Систематика живых организмов и теория эволюции
Систематика живых
организмов и теория эволюции
организм дарвин отбор линней
Первая классификация живых
организмов была построена Карлом Линнеем в 1761 г. Но у Линнея была так
сейчас называемая искусственная классификация - например, растения он
классифицировал только по числу тычинок. Естественно, классификация, основанная
только на одном признаке, не могла правильно отразить все эволюционные
взаимоотношения жизненных форм. Тем не менее, Линней выделил 67 семейств
растений, но, тем не менее, точного критерия деления на семейства Линней дать
не смог.
В 1803 году Жан-Батист
Ламарк впервые создал естественную систему
классификации живых организмов, основанную на многих признаках. Он выделил 114
семейств и 1597 родов живых организмов. Классификация Ламарка является основой
современной классификации, хотя она постоянно обновляется и усовершенствуется.
Но не обошлось и без казусов: например, Ламарк описал утку-самку (серое
оперение) и самца-селезня (оперение яркое и многоцветное) как разные виды.
До сих пор принята иерархическая
классификация, предложенная Ламарком: все живое делят на царства, типы,
классы (у растений - порядки), семейства, роды и виды.
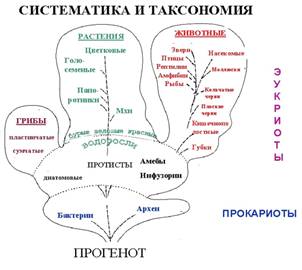
По современной классификации, все
жизненные формы разделяют на 5-6 царств: Бактерии и Цианобактерии - прокариоты,
не содержащие ядра, и эукариоты - Простейшие, Грибы, Животные и
Растения.
В последнее время выделяют еще одно
царство - Археобактерии. Это некоторые бактерии, живущие глубоко в толще земли.
У них не все в порядке с генетическим кодом - он слегка отличается от кода всех
других живых организмов, но, тем не менее, на 99% ему соответствует. Возможно,
они принадлежали к первым живым организмам, зародившимися на Земле, и их
отличия в генетическом коде - первичны и отражают еще не полностью
сформировавшуюся жизнь.
Вирусы. Вирусы сами по себе формой жизни не являются. Вирус лишен
основного свойства живого - самостоятельного самовоспроизведения. Вирус,
наряду с мобильными диспергированными генетическими элементами являются побочным
продуктом эволюции генома, своеобразной платой за эволюцию. Вирус -
это короткая молекула ДНК или РНК, покрытая белковой оболочкой, и содержащая
всего несколько генов: ген белка-капсида (оболочки вируса), ген белка,
обеспечивающего сорбцию на хозяине, и еще несколько генов, обеспечивающих
вирулентность. Вирусная ДНК содержит всегда очень сильный промотор,
обеспечивающий ей быструю транскрипцию в клетке. Происхождение вирусов полифилетическое
- все высшие организмы время от времени порождают новые вирусы.
Кроме вирусов существует еще один
вид экзотической инфекции, вызываемой неживым фактором биологического
происхождения. Это так называемые губчатые энцефалопатии человека и
домашних животных. К этим заболеваниям относятся куру-куру у человека -
смертельное заболевание, связанное с ритуальным каннибализмом, скрепи,
или вертячка у овец, коровье бешенство и ряд других инфекций человека и
животных. Вызываются эти смертельные заболевания в результате попадания в
организм прионов. Прионы представляют из себя нормальный белок,
содержащийся в нервных клетках данного организма, но находящийся в «неправильной»
конформации. У белков этого типа может быть два устойчивых конформационных
состояния: нормальное, с преобладанием альфа-спиралей, и аномальное,
прионовое, с преобладанием бета-складчатых структур. Попадая в организм,
белок с прионовой конформацией служит как бы кристаллической затравкой для
преобразования вторичной структуры нормального белка клеток. Все родственные
ему клеточные белки, находившиеся ранее в нормальной конформации, при контакте
с прионом меняют свою вторичную структуру, становясь сами прионами. Прионы, как
правило, видоспецифичны - только человеческий прион может заразить человека,
и относятся к классу медленных инфекций - так, инкубационный период куру-куру
может составлять 15-20 лет. Также как и вирусы, прионы являются порождением нас
самих, и представляют из себя своеобразную плату за совершенство, сложность
нашей организации.
