Биология ротана-головешки (Perccottus glenii,) в водоемах юга Западной Сибири
Содержание
Введение
. Обзор литературы
.1 Рыбы семейства головешковые
.2 Головешка-ротан - perccottus glenii Dybowski, 1877
.3 Распространение
.4 Возраст и размеры
.5 Экология ротана-головешки
.6 Особенности роста и образование эко-морфологических форм
.7 Питание и пищевое поведение
.8 Половой диморфизм
.9 Нерест
.10 Конкуренты и хищники
.11 Воздействие на аборигенные сообщества
.12 Возможный контроль
. Место и условия проведения работы
.1 Физико-географический очерк
.2 Характеристика организации
. Материалы и методы исследования
.1 Проведение биологического анализа и морфометрии
.2 Определение возраста по отолитам
.3 Жирность
.4 Изучение питания рыб
.5 Изучение полового состава
.6. Плодовитость
. Собственные исследования
.1 Общебиологический анализ разных половозрастных групп
ротана из Бердского пруда
.2 Морфометрия ротана-головешки разных половозрастных групп
.3 Питание ротана-головешки в различных водоемах
.4 Темп роста ротана в разных половозрастных группах и
водоемах
.5 Половозрастная структура популяции ротана-головешки
5.
Безопасность жизнедеятельности
.
Охрана природы
Выводы
Предложения
Библиографический список
Реферат
Квалификационная работа содержит 84 страницы, 10 таблиц, 9 рисунков, 5
графиков, 61 источник литературы.
БИОЛОГИЯ РОТАНА-ГОЛОВЕШК (PERCCOTTUS GLENII DYBOWSKI, 1877) В ВОДОЕМАХ ЮГА ЗАПАДНОЙ
СИБИРИ
Объектом исследований являлась рыба ротан-головешка. Были обследованы
самки и самцы рыб из Бердского пруда, оз. Улугуль и р. Алей. Общее количество
исследуемых рыб составило 250 экземпляров.
Целью исследования было изучение биологии ротана-головешки в водоемах юга
Западной Сибири.
Впервые в водоемах юга Западной Сибири изучена биология ротана-головешки.
Проведен анализ морфометрического анализа в условиях разнотипных районах, что
позволяет оценить адаптивность ротана. Изучено питание в условиях новой среды
его обитания. Выявлены закономерности динамики численности
(размерно-возрастная, половая), тенденции увеличения численности или ее
снижения и проведена оценка пресса вселенцев на кормовую базу водоемов.
Полученные данные необходимы для объективной оценки биологических
ресурсов водоемов с учетом не только комплекса промысловых, но и непромысловых
рыб.
Введение
Данная работа ориентирована на изучение роли представителей непромысловой
фауны, на примере ротана-головешки, в условиях естественных водоемов.
Проблема чужеродных видов - это одна из наиболее актуальных проблем
современности, известная так же, как проблема биологических инвазий. Инвазии
чужеродных видов нередко приводят к существенным перестройкам в структуре
сообществ и серьезным экономическим потерям (Biological invasions: economic, 2002; Lodge et al., 2006). Между вселенцами могут устанавливаться обратные
положительные связи, облегчающие взаимное освоение новых экосистем, однако наиболее
распространенными являются односторонние, когда лишь один из вселенцев
способствует натурализации другого (Симберлов, 2006). Познание влияния
положительных связей на формирование сообществ является необходимым условием
развития теории биологических инвазий (Bruno et al., 2003; Симберлов, 2006).
В последнее время процессы, связанные с появлением чужеродных видов и
воздействиями их на местные сообщества, принято именовать биологическими
инвазиями. Под биологическими инвазиями понимаются все случаи проникновения
живых организмов в экосистемы, расположенные за пределами их первоначального
(обычно, естественного) ареала. Таким образом, к биологическим инвазиям
относятся вселения чужеродных видов, произошедшие в результате:
естественных перемещений, связанных с флюктуациями численности и
климатическими изменениями;
интродукции и реинтродукции важных в хозяйственном отношении полезных
организмов;
случайных заносов с балластными водами, с импортной сельскохозяйственной
продукцией, багажом, с интродуцентами и т.п.
Чужеродные виды всегда оказывали существенное воздействие на экосистемы.
Особенно заметными эти воздействия стали со второй половины ХХ столетия, когда
расширения ареалов и проникновение живых организмов в новые сообщества
происходили на фоне общих климатических и антропических изменений.
Во многих случаях инвазийные виды, вступая в контакты с популяциями
видов-аборигенов, существенно преобразуют структуру биоценозов, и их появление
имеет глобальные экологические, экономические, а иногда, и социальные последствия.
Во всем мире в последние годы наблюдается интенсификация исследований в
области биологических инвазий. Разработка мер по их предотвращению, смягчению
последствий и мониторингу являются обязанностью всех стран (к ним относится и
Россия), подписавших в 1992 г. в Рио-де-Жанейро Конвенцию о биологическом
разнообразии.
В РФ фундаментальные и прикладные исследования инвазионного процесса
(анализ последствий проникновения чужеродных видов и разработка мер по его
контролю) ведутся спорадически и в небольшом объеме. В то же время проблема
чужеродных видов (биологических инвазий) имеет для России исключительно важное
социально-экономическое значение. Число случаев возникновения крупных
экологических катакликзмов, вызванных биологическими инвазиями чужеродных
видов, постоянно растет. Достаточно назвать несколько примеров инвазий,
непосредственно касающихся нашей страны: сорняки рода амброзия, колорадский
жук, золотистая нематода, фомопсис подсолнечника, гребневик мнемиопсис, рачек
церкопагис, рыба ротан.
Если попытаться посмотреть на проблему биологических инвазий в плане их
влияния на аборигенные виды и их разнообразие, то совершенно очевидными
представляются следующие формы воздействия чужеродных видов как лимитирующих
факторов:
. Инвазийные виды могут существенно изменить среду обитания аборигенных
видов (особенно в случаях, когда инвазийные виды являются «ключевыми видами»
сообщества) путем изменения структуры и функции экосистемы.
. Инвазийные виды могут стать конкурентами аборигенных видов и
способствовать их вытеснению.
. Инвазийные виды могут стать хищниками по отношению к аборигенным видам
и также способствовать их вытеснению.
. Инвазийные виды могут являться переносчиками возбудителей заболеваний
аборигенных видов или сами вызывать их заболевания.
Обозначенные формы воздействия, а также общее влияние инвазионного
процесса на структуру и функции экосистем фундаментальными научными и
прикладными организациями России изучены далеко не в полной мере. Особенно
досадно, что нет хорошей информационной поддержки этих работ. А она необходима
для оценки риска и разработки мер борьбы с чужеродными видами, наносящими ущерб
народному хозяйству, а также для создания прогнозов новых инвазий.
Изучение питания инвазионных видов позволит оценить функциональную роль в
сообществе аборигенного комплекса рыб. Изучение размерно-половой и возрастной
структуры популяции позволит выявить закономерности динамике численности,
тенденции, увеличения численности либо ее снижения и оценить пресс вселенцев на
кормовую базу водоема. Полученные данные необходимы для объективной оценки
биологических ресурсов водоемов с учетом не только комплексу промысловых, но и
непромысловых видов рыб.
Знание общей ситуации в стране, путей и механизмов инвазионного процесса
поможет разработать меры борьбы с чужеродными видами, трансформирующими
экосистемы и уничтожающими хозяйственно ценную продукцию.
Целью данной работы явилось изучение биологии ротана-головешки в водоемах
юга Западной Сибири.
В связи с поставленной целью были определены следующие задачи:
изучить места распространения и морфологию вида;
определить половую и возрастную структуру популяции;
изучить темп роста;
изучить питания рыб.
1. Обзор литературы
.1 Рыбы семейства головешковые
Рыбы семейства головешковые (Eleotrididae) входят в состав подотряда бычки (Gobioidei) отряд окунеобразные. Характерным
для рыб подотряда является наличие в спинном плавнике не более восьми слабых
колючек, образование у многих видов присоски в результате срастания брюшных
плавников, отсутствие у большинства видов плавательного пузыря. Это богатая
видами (≈600) группа небольших, главным образом морских прибрежных рыб,
обитателей преимущественно тропических, в меньшей степени умеренных вод. К
условиям жизни в пресных водах приспособилось небольшое число видов подотряда.
В пресных водах России из семейства головешковых обитает только два вида:
китайский элеотрис и головешка-ротан. Последний вид является случайным
акклиматизантом некоторых водоемов Сибири.
.2 Головешка-ротан - perccottus glenii Dybowski, 1877
.3 Распространение
Естественный ареал ротана - бассейн Амура, на юг до Владивостока и
полуострова Корея. В 1877 г. ротан был завезен в Европейскую часть России, в
1916 г. Его вселили в садковый пруд под Петербургом, где он успешно прижился.
Вторично завезен в 1948 г. в ряд прудов и озер Подмосковья. В конце ХХ в. Он
освоил многие водоемы бассейнов Балтийского, Белого, Каспийского и Черного
морей. В настоящее время встречается в мелких водоема Московской,
Ленинградской, Калининградской, Самарской и Нижегородской областей. В Волге
ротан расселился от верховьев до Иваньковского, Саратовского и Куйбышевского
водохранилищ, где активно питается аборигенными видами рыб. В 2001г. несколько
экземпляров ротана поймано в Онежском озере (Еловенко, 1985; Неелов, 1987; Рыбы
Подмосковья, 1988; Атлас пресноводных…, 2003; Бознак, 2004). Повсеместен ротан
в водоемах на юго-востоке Казахстана и на востоке Узбекистана (Пронин, Болонев
и др., 2005).
На территори Сибири ротан встречается в оз. Большое Миассово
(восточно-предгорный район Южного Урала) (Перескоков, 2004), в пойменных
водоемах р.Оми (Бабуева, 2001а), в озерах, имеющих сток в р. Тобол (Мухачев,
2002; Терентьев, Мухачев, 2006) и в ряде озер на западе Новосибирской области.
В некоторых из указанных озер ротан многочислен и является объектом
промыслового и любительского лова. Промысловым видом является он и в ряде озер
Челябинской и Курганской областей. Летом 1998 г. Несколько экземпляров ротана
было поймано в пойменных озерах Оби в 100 км ниже слияния Бии и Катуни
(Торопов, 2000). В настоящее время ротан встречается в пойменных водоемах на
всем протяжении Верхней Оби (Журавлев, Ломодуров и др., 2006).
В 1990 - 1992 гг. ротан впервые отмечен в ряде водоемов Томска и особенно
многочислен здесь в небольших, хорошо прогревающихся в летний период,
малопроточных прудах пригородной зоны, в пойменных озерах в низовьях р. Томи.
Помимо ротана, в этих водоемах обитают верховка, золотой и серебряный караси
(Петлина, Рябова, 2004). Известен ротан и в ряде водоемов Кемеровской области
(Острошабов, 2004).
В 1969 г. Ротан случайно завезен вместе с сеголетками сазана из бассейна
Амура в оз. Гусиное (бассейн Селенги). В настоящее время ротан населяет
практически все водоемы в окресностях Улан-Удэ, включая старицы селенги.
