Почвенная мезофауна окрестностей г. Ханты-Мансийска
Содержание
Введение
Глава 1. Литературный обзор
1.1 Принцип выделения мезофаунистической группы среди педобионтов
1.2 Показатели мезофауны
1.3 Роль почвенной мезофауны в устойчивости экосистем
1.4 Почвенная мезофауна как индикатор антропического воздействия на
биоценозы
Глава 2. Материалы и методы
2.1 Методы сбора и камеральной обработки материала
2.2 Физико-географическое описание окрестностей г. Ханты-Мансийска
Глава 3. Результаты исследования
3.1 Результаты исследования мезофауны
3.2 Результаты исследование почвы
Библиографический список
Введение
Актуальность темы. Недостаточность сведений
о почвенных беспозвоночных затрудняет выделение региональных и свойственных
зонам особенностей животного населения почв. Относительно полно изучены средне-
и южнотаежные леса Европейской части СССР (Козловская. 1957, 1959, 1962;
Гаврилов, Перель, 1958; Перель, 1965; Арнольди, Матвеев, 1973; Криволуцкий,
1979). Значительно меньше известно о почвенной фауне североевропейской,
Западносибирской тайги и Сибири в целом (Козловская, 1975; Стриганова, 1967;
Клюева, 1972; Бызова, 1978; Стриганова, 2005; Голованова с соавт., 2010).
Почвенные беспозвоночные играют важную роль в жизни лесных
экосистем, являются одним из важнейших показателей их состояния и устойчивости.
Велика роль почвенных беспозвоночных в переработке растительного опада
(Стриганова, 1980), процессах его гумификации и минерализации (Курчева, 1977;
Емец, 2002). В то же время среди них много вредителей сельского и лесного
хозяйства, видов, повреждающих искусственные насаждения, особенно молодняки
(культуры) (Гиляров, 1969). Весь комплекс почвенных животных в целом и
отдельные его элементы весьма чувствительны к большинству факторов биотической
и абиотической природы, специфично реагирует на изменения влажности, температуры,
химического состава почвы, что позволяет успешно использовать его для
диагностики и индикационных целей (Гиляров, Криволуцкий, 1980). Особенно
большой интерес почвенные беспозвоночные представляют в изучении биологической
продуктивности лесных сообществ и её изменений под влиянием антропогенных
факторов (Покаржевский, 1976).
Наиболее интересно использование почвенной мезофауны в
биоиндикационных исследованиях (Криволуцкий, 1985). При этом не всегда
учитывается комплексное изменение состава педобионтов, а берется отдельный тест
объект по состоянию показателей которого и ведётся оценка изменённости биома
или содержания тяжелых металлов в среде по накопляемости их в организмах
биоиндикаторов (Воробейчик, 1996, 1998; Воробейчик с соавт., 2007). Также существует
необходимость выделения типичных групп синантропов среди педобионтов и
использование ряда коэффициентов (Ефремов с соавт, 2010) для понимания и оценки
антропической нагрузки и трансформации всего сообщества в целом под ее
влиянием.
Цель работы: изучение почвенной мезофауны
естественных и нарушенных местообитаний окрестностей г Ханты - Мансийска.
Задачи:
1. изучить таксономический состав почвенной мезофауны;
2. установить характер и степень изменений сообществ
педобионтов нарушенных местообитаний;
. проанализировать основные почвенные характеристики
естественных и нарушенных мест обитания мезофауны (механический состав,
структурный состав, водопрочность, водопроницаемость, влагоемкость, рН,
содержание гумуса);
. изучить качественное и количественное соотношение
различных систематических групп мезофауны;
. установить плотность и биомассу почвенных организмов
естественных и нарушенных биотопов;
. изучить возрастной состав почвенных организмов;
. выявить вертикальное распределение почвенной биоты;
. проанализировать трофическую структуру мезофауны.
Объект исследований. Мезофауна почв
окрестностей г. Ханты-Мансийска.
Предмет исследований: зависимость видового
состава мезофауны от качественных характеристик почв естественных и нарушенных
биотопов окрестностей г. Ханты-Мансийска.
почвенная мезофауна педобионт антропический
Гипотеза: Существует зависимость между сообществами
педобионтов естественных и нарушенных местообитаний.
Научная новизна: Впервые получены данные о
видовом составе мезофауны окрестностей г. Ханты-Мансийска.
Глава 1.
Литературный обзор
1.1 Принцип
выделения мезофаунистической группы среди педобионтов
Почвенные животные, наряду с растениями и микроорганизмами,
играют немаловажную роль в создании биологической активности почвы, в
обогащении ее жизненно важными для роста и развития высших растений веществами
(Гиляров, 1965).
Для одних дышащих воздухом животных почва представляется как
система мелких пещер, для других - это плотная среда, оказывающая значительное
механическое сопротивление при движении. Таких почвенных животных объединяют
под названием мезофауна. Размеры представителей мезофауны почв от 1,5 до 20 мм.
Мезофауна самая многочисленная часть почвенных животных (Виноградский, 1952),
благотворно влияющих на качество и структуру почвы.
Почвенная фауна была классифицирована многочисленными
западными авторами, включая Kevan (1962), Schaller (1968) и Wallwork (1970).
Получили широкое признание классификации основанная на размерах тела, время,
проведенное в почву; место или места обитания в почвенном профиле; кормления; и
способ передвижения в почве, за основу берется размер тела (Wallwork, 1970).
Почвенная фауна в целом характеризуется небольшими размерами
(Kuhnelt, 1976). Длина тела колеблется от 0,0002 см до более чем 20 см).
Педобионтов на основании этой характеристики подразделяют на микрофауны,
мезофауны и макрофауны. Микрофауна (размер тела от 0,0002 до 0,002 см) состоит
из простейших. Мезофауна колеблется в диапазоне от чуть более 0,002 до 1 см, и
включает в себя клещей, ногохвосток, пауков, псевдоскорпионов, личинок
насекомых и многоножек и меньшей мере равноногих (Dunger, 1964; Brans, 1968).
Макрофауна (от 1 см и более) включают дождевых червей, насекомых и крупных
паукообразных, и живущих в почве позвоночных (Wallwork, 1970).
По классификации М.С. Гилярова к мезофауне относятся
почвенные беспозвоночные средних размеров, выбираемые из почвы методом ручной
разборки, размеры которых колеблются от нескольких милиметров до нескольких
сантиметров (дождевые черви, пауки, насекомые и их личинки и т.д.).
Подразделяясь по способу питания на сапрофагов, фитофагов и зоофагов. (Гиляров,
1941).
Согласно классификации Н.М. Черновой (2004) почва для
организмов разных размеров выступает как разная среда следовательно она
выделяет мелких почвенных животных, которых объединяют под названием микрофауна
(простейшие, коловратки, тихоходки, нематоды и др.), почва - это система
микроводоемов. По существу, это водные организмы. Они живут в почвенных порах,
заполненных гравитационной или капиллярной водой, а часть жизни могут, как и
микроорганизмы, находиться в адсорбированном состоянии на поверхности частиц в
тонких прослойках пленочной влаги. Многие из этих видов обитают и в обычных
водоемах.
Для дышащих воздухом несколько более крупных животных почва
предстает как система мелких пещер. Таких животных объединяют под названием
мезофауна. Размеры представителей мезофауны почв - от десятых долей до 2-3 мм.
К этой группе относятся в основном членистоногие: многочисленные группы клещей,
первичнобескрылые насекомые (коллемболы, протуры, двухвостки), мелкие виды
крылатых насекомых, многоножки симфилы и др. У них нет специальных
приспособлений к рытью. Они ползают по стенкам почвенных полостей при помощи
конечностей или червеобразно извиваясь.
Более крупных почвенных животных, с размерами тела от 2 до 20
мм, называют представителями макрофауны. Это личинки насекомых, многоножки,
энхитреиды, дождевые черви и др. Для них почва - плотная среда, оказывающая
значительное механическое сопротивление при движении. Эти относительно крупные
формы передвигаются в почве либо расширяя естественные скважины путем
раздвигания почвенных частиц, либо роя новые ходы.
И мегафауна почв - это крупные землерои, в основном из числа
млекопитающих. Ряд видов проводит в почве всю жизнь (слепыши, слепушонки,
цокоры, кроты Евразии, златокроты (Чернова, Былова, 2007).
Существуют различные классификации почвенных организмов,
составленные зарубежными и отечественными авторами, но при этом одни и те же
организмы часто оказываются отнесенными в разные категории, поэтому мы
придерживаемся классификации М.С. Гилярова, где педобионты четко разнесены.
Распределение педобионтов в почве весьма не однородно, виной
этому является множество факторов биотические, абиотические и антропогенные.
