Морфобиологическая характеристика семейства спаровых в восточной части Чёрного моря
Министерство
образования и науки Российской Федерации
Государственное
бюджетное образовательное учреждение
Высшего
профессионального образования
«КУБАНСКИЙ
ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ»
Кафедра
водных биоресурсов и аквакультуры
ДИПЛОМНАЯ
РАБОТА
Тема:
Морфобиологическая
характеристика семейства спаровых (sparidae) в восточной части Чёрного моря
Краснодар
2013
РЕФЕРАТ
Ключевые слова: СПАРОВЫЕ, МОРСКИЕ КАРАСИ,
МОРФОБИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА, СТРУКТУРА ПОПУЛЯЦИИ.
Дипломная работа посвящена изучению основных
морфо-биологических особенностей видов из семейства спаровых (Sparidae),
выловленных в восточной части Чёрного моря. Для достижения поставленной цели
проводилось изучение: возрастного и полового состава популяции, определение
линейно-массовых характеристик; оценка физиологического состояния и зрелости
половых продуктов; изучение особенностей питания особей.
Содержание
Введение
.
Обзор литературы
.1
Систематическое положение и географическое распространение семейства спаровых
.2
Морфологические признаки спаровых
.3
Размножение и развитие спаровых
.4
Питание спаровых
.5
Хозяйственное значение
.
Материалы и методы исследования
.
Морфо-биологическая характеристика семейства спаровых восточной части Черного
моря
.1
Сравнительная морфологическая характеристика видов
.2
Сравнение морфологических половых особенностей изученных видов семейства
спаровые
.3
Сравнительная морфологическая характеристика самцов и самок
.4
Морфологическая характеристика возрастных групп
.5
Темпы линейного роста
.6
Темпы весового роста
.7
Возрастная структура
.8
Половая структура
.9
Упитанность и ожирения внутринностей
.10
Динамика зрелости половых продуктов
.11
Питание
Заключение
Библиографический
список
Введение
В последние 30-40 лет структура экосистемы
Черного моря претерпела значительные изменения, связанные с человеческой
деятельностью и вселением новых видов морских организмов, таких как моллюск
рапан (Rapana venosa), гребневик мнемиопсис (Mnemiopsis leidyi), голубой краб
(Callinectes sapidus) и другие. Наибольший урон разнообразию жизни в Черном
море нанесли катастрофический перелов промысловых видов рыбы и переудобрение
(эвтрофикация) морских вод в результате стока речных вод, насыщенных солями
азота и фосфора с сельскохозяйственных угодий и неочищенных стоков канализации.
К 1988 году размножившийся мнепиопсис из-за
поедания планктона, фактически лишил основного объекта питания планктоноядных
рыб - хамсы и кильку (шпрота). В свою очередь из-за исчезновения сельдевых
лишились пищи хищные рыбы - скумбрия, ставрида, пеламида, мерланг. Однако в
данных условиях антропогенной деструкции отдельных видов черноморской
ихтиофауны и их массовой гибели, меньше всего пострадали морские караси,
сумевшие не только сохранить популяцию, но даже ее увеличить, за счет вселения
других видов из региона Средиземного моря.
В настоящее время из семейства Спаровых в Черном
море обитают 13 видов морских карасей, из которых 5-6 являются оседлыми видами,
а остальные считаются приходящими на откорм через Босфор из бассейна
Средиземного моря. Примечательно, что побережье от Анапы до Адлера заселено
разными видами карасей крайне неравномерно, что напрямую связано с
температурной средой обитания некоторых рыб семейства Спаровых. Например, у
побережья Аджарии и Абхазии, где средняя температура морской воды на 3-4
градуса выше, чем в районе Анапы, можно наблюдать практически все виды морских
карасей, обитающих в Черном море.
Новизна данной работы заключается в том, что
изученные виды из семейства спаровые являются малоизученными. Это связано с
тем, что они не являются объектами промысла в нашей стране. В других же странах
уже давно эти рыбы, и особенно вид Puntazzo puntazzo, являются объектами
аквакультуры. Для того чтобы это было возможно и в нашей стране необходимо
провести селекционную работу, которая требует изучения морфо-биологических
характеристик рыб.
Цель данной работы изучить видовой состав и
морфо-биологические характеристики рыб семейства спаровые (Sparidae) в
восточной части Чёрного моря.
Для достижения этого поставлены следующее
задачи:
Описать морфологические особенности видов
Diplodus annularis и D. Puntazzo, особенности формирования морфологических
признаков у возрастыных групп;
Дать сравнительную морфологическу характеристику
изученных видов;
Описать биологические особенности развития и
жизнедеятельности видов D. annularis и D. Puntazzo;
Дать сравнительную биологическую характеристику
изученных видов.
1. Обзор литературы
.1 Систематическое положение и
географическое распространение семейства спаровых
Тип Хордовые - Chordata
Подтип Позвоночные - Vertebrata
Надкласс Челюстноротые - Gnathostomata
Группа или ряд Рыбы - Pisces
Класс Костные рыбы - Osteichthyes
Подкласс Лучепёрые рыбы - Actinopterygii
Отряд Окунеобразные - Perciformes
Семейство Спаровые - Sparidae
Род Морские караси - Diplodus
Вид Морской карась, ласкирь - Diplodus annularis
Вид зубарик, хиэна - Diplodus puntazzo
Спаровые распространены в основном (92 вида) в
тропических и субтропических (116 видов) зонах Атлантического, Тихого и
Индийского океанов и лишь 25 видов в умеренных водах. Больше всего морских
карасей обитает в восточной части Атлантического океана, меньше всего - в
Восточной Пацифике. Более половины родов - эндемики той или иной области, а
среди видов лишь 10 встречается в двух областях и один - в трех.
Спаровые - типично морские рыбы.
Среди них нет пресноводных видов, лишь один вид
(Argyrops butcheri) является обитателем солоноватых вод. Живут они на шельфе и
в верхней части континентального склона, до глубины 500-800 м (однако
предпочитают глубину 200-250 м), встречаются на разных грунтах, как мягких,
илистых, так и скалистых. Род Diplodus
включает 14 видов, живущих в субтропической зоне Восточной Атлантики и западной
части Индийского океана.
В Черном море широко распространен ласкирь, или
морской карась (D. annularis).
Рис. 1
Вид Diplodus puntazzo распространен в водах
Атлантического океана у берегов Европы и Африки. Вдоль Европы на север у
берегов Франции до Аркашона. У берегов Африки на юг до Зеленого Мыса и далее у
самой южной границы. Средиземное и прилегающие к нему моря. В Чёрном море у
берегов Кавказа, Крыма и Турции.
1.2 Морфологические признаки
спаровых
Тело довольно высокое и сильно сжато с боков.
Передние зубы направлены несколько вперед, крупные, широкие, в виде резцов с
гладким режущим краем, расположены в один ряд, сзади которого несколько рядов
мелких, закруглённых зубов; боковые зубы жевательные, закруглённые, расположены
в 2-3 ряда. Рот небольшой. Предглазничная кость высокая. Ноздри крупные, задние
в виде щели. Плавательный пузырь без выростов у заднего конца, с каждой стороны
с несколькими небольшими боковыми выростами, прикреплёнными к рёбрам. Тело
умеренно удлинённое, сжатое с боков, спина более или менее горбатая. Спинной
плавник с ХI - ХII, 11 - 15 лучами. Анальный плавник с 11 - 14 разветвлёнными
лучами.

Рис. 2 -
Внешний вид морского карася (D. annularis)
Морской карась или ласкирь (Diplodus annularis).
В спинном плавнике Х - ХI, 11 - 12 лучей; в анальном III, 10 - 11 лучей. 4
пилорических придатка. Рот небольшой конечный. На обеих челюстях в переднем
ряду по 8 резцовых широких зубов с гладким режущим краем, жевательные зубы с
боков челюстей крупные, закруглённые, передние зубы позади резцовых мелкие,
также закруглённые. Тело покрыто большими зазубренными чешуйками, голова также
покрыта чешуёй, межглазничная область не покрыта чешуёй. В боковой линии 50 -
55 чешуй, несколько более мелких непрободенных чешуй на основании лучей
хвостового плавника.
Бока тела у морского карася - светло-желтые, с
серебристым блеском, а спина темнее и с золотистым отливом. Грудные плавники
желтые, а все остальные - светлые. На хвостовом стебле, сразу за спинным
плавником, проходит широкая темная полоса, почти полностью охватывающая кольцом
хвостовой стебель, маленькое темное пятно имеется и у начала грудных плавников.
Молодь карася окрашена более ярко: полоса на хвостовом стебле интенсивно черная
и полностью замкнутая, а на спине проходят пять узких поперечных черных полос с
каждой стороны. Морской карась - некрупная рыба, достигающая в длину не более
33 см, обычно встречаются особи длиной 7-15 см.

