхней и нижней
поверхностях листьях, поддерживая, таким образом концентрацию ионов в тканях на
оптимальном уровне [Филатова, Мельникова, 1987].
Устойчивость к засолению разных
сельскохозяйственных культур и сортов меняется в процессе онтогенеза. При этом
на ранних его этапах, а также в период формирования репродуктивных органов,
растение обычно характеризуется наименьшей солеустойчивостью [Бессонова, 1995].
Например, ячмень, сахарная свекла и хлопчатник относятся к числу наиболее
толерантных к засолению культур, но их прорастающие семена и проростки весьма
чувствительны к засолению [Хориков, 1990].
Таким образом, солеустойчивость является
результатом совокупного действия ряда защитных механизмов, детерминированных в
клетке, скоростью адаптивных реакций, а также многовариантностью путей
реализации онтогенетического развития растительного организма.
Существует большое количество прямых и косвенных
физиологических методов оценки солеустойчивости растений, основанных на учете
различных параметров. Большая группа методов включает оценку прорастания семян
в солевых растворах [Удовенко, Гончарова, 1982]. Тестирующим признаком служит
депрессия накопления биомассы и депрессия роста проростков в солевом растворе
по сравнению с проростками пресного контроля [Иванов, Удовенко, 1970; Семушина,
Лукьянова, 1979]. Многие исследователи подчеркивают, что детальная информация о
степени солеустойчивости различных растений в первый период роста может
способствовать рентабельному возделыванию их на засоленных землях [Быстров,
Коваль, 1996; Иванов, Удовенко, 1970]. Метод проростков позволяет изучать
способы повышения биологической солеустойчивости сортов; сопоставлять
солеустойчивость семян различных видов и сортов, выделять наиболее устойчивые
из них для возделывания на полях с учетом уровня засоления; определять уровень
солестойкости данного сорта; выявлять устойчивые формы растений для целей
селекции; устанавливать предел засоления, при котором возможно прорастание
семян, точно разграничивать сорта по интенсивности роста проростков при
засолении [Иванов, Удовенко, 1970]. При этом долгое время стоял вопрос о
возможности диагностики солеустойчивости сортообразцов путем их оценки в первую
фазу роста. В результате многочисленных исследований солеустойчивости
культурных растений физиологическим и вегетационным методом, Г. В. Удовенко [1975]
обнаружена пригодность метода проростков для сравнительной характеристики
солеустойчивости культурных растений.
Вегетационный и вегетационно-полевой методы
оценки солеустойчивости растений являются интегральными и наиболее точными, но
в то же время наиболее длительными и трудоемкими. В этом случае растения
выращивают по обычной методике вегетационного или полевого опыта на двух фонах
- незасоленном [контроль] и засоленном. Солеустойчивость растений определяют по
степени снижения урожайности сорта или культуры на засоленном фоне по сравнению
с урожайностью его на контроле [Удовенко, Гончарова, 1982].
Глава 2. МЕТОДЫ И
ОБЪЕКТЫ ИССЛЕДОВАНИЯ
.1. Объекты
исследования
Объектами исследования служили проростки
пшеницы, выращенные в условиях рулонной культуры на различных концентрациях
солей.
Яровая пшеница из всех зерновых культур наиболее
требовательна к условиям произрастания. Это объясняется некоторыми ее
биологическими особенностями. Прежде всего, после всходов пшеница растет
медленно. По приросту ассимиляционной поверхности в первые дни после всходов
она значительно уступает ячменю. Это обуславливает большую чувствительность
пшеницы к неблагоприятным факторам по сравнению с другими зерновыми культурами.
Кроме того, основная масса корней располагается в пределах пахотного слоя на
глубине до 30 см, предельная глубина их проникновения редко превышает 50-60 см.
Поэтому влагу и питательные вещества пшеница может использовать только верхних
горизонтов почвы [Андреева, Родман, 1994; Яковлева, Челомбитько, 2001].
Лучшие почвы для возделывания пшеницы −
черноземы хорошо обеспеченные влагой и питательными веществами. Неплохо растет
пшеница на каштановых, особенно темно-каштановых почвах, серых лесных, имеющих
пониженное потенциальное плодородие. Мало пригодны для пшеницы
дерново-подзолистые почвы, пшеница снижает урожаи на кислых почвах с реакцией
почвенного раствора pH 5,5-4,5.