Северной границей распространения этого вида в Забайкалье является бассейн р.
Турка, где ротан в массе встречается на мелководных участках. В 1978 г. Ротан
впервые отмечен в устье Селенги и в Истокском и Посольском ссорах Байкала; в
настоящее время особенно многочислен на заросших участках протока и пойменных
озер дельты Селенги и соров Селенгинского мелководья. Освоил ротан и
высокоминерализованные (до 6,5 г/л) щелочные озера Оронгой и Сульфатное,
расположенные в долине р. Селенга (Пронин, Болонев, 2005). В Байкале в районе
Селенгинского мелководья ротан обитает до 30-метровой изобаты (Пронин, Селгби и
др., 1998), на некоторых других участках озера - до глубины 75 м (Скрябин,
1997). В последние годы ротан начал активно осваивать западную часть озера, где
регулярно встречается в бухте Большие Коты, в заливе Песчанка, близ устья р.
Голоустная (Литвинов, 1990; Демин, 2001; Болонев, Пронин и др., 2002а,б;
Аннотированный список…, 2004). Но в целом по бассейну Байкала численность этого
вида рыб или стабилизировалась или (по западному побережью) снижается, основная
причина чего - низкая температура воды в озере (Болонев, Пронин и др., 2002а,
б; Пронин, Болонев, 2005). Вылавливаются отдельные особи ротана в Ирткутском
водохранилище (Купчинский, Купчинская ,2001).
1.4 Возраст и размеры
Максимальная длина тела ротана в разных водоемах естественного ареала
колеблется от 14 до 25 см. Темп его роста в природных водоемах довольно низкий
и весьма изменчив. В естественном ареале особи старше 4+ встречаются крайне
редко.
По данным Яковлева (1925) для ротана из небольшого озерца в бассейне
озера Ханка, к концу первого года жизни мальки едва достигали 1 г (в среднем
0.8 г) и 3.5 см длины. Двухлетние имеют длину тела в среднем около 5.5 см и вес
около 4 г., трехлетки соответственно 7.7 см и 11.6 г, семилетний ротан имел
13.6 см и 53.5 г. Автор находит этому объяснение в большой плотности популяции
данного водоема - до 3-4 взрослых особей на 1 кв. м площади водоема.
Для озера Глубокое под Москвой Решетников (2001) указывает длину 250 мм и
массу 250 г. в возрасте около 10 лет.
По данным Кудерского (1982), в пруду у Сестрорецка под Петербургом в
октябре 1981 г. были представлены особи трех возрастных групп: 1+, 3+ и 4+.
В отличие от естественных водоемов, при разведении в аквариуме
(Спановская и др., 1964) ротан в первый год жизни может достигать 5-6 см длины.
Таким образом, потенциальные возможности роста у него велики. В естественных
водоемах Московской области отмечены значительные колебания длины тела ротана в
одном и том же водоеме, что авторы объясняют очень неравномерным ростом.
Увеличение неравномерности линейного и весового роста популяции ротана связано
с ухудшением условий жизни в исследованном пруду. В первые годы существования в
этом водоеме ротан был немногочислен и условия питания, а, следовательно, и
роста были благоприятны. Темп его роста был значительно выше, чем в р. Суйфун и
в Нижнем Амуре. Амплитуда изменчивости длины и веса разных возрастных групп
была примерно одинакова. В дальнейшие годы, с возрастанием численности ротана в
пруду, условия питания стали хуже, снизилась упитанность рыб, а изменчивость
темпа роста заметно усилилась. Темп роста основной части популяции (по данным
за 1961-1962 гг.) замедлился и приблизился к темпу его роста в бассейне р. Суйфун.
В то же время появились особи, растущие очень быстро, гораздо быстрее, чем в
1955-1956 гг. Увеличение неравномерности роста проявилось не только в
разделении на быстро- и медленнорастущих рыб, но и в усилении
разнокачественности линейного, весового роста и накопления запасных питательных
веществ в печени, количества жира на внутренностях в пределах каждой возрастной
группы быстро- и медленнорастущих рыб, особенно у трех- и четырехлеток, т. е. у
особей, составляющих основу популяции.
В Мордовии (бассейн Волги) ротан в возрасте 4 - 5 лет достигает длины
8-10 см. Предельная длина - 25 см, масса тела - 350 г. В оз. Тростном (пойма р.
Суры) был пойман экземпляр массой 350 г в возрасте 8 лет (Вечканов, 2000).
Линейный и весовой рот в новом ареале могут заметно варьировать в
зависимости от обеспеченности пищей. Так, показатели линейного и весового роста
ротана в озере Гусином под Москвой были высокими. Максимальный вес ротанов
достигал 260-270 г (Болонев и др., 2002), тогда как в материнском водоеме (р.
Амур) и Ивлевском рыбхозе Горьковской области максимальный вес старшевозрастных
рыб в возрасте 7 лет не превышал 60-70 г (Кудерский, 1980). Это свидетельствует
о том, что ротан нашел очень хорошие условия обитания с высокой обеспеченностью
пищей на мелководьях озера Гусиное.
В Амуре и Байкале ротан живет до 8+ и достигает 25 см длины и 300 г массы
(Скрябин, 1996). К концу первого года жизни вырастает до 3 - 4 см и 0,8 г,
второго года - 6 - 7 см и 4 г, третьего года - 9 - 11 см и 11 - 12 г,
четвертого года - 12 - 13 см и 13 - 15 г. В оз. Круглое (Алтайский край) ротан
в 2+ имеет в среднем 12 - 15 см длины и 62 - 146 г массы (Журавлев, Ломодуров и
др., 2006). В водоемах г. Томска ротан живет до 5+ лет; в 1+ самки достигают
длины 7 - 9 см и массы 11 - 27 г, в 5+ - в среднем 12 см и 94 г (Петлина,
Рябов, 2004). Для сравнения: в некоторых водоемах в бассейне р. Вычегда особи
ротана в 2+ имеют 6 - 10 см, в 4+ - 9 - 15 см длины (Бознак, 2004).
.5 Экология ротана-головешки
Головешка - ротан обычно обитает в стоячих водоемах: старицах рек,
зарастающих и заболоченных озерах, прудах и болотах. Неприхотлив к условиям
среды. Хорошо переносит дефицит кислорода в воде. Выдерживает почти полное
высыхание и промерзание водоемов, зарываясь в ил. Ведет оседлый образ жизни. За
добычей охотится из засады - густых зарослей подводной растительности. В
придаточных водоемах Амура в конце декабря ротан скапливается в большом числе в
полостях льда, заполненных воздушно-ледовой влажной массой. Полость в виде
полусферы имеет диаметр от 20 см до 2 м. От верхней кромки льда до полусферы 30
- 60 см. Температура в ней около 0 - 1 0С. Рыбы в таких полостях находятся в
состоянии оцепенении и будучи вынутыми слегка шевелятся. При помещении в воду
они быстро отходят и начинают активно плавать. В природе ротан пробуждается от
спячки в конце апреля. В водоемах европейской части России ротан в спячку не
впадает и активен в течение всей зимы (Еловенко, 1985; Рыбы Подмосковья, 1988;
Соколов, 2001; Атлас пресноводных…, 2003).
В водоемах Томска самки ротана созревают в 3 года при длине в среднем 10
см и массе 42 г. Нерест в течение года однократный и протекает с начала июня до
конца июля - начала августа. ИАП колеблется от 560 до 2459 икринок (Петлина,
Рябова, 2004). В Амуре половозрелым становится в 2 - 3 года при длине около 6
см. Самки выметывают до 1000 икринок. Нерест порционный и происходит в мае -
июле при температуре воды 15 - 20 0С. Икра откладывается на корневища растений,
коряги, камни, на нижнюю поверхность плавающих предметов и листья водной растительности.
Икринки удлиненной формы, размером 3,8 × 1,3 мм. К субстракту икринка
приклеивается с помощью клейких ворсинок, расположенных на одном из ее концов.
Икринки располагаются в один ряд, обычно близко к поверхности воды. При
вылуплении личинки имеют длину 5,0 - 5,5 мм. Поскольку плавательный пузырь
наполняется воздухом еще у эмбрионов в оболочке, личинки ротана не опускаются
на дно и способны плавать в толще воды в более благоприятных условиях дыхания.
Самец охраняет и икринки и личинок (Никольский, 1956, 1971).
В оз. Круглое (Алтайский край) ротан половозрелым становится в 2+; в
преднерестовый период в яичниках рыб хорошо различимы икринки двух генераций
диаметром 0,4 - 0,7 и 0,9 - 1,3 мм; ИАП, судя по незрелой икре, составляет 4210
- 9200 икринок (Журавлев, Ломодуров и др., 2006).
В Байкале ротан приступает к размножению в 2+ при достижении 7 см длины и
в 6 г массы, в оз. Гусиное - в 1+ при 4,5 см длины и 2 г массы. Способность к
воспроизводству сохраняется до 8+. Первая порция икры выметывается в водоемах
Селенги и сорах Байкала в течение июня, вторая - в конце июля - начале августа,
третья порция, в отличие от ротана из Амура, не созревает. В первой порции у
самок байкальского ротана в 2+ длиной 4 - 5 см насчитывается до 250 икринок, в
5+ длиной 10 - 12 см - до 5,4 тыс. икринок. Нередко икра откладывается на
попавшие в водоем предметы: различные трубы, домашнюю посуду, обувь и т.п. Как
и в Амуре, самцы охраняют икру и личинок (Скрябин, 1997).
В водоемах Амура на вторые сутки после выклева личинки ротана начинают
активно питаться мелкими, а затем и более крупными беспозвоночными планктона.
Взрослый ротан питается животными бентоса, реже - планктона, поедает икру и
молодь рыб, в начале лета охотно поедает головастиков. Активно хищничает ротан
во многих водоемах Европейской части России (Еловенко, 1985; Неелов, 1987; Рыбы
Подмосковья, 1988; Евланов, Козловский и др., 1998). В оз. Круглое (Алтайский
край) 5 из 10 просмотренных желудков ротана оказались пустыми, в остальных
обнаружены остатки рыб, в том числе собственная молодь (Журавлев, Ломодуров и
др., 2006). В озерно-речной системе Байкала молодь ротана питается
зоопланктоном, особи в 1+ и старше - преимущественно личинками бентосных
беспозвоночных. По мере увеличения размеров ротана в его пище все больший
удельный вес занимают молоди и икры рыб. У взрослых рыб суточный рацион состоит
нередко пости на 100% из рыб, в том чмсле молоди омуля. Сам ротан играет
заметную роль в питании таких рыб Байкала, как щука, окунь, а также в питании
многих видов птиц (Демин, 2001).
В связи с высокими приспособительными свойствами, ротан может широко
распространиться как в самом Байкале (в его мелководной зоне), так и в других
водоемах в бассейне этого озера. В 1990-е гг. на отдельных участках Посольского
сора плотность этой рыбы достигала 95 экз./м2 (Скрябин, 1997). Ущерб, наносимый
ротаном в Байкале в результате поедания им икры и молоди рыб-аборигенов, а
также в результате конкуренции с байкальскими рыбами на почве питания,
оценивался в 1980-е гг. в 2200 - 5000 ц рыбной продукции (Скрябин, 1997; Демин,
2001). Такая же ситуация в связи с увеличением численности и расширением ареала
ротана может сложиться в недалеком будущем и во многих водоемах Западной
Сибири.
