Анализ наследования количественных признаков у гибридов риса F2 от скрещивания Айсберг*Вираж
МИНИСТЕРСТВО
СЕЛЬСКОГО ХОЗЯЙСТВА РОССИЙСКОЙ ФЕДЕРАЦИИ
ДЕПАРЛАМЕНТ
НАУЧНО-ТЕХНОЛОГИЧЕСКОЙ ПОЛИТИКИ И ИОБРАЗОВАНИЯ
ФЕДЕРАЛЬНОЕ
ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ
ВЫСШЕГО
ПРОФЕССИОНАЛЬНОГО ОБРАЗОВАНИЯ
«АЗОВО-ЧЕРНОМОРСКАЯ
ГОСУДАРСТВЕННАЯ АГРОИНЖЕНЕРНАЯ АКАДЕМИЯ»
Кафедра:
Селекции и Генетики с.-х. культур
Курсовая
работа
ПО
ДИСЦИПЛИНЕ: «Генетика популяций и количественных признаков»
НА
ТЕМУ: Анализ наследования количественных признаков у гибридов риса F2
от скрещивания Айсберг*Вираж
Выполнил: Кузнецов М.В.
Проверил: доктор с.-х. н., профессор
Костылева Л.М.
Зерноград
2013
СОДЕРЖАНИЕ
Введение
1. Литературный обзор
2. Материал и методика
исследований
. Результаты исследований
Выводы
Список литературы
Приложение
ВВЕДЕНИЕ
гибрид
рис скрещивание
Генетический анализ является одним из основных
профилирующих курсов при подготовке специалистов-генетиков независимо от их
специализации.
Понимание принципов генетического анализа
основано на знании закономерностей наследования признаков и свойство
наследственности, с одной стороны, и биологии объекта - с другой.
Освоение логики анализа необходимо для изучения
частной генетики видов, селекции сельскохозяйственных культур растений, пород
животных, штаммов промышленных микроорганизмов. Только в ходе анализа
наследования многих и разнообразных признаков организмов одного вида
складывается представление о генотипе как о системе, а не как о сумме генов.
Понимание основополагающего положения генетики, гласящего о том, что каждый
признак определяется многими генами, взаимодействующими между собой, а каждый
ген - плейотропен в своем проявлении, проходит так же только в ходе
теоретического и практического знакомства с генетическим анализом. Логика
генетического анализа необходима и для такого плодотворного этапа в работе
генетика, как генетический синтез, т.е., создание организмов с определенными
генотипами. Синтез широко применяется сейчас в биотехнологии и генетической
инженерии. Попытки использования целенаправленного синтеза генотипов делаются
уже и в селекционной сельскохозяйственной практике.
Создание особей с новыми генотипами, благодаря
использованию современных методов генетической инженерии, накладывает очень
большую ответственность на генетиков, которые должны уметь делать прогнозы,
т.е. предвидеть как судьбу синтезированных человеком генотипов, так и их роль в
популяциях своего вида и биогеоценоза в целом. А это требует развития
фундаментальных генетических исследований, опережающих развитие прикладной и
практической генетики. При этом первостепенное значение имеет совершенствование
методов генетического анализа, целенаправленное повышение его разрешающей
способности. Не меньшую роль в развитии генетики должно сыграть и овладение
принципами, методами и логикой генетического анализа.
Генетический анализ как самостоятельны курс
начал складываться в последние десятилетия, но это не значит, что у него не
было предшественников. Так, большой практику по генетике в Ленинградском
университете появился на только что созданной Ю. А. Филипченко кафедре генетики
в 1919 г. Практикум знакомил с методами генетических исследований и, прежде
всего с методами скрещиваний и анализа полученных результатов. Начиная с 1923
г. Выходит в свет ряд книг Ю. А. Филипченко - «Изменчивость и методы ее
изучения» (1923), «Частная генетика растений» (1927), «Частная генетика
животных» (1928), курс лекций «Генетика» (1929), а в 1934 г. - его же
монография по генетическому анализу наследования количественных и
морфологических признаков пшеницы, «Генетика мягкой пшеницы». Достижения цитологических
методов были обобщены в монографии И. И. Соколова «Половые клетки» (1926).