Также плохо переносит солонцеватые почвы со
щелочной реакцией почвенного раствора.
Культура в целом не требовательна к теплу,
хорошо растет и развивается при умеренных температурах. Потребность в воде
довольно высокая. На образование 1 г сухого вещества растения расходуют более
400 г воды.
Биологические особенности по периодам развития в
условиях Восточной Сибири заключаются в следующем.
Посев − всходы. Пшеница не требовательна к
теплу, особенно в первые периоды роста. Семена ее способны прорастать при
температуре +1-2˚С. энергичное прорастание пшеницы и появление всходов
проходит при температуре +8-12˚С. при влажности почвы более 40% от НВ
зерно быстро прорастает, вбирая в себя влагу, более высокая влажность ускоряет
процесс прорастания. Сумма среднесуточных положительных температур, необходимая
для появления всходов, составляет 111-159˚С. период от посева до всходов
колеблется от 11 до 18 дней в зависимости от увлажнения и температуры почвы.
При посеве во влажную, теплую почву всходы появляются на 6-8-ой день. Сортовые
особенности не влияют на продолжительность этого периода.
Для роста корневой системы оптимальная
температура ниже, чем для наземной части. Поэтому в прохладные весны всходы
получаются более крепкими за счет лучшего развития корневой системы, а при
быстром повышении температуры сразу после посева отношение массы корней к массе
наземной части ухудшается. Это приводит к ослаблению растений, их изреживанию.
Кроме того, резкое повышение температуры поверхности почвы до 35-40˚С
вызывает термический ожог всходов.
Всходы − кущение. Фаза кущения наступает
через 10-14 дней после всходов.
Сумма эффективных температур за период всходы−кущение
колеблется по различным зонам и сортам от 90 до 200˚С. количество осадков
от всходов до кущения составляет 13,4-33,4 мм.
В фазе кущения пшеница требует большого
количества влаги, в это время начинается формирование колоса и закладка
колосков. При недостатке влаги в этот период сильно уменьшается длина колоса и
число колосков. Из общего суммарного расхода влаги за всю вегетацию пшеница
использует от посева до кущения не более 5-7%, а в фазе кущение 15-20%. Период
кущения в зоне тайги и подтайги длится 20-22 дня, в лесостепи и степи -35-36
дней.
Наиболее благоприятная температура воздуха для
кущения составляет 12-15˚С.
На интенсивность кущения существенно влияет
минеральное питание. На повышенном фоне пшеница кустится лучше. Крупное зерно,
имея большой запас питательных веществ по сравнению с мелким дает большее число
побегов.
Кущение − выход в трубку −
колошение. Зачаточный стебель с узлами и зачаточным колосом развивается у
пшеницы в фазе кущения.
К моменту развития 4-6 листьев у пшеницы
начинает удлиняться нижнее междоузлие (отрезок стебля между стеблевыми узлами).
Одновременно начинает развиваться зачаточный колос, расположенный над
стеблевыми узлами. Когда нижнее междоузлие достигает длины 3-5 см, отмечается
фаза выхода в трубку.
За время выхода в трубку и колошения пшеница
потребляет 50-55% от общего расхода влаги за вегетационный период. Недостаток
элементов питания резко снижает урожай. Период 15-20 дней перед
колошением-цветением является критическим и определяет высоту урожая.
Длительность периода кущение-колошение по
различным зонам колеблется от 25 до 41 дня.
Колошение − восковая спелость. За начало
колошения принимают момент, когда из верхнего листового влагалища появляется
одна треть колоса. Через 2 дня после колошения начинается цветение с колосков
средней части колоса. В отличие от других зерновых культур цветки у пшеницы
раскрываются рано утром и закрываются вечером. В каждом колоске могут
развиваться 3-5 цветков и в благоприятные годы они формируют полноценное зерно.
В период цветения пшеница наиболее чувствительна
к пониженным и повышенным температурам воздуха. Заморозки (минус 1-2˚С)
могут повредить пыльцу и завязь пшеницы, в результате чего не произойдет
оплодотворение и зерно не сформируется. Опасны суховеи и недостаток влажности
почвы.
Колос цветет 4-12 дней. Через 1-2 дня после
цветения прекращается рост стебля в высоту, приостанавливается нарастание
вегетативной массы, идет интенсивный рост зерновки в длину и быстро
увеличивается в ней содержание воды, происходит налив зерна. В этот период
пшеница требовательна к влаге.