.6 Особенности роста и образование эко-морфологических форм
Как отмечено выше, Спановская с соавторами (1964) заметила у ротана из
Европейской части СССР дифференциацию по темпу роста и отнесли ее за счет
последствий акклиматизации. Крысанов и Еловенко (1981) также обратили внимание
на возрастающую изменчивость этих рыб при длительной изоляции водоема. Однако с
такой же дифференциацией авторы столкнулись и при изучении ротана из
естественных дальневосточных водоемов. В отличие от популяций пойменных озер
рек Амур, Уссури, Мо, Раздольная, в тех водоемах, которые не имеют связи с
рекой много лет, как, например, глиняные карьеры в Хабаровске, изолированные
озера Европейской части России и Восточного Казахстана, наблюдается четкий
экологический диморфизм. Две формы, которые авторы условно назвали светлой и
темной, различаются статистически достоверно по 12 из 45 изученных пластических
признаков. Ротан светлой быстрорастущей формы питается большей частью рыбой. Он
имеет более вытянутую форму тела, мощную голову, конечный рот, у них более
развиты и сдвинуты назад анальный и второй спинной плавники. Ротан темной формы
питается всеми группами водных беспозвоночных. Дифференциация становится
заметной к концу первого года жизни, при достижении III стадии зрелости гонад.
Соотношение численности этих форм в популяции меняется по мере роста. Рыбы
светлой формы в размерной группе 50-100 мм так же редки, как и темные среди
экземпляров длиннее 200 мм.
.7 Питание и пищевое поведение
В водоемах Дальнего Востока ротан питается личинками насекомых
(Chironomidae, Aedes, Chaoborus, Odonata); в меньшем количестве,
преимущественно у молоди, встречаются ракообразные Cladocera и Copepoda, а у
крупных - мелкие рыбы (Яковлев, 1925).
Наиболее подробно питание ротана в естественном ареале исследовано
Синельниковым (1976) в пойменных водоемах бассейна р. Раздольная (Суйфун).
Показано, что спектр питания ротана исключительно широк. В пище отмечено 76
компонентов. Ветвистоусые рачки представлены 18 видами, среди которых
преобладают Chydorus sphaericus, Eurycercus lamellatus, Simocephalus
elizabethae. Из веслоногих раков отмечено 8 видов (численно преобладают
ювенальные стадии). Из высших ракообразных ротан предпочитает молодь речного
рака и пресноводных креветок. Существенную роль в питании играют личинки
различных водных насекомых: Odonata, Ephemeroptera, Trichoptera, Coleoptera,
Diptera (Chirinomidae, Heleidae, Tabanidae), причем ведущей группой как по
численности, так и по биомассе, являются личинки хирономид. Молодь ротана
хищничает редко, взрослые кормятся преимущественно рыбой: молодью ротана, горчаков,
гольянов.
Так как характер питания ротана с возрастом существенно изменяется, этот
автор (Синельников, 1976) счел возможным выделить 7 размерных групп: 5 мм,
8-11, 12-25, 26-40, 41-60, 61-100, 101-217 мм.
У личинки ротана длиной 5 мм (с желточным мешком) пища не обнаружена.
Рыбы размером 8-11 мм - вполне оформившиеся и активно питающиеся мальки. Пищей
им служат ветвистоусые, веслоногие раки и личинки хирономид. По числу особей и
по весу преобладают ветвистоусые раки и особенно Chydorus sphaericus. Из веслоногих
раков встречаются в основном неполовозрелые формы. Личинки хирономид
представлены очень мелкими особями, поэтому хотя они и многочисленны (58%), вес
их не превышает 22,5 %. Единично отмечаются ракушковые раки, личинки поденок,
встречаются водоросли и детрит.
У молоди размером 12-25 мм пища более разнообразна. Ветвистоусые
встречаются еще в заметном количестве, однако, по весу (48,4 %) преобладающей
группой становятся личинки хирономид. У рыб этой размерной группы в желудках
впервые обнаружены остатки рыбной пищи. Отмечен случай, когда 18-миллиметровый
ротан заглотил рыбу того же вида длиной 7 мм. Этот факт свидетельствует не
только о наличии каннибализма, но и возможности перехода ротана уже на ранних
стадиях к хищничеству. Тем не менее, хищничество для ротанов этой размерной
группы скорее не правило, а исключение, так как отмеченные случаи единичны и,
как правило, рыбой питаются только половозрелые особи (свыше 50 мм).
Молодь длиной 26-40 мм питается в основном личинками различных водных
насекомых, среди которых преобладают личинки хирономид. Часто отмечаются имаго
насекомых. Роль низших ракообразных значительно снижается: они составляют по
весу около 30 %.
Рыбы длиной 41-60 мм, среди которых уже встречаются половозрелые особи,
почти не питаются низшими ракообразными и полностью переходят на более крупные
объекты - личинок и имаго насекомых. В пище у них появляются пресноводные
креветки, встречаются остатки рыб.
Ротан размером 61-100 мм питается преимущественно личинками водных
насекомых. Хирономиды используются им в меньшей степени (34,1 %). Заметную роль
в питании начинают играть рыбы (11,1 %).
На основании полученных данных в питании ротана можно выделить три
периода: планктоноядный (8-11 мм), бентосоядный (12-100 мм) и хищный (свыше 100
мм).
И в приобретенном ареале спектр питания ротана очень широк - от циклопов
и дафний до рыб, лишь немного уступающих ему по размерам. В пище ротана из
бассейна Верхней Волги было обнаружено 100 компонентов. При недостатке корма в
водоемах крупные особи ротана поедают более мелких, как и в естественном ареале
(Бандура, 1979; Болонев и др., 2002; мн. другие). Это является одним из
факторов, позволяющих его популяциям существовать в любых биоценозах и
поддерживать численность на постоянном уровне. Питаясь мальками ценных видов
рыб, ротан способен полностью вытеснить их из водоема и стать в нем
единственным видом.
По данным Спановской и др. (1964), питание ротана в пруду в Подмосковье в
первые годы после индродукции по качественному составу мало отличалось от пищи
в дальневосточных водоемах. Молодь длиной до 40 мм питалась преимущественно
ракообразными (Cladocera, Copepoda), а более крупные, помимо Cladocera (главным
образом, Daphnia cucullata), потребляли личинок насекомых (Chironomidae,
Ephemeroptera, Odonata, Coleoptera, Chaoborus, Heleidae, Trichoptera,
Hemiptera). В 1961 - 1962 гг. качественный состав пищи ротана по сравнению с
1955-1956 гг. сильно изменился. Процент Cladocera у рыб размером 40-60 мм
сохранился довольно высокий, но важное значение приобрели Chydoridae (Chydorus,
Camptocercus) - рачки зарослей; Daphnia и Bosmina, населяющие толщу воды,
встречались лишь у более крупных особей из этой группы и у рыб длиной 60-105
мм. Из пищи почти совсем исчезли личинки Odonata, Coleoptera, Hemiptera, сильно
уменьшилось значение личинок Chironomidae и изменился их видовой состав. В
1955- 1956 гг. преобладали личинки Tanytarsus, Polypedilum, Einfeldia gr.
carbonaria, Limnochironomus gr. nervosus, а в 1961-1962 гг. чаще встречались
личинки Orthocladiinae (Cricotopus gr. silvestris, Psectrocladius gr.
psilopterus) и Corynoneurinae, приуроченные к обрастаниям. Только с 1961 года
ротан в данном пруду стал потреблять собственную молодь. Каннибализм отмечен у
особей, достигших длины 45 мм. У рыб длиной свыше 60 мм более половины съеденной
пищи принадлежало собственной молоди. Кроме того, ротан поедал головастиков,
которых рыбы вообще потребляют редко.
Переход на питание собственной молодью и головастиками авторы (Спановская
и др., 1964) связывают с резким обеднением фауны крупных беспозвоночных (жуков,
их личинок, личинок стрекоз), отличающихся от планктонных форм низким темпом
воспроизводства, и с полным выеданием карася на стадии икры, личинки и молоди.
Питание личинками и взрослыми особями амфибий описано для ротана из
Подмосковья (Мантейфель, Решетников, 1997, 2001; Решетников, 2001) и
Закарпатья. Фрагменты личинок Вufo bufo были обнаружены в кишечниках у 6
экземпляров (3 самок и 3 самцов) из 31, имевших абсолютную длину от 100 до 161
мм. В среднем в желудках ротанов, съевших личинок жаб, находились по 4 ± 1.41
В. bufo (от 1 до 10 шт.). Ротан поедает личинок двух видов тритонов, Triturus
cristatus и T. vulgaris, и взрослых особей первого (Решетников, 2001), а также
личинок и взрослых особей T. dobrogicus.
Для водоемов Мордовии (бассейн Волги) также показано, что
многогенеративная популяция ротана использует практически всю кормовую базу
водоема. Например, в пойменных озерах реки Суры эта рыба потребляла до 70
различных представителей беспозвоночных, включая моллюсков и собственную молодь
в стадии личинки. За несколько лет сверхплотная популяция вида была способна
опустошить всю фауну водоема. Например, в оз. Пыжовка (сурская пойма), где
сформировалась мощная популяция ротана, было обнаружено только 12 форм
беспозвоночных вместо 40- 60 обычных представителей в таких озерах.
В бассейне Байкала в составе пищи ротана в различных водоемах
зарегистрировано около 100 видов кормовых организмов из 57 родов и 30 семейств
с наибольшим разнообразием личинок и куколок хирономид (28 видов) и ручейников
(Пронин и др., 1998). Во всех водоемах ротан питается животными кормами и
характеризуется как эврифаг. Молодь ротана в значительном количестве потребляет
зоопланктон - мелких животных, обитающих в толще воды. От года до
четырехлетнего возраста ротан - преимущественно бентофаг (питается организмами,
живущими на дне водоема) и частично хищник. Старшие возрастные группы (на
пятом-шестом году жизни) в значительном количестве потребляют молодь рыб,
особенно карповых (Болонев и др., 2002).
В рыбоводных прудах, по-данным Еловенко и Климовой (1983) и Еловенко
(1984), молодь ротана в первую неделю жизни питается фитопланктоном, затем
переходит на ветвистоусых и веслоногих ракообразных. При достижении рыбами
длины 12-15 мм преобладающей группой становятся личинки хирономид, которые
доминируют в желудках ротанов длиной до 110 мм. Видовой состав личинок
хирономид, как правило, представлен плавающими формами. Рыбы длиной до 40 мм,
кроме того, поедают моллюсков, личинок поденок, жуков, других двукрылых,
стрекоз (во время линьки), ручейников. Доля ракообразных в пищевых комках
сходит на нет. У особей длиной 20 мм в желудках начинает появляться икра и
личинки рыб, в том числе и своего вида; доля рыбной пищи возрастает по мере
роста и у особей длиной свыше 110 мм имеет преимущественное значение. Личинки
хирономид уступают по массе рыбной пище. Среди рыб (в рыбоводных прудах) ротаны
предпочитают заглатывать невоооруженных: гольянов, верховку, пескаря, горчака,
щуку, карася, собственную молодь по сравнению с ершом, окунем, щиповкой,
карпом. По данным Залозных (1982) для ротана из прудов Илевского рыбхоза, пища
особей старших возрастных групп до 97% состояла из рыбных объектов, причем в
конце лета и осенью до 82% составляла собственная молодь.