Основным фактором ограничивающим распространение педобионтов является
влажность. (Проканова, 2005; Семенова, 1967) Представители мезофауны очень
чувствительны к высыханию почвы. Основным средством спасения от колебания
влажности воздуха для них является передвижение вглубь. Но для некоторых видов
возможность миграции по почвенным горизонтам вглубь ограничивается изменением
структурного состава и диаметра пор, поэтому передвижения по скважинам доступны
только самым мелким видам. Более крупные представители мезофауны обладают приспособлениями,
позволяющими переносить временные неблагоприятные условия: защитными чешуйками
на теле, примитивной трахейной системой, частичной непроницаемостью покровов.
(Гиляров, 1949) Отмечено, что при снижении влажности почвы, дождевые черви
перестают питаться и инкапсулируются (Соколов, 1956; Роднянская, 1957).
Почвенные животные заселяют, в основном, верхние (глубиной до
20-40 см) горизонты почвы, в сухих местностях лишь отдельные виды проникают на
глубину нескольких метров (Новоселова, 2009) что чаще всего связанно с типом
сложения почвы, ее структурой (Антощенков, 1992). Содержание комплекса солее,
pH и содержание органического вещества или опада в почве также влияют на
локализацию почвенных организмов (Хотько и др., 1982; Криволуцкий, 1994) Так
например Сатчел (Сатчел, 1955) установил, что некоторые виды дождевых червей
имеют разную зависимость от pH почвы и потому разные виды часто обитают в
почвах с разной кислотностью. При этом в процессе жизнедеятельности сами живые
организмы влияют друг на друга. Почвенные животные часто обнаруживаются в таких
местах, где скапливаются определенные педобионты, выделяя специфические
вещества, организмы могут активировать или подавлять деятельность почвенных
животных (Курчева, 1971).
.2 Показатели
мезофауны
Хорошими показателями состояния благополучия или нет,
сообществ педобиотов, являются такие показатели как плотность, масса, видовое
разнообразие, возрастная структура, вертикальное распределение, трофическая
структура.
Изменения почвенной фауны, которая составляет до 95% по
биомассе числу видов всего населения ландшафта. Может служить индикатором
нарушенности среды на территориях, наиболее подверженных такому воздействию
(Криволуцкий, 1975). Изучение сезонной динамики численности и вертикального
распределения почвенных беспозвоночных позволяет более полно охарактеризовать
комплекс и экологический состав фауны почв, а также дает ценные сведения о
процессе почвообразования, водных, солевых, температурных режимах почвы
(Криволуцкий, 1977). В частности изменения численности, возрастного состава и
вертикального распределения почвенных организмов дают информацию о присутствие
полютантов в почве и сообществе в целом (Воробейчик, 1995, 1998). В работе Е.Л.
Воробейчика (Воробейчик, Голованова, 2007) говорится о том что с приближением к
источнику выбросов меняется характер вертикального распределения, таким образом
что на фоновой территории около 40% беспозвоночных сосредоточенно в почве. При
росте загрязнения для основных групп мезофауны прослеживается четкая
закономерность - перераспределения плотности в подстилку, в которой на
импактной территории сосредоточено около 90 % животных. Вероятным обьяснением
этого может быть изменение водно-воздушных свойств почвы (Кайгородова,
Воробейчик, 1996), что при высокой токсичности делает невозможным обитание в
ней почвенных животных. С приближением к СУМЗу (Среднеуральский медеплавильный
завод) численность, возрастной состав и вертикальное распределение дождевых
червей изменяется подтверждая ранее описанную закономерность (Воробейчик, 1995,
1998).
Важной характеристикой почвенной мезофауны является состав
трофических групп. М.С. Гиляров подразделяет по способу питания педобионтов на
сапрофагов, фитофагов и зоофагов, они играют важную роль в процессах
почвообразования и потребления органического вещества.
Зоофаги поедают других животных, выступая в роли хищников,
или паразитов (стафилины).
Сапрофаги питаются мертвыми остатками растений, трупами
животных, детритом, экскрементами (настоящие дождевые черви, личинки насекомых,
слизни и др.).
В почве беспозвоночные рождаются, питаются, размножаются и
умирают. Поэтому, естественно, они не могут не оказывать влияния на среду, в
которой обитают. Влияние это разностороннее, но наиболее существенно их участие
в разложении и гумификации растительных остатков. (Satchell, 1963). Свойственно
это сапрофагам, составляющим обычно очень высокий процент, как по численности,
так и по биомассе от всей почвенной фауны. Только там, где почва не имеет
выраженной подстилки и бедна перегноем, сапрофаги могут уступать по числу и
биомассе почвенным животным с другим характером питания (Гиляров, 1965)
К сапрофагом относят животных, питающихся тканями мертвой
растительности. Одни из них поедают непосредственно опад, другие - экскременты
потребителей растительного материала среди беспозвоночных и позвоночных,
гниющую древесину, детрит. (Курчева, 1971).
Среди почвенных беспозвоночных мезофауны довольно часто
встречаются факультативные сапрофаги. Личинки некоторых насекомых - вредителей
сельскохозяйственных и лесных культур, - являющихся фитофагами, при хороших
условиях увлажнения и богатстве почвы разлагающимися растительными остатками
могут переходить к питанию последними.
Преобладание той или иной трофической группы или наличие
одной говорит о нарушениях внутри сообщества. Преобладание зоофагов в
сообществе верный сигнал того что оно подверглось антропической нагрузке или
загрязнении (Покаржевский, 1976).
Показатели видового богатства также являются важным фактором
характеризующим сообщества педобионтов. В почве встречаются колоссальные
количества беспозвоночных животных: простейших, коловраток, тихоходок, нематод,
энхитреид, дождевых червей, брюхоногих моллюсков, мокриц, многоножек, клещей и
прочих членистоногих. (Курчева, 1971).
Помимо большого числа постоянно привязанных к почве насекомых
(ногохвостки, личинки двукрылых и жесткокрылых и др.) 98% видов свободно
живущих представителей класса насекомых в какой-то период своей жизни тоже
связаны с почвой. (Гиляров, 1965) Выявлено, что основными и наиболее обычными
группами почвенной мезофауны на исследованной территории являются: дождевые
черви, многоножки, личинки и имаго пластинчатоусых жуков, долгоносиков,
жужелиц, двукрылых, перепончатокрылых, проволочники. Представители других групп
встречались единично в отдельных пробах (мокрицы, личинки мертвоедов и др.)
(Антощенков, 1992. Следует отметить что наиболее типичными представителями
слагающими почвенную мезофауну территории Центральной России, Западной
Восточной Сибири и дальнего востока являются три типа беспозвоночных: кольчатые
черви, членистоногие и моллюски. (Стриганова, 2005; Криволуцкий, 1987; Курчева,
1977; Ганин, 1997; Валькова, 2007).
Пространственное распределение почвенных животных -
направление, разработкой которого занимались несколько поколений почвенных
зоологов, как в нашей стране, так и за рубежом. (Гиляров, 1965; Decaens, Rossi,
2001).
Неравномерность пространственного распределения организмов
известна давно и является одной из важнейших характеристик реакции организмов
на факторы окружающей среды. Уже в 1946 году пространственное распределение
почвенных животных было проанализировано к. Кроулом (Cole, 1946, 1949), который
показал, что случайным оно является только у пауков, тогда как для других
подстилочных обитателей оно определенно. Необходимость исследования пространственного
распределения почвенных организмов очевидна, и ее исследования и временной
динамики были поставлены на первое место среди приоритетных задач в почвенной
экологии (Eijsackers, 2001), фактически наравне с исследованиями
биоразнообразия и функционирования почвенных систем (Brussaard, 1997).
Изучение пространственного распределения почвенных организмов
остается на переднем плане современных исследований, о чем свидетельствуют
появляющиеся новые работы (Kessler, 2006; Schooley, 2006; Huhta, 2007). Хотя
изменчивость распределения почвенных организмов часто рассматривается как
случайный шум, понимание ведущих факторов такой изменчивости может
способствовать развитию представлений о факторах, регулирующих состав почвенной
биоты и о ее функциональной роли в экосистемах. (Покаржевский, 2005)
Пространственная иерархия факторов окружающей среды, внутренние популяционные
процессы и внешние воздействия влияют на пятнистость распределения популяций
почвенных организмов и экосистемные процессы, происходящие в масштабах от
миллиметров до сотен метров. (Савин, 2004) Характеристика таких процессов -
одна из основных целей дальнейших исследований для улучшения качества их оценки
и понимания факторов, управляющих почвенными организмами, как по
пространственной, так и по временной шкале (Мешалкина, 2001).
Пространственный подход может помочь в выявлении факторов,
ответственных за чрезвычайное высокое разнообразие организмов, живущих в почве.