Когда зерно в период налива примет присущую ему
форму и величину, запасные вещества зерновки переходят в нерастворимое
состояние, наступает созревание зерна.
Различают три фазы спелости: молочную, восковую
и полную.
Молочная спелость- начало фазы созревания зерна.
Растения в это время еще сохраняют зеленую окраску, стебли желтеют только
снизу. Зерно уже сформировано, но сохраняет зеленый цвет. При надавливании из
него выступает бело-молочная жидкость, содержащая 40-60% воды.
Восковая спелость- зерно желтеет, приобретает
восковидную консистенцию, содержание влаги снижается до 35-40%. В это время уже
прекращается накопление органического вещества в зерне.
Полная спелость- зерно твердеет, влажность
снижается до 20-22%. Чувствительность зерна к заморозкам падает по мере
созревания. В фазе молочной спелости пшеница может быть повреждена при
температуре минус 2˚С, а в середине восковой спелости− при минус
2,5-4˚С, а в более поздние фазы- при минус 13-16˚С.
В период созревания пшеница более требовательна
к теплу. В Восточной Сибири температура воздуха вслед за колошением от декады к
декаде снижается и пшеница не вызревает даже при длительном отсутствии
заморозков.
Яровая пшеница − растение длинного дня,
поэтому продуктивность вегетационного периода изменяется не только в
зависимости от тепла и влаги, но и при продвижении ее посевов с севера на юг.
Продуктивность периода колошение−восковая
спелость определяется температурным режимом и условиями увлажнения и не зависит
от длины дня.
2.2 Методика
выращивания пшеницы
Для лабораторной оценки солеустойчивости
исследуемых генотипов яровой пшеницы использовали рулонный метод оценки
проростков [Каталог мировой коллекции ВИР. Ячмень..., 1991]. В качестве тестирующего
признака смотрели депрессию накопления биомассы и депрессию величины
проростков, выращенных в солевом растворе по сравнению с проростками пресного
контроля. [Иванов, Удовенко, 1970; Семушина, Лукьянова, 1979; Быстров, Коваль,
1996].
Отобранные семена предварительно замачивали
водой в чашках Петри в течение трех суток в темном термостате при t=22oC. Затем
проросшие зерна переносили в рулоны из фильтровальной бумаги. Рулоны помещали в
вегетационные сосуды с дистиллированной водой (контроль) и с раствором хлорида
(сульфата) натрия при концентрации 1,68% (опыт). Выбор концентрации обусловлен
тем, что при таком уровне содержания хлорида натрия начинает проявляться
значимое влияние засоления. Еще в 1953 г. Эйерс (цит. по Иванов, Удовенко,
1970) в опыте с 30 сортами ячменя установил, что достоверные сортовые различия
можно обнаружить в течение 10 дней и дальнейшее пребывание семян в условиях
засоления существенно не меняет их относительной солеустойчивости. Поэтому
время экспозиции проростков в рулонах мы ограничили до семи суток. По
прошествии этого времени рулоны разворачивали и проводили необходимые измерения
(линейных размеров и сырой массы побегов и корней). Помещение в рулоны уже
пророщенных зерен обусловлено тем, что в случае непосредственного проращивания
семян в растворах солей за небольшой промежуток времени (7-10 дней) было бы
невозможно получить проростки. В ходе исследований было проведено два
эксперимента. В первом измеряли длины (мм) побегов и корней каждого отдельного
проростка в рулоне, а также рассчитывали варьирование этих характеристик среди
проростков в одном таком рулоне. Повторностями служили тридцать зерен в одном
рулоне.
Для измерения сырой массы надземной и подземной
части проростков (по 30 проростков в каждой из трех повторностей) в каждом
рулоне измеряли суммарную сырую массу побегов и отдельно корней проростков в
одном рулоне, затем высчитывали среднее. Средние значения по рулону служили
повторностями. Для взвешивания использовались технические весы ВЛКТ-160.
Измерения проводились с точностью до сотых грамма.
2.3 Биометрические
методы исследования
В работе рассчитывались:
1. Средняя арифметическая (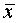 ) по
формуле:
) по
формуле:
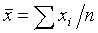
где
n - число
наблюдений.
2. Среднее квадратичное отклонение (σ),
по
формуле:
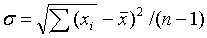
где
(n - 1) -
число степеней свободы.
