Биоразнообразие мицелиальных грибов
Введение
Грибы являются компонентами практически всех морских
сообществ. Они разлагают органическое вещество автохтонного и аллохтонного
происхождения. Они являются сапротрофами, паразитами, патогенами и симбионтами
морских животных и растений. Способны продуцировать вторичные метаболиты:
антибиотики, токсины и витамины.
Исследование мицелиальных грибов, ассоциированных с
двустворчатыми моллюсками, имеет как фундаментальный интерес, так и большое
практическое значение. С одной стороны, изучение таксономического состава
мицелиальных грибов - ассоциантов моллюсков и их распределения на поверхности
раковины и во внутренних органах животных является основой для выяснения
характера взаимоотношений грибов с моллюсками, установления их трофических и
топических связей и, в конечном итоге, определения роли мицелиальных грибов в
метаболизме двустворчатых моллюсков. Так как на настоящий момент нет никаких
данных по этой теме. С другой стороны, многие виды мицелиальных грибов являются
условно-патогенными организмами, вызывая микозы растений, животных и человека.
Кроме того, многие штаммы мицелиальных грибов продуцируют микотоксины,
способные вызывать микотоксикозы животных и человека.
Практический интерес обусловлен тем, что двустворчатые
моллюски - важнейший объект промысла и марикультуры, используются в пищевых,
лечебно-профилактических и других коммерческих целях. В связи с этим,
исследования патогенных и токсикогенных грибов, заселяющих внутренние органы
съедобных моллюсков, необходимы для безопасного использования моллюсков в пищу,
для предотвращения токсикоинфекций людей при употреблении зараженных грибами
морепродуктов, а также для успешного развития хозяйств марикультуры.
Целью нашей работы является изучение биоразнообразия
мицелиальных грибов, ассоциированных с двустворчатыми моллюсками.
Для достижения этой цели мы поставили следующие задачи:
1. Получить штаммы мицелиальных грибов - ассоциантов
двустворчатых моллюсков Modiolus modiolus, Crenomytilus grayanus, Mizuhopecten
yessoensis и Anadara broughtoni.
2. Изучить видовой состав мицелиальных грибов,
ассоциированных с двустворчатыми моллюсками Modiolus modiolus, Crenomytilus
grayanus, Mizuhopecten yessoensis и Anadara broughtoni.
3. Изучить распределение грибов во внутренних органах
моллюсков Modiolus modiolus, Crenomytilus grayanus и Mizuhopecten yessoensis.
. Определить степень расхождения грибных комплексов
ассоциированных с двустворчатыми моллюсками.
. Дать оценку биоразнообразию мицелиальных грибов,
ассоциированных с двустворчатыми моллюсками зал. Восток.
1. Мицелиальные грибы в морских сообществах
Грибы весьма разнообразны по внешнему виду, местам обитания и
физиологическим функциям. Однако у них есть и общие черты. Основой
вегетативного тела грибов является мицелий, или грибница, представляющая собой
систему тонких ветвящихся нитей, или гиф, находящихся на поверхности субстрата,
где живет гриб, или внутри него. Обычно грибница бывает весьма обильна, с
большой общей поверхностью. Через нее осмотическим путем происходит всасывание
питательных веществ. У грибов некоторых классов и многих организмов,
традиционно изучающихся микологами, в последние годы относящихся к царству
Chromista, грибница не имеет перегородок (ценоцитный мицелий).
Обширную группу составляют водные грибы. Они обитают во
всевозможных водоемах: океанах и морях, озерах, реках, болотах, термальных
источниках. По происхождению водные грибы можно разделить на две подгруппы
(Дудка, 1985): а) первичноводные (появились в водной среде и остались в ней);
б) вторичноводные (в ходе эволюции мигрировали на сушу, а затем вновь вернулись
в водную среду). Ко второй группе относятся водные гифомицеты, многие
аскомицеты и некоторые базидиомицеты.
1.1 Эколого-физиологическая группа морских
грибов
Наряду с истинными водными грибами, весь цикл которых связан
с водой, в водоемах в больших количествах обнаруживаются виды, которые способны
развиваться как в наземных, так и в водных местообитаниях. Они получили
название водно-наземных и водно-воздушных грибов (Дудка, Вассер, 1987).
Что касается морских грибов, то выделение этой группы часто
основывалось на их способности расти при определенных концентрациях морской
воды (Johnson, Sparrow, 1961; Tubaki, 1969). Было показано, что морские грибы
не могут быть определены строго на физиологической основе, и мы используем более
широкое, экологическое определение (Kohlmeyer, Kohlmeyer, 1979): Облигатные
морские грибы - грибы растущие и размножающиеся исключительно в морских и
эстуарных местообитаниях; факультативные морские грибы - грибы пресноводные или
наземные, способные расти (и иногда также образовывать репродуктивные
структуры) в морской среде.
Простое выделение гриба из морских образцов не доказывает,
что этот вид активен в морской среде. Такие грибы, возможно, покоятся в виде
спор или фрагментов гиф, пока среда не станет благоприятной для прорастания и
развития. Действительным критерием для определения морских грибов может служить
способность прорастать и формировать мицелий в естественных морских условиях.
Эти условия могут варьировать в зависимости от вида. Некоторые грибы требуют
постоянного погружения, тогда как другие, например, виды, встречающиеся в
песчаных пляжах, нуждаются в большем поступлении кислорода для прорастания
(Kohlmeyer, Kohlmeyer, 1979).
1.2 Видовое богатство морских грибов
Водные грибы находятся в сравнительно более стабильных
условиях существования, чем наземные: вода медленно остывает и медленно
нагревается, она хорошо растворяет и выводит различные продукты
жизнедеятельности гидробионтов, защищает их от прямого действия солнечной
радиации. Следовательно, водные грибы не испытывают резких колебаний основных
факторов среды - температуры, минерального и газового состава.