Рис.
3
- Внешний вид зубарика или хиэны (Diplodus puntazzo)
Передние зубы длинные и узкие, в виде резцов с
незазубренным режущим краем, направлены вперед, расположены в один ряд. Боковые
зубы очень малы, заостренные, расположены так же в один ряд. На обеих челюстях
спереди по 8 направленных вперед узких, длинных зубов, крайние из них более
короткие, со скошенным назад режущим краем. С каждой стороны на обеих челюстях
около 15 мелких жевательных зубов. Сошник и нёбные кости без зубов. Рот не
большой, рыло удлиненное, заостренное. Благодаря выступающим вперед большим и
крепким резцам, которые окрашены в светло-коричневый цвет, рот вытянут в
трубочку, а верхняя губа выпячена. Предглазничная кость высокая. Жаберная
крышка без шипов, край предкрышки гладкий. Ноздри крупные, задняя в виде щели.
В боковой линии 59 - 62 чешуйки, несколько более мелких чешуек на основании
лучей хвостового плавника.
Имеют семь пилорических придатков. Плавательный
пузырь с двумя короткими, но широкими выростами, без боковых выростов с боков,
прикрепленных к ребрам. Тело умеренно удлиненное, высокое, сжатое с боков,
спина горбатая. Спинной плавник с Х - ХI, 12 - 13 лучами, в анальном III 11-12
лучей. Межглазничная область не покрыта чешуёй. На щеках между предглазничной и
предкрышечной костями 6 - 8 рядов чешуи.
Окраска серовато-серебристая, на боках по 5 - 8
узких продольных тёмных полос, на хвостовом стебле с каждой стороны по большому
чёрному пятну. Молодые особи более светлые, серебристого цвета. Их тело покрыто
серыми поперечными полосами. Зубарь - стайная рыба, ее размеры не превышают
40-50 см. Наиболее часто попадаются экземпляры, средний размер которых 25-30
см. Может достигать веса 3-4 кг. Продолжительность жизни около 10 лет.
1.3 Размножение и развитие спаровых
У спаровых встречаются два типа
онтогенетического нефункционального гермафродитизма: протерогиния, при которой
с увеличением возраста часть самок превращается в самцов, ипротерандрия, когда
с возрастом часть самцов превращается с самок.
У вида Diplodus annularis половая железа
содержит зачатки мужские и женские, но протандрический гермафродитизм если и
имеет место, то лишь у части особей, в то время как у остальных созревает лишь
мужская или женская часть железы. Икрометание происходит в вечернее время, в
прибрежной, более прогретой области, начинается с июня у берегов Крыма, Кавказа
и в южной части Керченского пролива наиболее интенсивно с июля до середины
сентября. Икринки пилагические, развиваются при температуре 21,6 - 25,0°C и
солёности 12,8 - 18,2‰. Неодинаковые размеры икринок в зрелых ястыках и
растянутость периода нереста свидетельствуют, что икрометание происходит не
одновременно, а несколькими порциями. Личинки держаться в поверхностных слоях
воды, преимущественно на глубине 10 - 12 м, единично до 30 м. Мальки ведут
придонный образ жизни, на песчаном и ракушечном грунтах, длина их в июле 9 - 11
мм, в начале августа от 12 до 25 мм.
Предличинки выклевываются слабо развитыми, с
большим, чуть выдающимся за голову желточным мешком и плотно прижатой к нему
головой. Жировая капля находится в задней части желточного мешка. Длина тела
2,0-2,2 мм. Меланофоры в предглазничном и заглазничном отделах головы, на
кишечнике и на желточном мешке в области жировой капли. Серо-желтый пигмент
небольшими пятнами покрывает предглазничный и заглазничный отделы, конец
головы, область ануса и среднюю часть хвостового отдела.
В возрасте трех суток прорезается рот, глаза
интенсивно пигментируются, большие грудные плавники приобретают подвижность. По
мере рассасывания желточного мешка увеличивается оводненная
полость в передней части спинной плавниковой каймы. Четырехсуточные предличинки
достигают в среднем 2,7 мм длины. У них еще сохраняется небольшой остаток
желтка и маленькая жировая капля. Пигментация меняется: желтый пигмент
исчезает, появляется ряд мелких меланофоров вдоль вентральной стороны
хвостового отдела и звездчатые меланофоры на кишечнике.
В возрасте 5 суток желток полностью
рассасывается, сохраняется лишь маленькая жировая капелька. Кишечник петлеобразно
изгибается. Более старшие личинки имеют большую голову, короткое широкое
туловище и длинный хвостовой отдел. Меланофоры - на голове, верхней стороне
кишечника и вдоль вентральной стороны хвостового отдела. При длине 6 мм и более
на жаберной крышке имеются зубчики.
Постэмбриональное развитие морского карася
изображено на рисунке 4.
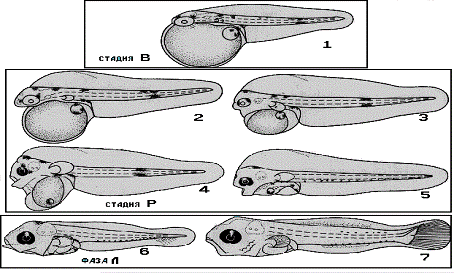
Рис.
4 - Постэмбриональное развитие морского карася
У вида D. puntazzo как и ласкиря половая железа
содержит зачатки мужские и женские, но развивается один из них. В
преднерестовый период образуют небольшие скопления. Нерест в водах Чёрного моря
происходит с августа по сентябрь.
1.4 Питание спаровых
По способу питания встречаются как типичные
растительноядные рыбы, так и планктофаги и хищники.
Представители рода морские караси (Diplodus) в
значительной степени потребляют водоросли и в связи с этим имеют своеобразное
строение зубного аппарата. Передние зубы у них направлены вперед, широкие, в
виде резцов, боковые - жевательные, расположены в 2-З ряда.
Взрослые особи Diplodus annularis питаются
водорослями, преимущественно диатомовыми, гидроидами, губками, полихетами, а
так же гаммаридами и креветками.puntazzo растительноядный вид, хотя и не
исключительно, но водоросли (Ulva, обрастания) в питании играют существенную
роль (Андрияшев, 1946). В желудках встречаются также баланусы, мелкие
ракообразные (изоподы, крабы), моллюски (Nassa, Clavatula и др.).
Рот конечный, он приспособлен к захвату пищевых
объектов, находящихся в различных положениях. Мальки длиной 0,8 - 1,2 см
питаются главным образом прибрежными планктонными организмами, которые
составляют в пищевом комке, в данный период, 84,5%. Переходя на придонное
питание, зубарики всё реже и реже потребляют подвижные организмы.
1.5 Хозяйственное значение
Многие спаровые имеют важное промысловое
значение, являясь объектами большого тралового океанического рыболовства. В
настоящее время во Франции, Италии и Японии ведутся работы по искусственному
разведению четырех видов морских карасей.
Мировая аквакультура не стоит на месте и
ихтиологи-рыбоводы Греции, Японии, Кипра, Израиля, Испании, Хорватии и других
стран постоянно занимаются вопросами внедрения новых объектов в уже
отработанный процесс искусственного выращивания морских видов рыб в садках и
при помощи системы УЗВ. Япония, являющаяся крупнейшим потребителем различного
типов и видов пресноводных и морских гидробионтов, с древних времен занималась
аквакультурой, выращивая на побережье о. Хонсю с середины 17-ого века большого
красного тая (Pagrus major) из рода Pagrus, семейства Спаровые (Sparidae).
Примечательно, что впервые в мире, разведение и выращивание дикого
представителя семейства Спаровых, большого красного тая, по инициативе
Токийского Института Рыбного хозяйства (создан в 1889 году) было начато в
Японии в 1930 году, в префектуре Кагава (остров Сикоку, мыс Мурото, Пацифик).
На данный момент следующие виды рыб из семества
Спаровые являются объектами аквакультуры: дорада (Sparus aurata), большой
красный тай (Pagrus major), обыкновенный пагр (Pagrus pagrus), обыкновенный
зубан (Dentex dentex), атлантический землерой или мормир (Lithognathus
mormyrus), зубарик (Diplodus puntazzo) и др. Следует отметить, что зубарик
широко распространен и в восточной части Чёрного моря наряду с морским карасём
(Diplodus annularis).
Обитающий в Черном море представитель рода
Diplodus вследствие незначительной численности и малых размеров (обычно длина
их менее 30 см) не имеет серьезного промыслового значения и попадается только в
виде прилова. Часто является объектом спортивного рыболовства.
2.
Материалы и методы исследования
Сбор материала проводился в с июня по август
2012 года в районе посёлка Лазаревский. Вылов осуществлялся с помощью ставных
неводов.