Традиции, заложенные основателями кафедры,
сохраняются до сих пор, Однако это не значит, что содержание курса и методы
работы остаются старыми.
В современном виде курс начал складываться в
50-е годы. Его становления много внимания уделял М. Е. Лобашев.
Основоположником курса был В. С. Федоров, который затем передал его в руки
своих учеников. Значительную лепту в разработку курса внесла К. В. Ватти. В
современной работе с сотрудником агрофизического института О. С. Журавлевым она
в 1978 г. Создала машинную задачу для генанализа.
Следует так же сказать, что гибридологическому
анализу очень часто сопутствуют исследования с помощью сложных молекулярных
методов.
Задачи генетического анализа можно коротко
сформулировать как определение системы генотипа организма или генотипической
структуры популяции. Методы генанализа очень разнообразны. В генетике анализе
используются скрещивания в последовательном ряду поколений: F1,
F2, F3
и т.д.
Цели курсовой работы:
1. Закрепление теоретических знаний,
полученных в курсе изучения дисциплины «Генетика популяций и количественных
признаков».
2. Формирование навыков по анализу
наследования и количественных признаков сельскохозяйственных культур в
расщепляющихся гибридных популяциях, что необходимо в селекционной работе.
Задачи:
1. Овладеть методами подсчетов и промеров
признаков сельскохозяйственных культур.
2. Представить результаты проведенного
анализа первичных данных в виде таблиц, графиков и текста, в котором
описываются полученные закономерности.
. Провести осмысление информации,
сравнить ее с литературными данными, сделать выводы.
. Определить число генов, отвечающих за
конкретные признаки, и характер их наследования.
. Провести биометрический анализ
признаков у растений (высота, длина метелки, число колосков.
. Провести генетический анализ с помощью
компьютерных программ и сделать окончательный вывод по исследованию признаков
данного гибрида.
I.
ЛИТЕРАТУРНЫЙ ОБЗОР
Рис - древнейшая и широко распространенная
продовольственная культура. В Китае, Индии, Японии, Вьетнаме и ряде других
стран он является основным продуктом питания. Рисовая крупа отличается
сбалансированным аминокислотным составом, имеет отличные вкусовые качества,
отличается высокой переваримостью и усвояемостью. Поэтому она широко
используется, в общем, и диетическом питании, а рисовый отвар обладает
целебными свойствами. Из зерна риса вырабатывают крахмал, который представляет
большую ценность для текстильной, парфюмерной и медицинской промышленности. Из
зародышей получают рисовое масло, применяемое для изготовления мыла и
стеариновых свеч. Рисовая солома используется для кормления скота, а также
производства высших сортов бумаги, картона, шляп, веревок, мешков, корзин и
других изделий домашнего обихода.
Районы возделывания и урожайность. Родина риса -
тропический и субтропический пояс Юго-Восточной Азии. В Индии, Китае, Японии
культура риса известна более 4000 лет. В Европе он возделывается с XV в. Его
выращивают более 60 стран Азии, Европы, Америки, Австралии и Африки на площади
более 140 млн. га.
В СССР древнейшими районами возделывания риса
являются Средняя Азия и Закавказье. За годы Советской власти посевные площади
под рисом и его урожайность возросли. Были созданы новые районы рисосеяния - на
Дальнем Востоке, Кубани, в Крыму, низовьях Волги, Дона, Днепра, Дуная. Основные
посевные площади сосредоточены в Краснодарском крае. Казахской и Узбекской ССР.
Среди зерновых культур это одна из самых урожайных культур. Передовые хозяйства
получают по 69-80 ц/га риса и даже больше. Наивысшая урожайность риса в
Советском Союзе была достигнута в колхозе «Кзыл-Ту» Кзыл-Ординской области
Казахской ССР дважды Героем Социалистического Труда И. Жахаевым - 171 ц/га.
