Изучение дискретной модели популяции при помощи программы Model Vision Studium
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ
РОССИЙСКОЙ ФЕДЕРАЦИИ
Федеральное государственное бюджетное
образовательное учреждение высшего профессионального образования
«КУБАНСКИЙ ГОСУДАРСТВЕННЫЙ
УНИВЕРСИТЕТ»
Физико-технический факультет
Кафедра
физики и информационных систем
ДИПЛОМНАЯ
РАБОТА
ИЗУЧЕНИЕ
ДИСКРЕТНОЙ МОДЕЛИ ПОПУЛЯЦИИ С ПОМОЩЬЮ ПРОГРАММЫ MODEL
VISION
STUDIUM
Специальность
200402 - Инженерное дело в медико-биологической практике
Работу выполнил Литвинов
Николай Александрович
Научный руководитель В. В.
Супрунов
Краснодар
2013
РЕФЕРАТ
Целью дипломной работы является исследование
дискретной модели популяции при помощи программы Model
Vision Studium
( MVS )
Объектом разработки дипломной работы является
интегрированная графическая оболочка для быстрого создания интерактивных
визуальных моделей сложных динамических систем Model
Vision Studium.
Основными задачами дипломной работы является
создание дискретной модели популяции и определение развития численности
популяции, в зависимости от начального числа особей
В результате выполнения дипломной работы была
создана дискретная модель численности популяции, а так же были построены
временные диаграммы, на которых показаны зависимости численности особей от
времени.
СОДЕРЖАНИЕ
ВВЕДЕНИЕ
.
СТРУКТУРА ПРОГРАММЫ MODEL
VISION STUDIUM
.1
Определение моделирования
.2
Model
Vision
Studium
.3
Карта поведения
.4
Блоки и связи
.5
Объективно-ориентированное моделирование
.6
Численное решение
.7
Интегрированная среда
.
ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ ПОПУЛЯЦИИ
.1
Определение популяции
.2
Динамика популяции
.3
Структура популяции
.
ДИСКРЕТНАЯ МОДЕЛЬ ПОПУЛЯЦИЙ
ЗАКЛЮЧЕНИЕ
СПИСОК
ИСПОЛЬЗУЕМЫХ ИСТОЧНИКОВ
ВВЕДЕНИЕ
В настоящее время нельзя назвать область
человеческой деятельности, в которой в той или иной степени не использовались
бы методы моделирования.
Компьютерное моделирование используют для
исследования системы до того, как она спроектирована, с целью определения
чувствительности ее характеристик к изменениям структуры и параметров объекта
моделирования и внешней среды. На этом этапе проектирования системы
компьютерное моделирование используют для анализа и синтеза различных вариантов
и выбора максимально эффективного при принятых ограничениях.
Программный комплекс Model
Vision
Studium
(MVS)
предназначен
для моделирования сложных динамических систем. MVS является представителем
подхода к решению проблемы моделирования сложных динамических систем,
основанного на использовании схемы гибридного автомата. Использование карты
поведения при описании переключений состояний, а также непосредственное
описание непрерывных поведений системы системами алгебро-дифференциальных
уравнений предоставляет большие возможности в описании гибридного поведения со
сложной логикой переключений.
Сложные по внутренним связям и большие по
количеству элементов системы экономически трудно поддаются прямым способам
моделирования и зачастую для построения и изучения переходят к имитационным
методам. Появление новейших информационных технологий увеличивает не только
возможности моделирующих систем, но и позволяет применять большее многообразие
моделей и способов его реализации. Совершенствование вычислительной и
телекоммуникационной техники привело к дальнейшему развитию методов машинного
моделирования, без которых невозможно изучение процессов и явлений, а также построение
больших и сложных систем.
Вычислительная техника облегчает задачи
исследователей, связанных с математическими расчетами. Построение
математической и компьютерной моделей системы, работать в этой области стало
проще, поскольку математические расчеты и выкладки без применения ЭВМ занимают
много времени.
Передо мной была поставлена задача: исследовать
дискретную модель популяции при помощи интерактивной графической оболочки Model
Vision Studium
( MVS )
1. СТРУКТУРА ПРОГРАММЫ MODEL
VISION
STUDIUM
.1 Определение моделирования
Замещение одного объекта другим с целью
получения информации о важнейших свойствах объекта-оригинала с помощью
объекта-модели называется моделированием.
Таким образом, моделирование может быть
определено как представление объекта моделью для получения информации об этом
объекте путем проведения экспериментов с его моделью. Теория замещения одних
объектов (оригиналов) другими объектами (моделями) и исследования свойств
объектов на их моделях называется теорией моделирования
Определяя гносеологическую роль теории
моделирования, т. е. ее значение в процессе познания, необходимо прежде всего
отвлечься от имеющегося в науке и технике многообразия моделей и выделить то
общее, что присуще моделям различных по своей природе объектов реального мира.
Это общее заключается в наличии некоторой структуры (статической или
динамической, материальной или мысленной), которая подобна структуре данного
объекта. В процессе изучения модель выступает в роли относительного
самостоятельного квазиобъекта, позволяющего получить при исследовании некоторые
знания о самом объекте.
Если результаты моделирования подтверждаются и
могут служить основой для прогнозирования процессов, протекающих в исследуемых
объектах, то говорят, что модель адекватна объекту.
При этом адекватность модели зависит от цели
моделирования и принятых критериев. Обобщенно моделирование можно определить
как метод опосредованного познания, при котором изучаемый объект-оригинал
находится в некотором соответствии с другим объектом-моделью, причем модель
способна в том или ином отношении замещать оригинал на некоторых стадиях
познавательного процесса. Стадии познания, на которых происходит такая замена,
а также формы соответствия модели и оригинала могут быть различными:
) моделирование как познавательный процесс,
содержащий переработку информации, поступающей из внешней среды, о происходящих
в ней явлениях, в результате чего в сознании появляются образы, соответствующие
объектам;
) моделирование, заключающееся в построении
некоторой системы-модели (второй системы), связанной определенными
соотношениями подобия с системой-оригиналом (первой системой), причем в этом
случае отображение одной системы в другую является средством выявления
зависимостей между двумя системами, отраженными в соотношениях подобия, а не
результатом непосредственного изучения поступающей информации. [12, 15]
Следует отметить, что с точки зрения философии
моделирование - эффективное средство познания природы. Процесс моделирования
предполагает наличие объекта исследования; исследователя, перед которым
поставлена конкретная задача; модели, создаваемой для получения информации об
объекте и необходимой для решения поставленной задачи. Причем по отношению к
модели исследователь является, по сути дела, экспериментатором, только в данном
случае эксперимент проводится не с реальным объектом, а с его моделью.
Такой эксперимент для инженера есть инструмент
непосредственного решения организационно-технических задач. Надо иметь в виду,
что любой эксперимент может иметь существенное значение в конкретной области
науки только при специальной его обработке и обобщении.
1.2
Model
Vision
Studium
Model Vision
Studium - это интегрированная графическая оболочка для быстрого создания
интерактивных визуальных моделей сложных динамических систем и проведения
вычислительных экспериментов с ними.
Пакет предназначен для численного моделирования
гибридных систем. Гибридная система - это специальная математическая модель
реальных объектов, обладающих одновременно "непрерывными" и
"дискретными" свойствами. Такие системы достаточно трудно
формализовать, так как необходим единый язык для описания как непрерывных и так
и дискретных аспектов поведения, но еще труднее корректно численно решить
возникающую при этом математическую задачу.
Моделирование является основным методом
исследований во всех областях знаний и научно обоснованным методом оценок
характеристик сложных систем, используемым для принятия решений в различных
сферах инженерной деятельности.
1.3
Карта поведения
Это ориентированный граф, в котором узлам
приписываются некоторые локальные поведения, а дуги интерпретируются как
переходы от одного поведения к другому. В каждый момент времени один из узлов
является текущим. Один из узлов должен быть помечен как начальный, он
становится текущим при создании карты состояний. Смена текущего узла происходит
в результате срабатывания переходов. Локальные поведения являются экземплярами
соответствующих классов. Когда узел становится текущим, создается экземпляр
приписанного ему локального поведения, который уничтожается, когда узел
перестает быть текущим. Начальное поведение проявляется при создании экземпляра
блока. Карта поведения представляет собой простую и наглядную форму визуального
представления смены поведений. В частном случае элемента с чисто дискретным
поведением, его карта поведения, всем узлам которой приписаны пустые поведения,
вырождается в обычную карту состояний с некоторыми упрощениями. Упрощения
связаны с запретом перехода в локальную карту поведения. Это связано с
трактовкой поведения как параметризованного класса, конкретный экземпляр
которого создается при входе в узел карты поведения и уничтожается при выходе.
Элемент с чисто непрерывным поведением трактуется как гибридный с картой
поведения, состоящей из единственного узла, которому приписано непрерывное
поведение.