Три этапа Великих
биологических объединений.
,5 млрд лет назад на Земле возникли первые эукариоты - ядерные организмы. До
этого Земля была заселена только прокариотами - в основном, бактериями и
синезелеными водорослями, не имеющими ядра, хроматина, а также механизмов
митоза и мейоза и являющихся гаплоидными. Возникновение эукариот было связано с
появлением у Земли кислородной атмосферы, и привело к первому мощному всплеску
видообразования.
Первый этап
биологической юниализации - это
симбиогенез эукариот. То, что эукариоты
возникли первоначально в результате симбиоза по крайней мере 4-х
различных организмов, сейчас не вызывает сомнений. Основные органеллы
эукариотической клетки - митохондрии, в которых происходит окислительное
фосфорилирование, хлоропласты и пластиды, в которых идет фотосинтез,
центриоли с тубулиновыми микротрубочками и жгутики, обеспечивающие
процессы митоза и мейоза и клеточное движение, клеточное ядро
- все это остатки первичных эндосимбионтов праэукариотической клетки.
Все эти органеллы имеют двойные цитоплазматические мембраны и содержат
свой геном. Правда, размеры геномов митохондрий и центриолей сейчас
весьма малы, только в геноме хлоропластов сохранилось немногим более 100 генов.
Основная масса генов симбионтов в процессе эволюции была захвачена ядром, то
есть перекочевала в ядерный геном. В эволюции происходило все более глубокое
подчинение первичных эндосимбионтов ядру. Митохондрии и хлоропласты содержат бактериальные
70S-рибосомы, тогда как цитоплазма эукариотической клетки имеет более
тяжелые 80S-рибосомы.
Возникновение эукариот было
гиганским эволюционным прорывом: впервые воедино в одной клетке были
собраны все мощнейшие энергетические механизмы, геном клетки был защищен
ядерной мембраной, возник эффективный механизм воспроизводства генетической
информации - митоз, клетка стала диплоидной, что обеспечило
дополнительную защиту генетической информации. Одновременно возник механизм мейоза
и кроссинговера, обеспечившие клетке эффективное размножение и
рекомбинационную изменчивость. Резко выросший в размерах геном нужно было
эффективно паковать в малом объеме ядра и эффективно передавать потомкам -
возник хроматин и хромосомы.
Хроматин - это сложный нуклеопротеиновый комплекс, содержащий,
кроме ДНК, массу белков. Белки хроматина подразделяются на два типа: гистоны
и негистоновые белки, последний тип является сборным, сюда также входят
многочисленные ферменты репликации, репарации и транскрипции. Именно гистоны
формируют структуру, необходимую для первого порядка упаковки ДНК - нуклеосому.
Гистоны являются сильноосновными белками, поэтому они имеют большое сродство к
фосфорным остаткам костяка цепей ДНК. Есть 5 типов гистонов: Н4, Н3, Н2а,
Н2b и Н1. Первые четыре гистона - по две молекулы каждого, итого, 8 молекул
- формируют кор нуклеосомы, имеющий шаровидную структуру. На
нуклеосомный кор наматывается молекула ДНК - приблизительно 2,5 оборота, или
около 200 нуклеотидов. Следующий участок ДНК наматывается на следующую
нуклеосому, но между ними остается линкерный район ДНК размером около 100
нуклеотидов. Этот район укрепляется гистоном Н1, который стаскивает соседние
нуклеосомы, сближая их между собой.