Основоположником гибридологического анализа по
праву считают Г. Менделя. Это не значит, что он впервые описал такие явления,
как единообразие гибридов F1,
или расщепление в их потомстве. Предшественники его И. Г. Кельрейтер, Т. Найт,
О. Сажрэ, Дж. Госс, Ш. Нодэн, Ч. Дарвин тоже описывали эти явления, но им не
удалось понять их значимость, так как эти явления не воспроизводились при
повторных экспериментах. Именно Г. Мендель в 1865 г. заложил основы
генетического анализа, сформулировав необходимую систему гибридологического
анализа, состоящую:
) в проверке признака в течение 2-3 поколений на
константность (гомозиготность исходного материала);
) в изучении отдельных альтернативных признаков;
) в количественном учете расщепления;
) в индивидуальном (посемейном) анализе всего
потомства гибридов (а не части его, как делали раньше). Следуя перечисленным
правилам, можно получать хорошо воспроизводимые результаты, а значит изучать
закономерности наследования признаков, вести генанализ. И действительно, в 1900
г. после переоткрытия законов Менделя Г. Де Фризом, К. Корренсом, Э. Чермаком
началось бурное развитие генетики.
По посевным площадям и валовым сборам зерна рис
занимает в мировом земледелии второе (после пшеницы) место. Основные площади
его культуры сосредоточены в Юго-восточной Азии, Индия, Пакистан, Бангладеш и
Китай. В дореволюционной России основными районами производства риса были
Средняя Азия (90% сбора) и Закавказье (10%).В России площадь под посевами риса
увеличилась с 350 (1960) до 377.3 тыс. га (1980), а урожайность соответственно
выросла с 2.02 до 3.96 т/га. Увеличение площадей связано с освоением
малопродуктивных плавневых и засоленных почв под культуру риса. Ростовская
область
один из самых северных районов выращивания риса.
В настоящее время в Ростовской области имеется 48 тыс. га инженерных рисовых
систем. Стебель злаковых растений также называется соломиной. На растениях риса
стебель - важный орган, тесно связанный с устойчивостью к полеганию и, в
конечном счете, влияющий на урожайность зерна. Сорта риса, выведенные в
последние три десятилетия, имеют короткие и прочные стебли, что способствует
высокой урожайности, устойчивости к болезням, хорошему качеству.
В Японии с 1908 до 1973 гг. средняя длина стебля
риса для пяти самых распространенных сортов стала короче от 100-120 см до 73-88
см (в среднем от 109 до 81 см). Таким образом, стебель всегда принимался во
внимание как важный признак в современной селекции риса. Поэтому генетические
исследования признаков, связанных со стеблем, были активно проведены многими
учеными. Многие из них связаны с длиной, а также толщиной, упругостью стеблей и
числом стеблей и метелок на растении.
«Высота растения» риса - наиболее важный в
селекционной практике морфологический признак, так как он тесно связан с
устойчивостью к полеганию, а, следовательно, способностью выдерживать высокие
нормы удобрений.
Ведущее положение среди рисопроизводящих стран
занимают Китай и Индия. Они обеспечивают 56,2 % мирового производства риса. В
этих странах находится 50,9% площади, занятой рисом в мире. Рис используется в
пищу практически во всех странах. Для народов Азии, Африки и Латинской Америки
он является важнейшим продуктом питания. Потребление ими риса составляет 100 кг
и более в год на душу населения.
Главный продукт изготовляемый из зерна риса,
крупа, обладает высокой питательностью, хорошими вкусовыми качествами,
лечебными, диетическими свойствами (рисовый отвар), легкой переваримостью и
усвояемостью. В зерне риса содержится: воды 12.2%, азотистых веществ 6.4%, жира
2.1%, безазотистых экстрактивных веществ 69.3%, клетчатки 6.5%, золы 3.5%.
Высокая биологическая ценность белка риса связана с повышенным содержанием в
нем незаменимых для человека аминокислот с малым содержанием танина в зерне. В
сравнительно небольших количествах из зерна риса получают: крахмал, рисовую
пудру, спирт, пиво.
Рисовая солома служит ценным кормом для скота,
из рисовой соломы вырабатывают высшие сорта бумаги, строительный картон,
канаты, изготавливают шляпы, обувь, сумки и т.д. Рисовые отруби (килак)
являются ценным кормом для молодняка. В них содержится от 10 до 13% белка и до
14.5% жира. Из них экстрагируют высококачественное масло, отличающееся большим
содержанием ненасыщенных жирных кислот и обладающее ценными лечебными
свойствами.