С картой поведения связан гибридный
интерпретатор поведений, объединяющий интерпретатор карт состояний,
интерпретатор интерактивных воздействий и менеджер численных методов. В
MVSTUDIUM имеется достаточно богатый набор численных методов предназначенных
для воспроизведения поведения гибридных систем. Это программные реализации
методов решения нелинейных алгебраических уравнений, систем обыкновенных
дифференциальных уравнений и систем алгебро-дифференциальных уравнений. Для каждой
группы задач имеется свой автоматический решатель, цель которого обеспечивать
получение решения на заданном временном участке, любыми доступными, из числа
имеющихся в пакете, методами.
Любой автоматический решать начинает решать
каждую новую задачу наименее трудоемких методом, а если при этом возникают
какие-либо сложности, пытается подобрать метод, способный их преодолеть. Каждая
смена поведения рассматривается как новая численная задача. Автоматический
решатель может не справится с поставленной задачей, и тогда пользователь может
попытаться управлять выбором метода самостоятельно.
Пользователю доступны те же программные
реализации численных методов, что и автоматическому решателю. Для отладки или
первоначального решения задач в форме обыкновенных дифференциальных уравнений,
когда у пользователя нет отчетливого представления о свойствах решаемой задачи
или в описании поведения допущена ошибка, которую и надо найти, предусмотрена
группа отладочных численных методов.
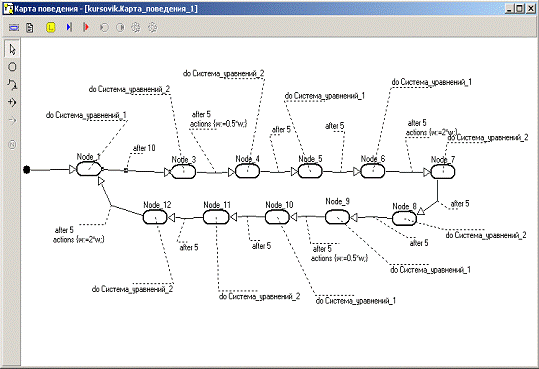
Рисунок 1 - Карта поведения системы
1.4 Блоки и связи
Основным « строительным элементом » описания в MVS
является блок. Блок - это некоторый активный объект, функционирующий
параллельно и независимо от других объектов непрерывном времени. Блок является
ориентированным блоком, все взаимодействия которого с окружающим миром
осуществляется только через его входы и выходы, осуществляющие интерфейс блока.
Таким образом блок информацию от входа к выходу. Все остальные свойства
инкапсулированы внутри него. В общем случае в описании блока следующие
элементы: входы, выходы, параметры конструктора переменные состояния,
поведение, локальная структура, анимация. Входы, выходы и переменные состояния
являются фазовыми переменными и все вместе составляют фазовый вектор блока.
Идентификаторы блоков и переменных записываются по привычным программистам
правилам, и могут содержать помимо латинских также и буквы русского алфавита.
Для описания модели разработан специальный язык
- Model Vision
Language (MVL).
Он имеет как графическую, так и текстовую формы. Все переменные это языка
должны быть явно описаны. В MVL
существуют следующие типы данных: скалярные, регулярные и комбинированные типы,
а также типы, определяемые пользователем.
Скалярные типы это -вещественные, целые,
булевский, перечислимые, символьный и строковый типы. Регулярные типы
представлены одномерными и двумерными массивами с произвольными элементами,
матрицами и векторами с вещественными элементами, а также списками. К
комбинированным типам относят записи с полями различных типов. Для передачи
информации о дискретных событиях используется специальный тип данных
"сигнал".
MYL
поддерживает объектно-ориентированную технологию. По аналогии с классом и.
используемыми в объектно-ориентированных языках, в MVL
введено новое понятие - класс блоков Описание нового блока всегда строится как
описание класса блока, а затем уже используются экземпляры того класса Для
конкретных экземпляров блока, как и обычно, могут быть указаны специальные
значения параметров конструктора Блоки могут соединяться между собой
однонаправленными функциональными связями и входить в состав других блок,
образуя иерархическую структуру.
При описании локальной структуры возможно
использование сложных многокомпонентных связей, агрегирование и расщепление
связей. Возможно использование регулярных структур блоков В версии 3.0
агрегирование и расщепление связей, а также массивы и списки блоков не
поддерживаются. Конкретная моделируемая система, с которой будет проводиться
вычислительный эксперимент. собирается в окне редактора виртуального стенда из
экземпляров классов, определенных в данном проекте, и или из библиотечных
классов.
1.5 Объективно-ориентированное
моделирование
В MVS
описание блока всегда строится как описание класса блок. Блоки могут быть
простыми, то есть не содержать внутренних или локальных блоков и составными.
Составные блоки образуют некоторую иерархическую структуру. Экземпляры блока
могут быть динамическими и статическими. Статическими называются экземпляры,
время жизни которых совпадает с временем проведения вычислительного
эксперимента над главным блоком. Динамические экземпляры создаются и
уничтожаются во время вычислительного эксперимента. Статический экземпляр
локального блока создается автоматически при создании экземпляра составного блока.
К проекту могут быть присоединены любые ранее
созданные библиотеки классов, таким образом при создании своей модели вы можете
использовать уже готовые классы блоков. Из любого разработанного проекта можно
автоматически получить библиотеку классов, а любой класс из вашего проекта
может быть добавлен в существующую библиотеку. Пакет поставляется со
стандартной библиотекой классов SysLib,
содержащей набор наиболее типовых линейных блоков, нелинейных блоков, и
источников сигналов.
1.6 Численное решение
На этапах редактирования исходного описания и
генерации кода выполняемой модели проверяется корректность непрерывных
поведений (нет ли присваиваний входам, не стоит ли одна и та же переменная
дважды в левой части формулы или дифференциального уравнения, не образуют ли
формулы алгебраические циклы, равно ли число неизвестных числу алгебраических
уравнений и т.п.), выполняется локальная сортировка формул и вырабатываются
некоторые признаки свойств для этапа выполнения. Поскольку мы используем
ориентированные связи, соединения не могут сделать локально корректные
поведения некорректными.
С картой поведения связан гибридный
интерпретатор поведений, объединяющий интерпретатор карт состояний,
интерпретатор интерактивных воздействий и менеджер численных методов. В MVS
имеется достаточно богатый набор численных методов предназначенных для
воспроизведения поведения гибридных систем. Это программные реализации методов
решения нелинейных алгебраических уравнений, систем обыкновенных
дифференциальных уровней и систем алгебро-дифференциальных уравнений. Для
каждой группы задач имеется свой автоматический решатель, цель которого
обеспечивать получение решения на заданном временном участке, любыми
доступными, из числа имеющихся в пакете, методами.
Любой автоматический решатель начинает решать
каждую новую задачу наименее трудоемких методом, а если при этом возникают
какие-либо сложности, пытается подобрать метод, способный их преодолеть. Каждая
смена поведения рассматривается как новая численная задача. Автоматический
решатель может не справится с поставленной задачей, и тогда пользователь может
попытаться управлять выбором метода самостоятельно. Пользователю доступны те же
программные реализации численных методов, что и автоматическому решателю.
Для отладки или первоначального решения задач в
форме обыкновенных дифференциальных уравнении, когда у пользователя нет
отчетливого представления о свойствах решаемой задачи или в описании поведения
допущена ошибка, которую и надо найти, предусмотрена группа отладочных
численных методов Экземпляр непрерывного локального поведения создается всякий
раз, когда узел становится текущим и уничтожается, когда он перестает быть
текущим, при ном изменяется состав совокупной глобальной системы уравнений
модели Всякий раз при перестройке совокупной системы уравнений производится
сортировка формул с учетом соединений, а также проверяется появление
алгебраических циклов. В случае нахождения такого цикла одна из связей
принудительно разрывается к заменяется алгебраическим уравнением.
При переключении совокупная система может
поменять тип (например, из чисто дифференциальной стать
алгебро-дифференциальной или чисто алгебраической). Следовательно, в тот момент
менеджер численных методов должен на основе анализа признаков, оставленных
компилятором, выбрать начальный численный метод. На непрерывном участке между
переключениями может иметь место внешнее воздействие, скачком меняющее значение
некоторых переменных, или переход условных выражений в правых частях уравнений
или формул на другую ветвь. В этом случае менеджер численных методов считает,
что имеет дело с новой численной задачей, и выполняем соответствующие действия.
