Особенности пространственно-функциональной организации лесной растительности лесостепной полосы Нижегородского Поволжья
Особенности
пространственно-функциональной организации лесной растительности лесостепной
полосы Нижегородского Поволжья
Боряков И. В., Глазунов П. А.
Дубравы в широком смысле являются наиболее
уязвимым типом растительности средней полосы России. В Поволжском регионе они -
один из ключевых компонентов лесостепных ландшафтов, но в настоящее время
подверглись глубокой деградации и находятся на грани исчезновения. Это
обусловливает актуальность мероприятий по их охране и восстановлению, что, в свою
очередь, невозможно без понимания их пространственно-функциональной
организации; изучение последней подразумевает анализ структуры как сообществ,
так и объектов инфраценотического уровня. Согласно современным представлениям,
структура упомянутых систем определяется в значительной степени экологическими
факторами (не умаляя, конечно, значения антропогенного пресса и
внутриценотических взаимодействий). При этом необходимо отметить, что изучение
именно экологических закономерностей связано с методическими трудностями
определения различных характеристик экологических режимов; эффективным и
относительно нетрудоемким решением здесь является косвенная оценка экотопа
методами фитоиндикации. С другой стороны, экологические связи отражаются на
пространственной структуре растительности и проявляются в форме ассоциации
видов в корреляционные плеяды. Поднимаясь с внутриценотического уровня на
инфраценотический, пространственные аспекты структуры приобретают форму
распределения типологических и структурных единиц мезомасштаба на более крупных
элементах ландшафта и могут изучаться средствами количественной классификации.
Опыт применения соответствующих методических подходов для анализа структуры
лесостепных дубрав Нижегородского Поволжья излагается в настоящей работе.
Материалы и методы
Материалом послужили серии геоботанических
описаний (53 пробные площади), проведенных в трех массивах широколиственного
леса, располагающихся в различных ботанико-географических подрайонах. Далее в
тексте для обозначения выборок будут использоваться названия административных
районов - Починковский, Лукояновский и Гагинский. Проводились
детально-маршрутные исследования с закладкой пробных площадей по стандартной
геоботанической методике. Для указания обилия использовалась шкала Браун-Бланке
в модифицированном варианте 1973 г., в пределах пробных площадей в большинстве
случаев закладывалось по 20 учетных площадок по 1 кв. м. Материалы были
переведены в электронный вариант с помощью программы Ecodat
(Боряков и др., 2005). Проводилась обработка методом фитоиндикации по
Элленбергу средствами этого же пакета. Статобработка проводилась в программе Statistica.
При обработке результатов возник методический
вопрос - как проводить обработку (расчет корреляций, кластерный анализ) на
материале, описанном в шкале Браун-Бланке. В качестве решения был предложен
перевод в числовую форму, использовалась следующая таблица:
Таблица. Перевод градаций шкалы Браун-Бланке в
числовую форму
|
Шкала
Браун-Бланке
|
Числовое
значение
|
|
5..3
|
5..3
|
|
2b
|
2,6
|
|
2a
|
2,3
|
|
2m
|
1,5
|
|
1
|
1
|
|
+
|
|
r
|
0,05
|
На основе первичной матрицы проводился
кластерный анализ с целью нахождения групп близких описаний. Использовалась
объединяющая стратегия, поэтому на ветвях дендрограммы помещались близкие по
составу сообщества. Вообще для кластерного анализа могут быть использованы
различные алгоритмы объединения и меры сходства (Факторный.., 1989), поэтому в
каждой работе такого рода традиционно следует обосновать выбор метода. В нашем
случае в качестве метрики было взято расстояние Эвклида - стандартная мера,
свойства которой хорошо известны (Розенберг, 1977). Она имеет недостаток, общий
для целой категории индексов, обобщаемых в виде метрики Минковского, - свойство
завышать значение сходства при скоррелированности осей, т. е. в данном случае -
при наличии положительной межвидовой корреляции. Тем не менее, из доступных
формул нет более подходящей, а идеальная в этом отношении мера расстояния
Махалонобиса сложна для расчетов. В связи со сказанным выбор формулы
представляется очевидным.
Результаты и их обсуждение
Серии описаний были обработаны методом
фитоиндикации по шкалам Элленберга и для каждого сообщества были получены
оценки четырех факторов - освещенности L, влажности F, почвенной реакции
(благоприятствование) R и богатства азотом N (из шести имеющихся в шкалах
факторов выбраны наиболее варьирующие в мезомасштабе). Визуализацию результатов
удобно производить в форме двумерных диаграмм, отражающих распределение
точек-описаний в экологическом пространстве выбранной пары факторов (этот прием
восходит к методу синэкологических диаграмм Л. Г. Раменского, 1912, цит. по:
Трасс, 1976). Такой подход, описательный по своей сути, является удобным
способом быстрой ориентации в материалах.