Нуклеосомы, да и хроматин в целом,
возникают в результате самосборки. Если смешать коровые гистоны в
эквимолярных количествах, то возникнут коры нуклеосом, а если к ним добавить
ДНК и гистон Н1, образуется нормальный хроматин.
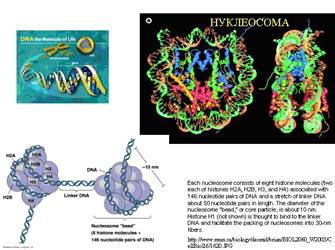
Структура гистоновых белков
настолько хорошо подогнана друг к другу и к структуре ДНК, а правильная
упаковка ДНК в клетке является настолько жизненно необходимой, что гистоны
являются одними из самых эволюционно консервативных белков: так, гистон Н4
коровы и гороха различается всего на одну аминокислотную замену!
Эукариотическое
объединение, сопровождавшееся
резким усложнением генома, и приобретением клеткой механизмов митоза
и мейоза подготовило почву для второго этапа юниализации:
возникновения многоклеточности.
Второй этап юниализации.
Это произошло около 600 млн лет назад, в конце архея -
начале фанерозоя, или эпохи явной жизни. Уже в первом периоде фанерозоя - кембрии
- появляются практически все современные типы животных, а также многоклеточные
растения. Это был второй резкий всплеск видообразования.
Прежде всего возникли колониальные
организмы, которые могут существовать в двух состояниях: свободноживущие клетки
и колонии слабодифференцированных клеток. Из простейших к колониальным формам
относится Fusarium policephalum - свободноживущая амеба, образующая в период своего размножения
ярко-розовые макроскопические колонии из дифференцировавшихся клеток на старых
пнях в наших лесах, из растений - Volvox, образующий колонии наподобие бластулы многоклеточных, из
животных - португальские кораблики и губки.
Иногда губок относят к
многоклеточным, но это не совсем так. Губки формируют сложный минеральный
скелет и содержат четыре типа слабодифференцированных клеток. Дифференциация у
губок обратимая: клетки при необходимости могут перестраиваться из одного типа
в другой. Более того, сама губка неделимым организмом, как таковым, не
является. Если ее измельчить, а затем оставить в покое в соответствующих
условиях, она возникнет заново. Здесь, фактически, работает тот же механизм
самосборки, только на более высоком уровне. Оставшиеся живые клетки губки
понемногу начнут формировать новый минеральный скелет ее тела, клетки будут размножаться
и дифференцироваться в соответствии с занимаемым ими местом в новом теле губки.
Первыми истинными многоклеточными
являются кишечнополостные, к которым относятся медузы, гидры, актинии,
кораллы. У кишечнополостных впервые начинается гаструляция - образование
первичной внутренней полости и первичного рта. Дифференциация
клеток у кишечнополостных уже глубокая, и организм представляет из себя
неделимое целое - из части медузы уже не получишь целую медузу. Но тем не
менее, тотипотентность клеток - их способность к дедифференцировке у
кишечнополостных еще остается - например, разрезав гидру на части, можно из
частей вырастить новых гидр.
Далее у многоклеточных возникает
жесткая иерархия клеток, подчиняющаяся возникшим системам нового уровня: нервной
и гуморальной (гормонной) регуляции. Дифференцированные клетки полностью
утрачивают свою самостоятельность и становятся подчиненными организму в целом.
На этом завершается второй этап биологической юниализации -
возникновение многоклеточности.
Третий этап юниализации. Уже у червей начинается ганглиолизация
- медленное преобразование сначала диффузной нервной системы кишечнополостных в
сначала частично, а потом полностью централизованную. Ганглиолизация - это
процесс дальнейшей цефализации, завершившейся только у позвоночных.
