Классы задач и математический аппарат
Введение
Современная биология активно использует
различные разделы математики: теорию вероятностей и статистику, теорию
дифференциальных уравнений, теорию игр, дифференциальную геометрию и теорию
множеств для формализации представлений о структуре и принципах
функционирования живых объектов.
Многие ученые высказывали мысль о том, что
область знаний становится наукой только тогда, когда выражает свои законы в
виде математических соотношений. В соответствии с этим самая
"научная" наука - физика - наука о фундаментальных законах природы,
математика для нее - естественный язык. В биологии, для которой предметом
изучения являются индивидуальные живые системы, дело обстоит сложнее. Только в
нашем веке появились экспериментальные биохимия, биофизика, молекулярная биология,
микробиология, вирусология, которые изучают воспроизводимые in vitro
явления и активно используют физические, химические и математические методы.
В связи с индивидуальностью биологических
явлений говорят именно о математических моделях в биологии (а не просто
о математическом языке). Слово модель здесь подчеркивает то
обстоятельство, что речь идет об абстракции, идеализации, математическом
описании скорее не самой живой системы, а некоторых качественных
характеристик протекающих в ней процессов. При этом удается сделать и количественные
предсказания, иногда в виде статистических закономерностей. В отдельных
случаях, например, в биотехнологии, математические модели, как в
технике, используются для выработки оптимальных режимов производства.
Классы
задач и математический аппарат
При разработки любой модели необходимо
определить объект моделирования, цель моделирования и средства моделирования. В
соответствии с объектом и целями математические модели в биологии можно
подразделить на три больших класса. Первый - регрессионные модели, включает
эмпирически установленные зависимости (формулы, дифференциальные и разностные
уравнения, статистические законы) не претендующие на раскрытие механизма
изучаемого процесса. Приведем два примера таких моделей.
1. Зависимость между
количеством производителей хамсы S и количеством молоди от каждого
нерестившегося производителя в большой
имитационной модели динамики рыбного стада Азовского моря выражается в виде
эмпирической формулы (Горстко и др, 1984)
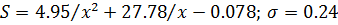
Здесь S - количество сеголеток
(штуки) на каждого нерестившегося производителя; x - количество зашедших
весной из Черного моря в Азовское производителей хамсы (млрд штук); -
среднеквадратичное отклонение.
1. Скорость поглощения кислорода
опадом листьев может быть достаточно хорошо
описывается формулой для логарифма скорости поглощения кислорода:
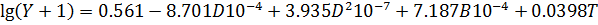
Здесь Y поглощение кислорода,
измеренное в мкл(0,25 г)-1ч-1.; D - число дней, в течение которых выдерживались
образцы; B - процентное содержание влаги в образцах; Т - температура,
измеренная в градусах С.
Эта формула дает несмещенные оценки для скорости
поглощения кислорода во всем диапазоне дней, температур и влажностей, которые
наблюдались в эксперименте, со средним квадратичным отклонением в поглощении
кислорода, равном =0.3190.321.
(Из книги: Д. Джефферс "Введение в
системный анализ: применение в экологии", М., 1981)
Коэффициенты в регрессионных моделях обычно
определяются с помощью процедур идентификации параметров моделей по
экспериментальным данным. При этом чаще всего минимизируется сумма квадратов
отклонений теоретической кривой от экспериментальной для всех точек измерений.
Т.е. коэффициенты модели подбираются таким образом: чтобы минимизировать функционал:
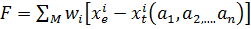
Здесь i - номер точки измерения; xe
- 'экспериментальные значения переменных; хt - теоретические значения
переменных; a1, a2... - параметры, подлежащие оценке; wi -
"вес" i-го измерения; N - число точек измерения.
Второй класс - имитационные модели конкретных
сложных живых систем, как правило, максимально учитывающие имеющуюся информацию
об объекте. Имитационные модели применяются для описания объектов различного
уровня организации живой материи - от биомакромолекул до моделей биогеоценозов.
В последнем случае модели должны включать блоки, описывающие как живые, так и
"косные" компоненты (См. Экология математическая).
Классическим примером имитационных моделей являются модели молекулярной
динамики, в которых задаются координаты и импульсы всех атомов,
составляющих биомакромолекулу и законы их взаимодействия. Вычисляемая на
компьютере картина "жизни" системы позволяет проследить, как
физические законы проявляются в функционировании простейших биологических объектов
- биомакромолекул и их окружения. Сходные модели, в которых элементами
(кирпичиками) уже являются не атомы, а группы атомов, используются в
современной технике компьютерного конструирования биотехнологических
катализаторов и лекарственных препаратов, действующих на определенные активные
группы мембран микроорганизмов, вирусов, или выполняющих другие направленные
действия.
Имитационные модели созданы для описания физиологических
процессов происходящих в жизненно важных органах: нервном волокне, сердце, мозге,
желудочно-кишечном тракте, кровеносном русле. На них проигрываются
"сценарии" процессов, протекающих в норме и при различных патологиях,
исследуется влияние на процессы различных внешних воздействий, в том числе
лекарственных препаратов. Имитационные модели широко используются для описания продукционного
процесса растений и применяются для разработки оптимального режима
выращивания растений с целью получения максимального урожая, или получения
наиболее равномерно распределенного во времени созревания плодов. Особенно
важны такие разработки для дорогостоящего и энергоемкого тепличного хозяйства.