По имеющимся данным были построены диаграммы для
большинства сочетаний. Ниже приведены диаграммы для факторов, априори
считающихся наиболее существенными для растительности (рис. 1, 2). На них
разными значками показано положение в экологическом пространстве сообществ,
относящихся к трем рассмотренным районам исследований (на легенде “1”
обозначает серию описаний Починковского р-на, 2 - Лукояновского, 3 -
Гагинского).

Рис. 1. Распределение сообществ из трех
рассмотренных серий описаний в пространстве факторов F - влажности и N -
богатства почвы (обозначения в тексте)
Несмотря на значительное перекрывание
экологических диапазонов выборок, хорошо заметны их различия. Так, на рис. 1
видно, что серия сообществ, описанных в Лукояновском р-не, располагается в
области бóльших значений
влажности и богатства почв.
График, приведенный на рис. 2, частично
совпадает с рис. 1, так же имеется ось влажности, но по оси абсцисс
откладывается освещенность. И тут достаточно заметен уход описаний
Лукояновского р-на в область больших значений. При этом, однако, следует
напомнить, что рассматриваемые значения факторов являются оценками, полученными
по составу травостоя, и отражают преимущественно микроклимат в нижних ярусах
леса, а не климатические условия и не экотоп сообщества в целом. Величина
индицированных значений в данном случае, по-видимому, в значительной степени
определяется сомкнутостью древостоя.
На следующем этапе работы было предпринято
детальное исследование серии описаний растительности Починковского района.
Выбор обусловливается тем, что исследованный там массив представляет самый
южный вариант лесостепных дубрав района исследований, интересный как своей
флорой (присутствуют уязвимые и охраняемые виды), так и благодаря относительной
ненарушенности, а также в силу вообще плохой сохранности лесной растительности
этих вторично почти безлесных территорий. Для рассматриваемой выборки был
предпринят кластерный анализ с целью выявления групп близких по составу
сообществ, что является первым этапом количественной классификации
растительности.
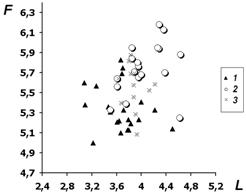
Рис. 2. Распределение сообществ в пространстве
факторов освещенности L и F - влажности (обозначения в тексте)
Прежде, чем приступить к обработке, необходимо
уточнить детали используемой техники кластерного анализа. Выбор алгоритма
объединения - метода одиночной связи, полной связи, Уорда и др. (Боровиков,
2001), - вопрос более сложный, чем выбор метрики сходства, обоснованный выше
(строгого критерия здесь пока еще не существует - Факторный.., 1989). Для
объективности нами были применены различные методы и на основе сравнения
полученных результатов был выбран способ, показавший себя как более корректный
в плане фитоценотической интерпретации.
На дендрограмме, построенной по методу одиночной
связи (не приводится за отсутствием места), имеются по меньшей мере два более
или менее крупных обособленных кластера. Рассмотрение схемы, построенной с
использованием взвешенного центроидного метода, позволяет заметить те же
группы, несмотря на иной общий вид дендрограммы. И, наконец, можно отметить,
что схема парно-группового объединения по составу принципиально не отличается
от предыдущих, но, при этом, является самой четкой, в связи с чем дальнейшее
рассмотрение велось по ней. Вообще опыт данного рода работ (Боряков и др.,
1999) приводит к выводу о принципиальном сходстве результатов кластерного
анализа, полученных с использованием различных алгоритмов объединения, исключая
метод Уорда, по-видимому, слишком сильно изменяющего структуру данных.
В данной также работе было проведено построение
трехмерных диаграмм, отражающих изменение обилия вида в экологическом
пространстве, что оказалось наглядным и информативным способом выявления
внутриценотических закономерностей. На рис. 3 показаны поверхности, описывающие
изменение обилия некоторых ключевых видов травяно-кустарничкового яруса
(пролесника многолетнего Mercurialis perennis и сныти Aegopodium podagraria) в
зависимости от сочетания значений двух факторов, в значительной степени
определяющих богатство эдафотопа: количества доступного азота и реакции почвы
(степени благоприятствования pH).