Органические субстраты, такие как водоросли, растения соленых
болот, растительная подстилка, сконцентрированы в основном вдоль побережий, где
они обеспечивают питание для грибов. Открытый океан - это грибная пустыня, где
могут быть найдены только дрожжи и низшие грибы, ассоциированные с планктонными
организмами и пелагическими животными (Kohlmeyer, Kohlmeyer, 1987). В
противоположность наземным местообитаниям, стабильные условия океана и
сравнительно небольшое количество хозяев и субстратов в морской воде не
оказывали достаточного выборочного давления в процессе эволюции, чтобы привести
к образованию большого количества различных типов грибов. И действительно, из
морских местообитаний известно около 1500 видов грибов (Hyde et al., 1998),
т.е. менее 1% от числа наземных видов.
1.3 Образ жизни и распространение морских
грибов
Изучение экологии такой специфической группы организмов, как
морские грибы, началось сравнительно недавно. Однако отдельные сообщение о
случайных находках грибов - мицелия и репродуктивных структур - начали
появляться в литературе уже в середине 19 столетия. Так, Кёлликер (Kölliker, 1860) сообщал о «трубочках», найденных им в различных
морских животных - губках, кораллах, моллюсках, рыбах и др. По его
предположению, эти организмы относились к «неклеточным грибам».
В отношении грибов, обитающих в воде, выяснены следующие
закономерности: распространение грибов в океане не зависит от широтности района
обитания - определенные виды грибов были обнаружены как в водах Гренландии и
Антарктики, так и в тропических районах у берегов Бразилии, Индии и островов
Тихого океана. Споры грибов и обрывки мицелия обнаружены в пробах воды, начиная
с поверхности вплоть до больших глубин (4610 м), даже в таком водоеме, как
Черное море (до 2000 м). Выделенные формы принадлежали к анаморфным грибам и
представителям царства Chromista. Плотность грибных пропагул убывала с
удалением от суши и с увеличением глубины (Артемчук, 1981).
Как сказано выше, первые сведения о присутствии грибов на
морских животных появились в 1860 году (Kölliker). Грибы - паразиты
рыб (сельди, горбуши, камбалы), а так же промысловых моллюсков, изучались чаще
других, что обусловлено хозяйственными потребностями (Артемчук, 1981). Большая
часть их относились к порядку Saprolegniales (Oomycota, Chromista).
До последнего времени имелись лишь спорадические сведения о
грибах других морских животных. Мортенсен (Mortensen, 1909) описал болезнь
иглокожих родов Rhynochocydaris и Ctenocidaris, которая вызывалась
грибоподобным организмом Echinophyces mirabilis. Позднее была обнаружена
способность грибов поражать иглы морских ежей Diadema antillarum (Mortensen,
1940) и Strongylocentrotus franciscanus (Johnson, Charman, 1970).
С недавнего времени возобновился интерес исследователей к
грибам, ассоциированными с морскими животными. Было изучено биоразнообразие
грибов мшанок восточного побережья Австралии (Sterflinger et al., 2001), губок
северной Атлантики (Höller et al., 2000) и
голотурий российского побережья Японского моря (Pivkin, 2000). У побережья
Белиза (Карибское море) были обнаружены и описаны два новых вида рода
Koralionastes, заселяющие губки и кораллы (Kohlmeyer, Volkman-Kohlmeyer, 1990).
Намикоши с соавторами (Namikoshi et al., 2002) изучали грибы, выделенные из
губок коралловых рифов индонезийских островов Палау и Банакен.
В последние годы выявлено, что факультативные морские грибы
вызывают заболевания морских животных: Fusarium solani Shlegel - болезнь
лобстеров возле побережья Великобритании (Alderman, 1981); Aspergillus
fumigatus Fres. - болезнь мягких кораллов в Карибском море (Kim et al., 2000).
Стерфлингер с соавторами (Sterflinger et al., 1999) изучали грибные инфекции колониальных
морских беспозвоночных в районе острова святого Хуана (Вашингтон, США). Их
исследования показали, что разрастание тканей мшанок - это защитная реакция на
поражение грибами родов Penicillium, Cladosporium и др. Токсины факультативных
морских грибов были обнаружены в моллюсках, культивируемых на Атлантическом
побережье Аргентины (Sallenave et al., 1999). Позднее продуценты этих токсинов
были выделены в чистую культуру из этих же моллюсков (Sallenave-Namont et al.,
2000). Недавно факультативные морские грибы были обнаружены в тканях
дальневосточного двустворчатого моллюска Corbicula japonica Prime (Зверева и
др. 2002).
2. Субстраты животного происхождения, на которых обитают
морские грибы
Морские грибы, обитающие на животных, редки, большинство из
них сапротрофы, заселяющие экзоскелеты, раковины или защитные трубки. Такие
субстраты животного происхождения состоят из целлюлозы, хитина, кератина и
карбоната кальция с органической матрицей (Kohlmeyer, Kohlmeyer, 1979; Пивкин и
др., 2006).
.1. Туницин - целлюлоза животных, которая напоминает
целлюлозу растений. Штамм гриба Halosphaeria quadricornuta был изолирован с
оболочников (Ritchie, 1954). Кроме того, Ceriosporopsis circumvestita и
Pleospora gaudefroyi хорошо растут на кусочках туницина из Phallusia
mammillata, на разлагающейся древесине бальзы и целлюлозной пленке (Kohlmeyer,
1962b; Kohlmeyer, Kohlmeyer, 1966). Эти наблюдения показывают, что грибы могут
разрушать целлюлозу туникат в природе.
.2. Хитин - содержащий азот полисахарид, входит в состав
экзоскелетов многих морских животных. Найдены септированные гифы
неидентифицированного гриба в экзоскелетах лангуста Palinurus elephas на
западном побережье Британских островов (Alderman, 1973). Мицелий интенсивно
повреждал покровы, проникал через них и развивался в тканях мускулов.
Бактериальный некроз следовал за грибной инфекцией.