В современном учении о наследственности рядом с
чрезвычайно хорошо разработанными отделами существует вопрос, который,
безусловно, оставляет желать еще многого: это вопрос о наследовании
количественных признаков.
При первых шагах менделизма все внимание
исследователей направлялось главным образом на передачу по наследству
особенностей качественного характера, как более доступных для изучения.
Относительно количественных признаков одно время существовало даже убеждение,
что они передаются не по типу менделистической, а промежуточной
наследственности, пока открытие однозначных факторов не заставило отказаться от
этого взгляда. В ряде последующих работ, посвященных вопросу о наследовании
количественных признаков, дело не шло обычно дальше доказательства того, что
при этом играют роль подобные однозначные факторы, сама природа которых, а
нередко даже и число оставались невыясненными.
В настоящее время, несмотря на огромнейшие
достижения генетики в познании структуры гена и генетических процессов,
протекающих на молекулярно-генетическом уровне, не уменьшился разрыв между
генетикой и селекцией, о котором вел речь еще Н.И.Вавилов в 30-е годы. Теория
селекции базируется на знаниях генетических процессов организменного
популяционного уровней, а они не удовлетворяют сильно возросшие запросы
селекционеров. Объясняется это в основном отсутствием представлений о
генетической природе количественных признаков и теорий, объясняющих явления
связанные с функционированием этих признаков.
В последние годы состояние рисоводства резко
ухудшилось в результате многократного подорожания гербицидов, удобрении, семян,
электроэнергии, горюче-смазочных мат е риал ой услуг авиации и г.д. Для
повышения урожайности культуры огромное значение имеет создание новых
продуктивных сортов. Такая работа и современных условиях невозможна без
селекционно-генетических исследований.
В селекционной работе с растениями для более
быстрого достижения цели. К количественным признакам относятся урожайность
зерна и листостебельной массы, которые в свою очередь определяются
количественными признаками: высота растений, размер метелок или колосьев, масса
семян, кустистость и т.п. Эти признаки контролируются 3-5 и более генами.
Изучение наследования количественных признаков
на фоне разнообразных экологических условий представляет собой важную
фундаментальную и прикладную проблему. В тех случаях, когда удалось
расшифровать генетическую составляющую во взаимодействии растений со средой,
решение селекционных задач многократно упростилось.
Выяснение генетических основ количественных
признаков у риса составляющих основные элементы структуры урожая, - одна из
наиболее важных задач, решение которой необходимо для ускорения селекционного
процесса.
Гены, определяющие развитие количественных
признаков, слабо изучены. Трудность выявления последних заключается в том, что
эти признаки контролируются большим количеством генов, каждый из которых в
отдельности имеет сравнительно небольшой эффект. Кроме того, проявление
большинства количественных признаков во многом зависит от действия внешних
факторов, в частности площади питания одного растения.
Для практической селекции большое значение имеет
необходимость знания поведения гибридов и закономерности наследования
хозяйственно-ценных признаков у растений риса и их изменчивость. Наследование
есть закономерность передачи родительских признаков и свойств потомству, а
наследуемость является материалом генотипической изменчивости популяции.
Функции мирового хранилища и обменного центра,
наряду с собственными изысканиями, несет с 1961 г. Международный
научно-исследовательский институт риса - IRRI
(Филиппины). Международный центр зародышевой плазмы при институте имеет
дубликаты большинства национальных коллекций, аборигенные образцы азиатских
стран, дикорастущие виды риса. Из зарегистрированных во всем мире 120 тысяч
сортов половина представлена в IRRI.
В странах Азии, Африки и Америки созданы
национальные и региональные коллекции. К числу наиболее крупных принадлежит
коллекция зародышевой плазмы при Институте тропических культур (IITA)
в Нигерии (5 тыс. образцов африканского риса), коллекция Национальной
лаборатории консервации семян в Форт-Коллинсе, штат Колорадо (13 тыс.
образцов).