Первая попытка 3-го
этапа юниализации, заключавшаяся в
объединении отдельных особей в новый организм, созданный по старому принципу
многоклеточности, где каждый индивид выступал в роли отдельной клетки
и полностью подчинялся единому центру, оказалась в целом малопродуктивной. По
такому принципу в эволюции возникли общественные насекомые: муравьи,
пчелы и термиты. Видимо, какой-то краткий миг эволюции это было выгодно, но
потом такая надорганизменная ассоциация потеряла свой эволюционный потенциал:
муравейник практически неизменен последние 150 млн лет.
Не исключено, что провал первой
попытки третьего этапа юниализации был связан как с недостаточным уровнем
цефализации индивидов-насекомых, так и с неразвитостью общих централизованных
систем управления суперорганизмом: действительно, централизованное управление
муравейником можно сравнить разве что с диффузной нервной системой
кишечнополостных. Напротив, очень высокая специализация
элементов-индивидов суперорганизма уменьшила его пластичность и затруднила
его дальнейшее эволюционное развитие.
Наконец, вторая, и, будем
надеяться, успешная попытка третьего этапа юниализации началась с
появлением позвоночных, особенно млекопитающих. Началось возникновение
социума, или сообщества. Практически у всех млекопитающих зафиксировано сложное
социальное поведение. Это и стая волков, и стадо слонов, и прайд львов, не
говоря уж о сообществах приматов. Наибольшего развития социализация достигла у человека.
У нас возникла сложная иерархия общества на многих уровнях организации: государство,
другие социальные институты. Возникновение сложной иерархии человеческого
общества, в первую очередь, была связана с возникновением истинно знакового
языка, чего ранее не было у других млекопитающих, и с возникновением коллективной
памяти, или культуры.
Но если сравнивать наше общество с
многоклеточным организмом или таким же сильно иерархизированным организмом,
каким является муравейник, то наше общество пока не пошло дальше клеток губок,
находящихся на колониальном уровне объединения.
Таким образом, возникновение
социума - это закономерное естественное продолжение всей предыдущей
биологической эволюции, и принципиальных различий между биологическими и
социальными процессами не существует. Это всего лишь новый уровень
организации. На нем продолжают работать прежние, биологические законы, но
появляются и новые, социальные - точно также, как при переходе с клеточного на
организменный уровень
Теория эволюции
В начале 19 в. Жан-Батист Ламарк
предложил первую концепцию эволюции органического мира. В 1809 г. вышла
его «Философия зоологии». До этого в биологии господствовал креационизм,
считалось, что виды неизменны с момента творения. Ламарк тоже не отрицал акт
творения, но он сочетался у него с последующим механистическим развитием, изменчивостью.
Природу Ламарк отождествлял с Богом - отсюда возникала движущая сила эволюции -
стремление природы к совершенству. Механизм эволюции по Ламарку был
следующий: на растения и низших животных влияет сама среда и формирует их, у
высших животных основная движущая сила - упражнение - неупражнение органов.
Благоприобретенные признаки по Ламарку наследуются. (Но очень
скоро Вейсман показал, что благоприобретенные признаки не наследуются -
известные эксперименты с хвостами у мышей.) Поскольку виды пластичны и
постоянно меняются под воздействием среды, между видами нет границ - Ламарк
пришел к отрицанию существования видов.
На протяжении первой половины 19 в.
Ламарк не был понят и о его теорию практически не вспоминали, потому что в
биологии продолжал господствовать креационизм. Ламаркизм как явление возник
гораздо позже, в качестве противовеса теории эволюции Ч. Дарвина. Для внедрения
идеи эволюционизма в биологию требовался иной подход - не механистический, как
у Ламарка, а статистический, нужно было рассматривать не отдельные особи, а
вид, как единое целое. Это было сделано только Ч. Дарвином.
В первой половине 19 в. креационизм
продолжает господствовать. Считается, что виды статичны и неизменны. Для
объяснения смены видов в геологической истории известный французский
палеонтолог Жорж Кювье выдвигает теорию катастроф: в истории было
множество актов творения, за каждой геологической катастрофой следовал новый
акт творения - всего Кювье насчитал 27 актов творения, по числу известных тогда
геологических слоев.