Ограниченное расселение многих представителей почвенной биоты, гетерогенность
почвенных ресурсов и местные нарушения ведут к стохастической динамике
пространственного распределения, что может играть важную роль в поддержании
широты почвенного биоразнообразия и его функционирования. Благодаря роли
пятнистости почвенной биоты в регулировании пространственного распределения
ресурсов питательных веществ для растений, мутуалистов и антагонистов корней,
пространственное распределение почвенных организмов, по видимому, действует как
ведущая сила в пространственном распределении растений и в конечном итоге в
структуре растительного разнообразия (Bever, 1997)
.3 Роль
почвенной мезофауны в устойчивости экосистем
Почвенные животные, наряду с растениями и микроорганизмами,
играют немаловажную роль в создании биологической активности почвы, в
обогащении ее жизненно важными для роста и развития высших растений веществами.
В процессе метаболизма животных происходит так же расщепление сложных
органически веществ на легко минерализирующиеся соединения: мочевину, мочевую и
гиппуровую кислоты, гуанин. Кроме того, они выделяют в свободном состоянии
многие соли: хлориды, фосфаты калия, магния, натрия, кальция и т.п. (Курчева,
1971).
Свежие экскременты представителей мезофауны имеют, как
правило, повышенное, по сравнению с окружающей почвой, содержание углерода,
азота, фосфора, калия и т.п. По данным Битюцкого и др. (2004) дождевые черви
экскретируют около 250 мкг орг. С/особь/сутки, в которых содержится 540 мкг/мл
глюкозы на особь. Дождевые черви играют в почве роль активных
средообразователей, так как элементы их экскрементов становятся прекрасным
энергетическим материалом для микроорганизмов и источником питательных веществ
для растений. (Олейник, Демин, Бызов, 2005)
Сапрофаги мезофауны обогащают почву кальцием, извлекаемым из
опада. Много углекислого кальция, аккумулирующегося в покровах тела поставляют
многоножки и насекомые при неоднократных линьках, а так же раковинные моллюски
при отмирании. (Курчева, 1971)
М.С. Гиляровым установлена тесная связь характера почвенного
профиля, содержания и распределения в нем гумуса и мощности гумусового
горизонта с видовым составом, характером миграции биомассой почвенных
сапрофагов; выявлено, что возрастание численности крупных сапрофагов и общей
численности всех почвенных беспозвоночных коррелирует с повышением содержания
гумуса в почве. Почвенные животные играют огромную роль в возвращении в почву
питательных веществ, выносимых из нее растениями. Они во много раз ускоряют
разложение растительных остатков не только как потребители опада, но и как
стимуляторы действий микроорганизмов.
По данным Гилярова, Криволуцкого и др. (1985), фитофаги
являются вредителями. При высокой плотности расселения и в почве, наряду с
положительным действием фитофагов на физические, водные, воздушные свойства,
вследствие их роющей деятельности отрицательное влияние их в целом на процесс
гумусообразования в целом превалирует. Это объясняется тем, что подгрызая корни
живых растений, они значительно снижают биомассу поступления растительных
остатков в почву.
Мезофауна влияет на аэробность почвы. Так, черви роют ходы и
тоннели, улучшая воздушный баланс слоев почвы. На характеристики почвы
оказывают значительное влияние муравьи. Однако влияние это строго локально и
ограничено гнездом, где муравьи осуществляют активную роющую и строительную деятельность,
а так же кормовой территорией. Для муравьев влияние отмечено в радиусе до 3 м
от гнезда и до 1 м в глубину. Муравьи оказывают максимальное воздействие на
процессы почвообразования в условиях, когда территории отдельных гнезд
сливаются, и муравьи сплошь контролируют отдельные пространства. (Гилев, 2005)
Населяющие почву беспозвоночные способствуют перемешиванию
почвенных горизонтов и увеличению скважности, а, следовательно, воздухо - и
водопроницаемости почвы, содействуют образованию почвенной структуры. От
состава и численности почвенного населения зависят скорость и характер
разложения лесной подстилки, а также физико-химические свойства и профиль почв.
(Хотько, Чумаков, 1980).
Полноценное функционирование комплекса почвенных
беспозвоночных обеспечивает благоприятные условия для существования и развития
биотопа. Дождевые черви и многоножки не слишком подходят для сравнения
различных экосистем по видовому составу, хотя они и доминируют по биомассе
среди других групп почвенной фауны, при этом их нельзя не учитывать. Наоборот,
насекомые и особенно жесткокрылые, - очень удачный объект для сравнения
экосистем по видовому составу обитающих в них животных. (Покаржевский, Богач,
Гусев, 1980).
Для устойчивого существования и функционирования экосистем
необходимо сохранение связей между всеми основными элементами этих систем.
Только в этом случае возможно нормальное течение многих процессов, в том числе
и основы существования экосистем - круговорота веществ. В цепи преобразования
органических веществ очень важным звеном является почва, так как в ней
начинается и заканчивается биогенный круговорот. Именно почва помогает
веществам и элементам снова включиться в биогенный круговорот через зеленые
растения. При этом почва обладает уникальной способностью накапливать, а затем
снабжать растения необходимыми для них элементами питания и водой. Таким
образом, продуктивность наземных экосистем очень сильно зависит от плодородия
почвы. В свою очередь, плодородие почвы и процессы почвообразования зависят от
обитающих в ней живых организмов. (Антощенков, Гиляров, Криволуцкий, 1985)
За последние годы среди различных направлений современной
почвенной зоологии довольно широкое распространение получило индикационное
направление. Ставящее задачей использовать отдельные виды или сообщества видов
в качестве показателей среды (Криволуцкий. 1985).
.4 Почвенная
мезофауна как индикатор антропического воздействия на биоценозы
Основные понятия, встречающиеся в индикационной почвенной
зоологии - это прежде всего индикаторы, индикация и индикационные признаки.
Индикаторами называются виды или сообщества почвенных беспозвоночных, связанные
с определенными экологическими условиями настолько, что данные условия могут
быть распознаны по присутствию этих видов или сообществ.
Определение экологических условий по присутствию тех или иных
индикаторов носит название почвенно-зоологической индикации. Индикационными
признаками называются те или иные особенности видов или сообществ, которые
могут быть показателями определенных условий среды, хотя сами эти организмы и
сообщества не являются индикаторами этих условий. При этом нельзя забывать, что
индикационное значение тех или иных видов или сообществ имеет относительный, географически
обусловленный характер и что один и тот же вид в разных частях своего ареала
характеризуется разным диапазоном требований в отношении режима почв, и
выделение индикаторов носит чисто региональный характер. (Криволуцкий, 1987)
Антропогенное воздействие на территории, осваиваемые
человеком, так или иначе приводит к изменению всего комплекса животных,
населяющих данный ландшафт. Известно, что наибольшие воздействие на
естественные сообщества и всю биосферу в целом оказывает человек, не является исключение
и почвенный компонент. Можно выделить следующие формы антропогенного
воздействия на почву:
. Физическое воздействие (рекреация, выпас скота,
тяжелая с/х техника, орошение, водная эрозия, открыта добыча полезных
ископаемых, строительство, военные действия и т.п.).
2. Химическое загрязнение (промышленные выбросы,
агрохимикаты, радиоактивные осадки, органические газы и жидкости и т.п.)
. Биологическое загрязнение (объекты
микробиологического производства белка и белково-витаминных концентратов,
энтомоппатогенные бактерии, используемые в биологической борьбе с разного рода
вредителями и т.п.).
Физические воздействия на почвы. Изменение почвенных
параметров касается прежде всего сложения и структуры почвы снижения
порозности, ухудшения аэрации, дренажа. Одним из примеров физического
воздействия на почвенный покров являются рекреационные нагрузки, выделяют
четыре стадии рекреационной дигрессии лесных сообществ. Основными
индикационными параметрами являются: посещаемость, площадь тропиночной сети,
проективное покрытие травянистого покрова, фаунистический состав и соотношение
лесных и сорных видов, запасы подстилки и ее фракционный состав (Практические
рекомендации., 1987). Высокие рекреационные нагрузки приводят к снижению
видового разнообразия и исчезновению подстилочных форм педобионтов, подавляют
общую биологическую активность.
При длительном сельскохозяйственном использовании почв наблюдается
глубокая деградация почвенного биоценоза (Бабьева И, Зенова, 1989).
Индикационными признаками являются: снижение общей биомассы и видового
разнообразия, упрощение педокомплекса, снижение обилия и видового разнообразия
сапрофагов, доминирование фитофагов, смена детритной пищевой цепи на
пастбищную, увеличение патогенных форм микроорганизмов, частые вспышка
массового размножения вредителей. Основными индикаторами нарушении такого типа
являются комплексы почвенных беспозвоночных - структурные перестройки переходят
в функциональные и приводят к локальным изменениям характеристик зонального
типа биологического круговорота.