.3. Кератин - содержащий серу белок, входит преимущественно в
состав эпидермы беспозвоночных животных. Сведения о разложении кератина в
морской среде обитания незначительны. Единственное сообщение о разрушении
кератиноподобных веществ морскими грибами принадлежит Кольмейеру (Kohlmeyer,
1972a), который нашел аскомицеты в трубках полихет Chaetopterus variopedatus.
Кожистые защитные трубки животного часто выбрасывает на берег. Трубки,
собранные у Калифорнийских берегов, содержат гифы и аскокарпы Lulworthia sp.,
которые расщепляют субстрат на многочисленные тонкие слои. Аскокарпы
поверхностные. Мицелий распространяется по всей трубке, всегда находится в
близком контакте с субстратом. Это указывает на то, что ферменты действуют
только при непосредственном контакте между гифами и субстратом. Наблюдения
Кольмейера демонстрируют, что облигатные морские грибы способны усваивать
кератиноподобные вещества, но необходимы дальнейшие исследования для выяснения
степени их участия в разрушении таких веществ в морской среде (Пивкин и др.,
2006).
.4. Карбонат кальция с органической матрицей. Только
несколько видов морских грибов встречались на известковых субстратах, таких как
раковины моллюсков и балянусов. Хенк (Hőhnk, 1967, 1969) показал
септированные гифы и плодовые тела неидентифицированных грибов с фрагментов
раковин. Известковые стенки пустых трубок корабельных червей, особенно в
тропических и субтропических водах, часто заселяются аскомицетами и анаморфными
грибами (Kohlmeyer, 1969d). После их внедрения твердый субстрат становится
мягким, ломким и приобретает коричневый цвет. В конечном счете, конидии или
аскокарпы развиваются в пределах известкового слоя и прорываются на
поверхность. Иногда плодовые тела развиваются в древесине под трубками,
формируя длинные шейки, которые проникают в известковый слой (Пивкин и др.,
2006). Аскомицеты Halosphaeria quadricornuta и Н. salina, а также анаморфные
грибы Cirrenalia pygmea, Humicola alopallonella и Periconia prolifica
встречаются на таких субстратах (Kohlmeyer, 1969d). Гифы и конидии С. pygmea и
Н. alopallonella были найдены только на поверхности известковых трубок, тогда
как другие грибы развивались внутри этих субстратов (Пивкин и др., 2006).
Аскомицет Pharcidia balani, часто вступающий в ассоциации с
различными видами микроскопических морских водорослей, регулярно находят в
раковинах живых моллюсков и усоногих раков литоральной зоны. В лихенологической
литературе ассоциацию рассматривают как лишайник Pyrenocollema sublitorale.
Черные аскокарпы и пикниды Р. balani наполовину погружены в известковый
субстрат (Kohlmeyer, Kohlmeyer, 1979). Гифы растут всюду по внешнему слою
раковин и вызывают размягчение известкового субстрата. Когда старые плодовые тела
распадаются, они оставляют вмятины в поверхности раковины, делая ее губчатой и
изъеденной. Поскольку гриб встречается только погруженным во внешнюю сторону
раковин, это не вредит животному. Вероятно, грибы используют конхиолин -
органическую матрицу раковин, тогда как кристаллы СаСО, или аморфный карбонат
кальция, не повреждаются.
Нити биссуса Mytilus galloprovincialis из Средиземноморья
регулярно содержат грибные гифы (Vitellaro-Zuccarello, 1973). Они состоят из
белковых веществ, включая некоторое количество коллагена, и довольно устойчивы
к химическому и физическому разрушению. Грибы не удалось идентифицировать.
Ультраструктура гиф предполагает близость гриба к аскомицетам
(Vitellaro-Zuccarello, 1973).
3. Грибные болезни
Всегда считалось, что возбудителями болезней морских животных
являются преимущественно страминипилы (Alderman, 1976). В последнее время стали
появляться сведения о болезнях, вызываемых вторичными морскими грибами.
Например, Fusarium solani (Mart.) Sacc. вызывает болезни лобстеров (Alderman,
1981). Недавние исследования заболевания мягких кораллов Gorgonia в Карибском
море показали, что вызывает болезнь Aspergillus sydowii (Bainier et Sartory)
Thom et Church. (Geiser et al., 1998). Penicillium corylophilum и Cladosporium
sphaerospermum повреждают почки и плавательный пузырь у красных люцианов
(Lutjanus campechanus) (Blaylock et al., 2001).
Токсичные метаболиты Trichoderma sp. и Penicillium sp. были
обнаружены в моллюсках (Sallenave et al., 1999). Позже из них были выделены и
продуценты этих токсинов (Sallenave-Namont et al., 2000).
4. Отношения животных и грибов
.1 Моллюски-сверлильщики
Двустворчатые моллюски семейств Teredinidae и Pholadidae -
важные разрушители древесины в морской среде обитания (Turner, 1971). Ранние
сообщения о грибах на моллюсках-древоточцах относятся ко второй половине XIX в.
(Kolliker, 1860a, b). Грибы не участвуют в пищеварительном процессе взрослых
древоточцев, потому что грибные гифы не найдены в их пищеварительных трактах и
не встречаются в древесине около скопления раковин. Однако грибы причастны к
поселению личинок. Личинки выходят из материнского животного, высвобождаясь
через сифон, и рассеиваются с потоками воды (Lane, 1961). Личинки древоточцев
плавают хаотично с помощью паруса, имеющего форму зонтика и составленного из
длинных ресничек. Затем они прикрепляются к любому субстрату, с которым они
входят в соприкосновение. Нога служит для исследования поверхности субстрата в
поиске подходящего места для внедрения. На этой стадии раковины личинок
неотвердевшие и не способны проникнуть в свежую древесину. В том случае, если
древесная поверхность размягчена под действием целлюлозолитических грибов или
бактерий, личинка начинает очищать субстрат, формируя в течение нескольких
минут вокруг насыпь кусочков. Личинки не привлекались древесиной или грибными
экстрактами, натрий-гидроксидом, яблочной и соляной кислотами, мицелием или
древесиной, как с грибами, так и без них. Однако они прикреплялись к
инфицированной грибом древесине, когда находились в контакте с ней. Результаты
экспериментов с Limnoria tripunctata (Becker et al, 1957; Becker, 1959)
показали, что этот вид не способен заселять неповрежденную древесину.