Кроме того, во время решения совокупной системы могут изменяться ее численные
свойства (например, система может оказаться жесткой), которые потребуют смены
численного метода "на ходу". В опциях модели может быть указан режим
«автомата» и тогда менеджер сам будет переключать численные методы и
настраивать их параметры пока не найдет решение или не исчерпает свои
возможности. В последнем случае решение будет остановлено с соответствующим
сообщением и пользователь должен будет сам выбирать новый метод
1.7 Интегрированная среда
Оболочка пакета MVS
представляет собой многооконную среду, позволяющую редактировать проект,
осуществлять экспорт в текстовое представление и импорт из текстового
представления, подключать библиотеки классов, создавать выполняемые мотели и
запускать их. В пакетe
MVS используются
специальное внутреннее представление информации о префекте в форме деревьев
объектов с перекрестными связями, хранящееся в объектно-ориентированной базе
данных MVBase (на диске
вся информация о проекте находится в одном файле с расширением *.mvb)
Редакторы интегрированной среды немедленно проверяют синтаксическую и
семантическую правильность вводимых конструкций. При генерации «кода»
выполняемой модели на промежуточном языке производится проверка правильности
всего проекта в целом и обнаружении ошибки ошибочная конструкция выводится в
окно соответствующего редактора.
Рисунок 2
- Редактор интегрированной среды
Пакет MVSTUDIUM
относится к компилирующим средам моделирования, т.е. выполняемая модель
создается как некоторый исполняемый модуль операционной среды, который может
запускаться и без интегрированной оболочки пакета. Возможны два вида
выполняемой модели:
) визуальная интерактивная модель с отладочными
возможностями;
) "скрытая" модель с максимальным
быстродействием.
Визуальная модель является имитацией некоторого
виртуального испытательного стенда, где все можно посмотреть и все изменить.
Пользователь может наблюдать динамику процессов на временных и фазовых
диаграммах в ходе моделирования, в окнах карт поведения, где отображаются
текущий узел и срабатывающие переходы, в окнах структурных схем, где отображается
прохождение сигналов по связям, в окнах фазового вектора, где динамически
выводятся текущие значения переменных, а также с помощью типовых 2D и
3D-анимационных элементов и анимационных окон, которые он сам может создать.
Пользователь может в любой момент изменить
значение любой переменной модели. а также менять значение непрерывно с помощью
интерактивных анимационных элементов. Можно задать план прогона модели, в
котором указывается последовательность изменений значений переменных и подач
сигналов во времени. Можно задать останов по времени, по истинности логического
условия, по срабатыванию перехода, по входу в указанный узел.
При появлении ошибки в ходе вычислений
диагностируется блок и поведение, где произошла ошибка, а в случае, если при
создании выполняемой модели были установлены отладочные опции, то и выводится
текст соответствующего уравнения или оператора. Имеется встроенный калькулятор,
в котором можно задавать любые выражения с именами переменных модели. В MVS
визуальная модель это модуль * . EXE.
* “ Скрытая ” модель предназначена исключительно для многократно повторяющихся
вычислений и не имеет ни окон, ни визуальных элементов управления, все, что она
может, это вычислить значение переменных модели в заданной временной точке. В MVS
это специальная DLL.
“ Скрытая ” модель нужна для специальных надстроек ( получение фазового
портрета системы, получение параметрической зависимости, оптимизация и т.д.).
Выполняемая модель одновременно является сервером автоматизации, поддерживающим
интерфейс IMVSAuto.
Этот интерфейс позволяет осуществлять чтение и запись значений переменных
моделей и управление прогоном модели из других приложений.
2. ОСНОВЫЕ ПОЛОЖЕНИЯ ТЕОРИИ
ПОПУЛЯЦИИ
.1 Определение популяции
Виды живых организмов обеспечивают устойчивое
поддержание биогенного круговорота в биогеоценозе на уровне популяции.
Изучением популяций занимаются достаточно давно, и к настоящему времени
сложилось определенное понимание особенностей их функционирования. Под
популяцией понимают исторически сложившуюся естественную общность особей живых
организмов одного вида, связанных генетически, населяющих общие места обитания
и реализующих закономерные функциональные взаимодействия.
По А.М. Гилярову (1990, с. 38): «популяция - это
любая, способная к самовоспроизведению, совокупность особей одного вида, более
или менее изолированная в пространстве и времени от других аналогичных
совокупностей того же вида». Особи в популяции скрещиваются друг с другом и
таким образом размножаются. Без этого условия вид из экосистемы исчезнет.
В современной популяционной экологии эту
общность организмов рассматривают как биологическую систему надорганизменного
уровня, обладающую собственными функциями и структурно обусловленную (С.С.
Шварц, 1980, И.А. Шилов, 2000).
Рассмотрение популяций обычно проводят с двух
главных позиций. Во-первых, популяция является формой жизни конкретного вида
живых организмов в условиях среды их обитания. В этом случае осуществляется
важнейшая функция популяции - обеспечение выживания живых организмов
составляющего ее вида и воспроизведение вида в данных условиях. Данная функция
обеспечивается ориентированными адаптациями (приспособительными изменениями)
отдельных особей вида, имеющих морфологическую общность, а также
сформированными закономерными взаимоотношениями. Все это поддерживает и
регулирует возможности размножения (воспроизводство) особей вида. Процесс этот
носит непрерывный характер, выражающийся в смене отдельных индивидов внутри
популяции, и поэтому при оптимальных условиях жизнедеятельности существование популяции
может продолжаться неограниченно долго - популяция бессмертна как структурная
единица.
Во-вторых, популяция, являясь структурной
единицей биогеоценоза (экосистемы), выполняет одну из главнейших его функций, а
именно участвует в биологическом круговороте. В данном случае реализуется
видоспецифическая особенность типа обмена веществ. Популяция представляет вид в
экосистеме и все межвидовые взаимоотношения осуществляются в ней на
популяционном уровне. Устойчивая реализация функции участия в биогенных
процессах определяется специфическими механизмами авторегуляции, которые
создают условия самоподдержания популяции как системы в изменяющихся внутренних
и внешних факторах среды обитания. [5]
В целом популяции обладают всеми признаками
самостоятельной функционирующей биологической системы. Популяция имеет
пространственное расположение входящих в нее элементов, они структурированы,
что позволяет оптимально использовать ресурсы среды, а также создает условия
для бесперебойного осуществления внутрипопуляционных взаимоотношений.
Однако особи, входящие в вид, формирующий данную
популяцию, не являются абсолютно идентичными в осуществлении
внутрипопуляционных взаимоотношений. Эти взаимоотношения образуют определенную
функциональную систему. Даже при условии общего морфофизиологического типа
(сходстве) отдельные особи неравноценны по своему участию в жизнедеятельности
популяции. Таким образом, популяция структурирована еще и функционально. Особи
в популяциях находятся в состоянии постоянного обмена информацией, что представляет
собой специфический механизм взаимодействия живых организмов На разных уровнях
развития живой материи продукционные процессы проявляют себя по-разному, но их
феноменологическое описание всегда включает рождение, рост, взаимодействие с
внешней средой, в том числе с другими особями своего вида или других видов,
смерть особей. Именно это обстоятельство позволяет применять сходный
математический аппарат для описания моделей роста и развития у таких, казалось
бы, удаленных друг от друга по лестнице уровней организации живой материи, как
клеточная популяция и сообщество видов в экосистеме. Описание изменения
численности популяции во времени составляет предмет популяционной динамики.
Популяционная динамика является частью биологии математической, наиболее
продвинутой в смысле формального математического аппарата, своего рода
"математическим полигоном" для проверки теоретических идей и
представлений о законах роста и эволюции биологических видов, популяций,
сообществ. Возможность описания популяций различной биологической природы
одинаковыми математическими соотношениями обусловлена тем, что с динамической
точки зрения, рост и отбор организмов в процессе эволюции происходит по
принципу "Кинетического совершенства" (Шноль, 1979) [2] Преимущества
математического анализа любых, в том числе популяционных, процессов, очевидны.
Математическое моделирование не только помогает строго формализовать знания об
объекте, но иногда (при хорошей изученности объекта) дать количественное
описание процесса, предсказать его ход и эффективность, дать рекомендации по
оптимизации управления этим процессом. Это особенно важно для биологических
процессов, имеющих прикладное и промышленное значение - биотехнологических
систем, агробиоценозов, эксплуатируемых природных экосистем, продуктивность
которых определяется закономерностями роста популяций живых организмов,
представляющих собой "продукт" этих биологических систем.
2.2
Динамика популяций
Численность и биомасса популяций обычно
подвержены большим колебаниям во времени. Изменение численности, биомассы
организмов во времени называют динамикой популяций. Существуют два основных
типа динамики численности - периодическая и непериодическая. Периодические
колебания происходят главным образом под влиянием закономерно изменяющихся факторов
среды. У некоторых видов млекопитающих, птиц, рыб, насекомых наблюдаются четкие
периодические изменения численности, то есть ее вспышки чередуются со спадами.