Измерение представителей семейства Sparidae
проводились по следующей схеме:
. Пол и стадия зрелости.
2. Вес рыбы, г.
3. Вес рыбы без внутренностей, г.
4. Возраст.
5. Длина рыбы без С.
6. Чешуи в боковой линии.
7. Лучей в D.
8. Лучей в А.
9. Тычинок на первой жаберной дуге.
10. Позвонков.
11. Длина рыла.
12. Диаметр глаза.
13. Заглазничный отдел головы.
14. Длина головы.
15. Высота головы у затылка.
16. Ширина лба.
17. Наибольшая высота тела.
18. Наименьшая высота тела.
19. Антедорсальное расстояние.
20. Постдорсальное расстояние.
21. Длина хвостового стебля.
22. Длина основания D.
23. Наибольшая высота D.
24. Длина основания А.
25. Наибольшая высота А.
26. Длина Р.
27. Длина V.
28. Расстояние между Р и V.
29. Расстояние между V и А.
30. Зубов на верхнечелюстной.
31. Зубов на нижнечелюстной.
Все линейно-массовые характеристики измерялись с
помощью мерной линейки и электронных весов, масса измерялась с точностью до
десятых долей грамма, длина - с точностью до миллиметра.
Степень наполнения пищеварительного тракта
определялась в баллах по схеме Н.В. Лебедева (1936):
баллов - желудок пустой;
балл - единичные пищевые организмы;
балла - малое наполнение;
балла - среднее наполнение;
балла - полный желудок или определённые отделы
кишечника;
баллов - желудок растянут, пища просвечивает
через стенки.
Обработка материала и его анализ осуществлялись
по стандартным методикам.
Для определения половой принадлежности и стадий
зрелости половых продуктов использовалась шкала Никольского(1974).
В ходе работы были определены следующие
показатели:- упитанность, определялась по формуле:
Q = 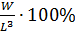
Где W - вес целой рыбы,- общая длина.
Для определения половой принадлежности и стадий
зрелости половых продуктов использовалась шкала Никольского(1974).
Оценка степени ожирения осуществлялась по шкале
М.Л. Прозоровской (1952):
баллов - рыба крайне истощена, жир полностью
отсутствует на петлях кишечника;
балл - тонкая шнуровидная полоска жира между 2 и
3 отделом кишечника. Иногда по верхнему краю второго отдела - узкая прерывистая
полоска жира;
балла - широкая полоска жира между 2 и 3
отделами кишечника. По верхнему краю второго отдела - узкая непрерывная полоска
жира, по нижнему краю 3 отдела - отдельные небольшие участки жира;
балла - широкая полоска жира между 2 и 3
отделами кишечника, в петле между ними полоска расширена. По верху 2 и низу 3
отделов - широкие жировые полосы. Анальный конец кишечника у большинства рыб
залит жиром;
балла - кишечник почти полностью покрыт жиром,
за исключением просветов, где видна кишка;
баллов - кишечник полностью залит жиром без
просветов, на петлях кишечника имеются толстые выросты жира.
Возраст особей определялся по отолитам и чешуе с
помощью лупы по методике Чугуновой(1952).
3. Морфо-биологическая
характеристика семейства спаровых восточной части Черного моря
.1 Сравнительная морфологическая
характеристика видов Diplodus annularis и D. puntazzo
Нами был проведен анализ данных полученный при
изучении двух видов. Нами были рассчитаны средние показатели признаков всех
изученных возрастных групп, а также средние данные не учитывая особей возрастом
5 лет для вида Diplodus annularis, результаты представлены в таблице 1.
Таблица 1 - Средние показатели морфологических
признаков видов D. annularis и D. puntazzo.
|
Diplodus
annularis (с 5∙)
|
Diplodus
annularis (без 5∙)
|
Diplodus
puntazzo
|
Diplodus
annularis (с 5∙)
|
Diplodus
annularis (без 5∙)
|
Diplodus
puntazzo
|
|
Вес
|
50
|
41
|
53
|
-
|
-
|
-
|
|
Вес
без внутренностей
|
41
|
34
|
44
|
-
|
-
|
-
|
|
Длина
рыбы
|
13,8
|
12,8
|
14,0
|
|
|
|
|
|
|
|
В
% общей длины тела
|
|
Длина
без С
|
11,0
|
10,3
|
11,2
|
79,7
|
80,5
|
80,0
|
|
Длина
по Смитту
|
12,3
|
11,7
|
12,4
|
89,1
|
91,4
|
88,6
|
|
Наибольшая
высота тела
|
5,0
|
4,7
|
5,2
|
36,2
|
36,7
|
37,1
|
|
Наименьшая
высота тела
|
1,1
|
1,0
|
1,2
|
8,0
|
7,8
|
8,6
|
|
Антедорсальное
расстояние
|
4,7
|
4,2
|
4,8
|
34,0
|
32,8
|
34,3
|
|
Постдосальное
расстояние
|
1,8
|
1,7
|
1,8
|
13,0
|
13,3
|
12,9
|
|
Длина
хвостового стебля
|
1,6
|
1,4
|
1,6
|
11,6
|
10,9
|
11,4
|
|
Длина
основания D
|
5,8
|
5,5
|
5,8
|
42,0
|
43,0
|
41,4
|
|
Наибольшая
высота D
|
1,5
|
1,4
|
1,3
|
10,9
|
10,9
|
9,3
|
|
Длина
основания А
|
2,4
|
2,2
|
2,4
|
17,4
|
17,2
|
17,1
|
|
Наибольшая
высота А
|
1,2
|
1,1
|
0,9
|
8,7
|
8,6
|
6,4
|
|
Длина
Р
|
3,3
|
3,0
|
3,2
|
23,9
|
23,4
|
22,9
|
|
Длина
V
|
2,1
|
2,0
|
2,0
|
15,2
|
15,6
|
14,3
|
|
Расстояние
между Р и V
|
1,2
|
1,1
|
1,1
|
8,7
|
8,6
|
7,9
|
|
Расстояние
между V и A
|
3,4
|
3,2
|
3,3
|
24,6
|
25,0
|
23,6
|
|
Длина
головы
|
3,2
|
2,9
|
3,3
|
23,2
|
22,7
|
23,6
|
|
|
|
|
В
% общей длины головы
|
|
Длина
рыла
|
1,1
|
1,0
|
1,3
|
37,9
|
34,5
|
39,4
|
|
d
глаза
|
0,9
|
0,8
|
0,8
|
28,1
|
27,6
|
24,2
|
|
Заглазничный
отдел головы
|
1,2
|
1,1
|
1,2
|
37,5
|
37,9
|
36,4
|
|
Высота
головы у затылка
|
3,1
|
2,8
|
3,1
|
96,9
|
96,5
|
93,3
|
|
Ширина
лба
|
1,0
|
1,0
|
1,0
|
31,2
|
31,4
|
30,3
|
|
Меристические
признаки
|
|
Зубов
на верхней челюсти
|
8
резцов
|
8
резцов
|
8
резцов
|
-
|
-
|
-
|
|
Зубов
на нижней челюсти
|
8
резцов
|
8
резцов
|
8
резцов
|
-
|
-
|
-
|
|
Чешуй
в боковой линии
|
52
|
52
|
62
|
-
|
-
- -
|
-
-
|
ХI
12
|
ХI
12
|
ХI13
|
-
|
-
|
-
|
|
Лучей
в А
|
III
12
|
III
12
|
III12
|
-
|
-
|
-
|
|
Количество
позвонков
|
23
|
23
|
20
|
-
|
-
|
-
|
|
|
|
|
|
|
|
|
При сравнении морфологического строения изученных
видов нами были выявлены следующие закономерности:
. Средние весовые значения совпадают у
представителей вида Diplodus annularis с учётом особей 5 лет и у представителей
вида Diplodus puntazzo, что подтверждает тугорослость вида Diplodus annularis.
. Похожая закономерность наблюдается и при
сравнении других признаков, таких как длина тела, высота тела, антедорсальное и
постдорсальное расстояние и др.
. Найдены отличия в строении спинных, анальных
плавников. Так наибольшая высота спинного плавника у ласкиря составляет (в % от
длины тела) 10,9 %, а у хиены 9,3%. Так же необходимо заметить, что у
представителей вида Diplodus annularis средние параметры данного признака у
четырёх возрастных групп так же равны 10,9% от длины тела. Это говорит о том,
что спинной плавник у данного вида формируется к 4 годам и затем рост его
значительно замедляется.
Наибольшая высота анального плавника так же
различается у изученных видов. Если у ласкиря его длина составляет 8,7 % от
длины тела, то у зубарика всего 6,4%.