Потребление кормовых объектов, сильно отличающихся по размерам, характеру
поведения, возможно потому, что ротану свойствен ряд морфологических
особенностей, характеризующих его как активного ловца и потенциального хищника.
Рот у ротана большой (отсюда его название); на нижнечелюстных, предчелюстных
костях, на нижних и верхних глоточных костях, на верхних элементах двух
последних жаберных дуг есть зубы. Жаберные тычинки (9-10 шт.) бугорковидные,
покрытые собранными в отдельные пучки щетинками, помогают удерживать добычу в
глотке. По мере роста увеличивается относительный размер головы, удлиняются
челюсти, и это облегчает захват крупной добычи. Махлин (1960) указывает, что
ротаны в аквариуме охотно захватывают гуппи, молодь карася. Результаты
аквариальных наблюдений по избирательности ротана к разному корму показывают,
что при наличии достаточного количества различных крупных беспозвоночных
(личинки стрекоз, жука-плавунца) ротаны размером около 60 мм гораздо охотнее
питаются ими, чем рыбой. Более крупные особи переходят на питание рыбой, но и у
них степень избирательности по отношению к крупным беспозвоночным остается
очень высокой.
По мнению Спановской и др. (1964) и Еловенко (1984), ротан не питается
молодью своего вида, если достаточно другого корма. Питание беспозвоночными,
по-видимому, более свойственно этому виду; переход на питание собственной
молодью, головастиками объясняется неблагоприятными условиями откорма в связи с
возросшей численностью популяции и ухудшением состояния кормовой базы.
Яковлев (1925) описывают "охоту" ротана в аквариуме. Добычу
замечает быстро и издали, но никогда не бросается на нее сразу, а начинает
подплывать постепенно и с остановками, ровным и спокойным ходом по прямой
линии, работая только грудными плавниками, не шевеля хвостом и не отрывая от
добычи глаз; только подойдя на короткое расстояние, схватывают добычу быстрым и
резким движением.
.8 Половой диморфизм
Самцы в период размножения имеют весьма выраженные внешнеморфологические
отличия от самок. Они интенсивно черного цвета и с большим "вздутием"
кожного покрова на верхней части головы. На боках тела четко выделяются
беспорядочно разбросанные яркие голубовато-зеленые пятнышки. Такого же цвета
пятнышки, но расположенные ровными горизонтальными рядами, имеются на спинных и
анальном плавниках. Сильное "вздутие" на голове напоминает жировую
подушку самцов некоторых видов сем. Cichlidae. Оно начиналось под кожей от
межглазничного пространства и, заходя на спину, оканчивается вблизи первого
спинного плавника. Когда самец заканчивает охранять кладку, окраска бледнеет, а
вздутие на голове исчезает, голова его постепенно приобретает обычную форму, а
тело - зеленовато-серую (несколько более темную, чем у самок) окраску.
.9 Нерест
Во время нереста и охраны кладки самец не питается. Он проявляет при этом
агрессивное поведение. На помещаемых в аквариум взрослых особей своего вида
(как самцов, так и самок) он набрасывается, ударяя их головой и пытаясь
прогнать со "своей" территории. Нерестовые игры довольно
продолжительные и длятся несколько дней (Болонев и др., 2002)
Икра откладывается на нижнюю поверхность плавающих предметов (досок и
т.п.) и растений (Залозных, 1984; Козловский, 2001), а также на камни и другие
предметы, лежащие на дне, вплоть до консервных банок (Болонев и др., 2002). Во
время инкубации кладка охраняется самцом. В случае гибели самца икра обычно на
вторые сутки уничтожается другими рыбами (Залозных, 1982, 1984). Самец
обмахивает кладку грудными плавниками, поэтому икринки хорошо омываются и
зародыши не испытывают недостатка в поступлении кислорода. Кроме того, самец
яростно отгоняет от кладки всех, бросается даже на руку человека. Однако, ротан
может поедать свою молодь после вылупления (Болонев и др., 2002).
.10 Конкуренты и хищники
Значительно ограничивают численность ротана хищники. На Амуре таких ротаном
питаются более 10 видов рыб, в том числе сомы-косатки и змееголов. Полагают,
что основным ограничителем численности ротана является змееголов (Болонев и
др., 2002).
Еловенко и Климова (1983) исследовали пищевые взаимоотношения ротана с
гидробионтами в рыбоводных прудах Ханкайского рыбокомбината, Хабаровского
рыбхоза (естуственный ареал) и рыбокомбината "Нара" Московской
области. В прудах ротаном питаются амурский и европейский сомы, щука, амурская
щука, косатки, змееголов, европейский окунь и ерш. Среди них наиболее
интенсивно ротан выедается змееголовом и окунем. Икра и молодь ротана
уничтожается клопами: Notonecta glauca, Nepa cinerea, Naucoris cimicoides,
Ranatra linearis, Sigara hellensii, S. falleni, Micronecta griseola и др.,
жуками Dytiscus marginalis, Hibius ater, личинками стрекоз Lestes sponea и L.
nympha. Среди хищных беспозвоночных наибольший ущерб потомству ротана
причиняется клопами, которые, в свою очередь, в незначительных количествах
поедаются взрослыми экземплярами ротанов. В вырастных прудах молодь ротана
является конкурентом в питании многих видов бычков, ерша, окуня, верховки,
гольянов, вьюна, щиповки. Горчака и других видов рыб по личинкам хирономид и
ракообразным, а среди разводимых рыб - с сеголетками карпа, карася, амурского сазана
и амурской щуки.
Пищевые взаимоотношения ротана с местными байкальскими видами рыб
довольно напряженные. Особенно наглядно это проявляется в водоемах дельты реки
Селенги, где по сходству пищи наибольший антагонизм наблюдается с язем (до 90
%), карасем (81,2 %) и плотвой (67,3 %), в меньшей степени с ельцом (49,4 %)
(Пронин и др., 1998).
Шляпкин и Тихонов (2001) исследовали 17 водохранилищ в Ярославской
области, заселенных ротаном. Естественные рыбные сообщества в них включали от 1
до 7 видов. В результате интродукции ротана, в сообществах с небольшим числом
видов ротан стал доминирующим видом. Сообществах с исходно могокомпонентной
структурой включающей хищников (щуку, окуня и др.) гораздо более устойчивы к
появлению вселенца (Еловенко, 1980; Кудерский, 1980).
По данным Залозных (1982) для Горьковской области (бассейн Волги), в
заливах реки Сарма и небольших водоемах, периодически соединяющихся с рекой,
ротан встречается очень редко, несмотря на то, что ежегодно в эту реку из
выростных прудов Илевского рыбхоза попадает масса молоди. Также случайный
характер носят встречи ротана в старицах и пойменных озерах реки Оки, регулярно
затапливаемых паводковыми водами, где хищники - окунь и щука - являются
обычными компонентами ихтиофауны. В то же время в изолированных от рек
небольших водоемах с часто возникающим дефицитом кислорода и преимущественно
плотвично-карасевыми ихтиоценозами ротан бывает доминирующим видом.
.11 Воздействие на аборигенные сообщества
Результаты исследования (Решетников, 2001) показывают, что присутствие
ротанов в малых водоемах ведет к существенному уменьшению разнообразия видов и
обилия личинок амфибий и беспозвоночных, питающихся этими личинками. Однако
разные виды амфибий в разной степени восприимчивы к воздействию Perccottus glenii. В водоемах, колонизованных
ротаном, как правило, не могут успешно размножаться тритоны Тriturus cristatus,
Т. vulgaris, лягушки Rana temporaria, R. arvalis и R. lessonae. Ротаны способны
нарушать нормальное развитие нерестового поведения тритонов обоих видов,
поедать взрослых Т. vulgaris и личинок обоих видов. Лягушки трех видов не
избегают нереститься в водоемах, колонизованных ротаном, но их личинки активно
поедаются ротанами и в большинстве случаев полностью уничтожаются ими до
метаморфоза. Жабы В. bufo успешно размножаются в заселенных ротанами водоемах.
Личинки этого вида амфибий сравнительно мало съедобны для Perccottus glenii и в массе достигают стадии
метаморфоза в таких водоемах. Возможно, что условия развития личинок В. bufo
даже улучшаются после заселения водоемов ротанами (Решетников, 2001). В малых
водоемах, населенных ротаном, автором отмечалось от 0 до 2 видов амфибий, в то
время как в водоемах без ротана этот показатель варьировал от 0 до 5. Выявлена
достоверная отрицательная корреляция между присутствием ротанов и разнообразием
видов (N=22; r = - 0.4619; p = 0.03), а также обилием (N=22; r = - 0.4455;
p=0.038) амфибий в 1997 г. Сходные результаты получены при анализе связи обилия
ротанов с разнообразием (r = - 0.4969) и обилием (r = - 0.4792) амфибий.
В ряде водоемов ротан может полностью выедать личинок тритонов. По
наблюдениям других авторов для бассейна Дуная, T. dobrogicus перестал
встречаться в некоторых водоёмах в окрестностях Чопа, Батево и Мукачево после
появления в них этого вида рыбы (в 1996 ротана-головёшки ещё нет, а в 2000 году
он уже многочислен).
Обнаружена (Решетников, 2001) также отрицательная корреляция между
присутствием в водоемах ротанов и разнообразием видов беспозвоночных (r =
-0.5528; N= 18; р - 0.017). В водоемах, заселенных ротаном, не встречались или
встречались крайне редко взрослые жуки сем. Dytiscidae и их личинки, жуки
Hydrous sp., личинки стрекоз Aeschna cyanea, Somatochlora aenea и Erythromma
viridulum, пауки Dolomedes sp., пиявки Haemopis sanguisuga. Эти виды были
отмечены в некоторых других водоемах, в которых нет ротана. Однако
клопы-гладыши Notonecta glauca в массе населяют один из водоемов с Perccottus glenii. Из моллюсков в одном из
водоемов, населенном ротанами, остаются многочисленными крупные прудовики Lymnea
stagnalis.
Пронин (1982) считает, что вселение в бассейн Байкала ротана, элодеи
канадской, рипуса, пеляди и завезенных с акклиматизантми ихтиопаразитов следует
рассматривать как биологическое загрязнение озера. Биологическое загрязнение
крупных водоемов практически не ограничено во времени. Специально или случайно
акклиматизировавшийся вид будет эволюционировать вместе с экосистемой. В другой
публикации, посвященной ротану в Байкале, специально проанализировали
взаимоотношения между ротаном и другими рыбами. Они показали, что присутствие
ротана снижает численность плотвы и ельца, поскольу он сильно конкурирует с
ними за пищевые ресурсы. Однако, по мнению этих авторов, наибольшую опасность
представляет выедание ротаном икры других видов, размножающихся в прибрежных
мелководьях, отмелях и разливах.