Качественные
(базовые) модели
В любой науке существуют простые модели, которые
поддаются аналитическому исследованию и обладают свойствами, которые позволяют
описывать целый спектр природных явлений. Такие модели называют базовыми. В
физике классической базовой моделью является гармонический осциллятор (шарик -
материальная точка - на пружинке без трения). Базовые модели, как правило,
подробно изучаются в различных модификациях. В случае осциллятора шарик может
быть в вязкой среде, испытывать периодические или случайные воздействия,
например, подкачку энергии, и проч. После того, как досконально математически
изучена суть процессов на такой базовой модели, по аналогии становится
понятными явления, происходящие в гораздо более сложных реальных системах.
Например, релаксация конфирмационных состояний биомакромолекулы рассматривается
аналогично осциллятору в вязкой среде. Таким образом, благодаря простоте и
наглядности, базовые модели становятся чрезвычайно полезными при изучении самых
разных систем.
Все биологические системы различного уровня
организации, начиная от биомакромолекул вплоть до популяций, являются
термодинамический неравновесными, открытыми для потоков вещества и энергии.
Поэтому нелинейность - неотъемлемое свойство базовых систем математической
биологии. Несмотря на огромное разнообразие живых систем, можно выделить
некоторые важнейшие присущие им качественные свойства: рост, самоограничение
роста, способность к переключениям - существование в двух или нескольких
стационарных режимов, автоколебательные режимы (биоритмы), пространственная
неоднородность, квазистохастичность. Все эти свойства можно продемонстрировать
на сравнительно простых нелинейных динамических моделях, которые и выступают в
роли базовых моделей математической биологии.
Неограниченный
рост. Экспоненциальный рост. Автокатализ
В основе любых моделей лежат некоторые
предположения. Модель, построенная на основе этих предположений, становится
самостоятельным математическим объектом, который можно изучать с помощью
арсенала математических методов. Ценность модели определяется тем, насколько
характеристики модели соответствуют свойствам моделируемого объекта. Одно из
фундаментальных предположений, лежащих в основе всех моделей роста -
пропорциональность скорости роста численности популяции, будь то популяция
зайцев или популяция клеток. В основе этого предположения лежит тот
общеизвестный факт, что важнейшей характеристикой живых систем является их
способность к размножению. Для многих одноклеточных организмов или клеток,
входящих в состав клеточных тканей - это просто деление, то есть удвоение числа
клеток через определенный интервал времени, называемый характерным временем
деления. Для сложно организованных растений и животных размножение происходит
по более сложному закону, но в простейшей модели можно предположить, что
скорость размножения вида пропорциональна численности этого вида.
Математически это записывается с помощью
дифференциального уравнения, линейного относительно переменной x,
характеризующей численность (концентрацию) особей в популяции:
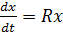 (1)
(1)
Здесь R в общем случае может быть
функцией как самой численности, так и времени, или зависеть от других внешних и
внутренних факторов.
Предположение о пропорциональности скорости
роста популяции ее численности было высказано еще в 18 веке Томасом Робертом
Мальтусом (1766-1834) в книге "О росте народонаселения" (1798).
Согласно закону (1), если коэффициент пропорциональности R=r=сonst
(как это предполагал Мальтус), численность будет расти неограниченно по
экспоненте.
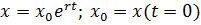
В своих работах Мальтус обсуждает последствия
этого закона в свете того обстоятельства, что производство продовольствия и
других товаров растет линейно, и следовательно, популяция, растущая
экспоненциально, обречена на голод.
Для большинства популяций существуют
ограничивающие факторы, и по тем или иным причинам рост популяции прекращается.
Единственное исключение представляет человеческая популяция, которая на
протяжении всего исторического времени растет даже быстрее, чем по экспоненте.
(См. Экология математическая, раздел Рост численности
человечества). Исследования Мальтуса оказали большое влияние как на
экономистов, так и на биологов. В частности, Чарльз Дарвин пишет в своих
дневниках, что положенные в основу модели Мальтуса предположения и
пропорциональности скорости роста популяции ее численности представляются
весьма убедительными, и из этого следует неограниченный экспоненциальный рост
численности. В то же время, ни одна из популяций в природе не растет до
бесконечности. Следовательно, существуют причины, препятствующие такому росту.
Одну из таких причин Дарвин видит в борьбе видов за существование.
Закон экспоненциального роста справедлив на
определенной стадии роста для популяций клеток в ткани, водорослей или бактерий
в культуре. В моделях математическое выражение, описывающее увеличение скорости
изменения величины с ростом самой этой величины, называют автокаталитическим
членом (авто - само, катализ - модификация скорости реакции, обычно
ускорение, с помощью веществ, не принимающих участия в реакции) Таким образом,
автокатализ - "само ускорение" реакции.