Все вышеперечисленные отличия не значительны и
визуально не определимы.
. При изучении строения головы нами выявлены
более значительные отличия, связанные со строением челюстей у данных видов.
Различия в форме зубов и их расположении показаны в приложении А.
При близких средних показателях длины головы 3,2
см и 3,3 см, длина рыла составляет 37,9% (у ласкиря) и 39,4% (у зубарика) от
длины головы. Это связано со строением резцов, которых насчитывается по 8 на
нижней и верхней челюстях. Следует отметить, что при этом диаметр глаза у
зубарика в среднем 0,8 см (24,2% от длина головы) против 0,9 см (28,1 % от
длины головы) у ласкиря. Тогда как длина заглазничного отдела и другие
показатели совпадают.
При изучении меристических признаков данных
видов выявлены следующие отличия:
Как было указано выше у изученных видов
одинаковое количество резцов как на верхней, так и на нижней челюстях - 8. Но
количество жевательных зубов не одинаково. Так у представителей вида D.
annularis на челюстях располагаются … зубов, а у вида D.puntazzo… зубов. Чешуй
в боковой линии также разнится, так у ласкиря их насчитывается в среднем 52,
тогда как у зубарика 62.
3.2 Сравнение морфологических
половых особенностей изученных видов семейства спаровые
Нами проведен сравнительный анализ морфологических
признаков самок и самцов видов Diplodus annularis и D. puntazzo. Сравниваемые
средние показатели представлены в таблице 2 (для вида D. annularis) и в таблице
3 (для вида D.puntazzo).
Таблица 2 - Сравнительная морфологическая
характеристика самок и самцов вида D. annularis
|
F
Diplodus annularis
|
M
Diplodus annularis
|
F
Diplodus annularis
|
M
Diplodus annularis
|
|
Вес
|
68
|
49
|
-
|
-
|
|
Вес
без внутренностей
|
57
|
41
|
-
|
-
|
|
Длина
рыбы
|
15,9
|
13,8
|
|
|
|
|
|
В
% общей длины тела
|
|
Длина
без С
|
12,7
|
11,1
|
79,9
|
80,4
|
|
Длина
по Смитту
|
14,2
|
12,5
|
89,3
|
90,6
|
|
Наибольшая
высота тела
|
5,9
|
5,1
|
37,1
|
37,0
|
|
Наименьшая
высота тела
|
1,3
|
1,1
|
8,2
|
8,0
|
|
Антедорсальное
расстояние
|
5,1
|
4,5
|
32,1
|
32,6
|
|
Постдосальное
расстояние
|
2,0
|
1,8
|
12,6
|
13,0
|
|
Длина
хвостового стебля
|
1,8
|
1,6
|
11,3
|
11,6
|
|
Длина
основания D
|
6,8
|
5,9
|
42,8
|
42,8
|
|
Наибольшая
высота D
|
1,7
|
1,5
|
10,7
|
10,9
|
|
Длина
основания А
|
2,8
|
2,4
|
17,6
|
17,4
|
|
Наибольшая
высота А
|
1,3
|
1,2
|
8,2
|
8,7
|
|
Длина
Р
|
3,8
|
3,3
|
23,9
|
23,9
|
|
Длина
V
|
2,3
|
2,1
|
14,5
|
15,2
|
|
Расстояние
между Р и V
|
1,4
|
1,2
|
8,8
|
8,7
|
|
Расстояние
между V и A
|
4,0
|
3,4
|
25,2
|
24,6
|
|
Длина
головы
|
3,8
|
3,3
|
23,9
|
24,0
|
|
|
|
В
% общей длины головы
|
|
Длина
рыла
|
1,4
|
1,2
|
36,8
|
36,4
|
|
d
глаза
|
1,0
|
0,9
|
26,4
|
27,2
|
|
Заглазничный
отдел головы
|
1,4
|
1,2
|
36,8
|
36,4
|
|
Высота
головы у затылка
|
3,7
|
3,2
|
97,4
|
97,0
|
|
Ширина
лба
|
1,2
|
1,1
|
31,6
|
33,3
|
При сравнении средних морфологических
показателей самок самцов вида D. annularis нами замечены следуещие
закономерности:
. Так как самки преобладают в старших возрастных
группах, а в младших отсутствуют значения их веса и длин значительно больше чем
у самцов. Так средний вес самок в выборке равен 68г, а самцов 49г, средняя
длина тела самок - 15,9 см, самцов - 13,8 см.
. Если рассматривать все показатели в процентах
от длины тела, то видно, что существенных отличий нет. Имеется незначительное
расхождение в длине плавников - плавники самцов немного длиннее, за исключением
грудных.
Таким образом, значительных морфологических
отличий у самцов и самок вида не отмечено, что говорит о том, что для вида
Diplodus annularis не характерен половой диморфизм.
Таблица 3 - Сравнительная морфологическая
характеристика самок и самцов вида Diplodus puntazzo
|
F
Diplodus puntazzo
|
M
Diplodus puntazzo
|
F
Diplodus puntazzo
|
M
Diplodus puntazzo
|
|
Вес
|
80
|
61
|
-
|
-
|
|
Вес
без внутренностей
|
66
|
50
|
-
|
-
|
|
Длина
рыбы
|
16,5
|
14,8
|
|
|
|
|
|
В
% общей длины тела
|
|
Длина
без С
|
13,4
|
11,8
|
81,2
|
79,7
|
|
Длина
по Смитту
|
14,7
|
13,1
|
89,1
|
88,5
|
|
Наибольшая
высота тела
|
6,0
|
5,5
|
36,4
|
37,1
|
|
Наименьшая
высота тела
|
1,4
|
1,2
|
8,5
|
8,1
|
|
Антедорсальное
расстояние
|
5,6
|
5,0
|
33,9
|
33,8
|
|
Постдосальное
расстояние
|
2,2
|
1,9
|
13,3
|
12,8
|
|
Длина
хвостового стебля
|
2,0
|
1,7
|
12,1
|
11,5
|
|
Длина
основания D
|
7,1
|
6,2
|
43,0
|
41,9
|
|
Наибольшая
высота D
|
1,4
|
1,4
|
8,5
|
9,9
|
|
Длина
основания А
|
2,9
|
2,6
|
17,6
|
17,6
|
|
Наибольшая
высота А
|
1,0
|
1,0
|
6,1
|
6,8
|
|
Длина
Р
|
3,8
|
3,4
|
23,0
|
23,0
|
|
Длина
V
|
2,3
|
2,1
|
13,9
|
14,2
|
|
Расстояние
между Р и V
|
1,3
|
1,2
|
7,9
|
8,1
|
|
Расстояние
между V и A
|
3,9
|
3,5
|
23,6
|
23,6
|
|
Длина
головы
|
3,7
|
3,3
|
22,4
|
22,3
|
|
|
|
В
% общей длины головы
|
|
Длина
рыла
|
1,5
|
1,3
|
40,5
|
39,4
|
|
d
глаза
|
0,9
|
0,8
|
24,3
|
24,2
|
|
Заглазничный
отдел головы
|
1,3
|
1,2
|
35,2
|
36,4
|
|
Высота
головы у затылка
|
3,6
|
3,2
|
97,3
|
97,0
|
|
Ширина
лба
|
1,2
|
1,1
|
32,4
|
33,3
|
Аналогично предыдущему виду средние показатели
самок вида Diplodus puntazzo значительно больше чем у самцов. Так средняя масса
самок в выборке - 80г, самцов - 61г; средняя длина самок - 16,5см, тогда как у
самцов - 14,8 см. В связи с этим, нами рассмотрено процентное соотношение
параметров к длине тела, а так же длине головы и сделаны следующие выводы:
. Все морфологические показатели имеют сходные
значения, различия небольшие и не превышают 2%.
. Небольшие отличия касаются длины плавников,
особенно это касается анального плавника, если у самок он составляет 6,1% от
длины тела, то у самцов 6,8%.
Как и в случае с видом Diplodus annularis, нами
сделан вывод, что для вида Diplodus puntazzo не свойственен половой диморфизм.
3.3 Сравнительная морфологическая
характеристика самцов и самок
Анализируя полученные данные, нами сделаны
выводы, что половой диморфизм данным видам не присущ. Меристические признаки
имеют одинаковые значения вне зависимости от пола, поэтому мы рассмотрим их
отдельно, а пластические сравним в таблице 4 (для самок) и в таблице 5 (для
самцов) видов Diplodus annularis и Diplodus puntazzo.