В 1859 г. Ч. Дарвин издает свое «Происхождение видов». Дарвин не публиковал его в
течение 15 лет, ждал благоприятного момента.
Основные предпосылки возникновения
теории Дарвина были следующие:
успехи селекционеров по выведению
пород животных - искусственный отбор;
достижения эмбриологии и
морфофизиологии;
Теория Мальтуса о народонаселении -
борьба за существование;
успехи морфологии - изменчивость;
Основные движущие силы эволюции по
Дарвину - наследственность, изменчивость и естественный отбор.
Изменчивость у Дарвина ненаправленная,
случайная. Эволюция, в противовес Ламарку, не имеет цели. Изменчивость сама
неадаптивна, но ее наличие - адаптивно.
Понятие естественного отбора
введено Дарвиным по аналогии с искусственным отбором и из его представлений о
борьбе за существование, основанных на огромной величине коэффициентов
размножения (теория Мальтуса). По Дарвину, естественный отбор имеет постепенный
характер: он приводит к постепенному накоплению у вида адаптивных изменений. Но
это приводит к скачкообразным изменениям в геологических масштабах времени.
Основное возражение против теории Дарвина было высказано вскоре после выхода
«Происхождения видов» английским клерком Дженкиным. Суть его проста: Если одна
особь приобрела адаптивное, полезное изменение, то при скрещивании с другой
особью, этого адаптивного признака не имеющей, полезное приспособление
разбавится вдвое, в следующем поколении - вчетверо, и т.д., и, таким образом,
исчезнет в ряду поколений. Это возражение получило название «Кошмара
Дженкина». Поскольку генетики тогда не существовало, а представления о
наследственности сводились к стихийной теории кровей, преодолеть кошмар
Дженкина Дарвин не смог до конца своей жизни. Чтобы хоть как-то избавиться от
этого кошмара, Дарвин предложил теорию пангенезиса, которая по сути
следовала ламаркистским представлениям о наследовании благоприобретенных
признаков.
Тем не менее, сразу с появлением
теории Дарвина идеи эволюционизма захватили практически все области
биологических наук. Появилось множество эволюционных теорий, рассматривающих, в
основном, отдельные факторы эволюционного процесса. Затем, в 1-й половине 20 в.
началось бурное развитие генетики, и кошмар Дженкина наконец-то был разрешен,
возникли новые представления о наследственности и изменчивости. В 20-60-е годы
20 в. произошло возникновение и формирование Синтетической теории эволюции
(СТЭ).
СТЭ вобрала в себя знания из
генетики, молекулярной биологии, систематики, палеонтологии и многих других областей
биологии. Основы СТЭ заложены Четвериковым, его учениками и единомышленниками.
Это Тимофеев-Ресовский, Добржанский, Фишер, Райт, Холдейн, Симпсон и многие
другие.
В 1926 г. Четвериков своими работами по генетическому анализу так называемых «волн
жизни» - регулярных колебаний численности популяций - положил начало новому
направлению в биологии - генетике популяций, или популяционной генетике.
При высокой численности популяции вид устойчив, в нем наблюдается постоянное соотношение
частот аллелей, но постепенно происходит увеличение внутривидовой изменчивости
за счет накопления мутаций в гетерозиготном состоянии. Гетерозиготность
нарастает, и признаки вида постепенно расшатываются.
При снижении численности популяции происходит резкий отбор адаптивных вариаций и снижение
общего разнообразия. Частоты аллелей значительно изменяются, и когда
численность популяции опять нарастает, наблюдаются уже совсем не те частоты
аллелей, что были раньше. Сдвиг частот происходит как за счет действия отбора,
так и за счет генетического дрейфа - эффекта основателя. Таким образом
происходит трансформация вида - видоизменение. Но для акта
видообразования необходима еще репродуктивная изоляция.