Ограниченный
рост. Уравнение Ферхюльста
Базовой моделью, описывающей ограниченный рост,
является модель Ферхюльста (1848):
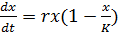 (2)
(2)
Здесь параметр K носит название
"емкости популяции" и выражается в единицах численности (или
концентрации). Он не имеет какого-либо простого физического или биологического
смысла и носит, системный характер, то есть определяется целым рядом
различных обстоятельств, среди них ограничения на количество субстрата для
микроорганизмов, доступного объема для популяции клеток ткани, пищевой базы или
убежищ для высших животных.
График зависимости правой части уравнения (2) от
численности x и численности популяции от времени представлены на
рис. 1 (а и б).
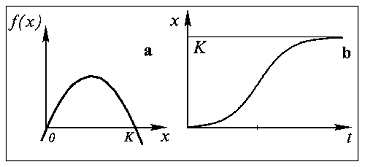
Рис.1. Ограниченный рост. Зависимость величины
скорости роста от численности (а) и численности от времени (б) для
логистического уравнения.
В последние десятилетия уравнение Ферхюльста
переживает вторую молодость. Изучение дискретного аналога уравнения (2) выявило
совершенно новые и замечательные его свойства [3,5]. Рассмотрим численность
популяции в последовательные моменты времени. Это соответствует реальной
процедуре пересчета особей (или клеток) в популяции. В самом простом виде
зависимость численности на временном шаге номер n+1 от
численности предыдущем шаге n можно записать в виде:
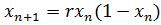 (3)
(3)
Поведение во времени переменной xn
может носить характер не только ограниченного роста, как было для непрерывной
модели (2), но также быть колебательным или квазистохастическим (рис.2).
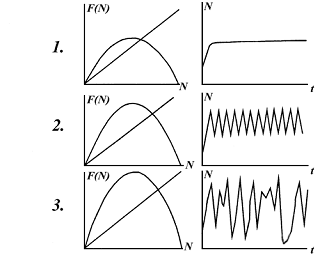
Рис.2. Вид функции зависимости численности на
последующем шаге от численности на предыдущем шаге (а) и поведение численности
во времени (б) при разных значениях параметра r: 1 - ограниченный
рост; 2 - колебания, 3 - хаос.
Тип поведения зависит от величины константы
собственной скорости роста r. Кривые, представляющие вид
зависимости значения численности в данный момент времени (t+1) от
значений численности в предыдущий момент времени t представлены
на рис. 2 слева. Справа представлены кривые динамики численности - зависимости
числа особей в популяции от времени. Сверху вниз значение параметра собственной
скорости роста r увеличивается.
Характер динамики численности определяется видом
кривой зависимости F(t+1) от F(t). Эта кривая
отражает изменение скорости прироста численности от самой численности. Для всех
представленных на рис. 2 слева кривых эта скорость нарастает при малых
численностях, и убывает, а затем обращается в нуль при больших численностях.
Динамический тип кривой роста популяции зависит от того, насколько быстро
происходит рост при малых численностях, т.е. определяется производной
(тангенсом угла наклона этой кривой) в нуле, который определяется коэффициентом
r - величиной собственной скорости роста. Для небольших r
(r<3) численность популяции стремится к устойчивому равновесию.
Когда график слева становится более крутым, устойчивое равновесие переходит в
устойчивые циклы. По мере увеличения численности длина цикла растет, и значения
численности повторяются через 2, 4, 8,..., 2n поколений. При
величине параметра r>5,370 происходит хаотизация решений. При
достаточно больших r динамика численности демонстрирует хаотические всплески
(вспышки численности насекомых).
Уравнения такого типа неплохо описывают динамику
численности сезонно размножающихся насекомых с неперекрывающимися поколениями.
При этом некоторые достаточно просто измеряемые характеристики популяций,
демонстрирующих квазистохастическое поведение, имеют регулярный характер. В
некотором смысле, чем хаотичнее поведение популяции, тем оно предсказуемое.
Например, при больших x амплитуда вспышки может быть прямо
пропорциональна времени между вспышками.
Дискретное описание оказалось продуктивным для
систем самой различной природы. Аппарат представления динамического поведения
системы на плоскости в координатах [xt, xt+T]
позволяет определить, является наблюдаемая система колебательной или
квазистохастической. Например, представление данных электрокардиограммы
позволило установить, что сокращения человеческого сердца в норме носят
нерегулярный характер, а в период приступов стенокардии или в предынфарктном
состоянии ритм сокращения сердца становится строго регулярным. Такое
"ужесточение" режима является защитной реакцией организма в
стрессовой ситуации и свидетельствует об угрозе жизни системы.
Отметим, что решение разностных уравнений лежит
в основе моделирования любых реальных биологических процессов. Богатство
динамического поведения модельных траекторий разностных уравнений является
основой их успешного применения для описания сложных природных явлений. При
этом ограниченность параметрических областей существования определенного типа
режимов служит дополнительным основанием для оценки адекватности предлагаемой
модели.
Еще более интересные математические объекты
получаются, если переписать уравнение (3) в виде:
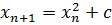 (4)
(4)
и рассматривать константу с в
комплексной области. При этом получаются объекты, называемые множествами
Мандельброта Подробнее об этих множествах можно прочитать в книге
"Красота фракталов" (Образы комплексных динамических систем)[2], там
же приведены их многочисленные красочные изображения. Имеют ли эти объекты
биологическую интерпретацию, имеющую под собой глубокий смысл, или это просто
красивый "сюрприз", который нам преподносит базовая система? Пока на
этот вопрос нет окончательного ответа.