семейство спаровые размножение
питание
Таблица 4 - Морфологическая характеристика самок
|
Diplodus
annularis
|
Diplodus
puntazzo
|
Diplodus
annularis
|
Diplodus
puntazzo
|
|
Вес
|
68
|
80
|
-
|
-
|
|
Вес
без внутренностей
|
57
|
66
|
-
|
-
|
|
Длина
рыбы
|
15,9
|
16,5
|
|
|
|
|
|
В
% общей длины тела
|
|
Длина
без С
|
12,7
|
13,4
|
79,9
|
81,2
|
|
Длина
по Смитту
|
14,2
|
14,7
|
89,3
|
89,1
|
|
Наибольшая
высота тела
|
5,9
|
6,0
|
37,1
|
36,4
|
|
Наименьшая
высота тела
|
1,3
|
1,4
|
8,2
|
8,5
|
|
Антедорсальное
расстояние
|
5,1
|
5,6
|
32,1
|
33,9
|
|
Постдосальное
расстояние
|
2,0
|
2,2
|
12,6
|
13,3
|
|
Длина
хвостового стебля
|
1,8
|
2,0
|
11,3
|
12,1
|
|
Длина
основания D
|
6,8
|
7,1
|
43,0
|
|
Наибольшая
высота D
|
1,7
|
1,4
|
10,7
|
8,5
|
|
Длина
основания А
|
2,8
|
2,9
|
17,6
|
17,6
|
|
Наибольшая
высота А
|
1,3
|
1,0
|
8,2
|
6,1
|
|
Длина
Р
|
3,8
|
3,8
|
23,9
|
23,0
|
|
Длина
V
|
2,3
|
2,3
|
14,5
|
13,9
|
|
Расстояние
между Р и V
|
1,4
|
1,3
|
8,8
|
7,9
|
|
Расстояние
между V и A
|
4,0
|
3,9
|
25,2
|
23,6
|
|
Длина
головы
|
3,8
|
3,7
|
23,9
|
22,4
|
|
|
|
В
% общей длины головы
|
|
Длина
рыла
|
1,4
|
1,5
|
36,8
|
40,5
|
|
d
глаза
|
1,0
|
0,9
|
26,4
|
24,3
|
|
Заглазничный
отдел головы
|
1,4
|
1,3
|
36,8
|
35,2
|
|
Высота
головы у затылка
|
3,7
|
3,6
|
97,4
|
97,3
|
|
Ширина
лба
|
1,2
|
1,2
|
31,6
|
32,4
|
|
Меристические
признаки
|
|
Зубов
на нижней челюсти
|
8
резцов
|
8
резцов
|
-
|
-
|
|
Зубов
на верхней челюсти
|
8
резцов
|
8
резцов
|
-
|
-
|
|
Чешуй
в боковой линии
|
52
|
62
|
-
|
-
|
|
Лучей
в D
|
ХI
12
|
ХI
13
|
-
|
-
|
|
Лучей
в А
|
III
12
|
III
12
|
-
|
-
|
|
Количество
позвонков
|
23
|
20
|
-
|
-
|
|
|
|
|
|
|
|
Сравнивая средние морфологические показатели
самок изученных видов, нами были сделаны следующие выводы:
. Размеры самок вида Diplodus puntazzo превышают
размеры самок вида Diplodus annularis, так масса самок зубарика - 80г, при
длине тела 16,5см, тогда как средняя масса самок ласкиря равна 68г, при средней
длине тела 15,9см.
. При сравнении показателей в процентах от длины
тела было замечено, что тело самок ласкиря выше, то есть наибольшая высота тела
составляет 37,1% против 36,4% у самок зубарика. Учитывая, при этом
антедорсальное и постдорсальное расстояния, как и длину тела без хвостового
плавника, тело самок ласкиря уступает соотношению этих параметров у самок
зубарика. Таким образом, тело зубарика более прогонистое.
. В строении головы и плавников замечаются те же
тенденции, что и рассмотренные ранее при общем сравнении видов.
Таблица 5 - Морфологическая характеристика
самцов
|
Самцы
Diplodus annularis
|
Самцы
Diplodus
puntazzo
|
Diplodus
annularis
|
Diplodus
puntazzo
|
|
Вес
|
49
|
61
|
-
|
-
|
|
Вес
без внутренностей
|
41
|
50
|
-
|
-
|
|
Длина
рыбы
|
13,8
|
14,8
|
|
|
|
|
|
В
% общей длины тела
|
|
Длина
без С
|
11,1
|
11,8
|
80,4
|
79,7
|
|
Длина
по Смитту
|
12,5
|
13,1
|
90,6
|
88,5
|
|
Наибольшая
высота тела
|
5,1
|
5,5
|
37,0
|
37,1
|
|
Наименьшая
высота тела
|
1,1
|
1,2
|
8,0
|
8,1
|
|
Антедорсальное
расстояние
|
4,5
|
5,0
|
32,6
|
33,8
|
|
Постдосальное
расстояние
|
1,8
|
1,9
|
13,0
|
12,8
|
|
Длина
хвостового стебля
|
1,6
|
1,7
|
11,6
|
11,5
|
|
Длина
основания D
|
5,9
|
6,2
|
42,8
|
41,9
|
|
Наибольшая
высота D
|
1,5
|
1,4
|
10,9
|
9,9
|
|
Длина
основания А
|
2,4
|
2,6
|
17,4
|
17,6
|
|
Наибольшая
высота А
|
1,2
|
1,0
|
8,7
|
6,8
|
|
Длина
Р
|
3,3
|
3,4
|
23,9
|
23,0
|
|
Длина
V
|
2,1
|
2,1
|
15,2
|
14,2
|
|
Расстояние
между Р и V
|
1,2
|
1,2
|
8,7
|
8,1
|
|
Расстояние
между V и A
|
3,4
|
3,5
|
24,6
|
23,6
|
|
Длина
головы
|
3,3
|
3,3
|
24,0
|
22,3
|
|
|
|
В
% общей длины головы
|
|
Длина
рыла
|
1,2
|
1,3
|
36,4
|
39,4
|
|
d
глаза
|
0,9
|
0,8
|
27,2
|
24,2
|
|
Заглазничный
отдел головы
|
1,2
|
1,2
|
36,4
|
36,4
|
|
Высота
головы у затылка
|
3,2
|
3,2
|
97,0
|
97,0
|
|
Ширина
лба
|
1,1
|
1,1
|
33,3
|
33,3
|
|
Меристические
признаки
|
|
Зубов
на нижней челюсти
|
8
резцов
|
8
резцов
|
-
|
-
|
|
Зубов
на верхней челюсти
|
8
резцов
|
8
резцов
|
-
|
-
|
|
Чешуй
в боковой линии
|
52
|
62
|
-
|
-
|
|
Лучей
в D
|
ХI
12
|
ХI
13
|
-
|
-
|
|
Лучей
в А
|
III
12
|
III
12
|
-
|
-
|
|
Количество
позвонков
|
23
|
20
|
-
|
-
|
При сравнении морфологических особенностей
самцов представленных видов нами были отмечены следующие различия:
. Так же как и самки, самцы вида D. puntazzo в
среднем крупнее (средняя масса - 61г при длине - 14,8см), чем самцы вида D.
annularis (средняя масса - 49г, при длине тела - 13,8см).
. В отличии от самок пропорции тела самцов
совпадают, так наибольшая высота тела обоих видов составляет 37% от общей длины
тела.
. У самцов ласкиря лучи плавников длинее чем у
самцов зубарика, что отмечено и у самок.
. В строении головы наблюдаются общие
закономерности: длина рыла больше у самцов вида D. puntazzo и составляет 39,4%
от длины головы, а диаметр глаза больше у вида D. annularis, другие показатели
совпадают.