Биологическое загрязнение почв чужеродными микроорганизмами
происходит в результате попадания в почву бытовых и сельскохозяйственных
отходов и отбросов, а также за счет аэрозолей микробиологических производств. С
бытовыми отбросами в почву могут попадать потенциально опасные микроорганизмы -
патогенные и токсикогенные, способные вызывать кишечные инфекции и пищевые
отравления у человека, эпидемические заболевания у животных, токсикозы
растений.
Химическое загрязнение почв по своему воздействию
значительно превосходит все виды физических воздействий. Изменение химических
параметров почвы отражается прежде всего на росте и продуктивности отдельных
видов и всего сообщества почвенных организмов в целом. Консументы и редуценты
чаще всего испытывают косвенное влияние в результате изменения структуры
фитоценозов, количественных и качественных перемен в доступности пищи. Это
отражается на их активности, обилии разнообразии и возрастной структуре
популяций. Химические изменения в силу своей сложности часто приводят к структурным
изменениям на нескольких трофических уровнях (Яковлева, 1992).
Загрязнение почв тяжелыми металлами. Загрязнение почв
металлами происходит в результате их поступления из атмосферы в виде сухих
выпадений и с осадками (Бериня с соавт, 1980, 1983; Boutron, 1988). Особенно
это относится к организмам, потребляющим тяжелые металлы вместе с пищей
непосредственно из почвы и включающим их в свой обмен веществ. Внесение в почву
тяжелых металлов в конечном итоге вызывает снижение численности и разнообразия
педобионтов, при этом возрастает доля один видов, которые приспособившись
полностью занимают освободившуюся нишу.
Почва способна к самоочищению от несвойственных ей
микроорганизмов. Они могут быть разной природы. В первую очередь это связано с
отсутствием в почвенной среде условий, необходимых для развития попадающих
извне микроорганизмов, а также неблагоприятного действия физических и
химических факторов (кислотности, высушивания, солнечной радиации и т.д.).
Другой механизм элиминации микроорганизмов - взаимодействие с членами почвенной
биоты - выедание, лизис и др. (Гиляров, 1965).
При всем многообразии антропогенных факторов некоторые из них
сходны по своей природе, характеру воздействия на живые организмы с
естественными биотическими и абиотическими факторами, например, пастьба скота.
Другие же совсем не характерны для природных комплексов, так как искусственно
созданы человеком, например, часть минеральных удобрений, а также
сельскохозяйственная деятельность отрицательно воздействует на природные сообщества
и на организм каждого почвенного животного в частности. Замечено, что внесение
в почву умеренных норм минеральных удобрений может оказать положительный эффект
не только на растения, но прямо или косвенно положительно отразиться и на
некоторых, играющих значительную роль в почвообразовательных процессах, группах
животных (Тишлер, 1971). Однако внесение высоких концентраций минеральных
удобрений в почву может отрицательно сказаться на тех же животных. При этом
действие такого, очень естественного антропогенного фактора, как выпас скота на
почвообитающих животных, тоже не однозначно.
Крупные почвенные беспозвоночные имеют большую индикационную
ценность. (Лебедева, 2004; Чернова, Стриганова, 1999) Для экологического
мониторинга применимы в первую очередь именно объекты мезофауны. (Захаров,
Бызова, 1989) Почвенная мезофауна чувствительна к реакции почвенных растворов.
Изменение химизма среды обитания, физических свойств почв приводит к изменению
численности и видового состава животных. В связи с этим многие исследователи
считают мезофауну одним из лучших биоиндикаторов, так как все активно
передвигающиеся виды реагируют на малейшее изменение среды варьированием
численности и нарушением соотношений трофических групп. (Иванов, 2009) Наиболее
удобными тест-объектами являются дождевые черви, щелкуны и их личинки, крупные
жужелицы, некоторые виды мокриц, чернотелки и их личики. (Смольникова, 2008)
Глава 2.
Материалы и методы
2.1 Методы
сбора и камеральной обработки материала
Материалом для данной работы послужили исследования
проведенные летом 2011 г. В окрестностях г. Ханты-Мансийска.
Численность в пробах пересчитывалась на м2. Отбор
и фиксация материала осуществлялись по стандартной методике, путем ручной
переборки почвы (Бызова с соавт., 1987). Установление видов осуществлялось по
определителю пауков В.П. Тыщенко (Тыщенко, 1971), определителю семейств личинок
жесткокрылых Е.А. Негробова (Негробова, 2008), определитель насекомых
европейской части СССР Б.М. Мамаева (Мамаев, 1976), определителю насекомых по
личинкам Б. М Мамаева (Мамаев, 1972), жужелиц рода Carabus южной Сибири В.Г.
Шиленкова (Щеленков. 1996), определителю многоножек-костянок СССР Н.Т.
Залесской (Залесская, 1978), определителю дождевые черви фауны России Т.С.
Всеволодовой-Перель (Всеволодова - Перель, 1998).
Всего проанализировано 50 проб почвенной мезофауны и 10
сборных проб почвы. Материал для исследования отбирался - методом почвенных
раскопок по М.С. Гилярову (1975). В каждом биоценозе было заложено по 10
экспериментальных площадок размерами 25 х 25 см. При этом можно увеличить
количество проб в 2-3 раза с одного участка. (Бызова, 1987). На каждой
экспериментальной площадке изымалось по четыре почвенных пробы до встречаемости
видов из слоев 0-10, 10-20, 20-30. Грунт выкладывался на полиэтиленовую пленку
и из него выбирались все беспозвоночные организмы, которые составляли мезофауну
в месте экспериментальной площадки. Собранные экземпляры фиксировались в спирте
и помещались в емкостях соответственно "проба - емкость" для
дальнейшей идентификации, определения таксона и составления коллекции.
Камеральная обработка материала проводилась на базе кафедры
зоологии и физиологии ОмГПУ.
Изучались 5 видов местообитаний. В качестве естественных
местообитаний выбраны:
. хвойная тайга, преимущественно представленная
разнотравными и папоротниковыми сосняками и ельниками;
2. пойменные луга на берегу реки Щучье.
В качестве нарушенных местообитаний рассматривались:
. смешанная тайга, которая возникает на месте вырубок
дорог, пожаров, гарей;
2. пойменный луг под действием рекреационной нагрузки в
черте города;
. пойменный луг под действие мусороперерабатывающего
завода за чертой города.
Сбор материала производился в каждом биотопе по методике
описанной выше. Согласно провинциальному делению пробы брались в наиболее
типичных биотопах (тайга, заливные луга рек) а также на территориях
подверженных антропогенному воздействию (заливные луга, под действием
рекреационной нагрузки, и под действием мусороперерабатывающего завода) и
сосновых лесах. Далее осуществлялось определение каждого объекта и включение
его в коллекцию. При обработке материала изучались следующие показатели
почвенной мезофауны: видовое разнообразие на уровне точки сбора, биомасса,
трофические группы, плотность, возрастной состав. Нами предлагается при оценке
состояния сообществ использовать анализ доли фоновых видов педобионтов,
видов-синантропов и видового разнообразия. В качестве основных показателей
степени дигрессии сообществ использовались таксономическое разнообразие,
плотность.
Изучение структуры почвы имеет большое значение, так как она
обуславливает создание водно-воздушного и пищевого режимов. Наиболее
благоприятна с экологической точки зрения зернистая структура диаметром от 3 до
5мм. В почвах с такой структурой создается оптимальное соотношение между капиллярными
и некапиллярными порами, следовательно, и благоприятное соотношение воды,
находящейся в капиллярах, и воздуха, содержащегося в крупных некапиллярных
порах. (Пирогова 2005)
С целью изучения структурного состава почвы и выявления
процента полезной фракции мы использовали метод просеивания почвы через
специальный набор сит с различными по диаметру отверстиями. Массу структурных
фракций определяли на электронных весах. Полученные данные вносили в таблицу.
На основании данных таблицы построили диаграмму Рис 3.
Расчет процентного содержания фракции 3-5мм от общей массы
пробы произвели по формуле:
Х = (А х 100) / Р,
где
Х - процентное содержание структурных отдельностей данной
фракции,
А - масса структурных отдельностей данной фракции,
Р - общая масса пробы почвы.
Так как ценность структуры зависит от способности элементов
противостоять размывающему действию воды, мы определили водопрочность элементов
фракций методом Н.Н. Никольского. Из каждой фракции выбирали по 10 агрегатов,
помещали и в чашки Петри, приливали в чашку дистиллированную воду до уровня 2см
над агрегатами. Через 20 минут подсчитывали количество сохранившихся элементов.
Прочными считаются агрегаты, которые после 20-минутного размачивания при слабом
и осторожном перемешивании стеклянной палочкой не распадались.
Процент водопрочных агрегатов вычисляли по формуле:
А = (В х 100) /С, где
А - процентное содержание водопрочных агрегатов данной
фракции,
В - количество сохранившихся агрегатов,
С - количество взятых для анализа агрегатов.