Ограничения
по субстрату. Модели Моно и Михаэлиса-Ментен
Одной из причин ограничения роста может быть
недостаток пищи (лимитирование по субстрату на языке микробиологии).
Микробиологи давно подметили, что в условиях лимитирования по субстрату
скорость роста растет пропорционально концентрации субстрата, а если субстрата
вдоволь - выходит на постоянную величину, определяемую генетическими
возможностями популяции. В течение некоторого времени численность популяции
растет экспоненциально, пока скорость роста не начинает лимитироваться
какими-либо другими факторами. Это означает, что зависимость скорости роста R
в формуле (1) от субстрата может быть описана в виде:
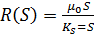 (5)
(5)
Здесь КS - константа,
равная концентрации субстрата, при которой скорость роста равна половине
максимальной. 0 - максимальная скорость роста, равная
величине r в формуле (2). Это уравнение было впервые написано
крупнейшим французским биохимиком. Жаком Моно (1912-1976). Совместно с Франсуа
Жакобом им были разработаны представления о роли транспортной рибонуклеиновой
кислоты (mRNA) в аппарате размножения клетки. В развитие представлений о генных
комплексах, которые были ими названы оперонами, Жакоб и Моно постулировали
существование класса генов, которые регулируют функционирование других генов
путем воздействия на синтез транспортной РНК. Такой механизм генной регуляции
впоследствии полностью подтвердился для бактерий, за что обоим ученым (а также
Андре Львову) была присуждена Нобелевская премия 1965 г. Ниже рассмотрена
знаменитая модель генной регуляции синтеза двух ферментов, названная триггерной
моделью Жакоба и Моно.
Жак Моно был также философом науки и незаурядным
писателем. В своей знаменитой книге "Случайность и необходимость",
1971 Моно высказывает мысли о случайности возникновения жизни и эволюции, а
также о роли человека и его ответственности за происходящие на Земле процессы.
Любопытно, что модель Моно (5) по форме совпадает
с уравнением Михаэлиса-Ментен (1913), которое описывает зависимость скорости
ферментативной реакции от концентрации субстрата при условии, когда общее
количество молекул фермента постоянно и значительно меньше количества молекул
субстрата:
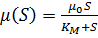 (6)
(6)
Здесь КМ - константа
Михаэлиса, одна из важнейших для ферментативных реакций величина, определяемая
экспериментально, имеющая смысл и размерность концентрации субстрата, при
которой скорость реакции равна половине максимальной.
Закон Михаэлиса-Ментен выводится на основании
уравнений химической кинетики и описывает скорость образования продукта в
соответствии со схемой:
+S [ES] E+P.
Сходство уравнений (5) и (6) не случайно.
Формула Михаэлиса-Ментен (5) отражает более глубокие закономерности кинетики
ферментативных реакций, которые в свою очередь определяют жизнедеятельность и
рост микроорганизмов, описываемые эмпирической формулой (5).
Базовая
модель взаимодействия. Конкуренция. Отбор
Биологические системы вступают во взаимодействие
друг с другом на всех уровнях, будь то взаимодействие биомакромолекул в
процессе биохимических реакций, или взаимодействие видов в популяциях.
Взаимодействие может протекать в структурах, тогда система может быть
охарактеризована определенным набором состояний, так происходит на уровне
субклеточных, клеточных и организменных структур. Кинетика процессов в
структурах в математических моделях как правило описывается с помощью систем
уравнений для вероятностей состояний комплексов.
В случае, когда взаимодействие происходит
случайно, его интенсивность определяется концентрацией взаимодействующих
компонентов и их подвижностью обобщенной диффузией. Именно такие представления
приняты в базовых моделях взаимодействия видов. Классической книгой, в которой
рассматриваются математические модели взаимодействия видов стала книга Вито
Вольтерра "Математическая теория борьбы за существование" (1931) [1].
Книга, построена как математический трактат, в ней постулированы в
математической форме свойства биологических объектов и их взаимодействий, а
затем эти взаимодействия исследуются как математические объекты, Именно с этой
работы В. Вольтерра начались современная математическая биология и
математическая экология.
Вито Вольтерра (1860-1940) завоевал мировую
известность своими работами в области интегральных уравнений и функционального
анализа. Кроме чистой математики его всегда интересовали вопросы применения
математических методов в биологии, физике, социальных науках. В годы службы в
ВВС в Италии, он много работал над вопросами военной техники и технологии
(задачи баллистики, бомбометания, эхо локации). В этом человеке сочетался
талант ученого и темперамент активного политика, принципиального противника
фашизма. Он был единственным итальянским сенатором, проголосовавшим против
передачи власти Муссолини. Когда в годы фашистской диктатуры в Италии Вольтерра
работал во Франции, Муссолини, желая привлечь на свою сторону всемирно
известного ученого, предлагал ему различные высокие посты в фашистской Италии,
но всегда получал решительный отказ. Антифашистская позиция привела Вольтерра к
отказу от кафедры в Римском университете и от членства в итальянских научных
обществах.
Серьезно вопросами динамики популяций В.