3. Ошибка среднего (mx), по формуле
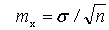
4. Существенность различия (t)
по формуле:
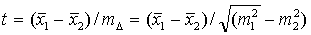
Если t
≥
3, то различие существенно, доказано и данное мероприятие вызвало существенное
изменение. Обе сравниваемые выборочные совокупности являются представителями
качественно разных генеральных совокупностей. Если t < 3, то можно
утверждать, что расхождение оказалось случайным, недостоверным и во всяком
случае целесообразность данного мероприятия осталась недоказанной а обе
совокупности являются варьирующими представителями одной и той же генеральной
совокупности (Дворецкий, 1971).
5. Коэффициэнт вариации - среднее
квадратическое отклонение, выраженное в процентах от среднего значения
признака, т.е.
При помощи коэффициента изменчивости можно
судить о степени различия в колебании разных признаков как в пределах одной
совокупности, так и разных совокупностей независимо от единиц измерения разных
сопоставляемых признаков.
На основании величины коэффициента изменчивости
можно установить характеристику изменчивости, например, по следующей схеме:
коэффициент изменчивости до 5% - изменчивость слабая; 6-10% - умеренная; 11-20%
- значительная; 21-50%- большая; 50% и больше - очень большая
Глава 3. ОХРАНА ТРУДА
В соответствии с уставом всемирной организации
здравоохранения ″здоровье - это состояние физического, духовного и
социального благополучия, а не только отсутствие болезней и физических дефектов″.
Охрана труда - система обеспечения безопасности
жизни и здоровья работников в процессе трудовой деятельности, включая правовые,
социально-экономические, организационно-технические, реабилитационные и другие
мероприятия. С охраной труда тесно связана пожарная безопасность. Охрана труда
решает задачи идентификации опасностей, профилактики, действия в условиях
чрезвычайных ситуаций [Никитин, Щербаков, 1985].
Безопасность. Работы проводились на базе
лаборатории физиологии и биохимии сельскохозяйственных растений кафедры
агроэкологии и природопользования ЭБТФ ФГОУ ВПО КрасГАУ. В технологический
процесс входили следующие операции:
обработка данных на ЭВМ,
работа с литературой,
работа с химреактивами и химпосудой.
Для разработки мероприятий по охране труда,
необходимо выявить опасные и вредные производственные факторы в
производственных процессах.
Анализ факторов ведется с учетом технологии
работ и классификаций, в соответствии с положениями ГОСТ 12.003-90.
Классификация опасностей и выявленные при этом вредные производственные факторы
технологического процесса, приведены в таблице 1.
Мероприятия по улучшению условий труда. Для
создания оптимальных условий труда предусмотрены организационные мероприятия,
мероприятия по защите органов зрения, мероприятия по электробезопасности,
мероприятия по пожарной безопасности и мероприятия по технике безопасности при
обращении с химреактивами.
Таблица 1. Опасности в технологическом процессе
|
Процесс
операция
|
Опасные
и вредные производственные факторы
|
Продолжительность
|
|
вид
|
характеристика
|
|
|
Работа
с ЭВМ
|
физические
|
повышенная
напряженность магнитного поля, повышенный уровень шума.
|
постоянные
|
|
психофизиологические
|
перенапряжение
глаз, гиподинамия, монотонность труда
|
временные
|
физические
|
недостаточная
освещенность
|
постоянные
|
|
психофизиологические
|
умственное
перенапряжение, перенапряжение глаз
|
временное
|
|
Работа
с химреактивами
|
физические
|
недостаток
вентиляции
|
временное
|
|
психофизиологические
|
умственное
перенапряжение
|
временное
|
|
Обращение
с химической посудой
|
физические
|
острые
кромки
|
временное
|
Организационные мероприятия. В соответствии с
ГОСТ 12.0.004-90 обучение рабочих безопасности труда проводят на всех
предприятиях и организациях, независимо от характера и степени опасности
производства при:
· подготовке новых рабочих (вновь принятых рабочих
не имеющих профессии или меняющих профессию);
· проведение различных видов
инструктажа ;
· повышение квалификации;
Вводный инструктаж проводит инженер по охране
труда или лицо, на которое возложены обязанности по охране труда. Он проводится
со всеми принимаемыми на работу, не зависимо от образования, стажа работы или должности,
а также студентами прибывшими на производственное обучение или практику.