Полученные результаты заносили в таблицу, по данным таблицы
построили диаграмму Рис 4.
Для выявления качества почвенных проб определяли
характеристики: водопроницаемость, влагоемкость, водоотдачу.
Водопроницаемость - способность почвы пропускать сквозь себя
воду. На водопроницаемость значительное влияние оказывают строение, структурный
состав почвы. (Пирогова 2005)
Влагоемкость - способность почвы удерживать в себе воду.
Водопроницаемость - способность почвы после полного
промачивания и насыщения отдавать то или иное количество воды путем стекания.
Для определения этих характеристик собирали установку из
стеклянной трубки диаметром 3-4 см и длиной не менее 25 см. Один конец трубки
завязывали марлей, предварительно закрыв его фильтровальной бумагой. Заполняли
трубку на 20 см от основания почвой, укрепляли ее на штативе, подставив под нее
мерный стакан. Точно замерив время, наливали в трубку 100 мл воды. Момент
появления первой капли из почвенного слоя (сек), время появления последней
капли (мин), объем вылитой из трубки воды (мл) заносили в таблицу. Вычисляли
влагоемкость как разницу между количеством влитой и вытекшей воды.
Для определения в почве легкорастворимых веществ, проводили
качественный анализ водной вытяжки. Материал тщательно растирали пестиком в
фарфоровой ступке. Брали навеску почвы 10 г и заливали ее 50 мл
дистиллированной воды. Содержимое оставляли на 10 минут и фильтровали через
стеклянную воронку с бумажным фильтром в колбу.
Для качественного определения хлоридов из отфильтрованной
жидкости пипеткой брали 5 мл в пробирку, добавляли 3-4 капли 10% -ной азотной
кислоты и по каплям добавляли 0,1Н раствор нитрата серебра. Выпадение белого
осадка, указывая на наличие в фильтрате хлоридов.
Для качественного определения сульфатов брали 3 мл водной
вытяжки в пробирку, 3-4 капели 10% -ной соляной кислоты и 20 мл 20% -ного
раствора хлорида бария. Раствор в пробирке нагревали до кипения, сульфат бария
выпадал в виде белого мелкокристаллического осадка, что свидетельствовало о
содержании сульфатов.
Наличие в почве кальция устанавливали качественной реакцией 5
мл фильтрата, подкисленного 1-2 каплями 10% соляной кислоты, и 2 мл 4% -ного
раствора оксалата аммония. Выпадающий белый осадок щавелевокислого кальция
свидетельствовал о содержании кальция в почве.
Одной из важнейших характеристик почвы является кислотность.
Кислотность почв вызывается водородными ионами, которые находятся в почвенном
растворе в свободном состоянии или поглощены почвенными коллоидами. Наличие в
почве поглощенного водорода значительно ухудшает ее свойства: она теряет
структурность, из нее легко вымываются питательные соединения, резко снижается
плодородие. (Пирогова 2005)
Пробу почвы тщательно растирали пестиком в фарфоровой ступке,
брали навеску 20 г и помещали ее в стакан емкостью 200 мл. В стакан наливали
100 мл дистиллированной воды, несколько раз взбалтывали и оставляли на 5 минут.
Определение кислотности было проведено с помощью прибора - мультитеста.
Гумус является основной составной частью органического
вещества почвы. Под органическим веществом почвы понимают совокупность живой
биомассы и органических остатков растений, животных, микроорганизмов, продуктов
их метаболизма и специфических новообразованных органических веществ почвы -
гумуса. Анализируя количество гумуса в почве, можно установить потенциал,
продуктивность биотопа, сбалансированность использования его ресурсов.
Определение гумуса в почве проводилось методом И.В. Тюрина, в
модификации В.Н. Симакова с дополнениями Б.А. Никитина. Из образцов почвы брали
навеску от 0,1-0,2 г в зависимости от предполагаемого содержания гумуса в
почве, переносили ее в коническую колбу емкостью 100 мл и приливали в нее по
каплям из бюретки 10 мл 0,4 Н раствора бихромата калия в разведенной серной
кислоте. Колбу нагревали до кипения в течении пяти минут. После этого осторожно
приливали дистиллированную воду, доведя объем в ней до 30-40 мл. Добавляли в
колбу 3-4 капель фенилантраниловой кислоты и титровали 0,2 Н раствором соли
Мора. Конец титрования определяли переходом вишнево-фиолетовой окраски в зеленую.
Содержание углерода определяли по формуле:
С = (А х Н1 - В х Н2) х 0,003 х 100 /Г, где
С - содержание углерода, %;
А - количество раствора бихромата калия, мл;
Н1 - нормальность раствора бихромата калия;
В - количество соли Мора, мл;
Н2 - нормальность раствора соли Мора;
,003 - величина мг-экв. углерода,
Г - навеска почвы, г.
После подсчета углерода вычисляют процентное содержание
гумуса:
гумус % = С%*1,724
На основании данных о кислотности почвы и процента содержания
гумуса построили диаграмму Рис 6.
Установка видовой принадлежности проводилась с помощью
определителей пауков В.П. Тыщенко, личинок жесткокрылых Е.А. Негробова,
насекомых европейской части СССР Б.М. Мамаева, жужелиц рода Carabus южной
Сибири В.Г. Шиленкова, определителю многоножек-костянок СССР Н.Т. Залеской,
определителю дождевых червей фауны России Т.С. Всеволодовой-Перель.
Помимо видовой идентификации, определялся также возраст
представителей почвенной биоты, подсчитывалось количество половозрелых и
неполовозрелых особей, принадлежность к трофическим группам. Половозрелость
определялась по внешним морфологическим признакам, у дождевых червей в первую
очередь - по наличию у них "пояска", у других животных - по стадии
развития.
Биомасса мезофауны определялась взвешиванием почвенных животных
с помощью электронных весов. При обработке результатов использовались
стандартные статистические методы (Лакин, 1990)
Анализ фаунистического сходства проводился через расчет
коэффициента Жаккара Кj
А и В - число видов в группировках, С - число общих семейств.
2.2
Физико-географическое описание окрестностей г. Ханты-Мансийска
Г. Ханты-Мансийск находится в бореально-лесной области
западно-сибирской равнины в климатическом отношении. В бореально-лесной области
климат имеет континетальный характер. Летний период характеризуется высокой
температурой и низкой влажностью.
Весной переход температуры через 0°С наблюдается в апреле. В
этот период облачность низкая, и суточная амплитуда температур относительно
широка, что определяется в основном радиационными факторами. В мае преобладают
дни с положительной температурой. Но повышение ее весной замедляется таянием
глубокого снежного покрова и постепенным размораживанием обширных болотных
пространств. Поэтому испарение оказывается ниже суммы выпадающих осадков, и
влажность почвы и воздуха и почвы остается высокой. Лишь при переходе
температуры +5°С (с середины мая) испарение начинает превышать поступление
осадков, и в этот период начинается подсыхание верхнего слоя почвы, что
обусловливает начало зоогенной активности. Переход к суточным температурам выше
10°С, совпадающий с прекращением весенних заморозков и началом вегетационного
периода в конце мая.
Первая половина лета сухая осадков немного, и испарение
превышает поступление осадков. В конце лета температура снижается, а выпадение
осадков повышается в связи с прохождением фронтов и высоким влагосодержанием
воздушных масс приходящих с юга. Поэтому вторая половина лета оказывается
избыточно влажной на фоне недостатка тепла. Летом температура колеблется в
среднем в пределах 10-20°С.
В сентябре появляются дни со сменой положительных и
отрицательных температур, в октябре число таких дней достигает 50%, в ноябре
устанавливается температура ниже - 10°С. Температура наиболее холодных дней
зимой ниже - 20°С, число таких дней в декабре-январе составляет треть от общего
периода.
Г. Хант-Мансийск относится к зоне средней тайги (Стриганова,
Порядина, 2005). Средняя тайга представлена елово-кедровыми лесами,
занимающими, дренированные склоновые позиции водоразделов и высокие приречные
террасы со слабоподзолистыми суглинистыми почвами (Рысин, Савельева, 2002). Эти
леса отличаются высокой продуктивностью, большой высотой (17 - 20 м) и
сомкнутостью (0,6 - 0,7). Для темнохвойных лесов в подзоне средней тайги создаются
оптимальные условия, поэтому они занимают широкий спектр местообитаний в
междуречьях (Крылов, 1961). Речные воды относятся к гидрокарбонатно-кальциевому
типу. На плоских равнинах, сложенных горизонтально залегающими рыхлыми
отложениями, (пески, глины, суглинки) распространены ельники и елово-кедровые
леса. Слабый дренаж поверхностного слоя почвы приводит к оторфыванию лесной
подстилки, развитию глеевых процессов, ухудшению состояния древостоя. В местах
с более расчлененным и возвышенным рельефом заболоченность снижается, а густота
и высота древостоя повышается. На дренированных увалистых равнинах развиваются
слабо-торфянистые подзолистые почвы под еловыми лесами.