Однако, по мнению, при умеренной и низкой численности в заморных
водоемах, где прочие хищники погибают, ротан может замещать окуня, выполняя его
функцию ограничения хищных беспозвоночных и улучшая тем самым условия
существования молоди других рыб.
.12 Возможный контроль
рыба головешковый ротан водоём
В бассейне оз. Байкал реальным регулятором численности ротана могут быть
щука и окунь. Они уже проявляет себя в этом качестве в водоемах дельты р.
Селенги, однако численность их низка. Повышение численности щуки вполне
возможно путем разведения на временных рыбоводных пунктах и существующих
рыборазводных заводах. На естественное же воспроизводство щуки существенное
влияние оказывает динамика уровня воды в Байкале в период нереста (апрель-май).
В свою очередь на естественные колебания уровня Байкала оказало влияние
зарегулирование его плотиной ГЭС (1958). До этого минимальный уровень воды в
его годовой динамике приходился на апрель. После зарегулирования и при поднятии
общего уровня на 1 метр сезонный минимум переместился на май. Генетически же
детерминированный годовой физиологический цикл щуки остался неизменным.
Снижение уровня воды в мае приводит к значительному снижению нерестовых
площадей. Поэтому режим работы Иркутской ГЭС должен обеспечивать минимальный
уровень воды в апреле, как и до.
На основании проведенных опытов (Залозных, 1984) было установлено, что из
всех видов сорных рыб ротан является наиболее стойким к действию хлорной
извести и аммиачной воды. 100%-ная гибель ротанов наступает при концентрации
хлорной извести 0.3 г на 1 л воды и экспозиции не менее 6 часов. В реальных
условиях это можно сделать лишь в небольших по площади лужах. При обработке
бочагов и водосборных каналов большой эффект дает применение аммиачной воды.
При температуре воды 7-8 градусов и pH 9.0 100%-ная гибель ротанов наступала
при разведении 1 мл аммиачной воды на 1 л уже через 1 час 5 минут.

Рис. 1. Головешка-ротан - perccottus glenii Dybowski, 1877
2. Место и условия проведения работы
.1 Физико-географический очерк
Площадь Новосибирской области 178 тысяч кв. км. или 1% территории России.
Протяжённость области с запада на восток более 600, а с севера на юг до 400
километров.
География юго-западной Сибири. Западно сибирская равнина одно из немногих
физико-географических мест, границы которого отчетливо выражены в рельефе. Ее
рубежами на западе являются восточные предгорья Урала. На востоке равнина
ограничена уступом Енисейского кряжа и Среднесибирского плоскогорья, вдоль
которого заложилась долина реки Енисей, на севере омывается водами Карского
моря. южная часть равнины уходит за пределы России в Казахстан и лишь на
крайнем юго-востоке граничит с Алтаем.
Рельеф Западной Сибири обусловлен геологическим развитием, тектоническим
строением и влиянием разнообразных экзогенных рельефообразующих процессов.
Основные орографические элементы находятся в тесной зависимости от
структурно-тектонического плана плиты, хотя длительное мезокайнозойское
прогибание и накопление мощной толщи рыхлых отложений в значительной мере
снивелировали неровности фундамента. Малой амплитудой неотектонических движений
обусловлено низкое гипсометрическое положение равнины. Специфические черты
природы Западной Сибири, определяющие ее своеобразие и уникальность среди
других физико-географических стран - довольно однообразный, слабо пересеченный
рельеф с малыми абсолютными и относительными высотами, исключительная
заболоченность и ярко выраженная широтная зональность природных условий. Южная
часть равнины - наиболее освоенные и обжитые районы Сибири, где природа в
значительной мере изменена хозяйственной деятельностью.
Климат Западной Сибири - континентальный, достаточно суровый. Он более
суров, чем климат Русской равнины, но мягче остальной территории Сибири
(Челазнова,2003).
По
территории Новосибирской области протекает около 380 рек, в том числе одна из
крупнейших рек мира - Обь, берущая начало в ледниках Алтайских гор и впадает в
Северный Ледовитый океан. Из рек области стоит отметить также Иню,
<#"650078.files/image002.jpg">
Рис.2. Спектр питания двухлетних самок ротана в Бердском
пруду
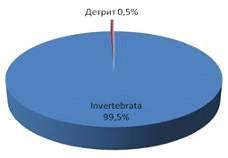
Рис. 3. Состав пищевого комка самок ротана из Бердского пруда
Спектр питания самцов ротана в этом же водоеме в отличие от самок был
менее широк. Но также основную массу рациона составляли двукрылые dipteral - 80% и ручейники trichoptera - 14%. Малую долю рациона составляли
личинки жуков coleoptera 2% и стрекозы odonata 1% (рис 4).
Пищевой комок самцов выловленных из Бердского пруда состоял на 100% из
беспозвоночных invertebrate.
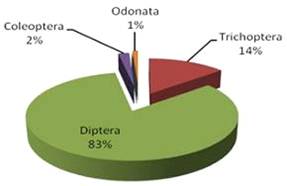
Рис. 4. Спектр питания двухлетних самцов ротана в Бердском пруде
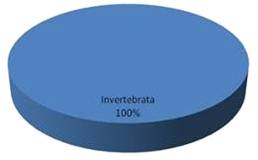
Рис. 5. Состав пищевого комка самцов ротана из Бердского пруда
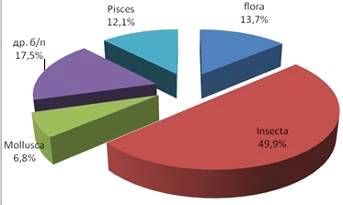
Рис. 6. Спектр питания ротана в оз. Улугуль
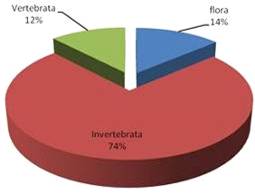
Рис. 7. Состав пищевого комка ротана в оз. Улугуль
При исследовании желудочно-кишечного тракта ротана пойманного в озере
Улугуль было выявлено, что 49,9% занимали насекомые insectа, из них клопов heteroptera - 6,8, двукрылых dipteral - 32,6, личинки жуков coleopteran - 10,5%. Также в спектр питания
входили моллюски gastropoda (6,8%);
рыбы pisces (12,1%); макрофиты (1,1%), семена
растений (13,7%) и прочие беспозвоночные 17,5% (рис.6).
Общий пищевой ком подразделялся на три составляющие: беспозвоночные invertebrate 74%; флора flora 14% и позвоночные vertebrata 12% (рис. 7).
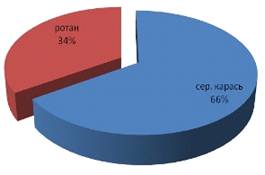
Рис. 8. Спектр питания ротана из р. Алей
Рис. 9. Состав пищевого комка ротана из р. Алей
Пищевой комок ротана из реки Алей на 100% состоял из позвоночных и был
представлен исключительно рыбой (серебряным карасем 66% и ротаном 34%).
Из такого сравнительно не одинакового питания в разных водоемах, размеры
ротана значительно отличаются. В р. Алей, где кормовая база представлена только
рыбой, ротан имеет более короткое (взбитое) широкое тело, а в Бердском
пруду и оз. Улугуль, где спектр питания представлен в основном беспозвоночными,
рыба имеет более вытянутое и худое тело.
.4 Темп роста ротана в разных половозрастных группах и
водоемах
В первые годы жизни темп роста наиболее интенсивен, от годовалого до
двухлетнего возраста самки ротана из Бердского пруда увеличили свою массу на
98%, и весили 6,92 г, а длина тела на 26%, и составляла 62,6 мм. Во второй год
жизни за лето прирост увеличился еще в 1,5 раза, и составил 15,02 г; длина
возросла на 41% (82,57 мм). С трехлетнего возраста до трехгодовалого самки
ротана набрали порядка 26,13 г (174%), и весили 41,15 г, длина тела
увеличивалась на 27,78 мм (33,6%), и составила 110,35 мм. Существует прямая
взаимосвязь между массой тела и ее длиной, при этом r= 0,97; 0,85; 0,95 для годовиков, двухгодовиков и
трехгодовиков, соответственно.
У самцов наблюдалась примерно такая же динамика роста; до трехгодовалого
возраста интенсивно набирали вес. За второе и третье лето в жизни увеличили
свою массу в 3 раза, которая составила 8,7 и 13,5 г, соответственно, а длина
тела возросла на 20 мм, это 40% (67,05 мм) и 37% (74,68 мм), соответственно.
Пик роста приходился на период жизни от трехлетнего возраста до трехгодовалого,
масса увеличилась в 5 раз, и стала 67,3г, а длина на 70% (127,25 мм). Далее
рост остался на одном уровне. Существует прямая взаимосвязь между массой тела и
ее длиной, при этом r= 0,93; 0,94;
0,92 для двухлеток, трехлеток и четырехлеток, соответственно.
Темпы роста и размеры, которых достигает рыба, в различных водоемах
неодинаковы и зависят в основном от гидробиологических условий среды, которые,
в свою очередь, связаны с кормовой базой, физико-химическими и климатическими
особенностями водоема. Так, при сравнении двух выборок ротана из реки Алей и
Бердского пруда, можно заметить, что более благоприятные условия в первом
водоеме; масса двухлеток в нем в 2,5 раза превосходила массу рыб из Бердского
пруда, и составляла 27,3 г, длина больше на 50% (96,14 мм). Трехлетки ротана из
реки Алей были крупнее своих сверстников из Бердского пруда в таких же
соотношениях, что и двухлетки (75,66 : 108,48 мм и 13,65 : 47,9 г). У
четырехлеток длина тела в обоих водоемах примерно на одном уровне 113,8 мм
(Бердский пруд) и 117,2 мм (р. Алей), что касается массы, то ротаны из р. Алей
больше на 61%, и весили 63,08 г.
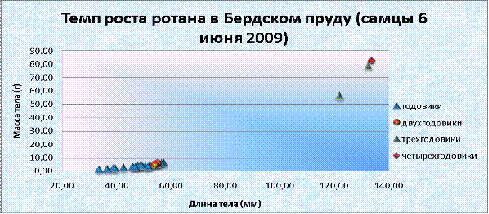
График 1. Темп роста самцов ротана в Бердском пруду (6 июня)
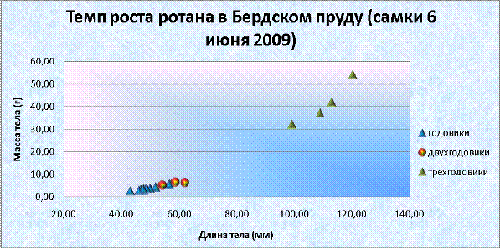
График 2. Темп роста самок ротана в Бердском пруду (6 июня)
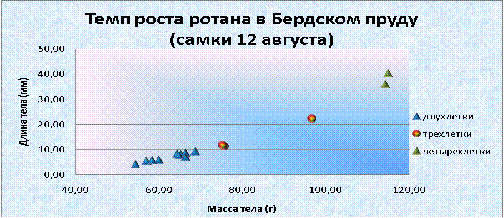
График 3. Темп роста самок ротана в Бердском пруд (12 августа)

График 4. Темп роста самцов ротана в Бердском пруду (12 августа)
4.5 Половозрастная структура популяции ротана-головешки
Из таблицы 10 видно, что из выборки сделанной 6 июня в Бердском пруду,
соотношения полов годовиков равно 63% самцов к 37% самок, двухгодовиков - 40 :
60; трехгодовиков - 33 : 67. Соотношение полов из улова от 12 августа равно 47
: 53; 83 : 17; 50 : 50, двухлеток, трехлеток и четырехлеток, соответственно.