Вводный инструктаж проводится в кабинете по охране труда, или специально
оборудованном помещении с использованием современных технических средств
обучения и пропаганды, а также наглядных пособий. О проведении вводного
инструктажа и проверки знаний делают запись в журнале регистраций, с
обязательной подписью инструктирующего и инструктируемого.
Первый инструктаж на рабочем месте, повторный,
внеплановый, текущий проводит непосредственно руководитель работ. Он проводится
с каждым работником индивидуально с практическим показом безопасных приемов и
методов работы.
Повторный инструктаж проводят в целях проверки и
повышения уровня знаний, правил и инструкций по охране труда индивидуально или
с группой работников, он проводится не реже чем через шесть месяцев.
Внеплановый инструктаж проводится индивидуально
или с группой работников в объеме первичного инструктажа на рабочем месте при:
изменении правил по охране труда, изменении технологического процесса,
модернизации оборудования, приспособлений и других факторов, влияющих на
безопасность труда.
Текущий инструктаж проводят с работниками перед
выполнением работ, на которые оформляется наряд- допуск.
Знания, полученные при инструктаже, проверяет
работник, проводивший инструктаж. Работник, получивший инструктаж и показавший
неудовлетворительные знания, к работе не допускается. Он обязан вновь пройти
инструктаж. О проведении первичного, повторного и вне планового инструктажа,
лицо проводившее инструктаж, делает запись в журнале регистрации на рабочем
месте с обязательной подписью инструктируемого. При регистрации внепланового
инструктажа указывают причину, вызвавшую их проведения.
Контроль за выполнением работ по охране труда
возлагается на инструкторов, которые должны постоянно осуществлять контроль за
соблюдением работниками безопасных приемов и методов труда. К лицам,
систематически нарушающим безопасные приемы труда, применяют меры воздействия,
предусмотренные правилами внутреннего трудового распорядка, правилами охраны
труда и трудовым законодательством.
Защита органов зрения. При работе с компьютером,
а также с литературой основная нагрузка ложится на зрительные органы.
Недостаточная освещенность, пониженная контрастность оказывают вредное
воздействие на глаза, что в конечном итоге приводит к ухудшению или полной
потере зрения. Исследования показывают, что 90% людей, работающих на ЭВМ,
жалуются на утомляемость, боли в области затылка, шеи, плечевого пояса, но чаще
всего страдает зрение, наблюдается покраснение век, слезоточение, жжение и боли
в области глаз. Но влияние всех этих факторов можно избежать или снизить их
воздействие. При работе за письменным столом рекомендуется освещенность стола
300-500лк, освещенность монитора не более 300лк. При работе с текстовой
информацией предпочтение отдается позитивному контрасту (темные знаки на
светлом фоне). Для уменьшения зрительного напряжения необходимо, чтобы
изображение на экране было четким и контрастным, не имело бликов и отражений
рядом стоящих предметов. Необходимо также исключить, возможность засветки
экрана, поскольку это снижает контрастность и яркость изображения. Для этого
можно использовать шторы или жалюзи. Расстояние, от монитора до глаз
работающего за ним, рекомендуется выдерживать в приделах 50-70 см. Для
предохранения глаз оператора от попадания в них наэлектризованной пыли, на
мониторе рекомендуется использовать защитный экран. Для снижения статического
напряжения за дисплеем сидеть следует так, чтобы линия взора приходилась на центр
экрана, а расстояние от глаз до рабочих поверхностей (клавиатура, дисплей,
стол) были близки по значению, иначе чрезмерное напряжение аккомодационной
мышцы может привести к развитию близорукости. Рабочий стол имеет пространство
для ног высотой не менее 600мм, шириной не менее 500мм, глубиной на уровне
колен не менее 450мм и на уровне вытянутых ног не менее 650мм (согласно СанПИН
2.2.2.542-96).
Электробезопасность. Большинство приборов,
используемых при обработке материалов дипломной работы, имеют питание от
электросети и должны соответствовать ГОСТ 12.1.019-79. При обращении с ними
необходимо соблюдать следующие правила техники безопасности:
· перед началом работы следует проверить
исправность электропроводки;
· не включать прибор не имеющий
заземления;
· не оставлять прибор без присмотра;
· не пытаться самостоятельно устранять
неисправности в электроприборах и электросети, все неисправность должны
устранять только электромонтеры;
· все нагревательные приборы (в данном
случае термостат) независимо от их мощности имеют постоянное место с
достаточной тепловой изоляцией как снизу так и со стороны стены.