В нашей стране многообразие форм риса
сосредоточено в коллекциях Всесоюзного научно-исследовательского института
растениеводства им. Н.И.Вавилова и Всероссийского научно-исследовательского
института риса. Большую часть коллекции составляют образцы O.sativa
из СНГ и азиатских стран.
Знание природы генов, определяющих развитие
количественных признаков, их количества и локализации в хромосомах позволит
более целенаправленно и эффективно проводить отбор нужных линий, являющихся предшественниками
новых сортов. Поэтому изучение генетики количественных признаков риса является
весьма актуальным и имеет очень большое теоретическое и практическое значение.
. Материал и методика исследований В качестве
исходного материала были использованы сорта Айсберг и Вираж. Гибридизацию
проводили на привезенных с поля растениях. В качестве материнской формы
использовали сорт Айсберг, а отцовской - Вираж. Выращивания гибридов F2
и родительские формы проводили на ряду расположенных делянок площадью 20 м2 с
одинаковой густотой стояния растений. Для учета брали центральный стебель с
метелкой и остатками корней. Измеряли длину растения, метёлки, подсчитывали
общее количество колосков. Определяли частоты различных значений признаков и
группировали их по классам, на основе которых строили графики распределения.
II. МАТЕРИАЛЫ
И МЕТОДИКА ИССЛЕДОВАНИЙ
В качестве материала для исследования были
использованы растения гибридов риса F2
Айсберг*Вираж и их родительские формы Айсберг и Вираж.
Айсберг.
Родословная: инд. Отбор из сорта ВНИИР 8847.
Включен в Госреестр по Северо-Кавказскому (6) региону. Рекомендован для
возделывания по Западно-дельтовой зоне Краснодарского края. Разновидность
италика. Лист без антоциановой окраски. Опушение пластинки предпоследнего листи
отсутствует или очень слабое, антоциановая окраска ушек отсутствует. Метелка
желтоватая, длинная, изогнутость главной оси отсутствует или очень слабая. Ости
отсутствуют или очень короткие. Антоциановая окраска верхушки нижней цветковой
чешуи отсутствует или очень слабая. Стебель длинный. Обрушенное зерно белое,
полуветереновидное - веретеновидное, средней ширины. За годы испытаний в
регионе средняя урожайность составила 51.7 ц/га, на уровне среднего стандарта.
По Западно-Дельтовой зоне Краснодарского края при урожайности 49.8 ц/га
превысил стандарт Лиман на 7.6 ц/га. Максимальная урожайность 84.4 ц/га
получена в 2003 г. В Краснодарском крае. Высота растений 105-130 см, в среднем
на 20 см выше стандарта. Устойчив в полеганию и осыпанию. Масса 1000 зерен 28-33
г. Технологические и кулинарные качества высокие. Характеризуется высокими
стекловидностью и выходом целого ядра. Зерно удлиненное (2.8). Включен в список
ценных по качеству сортов. Морфологические особенности сорта: высокорослые
широколистные растения с эректоидной метелкой.
Вираж.
Родословная: Буденовский * Приманычский. Включен
в Госреестр по Северо-Кавказскому (6) региону. Рекомендован для возделывания в
Ростовской области. Разновидность нигроапикулята. Лист без антоциановой
окраски. Опушение пластинки предпоследнего листа слабое, антоциановая окраска
ушек отсутствует. Метелка соломенно-желтая, короткая, плотная, изогнутость
главной оси отсутствует или очень слабая. Ости отсутствуют. Антоциановая
окраска верхушки нижней цветковой чешуи сильная. Обрушенное зерно белое,
полуверетеновидное, среденей длины, узкое. За годы среднего стандарта.. В
Ростовской области на Пролетавском ГСУ по предшественнику многолетние травы в
2000 году при урожайности 61,7 ц/га превысил стандарт на 7.5 ц/га. Максимальная
урожайность 74.0 ц/га получена в 1999 году в Ростовской области. Раннеспелый,
созревает одновременно с сортом Контакт. Высота растений 85-105 см, в среднем
на 25 см короче, чем у сорта Кубань 3. Устойчив к полегания и осыпанию.