Однако численность особей в популяциях может колебаться во времени без
определенной периодичности. Большое влияние на популяции, такие их свойства как
продолжительность жизни особей, плодовитость, которые определяют численность,
оказывают температура, освещенность, влажность. Действие многих факторов
становится более жестким с увеличением плотности популяций: это - трофические
условия, в том числе обостряющиеся конкуренция, хищничество, паразитизм,
заболеваемость. Почти всегда вызывают изменения численности популяций
антропические воздействия: в сельском и лесном хозяйствах, при рыболовстве и других
видах промысла, при разрушении местообитаний человек способствует уменьшению их
численности; при охране каких-то видов, наоборот, их численность возрастает.
Эти колебания («волны жизни») вызываются, таким образом, многими воздействиями
со стороны как живой, так и неживой природы. [10]
Динамика численности популяций складывается при
взаимодействии основных популяционно-динамических процессов: 1) рождаемости, 2)
смертности, 3) скорости роста, 4) иммиграции новых особей из других популяций,
5) эмиграции некоторых особей за пределы ареала данной популяции.
Рождаемость характеризует частоту появления
новых особей. Под рождаемостью понимают количество особей (яиц, семян,
эмбрионов), производимых в единицу времени в расчете на одну самку. Близкое к
приведенному определение приводится А. М. Гиляровым: «рождаемость определяют
как число особей (яиц, семян и т. д.), родившихся (отложенных, продуцированных)
в популяции за некоторой промежуток времени». Различают максимальную
(абсолютную, физиологическую, предельно-возможную) рождаемость и реализуемую
(экологическую) рождаемость, или просто рождаемость.
Максимальная рождаемость - это образование
теоретически максимально возможного количества новых особей в идеальных
условиях, когда отсутствуют лимитирующие факторы и размножение ограничивается
лишь физиологическими факторами. У каждой данной популяции эта величина
постоянная, она характеризует динамическую, эволюционно приобретенную силу
вида. Реализуемая рождаемость - это увеличение популяции за счет появления на
свет новых особей при фактических, реальных условиях среды. Данная величина
может варьировать в зависимости от физических, химических и прочих условий
среды.
Показатель смертности характеризует гибель
особей в популяциях. По определению, смертность - это количество особей,
умирающих в единицу времени в расчете на особь в популяции. Учитываются все
погибшие особи независимо от причины смертности (старость, элиминация
хищниками, болезнями и т. д.) Существует некая теоретическая максимальная
смертность - постоянная величина, которая характеризует гибель особей в
идеальных условиях, когда популяция не подвергается воздействию лимитирующих
факторов. Практически более важна реализуемая (экологическая) смертность, т. е.
величина, которая подобно экологической рождаемости, зависит от реальных
условий биотической и абиотической среды.
Представляет интерес величина, связанная со
смертностью, обратная ей - выживаемость, т. е. число или доля выживших особей.
Разность между рождаемостью и смертностью есть
некий результирующий параметр, который определяет реальную динамику численности
у данной популяции. Популяция может находиться в состоянии динамического
равновесия, если естественная убыль особей равна их возобновлению. Существенно
то, что антропические воздействия на популяцию могут изменять как рождаемость,
так и смертность (например, увеличивать смертность особей данного вида). [14]
Величина прироста популяции за единицу времени в
расчете на одну особь представляет скорость роста популяции. По мере роста
популяции происходит снижение доступных каждой особи ресурсов среды. При
истощении ресурсов рост популяции тормозится и в конце концов прекращается.
Популяции разных видов обладают удивительной способностью к быстрому росту
численности. Этот вопрос рассматривали Аристотель (4 в. до н.э.), Макиавелли
(около 1525 г.), позднее Бюффон (1751 г.). Ч. Дарвин обратил внимание на
многочисленные случаи поразительно быстрого размножения некоторых животных в
природном состоянии, когда условия особенно благоприятствовали. Он
распространил идею геометрического роста, когда численность популяции растет в
геометрической прогрессии (в этом случае график увеличения числа особей в ряде
поколений представляет собой экспоненциальную, или логарифмическую, кривую) на
все виды животных и растений, положив постулат о высоком репродуктивном
потенциале видов в основу своей теории естественного отбора.
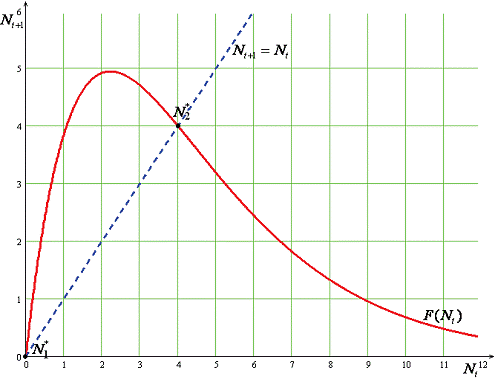
Рисунок 3 - Зависимость роста популяции от
времени
Заслуживает внимания рост народонаселения в
глобальном масштабе. В ранние исторические времена прирост населения за
поколение (20 лет) составлял 1,2%, в 17 в. он повысился до 7,2%, к 1930 г.
достиг - 36%, причем в наше время нет признаков того, что кривая роста
приближается к какому-то уровню насыщения. Поскольку с увеличением числа людей
на нашей планете также увеличивается потребление продуктов питания,
использование естественных источников сырья, загрязнение среды обитания и т.
д., все изменения в динамике численности человечества обусловливают
вышеназванные явления. Ограничение роста населения является важным шансом
выживания человечества.[9]
Кроме рассмотренных характеристик - рождаемости,
смертности, скорости роста на величину популяцию влияют эмиграция, иммиграция и
общая миграция.
Миграция - это особый случай перемещения особей,
когда почти вся популяция на время уходит из определенного района. Сезонные или
суточные миграции позволяют организмам использовать оптимальные условия среды в
таких местах, где они не могли бы жить постоянно. Перебираясь с места на место
вслед за перемещением оптимальных условий, такие виды могут сохранять высокую
активность, поддерживать большую плотность популяции и в те периоды, когда
немигрирующие виды переходят в неактивное состояние (в состояние диапаузы или
зимней спячки).
В динамике численности популяций большую роль
играют межвидовые отношения, что издавна вызывало у экологов большой интерес и
способствовало разработке теорий взаимоотношений конкурирующих видов, хищника и
жертвы, паразита и хозяина. Широкую известность получили работы А. Лотки (1923,
1925 гг.) и В. Вольтерры (1926 г.). Вольтерра проанализировал взаимоотношения
между конкурирующими видами, разработал математическую модель в системе жертва
- хищник. Пользуясь уравнениями, предложенными Вольтеррой, можно определять
условия, при которых устанавливается равновесие между плотностями популяций
жертвы и хищника, то есть система жертва - хищник становится стабильной. Лотка
предложила математическую модель взаимодействия животных в системе паразит -
хозяин, показала, что истребление особей хозяина паразитами является функцией
численности не только паразитов, но и хозяев. Лотка сделала заключение, что той
или иной численности хозяина соответствует определенная численность паразита.
По мере возрастания плотности популяции хозяина увеличивается плотность
популяции паразита. Повышение же численности паразита приводит к снижению
численности хозяина, а последнее опять снижает количество паразитов. И так
волна за волной происходят периодические колебания численности популяций
хозяина и паразита с небольшими отклонениями от какого-то оптимального уровня.
Здесь действует динамическая саморегулирующаяся система. Модель взаимодействия
популяций в системе паразит - хозяин соответствует модели взаимодействия
хищника и его жертвы. Уравнения, предложенные Лоткой и Вольтеррой можно
использовать для моделирования взаимоотношений в указанных системах,
определять, какой должна быть плотность каждой популяции, чтобы другая не имела
возможности увеличивать свою численность.
Изучение хищничества, паразитизма как факторов,
регулирующих численность популяций, влияющих на их величину, показывает, что
отрицательное влияние хищников, паразитов обычно не велико, если оба вида -
хищник и его жертва, хозяин и паразит - существуют совместно уже на протяжении
длительного времени. Известно много ярких примеров, касающихся паразитизма.
Так, у коренных жителей Африки выработался относительный иммунитет к малярии, и
это обеспечивает выживание как паразита - малярийного плазмодия, так и хозяина
- человека. Трипаносома, вызывающая сонную болезнь у человека, живет в крови
своих основных хозяев - крупных травоядных животных, не причиняя им вреда.
Заражение человека лентецами в обычных условиях не приводит к смерти. Однако в
любом из этих случаев установившееся тонкое равновесие может быть нарушено в
результате каких-то изменений в экосистеме или иных событий. Серьезные
последствия возникают, в частности, когда хищник и жертва (или паразит и
хозяин) встречаются друг с другом впервые. Если экосистема, в которую они
входят, будет выведена из равновесия, хищники и паразиты могут вызвать резкие
изменения численности популяции.