Вольтерра стал интересоваться с 1925 г. после бесед с молодым зоологом Умберто
Д'Анкона, будущим мужем его дочери, Луизы. Д'Анкона, изучая статистику рыбных
рынков на Адриатике, установил любопытный факт: когда в годы первой мировой
войны (и сразу вслед за ней) интенсивность промысла резко сократилась, то в
улове увеличилась относительная доля хищных рыб. Такой эффект предсказывался
моделью "хищник-жертва", предложенной Вольтерра. Эту модель мы
рассмотрим ниже. По сути дела это был первый успех математической биологии.
Вольтерра предположил по аналогии со
статистической физикой, что интенсивность взаимодействия пропорциональна
вероятности встречи (вероятности столкновения молекул), то есть произведению
концентраций. Это и некоторые другие предположения (См. Популяционная
динамика) позволили построить математическую теорию взаимодействия
популяций одного трофического уровня (конкуренция) или разных трофических
уровней (хищник-жертва).
Системы, изученные Вольтерра, состоят из
нескольких биологических видов и запаса пищи, который используют некоторые из
рассматриваемых видов. О компонентах системы формулируются следующие допущения.
. Особи каждого вида отмирают так, что в единицу
времени погибает постоянная доля существующих особей.
. Хищные виды поедают жертвы, причем в единицу
времени количество съеденных жертв всегда пропорционально вероятности встречи
особей этих двух видов, т.е. произведению количества хищников на количество
жертв.
. Если имеются пища в неограниченном количестве
и несколько видов, которые способны ее потреблять, то доля пищи, потребляемая
каждым видом в единицу времени, пропорциональна количеству особей этого вида,
взятого с некоторым коэффициентом, зависящим от вида (модели межвидовой конкуренции).
. Если вид питается пищей, имеющейся в
неограниченном количестве, прирост численности вида за единицу времени
пропорционален численности вида.
. Если вид питается пищей, имеющейся в
ограниченном количестве, то его размножение регулируется скоростью потребления
пищи, т.е. за единицу времени прирост пропорционален количеству съеденной пищи.
Перечисленные гипотезы позволяют описывать
сложные живые системы при помощи систем обыкновенных дифференциальных
уравнений, в правых частях которых имеются суммы линейных и билинейных членов.
Как известно, такими уравнениями описываются и системы химических реакций.
Действительно, согласно гипотезам Вольтерра,
скорость процесса отмирания каждого вида пропорциональна численности вида. В
химической кинетике это соответствует мономолекулярной реакции распада
некоторого вещества, а в математической модели - отрицательным линейным членам
в правых частях уравнений. Согласно представлениям химической кинетики,
скорость бимолекулярной реакции взаимодействия двух веществ пропорциональна
вероятности столкновения этих веществ, т.е. произведению их концентрации. Точно
так же, согласно гипотезам Вольтерра, скорость размножения хищников (гибели
жертв) пропорциональна вероятности встреч особей хищника и жертвы, т.е.
произведению их численностей. И в том и в другом случае в модельной системе
появляются билинейные члены в правых частях соответствующих уравнений. Наконец,
линейные положительные члены в правых частях уравнений Вольтерра, отвечающие
росту популяций в неограниченных условиях, соответствуют автокаталитическим
членам химических реакций. Такое сходство уравнений в химических и
экологических моделях позволяет применить для математического моделирования
кинетики популяций те же методы исследований, что и для систем химических реакций.
Можно показать, что вольтеровские уравнения могут быть получены не только из
локального "принципа встреч", ведущего свое происхождение из
статистической физики, но и исходя из баланса масс каждого из компонентов
ценоза и энергетических потоков между этими компонентами.
Рассмотрим простейшую из Вольтерра моделей
модель отбора на основе конкурентных отношений. Эта модель работает при
рассмотрении конкурентных взаимодействий любой природы биохимических соединений
различного типа оптической активности, конкурирующих клеток, особей, популяций.
Ее модификации применяются для описания конкуренции в экономике.
Пусть имеется два совершенно одинаковых вида с
одинаковой скоростью размножения, которые являются антагонистами, то есть при
встрече они угнетают друг друга. Модель их взаимодействия может быть записана в
виде:
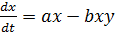
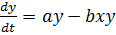 (7)
(7)
Согласно такой модели, симметричное состояния
сосуществования обоих видов является неустойчивым, один из взаимодействующих
видов обязательно вымрет, а другой размножится до бесконечности.
Введение ограничения на субстрат (типа 5) или
системного фактора, ограничивающего численность каждого из видов (типа 2)
позволяет построить модели, в которых один из видов выживает и достигает
определенной стабильной численности. Они описывают известный в
экспериментальной экологии принцип конкуренции Гаузе, в соответствии с которым
в каждой экологической нише выживает только один вид.
В случае, когда виды обладают различной
собственной скоростью роста, коэффициенты при автокаталитических членах в
правых частях уравнений будут различными, а фазовый портрет системы становится
несимметричным. При различных соотношениях параметров в такой системе возможно
как выживание одного из двух видов и вымирание второго (если взаимное угнетение
более интенсивно, чем само регуляция численности), так и сосуществование обоих
видов, в случае , когда взаимное угнетение меньше, чем самоограничение
численности каждого из видов.