3.4 Сравнительная морфологическая
характеристика возрастных групп видов Diplodus annularis и D. puntazzo
Нами были определены средние показатели
морфологических признаков для возрастных групп видов D. annularis (таблица 6) и
D. puntazzo (таблица 7).
Таблица 6 - Морфологическая характеристика
возрастных групп видов D. annularis
|
1∙
|
2∙
|
3∙
|
4∙
|
5∙
|
1∙
|
2∙
|
3∙
|
4∙
|
5∙
|
|
Вес
|
16
|
31
|
51
|
65
|
86
|
|
|
|
|
|
|
Вес
без внутренностей
|
14
|
25
|
42
|
53
|
71
|
|
|
|
|
|
|
Длина
рыбы
|
9,7
|
11,8
|
13,9
|
15,7
|
17,8
|
|
|
|
|
|
|
|
|
|
|
|
В
% общей длины тела
|
|
Длина
без С
|
7,7
|
9,6
|
11,3
|
12,4
|
14,1
|
79,4
|
81,4
|
81,9
|
79,0
|
80,1
|
|
Длина
по Смитту
|
8,9
|
11,0
|
12,6
|
13,8
|
15,7
|
91,7
|
93,2
|
91,3
|
87,9
|
89,2
|
|
Наибольшая
высота тела
|
3,3
|
4,4
|
5,3
|
5,9
|
6,1
|
34,0
|
37,3
|
38,4
|
37,5
|
34,6
|
|
Наименьшая
высота тела
|
0,8
|
1,0
|
1,2
|
1,2
|
1,4
|
8,2
|
8,4
|
8,7
|
7,6
|
7,9
|
|
Антедорсальное
расстояние
|
3,2
|
3,9
|
4,6
|
5,0
|
5,6
|
32,9
|
33,0
|
33,3
|
31,8
|
31,8
|
1,4
|
1,6
|
1,8
|
1,9
|
2,3
|
14,4
|
13,5
|
13,0
|
12,1
|
13,0
|
|
Длина
хвостового стебля
|
1,1
|
1,3
|
1,5
|
1,8
|
2,0
|
11,3
|
11,0
|
10,9
|
11,4
|
11,4
|
|
Длина
основания D
|
4,1
|
5,1
|
6,1
|
6,6
|
7,3
|
42,3
|
43,2
|
44,2
|
42,0
|
41,4
|
|
Наибольшая
высота D
|
1,2
|
1,3
|
1,5
|
1,6
|
1,8
|
12,4
|
11,0
|
10,8
|
10,2
|
10,2
|
|
Длина
основания А
|
1,7
|
2,1
|
2,5
|
2,7
|
2,9
|
17,5
|
17,8
|
18,1
|
17,2
|
16,5
|
|
Наибольшая
высота А
|
1,0
|
1,1
|
1,2
|
1,2
|
1,4
|
10,3
|
9,3
|
8,7
|
7,9
|
7,6
|
|
Длина
Р
|
2,3
|
2,9
|
3,4
|
3,6
|
4,2
|
23,7
|
24,5
|
24,6
|
22,9
|
23,8
|
|
Длина
V
|
1,6
|
1,9
|
2,2
|
2,3
|
2,5
|
16,5
|
16,1
|
15,9
|
14,6
|
14,2
|
|
Расстояние
между Р и V
|
0,7
|
1,0
|
1,2
|
1,4
|
1,5
|
7,2
|
8,5
|
8,7
|
8,9
|
8,5
|
|
Расстояние
между V и A
|
2,3
|
2,9
|
3,5
|
4,0
|
4,2
|
23,7
|
24,6
|
25,4
|
25,5
|
23,9
|
|
Длина
головы
|
2,3
|
2,8
|
3,2
|
3,6
|
4,3
|
23,7
|
23,7
|
23,2
|
22,9
|
24,4
|
|
|
|
|
|
|
В
% общей длины головы
|
|
Длина
рыла
|
0,7
|
0,9
|
1,1
|
1,3
|
1,7
|
30,4
|
32,1
|
34,4
|
36,1
|
39,5
|
|
d
глаза
|
0,7
|
0,8
|
0,9
|
1,0
|
1,1
|
30,4
|
28,6
|
28,1
|
27,8
|
25,6
|
|
Заглазничный
отдел головы
|
0,9
|
1,1
|
1,2
|
1,3
|
1,5
|
39,1
|
39,3
|
37,5
|
36,1
|
34,9
|
|
Высота
головы у затылка
|
2,2
|
2,7
|
3,1
|
3,5
|
3,9
|
95,7
|
96,4
|
96,9
|
97,2
|
90,7
|
|
Ширина
лба
|
0,7
|
0,8
|
1,0
|
1,2
|
1,4
|
30,4
|
28,6
|
31,2
|
33,3
|
32,6
|
Рассмотренные нами средние морфологические
показатели пяти возрастных групп вида D. annularis имели следующие особенности
и различия:
. средние весовые показатели изученных
возрастных групп имели тенденцию к плавному увеличению, так у годовиков средняя
масса была равна 16г, а у особей в возрасте 5 лет - 86г. Аналогично
увеличивалась и средняя длина тела.
. При изучении морфологических параметров в
процентном соотношении от общей длины тела нами отмечены следующие
закономерности развития особей:
Наиболее активно особи данного вида растут в
длину в первые три года жизни, далее рост замедляется. Максимальное процентное
соотношение хвостового плавника к длине тела наблюдается в первый и четвертый
год жизни (20,6% и 21%). Это может быть связано как с развитием самого
хвостового плавника, так и с общим ростом рыб.
Основной рост в высоту у данного вида происходит
в первые три года жизни и затем уступает росту рыб в длину. Так высота тела в 3
года является максимальной и составляет 38,4% от длины тела, а наименьшая
высота тела в три года составляет 8,7%, тогда как наименьшей она является в
четыре года и равняется 7,6% от длины тела. Та же закономерность наблюдается
при рассмотрении развития и роста спинного и анального плавников. При этом лучи
в плавниках развиваются в первый год. Так у годовиков наибольшая высота
спинного плавника составляет 12,4% от длины тела, у особей возрастом 2 года -
11,0%, 3 года - 10,8%, в возрасте 4 года и 5 лет значения совпадаю и равны
10,2%. Такая же закономерность и в развитии лучей в анальном плавнике, у
годовиков наибольшая высота лучей в анальном плавнике - 10,3%, у особей в
возрасте 2 года - 9,3%, 3 года - 8,7%, 4 года - 7,9% и 5 лет - 7,6%. Так же
развиваются грудные и брюшные плавники.
При развитии головы длина рыла изменяется так: в
первый год составляет 30,4% от длины головы, в 2 года - 32,1%, в 3 года -
34,4%, в 4 года - 36,1% и в 5 лет достигает 39,5% от длины головы. Глаза,
напротив, формируются на первый год жизни, и их максимальное значение достигает
30,4% от длины головы и затем скорость роста уменьшается по сравнению с ростом
головы и к 5 годам составляет 25,6%.
Таблица 7 - морфологическая характеристика
возрастных групп видов D. puntazzo
|
1∙
|
2∙
|
3∙
|
4∙
|
1∙
|
2∙
|
3∙
|
4∙
|
|
Вес
|
18
|
35
|
65
|
95
|
|
|
|
|
|
Вес
без внутренностей
|
14
|
27
|
53
|
80
|
|
|
|
|
|
Длина
рыбы
|
10,2
|
12,3
|
14,9
|
18,4
|
|
|
|
|
|
|
|
|
|
В
% общей длины тела
|
|
Длина
без С
|
8,1
|
9,7
|
12,2
|
14,7
|
79,4
|
78,9
|
81,9
|
79,8
|
|
Длина
по Смитту
|
9,2
|
11,0
|
13,2
|
16,3
|
90,1
|
89,4
|
88,5
|
88,5
|
|
Наибольшая
высота тела
|
3,8
|
4,8
|
5,4
|
6,7
|
37,3
|
39,0
|
36,2
|
36,4
|
|
Наименьшая
высота тела
|
0,9
|
1,1
|
1,2
|
1,6
|
8,8
|
8,9
|
8,1
|
8,7
|
|
Антедорсальное
расстояние
|
3,6
|
4,2
|
4,9
|
6,4
|
35,3
|
34,1
|
32,9
|
34,8
|
|
Постдосальное
расстояние
|
1,2
|
1,6
|
1,8
|
2,5
|
11,8
|
13,0
|
12,1
|
13,6
|
|
Длина
хвостового стебля
|
1,1
|
1,5
|
1,7
|
2,3
|
10,8
|
12,2
|
11,4
|
12,5
|
|
Длина
основания D
|
3,8
|
4,9
|
6,2
|
8,1
|
37,2
|
39,8
|
41,6
|
44,0
|
|
Наибольшая
высота D
|
1,0
|
1,2
|
1,4
|
1,5
|
9,8
|
9,7
|
9,4
|
8,2
|
|
Длина
основания А
|
1,7
|
2,2
|
2,5
|
3,4
|
16,7
|
17,9
|
16,8
|
18,5
|
|
Наибольшая
высота А
|
0,7
|
0,8
|
1,0
|
1,1
|
6,8
|
6,5
|
6,7
|
6,0
|
|
Длина
Р
|
2,2
|
2,7
|
3,3
|
4,4
|
21,6
|
22,0
|
22,1
|
23,9
|
|
Длина
V
|
1,4
|
1,8
|
2,1
|
2,5
|
13,7
|
14,6
|
14,1
|
13,6
|
|
Расстояние
между Р и V
|
0,8
|
1,1
|
1,2
|
1,5
|
7,8
|
8,9
|
8,1
|
8,2
|
|
Расстояние
между V и A
|
2,4
|
2,9
|
3,6
|
4,3
|
23,5
|
23,6
|
24,2
|
23,4
|
|
Длина
головы
|
2,7
|
2,9
|
3,3
|
4,2
|
26,5
|
23,6
|
22,1
|
|
|
|
|
|
В
% общей длины головы
|
|
Длина
рыла
|
1,0
|
1,1
|
1,3
|
1,8
|
37,0
|
33,3
|
39,4
|
42,8
|
|
d
глаза
|
0,7
|
0,7
|
0,8
|
1,0
|
25,9
|
24,2
|
24,1
|
23,8
|
|
Заглазничный
отдел головы
|
1,0
|
1,1
|
1,2
|
1,4
|
37,0
|
37,9
|
36,4
|
33,3
|
|
Высота
головы у затылка
|
2,6
|
2,8
|
3,2
|
4,1
|
96,3
|
96,5
|
97,0
|
97,6
|
|
Ширина
лба
|
0,7
|
0,9
|
1,1
|
1,4
|
26,0
|
31,0
|
33,3
|
33,3
|
Рассмотренные нами средние морфологические
показатели четырёх возрастных групп вида D. puntazzo имели следующие
особенности и различия:
. средние весовые показатели изученных
возрастных групп имели тенденцию к плавному увеличению, так у годовиков средняя
масса была равна 18г, а у особей в возрасте 4 года - 95г. Аналогично
увеличивалась и средняя длина тела у годовиков в среднем она равнялась 10,2 см,
а у особей в возрасте 4 года - 18,4 см.