В природе действуют другие факторы, влияющие на
динамику численности популяций. Связано это со следующими причинами. Для
некоторых видов решающее значение имеют физические факторы. Численность особей
в популяциях могут лимитировать такие факторы, как нехватка природных ресурсов
(например, пищи или мест, пригодных для размножения), недоступность этих
ресурсов и недостаток времени для размножения (короткий влажный сезон, короткий
день, например в Арктике).
У крупных организмов, жизненные циклы которых
довольно продолжительны, размеры популяций определяются не столько физической
средой, сколько взаимодействием между отдельными особями или взаимоотношениями
их с конкурентами, хищниками и паразитами.
Из внутренних факторов на величину популяции
могут оказывать влияние различные физиологические или поведенческие факторы, а
иногда те и другие одновременно. Если, например, плотность популяции
какого-нибудь грызуна чрезмерно возрастает, то животные чаще встречаются между
собой. Возникают драки, условия жизни в целом становятся более напряженными
(«стрессовыми»), и это ведет к увеличению надпочечников; связанное с этим
нарушение гормонального баланса отрицательно сказывается на спаривании и
размножении; кроме того, при скученности возрастает смертность. [17]
Любой фактор как регулятор численности популяций
- лимитирующий или благоприятный - является либо независимым от плотности (НП),
либо зависимым от плотности (ЗП). Влияние факторов ЗП может быть прямым, то
есть усиливаться с увеличением плотности, и обратным. Факторы, для которых
характерно прямое влияние, еще называют «управляющими плотностью» (в частности,
это один из главных механизмов, предотвращающих перенаселение). Как правило, НП
- абиотические факторы (климатические, химические, физические и т. п.), ЗП -
биотические факторы (конкуренция, паразиты, патогенные организмы, влияние эндо-
и экзометаболитов). НП-факторы могут вызывать драматические изменения
плотности, смещение уровня емкости экологической ниши. Роль НП-факторов более
выражена в нестабильных системах, а ЗП-факторов - в стабильных. Одновременное
действие НП- и ЗП-факторов обнаружено при изучении динамики популяций моллюска
Acmaea, живущего на камнях в литоральной зоне морей. Динамика популяции этого
моллюска регулируется, главным образом, ее плотностью (ЗП). Однако известны
случаи увеличения смертности после суровых зим (НП), когда разрушается
субстрат, на котором оседают моллюски, что является основной причиной
смертности.
Таким образом, колебания численности природных
популяций («волны жизни»), их величина обусловлены сложным взаимодействием
факторов - естественного темпа размножения того или иного вида, «сопротивления»
среды, отношениями между хищником и жертвой, паразитом и хозяином,
физиологической и поведенческой реакцией на перенаселенность и др., в целом,
многими воздействиями со стороны как живой, так и неживой природы. В итоге,
регуляция численности популяций осуществляется факторами внешней среды и внутрипопуляционными
факторами, преимущественно через рождаемость и смертность, представляя собой
результат взаимодействия их со всеми условиями существования.
«Волны жизни» резко осложняют планирование
эксплуатации данной популяции, поскольку ежегодное изъятие (отстрел, промысел)
одного и того же числа особей может означать, что в один год будет изъято,
скажем, лишь 5% особей, а в другой год, когда численность популяции упадет в 10
раз, - 50% особей от существующего состава популяции. Кроме того, колебания численности
призывают человека увеличить минимальную теоретически допустимую численность
популяции.
Популяции животных, растений, грибов и
микроорганизмов обладают способностью к естественному регулированию
численности, то есть при более или менее значительных колебаниях они остаются в
состоянии динамического равновесия, на каком-то уровне между верхним и нижним
пределами. Это обеспечивается действием специфических приспособительных
механизмов, основанных на том, что поступление энергии, необходимой для выживания
популяции, не превышает некоторого уровня и обеспечивает, таким образом,
размеры данной популяции. Способность популяции поддерживать устойчивость
благодаря способности к саморегулированию через собственные регулирующие
механизмы называется гомеостазом популяции. Так, рост численности популяции
приводит к истощению запасов пищи, за которым следует снижение рождаемости
организмов, увеличение их смертности (отрицательные связи), а, следовательно, и
снижение численности. Последнее, в свою очередь, увеличивает запасы пищи, что
вызывает рост рождаемости и численности популяции (положительные связи).
Равновесное состояние популяции (состояние динамического равновесия) является
кратковременным и достигается за счет быстрого чередования положительных и
отрицательных обратных связей.
Для оптимизации отношений человека с природой
важно учитывать численность популяции, принимать во внимание то, что на
численность популяции может повлиять истощение нужных ей ресурсов из-за
сокращения кормовой базы, конкуренция со стороны домашних животных,
вытаптывание почвы и ухудшение ее аэрации, снижение кислорода в воде при
загрязнении и евтрофировании. Человек может искусственно регулировать
численность популяций, например, животных путем запрещения охоты или
ограничения ее сроков на некоторые виды, ввода лицензий.
В целом, численность популяции, скорость ее
роста (в более общем смысле - скорость ее изменения, динамика численности)
являются весьма лабильными параметрами, высокочувствительными к воздействию
абиотических, биотических, антропических факторов. Поэтому человек должен
хорошо представлять все особенности той популяции, которая эксплуатируется,
чтобы обеспечить воспроизводство, стабильное длительное ее существование.
Сложность этой задачи увеличивается в силу многочисленных связей между
популяциями разных видов, населяющих одну территорию.
Совокупность особей одного вида, длительно
обитающих на определенной территории, обладающих общим генофондом и
способностью к обмену генетической информацией, отграниченных пространственно
от других групп, называется популяцией. Термин «популяция», введенный в
экологию В. Иогансеном в 1903 г. (от лат. populus - народ, население), получил
широкое распространение.
Однако такое определение популяции справедливо
для особей, размножающихся половым путем. Для организмов, не имеющих полового
способа размножения (некоторые грибы), оно также справедливо, за исключением
условия об обмене генетической информацией. Например, если сосновый лес по
берегам реки будет представлен одной популяцией сосны, так как река не является
препятствием для переноса пыльцы ветром, то для многих грибов река будет
непреодолимой границей, и особи грибов одного вида, но произрастающие на разных
берегах, будут разными популяциями.
В основу классификации популяций могут быть
положены разные принципы. По классификации Н.П.Наумова (1963), основанной по
ландшафтно-биотопическому принципу, различают популяции:
. Локальные, или элементарные, заселяющие
элементарные (однородные) участки рельефа. Во многих районах Беларуси хорошо заметны
всхолмления, оставленные отступившим ледником. Часто они покрыты сосновыми
лесами. Но сосны, произрастающие на вершинах холмов (дюн), заметно отличаются
по высоте стволов, их диаметру, густоте и развитию кроны от сосен, формирующих
участки сосновых лесов, расположенных в междюнных понижениях. Расположенные
рядом, такие участки представлены элементарными популяциями сосны обыкновенной.
Граница между элементарными популяциями невыразительна, размыта.
. Экологические популяции занимают территории с
более или менее однородными экологическими условиями, формируются как
совокупность элементарных популяций. Часто их называют экотопами. Например,
белка (Sciurus vulgaris) заселяет разные типы леса. Поэтому могут быть четко
выделены сосновые, елово-пихтовые и другие ее популяции. Они слабо изолированы
одна от другой, и обмен генетической информацией между ними происходит довольно
часто, но реже, чем между элементарными популяциями.
. Географические, занимающие в пределах ареала
крупные ландшафтно-географические регионы, слагаются из экологических
популяций. Особи географических популяций различаются морфологически,
физиологически, временем прохождения фенофаз. Географические популяции
достаточно четко разграничены и относительно изолированы. Например, в границах ареала
сосны обыкновенной (от Балтийского моря до Тихого океана) выделяют лапландскую
(на Кольском полуострове), кулундинскую (Казахский мелкосопочник) и ангарскую
(в бассейне р. Ангара) популяции. Различия между особями этих популяций столь
существенны, что их часто выделяют как подвиды.
По времени существования различают следующие
популяции.
Постоянные - возникают в оптимальных условиях и
способны к самовоспроизведению. Примером могут служить популяции ели на севере
Беларуси, популяции коренных народностей Севера, популяции волка.
Временные - существуют и восстанавливаются в
большей степени за счет притока особей извне. Например, популяции европейцев на
Крайнем Севере, в Африке и других регионах.
Можно выделять популяции по типу размножения,
возрастной структуре и т.д. Знание типа популяции очень важно, так как его
анализ подсказывает человеку, каким образом и какими особями следует заселять
тот или иной район, какие меры предпринимать для сохранения видов, какие
способы применять для их использования.
Любой вид представлен отдельными группировками -
популяциями, образующими элементарную биологическую надорганизменную систему.