На заболоченных почвах, занимающих значительную часть
территории, также развиваются елово-кедровые леса с мохово-кустрарничковым
покровом (брусника, черника, багульник, клыква). В составе покрова от
придолинных позиций к границам заболоченных водоразделов наблюдается сменяя
доминантов в напочвенном покрове гидрофитными формами и формирование долгомошно-сфагновых
типов темнохвойных лесов. На фоне доминирования зеленых мхов и таежных
кустарничков встречаются южно-таежные виды (кислица, седмичник, звездчатка
Бунге, папоротники).
Глава 3.
Результаты исследования
3.1
Результаты исследования мезофауны
Всего нами было обнаружено 20 таксонов, принадлежащим к 2
типам: тип кольчатые черви (2 вида), тип членистоногие (18 видов). Из них
наибольшим разнообразием отличался класс насекомые отряд жесткокрылые семейство
жужелицы (7 видов).
Список таксонов
Тип Кольчатые черви (Annelides)
Сем. Дождевые черви (Lumbricidae)
1. Eisenia nordenskioldi nordenskioldi (Eisen, 1879)
2. Пашенный червь Allolobophora caliginosa (Savigny, 1826)
Тип Членистоногие
Класс Многоножки (Myriapoda)
Сем. Костянки (Lithobiidae)
3. Monotarsobius curtipes (C. L. Koch, 1847)
4. Lithobius forficatus (Linnaeus, 1758)
Класс Паукообразные (Arachida)
5. Сем. Пауки-крабы (Thomisidae)
6. Сем. Пауки-волки (Licosidae)
. Сем. Пауки-скакуны (Salticidae)
Класс Насекомые (Insecta)
Отряд Полужесткокрылые (Hemiptera)
. Сем. Мезовелии (Mesoveliidae)
9. Сем. Хищнецы (Reduvidae)
Отряд Жесткокрылых (Coleoptera)
Сем. Жужелицы (Carabidae)
10. Pterostichus niger
11. Pterostichus lepidum Leske
. Calathus micropterus (Duftschmid,1812)
13. Pterostichus menalarius
. Harpalus serripes
. Pterostichus oblongopunctatus (Fabricius, 1787)
16. Pterostichus strenuus Panz
Сем. Стафилиниды (Staphylinidae)
17. Xantholinus linearis (Olivier, 1794)
Отряд Перепончатокрылые (Hymenoptera)
Семейство Муравьи (Formicida)
18. Formica rufa (Linnaeus, 1761)
19. Lasius flavus (Fabricius, 1781)
20. Camponotus vagus Scop.
Дождевые черви представлены (2 видами), преобладает Eisenia
nordenskioldi palida (Eisen, 1879), этот вид был отмечен как фоновый для
сибирской тайги (Перель, 1979), A. caliginosa предпочитает
теплообеспеченные территории. В классе многоножек следует отметить типичный
европейский вид Lithobius forficatus (Linnaeus, 1758), который
в связи с относительным смягчением климата в Южной Сибири стал продвигаться
на восток. Среди насекомых наибольшим разнообразием отличались представители
отряда жесткокрылых, представленные 1 семейством и 7 таксонами.
В естественных местообитаниях папоротниковом ельнике найдено
11 таксонов. В них доминировала Eisenia nordenskioldi nordenskioldi (Eisen,
1879), массовая доля которой составляет 29%, также значительный часть занимают
многоножки - 14% Monotarsobius curtipes (C. L. Koch, 1847). Имеет место
доля жужелиц и пауков-крабов по 10% (см. рис.1).

Рис. 1. Видовое разнообразие естественных местообитаний
на примере хвойной тайги, представленной папоротниковым ельником.
Смешанная тайга, возникающая на месте вырубок сокращает
таксономическое богатство с 11 до 8 видов (см. рис.2).
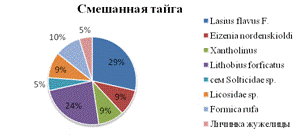
Рис.2. Видовое разнообразие вторичнонарушенных
местообитаний на примере смешанной тайги.
В качестве доминантов выходят семейство муравьи Lasius
flavus (Fabricius, 1781) - 29% и семейство костянки Lithobius forficatus
(Linnaeus, 1758), составляющие 24%. Снижается доля дождевых червей до 9%.
Меньшую же часть составляют пауки если в первом случае они составляли примерно
равное количество среди недоминантных видов.
На заливном лугу реки Щучье отмечено 8 таксоном среди них
преобладает семейство жужелицы птеростих красивый Pterostichus lepidum Leske
- 37%, и Harpalus serripes 26%. Кроме того значительная доля
Птеростиха черного - 18% (см. рис.3).
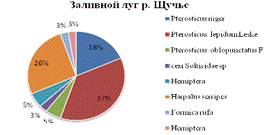
Рис.3. Видовое разнообразие естественных местообитаний
на примере заливного луга р. Щучье.
Исследование проведенное на заливном лугу под действием
рекреационной нагрузки показало что количество таксонов сокращается с 8 до 4.
Доминирующую позицию занимает Pterostichus niger - 40%, и хищные
стафилиниды Xantholinus (см. рис.4). На заливном лугу около
мусороперерабатывающего завода не было найдено ни одного таксона.
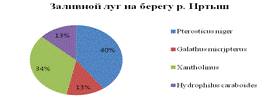
Рис. 4. Видовое разнообразие местообитаний, подверженных
рекреационной нагрузке, на примере заливного луга р. Иртыш.
Сравнительный анализ видового состава педобионтов разных
подзон и провинций на основании использования индекса общности фаун Жаккара
(Песенко, 1982) низкая 0 - 0,19 (максимально 1). Наибольшее сходство 0,19
наблюдается между естественными местообитаниями. Среднее значение 0,12 между
генетически родственными биотопами хвойная тайга и смешанная тайга (табл. 1).
Минимальное значение между естественными местообитаниями это заливные луга реки
Щучье и заливные луга под действием рекреационной нагрузки и под действием
мусороперерабатывающего завода.
Таблица 1. Сходство биотопов по фаунистическому составу
окрестностей г. Ханты-Мансийска
|
|
заливной луг р. Иртыш
|
заливной луг р. Шучье
|
Хвойная тайга
|
Смешанная тайга
|
заливной луг р. Иртыш около завода
|
|
заливной луг р. Иртыш
|
1
|
0,09
|
0,07
|
0,09
|
0
|
0,09
|
1
|
0, 19
|
0
|
0
|
|
Хвойная тайга
|
0,07
|
0, 19
|
1
|
0,12
|
0
|
|
Смешанная тайга
|
0,09
|
0
|
0,12
|
1
|
0
|
|
заливной луг р. Иртыш около завода
|
0
|
0
|
0
|
0
|
0
|
Максимальная плотность достигает на пойменном лугу р. Щучье.
Рекреационная нагрузка снижает показатель в 2,53 раза. Влияние
мусороперерабатывающего завода снижает этот показатель до нуля. Образование
вторичного сообщества на месте вырубки лесов оставляет этот показатель на
одинаковом уровне (см. рис.5).
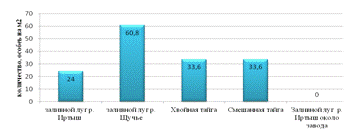
Рис. 5. Плотность почвенной мезофауны различных
местообитаний.
Биомасса приобретает максимальное значение в хвойной тайге
6,8 грамм на см2. Образование вторичного сообщества на месте вырубки
лесов снижает биомассу в 2,97 раза. Влияние рекреационной нагрузки снижает биомассу
в 2,05 раза. Влияние мусороперерабатывающего завода до исчезновения (рис. 6).
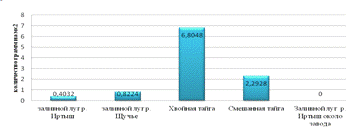
Рис.6. Биомасса педобионтов различных местообитаний.
На всей территории округа педобионты локализуются в верхних
слоях (Стриганова, Порядина 2005). Почвенные организмы в основном занимают
подстилку и верхний слой (0-10 см) в тайге, в заливных лугах педобионты
сосредоточены на подстилке, возможно это объясняется близким расположением
грунтовых вод (рис.7).
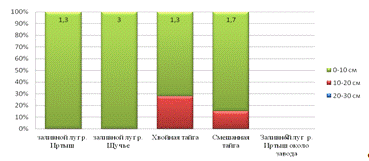
Рис.7. Вертикальное распределение педобионтов в почве.