При вылове ротана из реки Алей соотношение полов составило: двухлетних самцов
60% и самок 40%; трехлеток - 40 : 60; четырехлеток - 39 : 61; пятилеток - 33 :
67.
Таблица 10. Половозрастное соотношение самок к самцам
|
Возраст
|
Бердский пруд
|
р. Алей
|
|
|
6 июня
|
12 августа
|
|
|
1
|
30 (19♂;
11♀)
|
15 (7♂; 8♀)
|
5 (3♂; 2♀)
|
|
2
|
5 (2♂; 3♀)
|
30 (25♂;
5♀)
|
25 (10♂;
15♀)
|
|
3
|
6 (2♂; 4♀)
|
6 (3♂; 3♀)
|
70 (27♂;
43♀)
|
|
4
|
1 (1♂)
|
|
3 (2♂; 1♀)
|
июня из Бердского пруда было выловлено всего 42 экземпляра, из них
годовиков 71,4%; двухгодовиков - 11,9; трехгодовиков - 14,3 и четырехгодовиков-
2,4%.
августа улов состоял из 51 экземпляра рыб, из них двухлеток было 29,4%;
трехлеток - 58,8 и четырехлеток 11,8%.
Из р. Алей всего было поймано 103 рыбы: из них двухлеток 4,9%; трехлеток
-24,3, четырехлеток - 68 и пятилеток 2,9%.
5.
Безопасность жизнедеятельности
Рыбоводство - достаточно сложная отрасль сельского
хозяйства со специфическими производственными процессами и видами оборудования.
Многие работы выполняются на воздухе, на водоемах, что заставляет уделять
особое внимание вопросам безопасности.
В период пропуска паводков (до их начала) проводят
осмотр всех гидротехнических сооружений, проверяется их состояние и готовность
к эксплуатации, очищаются все водопропускные отверстия от снега, льда и
посторонних предметов.
До пропуска паводка в рыбоводном хозяйстве
составляется план оргтехмероприятий, учитывающий все возможные осложнения,
связанные с паводком и ледоходом, организуются рабочие бригады, которые
проходят инструктаж по безопасности труда, знакомятся с планом мероприятий по
пропуску паводковых вод, получают все необходимые материалы и спасательные
средства. В период паводка на водоемах устанавливается круглосуточное
дежурство, ведется постоянный надзор за состоянием плотин, дамб, водопропускных
сооружений.
Эксплуатируемые на рыбоводных предприятиях
грузоподъемные машины и устройства отвечают всем требованиям правил
Госгортехнадзора России: это относится и к подготовке обслуживающего персонала
- машинистов кранов, стропальщиков.
Лица, обслуживающие плавсредства, должны уметь плавать
и знать приемы спасения утопающих.
Облов рыбы в спускных и не спускных прудах можно
проводить только при высоте волны не более 0,5м. Применяемые для этого
плавсредства (лодки, неводники) должны иметь достаточную устойчивость.
В местах выгрузки рыбы из рыбоуловителей предусмотрены
служебные мостики, удобные огражденные сходы, устроены дорожки для передвижения
рыбаков, перемещающих концентрирующие устройства вдоль рыбоуловителей.
В институте приказом администрации назначен инженер по
охране труда и ответственный за пожарную безопасность, который проводит
инструктаж с вновь принятыми сотрудниками согласно ГОСТ 12.0.004-90.
На предприятии разработана следующая документация по пожарной
безопасности:
.Общеобъектовая инструкция о мерах пожарной безопасности предприятия.
.Инструкция по пожарной безопасности зданий, помещений и сооружений.
.Инструкция по обслуживанию установок, пожаротушения.
.Инструкция по обслуживанию установок пожарной сигнализации.
.Оперативный план пожаротушения для предприятия, корпуса, здания или
сооружения.
.План ликвидации возможных чрезвычайных происшествий (взрыв, авария,
пожар) с привлечением работников служб главного энергетика, главного механика,
главного технолога, пожарной и военизированной (войсковой) охраны.
.Планы и графики проведения противопожарных тренировок, обучения, и
проверки знаний персонала, технического надзора за системами пожарной защиты.
После инструктажа сотрудники расписываются в журнале,
что ознакомлены с правилами труда на рабочем месте и безопасными приемами
работы.
Лицом, отвечающим за безопасность труда на каждом
участке производства, является бригадир, который проводит инструктаж с
работниками своего участка. На каждом участке имеется журнал выдачи
удостоверений, нарядов-допусков.
Администрация рыбоводного предприятия осуществляет
специальную подготовку лиц, обслуживающих плавучие самоходные камышекосилки,
разрабатывает соответствующие инструкции по охране труда согласно ГОСТ
12.2.003-91 «Оборудование на производстве, общие требования безопасности», ГОСТ
12.3.002-75 «Процессы производства, общие требования безопасности», ГОСТ
12.4.011-75 «Средства защиты от прохождения электрического тока».
При производстве работ необходимо исключить
возможность травмирования рук о колючие плавники, зубы и костные образования на
теле рыб. Для предохранения рук от раздражения рыбной слизью работающим
предоставляются дезинфицирующие растворы.
При лечебно-профилактической обработке рыбы, так как
этот процесс связан с использованием веществ, вредно влияющих на кожу,
используют резиновые перчатки и очки (ГОСТ 12.4.016-83, ГОСТ 12.4.011-87).
Работающих с удобрениями и ядохимикатами информируют
об их вредных свойствах и необходимости мер предосторожности. Согласно ГОСТ
12.4.016-83, ГОСТ 12.4.011-87 спецодежда, выдаваемая лицам, работающим с
ядохимикатами, не должна иметь карманов, застегиваться и завязываться должна
сзади. Продолжительность работ с ядохимикатами ограничивается 4-6 часами в
зависимости от вида ядов. Ряд специальных требований предъявляется к
организации хранения и выдачи ядохимикатов, их транспортированию.
Основные случаи заболевания рабочих связаны с
переохлаждением и мелким травматизмом. Профессиональными заболеваниями для этой
группы являются ревматизмы, остеохондрозы, острые респираторные заболевания.
Каждый год сотрудники сдают экзамен по технике безопасности: о работе с
электроприборами, о работе на воде, экзамен по оказанию первой медицинской
помощи.
Организуют систему связи в удаленных районах. В четко установленное время
дается отчет о состояние дел, здоровья сотрудников.
Государственный профсоюзный контроль за охраной труда возложен на
профсоюзы РФ. В их аппарате имеются специальные органы, которые осуществляют
только государственный (не общественный) контроль за охраной труда. Органами
этого вида контроля являются технические инспекции профсоюзов.
Ведомственный контроль за охраной труда проводят министерства и
ведомства. Они контролируют внутриведомственное соблюдение законодательства о
труде. Для этого создают специальные службы охраны труда в виде отделов с
аппаратом инженеров по охране труда, санитарных врачей и других специалистов.
Профсоюзный общественный контроль за охраной труда осуществляют
общественные инспектора и комиссии по охране труда комитетов профсоюзов.
Комиссия по охране труда состоит из членов профсоюзов и возглавляет ее
член профсоюзного комитета, а в цехе - член цехового комитета профсоюза.
Председатель этой комиссии одновременно является старшим общественным
инспектором предприятия по охране труда. Председатель комиссии не может быть
лицом из администрации. Эта комиссия контролирует соблюдение администрацией
законодательства о труде, участвует в подготовке и проверке выполнения
соглашения по охране труда: изучает причины производственного травматизма,
профессиональных заболеваний и т. д.
Для усиления контроля на производстве за соблюдением законов о труде на
общих собраниях профгрупп избираются общественные инспектора по охране труда из
числа членов профсоюза сроком на один год.
Медицинские осмотры - контрольные медицинские обследования состояния
здоровья работников; один из важнейших компонентов первичной профилактики
профессиональных и профессионально обусловленных заболеваний. Предварительный
медицинский осмотр человек проходит при поступлении на работу. Задача
медицинского персонала - определить, можно ли данному человеку по состоянию
здоровья работать в условиях данного производства или профессии.
Итоги всех медицинский осмотр оформляют в виде индивидуальных заключений
о годности человека к работе по данной специальности либо о его временном или
постоянном переводе на другую работу (трудовые рекомендации).
По результатам периодических медицинский осмотр составляются
заключительные акты и принимаются решения не только о продолжении и прекращении
производственной деятельности в предлагаемых условиях, но и о проведении
оздоровительных мероприятий среди работников, а также санитарно-гигиенических
оздоровительных мероприятий на производстве.
За последние года был отмечен один случай травматизма: рабочий при
погрузки рыбы, находящийся в алкогольном опьянение, выпал из кузова и сломал
ногу.
В феврале 2010 года в здание Института Систематики и Экологии Животных
были установлены на каждом этаже: пожарная сигнализация, пожарные краны и
приобретены огнетушители; вывешаны схемы по эвакуации при чрезвычайных
ситуациях. Оборудован угол по безопасности труда на предприятии.
В данном институте нет достаточного количества спецодежды для ихтиологов
(резиновые сапоги, перчатки, прорезиненные костюмы).
Предложения: необходимо приобрести для защиты рыбоводов от опасных и
вредных производительных факторов средства индивидуальной защиты, а именно:
фартук хлопчатобумажный с водоотталкивающей пропиткой;
рукавицы комбинированные;
сапоги резиновые;
плащ прорезиненный.
И вовремя проверять спецодежду на пригодность использования.
6. Охрана природы
Охрана природы на рыбохозяйственных водоемах ведется на основании «Водного
кодекса» Российской Федерации #G1Принятого Государственной Думой 18 октября
1995 года.
На основании его «#G0Воды являются важнейшим компонентом окружающей
природной среды, возобновляемым, ограниченным и уязвимым природным ресурсом,
используются и охраняются в Российской Федерации как основа жизни и
деятельности народов, проживающих на ее территории, обеспечивают экономическое,
социальное, экологическое благополучие населения, существование животного и
растительного мира».
Отношения по поводу вод регулируются настоящим Кодексом путем
установления правовых основ использования и охраны водных объектов.
ГЛАВА 11. Охрана водных объектов
Статья 94. Общие требования к охране водных объектов
Органы государственной власти Российской Федерации и органы государственной
власти субъектов Российской Федерации в соответствии с законодательством
Российской Федерации принимают совместимые с принципом устойчивого развития
меры по сохранению водных объектов, предотвращению их загрязнения, засорения и
истощения, а также по ликвидации последствий указанных явлений.