Это обусловлено по крайней мере двумя причинами: индивидуальной и групповой
адаптивной изменчивостью (физиологической, морфологической, экологической,
поведенческой), определяющей формирование специфических особенностей;
разнообразием экологических условий, в которых обитает та или иная группа
особей вида. При этом чем сложнее расчленение территории, которую занимает вид,
тем более четко обособляются популяции. [ 7]
Таким образом, вид - это система популяций.
Однако в формировании популяционной структуры вида не меньшее значение имеют и
биологические особенности: подвижность особей, степень оседлости и
привязанности к территории, способность преодолевать естественные преграды.
Если особи постоянно перемещаются в пределах ареала, то вид имеет небольшое
число крупных популяций (северные олени, песцы). Границы между популяциями
обычно проходят по естественным рубежам - проливы, реки, горные хребты.
Например, Татарский пролив отграничивает сахалинскую популяцию белки от
материковой. Если ареал невелик, то вид состоит из одной популяции (например,
ель Глена на острове Сахалин).
Очень четко прослеживается популяционная
структура вида при обособленности условий обитания, например, если сравнивать
группы особей одного и того же вида рыб в разных озерах. Наоборот, сплошное
заселение затрудняет выделение популяций (например, у колорадского жука). В
этом случае границы между популяциями обычно смазанные, нечеткие. Между
популяциями осуществляется обмен отдельными особями, причем это может
происходить регулярно или эпизодически. Например, часть молодых ворон остается
на местах зимовок, образуя пары с оседлыми птицами этого вида. При высоком
уровне весенних паводковых вод многочисленные водоемы на какое-то время
образуют единую систему, в которой происходит обмен особями популяций. Таким
образом происходит обмен генетическими программами между отдельными
популяциями, адаптации особей к изменяющимся условиям среды, их выживание или
гибель (естественный отбор).
Связи между популяциями поддерживают вид как
единое целое. Различия могут затрагивать все стороны биологии вида:
морфологические, физиологические, размножение. Например, особи популяций соболя
на Сахалине, Сихотэ-Алине и Верхоянском хребте различаются величиной, длиной
шерсти и ее окрасом. Самки зайца-беляка в Карелии приносят зайчат 2 раза в год,
а в Беларуси - до 4 раз.
Установление численности популяции важно для
определения величины заказников, заповедников, организуемых для сохранения того
или иного вида. Именно с этой точки зрения многие заповедники не выполняют
своей главной задачи. Например, Воронежский заповедник не решил бы задачи
расселения бобра по всему его ареалу, если бы там не создали бобровой фермы, на
которой этих животных выращивают в неволе. Вся площадь заповедника позволяет
жить в естественных условиях лишь нескольким семьям.
Вот почему в настоящее время ставится вопрос об
установлении заповедного режима не на ограниченных территориях, а в целых
ландшафтно-географических регионах.
Любая популяция характеризуется определенными
показателями или свойствами. А так как популяция - это надорганизменная
структура, то ей свойственны показатели, характерные как для отдельных
организмов (например, рост, развитие), так и характеризующие ее как группу
особей (рождаемость, смертность, плотность). Наиболее существенными
характеристиками являются численность, плотность, рождаемость, смертность,
рост, прирост, структура.
Численность популяции - это общее число особей
(или их биомасса, или связанная в биомассе энергия) на данной территории или в
данном объеме. Этот показатель варьирует и колеблется в широких пределах.
Зависит от условий обитания, наличия пищи, врагов, соотношения полов в
популяции, размеров, составляющих ее особей. В определенный период времени
численность дает возможность судить в сравнительном плане о преобладании той
или иной популяции, но совершенно не характеризует ее состояние и перспективы
развития. [ 6]
Плотность - это количество особей популяции
(либо биомасса), отнесенное к единице занимаемого пространства или объема
(количество деревьев на 1 га, масса циклопов в 1 м3 воды). Различают среднюю
плотность, то есть количество особей, отнесенное к единице площади всего ареала
или объема, и экологическую - количество особей, отнесенное к единице
заселенного пространства или объема. Последняя обычно выше (ведь ни одно
растение не образует сплошного покрова на площади всего ареала!). Этот
показатель свидетельствует о различиях в условиях обитания организмов: чем они
благоприятнее, тем выше плотность. Существуют верхние и нижние пределы
плотности, обусловливающие возможность существования популяции, так как высокая
плотность вызывает отмирание особей, а низкая - препятствует своевременному
спариванию, опылению и появлению потомства. Зная динамику плотности популяции,
можно планировать заготовку сырья, древесины, отлов рыбы и т.д.
Смертность характеризует гибель особей популяции
и показывает число погибших за единицу времени, рассчитанное на определенное
количество живущих особей (антипод рождаемости). Минимальная смертность
наблюдается в идеальных условиях, экологическая - в конкретных. Смертность
может быть специфичной для определенной возрастной группы. Высокая смертность
говорит о неблагоприятных условиях среды. Однако большее значение в практике
имеет другой показатель - количество выживших особей, или выживаемость. Если
графически выразить количество выживших особей во времени, учитывая при этом
продолжительность жизни популяции в процентах к средней величине ее, что дает
возможность сравнивать популяции с разной продолжительностью жизни, то получим
кривые выживаемости. [1] Прирост популяции - это разница между рождаемостью и
смертностью. Его считают показателем жизненности популяции. При высокой
рождаемости высокая смертность не всегда является для популяции отрицательным
явлением. Прирост говорит о перспективности развития популяции, о возможности
человека эксплуатировать такую популяцию без риска ее уничтожения.
Благоприятные условия обитания, особенно питания, способствуют более быстрому
приросту популяции. Давно известен факт, что высокий урожай семян сосны, ели и
других деревьев в лесу определяет повышение плодовитости белок, других грызунов
и увеличение их прироста.
Рост популяции означает увеличение ее
численности. На этот показатель оказывают влияние многочисленные внешние и
внутренние факторы, и прежде всего - соотношение рождаемости и смертности. Рост
популяции сдерживают неблагоприятные условия, недостаток корма, хищники. В процессе
роста новое поколение не всегда может «увидеть» родителей. Такой рост
называется дискретным, или неперекрывающимся, что наблюдается у однолетних
растений. Поколение, принесшее семена, отмирает, и новое поколение, которое
образуется из семян, не застанет предыдущее. У многолетних растений и животных
рост непрерывный или с перекрывающимися поколениями, так как каждое новое
поколение некоторое время «живет» с родителями. По характеру увеличения
численности различают два типа роста популяции.
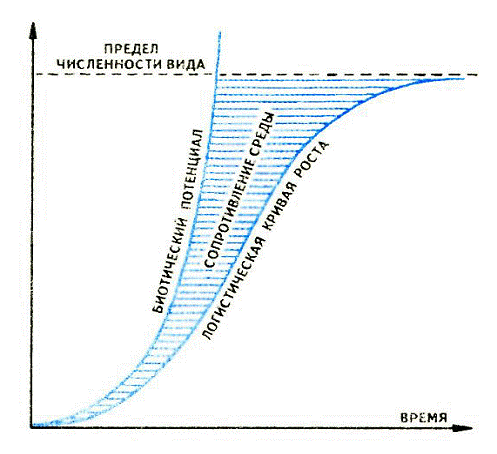
Рисунок 4 - Кривые роста численности популяций
Пространство между экспоненциальной кривой
(биотический потенциал) и логистической кривой соответствует сопротивлению
среды.
Экспоненциальный тип характеризуется быстрым,
постоянным, нелимитированным средой увеличением числа особей. Этот тип роста
теоретически может наблюдаться в любой популяции на начальном этапе, когда нет
хищников и достаточно пищи. Таким образом происходило увеличение численности
кроликов, когда их завезли в Австралию. Так же росла и популяция опунции,
завезенной на этот же континент, пока не появился вредитель (кактусовая
огневка), сдерживающий ее рост.
Если бы не было сдерживающих факторов, то пара
домашней мухи за один сезон дала бы потомство 5,6∙1012 особей. Однако на
определенном этапе роста популяции вступают в силу внутрипопуляционные
механизмы регуляции численности, сопротивление среды (недостаток пищи и убежищ,
враги), и рост приостанавливается. Таким образом, в природных условиях в чистом
виде популяции не имеют такого типа роста. При логистическом росте численность
популяции увеличивается в соответствии с логистическим уравнением (с
непостоянной скоростью снижается по мере возрастания рождаемости). В процессе
роста различают фазу положительного ускорения, логарифмического роста, фазу
отрицательного ускорения и фазу развития. Но и такой тип роста в природных
условиях часто нарушается в силу стихийных бедствий (пожары, засуха, извержения
вулканов и пр.) [13]
2.3
Структура популяции
Под структурой популяции понимают соотношение
особей по какому-либо признаку. Исходя из этого различают половую, возрастную,
генетическую, экологическую и пространственную структуры.