Были проанализированы признаки:
Высота растений
Длина метелки
Общее число колосков
Биометрический анализ проведен в лабораторных
условиях с помощью линейки. Генетический анализ проведен с помощью компьютерной
программы Мережко А. Ф., мо методике А. Ф. Мережко и программы Exel,
Полиген экспесс и Ген-3. Математическая обработка данных производилась по
методике Б. А. Доспехова.
III. РЕЗУЛЬТАТ
ИССЛЕДОВАНИЯ
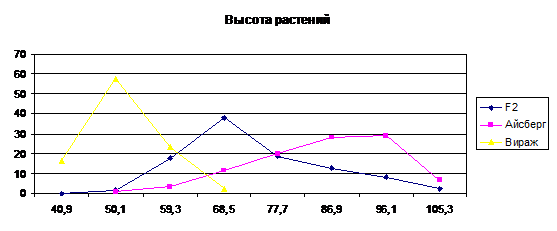
Рис. 1. Кривая распределения частот родительских
форм и гибрида по признаку высота растений
Высота растений принимается во внимание как
важный морфологический признак в современной селекции риса, так как он тесно
связан с устойчивостью к полеганию, а, следовательно, способностью выдерживать
высокие нормы удобрений.
Родительские формы сильно отличаются по средним
значениям (60,1 и 94,3). Среднее значение гибрида составляет 83,2 см. Кривая
распределения имеет правостороннюю ассиметрию (As=0.28).
Сопоставление краевых частот гибрида и рецессивного сорта Вираж показало их
совпадение при уменьшении частот родителей в 64 раза, это свидетельствует о
том, что родительские формы различаются по аллельному состоянию по 3 парам
генов. По скольку различия между родительскими формами составило 34,7 см, то
средняя сила одного гена составляет 11,4 см. Один из трех генов является
доминантным, а остальные наследуются промежуточно.
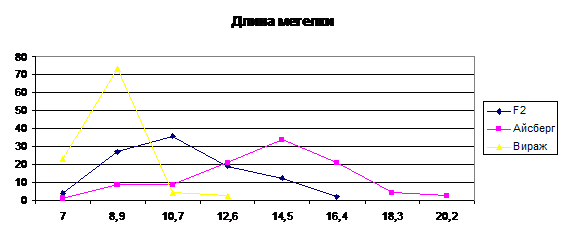
Рис. 2. Кривая распределения частот родительских
форм и гибрида по признаку длина метелки.
Длина метелки, по мнению ряда исследователей,
имеет самую высокую наследуемость и корреляцию с родительскими значениями среди
других признаков метелки (Matsuo
и др., 1960, Yamamato,
Toriyama, 1971).
По длине метелки родительские формы значительно
отличаются друг от друга. Кривая распределения в промежутке между размерами
родительских форм имеет правостороннюю ассиметрию (As=0.42).
Поскольку на долю родительской формы Вираж приходится очень мало частот
гибридов, а в первых двух классах такие формы не встречаются (10-14 см.) Эти
различия обуславливаются не менее чем по 4-м парам генов, в этом случае средняя
сила генов составила 2,4 см.

Рис. 3. Кривая распределения частот родительских
форм и гибрида по признаку число колосков
Число колосков на метелке подсчитывают в
лабораторных условиях. Оно имеет высокую корреляцию с продуктивностью растений.
Родительские сорта значительно отличаются друг
от друга (71,2 и 114,2). У гибрида наблюдается гетерозис (в среднем 161
колосок). Наблюдается положительная трансгрессия. Наличие трансгрессии
свидетельствует о том что родительские сорта отличаются по аллельному состоянию
генов (имеются и доминантные и рецессивные аллели разных локусов.) Минимальное
различие между родителями составляет три пары генов, при этом генная формула
Виража AAbbcc, а Айсберга
aaBBCC. В результате
рекомбинаций выщепляются формы, имеющих все три гена доминантном состоянии,
которые и составляют трансгрессионный ряд. По условии равной силы генов вклад
каждого из них составляет 53 зерна на одну метелку.
ВЫВОДЫ
После генетического анализа данных по
количественным признакам можно сделать следующие выводы:
По признаку высота растений родительские формы
различаются по 3 парам генов. Сила гена 3,3 см.