. При изучении морфологических параметров, в
процентном соотношении от общей длины тела, нами отмечены следующие
закономерности развития особей:
Наиболее активно особи данного вида растут в
высоту первые два года, если в первый год наибольшая высота тела достигает
37,3% от длины тела, то ко второму году этот параметр достигает 39,0%. Далее
рост замедляется. То же наблюдается и с наименьшей высотой тела. У годовиков
она равна 8,8%, у особей возрастом 2 года - 8,9, в 3года - 8,1 и в возрасте 4
лет увеличивается до 8,7%.
Развитие плавников проходит постепенно, так
длина основания спинного плавника у годовиков равна 37,2%, а у особей 4 лет
достигает 44,0%. Так же развивается и анальный, грудные и брюшные плавники.
В процентном соотношении длина головы у
годовиков составляет 26,5% от длины тела, что является максимальным значением.
Далее это значение уменьшается. Длина рыла у данного вида изменяется аналогично
предыдущему, как и диаметр глаза.
3.5 Темпы линейного роста
Размеры исследованных особей морского карася или
ласкиря варьировали от 8,9 см (годовики) до 18, 8 см. Средний размер годовиков
составил 9,7 см
Таблица 8
- Темпы линейного роста ласкиря (Diplodus annularis)
|
Возраст
|
L,
ср
|
L,
min - max
|
Прирост,
см
|
%
|
N,шт
|
|
1∙
|
9,7
|
8,9
- 10,4
|
-
|
-
|
20
|
|
2∙
|
11,8
|
11,0
- 12,8
|
2,1
|
18
|
20
|
|
3∙
|
13,9
|
13,1
- 14,9
|
2,0
|
14
|
20
|
|
4∙
|
15,7
|
15,3
- 16,7
|
1,6
|
10
|
20
|
|
5∙
|
17,8
|
17,1
- 18,8
|
2,1
|
12
|
20
|
В отличие от ласкиря зубарик менее тугорослый,
что позволяет использовать как объект аквакультуры. Размеры в выборке
варьировали от 9,1 см (годовики) до 18,9 см. Средний размер годовиков составил
10,2 см.
Таблица 9
- Темпы линейного роста зубарика (Diplodus puntazzo)
|
Возраст
|
L,
ср
|
L,
min - max
|
Прирост,
см
|
%
|
N,шт
|
|
1∙
|
10,2
|
9,1
- 11,2
|
-
|
-
|
20
|
|
2∙
|
12,3
|
11,2
- 13,1
|
2,1
|
16
|
20
|
|
3∙
|
14,9
|
14,1
- 16,2
|
2,6
|
15
|
20
|
|
4∙
|
18,4
|
17,7
- 18,9
|
3,5
|
21
|
20
|
3.6 Темпы весового роста
В изученной выборке ласкиря нами выделены
следующие значеня: минимальная масса - 13 г, максимальная масса - 95г.
Средний вес рыб, а так же максимальные и
минимальные значения для каждой возрастной группы указанны в таблице 10.
Таблица 10
- Показатели массы (г) морского карася
|
Возраст
|
M,
ср
|
M,
min - max
|
Прирост,
грамм
|
%
|
N,
шт.
|
|
1∙
|
16
|
13
- 19
|
-
|
-
|
20
|
|
2∙
|
31
|
25
- 39
|
15
|
48
|
20
|
|
3∙
|
51
|
43
- 59
|
20
|
39
|
20
|
|
4∙
|
65
|
63
- 71
|
14
|
21
|
20
|
|
5∙
|
86
|
83
- 95
|
21
|
24
|
20
|
У изученных особей в выборке зубарика были
выделены следующие значения: минимальная масса - 12г, максимальная масса -
100г. Приросты массы плавно увеличивались от годовиков к старшим возрастным
группам. Средний вес рыб, а так же максимальные и минимальные значения для
каждой возрастной группы указанны в таблице 11.
Таблица 11
- Показатели массы (г) зубарика
|
Возраст
|
M,
ср
|
M,
min - max
|
Прирост,
грамм
|
%
|
N,
шт.
|
|
1∙
|
18
|
12
- 24
|
-
|
-
|
20
|
|
2∙
|
35
|
29
- 43
|
17
|
|
20
|
|
3∙
|
65
|
55
- 73
|
30
|
|
20
|
|
4∙
|
95
|
89
- 100
|
32
|
|
20
|
3.7 Возрастная структура видов из
семейства спаровых восточной части Чёрного моря
Был проведен биологический анализ выборки 100
особей вида Diplodus annularis. По результатам исследований особей данного вида
было выделено пять возрастных групп. Каждая группа в выборке насчитывала 20
особей.
В уловах же преобладали особи возрастом два года
(41%), немного меньше было особей трёхгодовалого возраста (31%). Наименьшее
количество было особей возраста 5 лет.
Возрастная структура особей морского карася в
уловах показана на рисунке 5.
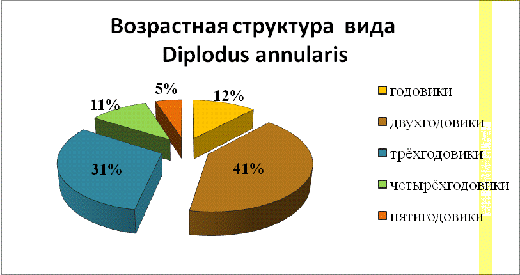
Рис.
5
- Возрастная структура морского карася
Так же нами была взята выборка из 80 особей вида
Diplodus puntazzo. В ней было выделено четыре возрастные группы, которые
насчитывали по 20 особей. Так же нами было изучено среднее процентное
соотношение возрастных групп рыб в уловах.
Наименьшее количество особей четырёхлетнего
возраста - 10%. Чаще встречались двухлетние особи - 42%.
Возрастная структура особей зубырика в уловах
показана на рисунке 6.
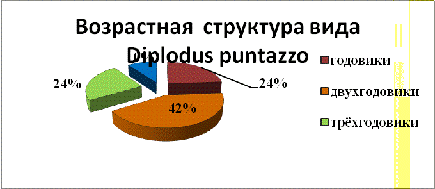
Рис. 6
- Возрастная структура зубарика или хиэны
3.8 Половая структура
У всех 100 исследованных рыб вида Diplodus
annularis был определен пол и стадия зрелости половых продуктов, таким образом,
среди всех возрастных групп было отмечено количество самок - 40 и самцов - 37 и
23 особи на ювенальной стадии. У годовиков все особи оказались на ювенальной
стадии развития. На втором году жизни так же имелись особи на ювенальной стадии
- 3, а преобладали самцы. Таким образом, можно сделать вывод, что самки
созревают на 2-3 году жизни, а самцы на 2.
Таблица 12 -
Половая структура морского карася (Diplodus annularis)
|
Возраст
|
Численность,
%
|
Соотношение
полов
|
Количество
рыб, шт.
|
|
самки
|
самцы
|
|
|
|
2∙
|
-
|
17
|
|
20
|
|
3∙
|
12
|
8
|
1,5:1
|
20
|
|
4∙
|
13
|
7
|
1,9:1
|
20
|
|
5∙
|
15
|
5
|
3:1
|
20
|
У всех 80 исследованных рыб вида Diplodus
puntazzo был определен пол и стадия зрелости половых продуктов, таким образом,
среди всех возрастных групп было отмечено 21 самки и 33 самца и 26 рыб
находились на ювенальной стадии, среднее соотношение полов - 1 к 1,6. Все
годовалые особи находились на ювенальной стадии развития половых продуктов. Как
и предыдущего вида, мы можем сделать вывод, что самцы созревают на 2 году
жизни, тогда как самки только на 3 год.