Половая структура отражает соотношение в
популяции особей разного пола. В идеальных условиях половая структура должна
быть оптимальной, что не означает соотношения особей по полу 1:1. Прежде всего
половая структура популяции зависит от биологических особенностей особи. Если
создаются моногамные семьи, то соотношение 1:1 можно считать нормальным явлением
(у лебедей). У полигамных особей такого быть не может (гаремы котиков, моржей).
В момент оплодотворения у большинства видов
наблюдается равное соотношение зигот по признаку пола, однако физиологические,
экологические, поведенческие и другие различия самцов и самок нарушают это
соотношение уже на момент рождения. Как правило, вероятность гибели особей
одного пола может быть выше, чем другого. Например, у комаров выше гибель
самок, поскольку они кровососущие, а самцы питаются нектаром. У пингвинов при появлении
птенцов из яиц нет разницы в количестве особей разного пола, но к 10-летнему
возрасту самок становится наполовину меньше. У фазанов, большой синицы,
утки-кряквы - более низкая смертность среди самцов. У рыжих лесных муравьев на
появление самцов и самок оказывает влияние температура среды. При температуре
ниже +20°С появляются самцы, а при более высокой - самки. У многих двудольных
на одном растении могут быть обоеполые цветки, а на другом только женские. На
появление особей того или иного пола у растений также оказывают влияние
температура, запас питательных веществ и другие экологические факторы.
Половая структура влияет на численность
популяции, ее плотность, рождаемость, возрастную структуру. Достаточно
вспомнить, какие демографические сдвиги повлекла за собой гибель значительной
части мужского населения во второй мировой войне.
Возрастная структура отражает не только
распределение особей популяции по возрастным группам, но и то, как меняется их
соотношение в разные сезоны года, при смене биогеоценозов, какова скорость
обновления популяции и каково взаимоотношение возрастных групп со средой.
Изучение возрастной структуры имеет большое значение для практической
деятельности, так как от принадлежности особи к той или иной возрастной группе
зависит степень изъятия растений и животных для использования человеком.
В демографии возрастные группы называют
когортами. Изучение их важно для решения социальных проблем (строительства
школ, детских садов, определения объема пенсионного фонда и т.д.). Определение
возрастной структуры в популяциях разных видов - у животных и растений,
однолетних и многолетних организмов имеет свою специфику. Но, независимо от
специфических особенностей, методика определения возрастной структуры
однотипна. Определяют возраст каждой особи, а если популяция большая, то делают
выборку: закладывают пробные площадки в лесу, проводят контрольные отловы рыбы
и т.д. Затем устанавливают величину класса возраста - временной промежуток в
днях, часах, годах, поделив возраст самой старой особи на число принятых
(произвольно) классов, и устанавливают границы классов. Допустим, после
переписи жителей обнаружилось, что их максимальный возраст равен 90 годам.
Принимаем условный раздел жителей на 9 классов. Поделив 90 на 9, получаем, что
возрастной класс составляет 10 лет. Значит, первый класс имеет границы от 0 до
10, второй - от 11 до 20, третий - от 21 до 30 лет и так далее до 90. После
этого всех особей распределяют по классам возраста и подсчитывают число их в
каждом классе. Это и будет возрастной структурой. Определяют также численность
каждого класса в процентах от общей численности популяции. Для большей
наглядности полученные цифровые данные представляют графически в виде
гистограмм. Естественно, что каждая популяция будет иметь свою гистограмму, но
тем не менее все их множество можно свести к трем типам, которые и являются
типами популяции в зависимости от возрастной структуры. [ 3, 16]
Помимо распределения особей животных по
возрастным классам обычно для удобства классы объединяют в возрастные группы:
молодые, средневозрастные и старовозрастные. Количество классов, объединенных в
эти группы, безусловно, различно у разных популяций. По преобладанию той или
иной возрастной группы различают популяции:
инвазионные, или внедряющиеся (предрепродуктивные),
в которых преобладают особи первых нескольких классов (молодняки);
репродуктивные, полноценные, или уравновешенные,
в которых снижение численности от самого молодого класса к последнему
происходит равномерно;
регрессивные, или вымирающие, в которых преобладают
особи последних возрастных классов (старовозрастные).
Выявление типа популяции имеет огромное значение
для проведения биотехнических мероприятий. Нельзя эксплуатировать внедряющиеся
и старовозрастные популяции, необходимо принимать меры для их сохранения
(подкармливать животных, устранять врагов и источники болезней). В
уравновешенных популяциях можно проводить обоснованные нормы отстрела, отлова,
вырубки. У однолетних растений подсчет особей производят не по возрастным
классам, а по возрастному состоянию. Выделяют периоды латентный, виргинильный
(от проростков до генеративных органов), генеративный и сенильный (старение).
Такое распределение растений очень важно для определения объемов и сроков
заготовки лекарственного сырья, начала сенокоса т.д. Известно, что с возрастом
у растений ослабевает способность к вегетативному размножению, и это
учитывается при восстановлении леса, выпасе скота на лугах.
У животных организмов наиболее устойчивы
популяции с длительным сосуществованием разных поколений (лоси, олени, слоны,
медведи). В зависимости от показателей рождаемости возрастная структура
значительно варьирует. Сокращение численности какой-то возрастной группы может
привести к деградации популяции или, наоборот, к чрезмерному ее увеличению.
У организмов, которые имеют одну генерацию в
течение года, все особи принадлежат к одной возрастной группе (некоторые виды
насекомых). Весной обычно из яиц бабочек появляются гусеницы, и в зависимости
от условий они могут иметь разные возрасты, но в конце концов окукливаются и
переходят во взрослую стадию. К зиме, после откладывания яиц, они погибают.
Самки майских жуков (очень опасные вредители,
поедающие корни сосны обыкновенной), в мае после откладывания яиц погибают.
Личинки живут 3-4 года и окукливаются. Поэтому в популяции присутствуют личинки
четырех генераций. Соотношение их численности будет зависеть от конкретных
условий во время лёта жуков.
Анализ возрастной структуры позволяет
прогнозировать численность популяции на протяжении ряда ближайших поколений.
Это важно для расчета динамики развития промысловых рыб, насекомых-вредителей.
С использованием математических моделей, учитывающих количественные воздействия
факторов среды на отдельные возрастные группы, получают достоверные прогнозы.
Генетическая структура определяет соотношение в популяции особей - носителей
определенных аллелей. Аллель (от греч. allelon - друг друга, взаимно) - одно из
возможных структурных состояний гена; определяет развитие того или иного
признака. Число организмов, несущих тот или иной аллель, называют частотой
данного аллеля. Например, у человека частота доминантного (господствующего)
аллеля, определяющего пигментацию кожи, волос и глаз, равна 99%, а рецессивный
аллель (подавленный) встречается с частотой 1% (общая частота 100% , или 1).
Если частота доминантного аллеля в генофонде данной популяции - Р, рецессивного
- Q, значит, Р + Q = 1. Таким образом, зная частоту одного аллеля, можно
определить частоту другого. Частота отдельных аллелей в генофонде позволяет
вычислить частоту генотипов, а генотип определяет фенотип (совокупность всех
признаков и свойств особи, формирующаяся на основе наследственности и влияния
внешней среды). В 1908 г. английский математик Дж. Харди и немецкий врач В.
Вайнберг вывели математическую зависимость между частотами аллелей и генотипов
в популяциях. Эта зависимость известна как закон Харди-Вайнберга: частоты
доминантного и рецессивного аллелей в данной популяции будут оставаться
постоянными из поколения в поколение при наличии следующих условий: размеры популяции
достаточно велики, скрещивание происходит случайно, новых мутаций не возникает,
все генотипы одинаково плодовиты, то есть не происходит отбора, поколения не
перекрываются, отсутствует обмен генетической информацией с другими
популяциями. А так как в природных условиях эти требования не могут быть
удовлетворены, то происходит и изменение частот аллелей. Генетическое же
равновесие выражается уравнением
Р2 + 2PQ + Q2 = 1. (1)
Так как для природных популяций характерно
стремление к стабильности, то очень важно соблюдать целый ряд условий при
отборе групп растений или животных по доминантному гену (высокая удойность
коров, высокая жирность молока, длинная шерсть у овец), чтобы не допустить
случайного скрещивания. В противном случае в группе высокопородных животных
появятся особи и гомозиготные по рецессиву, и гетерозиготные.