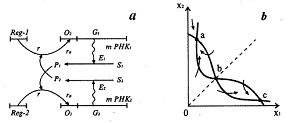
Рис.4. Схема синтеза двух ферментов Жакоба и
Моно (а) и фазовый портрет триггерной систем (б) [6].
Еще одной классической триггерной системой
является модель альтернативного синтеза двух ферментов Жакоба и Моно. Схема синтеза
приведена на рис. 4а. Ген-регулятор каждой системы синтезирует неактивный
репрессор. Этот репрессор, соединяясь с продуктом противоположной системы
синтеза ферментов, образует активный комплекс. Активный комплекс, обратимо
реагируя с участком структурного гена опероном, блокирует синтез mРНК. Таким
образом, продукт второй системы Р2 является корепрессором первой
системы, а Р1 - корепрессором второй. При этом в процессе
корепрессии могут участвовать одна, две и более молекул продукта. Очевидно, что
при таком характере взаимодействий при интенсивной работе первой системы вторая
будет заблокирована, и наоборот. Модель такой системы предложена и подробно
изучены в школе проф. Д.С.Чернавского [4] После соответствующих упрощений,
уравнения, описывающие синтез продуктов Р1 и
Р2 имеют вид:
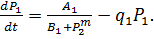
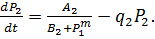 (8)
(8)
Здесь P1, P2
- концентрации продуктов, величины A1, A2, B1,
B2, выражаются через параметры своих систем.
Показатель степени m показывает, сколько молекул активного
репрессора (соединений молекул продукта с молекулами неактивного репрессора,
который предполагается в избытке) соединяются с опероном для блокировки синтеза
mRNK.
Фазовый портрет
системы, (изображение траекторий системы при разных начальных условиях на
координатной плоскости, по осям которой отложены величины переменных системы),
для m=2 изображен на рис.4б. Он имеет тот же вид, что и фазовый портрет системы
двух конкурирующих видов. Сходство свидетельствует о том, что в основе способности
системы к переключениям лежит конкуренция - видов, ферментов, состояний.
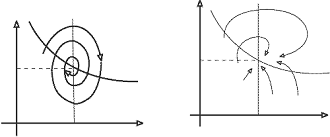
Рис.5. Модель химических реакций Лотки. Фазовый
портрет системы при значениях параметров, соответствующих затухающим
колебаниям.
Классические
модели Лотки и Вольтерра
Первое понимание, что собственные ритмы возможны
в богатой энергией системе за счет специфики взаимодействия ее компонентов
пришло после появления простейших нелинейных моделей взаимодействия -
химических веществ в уравнениях Лотки, и взаимодействия видов - в моделях
Вольтерра [1,3].
Уравнение Лотки рассмотрено им в 1926 г. в книге
и описывает систему следующих химических реакций
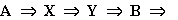
В некотором объеме находится в избытке вещество А.
Молекулы А с некоторой постоянной скоростью 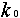 превращаются
в молекулы вещества X (реакция нулевого порядка). Вещество X
может превращаться в вещество Y, причем скорость этой реакции тем
больше, чем больше концентрация вещества Y - реакция второго порядка. В
схеме это отражено обратной стрелкой над символом y. Молекулы Y в
свою очередь необратимо распадаются, в результате образуется вещество B
(реакция первого порядка).
превращаются
в молекулы вещества X (реакция нулевого порядка). Вещество X
может превращаться в вещество Y, причем скорость этой реакции тем
больше, чем больше концентрация вещества Y - реакция второго порядка. В
схеме это отражено обратной стрелкой над символом y. Молекулы Y в
свою очередь необратимо распадаются, в результате образуется вещество B
(реакция первого порядка).
Запишем систему уравнений, описывающих реакцию:
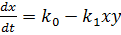
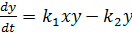
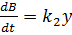 (9)
(9)
Здесь X, Y, B - концентрации химических
компонентов. Первые два уравнения этой системы не зависят от B, поэтому
их можно рассматривать отдельно. При определенных значениях параметров в
системе возможны затухающие колебания.
Базовой моделью незатухающих колебаний служит
классическое уравнение Вольтерра, описывающее взаимодействие видов типа
хищник-жертва. Как и в моделях конкуренции (8), взаимодействие видов
описывается в соответствии с принципами химической кинетики: скорость убыли
количества жертв (x) и скорость прибыли количества хищников (y)
считается пропорциональными их произведению
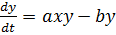 (10)
(10)
На рис. 6 представлены фазовый портрет
системы, по осям которого отложены численности жертв и хищников - (а) и
кинетика численности обоих видов - зависимость численности от времени-(б).
Видно, что численности хищников и жертв колеблются в противофазе.
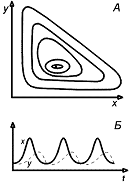
Рис.6. Модель хищник-жертва Вольтерра,
описывающая незатухающие колебания численности. А. Фазовый портрет. Б.
Зависимость численности жертвы и хищника от времени.
Модель Вольтерра имеет один существенный
недостаток. Параметры колебаний ее переменных меняются при флуктуациях
параметров и переменных системы. Такую систему называют негрубой.