Рис. 3. Изменение обилия доминантов
травяно-кустарничкового яруса - пролесника (Mercurialis perennis, А) и сныти
(Aegopodium podagraria, Б) в зависимости от сочетания значений факторов
богатства почвы азотом (левая ось) и реакции почвы (правая ось). На
вертикальной оси - обилие в условных единицах.
Заметно общее сходство поверхностей
распределения, с максимумом в области почв, более богатых азотом, и в то же
время, относительно более кислых. Интересно то, что поверхности распределения
других видов часто демонстрируют явление вытеснения из указанной области
экологической ниши (рис. 4).

Следует отметить, что показанные на рис. 4 виды
выбраны не случайно. Carex pilosa - один из ключевых видов, нередко
доминирующий в дубо-липняках и формирующий соответственно сообщества
волосистоосоковых и волосистоосоково-снытевых ассоциаций. Pulmonaria obscura,
как правило, выступает в качестве содоминанта указанных видов; есть данные,
свидетельствующие о центральном положении этого вида в неморальном комплексе
лесостепных дубрав (Воротников и др., 2003).
Описанный подход в настоящем варианте имеет
методологический недостаток, связанный с наличием скрытой зависимости
индицируемых значений от обилия вида, распределение которого изучается.
Решением этого вопроса является исключение рассматриваемого вида при расчете
значений оценок факторов, однако в данной работе по техническим причинам эта
поправка к вычислениям не вносилась. Тем не менее, предположительная величина
искажений вряд ли должна быть большой; более детально этот вопрос планируется
рассмотреть позднее.
Для выявления состава корреляционных плеяд
исследуемой растительности был проведен кластерный анализ (R-техника,
т. е. группировка видов в отличие от кластеризации описаний, обозначаемой как Q-анализ)
на основе канонического коэффициента корреляции Пирсона.
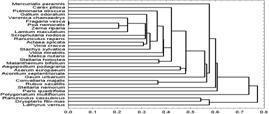
Рис. 5. Корреляционная структура изучаемой
растительности
На схеме (рис. 5) видно, что наиболее выраженную
группу образуют лесоопушечные виды, что объясняется в целом скорее лесостепным,
чем неморальным характером растительности.
Заключение
экотоп растительность кластерный
анализ
Косвенная характеристика экотопа по шкалам Г.
Элленберга применительно к лесной растительности является методом с достаточной
разрешающей способностью, позволяющий выявлять группы сообществ сходной
экологии и дифференциировать геоботанические объекты в рамках выборок
достаточно широкой экологической амплитуды.
Анализ структуры растительности способом
фитоиндикации дает возможность выявлять экологические закономерности как на
уровне инфраценотическом, (оперируя фитохорами мезомасштаба, описываемых
стандартными пробными площадями), так и внутриценотическом. В последнем случае
эффективным приемом визуализации синэкологических закономерностей является
построение экоклинов в форме поверхностей, отражающих изменение обилия вида в
зависимости от значений ведущих факторов (определяемых фитоиндикационным
методом) экотопа. Таким образом удается проследить расхождение видов по
экологическим нишам и эффект вытеснения второстепенных компонентов ценоза доминантами.
Литература
Боровиков
В. П. Программа Statistica для студентов и инженеров. М.: КомпьютерПресс, 2001.
301 с.
Боряков
И. В., Воротников В. П., Борякова Е. Е. Использование информационных технологий
для организации фитоценариев и обработки геоботанических данных // Бот. журн.
2005. Т. 90. № 1. С. 95-104.
Боряков
И. В., Воротников В. П., Казупица Н. А., Лютова О. В. Количественная
классификация и анализ структуры производных типов леса Керженского
заповедника. Н.Новгород, 1999. 25 с. Деп. в ВИНИТИ 20.09.99, № 2874-В99.
Воротников
В. П., Боряков И. В., Борякова Е. Е., Краснобаева О. С. О корреляционной
структуре сообществ и инфраценотической организации широколиственных лесов юга
Нижегородской области (на примере дубрав Починковского района) // Лесоэкологические
проблемы Поволжья. Н. Новгород, 2003. С. 71-79.
Розенберг
Г. С. Обзор методов статистической геоботаники. 2. Индексы сходства. Деп. в
ВИНИТИ N 20-77. М., 1977. 30 с.
Трасс
Х. Х. Геоботаника. История и современные тенденции развития. Л.: Наука, 1976.
252 с.
Факторный,
дискриминантный и кластерный анализ. М.: Финансы и статистика, 1989. 215 с.