В лаборатории необходимо иметь один общий
рубильник, для включения и выключения всей внутрилабораторной сети. От этого
рубильника ведут постоянную проводку к рабочим местам и распределительным
щитам. Приборы с мощностью до 800Вт подключаются в штепсели применяемые для
обычной сети, более мощные приборы необходимо присоединять к рубильникам и
распределительным щитам. Для обесточивания каждая лаборатория имеет два места
выключения тока: один на щите внутри лаборатории и второй вне лабораторного
помещения. Оба они имеют внешние отличительные признаки, доступны и известны
всем работающим в лаборатории.
Несоблюдение правил техники безопасности при
обращении с электроприборами может привести к поражению электротоком, а также к
ожогам разной степени тяжести. При термостатических ожогах пораженное место
следует смочить этиловым спиртом или раствором марганцовокислого калия или
наложить повязку с мазью от ожогов. При тяжелых ожогах помощь должна быть
оказана медицинским персоналом. При поражении электрическим током, если человек
остается в соприкосновении с токоведущими частями, необходимо немедленно
отключить ток или изолировав свои руки токонепроводящим материалом отделить
пострадавшего от токопроводящих частей к которым он прикасается. Если
пострадавший потерял сознание, нужно немедленно применить искусственное
дыхание.
Пожарная профилактика. В пожарной профилактике
наибольшее значение имеют предупредительные мероприятия, цель которых создать
такие условия, при которых возможность пожара была бы минимальной.
Каждый сотрудник лаборатории ознакомлен с
противопожарными требованиями, предъявляемыми к его работе, возможными
причинами возникновения пожара в лаборатории, способами их тушения, наличием и
размещением средств пожаротушения в рабочей комнате и коридоре, умеет
пользоваться ими в случае пожара, а также умеет обесточить комнату.
В рабочей комнате назначается лицо ответственное
за пожарную безопасность, требования которого в этом вопросе обязательны для
всех сотрудников, работающих в данной комнате.
Для того чтобы не допустить пожара, все
сотрудники лаборатории обязаны строго выполнять следующие правила пожарной
безопасности:
· не накапливать ненужных приборов и материалов на
рабочем столе, не загромождать проходы выход из комнаты и средства
пожаротушения;
· особое внимание уделять соблюдению
правила безопасного применения и хранения легковоспламеняющихся жидкостей;
· в лабораторных комнатах находятся
следующие первичные средства пожаротушения: углекислотные огнетушители (УО2 или
УО5), асбестовое или суконное одеяло, песочницы с песком;
· при проливах огнеопасных жидкостей
немедленно обесточить помещение, место пролива засыпать песком, собрать песок
совком и вынести из здания на площадку для сжигания органических остатков;
· запрещается оставлять работающие
установки и приборы без присмотра;
· курение разрешается только в
специально отведенных для этого местах;
· в случае загорания необходимо
немедленно принять меры к его ликвидации имеющимися средствами, поставить в
известность администрацию, в случае невозможности ликвидировать загорание
собственными силами - вызвать пожарную бригаду.
Техника безопасности при обращении с
химреактивами. Организация работ по технике безопасности в лаборатории
возлагается на заведующего лабораторией.
К работе с химреактивами допускаются лица не
моложе 18 лет, годные по состоянию здоровья. Во всех помещениях лаборатории
устраивается приточно-вытяжная вентиляция, обеспечивающая такой воздухообмен,
чтобы фактическая концентрация вредных веществ и пыли в воздухе не превышали
предельно допустимых концентраций. Работы, связанные с выделением вредных паров
или газов, проводятся в вытяжных шкафах.
Каждый работник снабжается спецодеждой,
полотенцем и мылом. В помещении лаборатории не допускается: хранить личную
одежду, хранить и употреблять продукты питания. В каждой лаборатории находится
аптечка первой медицинской помощи.
При обращении со стеклянной посудой, имеют место
следующие несчастные случаи: ожоги и ранения рук. Для избежания ожогов нагретые
сосуды следует брать специальными приспособлениями. При порезах рук в первую
очередь необходимо удалить из раны осколки стекла, а затем промыть рану 2%
раствором перманганата калия, смазать раствором йода и забинтовать.