Этот недостаток устранен в более реалистичных
моделях. Модификация модели Вольтерра с учетом ограниченности субстрата в форме
Моно (уравнение 5) и учет самоограничения численности (как в уравнении 2)
приводит к модели, подробно изученной А.Д. Базыкиным в книге "Биофизика
взаимодействующих популяций" (1985).
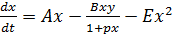
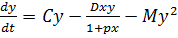 , (11)
, (11)
Система (11 представляет собой некий кентавр,
составленный из базовых уравнений (1, 2, 5, 10)и объединяющий их свойства.
Действительно, при малых численностях и в отсутствие хищника жертва (x)
будет размножаться по экспоненциальному закону(1). Хищник (y) в
отсутствие жертв будут вымирать также по экспоненте. Если особей того или иного
вида много, в соответствии с базовой моделью (2) срабатывает системный
ферхюльстовский фактор (член -Ex2 в первом
уравнении, и -My2 - во втором). Интенсивность
взаимодействия видов считается пропорциональной произведению их численностей
(как в модели (10)) и описывается в форме Моно (модель 5). Здесь роль субстрата
играет вид-жертва , а роль микроорганизмов - вид-хищник. Таким образом, модель
(11) брала в себя свойства базовых моделей (1), (2), (5), (10).
Но модель (11) представляет собой не просто
сумму свойств этих моделей. С ее помощью можно описать и гораздо более сложные
типы поведения взаимодействующих видов: наличие двух устойчивых стационарных
состояний, затухающие колебания численностей и проч. При некоторых значениях
параметров система становится автоколебательной. В ней с течением
времени устанавливается режим, при котором переменные изменяются периодически с
постоянным периодом и амплитудой независимо от начальных условий.
Волны
жизни
До сих пор мы говорили о базовых моделях поведения
живых систем во времени. Стремление к росту и размножению ведет к
распространению в пространстве, занятию нового ареала, экспансии живых
организмов. Жизнь распространяется так же как пламя по степи во время степного
пожара. Эта метафора отражает тот факт, что пожар (в одномерном случае -
распространение пламени по бикфордову шнуру) описывается с помощью той же
базовой модели, что и распространение вида. Знаменитая в теории горения модель
ПКП (Петровского - Колмогорова - Пискунова) впервые была предложена ими в 1937
г. именно в биологической постановке как модель распространения доминирующего
вида в пространстве. Все три автора этой работы являются крупнейшими
российскими математиками. Академик Иван Георгиевич Петровский (1901-1973) -
автор фундаментальных трудов по теории дифференциальных уравнений, алгебре,
геометрии, математической физике, в течение более 20 лет был ректором
Московского Государственного университета им. М.В. Ломоносова. (1951-1973).
Андрей Николаевич Колмогоров (1903 - ) глава российской математической школы по
теории вероятностей и теории функций, автор фундаментальных трудов по
математической логике, топологии, теории дифференциальных уравнений, теории
информации, организатор школьного и университетского математического
образования, написал несколько работ, в основу которых положены биологические
постановки. В частности в 1936 г. он предложил и подробно исследовал обобщенную
модель взаимодействия видов типа хищник-жертва (исправленный и дополненный
вариант 1972).(См. Популяционная динамика)
Рассмотрим постановку задачи о распространении
вида в активной - богатой энергией (пищей) среде. Пусть в любой точке прямой r>0
размножение вида описывается функцией f(x) = x(1-x). В начальный
момент времени вся область слева от нуля занята видом x,
концентрация которого близка к единице.. Справа от нуля - пустая территория. В
момент времени t=0 вид начинает распространяться (диффундировать)
вправо с константой диффузии D. Процесc описывается уравнением:
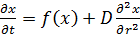 (12)
(12)
При t>0 в такой системе
начинает распространяться волна концентраций в область r>0,
которая является результатом двух процессов: случайного перемещения особей
(диффузии частиц) и размножения, описываемого функцией f(x). С
течением времени фронт волны перемещается вправо, причем его форма приближается
к определенной предельной форме. Скорость перемещения волны определяется
коэффициентом диффузии и формой функции f(x), и для функции f(x),
равной нулю при x=0 и x=1 и положительной в промежуточных
точках, выражается простой формулой: =2Df'(0).
Изучение пространственного перемещения в модели
хищник-жертва (10) показывает, что в такой системе в случае неограниченного
пространства будут распространяться волны "бегства и погони", а в
ограниченном пространстве установятся стационарные пространственные
неоднородные структуры (диссипативные структуры), или авто волны, в зависимости
от параметров системы.
модель имитационный базовый
математический
Автоволны
и диссипативные структуры. Базовая модель "брюсселятор"
На рассмотренной выше одномерной модели (14)
видно, что взаимодействие нелинейной химической реакции и диффузии приводит к
нетривиальным режимам. Еще более сложного поведения следует ожидать в двумерных
моделях, описывающих взаимодействие компонентов системы. Первая такая модель
была изучена Тьюрингом в работе под названием "Химические основы
морфогенеза". Алан М. Тьюринг (1912-1954) английский математик и логик,
прославился своими работами по компьютерной логике и теории автоматов. В 1952
г. он опубликовал первую часть исследования, посвященного математической теории
образования структур в первоначально однородной системе, где одновременно
проходят химические реакции, в том числе автокаталитические процессы,
сопровождаемые потреблением энергии, и пассивные процессы переноса - диффузия.