По признаку длина метелки родительские формы
различаются по 2 парам генов. Сила гена 2,5 см
По признаку общее количество колосков
родительские формы различаются по 1 паре генов. Сила гена 24 шт.
СПИСОК ЛИТЕРАТУРЫ
1. Брюбейкер
Дж. Л. Сельскохозяйственная генетика: Пер. с англ. Б. Н. Сидорова / Дж. Л.
Брюбейкер. - М.: Наука, 1966. - 223 с.
2. Генетические
методы в селекции растений / В. К. Шумный, А. Н. Сидоров, В. И. Коваленко и
др.; Под ред. Н. В. Турбина. - М.: Колос, 1974. - 208 с.
. Гуляев
Г. В. Селекция и семеноводство полевых культур с основами генетики / Г. В.
Гуляев, А. П. Дубнин. - М.: Колос, 1980 - 375 с.
. Доспехов
Б. А. Методика полевого опыта / Б. А. Доспехов. - 5-е изд., перераб. и. доп. -
М.: Агропромиздат, 1985. - 351 с.
. Иванова
Д. И., Костылев П. И. Генетика риса / Генетика культурных растений: кукуруза,
рис, просо, овес. - Ленинград, 1988. - с. 124-147.
. Костылев,
А. А. Парфенюк, В. И. Степовой. - Ростов на Дону, ЗАО «Книга», 2004. - 576 с.
. Мережко
А. Ф. Система генетического изучения исходного материала для селекции растений
/ А. Ф. Мережко. - Л.: ВИР, 1984. - 70 с.
. Рис.
Под ред П. С. Ерыгина и Н. Б. Натальина. М. «Колос», 1968. - 180 с.
. Федин
М. А. Статистические методы генетического анализа / М. А. Федин, Д. Я. Силис,
А. В. Смиряев. - М.: Колос, 1980. - 207 с.
ПРИЛОЖЕНИЕ
Таб. 1. Высота растений
|
Статистические
параметры родительских форм и гибрида F2
|
|
Параметры
|
Pmin
|
Pmax
|
F2
|
|
Численность
выборки
|
-
п
|
120
|
120
|
407
|
|
Среднее
значение выборки
|
-X
|
60.1
|
94,3
|
83,2
|
|
Среднее
квадратическое отклонение
|
-Sx
|
5,68
|
11,04
|
11,71
|
|
Ошибка
выборочной средней
|
-Souj.
|
0,52
|
1,01
|
0,58
|
|
Коэффициент
вариации
|
-Cv%
|
11.7
|
14,1
|
|
Максимальная
варианта
|
-
Max
|
77,0
|
115,0
|
115,0
|
|
Минимальная
варианта
|
-
Min
|
46,0
|
62,0
|
52,0
|
|
Нижний
доверительный интервал
|
-
X-3Sx
|
43,1
|
61,2
|
48,1
|
|
Верхний
доверительный интервал
|
-
X+3Sx
|
77,2
|
127,5
|
118,4
|
|
Показатель
доминирования в F2
|
|
|
0,35
|
|
n
|
Частоты
вариант (в % и числах) в классах со средними значениями
|
|
40,9
|
50,1
|
59,3
|
68,5
|
77,7
|
86,9
|
96,1
|
105,3
|
114,5
|
123,7
|
132,9
|
|
100
|
Вираж
|
16,7
|
57,5
|
23,3
|
2,5
|
|
|
|
|
|
|
|
100
|
F2
|
0,2
|
1,5
|
17,7
|
36,3
|
18,7
|
12,8
|
8,4
|
2,5
|
|
|
|
100
|
Айсберг
|
|
0,8
|
3,3
|
11,7
|
20
|
26,3
|
29,2
|
6,7
|
|
|
|
20
|
69
|
26
|
3
|
|
|
|
|
|
|
|
407
|
|
1
|
6
|
72
|
156
|
76
|
52
|
34
|
10
|
|
|
|
120
|
|
|
1
|
4
|
14
|
24
|
34
|
35
|
8
|
|
|
Таб. 2. Длина метелки
|
Статистические
параметры родительских форм и гибрида F2
|
|
|
|
|
Папаметм
|
Pmin
|
Pmax
|
F2
|
|
|
Численность
выборки
|
-
п
|
119
|
119
|
407
|
|
|
Среднее
значение выборки
|
-X
|
10.1
|
19.8
|
16,8
|
|
|
Среднее
квадратическое отклонение
|
-Sx
|
0,89
|
2,72
|
2,25
|
|
|
Ошибка
выборочной средней
|
-Soui.