пшеница солеустойчивость корень побег
Глава 4. Результаты и
обсуждение
.1 Изменение массы
проростков пшеницы по действием натриево-сульфатного засоления
Данные результатов исследования депрессии массы
проростков пшеницы представлены в таблице 2.
Таблица 2 Масса проростков пшеницы, г.
|
Контроль
|
1,68
%
Na2 SO4
|
|
М
± m
|
С*
|
М
± m
|
С
|
|
проросток
|
2,56
± 0,079
|
12,33
|
1,71
± 0,036
|
8,52
|
|
побеги
|
1,37
± 0,044
|
0,93
± 0,018
|
7,75
|
|
корни
|
1,19
± 0,068
|
22,73
|
0,78
± 0,019
|
9,89
|
* коэффициент вариации
Полученные результаты свидетельствуют о том, что
сырая масса надземной и подземной пшеницы зависит от среды выращивания и
изменяется под действием натриево-сульфатного засоления (табл.2).
При выращивании пшеницы на растворе сульфата
натрия с концентрацией 1,68 % наблюдается уменьшение массы проростков пшеницы
на 33,2 % по сравнению с контролем. При этом отмечено, что масса наземной и
подземной части изменяется неравномерно. Так, если масса побегов уменьшилась по
сравнению с контролем на 32,1% (рис.2.), то депрессия массы корней составляет
34,4%, то есть на 2,3% больше, чем у надземной части.
Такое изменение связано с негативным влиянием
натриево-хлоридного засоления. При чем отрицательное влияние более выражено при
действии на корневую систему проростков.
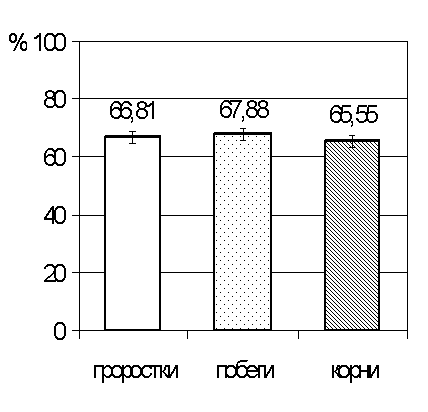

Таким образом негативное влияние
натриево-сульфатного засоления в большей степени отражается на корневой системе
проростков.
4.2 Изменение длины
корней и проростков пшеницы по действием натриево-сульфатного засоления
Данные результатов исследования изменения длины
корней и побегов пшеницы представлены в таблице 3.
Таблица 3. Изменение длины корней и проростков
|
Контроль
|
1,68
% Na
2SO4
|
|
М
± m
|
С
|
М
± m
|
С
|
|
Длина
корней
|
361,40
± 20,001
|
29,81
|
186,01
± 9,224
|
26,75
|
|
Длина
проростков
|
151,61
± 5,322
|
15,42
|
97,72
± 4,970
|
27,13
|
Полученные результаты показывают, что длина
корней и побегов зависит от среды выращивания и изменяется под действием
засоления. Изменение длины проростков под действием натриево-сульфатного
засоления видно невооруженным глазом.
При выращивании пшеницы на растворе сульфата
натрия с концентрацией 1.68 % наблюдается уменьшение длины побегов пшеницы на
35,54% и длины корней на 48,53 % (рис. 4) по сравнению с контролем. То есть
депрессия длины корней на 12,99 % выше чем депрессия побегов.
Причем отрицательное влияние наиболее выражено
при действии на длину корней.

Такое действии натриево-сульфатного засоления
связано с нарушением водно-солевого обмена проростков пшеницы. Наряду с
изменением уровня осмотического потенциала при засолении повышается и
проницаемость протоплазмы клеток, отхождение протоплазмы от клеточных стенок.
При этом происходит разобщение плазмодесм и нарушение межклеточных связей, а
следовательно транспорта веществ и воды.
Учитывая, что ионы натрия аккумулируются в
основном в надземных органах, прежде всего в стеблях, а сульфат-ионы в корнях
[Удовенко, 1977] и концентрация ионов натрия (более осмотически активны) в
растворе выше, чем концентрация сульфат-ионов, можно предположить, что большая
длина побегов по сравнению с корнями является результатом повышения концентрации
клеточного сока. Это приводит к повышению сосущей силы тканей пшеницы и как
следствие более интенсивному поступлению воды. При этом наблюдается увеличение
интенсивности роста растяжением и, как следствие, увеличение длины побегов.