Это исследование осталось незаконченным, так как он покончил жизнь
самоубийством, находясь под действием депрессантов, которыми его принудительно
лечили в тюрьме, где он отбывал срок по обвинению в гомосексуализме.
Работа Тьюринга стала классической, ее идеи
легли в основу современной теории нелинейных систем, теории самоорганизации и
синергетики. Рассматривается система уравнений:
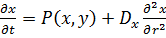
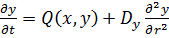 (13)
(13)
Уравнения такого типа называются уравнениями
"реакция-диффузия". В линейных системах диффузия процесс, который
приводит к выравниванию концентраций во всем реакционном объеме. Однако в
случае нелинейного взаимодействия переменных x и y,
в системе может возникать неустойчивость гомогенного стационарного состояния и
образуются сложные пространственно-временные режимы типа авто волн или диссипативных
структур - стационарных во времени и неоднородных по пространству распределений
концентраций, существование которых поддерживается в активных средах за счет
потребления энергии системы в процессах диссипации. Условием возникновения
структур в таких системах является различие коэффициентов диффузии реагентов, а
именно, наличие близкодействующего "активатора" с малым коэффициентом
диффузии и дальнодействующего "ингибитора" с большим коэффициентом
диффузии.
Такие режимы в двухкомпонентной системе были
изучены в деталях на базовой модели под названием "брюсселятор"
(Пригожин и Лефевр, 1968), названной в честь брюссельской научной школы под
руководством И.Р. Пригожина, в которой наиболее интенсивно проводились эти
исследования. Илья Романович Пригожин (род 1917 г. в Москве) - всю жизнь
работал в Бельгии. С 1962 г. он - директор Международного Сольвеевского
института физической химии в Брюсселе, а с 1967 г. - директор Центра
статистической механики и термодинамики Техасского университета (США). В 1977
г. он получил Нобелевскую премию за работы по нелинейной термодинамике, в
частности по теории диссипативных структур - устойчивых во времени неоднородных
в пространстве структур. Пригожин является автором и соавтором целого ряда книг
["Термодинамическая теория структуры, устойчивости и флуктуаций",
"Порядок из хаоса", "Стрела времени", и др.], в которых он
развивает математические, физико-химические, биологические и философские идеи
теории самоорганизации в нелинейных системах, исследует причины и закономерности
рождения "порядка из хаоса" в богатых энергией открытых для потоков
вещества и энергии системах, далеких от термодинамического равновесия, под
действием случайных флуктуаций.
Классическая модель "брюсселятор"
имеет вид
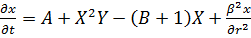
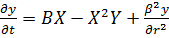 (14)
(14)
и описывает гипотетическую схему химических
реакций:
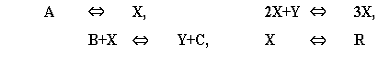
Ключевой является стадия превращения двух
молекул x и одной молекулы y в x так
называемая тримолекулярная реакция. Такая реакция возможна в процессах с
участием ферментов с двумя каталитическими центрами. Нелинейность этой реакции
в сочетании с процессами диффузии вещества и обеспечивает возможность
пространственно-временных режимов, в том числе образование пространственных структур
в первоначально однородной системе морфогенез.
Заключение
Современная математическая биология использует
различный математический аппарат для моделирования процессов в живых системах и
формализации механизмов, лежащих в основе биологических процессов.
Имитационные модели позволяют на компьютерах
моделировать и прогнозировать процессы в нелинейных сложных системах, каковыми
являются все живые системы, далекие от термодинамического равновесия. Базовые
модели математической биологии в виде простых математических уравнений отражают
самые главные качественные свойства живых систем: возможность роста и его
ограниченность, способность к переключениям, колебательные и стохастические
свойства, пространственно-временные неоднородности. На этих моделях изучаются
принципиальные возможности пространственно-временной динамики поведения систем,
их взаимодействия, изменения поведения систем при различных внешних
воздействиях - случайных, периодических и т.п.
Любая индивидуальная живая система требует
глубокого и детального изучения, экспериментального наблюдения и построения
своей собственной модели, сложность которой зависит от объекта и целей
моделирования.
Литература
2. Пайтген
Х.-О., Рихтер П.Х. Красота фракталов. Образы комплексных динамических систем.
М., Мир, 1993, 176 с.
. Ризниченко
Г.Ю., Рубин А.Б. Математические модели биологических продукционных процессов.
М., Изд. МГУ, 1993, 301 с.
. Романовский
Ю.М., Степанова Н.В., Чернавский Д.С. Математическая биофизика. М., Наука,
1984, 304 с.
. Рубин
А.Б., Пытьева Н.Ф., Ризниченко Г.Ю. Кинетика биологических процессов. М., МГУ.,
1988
. Свирежев
Ю.М., Логофет. Устойчивость биологических сообществ М., Наука, 1978, 352c
. Базыкин
А.Д. Биофизика взаимодействующих популяций. М., Наука, 1985, 165 с.
8. J.D.
Murray "Mathematical Biology", Springer, 1989, 1993.