Соотношение почвенной мезофауны по возрастным состояниям
характеризуется стабильным типом возрастного спектра в возделываемых и
заброшенных агроценозах, т.е. преобладанием неполовозрелых (личиночных стадий)
и коконов. Регресивным типом, т.е. увеличением численности половозрелых особей
(Стриганова, 2005). Заливные луга характеризуются полностью регрессивным типом,
возможно это объясняется тем, что стадии кокона трудно обнаруживаются (рис.8).
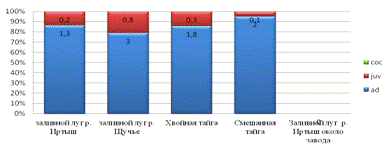
Рис.8. Возрастной состав педобионтов.
3.2
Результаты исследование почвы
При сборах данных было обработано 10 проб почвы.
Рассматривались следующие показатели - механический состав, водопрочность,
водопроницаемость, анализ водной вытяжки, РН и гумус.
Рис.9. Механический состав почв.
Доля наиболее ценной с агрономической точки зрения фракции (1
- 5 мм) с учётом всех слоёв почвы максимальна в естественных биотопах - хвойная
тайга и заливной луг на берегу р. Щучье (см. рис.9). Вторичное сообщество,
рекреационная нагрузка, а также действие завода приводит к увеличению доли
крупных фракций и пыли, особенно в нижних горизонтах; снижение доли ценных
фракций в 1,3 в первом случае и в 1,8 во втором.
Водопрочность крупных агрегатов исследуемых образцов
колеблется от 0 до 100%. На всех площадках наблюдается уменьшение водопрочности
нижних горизонтов. Наибольшей водопрочностью отличаются агрегаты почв
естественных местообитаний - хвойной тайги в нижнем горизонте. Учитывая, что
способность противостоять размывающему действию воды благотворно влияет на
качество, почвы естественных биотопов березовых лесов и луга сохраняют
биологически оптимальный механический состав. Тогда как структуры почв
приусадебных участков, обладающие более низким уровнем водопрочности, склонны к
разрушению при увлажнении, в результате чего ухудшается их водно-воздушный
режим (см. рис.10).
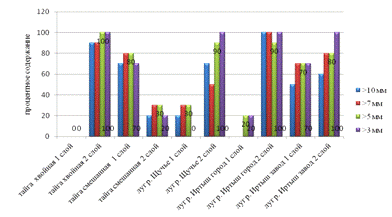
Рис.10. Показатели водопрочности.
Таким образом, особенно наглядно прослеживается зависимость
между установленными показателями водопроницаемости, водоемкости и структурным
составом исследуемых образцов почв: сохранившие оптимальный механический состав
почвы естественных биотопов имеют более качественные показатели физических
характеристик.
В естественных биотопах проницаемость больше в первом слое,
нежели во втором, то есть первый слой удерживает больше воды чем второй.
Образование вторичного биотопа не изменяет этой тенденции. Биотоп, под
действием рекреационной нагрузки, изменяет ситуацию. Почва первого слоя снижает
свою способность удерживать влагу. Первый слой луга, находящегося под действием
мусороперерабатывающего завода не способен удерживать влагу. Это явление может
объяснять отсутствие на этом лугу педобионтов и произрастание на нем сорняковых
видов растительности (см. табл.2).
Таблица 2. Показатели водопроницаемости.
|
|
время появления первой капли. С
|
время истечения последней капли. С
|
количество вытекшей воды
|
количество задержанной воды
|
|
тайга хвойная 1 слой
|
15 сек
|
1 мин
|
50
|
50
|
|
тайга хвойная 2 слой
|
5 сек
|
3 мин 30 с
|
70
|
30
|
|
тайга смешанная 1 слой
|
10 с
|
45 с
|
40
|
60
|
|
тайга смешанная 2 слой
|
1 мин 12 с
|
9 мин 28 с
|
60
|
40
|
1 мин
|
11 мин
|
50
|
50
|
|
луг р. Щучье 2 слой
|
50 с
|
7 мин
|
60
|
40
|
|
луг р. Иртыш город 1 слой
|
1 с
|
35 с
|
80
|
20
|
|
луг р. Иртыш город 2 слой
|
5 с
|
1 мин 30 с
|
60
|
40
|
|
луг р. Иртыш завод 1 слой
|
15 с
|
3 мин 15 с
|
100
|
0
|
|
луг р. Иртыш завод 2 слой
|
35 с
|
5 мин 55 с
|
60
|
40
|
Сопоставление результатов анализа структуры, водопрочности и
водопроницаемости образцов почв позволяет сделать вывод, что почва нарушенных
местообитаний в большей степени деструктурирована, имеет ярко выраженный
непропорциональный состав: с одной стороны глыбистый характер, с другой -
распыленность. Так же почвы вторичнонарушенных биотопов и биотопов, находящихся
под действием нугрузки, не способны удерживать свою структуру под действием
воды, и обладают большой влагоудерживающей способностью, что негативно
сказывается на качестве данных почв как среды обитания: они характеризуются
слабой дренажной системой, наличием излишней влаги, застойными явлениями. Эти
результаты являются последствием негативного воздействия на почвы в ходе их
механической обработки.
При воздействии воды на почву большинство из образующихся
минеральных солей растворяются и переходят в водную вытяжку, в которой они
могут быть определены соответствующими методами. Таким образом, по данным
анализа водной вытяжки из почвы можно судить о санитарном состоянии почвы, т.е.
о характере и степени загрязнения почвы органическими веществами, интенсивности
их минерализации и завершенности процесса самоочищения почвы. Анализ водной
вытяжки позволяет определить уровень и тип естественного засоления почв (Почва
и почвообразование, 1988).
Качественный анализ водной вытяжки показал что, естественный
биотоп - хвойная тайга и вторичнообразованное вообщество - смешанная тайга,
находятся в состоянии вторичного засоления. Вторичное засоление хлоридами в
первом слое испытывают также и остальные биотопы, самое высокое значение
достигает на заливном лугу около мусороперерабатывающего завода (до 50 мл/л).
Действие рекреационной нагрузки и мусороперерабатывающего завода приводит к
засолению хлоридами и второго слоя почв (см. табл.3). Это может объяснить то, что
педобионты были найдены только в 1 слое. На заливном лугу около завода не
найдено ни одного таксона.
Таблица 3. Показатели водной вытяжки.
|
Биотоп
|
Содержание хлоридов (мл/л)
|
Содержание сульфатов (мл/л)
|
Содержание кальция (мл/л)
|
|
тайга хвойная 1 слой
|
1,0-10
|
1,0-10
|
0
|
|
тайга хвойная 2 слой
|
0
|
0
|
0
|
|
тайга смешанная 1 слой
|
1,0-10
|
1,0-10
|
0
|
|
тайга смешанная 2 слой
|
0
|
0
|
0
|
|
луг р. Щучье 1 слой
|
1,0-10
|
0
|
0
|
|
луг р. Щучье 2 слой
|
0
|
0
|
0
|
|
луг р. Иртыш город 1 слой
|
1,0-10
|
0
|
0
|
|
луг р. Иртыш город 2 слой
|
1,0-10
|
0
|
0
|
|
луг р. Иртыш завод 1 слой
|
0
|
1,0-10
|
|
луг р. Иртыш завод 2 слой
|
1,0-10
|
0
|
0
|
Уровень рН почв естественного заливного луга и луга, под
действием рекреационной нагрузки близок к нейтральному показателю и в среднем
составляет рН 6,27 и 6,08 соответственно. Почвы луга под действием
рекреационной нагрузки имеет слабощелочную реакцию рН 7,2. Хвойная тайга
характеризуется слабокислой рН 5,53. Почвы вторично-образованного биотопа и
луга под действием завода смещены в более кислую сторону 4,43 и 4,27
соответственно. Полученные данные уровня реакции среды можно считать нормой для
черноземных почв (Мищенко, 1991), и оптимальными условиями для мезофауны
исследуемых биотопов (см. рис.11).
Средним показателем содержания гумуса для таежных почв
являются 2 - 4 %. Полученные нами данные содержания гумуса в таежных лесах
превышают норму (среднее 7,5), что можно объяснить характером почвы и условиями
процесса ее формирования (разнотравье, увлажнение, теплообеспеченность). Во
вторых слоях таежных участков очень низкое содержание гумуса 0,18 и 0,9, так
как эти почвы глинистые. Гумус в естественном луге имеет низкое значение 1,5 и
1,24. Луг, находящийся под действием завода, также не изменяет тенденцию. На
лугу, под действием рекреационной нагрузки, происходит увеличение доли гумуса в
5,5 раз. Процент содержания гумуса в почве зависит от водопроницаемости и
водопрочности фракций, согласно показателям этих характеристик можно
предположить, что гумус поля подвержен вымыванию в нижние слои. Кроме того,
большая часть органического вещества, продуцируемая агроценозом, не включается
в оборот веществ, а выводится из него. Эти факторы неизбежно ведут к уменьшению
содержания гумуса в почве искусственно созданного сообщества.