В свою очередь, некоторые сообщения указывают, что Limnoria
оказывает положительный эффект на рост морских грибов. Корт (Corte, 1975)
отмечал, что аскокарпы Lulworthia sp. развиваются только после того, как
древесина перфорирована раками-древоточцами. Конидии Cirrenalia macrocephala
образуются предпочтительно в оставленных туннелях Limnoria (Kohlmeyer, 1958, 1962а).
Вполне возможно эндозоотическое распространение конидий грибов моллюском и
другими населяющими древесину беспозвоночными, поскольку они сохраняют
жизнеспособность после прохождения через пищеварительный тракт (Kohlmeyer et
al., 1959), подобно амброзиевым грибам в наземных условиях. Изучение
совместного заселения древесины древоточцами Martesia striata и Lyrodus
pedicellatus и грибами показало специфичность заселения древесины этими
моллюсками и грибами Periconia prolifica и Antennospora quadricornuta
(Vishwakiran et al., 2001).
.2 Амфиподы солевых маршей
Исследования на пресноводных детритофагах показали, что
животные предпочитают листья, колонизированные микроорганизмами, особенно
области листа с высокой концентрацией гиф, по сравнению с недавно упавшими,
незараженными листьями (Barlocher, Kendrick, 1973, 1975а, б). Подобная ситуация
может ожидаться в литоральных соленых болотах, где накапливается большее
количество растительного детрита, служащего пищей для детритофагов. Пищевая
ценность детрита из растений болот увеличивается за счет грибов, и они входят в
рацион беспозвоночных, например, амфипод Gaminurits mucronutus. В
предварительных экспериментах амфипод содержали в течение 2 - 3 месяцев на
диете из 17 различных видов морских грибов. Животные поедали гифы и грибные
конидии, кроме того, конидии были найдены в фекальных шариках. Грибная диета
обеспечивает животных всеми основными питательными веществами, так как амфиподы
в этих экспериментах жили и размножались (Kohlmeyer, Kohlmeyer, 1979).
4.3 Нематоды
В морских бентосных экосистемах содержится большое число
нематод, которые являются, вероятно, самыми обильными животными в сублиторали.
Пищевые потребности для многих видов неизвестны, но Meyers с соавторами
(1963,1964) показали, что нематода Aphelenchoides sp. активно питается грибными
гифами, иссушая цитоплазму. Эти животные росли и размножались в культуре на
диете из мицелия, полученного из разнообразных грибов. Среди изученных грибов
морские аскомицеты и анаморфные грибы Halosphaeria mediosetigera var.
mediosetigera, Lulworthia sp., Scolecobasidium arenarium, Trichocladium
achrasporum и Zalerion maritima были чрезвычайно эффективными как источники
питания, тогда как изоляты грибов аквапочв родов Aspergillus, Cladosporium,
Fusarium и Pestalotia отличались очень низкой утилизацией (Meyers et al, 1964).
Очевидно, у аквапочвенных грибов имеются механизмы, защищающие их от нематод. В
других экспериментах с нематодами не наблюдалось поедание грибов, но животные
привлекались грибными матами, растущими на целлюлозе (Meyers и Hopper 1966,
1967). Отдельно грибной мицелий неэффективен. Авторы объяснили эту
привлекательность как ответную реакцию на вещества, выделяемые комплексом гриба
и целлюлозы или другими ассоциированными микроорганизмами, такими как бактерии,
диатомовые водоросли или простейшие. Нематоды накапливаются и размножаются в
хлопковых фильтрах, покрытых грибными гифами (Meyers, Hopper, 1967), или в
бумажных фильтровальных дисках, погруженных в естественной среде обитания
(Meyers, Hopper, 1966). Популяции нематод в этих экспериментах состояли почти
полностью из оплодотворенных самок, главным образом одного вида Metoncholaimus
scissus. Скопление нематод иногда можно наблюдать в разлагающихся плодовых
телах морских грибов. Нематоды развивались в старых плодовых телах Lulworthia
sp. и, вероятно, внесли вклад в разложение грибного материала (Kohlmeyer,
1958).
4.4 Клещи
Сообщается только о нескольких случаях питания клещей грибным
мицелием, например, на детрите на берегу (Schuster, 1966) и при распаде стеблей
и листьев Spartina (Gessner et al., 1972). Мертвые клещи могут разрушаться
грибами, например, Periconia prolifica растет и образует конидии на этих
животных, прикрепленных к старым туннелям древоточцев (Kohlmeyer, Kohlmeyer,
1977).
Немногочисленные примеры взаимодействия грибов и животных
показывают, что необходимы подробные исследования грибных частиц в
пищеварительных трактах животных. Эти сведения могут иметь не только
экологическое, но и экономическое значение. В настоящее время известно единственное
сообщение об использовании морских грибов, выращенных на сельскохозяйственных
отходах, для корма креветок в марикультуре (Newell, Fell, 1975).
.5 Кораллы
Несколько видов морских аскомицетов ассоциированы с
кораллами. Плодовые тела Corallicola папа Volkm.-Kohlm. et Kohlm. находят на
мертвых кораллах возле берегов Белиза (Kohlmeyer, Volkmann-Kohlmeyer, 1992).