При использовании водных объектов граждане и юридические лица обязаны
осуществлять производственно-технологические, мелиоративные, агротехнические,
гидротехнические, санитарные и другие мероприятия, обеспечивающие охрану водных
объектов.
Использование водных объектов должно осуществляться с минимально
возможными негативными последствиями для водных объектов.
Статья 95. Охрана водных объектов от загрязнения
В целях предупреждения и устранения загрязнения водных объектов
определяются источники их загрязнения.
Источниками загрязнения признаются объекты, с которых осуществляется
сброс или иное поступление в водные объекты вредных веществ, ухудшающих
качество поверхностных и подземных вод, ограничивающих их использование, а
также негативно влияющих на состояние дна и берегов водных объектов.
Охрана водных объектов от загрязнения осуществляется посредством
регулирования деятельности как стационарных, так и других источников
загрязнения.
Федеральные органы исполнительной власти и органы исполнительной власти
субъектов Российской Федерации осуществляют охрану водных объектов от всех
видов загрязнения, включая диффузное загрязнение (загрязнение через поверхность
земли и воздух).
Статья 96. Охрана водных объектов от засорения
Запрещаются сброс в водные объекты и захоронение в них производственных,
бытовых и других отходов.
Поступление взвешенных частиц в водный объект допускается только в
соответствии с требованиями водного законодательства Российской Федерации.
Статья 97. Аварийное загрязнение водных объектов
Аварийное загрязнение водных объектов возникает при залповом сбросе
вредных веществ в поверхностные или подземные водные объекты, который причиняет
вред или создает угрозу причинения вреда здоровью населения, нормальному осуществлению
хозяйственной и иной деятельности, состоянию окружающей природной среды, а
также биологическому разнообразию.
Меры предупреждения и ликвидации аварийного загрязнения водных объектов
определяются водным законодательством Российской Федерации и законодательством
Российской Федерации о недрах.
Статья 98. Охрана водных объектов от загрязнения и засорения из
источников загрязнения, находящихся на суше
Источники загрязнения, находящиеся на суше, не должны вызывать
загрязнения и засорения водных объектов сверх установленных нормативов
воздействия на водные объекты.
Соблюдение требований части первой настоящей статьи обеспечивается
приоритетным применением технологий, не оказывающих негативного воздействия на
окружающую природную среду, ограничением использования токсичных веществ и
тяжелых металлов, введением научно обоснованных методов измерения и расчетов
сбросов сточных вод в водные объекты и выбросов в атмосферу.
Статья 99. Охрана водных объектов от загрязнения и засорения, вызываемых
деятельностью на дне водных объектов
Добыча полезных ископаемых, торфа и сапропеля со дна водных объектов или
возведение сооружений с опорой на дно должны осуществляться такими способами,
которые не оказывали бы вредного воздействия на поверхностные воды, дно, берега
водных объектов и водные биоресурсы.
При осуществлении на дне водных объектов работ, которые направлены на
использование недр, но не связаны с добычей полезных ископаемых,
недропользователи обязаны не допускать загрязнения, засорения и истощения
водных объектов.
Надзор за соблюдением законодательства возложен на районный комитет по
экологии и охране окружающей среды, а также на санитарно- эпидемиологическую
станцию.
В ходе исследовательских работ были использованы рыб из трех водоемов:
Бердского пруда, озера Улугуль и реки Алей.
Бердский пруд
По результатам химических анализов проб воды Бердского залива на трех
створах наблюдения (6 проб), показали, что вода, за исключением пробы воды с
водозабора г.Бердска (3 класс качества - умеренно загрязненная), относится ко 2
классу качества и классифицируется как чистая. Кислородный режим
удовлетворительный (7,64-12,29 мг/л). Во всех пробах содержание легкоокслиямого
органического вещества (по БПК5) превышает ПДК до 1,5 раз. Концентрации
нефтепродуктов, органических веществ (ХПК, АПАВ) не превышает ПДК ни в одной из
проб. Содержание аммоний- и нитрат- ионов также в пределах ПДК. Для всех проб
характерно сезонное повышенное содержание железа - 2-6 ПДК. Содержание меди -
1-3 ПДК, что сопоставимо с фоновыми концентрациями для данного водоема. Во всех
пробах отмечается повышенное содержание марганца, особенно в воде поселка
Агролес - 0,26 мг/л.
Озеро Улугуль
Ихтиофауна озер и рек бедна в видом отношении. Основными обитателями
водоемов являются карась золотой, серебряный, гальян и пескарь, так же
встречаются плотва, окунь и ротан. На природу области отрицательное воздействие
оказывают предприятия индустриального и агропромышленного комплексов,
транспорт. В результате загрязняются воздух, вода, почвы; исчезают редкие виды
растений и животных; снижается продуктивность естественных кормовых угодий;
исчезают малые реки, болота. В целях улучшения экологической обстановки
проводится работа по созданию системы охраняемых природных территорий.
Охраняемые территории составляют около 8% от всей площади области. Большое
количество памятников природы, разнообразны и значительны рекреационные
ресурсы, из которых широко известна Красноярско-Чернолучинская
курортно-санаторная зона.
Основным фактором, влияющим на качество воды в реке Алей является
попадание загрязняющих веществ на территории города Рубцовска.
Большое количество источников техногенного загрязнения реки расположено
выше города Рубцовска (горнорудный промышленный комплекс г. Горняк, Змеиногорск
и др.).
По классу качества воды река Алей относится к следующим категориям:
р. Алей - г. Рубцовск, выше города, - грязная,
р. Алей - г. Рубцовск, ниже города, - очень грязная.
Основными источниками загрязнения р. Алей в черте города Рубцовска
являются хозяйственно-бытовые и производственные сточные воды (Северный и Южный
промузлы), неорганизованные свалки бытовых отходов, строительного мусора в
районах жилой и гаражной застройки, расположенные в непосредственной близости
от реки, поверхностные загрязненные воды (талые и дождевые) с неблагоустроенных
прибрежных территорий города, сельхозугодий. Несоблюдение экологических
требований при обращении с отходами производства и потребления.
В настоящее время одним из трудно решаемых вопросов на территории
Рубцовского управленческого округа является проблема утилизации промышленных и
бытовых отходов. Большинство свалок населенных пунктов не оформлено должным
образом, кроме того на территории округа существуют несанкционированные свалки,
которые подлежат ликвидации.
В городе отсутствует полигон промышленных отходов.
Как правило, на территории предприятий площадки временного хранения
отходов не соответствуют требованиям санитарного и природоохранного
законодательства.
Так в г. Рубцовске полигон ТБО введен в эксплуатацию в 1993 году.
Вместимость 784,0 тыс. тонн, мощность полигона 60,0 тыс.тонн/год. Процент
заполнения более 80%. Год окончания эксплуатации - 2010 год.
Необходимость перезатаривания и организации хранения устаревших
пестицидов.
До настоящего времени остаются не перезатаренными жидкие пестициды и тара
из-под них:
в Курьинском районе - 1800 л.
В Рубцовском районе - 762 л.
В целях упорядочения хранения запасов устаревших пестицидов и принятия
мер по предотвращению загрязнения природной среды на территории Курьинского
района был организован полигон захоронения 624 тонн ядохимикатов. В настоящее
время полигон не действует, вся его поверхность имеет естественный травостой.
Выводы
. Впервые проведен морфометрический анализ
ротана-головешки в водоемах юга Западной Сибири, который находился в условиях
разнотипных биотопов, что позволило оценить их адаптацию в водоемах.
2. Выявлено, что в Бердском пруду и в р. Алей самцы по росту превосходят
самок.
. Изучено, что спектр питания ротана-головешки в разных водоемах
неодинаков, в проточной воде он представлен рыбой, а в стоячих водоемах в
основном беспозвоночными.
. Выявлена закономерность динамики численности, в исследуемых водоемах
популяция ротона-головешки и она достигает средних показателей, что приводит к
снижению выхода промысловой рыбы, т.к. ротан поедает молодь других видов рыб.
. Определена прямая зависимость между массой тела и ее длиной у всех
возрастных групп, при r=
0,97; 0,85; 0,95 для годовиков, двух- и трехгодовиков, соответственно. И при r= 0,93; 0,94; 0,92 двух-, трех- и
четырехлеток, соответственно.
Предложения
1. Результаты исследований свидетельствуют о необходимости проведение
контроля за состоянием вселенцев интродуцентов входящих в комплекс
непромысловой фауны.
2. На основании данных мониторинга необходимо осуществлять комплекс мероприятий
по регулированию численности популяции ротана-головешки в рыбохозяйственных
водоемах.
. Необходимо проводить контроль за состоянием паразитофауны вселенцев,
чтобы оценить возможность поражения местных рыб и разработать систему
мероприятий для предотвращения паразитарных инвазий.
Библиографический список
1.Бабуева Р.В. Ихтиофауна водоемов Обь-Иртышского междуречья
// Современные проблемы гидробиологии Сибири. Томск, 2001а. с. 83 - 84
. Бознак ЭИ. Головешка-ротан Perccottus glenii (Eleotrididae) из бассейна реки Вычегда // Там же. 2004. Т. 44, вып.5. с. 712 - 713.
.Болонев Е.М., Пронин Н.М., Дугаров Ж.Д. Ротан - амурский
“завоеватель” в байкальском регионе. Улан-Удэ. Изд-во Бурятского научного
центра СО РАН. 2002. 45 с.
.Болонев Е.М. Пронин Н.М. Особенности размерно-возрастной и
половой структуры локальных популяций ротана Perccottus glenii Dybowskii (Perciformes: Eleotrididae) в водоемах и водотоках бассейна
озера Байкал // Вестник Бурятского государственного университета. - Вып. 7. -
2005. - С. 138-144.
.Васильева Е.Д., Макеева А.П. Морфологические особенности и
таксономия головешковых рыб (Pisces,
Eleotridae) фауны СССР. Зоол. журн. Т. 67. Вып.
8. С.1988. 1194-1204.
.Гладких К.К., Делицын В.В., Делицына Л.Ф. Нахождение
ротана-головешки в водоемах Воронежской области. Тр. Биол. Учеб.-Науч. центра
Воронеж. Гос. Ун-та. Вып. 14.2000. С. 8-10.
.Голубцов А.С. Эколого-генетический анализ популяций ротана Perccottus glenii в естественном и приобретенном ареалах:
Автореф.дис.…канд.биол.наук. М., 1990. с.24.
.Голубцов А.С., Ильин И.И. Структура внутрипо-пуляционной
генетической изменчивости у ротана (Еleotridae, Pisces): различия между самцами
и самками по частотам генотипов двух ферментных локусов. Фенетика популяций.
Матер. 3-го Всесоюзн. совещ. Саратов. 1985. M. с. 149-150.
.Делицын В.В. Новый список ихтиофауны водоемов бассейна реки
Дон (Воронежская область). В сб.: Ихтиологические и рыбохозяйственные
исследования на реках и водохранилищах. Воронеж. Воронеж. Гос. Ун-т. 2001. С.
20-26.
.Демин А.И. Пришельцы . Интродукция новых видов флоры и фауны
в бассейне Байкал: последствия и уроки // Волна.2001. №3. С.10 -24.