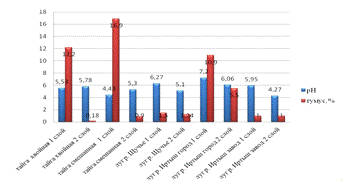
Рис.11. Содержание РН и процентное соотношение гумуса в
различных биотопах.
Выводы
1. Почвы ненарушенных биотопов окрестностей г.
Ханты-Мансийска имеют оптимальный процент экологически ценной фракции,
качественные физико-химические характеристики, высокий процент гумуса,
слабощелочную реакцию. Почва нарушенных биотопов деструктурирована, подвержена
вымыванию, испытывает вторичное засоление, содержание гумуса ниже нормы
черноземных почв или превышает ее во много раз.
2. Исследованная почвенная мезофауна относится к 8
семействам, обнаружено 25 таксонов, принадлежащих к 2 типам: тип кольчатые
черви (2 вида), тип членистоногие (18 видов). Из них наибольшим разнообразием
отличался класс насекомые, отряд жесткокрылые, семейство жужелицы (7 видов).
. Оценка вертикального распределения мезофауны в
почвах показала концентрацию педобионтов в верхних слоях почвы на лугах. В
таежных зонах (хвойная и смешанная тайга) отмечалось проникновение в нижние
слои.
. Фаунистическое сходство, рассчитанное по индексу
Жаккара, низкое от 0 до 0, 19. Максимально между естественными местообитаниями,
а также между генетически родственными биотопами.
. Максимальная плотность на пойменном лугу р. Щучье.
Рекреационная нагрузка снижает в 2,53 раза, влияние завода снижает до 0.
Биомасса колеблется от 0,81 до 13,6. Максимальное значение в хвойной тайге, в
2,97 раза снижает образование вторичного сообщества на месте вырубки,
рекреационная нагрузка снижает биомассу в 2,05 раза.
. При усилении антропогенного воздействия на биотоп
уменьшается биологическое разнообразие мезофауны. Нарушение структуры почвы и
разрушение естественных растительных сообществ ведет к обеднению почвенного
населения в количественном и качественном аспекте.
Библиографический
список
1. Акимушкин
И.А. Мир животных: беспозвоночные. - М.: Молодая гвардия, 1992. - 365 с.
2. Атлавините
О. Влияние дождевых червей на агроценозы. - Вильнюс: Мокслас, 1990. - 179 с.
. Балуев
В.К. Дождевые черви основных почвенных разностей Ивановской области //
Почвоведение. - 1950. - № 4. - С.219-227.
. Богатов
В.В., Стороженко С.Ю., Баркалов В.Ю., Холин С.К., Макарченко Е.А., Прозорова
Л.А. Биогеография острова Сахалин на примере распространения наземной и
пресноводной биоты // Теоретические и практические проблемы изучения сообществ
беспозвоночных: памяти Я.И. Старобогатова / под ред.А.И. Кафанова, П.В. Кияшко,
Б.И. Сиренко. - М.: Товарищество научных изданий КМК, 2007. - С. 193-224.
. Бызова
Ю.Б. Дыхание почвенных беспозвоночных // Экология почвенных беспозвоночных. -
М.: Наука, 1973. - С. 3-40.
. Воробейчик
Е.Л. Население дождевых червей (Lumbricidae) лесов Среднего Урала в условиях
загрязнения выбросами медеплавильных комбинатов // Экология. - 1998. - № 2. -
С. 102-108.
. Всеволодова-Перель
Т.С. Дождевые черви фауны России: Кадастр и определитель. - М.: Наука, 1997. -
102 с.
. Ганин
Г.Н. Почвенные животные Уссурийского края. - Владивосток-Хабаровск: Дальнаука,
1997. - 160 с.
. Гиляров
М.С. Методы количественного учета почвенной фауны // Почвоведение. - 1941. - №
4. - С.47-76.
. Гиляров
М.С. (ред.). Определитель обитающих в почве личинок насекомых. - М.: Наука,
1964. - 920 с.
. Гиляров
М.С. Учет крупных почвенных беспозвоночных (мезофауны) // Методы почвенных
зоологических исследований. - М.: Наука, 1975. - С.12-29.
. Гиляров
М.С., Криволуцкий Д.А. Жизнь в почве. - М.: Молодая гвардия, 1985. - 191 с.
. Голованова
Е.В. Население дождевых червей (Lumbricidae)"Птичьей Гавани" в
условиях загрязнения свинцом // Изучение экосистемы природного парка
"Птичья Гавань". - Омск: Изд-во ОмГПУ, ООО "Издательский дом
"Наука", 2003. - С.99-105.
. Голованова
Е.В. Популяции дождевых червей придорожных полос в условиях загрязнения свинцом
// Экологические механизмы динамики и устойчивости биоты. - Екатеринбург:
Академкнига, 2004. - С.54-60.
. Догель
В.А. Зоология беспозвоночных. - М.: Высшая школа, 1981. - 606 с.
. Иванов
А.В. Система животного мира // Протисты: Руководство по зоологии. - СПб: Наука,
2000. - С.97-114.
. Иоганзен
Б.Г. Основы экологии. - Томск: Изд-во ТГУ, 1959. - 390 с.
. Звягинцев
Д.Г., Бабьева И.П., Зенова Г.М. Биология почв. - М.: Изд-во Московского
университета, 2005. - 445 с.
. Зражевский
А.И. Дождевые черви как фактор плодородия лесных почв. - Киев: Изд-во АН
УкрССР, 1957. - 271 с.
. Криволуцкий
Д.А. Животный мир почвы. - М.: Знание, 1969. - 47 с.
. Криволуцкий
Д.А. Почвенная фауна в экологическом контроле. - М.: Наука, 1994. - 269 с.
. Курчева
Г.Ф. Роль животных в почвообразовании. - М.: Знание, 1973. - 64 с.
. Курчева
Г.Ф. Роль почвенных животных в разложении и гумификации растительных остатков.
- М.: Наука, 1971. - 156 с.
. Малевич
И.И. Собрание и изучение дождевых червей - почвообразователей. - М.: Изд-во АН
СССР, 1950 - 37 с.
. Орлов
В.А. Элективные курсы по физике и их роль в организации профильного и
предпрофильного обучения // Физика в школе. - 2003. - № 7. - с.36-38.
. Перель
Т.С. Жизненные формы дождевых червей (Lumbricidae) // Журнал общей биологии. -
1975. - Т.36. - № 2. - С.189-202.
. Перель
Т.С. Различия организации разных представителей дождевых червей в связи с
особенностями их экологии // Адаптация почвенных животных к условиям среды. -
М.: Наука, 1977. - С.129-144.
. Перель
Т.С. Распространение и закономерности распределения дождевых червей фауны СССР.
- М.: Наука, 1979. - 272 с.
. Петунин
О. Элективные курсы для профильного биологического образования // Народное образование.
- 2003. - № 9. - С.107-111.
. Пирогова
Т.И. Полевое и лабораторное изучение почв. - Омск, 2004. - 128 с.
. Пирогова
Т.И. Практикум по почвоведению. Омск: Изд-во ОмГПУ, 2006 - 41с
. Побережная
Т.М. Ландшафтно-геохимические исследования на Сахалине // Вестник ДВО РАН. -
2006. - № 1. - С.109-114.
. Польский
Б.Н. Практические занятия и межсессионные задания по географии почв с основами
почвоведения. - М.: Просвещение, 1972. - 188 с.
. Радкевич
В.А. Экология. - Мн.: Вышейша школа, 1998. - 159 с.
. Раковская
Э.М., Давыдова М.И. Физическая география России. Ч.2. Азиатская часть, Кавказ и
Урал. - М.: Владос, 2001. - 304 с.
. Рягин
С.Н. Проектирование содержания профильного обучения в современной школе:
Монография. - Омск: ООИПРО, 2003. - 155 с.
. Синько
Т.П. Элективные курсы // Естественнонаучное образование в школе: теория и
практика. - 2005. - № 1. - С.5-9.
. Соколов
А.А. Значение дождевых червей в почвообразовании. - Алма-Ата: 1956. - 261 с.
. Стриганова
Б.Р. Питание почвенных сапрофагов. - М.: Наука, 1980. - 243 с.
. Стриганова
Б.Р. Порядина Н.М. Животное население почв бореальных лесов Западно - Сибирской
равнины. М.: Товарищество научных изданий КМК. 2005 г.234с.
. Фасулати
К.К. Полевое изучение наземных беспозвоночных. - М.: Высшая школа, 1971. - 424
с.
. Чекановская
О.В. Дождевые черви и почвообразование. - М.: Изд-во АН СССР, 1960. - 203 с.