Пять видов рода Koralionastes также встречаются на кораллах: К. angustus Kohlm.
et Volkm.-Kohlm. и К. giganteus Kohlm. et Volkm.-Kohlm. в Белизе; К. ellipticus
Kohlm. et Volkm.-Kohlm. и К. ovalis Kohlm. et Volkm.-Kohlm. в Белизе и
Австралии; К. violaceus Kohlm. et Volkm.-Kohlm. в Австралии и на Фиджи
(Kohlmeyer, Volkmann-Kohlmeyer, 1987, 1990, 1992). Lulworthia calcicola Kohlm.
et Volkm.-Kohlm. - еще один вид, обитающий на кораллах в Белизе (Kohlmeyer,
Volkmann-Kohlmeyer, 1989a).
4.6 Голотурии
Определенный интерес с точки зрения экологии и физиологии
грибов представляют грибы голотурий (Holothurioidea, Echinodermata) (Pivkin,
2000). Эти животные, как известно, содержат тритерпеновые гликозиды, которые
обладают фунгицидной активностью. Тем не менее, грибы были изолированы из всех
органов голотурий. Двадцать семь видов анаморфных вторичных морских грибов были
обнаружены на голотуриях трех видов (Apostichopus japonicus, Eupentacta
fraudatrix и Cucumaria japonica). Штаммы грибов, выделенные с поверхности
животных, выдерживали в 5 раз большие концентрации гликозидов, чем грибы,
выделенные из других морских мест обитания. Грибы, выделенные из внутренних
органов голотурий, росли при концентрации суммарной фракции гликозидов Е.
fraudatrix 40 мг/мл, что значительно превышает их концентрацию в теле
животного. По мнению автора (Pivkin, 2000), фунгитоксичность не только не
снижает разнообразия грибов голотурий, но и стимулирует биоразнообразие
микобиоты животных.
5. Ферменты морских грибов
Адаптацию грибов в море обеспечивают в первую очередь
ферментные системы, свойства которых могут отличаться от свойств ферментных
систем наземных видов (Пивкин и др., 2006). Морские грибы, обладающие
эффективным и разносторонним ферментным аппаратом, осуществляют деградацию
древесины, растительных и животных остатков (Hyde et al., 1998).
В работе Молиторса и Шауманна (Molitoris, Schaumann, 1986)
были приведены обобщенные сведения о ферментах грибов, участвующих в
метаболизме, и их промышленном использовании (табл. 1). К настоящему времени
данные о грибах, в том числе из морских мест обитания, как биотехнологически
важных источниках различных ферментов, значительно расширены.
Таблица 1. Экологическое значение и промышленное
использование ферментов грибов (по: Molitoris, Schaumann, 1986)
|
Фермент
|
Расщепление
природных субстратов
|
Промышленное
использование (или потенциальная возможность)
|
|
Окислительный
метаболизм Лакказа Тирозиназа Пероксидаза
|
Древесина
(лигнин, целлюлоза) Древесина (лигнин)
|
Контроль
загрязнения, разрушение промышленных отходов
|
|
Азотный
метаболизм Протезы Желатиназа Казеиназа Нитратредуктаза Липидный метаболизм
Липаза
|
Белок
(животные, растения)
|
Детергенты,
добавки к пиву, лекарства, косметика, контроль загрязнения, утилизация
промышленных отходов
|
|
NO3
в воде как источник азота
|
-
|
|
Липиды
(животные, растения)
|
Пищевая
промышленность, добавки при экстракции липидов
|
|
Углеводный
метаболизм Амилаза
|
Крахмал
(растения)
|
Сахар,
пекарная, текстильная, бумажная промышленность
|
|
Целлюлаза
|
Клеточная
стенка (целлюлоза, растенияния)
|
Бумажная
промышленность, маркер загрязнения, утилизация промышленных отходов
|
|
Пектиназа
Полигалактуроназа Трансэлиминаза
|
Клеточная
стенка (пектин, растения)
|
Пищевая
промышленность, пектин напитков
|
|
Хитиназа
|
Полисахариды
(хитин, животные)
|
Биологический
контроль патогенных грибов
|
|
Ламинариназа
|
Полисахариды
(ламинарии, биомасса морских грибов, водоросли)
|
|
6. Вторичные метаболиты изолятов морских грибов
Грибы, выделенные из морской среды, являются перспективными
источниками уникальных химических соединений с разнообразной биологической
активностью (Михайлов и др., 1999; Blant et al., 2004; Kobayashi et al., 1993;
Faulkner, 2001). К настоящему времени описаны более чем 500 видов облигатных
морских грибов, тем не менее, существует точка зрения, что многие из них
являются космополитами и охотно адаптируются к жизни, как на суше, так и на
море. Эти космополиты могут быть выделены с поверхности и внутренней части
водорослей, а также из внутренних органов морских беспозвоночных-фильтраторов,
таких как губки и асцидии. Хотя экологическая роль грибов в море не выяснена,
космополитные роды, такие как Aspergillus, идентифицированы как агенты,
вызывающие массовую гибель морских беспозвоночных, т. е. грибы, могут оказывать
значительное экологическое воздействие. Известно, например, что гриб Fusarium
solani может инфицировать различных морских ракообразных, а оомицет Lagenidium
callinectes (в настоящее время состоящий в царстве страминипил) оказывает
патогенное воздействие на яйца крабов и лобстеров. В то же время показано, что
существуют мутуалистические взаимоотношения между организмом хозяина и грибами.
Так, присутствие аскомицета Turgidosculum ulvae в талломе зеленой водоросли
Blidingia minima защищает ее от поедания беспозвоночными.
Долгое время считали, что наземные грибы имеют высокую
степень толерантности к соли, и поэтому не удивительно, что они могут жить как
на суше, так и в море. Однако такая адаптация требует от грибов выработки
соединений, которые способствуют сохранению целостности клеток грибов и
защищают грибы и организм хозяина от различных внешних патогенных
микроорганизмов. Поэтому неудивительно, что изучение биосинтетических
возможностей морских изолятов грибов приводит к открытию ранее не описанных для
наземных грибов метаболитов. Одни и те же виды грибов, выделенные из наземных и
морских источников, синтезируют различные биологически активные соединения.