.Дирипаско О.А. Первый случай поимки бычка-ротана Percottus glenii (Eleotridae) в Калиниградской области. Вопр. ихтиол. Т. 36. Вып. 6. 1996.C. 842.
.Дмитриев М.А. Осторожно, ротан. Рыбоводство и рыболовство.
№1. 1971. C. 26-27.
.Дубовский Б. Рыбы системы вод Амура. Изв. Сиб. отд. Импер.
Русск. геогр. общ. Т. 8. № 1-2. 1877.С. 1-29.
.Еловенко В.Н. Систематическое положение и географическое
распространение рыб семейства Eleotridae (Gobioidei, Perciformes), интродуцированных в
водоемы Европейской части СССР, Казахстана и Средней Азии. Зоологический
журнал. T. 60. Вып. 10.1981. С. 1517-1522.
.Еловенко В.Н. Питание ротана в прудах Хабаровского рыбхоза.
Тез. докладов Всесоюзн. Конф. молодых ученых “Mетоды интенсификации прудового рыбоводства”. М. ВНИИПРХ.
1984. С. 11.
.Еловенко В.Н., Климова С.Е. Пищевые взаимоотношения ротана с
гидробионтами рыбоводных прудов. Первый Всесоюзн. Симп. “Теоретические основы аквакультуры”.
Тез. докл. М.1983. С. 110-111.
. Еловенко В.Н. Морфо-экологическая характеристика ротана Perccottus glenii в границах естественного ареала и за его пределами:
Автореф. дис….канд.биол.наук. М., 1985. С.24.
.Журавлев В.Б., Ломодуров Е.И., Лукоянов Д.П. Вселение
ротана-головешки в пойменные водремы бассейна Верхней Оби // IX съезд гидробиол. об-ва РАН: Тез.
докл. Тольятти, 2006, Т.1 с. 163.
.Залозных Д.В. Некоторые аспекты биологии ротана в водоемах
Горьковской области. В: Наземные и водные экосистемы. Межвузовский сборник.
Вып. 5. Горький. Горьковский Гос. Университет.1982. С. 44-47.
.Залозных Д.В. Контроль за ротаном в рыбоводных прудах
Горьковской области. Сборник трудов ГосНИОРХа. Вып. 217. 1984. С. 95-102.
.Ильин И.И. Биохимический полиморфизм ротана Percottus glenii Dyb. (Eleotridae) в исходном ареале и районах интродукции. Автореферат дис-ции ...
кандидата биологических наук. М. 1987. 25 с.
.Коробейник А. Рыбы России. Ростов-на-Дону. Изд-во “Феникс”.
2001. 352 с.
.Крысанов Е.Ю., Еловенко В.Н. Структурные особенности
кариотипов головешки Perccottus glehni Dybowski (Eleotridae) в разных частях ареала. Вопр.
ихтиол. Т. 21. Вып. 5. 1981. С. 950-952.
.Кудерский Л.А. Ротан в прудах Горьковской области. В:
Вопросы акклиматизации рыб и кормовых беспозвоночных. Рыбохозяйственное
изучение внутренних водоемов. Сб. трудов № 25. Ленинград.1980. С. 28-33.
.Кудерский Л.А. Ротан в прудах Ленинградской области. В:
Рыбохозяйственные исследования на внутренних водоемах. Сб. трудов ГосНИИОРХа №
191. Ленинград. 1982. С. 70-75.
.Кудерский Л.А. Биологическое загрязнение водоемов и
экологическая безопасность. Жизнь и безопасность. № 2-3.1998. С. 602-609.
.Курдяева В.П. Некоторые особенности гаметогенеза ротана (Percottus glenii Dyb.). В: Биология рыб Дальнего Востока. Владивосток.
1976. С. 91-95.
. Купчинский А.Б., Купчинская Е.С. состав ихтеофауны и
морфологические особенности ельца Иркутского водохранилища // Современные
проблемы гидробиологии Сибири. Томск. 2001 с. 95 - 97.
.Литвинов А.Г. Морфо-биологическая характеристика
ротана-головешки из водоемов бассейна оз. Байкал и р. Амур // Биологические
ресурсы и проблемы экологии Сибири. Улан-Удэ. 1990. С. 129 - 130.
.Литвинов А.Г. Экология ротана-головешки (Perccottus glehni Dyb.) в бассейне оз. Байкал и его влияние на промысловых
рыб. Автореферат дис-ции ... кандидата биологических наук. Санкт-Петербург.
ГосНИОРХ. 1993. С. 25.
.Матвеев А.Н. Рыбы. В: Флора и фауна водоемов и водотоков
Байкальского заповедника. М. Вып. 92. 2001. C. 56-65.
.Махлин М.Д. Семейство Головешковые, или Элеотровые
(Eleotridae). Жизнь животных. Т. 4. Рыбы. Под редакцией Т.С. Расса. М.
Просвещение. 1983. С. 446-447.
.Мошу А.Я., Гузун А.А. Первая находка ротана-головешки Percottus glenii (Perciformes, Odontobutidae) в реке Днестр. Вестник зоологии. Т.
36. № 2. 2002. С. 98.
.Мухачев И.С. Увеличение биоразнообразия фауны рыб Обского
бассейна // Новые технологии в защите биоразнообразия в водных экосистемах. М.,
2002. С. 149.
.Науменко А.Н. Новые виды в составе ихтиофауны и высших
ракообразных Волгоградского водохранилища (ротан и краб). В сб.:
Рыбохозяйственные исследования в бассейне Волго-Донского междуречья на
современном этапе. Санкт-Петербург. Изд-во ГосНИОРХ. 2002. С. 184-190.
.Неелов А.В. Природа Ленинградской области: Рыбы. Л., 1987.
С. 157.
.Острошабов А.А. Распространение Perccottus glenii в Кемеровской области и перспективы проникновения его
в Алтайский край // Экология Южной Сибири и сопредельных территорий: Материалы
Междунар. науч. шк. - конф. студентов и молодых ученых. Абакан, 2004. Т.1. с.
98 - 99.
.Перескоков А.В. Экология рыб. В: Экология озера Большое
Миассово. Миасс. 2000. С. 201-220.
.Петлина А.П., Рябова Т.С. К экологии ротана водоемов
окресности Томска // Сиб. зоол. конф.: Тез. докл. Новосибирск, 2004. С. 303 -
304.
.Пронин Н.М. Об экологических последствиях акклиматизационных
работ в бассейне озера Байкал. В: Биологические ресурсы Забайкалья и их охрана.
Улан-Удэ. БФ СО АН СССР. 1982. С. 3-18.
.Пронин Н.М., Болонев Е.М. Расселение ротана в водоемах и
водотоках Байкальского региона и анализ факторов, ограничивающих его
численность в озере Байкал // Чужеродные виды в Голарктике. Борок, 2005. С. 165
- 166.
.Пронин Н.М., Болонев Е.М., Дугаров Н.А. Распространение
амурского вселенца ротана-головешки в водоемах северной Евразии и ситуации с
экспансией его в экосистему озера Байкал //Использование и охраны природных
ресурсов в России, 2005. №2. С. 80 - 84.
.Пронин Н.М., Болонев Е.М. О современном ареале вселенца
ротана Perccottus glenii (Perciformes: Odontobutidae) в Байкальском регионе и проникновении
его в экосистему открытого Байкала. // Вопросы ихтиологии. - 2006. - Т. 46, №4.
- С. 564-566.
.Пронин Н.м., Селгби Д.Х., Литвинов А.Г., Пронина С.В.
Сравнительная экология и паразитофауна экзотических вселенцев в Великие озера
Мира: ротана-головешки в озере Байкал и ерша в озеро Верхнее // Сиб. экол.
журн.,1998. №5. С. 397 - 406.
.Рыбы Подмосковья. М., 1988. С. 181.
.Cинельников A.M. Питание ротана в пойменных водоемах бассейна р. Раздольная
(Приморский край). Биология рыб Дальнего Востока. Владивосток. 1976. С. 96-99.
.Скрябин А.Г. Характеристика популяций ротана - нового вида в
фауне Байкала. В: Проблемы экологии Прибайкалья. Тез. Докл. 3 Всесоюзн. Научн.
Конф. Иркутск.1988. С. 142.
.Скрябин А.Г. Экология и морфология Восточной Сибири:
Автореф. дис. …д-ра биол. наук. Иркутск, 1997. С. 39.
.Смирнова К.В. Материалы по паразитофауне рыб - случайных
вселенцев в Балхаш-Илийский бассейн. В: Рыбные ресурсы водоемов Казахстана и их
использование. Вып. 8. Алма-Ата. Кайнар. 1974. C. 192-194.
.Соколов Л.И. О зимовке ротана в Амурских водоемах. Вопр.
ихтиол. Т. 41. № 4. 2001. C. 572-573.
.Cпановская В.Д.,
Савваитова К.А., Потапова Т.Л. Об изменчивости ротана (Percottus glehni Dyb., fam. Eleotridae) при акклиматизации. Вопр. ихтиол.
Т. 4. Вып. 4(33). 1964. С. 632-643.
.Спановская В.Д. Распределение ротана в прудах Московской
области. В: Растительность и животное население Москвы и Подмосковья. М. Изд-во
МГУ. 1978. С. 124-126.
.Торопов А.В. Рыбы - акклиматизанты реки Бия и их влияния на
местную ихтиофауну // региональные проблемы экологии и природопользования.
Томск. 2000. С. 35 - 36.
.Травкина Г.Л. Особенности оогенеза и динамики развития
ооцитов у ротана-головешки Percottus glehni Dyb. В: Проблемы надежности
функционирования репродуктивной системы у рыб (Труды Биологического
научно-иссл. инст., вып. 44). Санкт-Петербург. Изд-во С.-Петербургского
Университета. 1997. С. 29-38.
.Е.А.Челазнова. Западная Сибирь - краткий
физико-географический обзор - 2003г.
.Цепкин Е.А. О брачном наряде самцов ротана Percottus glehni Dyb. Вопр. ихтиол. Т.17. Вып. 5(106). 1977. C. 945-946.
.Шатуновский М.И., Огнев Е.Н., Соколов Л.И., Цепкин Е.А. Рыбы
Подмосковья. М. Наука. 1988.143 с.
.Шляпкин И.В., Тихонов С.В. 2001. Распространение и
биологические особенности ротана Perccottus glenii Dybowski в водоемах Верхнего Поволжья. Тезисы
докл. американо-российского симпозиума по инвазионным видам. Борок. 2001. С.
203-204.
.Яковлев Б.П. К биологии Percottus glehni Dybowski бассейна р. Сунгари. Тр. Сунгарийской речн. биол. станции. Т. 1. Вып. 1. 1925. С. 30-41.
.Lodge D. M., Williams S., MacIsaac H. J., Hayes K.R.,
Leung B., Reichard S., Mack R. N., Moyle P.B., Smith M., Andow D.A., Carlton
J.T., McMichael A. Biological invasions: Recommendations for U.S. policy and
management // Ecological applications. - 2006. - Vol.16, № 6. - P. 2035- 2054.