Таблица 13 -
Половая структура зубарика (Diplodus puntazzo)
|
Возраст
|
Численность,
%
|
Соотношение
полов
|
Количество
рыб, шт.
|
|
самки
|
самцы
|
|
|
|
2∙
|
-
|
14
|
-
|
20
|
|
3∙
|
11
|
9
|
1,2:1
|
20
|
|
4∙
|
10
|
10
|
1:1
|
20
|
3.9 Упитанность и ожирения
внутренностей
Упитанность особей оценивалась по двум
показателям - по Фультону и по Кларк. Показатели упитанности по всем возрастным
группам имели относительно близкие значения. Наименьшими они были у пятилетних
особей, а наибольшими у годовиков. Средняя упитанность по всем возрастным
группа равнялась по Фультону - 3,35, по Кларк - 2,84.
Показатели упитанности особей морского карася
представлены в таблице 14.
Таблица 14
- Упитанность особей морского карася
|
Возраст
|
Коэффициент
упитанности по Фультону
|
Коэффициент
упитанности по Кларк
|
N,
шт.
|
|
1∙
|
3,71
|
3,23
|
20
|
|
2∙
|
3,48
|
2,94
|
20
|
|
3∙
|
3,51
|
2,95
|
20
|
|
4∙
|
3,57
|
2,99
|
20
|
|
5∙
|
2,48
|
2,1
|
20
|
У вида Puntazzo puntazzo показатели упитанности
по всем возрастным группам, как и у вида Diplodus annularis, имели относительно
близкие отношения. Средняя упитанность по всем возрастным группа равнялась по
Фультону - 3,37, по Кларк - 2,60.
Показатели упитанности особей зубарика (Puntazzo
puntazzo) представлены в таблице 15.
Таблица 15
- Упитанность особей зубарика
|
Возраст
|
Коэффициент
упитанности по Фультону
|
Коэффициент
упитанности по Кларк
|
N,
шт.
|
|
1∙
|
3,34
|
2,59
|
20
|
|
2∙
|
3,84
|
2,84
|
20
|
|
3∙
|
3,29
|
2,46
|
20
|
|
4∙
|
3,02
|
2,49
|
20
|
3.10 Динамика зрелости половых
продуктов
Степень зрелости гонад оценивали у возрастных
групп морского карася, исключая первую, особи которой имели гонады ювенальной
стадии зрелости. Это связано с тем, что особи данного вида имели достаточно
развитые половые продукты, уже на первом году жизни.
Степень
зрелости зубарика оценивалась у возрастных групп, исключая первую, особи
которой имели гонады ювенальной стадии зрелости. Следует
отметить, что для особей морского карася и зубарика характерно порционное
икрометание. Внешний вид гонад представлен в приложении В.
Заключение
В результате данного исследования нами были
изучены два вида рыб семейства спаровых наиболее часто встречаемых в уловах
восточной части Чёрного моря. Это виды - морской карась или ласкирь (Diplodus
annularis) и зубарик или хиэна (D. puntazzo).
При анализе полученных данных морфологических
характеристик, нами сделаны следующие вывода: основные особенности и различия
изученных видов касаются в наибольшей степени линейных размеров, так представители
вида D. puntazzo к четырем годам достигают размеров, которые представители вида
D. annularis достигают к пяти годам. Значительное расхождение у данных видов в
строении и пропорциях головы, особенно это касается длины рыла и диаметра
глаза. Эти различия связаны с формой челюсти и зубов, при том условии, что
количество резцов у изученных видов идентично. Меристические признаки довольно
близки по значению. Немного расходится количество жевательных зубов на
челюстях, число позвонков, чешуй в боковой линии. Значительно отличается
окраска тела и плавников данных видов. Так же следует отметить, что у вида D.
annularis лучше развиты спинной и анальный плавники, что скорее всего является
средством защиты. Так D. puntazzo имеют большие размеры тела, но менее развиты
плавники с колючками. Половой диморфизм данным видам не свойственен.
При анализе биологических параметров нами
выделены следующие закономерности, характерные для изученных видов семейства
спаровые: при установлении возраста нами были выделены пять возрастных групп у
вида D. annularis и четыре возрастные группы у вида D. puntazzo. При изучении
темпов линейного роста была определена закономерность - линейный рост особей
ласкиря достаточно замедляется от годовиков к особям достигших возраста 3 года,
а затем происходит скачок, что может быть связано с климатическими изменениями
и миграциями. У представителей вида D. puntazzo наблюдается та же
закономерность. При изучении возрастной структуры нами был изучены средние
соотношения возрастных групп в уловах, где преобладали особи возрастом два и
три года, это может быть связано как с экологическими особенностями данных
видов, так и с селективностью орудий лова.
Библиографический список
1.
Архипов А.Г., Кирносова И.П., Серобаба И.И. и др.Многолетний мониторинг рыбных
ресурсов Чёрного моря // Исследования шельфовой зоны Азово-Черноморского
бассейна: Сб. науч. тр. МГИ, 1991.
.
Брем А. Семейство спаровые, или морские караси // Жизнь животных. - М.:
Государственное издательство географической литературы, 1958.
.Васильева
Е.Д. Рыбы Черного моря // Определитель морских, солоноватоводных, эвригалинных
и проходных видов с цветными иллюстрациями, собранными С. В. Богородским. - М.:
Изд-во ВНИРО, 2007.-238 с.
4.
Вылканов А., Данов X., Маринов X., Владев П. и др. Черное море. - Л.:
Гидрометиздат, 1983. -408 с.
.
Данилевский Н.Н., Иванов Л.С, Каутиш И., Вериоти-Маринеску Ф. Промысловые
ресурсы // Основы биологической продуктивности Черного моря.
-
К.:
Наук, думка, 1979.-С. 291 -299.
.
Емтыль М.Х., Иваненко А.М. Рыбы Юго-запада России. Краснодар, 2002. - С. 219 -
223.
.
Ильин Б.С. 1949 к. Морской карась Sargus annularis (Linne) // Промысловые рыбы
СССР: Описания рыб. - М.: Пищепромиздат.-С. 598-599.
.
Козлов Ю.И., Елецкий Б.Д., Вишневский C.JI. и др. 1998. Современное состояние
экосистем восточного шельфа Чёрного моря // Вестник Краснодарского отдела ВГО.
Вып. 1. - С. 47-56.
.
Моисеев П.А., Азизова Н.А., Куранова И.И. 1981. Ихтиология. -М.: Лёгкая и пищ.
пром-ть. 384 с.
.
Остроумов А.А. 1896. Определитель рыб Чёрного и Азовского морей: с указанием
географического распространения и местонахождения // Вестник
рыбопромышленности. Т.9. - № 7-9. - С. 278-332.
.
Правдин И.Ф. 1966. Руководство по изучению рыб (преимущественно пресноводных).
М.: Пищевая пром-ть. - 376 с.
.
Расс Т.С. Ихтиофауна Черного моря и ее использование // Тр. ИОАН. - 1949. - 4 .
- С . 103 - 123.
.
Расс Т.С. Ихтиофауна Черного моря и некоторые этапы ее истории // Ихтиофауна
Черноморских бухт в условиях антропогенного воздействия. - Киев: Наук, думка,
1993.-С. 6-16.
.
Расс Т.С. Рыбные ресурсы Черного моря и их изменения // Океанология. - 1992. -
32, № 2. - С. 293 -302.
.
Световидов А.Н. 1964. Рыбы Чёрного моря. М.; Л.: Наука. -552 с
.
Степанов В.Н., Андреев В.Н. 1981. Чёрное море (Ресурсы и проблемы). -Л.:
Гидрометеоиздат. 161 с.
.
Сорокин Ю.И. 1982. Чёрное море: Природа, ресурсы. М.: Наука. -216с.
.
Тихонов В.Н. 1951. Рыбы Чёрного моря. Симферополь. - 78 с.
.
Фащук Д.Я., Самышев Э.З., Себах Л.К., Шляхов В.А. 1991. Формы антропогенного
воздействия на экосистему Чёрного моря и её состояние в современных условиях //
Экология моря. Вып. 38. - С. 19-28.
.
Чугунова Н.И. 1952. Методика изучения возраста и роста рыб. -М.: Советская
наука. 115 с.
.
Яковлев В.Н. Состояние биологических ресурсов Черного и Азовского морей (справочное
пособие).- Керчь: ЮгНИРО,
1995. - 64 с.
.
Erdogan N, Duzgunes E., Ogut H. Black Sea fisheries and climate change //
Climate forcing and its impacts on the Black Sea Marine Biota. - CIESM Workshop
Monographs, No 39. - Monaco: CIESM, 2010. - P. 113-120.