Так, грибы рода Trichoderma, известные почвенные обитатели, долгое время были
предметом интенсивного изучения. Это привело к выделению различных биологически
активных соединений, таких как глиотоксин (Erchner et al., 1986), глиоверин
(Stipanovic, Howell, 1982), стероидный антибиотик виридин, сесквитерпены,
гептилидиновые кислоты. Однако изучение морских изолятов этого рода грибов
способствовало открытию других необычных биологически активных веществ. Морские
изоляты грибов могут быть перспективными источниками новых антибиотиков. При
изучении антимикробной активности 1500 видов морских изолятов грибов и 1450
изолятов наземных грибов было показано, что морские изоляты грибов продуцируют
антимикробные соединения чаще, чем наземные (Cuomo et al., 1995). На
антибиотическую активность были тестированы 227 изолятов грибов, принадлежащих
к родам Eupenicillium, Penicillium, Aspergillus, Eurotium, Fusarium,
Emericella, Alternaria, Gliocladium (Christophersen et al., 1999). Из них 61
изолят (27 %) показал антибактериальную активность, 55 штаммов - активность по отношению
к Staphylococcus aureus, 7 изолятов были активны по отношению к Vibrio
parahaemolyticus. Ни одна из выделенных культур не ингибировала роста
Pseudomonas aeruginosa n Escherichia coli. Процентное отношение культур,
продуцирующих антибактериальные соединения, к общему числу протестированных
штаммов, совпадало с данными, опубликованными Cuomo с соавторами (Cuomo et al.,
1995).
7. Краткая характеристика обследованных двустворчатых
моллюсков
.1 Приморский гребешок (Mizuhopecten yessoensis)
Приморский гребешок - тихоокеанский, приазиатский
низкобореальный вид (Скарлато, 1981). Его ареал занимает обширную акваторию от
северных берегов п-ова Корея до южных Курильских островов, также он встречается
вдоль берегов Японии - у о-ва Хоккайдо и у северного побережья о-ва Хонсю
(Kira, 1962). Ареал распространения приморского гребешка изображен на Рис. 2
(Мотавкин, 1986).
На Южно-Курильском мелководье гребешок распространен широко,
но крупные скопления обнаружены только у юго-восточного прибрежья о-ва Кунашир,
между островами Танфильева, Юрий и Зеленый и у этих же островов со стороны
Южно-Курильского пролива (Скалкин, 1966).
У берегов о-ва Сахалин гребешок распространен вдоль западных
берегов, в основном он встречается в зал. Анива: южнее пос. Атласово, на траверзе
от пос. Крестьяновка до пос. Таранай, у пос. Пригородное в бух. Лососей и от г.
Корсаков до лагуны Буссе. Гребешок встречается у восточных берегов к югу от
зал. Терпения, а также у о-ва Монерон. Плотность скоплений и площади поселений
от одного участка к другому существенно различаются (Скалкин, 1966).
Интенсивный промысел в 60-е годы прошлого века существенно подорвал запасы
приморского гребешка и в прибрежье о-ва Сахалин (Жюбикас, 1969).
В прибрежье Приморья наибольшее количество гребешка отмечено
в заливах Посьета и Петра Великого, где он распространен неравномерно.
Наибольшая плотность поселения гребешка установлена в бухтах и заливах, а
значительно меньшая - у открытых берегов и островов (Бирюлина, 1972а).
Диапазон температур воды, при которых встречен приморский
гребешок, составляет от -2 до 26° С. Чаще и в большем количестве гребешок
встречается в тех районах, где зимой вода не ниже -1,5, а летом не выше 18 -
20° С.
Приморский гребешок, в отличие от некоторых других
двустворчатых моллюсков, даже в течение нескольких дней не переносит
значительного опреснения, поэтому не поселяется вблизи устьев рек. Зимой
практически во всех местах обитания гребешка соленость близка к 34‰. Летом
значение этого показателя меняется в зависимости от выпадения осадков и других
факторов.
Известно, что приморский гребешок, особенно его молодь,
чувствителен к дефициту кислорода. Он предпочитает хорошо аэрируемые районы с
постоянными довольно сильными течениями, периодическими приливами и отливами.
Для приморского гребешка (Брегман., 1973) установлена нижняя граница
оптимального содержания растворенного кислорода в воде - 6 мл/л.
Чаще всего приморский гребешок встречается на илисто-песчаном
и песчаном грунте, даже если такие участки располагаются островками среди
камней; на илистом грунте с примесью гальки, гравия или битой ракушки,
благоприятных для образования водорослевых зарослей - субстрата для молоди.
Кроме этого, он живет и на чисто галечном, гравийном и песчаном грунтах (Разин,
1934). На чистом иле и камнях гребешок редок, исключение составляет лишь
многочисленная популяция из оз. Второе залива Находка Японского моря, обитающая
на иле. По данным Скалкина (Скалкин, 1966), граница распространения гребешка по
отдельным районам совпадает с границами распределения предпочитаемого им
грунта.
Приморский гребешок отмечен в Японском море на глубине 1-80
м, в частности в зал. Петра Великого - на 4 - 80 м, в зал. Посьета - на 1 - 20
м (Скарлато, 1981), но наиболее массовые скопления образует на глубине 6 - 30
м.
Продолжительность жизни гребешка приморского около 15 лет.
Средняя продолжительность жизни гребешка - 10 лет. Однако встречаются
экземпляры в возрасте 15 - 17 лет и более.
Наибольшие темпы роста наблюдаются в первые 3-4 года при
температуре воды 10 - 16°С (май - июнь, октябрь - ноябрь). Для роста наиболее
благоприятны полуоткрытые бухты и приостровные зоны с интенсивным водообменом.
Зимой рост моллюска приостанавливается (Временная инструкция по технологии
выращивания приморского гребешка после годичного подращивания в садках, 1987).
Половозрелым моллюск становится на третьем году жизни, достигая в длину 9 см
Длина раковины изредка достигает 22 см, а живая масса - 1 кг.