|
0,08
|
0,25
|
0,11
|
|
|
Коэффициент
вариации
|
-
Cv%
|
8,8
|
13,7
|
13,4
|
|
|
Максимальная
варианта
|
-
Max
|
12,0
|
26,0
|
23,0
|
|
|
Минимальная
варианта
|
-
Min
|
8,0
|
13,0
|
12,0
|
|
|
Нижний
доверительный интервал
|
-
X-3Sx
|
11,6
|
10,0
|
|
|
Верхний
доверительный интервал
|
-
X+3Sx
|
12,8
|
28,0
|
23,5
|
|
|
Показатель
доминирования в F2
|
|
|
0,38
|
|
|
n
|
Частоты
вариант (в % и числах) в классах со средними значениями
|
|
7
|
8,9
|
10,7
|
12,6
|
14,5
|
16,4
|
18,3
|
20,2
|
22,1
|
24
|
25,9
|
|
100
|
Вираж
|
22,7
|
73,1
|
4,2
|
|
|
|
|
|
|
|
|
100
|
F2
|
|
|
3,9
|
27
|
35,9
|
18,9
|
12,5
|
1,7
|
|
|
|
100
|
Айсберг
|
|
|
0,8
|
8,4
|
8,4
|
21
|
33,6
|
21
|
4,2
|
2,5
|
|
119
|
0
|
27
|
87
|
5
|
|
|
|
|
|
|
|
|
407
|
0
|
|
|
16
|
110
|
146
|
77
|
51
|
7
|
|
|
|
119
|
0
|
|
1
|
10
|
10
|
25
|
40
|
25
|
5
|
3
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Таб. 3. Общее число колосков
|
Статистические
параметры родительских форм и гибрида F2
|
|
Параметры
|
Pmin
|
Pmax
|
F2
|
|
Численность
выборки
|
-
п
|
120
|
119
|
345
|
|
Среднее
значение выборки
|
-X
|
71,2
|
114,2
|
161,0
|
|
Среднее
квадратическое отклонение
|
-Sx
|
23,60
|
39,95
|
46,26
|
|
Ошибка
выборочной сродней
|
-SOUL
|
2,15
|
3,66
|
2,49
|
|
Коэффициент
вариации
|
-Cv%
|
33,2
|
35,0
|
28,7
|
|
Максимальная
варианта
|
-
Max
|
141,0
|
232,0
|
291,0
|
|
Минимальная
варианта
|
-
Min
|
29,0
|
23,0
|
30,0
|
|
Нижний
доверительный интервал
|
-
X-3Sx
|
0,4
|
-5,6
|
22,2
|
|
Верхний
доверительный интервал
|
-
X+3Sx
|
142,0
|
234,1
|
299,8
|
|
Показатель
доминирования в F2
|
|
|
3,17
|
|
n
|
Частоты
вариант (в % и числах) в классах со средними значениями
|
|
3,3
|
39,1
|
74,8
|
110,5
|
146,3
|
182
|
253,5
|
289,2
|
324,9
|
360,7
|
|
100
|
Вираж
|
29,2
|
50,8
|
19,2
|
0,8
|
|
|
|
|
|
|
|
100
|
F2
|
0,6
|
4,6
|
19,4
|
30,1
|
25,5
|
13,6
|
4,9
|
1,2
|
|
|
|
100
|
Айсберг
|
4,2
|
26,1
|
34,5
|
26,9
|
5,9
|
2,5
|
|
|
|
|
|
120
|
0
|
35
|
61
|
23
|
1
|
|
|
|
|
|
|
|
407
|
0
|
2
|
16
|
67
|
104
|
88
|
47
|
17
|
4
|
|
|
|
120
|
0
|
5
|
31
|
41
|
32
|
7
|
3
|
|
|
|
|