Такую разницу можно объяснить заметным
изменением содержания и соотношения эндогенных стимуляторов и ингибиторов роста
на что указывается в ряде литературных источников [Подлиток, 1987; Рейтарова,
Черников, 1991].
Таким образом полученные данные не противорячат
литературным, где отмечено также, что в условиях засоления у большинства
культивируемых видов растений изменяется интенсивность дыхания, снижается
интенсивность фотосинтеза [Белецкий, 1990], синтеза белков и нуклеиновых кислот
[Филатова, Мельникова, 1987], фосфорного обмена [Строганов, 1962], поглощения
элементов минерального питания и их метаболического использования в надземных
органах [Хориков, 1990; Климашевский, 1991], а также доступность воды для
растения (состояние «физиологической засухи»), что в целом приводит к заметному
снижению роста.
Выводы
1. Под воздействием натриево-сульфатного
засоления наблюдается заметное снижение интенсивности роста проростков пшеницы,
как побегов, так и корней, по сравнению с пресным контролем.
2. Натриево-сульфатное засоление наибольшее
негативное воздействие оказывает на рост корней, что подтверждается большей
депрессией их массы и длины по сравнению с побегом.
Литература
1. Андреева
И.И., Родман Л.С. Ботаника - М.: Колос, 1994. - 527 с.
2. Дворецкий
М.Л. Пособие по вариационной статистике. - М.: Изд-во «Лесн. пром.», 1971. -
с.104
3. Жученко
А.А. Экологическая генетика культурных растений. Кишинев: Штиинца, 1980. С.
77-79.
4. Зауралов
О.А. Физиологические основы устойчивости растений. Саранск:Изд-во Саранск.
ун-та, 1989. 44 с.
. Каталог
мировой коллекции ВИР. Ячмень. Выпуск 605. Характеристика Селекционных и
местных сортов на солеустойчивость. Сост.: Г.В. Давыдова, В.С. Коваль, М.В.
Лукьянова. Л.: ВИР, 1991. 32с
. Климашевский
Э.Л. Генетический аспект минерального питания растений. М.ВО: Агропромиздат,
1991. С.416.
. Методы
оценки устойчивости растений к неблагоприятным условиям среды /Ред.
Г.В.Удовенко; Всесоюз.акад.с.-х.наук.-Л.: Колос, 1976.-318с.
8. Никитин
Л.И., Щербаков А.С. Охрана труда в лесном хозяйстве, лесной и
деревообрабатывающей промышленности. - М.: Лесн. пром-сть, 1985. - 352с.
9. Охрана природы/ А.В.Михеев,
В.М.Галушин, Н.А.Гладков и др. - М.: Просвещение, 1987. - 256с.].
10. Плохинский
И.А. Биометрия. М.: изд-во МГУ, 1970.
11. Строганов
Б.П. Физиологические аспекты солеустойчивости растений. - М., 1962
12. Удовенко
Г.В. Механизмы адаптации растений к засолению почвы: физиологические и
генетические аспекты солеустойчивости растений // Проблемы солеустойчивости
растений. - Ташкент: ФАН, 1989. С.113-142.
. Удовенко
Г.В. Механизмы адаптации растений к стрессам. // Физиология и биохимия
культурных растений, 1979 , т 11, №2, 99-106.
. Удовенко
Г.В. Солеустойчивость культурних растений.-Л.:Колос, 1977.-215 с.
. Удовенко
Г.В., Семушина Л.А. Продуктивность, накопление солей и водноосмотические
свойства растений при чистом и смешанном засолении почвы // Агрохимия. 1970.
-11. С.90-101.
. Физиология
и биохимия растений. Сборник научных трудов / Под редакцией И.П. Елисеева. Ниж.
Новгород: Нижегородск. с-х. ин-т, 1992. 88 с.
. Физиология
и биохимия сельскохозяйственных растений./ Н.Н.Третьяков, Е.И.Кошкин,
Н.М.Макрушин; Под ред. Н.Н.Третьякова - М.: Колос, 2000. - 640 с.
. Яковлева
Г.П., Челомбитько В.А. Ботаника. - Спб.: СпецЛит, Изд-во СПХФА, 2001. - 680 с.
. Якушкина
Н.И. Физиология растений. - М.: Просвещение, 1993. - 335с.