Средний размер взрослого гребешка 12 - 17 см, масса 200 - 400 г. Вес мускула
крупных особей около 40 г. Промысловый размер составляет - 120 мм по длине
раковины (Правила рыболовства, 2007).
Раковина гребешка составляет в среднем 50% от общей массы
тела; мускул является наиболее ценной съедобной частью, доля которого
составляет 10 - 16% в зависимости от сезона и района обитания. Абсолютная масса
мускула взрослого гребешка колеблется от 20 до 200 г.
.2 Мидия Грея (Crenomytilus grayanus (Dunker, 1853))
Тихоокеанский, приазиатский, низкобореальный, сублиторальный
вид. Образует промысловые скопления. Ведется промысловый лов мидии. Максимально
установленные: возраст- 150 лет, размер 226мм, масса особи- 1,6 кг.
Половозрелость у моллюсков наступает в 6 лет при длине60-70
мм, на искусственных субстратах - в 3 года. Нерест в июне-августе. Обитает на
различных грунтах, прикрепляясь к субстрату биссусными нитями на глубинах 1 -
60 м. Основные поселения приурочены к глубинам до 20 м. Живет мидия как
одиночно, так и в друзах. Моллюски образуют скопления с плотностью поселения до
500 экз./м2 и биомассой 40 кг/м2. Скопления мидии
представлены в виде банок или полей.
В зал. Петра Великого мидия обитает как у открытых берегов,
так и в бухтах на скалисто-каменном грунте, на заиленной гальке в виде друз.
У западного и южного Сахалина мидия друз не образует. В
лагуне Буссе мидия обитает на глубинах 5-10 м.
Отмечена на южнокурильском мелководье у о-вов Полонского,
Шикотан, Итуруп (зал. Касатка), в Татарском проливе. Соотношение к общей массе
тела: мягкие ткани- 30%, раковина- 55%. В период нереста масса мягких тканей
возрастает. Раковина большая, толстостенная, выпуклая. Периостракум
черно-коричневый. Створки имеют грубые линии нарастания. Массивные макушки
оттянуты книзу. Отпечаток переднего мускула-замыкателя вдавлен. Отпечаток
заднего мускула-замыкателя не вдавлен. Лигамент закрытый, мощный, равный
половине длины раковины. (Явнов, 2000).
.3 Модиолус длиннощетинистый (Modiolus modiolus (Kuroda et
Habe, 1950))
Тихоокеанский, приазиатский, субтропическо-бореальный,
сублиторальный вид, встречающийся в литорали. Максимально установленные:
возраст- 43 года, длина раковины - 141 мм, вес- 247 г. Соотношение к общей
массе тела: мягкие ткани - 29%, раковина- 51%. Половозрелость с 4-го года
жизни. В зал. Посьета нерест в июне-августе, в зал. Восток - в
сентябре-октябре.
Моллюск образует скопления на заиленных песке и гальке;
единичные особи отмечены на каменистом и скалистом грунтах, в зарослях
водорослей и морской травы на глубинах 2-20 м. Модиолус образует друзы, в
состав которых входит 2-3 десятка взрослых моллюсков и много молоди. Обычен в
сообществе мидии Грея. Ведет прикрепленный образ жизни. Живет как в бухтах, так
и у открытых берегов.
Обитает в зал. Петра Великого, лагуне Буссе и зал. Анива у
юго-западного Сахалина, у Курильских (о. Парамушир) и Командорских островов.
Створки покрыты черно-коричневым периостракумом, выросты
которого длинные и густые; изнутри створки обычно интенсивно окрашены в
фиолетовый цвет. Биссус хорошо развит.
Общий запас в зал. Петра Великого может составлять порядка
2000 т. (Явнов, 2000).
.4 Анадара Броутона (Anadara broughtoni (Schrenck, 1867))
Тихоокеанский, приазиатский, субтропический,
верхнесублиторальный вид. Максимально установленные: возраст- 65 лет, длина
раконины- 128 мм, высота- 96 мм, толщина- 78 мм, общая масса особи- 380 г.
Раковина сильно выпуклая, створки покрыты 42-43-мя
радипш.ными ребрами. Периостракум темно-коричневый. Замок таксодонтный с
многочисленными зубами, расположенными в одну прямую линию. Мягкие части тела
моллюска окрашены в оранжево-кирпичный цвет. Кровь красного цвета из-за присутствия
гемоглобиноподобных веществ. Соотношение к общей массе тела: мягкие ткани -
30%, пищевая часть - 20%, створки - 47%, свободная жидкость - 23%.
Половозрелыми моллюски становятся в 3-5 лет. Нерест особей с конца июня до
начала августа.
Анадара обитает в илисто-песчаном грунте, зарываясь в него на
10-25 см. Моллюски населяют грунты в закрытых и полузакрытых бухтах, заливах на
глубинах от 1,5 до 20 м.
Анадара отмечена в Амурском, Уссурийском заливах, зал.
Восток, бухтах Нарва, Рейд Паллады, Экспедиции, Новгородская.
Значительные промысловые скопления моллюска расположены в
северной части Амурского, Уссурийского заливов и в бухте Рейд Паллады зал.
Петра Великого на глубинах 4-12 м с плотностью до 1,5 экземпляра на 1 м2.
Общий запас в зал. Петра Великого может составлять порядка 9000 т.
8. Физико-географическое описание залива Восток
Залив Восток находится в юго-восточной части зал. Петра
Великого и обращен к открытому морю. Акваторию залива ограничивают на западе
мыс Пещурова, на востоке - мыс Подосёнова. Западный берег, за исключением бухт
Тафуин, Гайдамак и Средней, как правило, скалистый, часто окаймлен камнями и
кекурами. Берег, вершины залива низкий и песчаный, лишь в средней его части
выступает в залив остроконечный обрывистый мыс. Восточный берег залива по мере
продвижения с севера на юг постепенно повышается и становится утесистым, с
красноватыми осыпями.