Экологическая структура отражает соотношение в
популяции особей различных экологических групп. Известно, что довольно
распространенный вредитель плодовых деревьев - кольчатый шелкопряд откладывает
яйца на яблоне, груше, вишне. Вышедшие из яиц гусеницы вредителя, обитающего на
вишне, никогда не переходят на яблоню, так как биохимический состав веществ в
листьях яблони отличается от листьев вишни и ферментные системы гусениц не
приспособлены их перерабатывать. [8, 9]
Лесоводам давно известно существование двух
фенологических форм дуба черешчатого: зимней и летней (или поздней и ранней). В
одном и том же лесном массиве деревья дуба в низких местах рельефа распускаются
значительно позже, чем на ровном месте или на возвышенностях, так как весной по
ночам в понижениях скапливается холодный воздух и образуются так называемые
морозобойные ямы. От пониженных температур в них листья погибают...
Знание биологических особенностей растений и
животных позволяет более эффективно использовать условия среды. Интересно, что
в Англии, которая по площади посевов пшеницы уступает Николаевской области
(Украина) почти в два раза, возделывается более 20 сортов пшеницы, а в
Николаевской области - всего 3. Английские фермеры учитывают и экспозицию
склона, и крутизну, и высоту холмов для подбора сортов, что позволяет с
максимальной эффективностью использовать природные условия. Пространственная
структура - это упорядоченное размещение особей популяции по занимаемой территории.
Такое размещение обусловлено тем, что на конкретном участке ареала популяции
может прокормиться лишь определенное количество особей. По территории ареала
особи могут распределяться двояко: одиночно и группами. Одиночное распределение
присуще немногим животным и в определенный период жизни (божья коровка,
жужелицы). Но даже одиночно проживающие животные на период спаривания и
выведения потомства образуют временные внутрипопуляционные группы. Групповое
распределение более распространено. Это объясняется прежде всего
неоднородностью условий обитания и биологическими особенностями каждого вида (у
растений при вегетативном размножении, при слабом распространении семян и
прорастании их вблизи материнской особи - дуб, бук, кедровые сосны).
И одиночное, и групповое распределение особей в
ареале популяции может быть равномерным и неравномерным. При равномерном
распределении особи и их группы расположены на одинаковом расстоянии друг от
друга. У растений в естественных условиях это наблюдается крайне редко. Примером
равномерного пространственного распределения могут служить агроценозы
(квадратно-гнездовые посевы). У животных такое распределение встречается при
оседлом образе жизни, когда каждая группа особей (семья, стая, табун) занимает
строго определенную территорию.
Неравномерное распределение присуще большинству
растений и животных и объясняется теми же причинами, которые обеспечивают
групповое распределение. В любом случае на характере распределения особей
сказываются приспособительные особенности, обеспечивающие оптимальное
использование среды.
Наиболее четко неравномерное распределение
выражено у растений. Скопления особей внутри популяций называют
микропопуляциями, субпопуляциями, отличающимися числом особей, плотностью,
возрастной структуры. Животные по типу использования жизненного пространства
делятся на оседлых и кочевых. При оседлом образе жизни используется
ограниченный участок территории, это имеет свои положительные стороны: животные
в нем свободно ориентируются, по кратчайшему пути следования находят корм,
делают запасы. Например, у белок есть основное гнездо и несколько
дополнительных. Однако если плотность популяции высока, реальна угроза
истощения ресурсов. Поэтому вся территория делится на участки, занимаемые одной
или группой особей. Распределение группами распространено гораздо чаще.
3. ДИСКРЕТНАЯ МОДЕЛЬ ПОПУЛЯЦИЙ
Численность популяции не изменяется непрерывно,
а представляет собой дискретную величину, что соответствует экспериментальным
данным по переписи реальных популяций. Существует некоторая популяция. Она
растет и размножается, но ее численность не может быть выше определенного числа
особей, (т. е. максимального числа, которые могут прокормиться). Требуется
определить развитие численности популяции, в зависимости от начального числа особей.
Математическая модель :
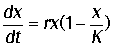 (2)
(2)
где х - численность популяции,- "емкость
популяции" определяется ограниченностью пищевых ресурсов, мест для
гнездования, и многими другими факторами, которые могут быть различными для
разных видов, r - скорость роста
численности.
Эксперимент 1.
Компьютерная модель
Первоначально задаем :
X=3, k=20,
r=2.5
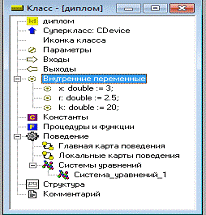
Рисунок 5 - Окно ввода переменных и уравнений
Получаем результат :
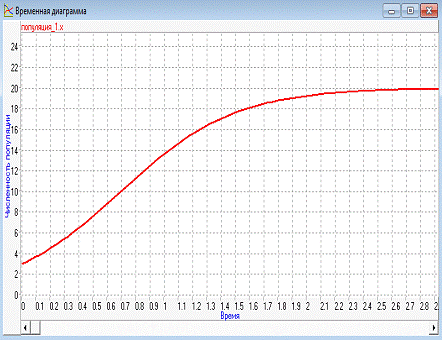
Рисунок 6 - Временная диаграмма при начальных
значениях
Эксперимент 2.
При изменении начального значения численности
популяции на x=10, получаем
следующий результат:
дискретный популяция особь информационный
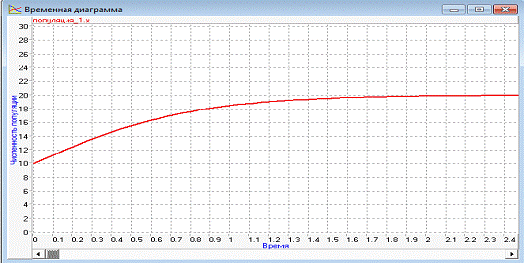
Рисунок 7 - Временная диаграмма при x=10
Эксперимент 3.
Изменяем начальное значение численности
популяции на х=40. Получаем следующие результаты:

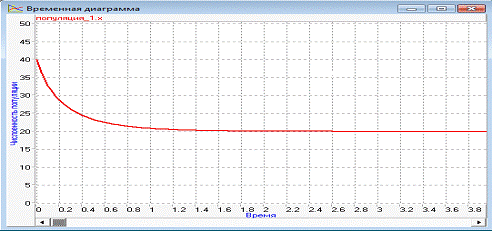
Рисунок 8 - Временная диаграмма при x=40
Численность популяции была больше
ограничивающего параметра, и уменьшилась до его размера
ЗАКЛЮЧЕНИЕ
) Процесс проектирования информационных систем,
реализующих новую информационную технологию, непрерывно совершенствуется.
В центре внимания инженеров оказываются все
более сложные системы, что затрудняет использование физических моделей и
повышает значимость математических моделей и машинного моделирования систем.
Машинное моделирование стало эффективным инструментом исследования и
проектирования сложных систем. Актуальность математических моделей непрерывно
возрастает из-за их гибкости, адекватности реальным процессам, невысокой
стоимости реализации на базе современных ЭВМ.
) Современные вычислительные средства позволили
существенно увеличить сложность используемых моделей при изучении систем,
появилась возможность построения комбинированных моделей, учитывающих все многообразие
факторов, имеющих место в реальных системах.
3) После создания дискретной
модели популяции из временной диаграммы следует, что при начальных значениях
численность популяции возрастет с течением времени и достигает определенной
ёмкости особей.
) Из проведенного ряда
экспериментов видно, что как бы мы не изменяли начальное значение особей,
количество особей не превысит определенного придела емкости популяции.
СПИСОК
ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
1. Бенькович,
Е.С. Практическое моделирование сложных динамических систем / Е.С. Бенькович,
Ю.Б. Колесоа, Ю.Б. Сениченко. - СПб.: БХВ, 2001.
. Ермаков,
С. М. Математический эксперимент с моделями сложных стохастических систем. -
СПб.: Изд. ГУ, 1993.
. Информационное
моделирование //
Информатика. - 2007. - №13. - С. 28-38.
. Клеймен,
Дж. Статистические методы в имитационном моделировании. - М.: Статистика, 1988.
. Колесов
Ю.Б., Сениченков Ю.Б. Имитационное моделирование сложных динамических систем-
М., 2009.
. Могилев,
Н.Н Элементы математического моделирования / А.В.Могилев, И.Я. Злотникова. -
Омск: ОмГПУ, 1995.
. Моделирование
экологических систем и процессов // Информатика. - 2007. - №14. - С. 2-4.
. Моисеев
Н.Н. Математические задачи системного анализа. - М.: Наука, 1981.
. Раскина,
И.И. Компьютерное моделирование: Учебно-методическое пособие / И.И. Раскина,
М.С. Сидоренко - Омск: Изд-во ОмГПУ, 2003.
. Савин,
Г.И. Системное моделирование сложных процессов. - М.: Фазис, 2000.
. Системное
обеспечение пакетов прикладных программ / Под ред. А. А.
Самарского.
- М.: Наука, 1990.
. Шилов
И.А. Экология. - М., 2006.
. Шеннон
Р. Имитационное моделирование систем. Искусство и наука. - М.: Мир, 1978.