Далее, на основе генетики популяций,
с привлечением массы данных из многих других областей биологии формируются
основные представления Синтетической теории эволюции.
Основные положения
Синтетической теории эволюции
1. Материалом для эволюции
служат мелкие дискретные изменения наследственности - мутации.
2. Мутационная изменчивость
носит случайный характер.
. Основным движущим фактором
эволюции является естественный отбор.
. Поток генов возможен лишь
внутри вида.
. Вид - это генетически
целостная система, состоящая из генетически отличных, но репродуктивно
неизолированных единиц - популяций.
. Между разными видами
существуют биологические репродуктивные барьеры.
. Наименьшей эволюционирующей
единицей является популяция, а не особь.
. Эволюция носит постепенный,
градуалистический характер.
. Эволюция имеет дивергентный
характер - одна предковая популяция может породить несколько новых видов, и у
каждой пары видов есть единая предковая популяция.
Формы естественного
отбора
Различают три основные формы
естественного отбора: движущий, стабилизирующий и дизруптивный.
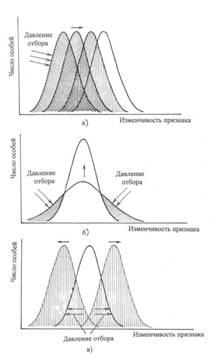
Рис. а - движущий, б -
стабилизирующий и в-дизруптивный.
Движущий отбор вызывает прогрессивное и направленное изменение признака или
свойства, переставшего соответствовать новым условиям среды. Изменение признака
может происходить как в сторону его усиления, так и в сторону ослабления. В
результате действия движущей формы отбора может происходить приобретение нового
признака (возникшего, например, в результате мутации и подхваченного отбором),
или его утрата. Движущий отбор действует при изменении условий внешней среды,
или когда популяция находится в процессе приспособления к новой среде
(например, при миграции и заселении новых пространств). Классический пример
движущего отбора - промышленный меланизм у бабочек березовой пяденицы
(Эдинбург, Великобритания).
Дизруптивный отбор благоприятствует
сохранению крайних форм и элиминирует промежуточные. Это приводит к разделению
популяции (по данному признаку) на две или несколько групп. Дизруптивный отбор
проявляется в тех случаях, когда популяция занимает гетерогенное местообитание,
и уклоняющиеся формы приспосабливаются к различным экологическим условиям.
Дизруптивный отбор фактически является разновидностью движущего. Движущий и
дизруптивный типы естественного отбора лежат в основе видообразования, процесса
появления новых видов. Пример дизруптивного отбора - выживание рано- и
поздноцветущих форм погремка на покосах.
При стабилизирующей форме
отбора преимущество в размножении имеют особи со средним выражением
признака. Сохранение стандартных форм и элиминация уклонений обуславливают
стабильное существование вида на протяжении длительных исторических периодов
(например, живые ископаемые - латимерия, гаттерия и многие другие виды живых
организмов). Стабилизирующий отбор происходит при постоянных условиях среды,
когда заурядные особи оказываются более приспособленными, чем уклоняющиеся.
Классический пример стабилизирующего отбора - выживание воробьев со средним
размахом крыльев после бури.
Микроэволюция и
макроэволюция.
Процесс эволюции делится на две
части: микроэволюцию и макроэволюцию.
Микроэволюция включает в себя как адаптивные, так и нейтральные генетические
процессы, происходящие в популяциях. Многие мутации, адаптивные в одном, могут
быть вредными в другом (пример - серповидно-клеточная анемия).
Макроэволюция - это процессы видообразования.
Для видообразования прежде
всего необходима генетическая изоляция эволюционирующих популяций в
течение некоторого времени.
На первых этапах разобщения популяций
обычно работают пространственные или временные механизмы
изоляции. К пространственным относятся географические причины: горные хребты,
реки; экологические: кислые или щелочные почвы. Временные механизмы связаны с
разным временем цветения или брачного периода у изолирующихся популяций. Еще
бывают поведенческие механизмы изоляции - разные ритуалы ухаживания.
В дальнейшем вступают в действие биологические
механизмы изоляции: это стерильность или нежизнеспособность гибридов
первого поколения, вызванная либо различными хромосомными перестройками,
либо другими серьезными генетическими процессами, создающими барьер
нескрещиваемости.
Аллопатрическое и
симпатрическое видообразование.
Чаще всего новые виды образуются аллопатрически
- т.е. на разных, изолированных территориях. После такого длительного
разобщения часто возникает биологическая репродуктивная изоляция, и происходит
видообразование. Если же биологической изоляции не происходит, то при
объединении территорий популяции опять скрещиваются, генофонды объединяются,
различия между ними исчезают, и видообразования не происходит.
Симпатрическое
видообразование - гораздо более редкое
явление у животных, но часто встречающееся у растений. Достаточно вспомнить
распространение у растений автополиплоидии и аллопополиплоидии. У
животных изредка тоже происходит симпатрическое видообразование. Известный
пример - черная и серая вороны в Шотландии. На севере живет серая, на юге -
черная. Короткая изоляция между этими популяциями привела к частичному
понижению жизнеспособности гибридов - поэтому зона гибридизации между ними
сейчас достаточно узкая и слабозаселенная. Со временем жизнеспособность
гибридов может еще уменьшиться, и тогда произойдет фактическое видообразование.
Симпатрическое видообразование на
основе возникновения хромосомных перестроек также давно известно у насекомых:
например, Drosophila melanogaster и D. simulans - известные виды-двойники, неспособные скрещиваться, но
практически неотличимые фенотипически.
Процесс видообразования.
Теория прерывистого равновесия.
Палеонтолог Вильямсон очень подробно
изучил ископаемую фауну моллюсков озера Туркана в Кении. На протяжении
последних нескольких десятков млн лет уровень воды в этом озере несколько раз
резко менялся - озеро частично пересыхало и разбивалось на многочисленные
маленькие озерки, а затем его уровень вновь восстанавливался. Всякий раз
понижение уровня озера было сопряжено с резким всплеском изменчивости
морфологии во всех филетических линиях брюхоногих и двустворчатых моллюсков,
происходило значительное возрастание дисперсии по многим признакам, а затем
признаки начинали стабилизироваться на новом уровне. Еще бы немного, и
произошла бы видовая трансформация - образовались бы новые виды. Но быстро
происходила регрессия уровня водоема - при этом новообразовавшиеся формы
вымирали и сохранившиеся старые формы быстро размножались до прежнего уровня
численности. В Палеонтологической истории оз. Туркана такие события происходили
несколько раз.
Сейчас считается, что период стазиса
для каждого вида составляет в среднем 5-10 млн. лет. Период видовой
трансформации, или видообразования, по сравнению с этим временем - краткий
миг: он длится от 5 до 50 тыс. лет.
Литература
1. Горбачев В.В. Концепции современного естествознания: Учеб.
пособие для студентов вузов. - М.: ООО «Издательский дом «ОНИКС 21 век»: ООО
«Издательство «Мир и образование», 2009. - 592 с.
. Грушевицкая Т.Г., Садохин А.П. Концепции современного
естествознания: Учеб. пособие для вузов. - М.: ЮНИТИ-ДАНА, 2009. - 670 с.
. Дубнищева Т.Я. Концепции современного естествознания. - Основной
курс в вопросах и ответах: Учебное пособие. 2-е изд. испр. и доп. -
Новосибирск: Сиб-е ун-е изд-во, 2006. - 592 с.
. Рузавин Г.И. Концепции современного естествознания: Учебник для
вузов. - М.: ЮНИТИ, 2000, 2007. - 287 с.