На линии входных мысов залива Восток глубины 15-31 м, грунт -
ил, песок. В средней части залива грунт - ил. К вершине залива глубины
постепенно уменьшаются, а грунт становится песчаным.
Первое появление льда в заливе наблюдается в конце ноября. В
начале декабря покрываются сплошным льдом сначала гавань Гайдамак и бухта
Восток, а затем западная часть бухты Гайдамак. Полное очищение залива Восток от
льда происходит в конце марта (Лоция, 1981).
Распределение океанологических параметров в придонном слое
обусловлено в основном глубиной места: температура с глубиной понижается, а
солёность растёт. По этим параметрам различия между разными бухтами залива незначительны:
диапазон солёности у дна составляет от 33,45‰ на глубине 4 м до 33,75‰ на
глубине 17 м, а температуры от 20ºС на глубине 4 м до 17,3ºС на глубине 17 м. Судя по распределению солёности на поверхности
моря, вдоль западного берега залива происходит сток распреснённых эстуарных
вод, но на глубинах свыше 10 м это явление не прослеживается.
Распределение кислорода с глубиной имеет более сложный
характер. На глубинах 10-20 м содержание кислорода примерно постоянно: в
среднем 4 мл/л, прослеживается тенденция к росту кислородосодержания с
уменьшением глубины, но на мелководной станции в бухте Восток с глубиной 5 м
отмечено минимальное содержание кислорода: около 3,4 мл/л. Подобный дефицит
кислорода вполне обычен для придонного слоя эстуарных зон, и обусловлен
активным его потреблением на окисление органического вещества. То есть,
наблюдаемый недостаток кислорода является признаком чрезвычайно высокой
продуктивности вод северо-западного района зал. Восток, куда впадает р.
Волчанка. Дополнительным фактором, способствующим развитию дефицита кислорода,
является хорошая защищённость бухты от волнения. В менее защищенной
северо-восточной части залива, куда впадает р. Литовка (бухта Литовка),
дефицита кислорода не отмечено, правда, самые малые глубины в этом районе не
обследовались (по устному сообщению Ю.И. Зуенко).
Рекреационная нагрузка на отдельных участках побережья залива
Восток выше допустимых норм. Действие антропогенного фактора отражается на
ухудшении качества водной среды в летний сезон по кислородным показателям,
количеству фосфатов и детергентов, а также санитарно-микробиологическим
характеристикам. Наиболее значительные изменения в структуре и составе отмечены
для сообществ макробентоса мягких грунтов. Наиболее сильные изменения произошли
в бухте Восток и бухте Гайдамак (Галышева Ю. А., Христофорова Н. К., 2007).
Несмотря на наличие небольшого числа локальных участков загрязнения, заметно
возросший за последнее десятилетие антропогенный пресс на залив изменил
экологическую ситуацию, существенно отклонив ее от фоновых условий. В то же
время медленно накапливающиеся изменения в целом еще не достигли тех границ, с
которых начинают сказываться негативные биологические эффекты (Христофорова и
др., 2004).
9. Материалы и методика
Материалом для микобиотических исследований служили
двустворчатые моллюски: приморский гребешок Mizuhopecten yessoensis, модиолус
Modiolus modiolus, мидия Грея Crenomytilus grayanus, анадара броутони Anadara
broughtoni.
Двустворчатые моллюски были собраны в заливе Восток (залив Петра
Великого, Японское море) в августе 2006 - 2007 гг.
Выбор вида моллюска обусловлен его промысловым значением и
массовостью вида.
Выборка двустворчатых моллюсков составляла по 3 особи каждого
вида.
Для изучения у моллюсков мы отпрепарировали внутренние
органы. Отпрепарированные внутренние органы: жабры, мантия, почки,
пищеварительная железа, мускул, женские гонады в течение двух часов вымачивали
в растворе антибиотиков (с добавлением 500000 ед. пенициллина и 0,5г.
стрептомицина на 1 л стерильной морской воды (Артемчук, 1981)) для подавления
роста бактерий, затем промывали в стерильной морской воде. Использовали глюкозо
- дрожжевую, сусло - агаровую, среду Чапека и среду Тубаки, приготовленные на
профильтрованной морской воде. Инокулированные среды инкубировали при
температуре 20oС. Выросшие на жидких питательных средах колонии
пересевали на агаризованные среды аналогичного состава и идентифицировали.
Пересев производили с помощью микологического крючка. (Методы…, 1982)
Идентификацию грибов проводили с помощью общепринятых ключей
и определителей (Ames, 1961; Ellis, 1971; Gams, 1971; Kohlmeyer, Kohlmeyer,
1987; Pitt, 1979; Ramirez, 1982; Rifai, 1969; Билай, Коваль, 1988; Егорова,
1986; Черепанова, 1989; и др.). Сокращения авторов таксонов приведены согласно Кирку
и Анселлу (Kirk, Ansell, 1992).
Число пропагул грибов рассчитывали на 1 г тела моллюска.
Экологическое биоразнообразие грибов каждого вида моллюсков было оценено с
помощью индекса Шеннона, доминирование - с помощью индекса Бергера-Паркера
(Мэгарран, 1992), расхождение грибных комплексов разных видов моллюсков между
собой - при помощи программы: STATISTICA 6.
10. Результаты и обсуждение
.1 Таксономический состав мицелиальных грибов,
ассоциированных с двустворчатыми моллюсками
В ходе проведённого исследования нами выделено около 200
штаммов мицелиальных грибов, идентифицировано 42 вида, относящихся к 14 родам,
из них 7 видов сумчатых грибов, 33 вида анаморфных грибов и 2 вида зигомицетов.
Во внутренних органах приморского гребешка обнаружено 18
видов мицелиальных грибов, из них 16 видов анаморфных грибов и 2 вида
зигомитетов; из мидии Грея выделено 22 вида грибов, из них 19 видов анаморфных
и 3 вида сумчатых грибов; у модиолуса длиннощетинистого - 23 вида, из них 18
видов анаморфных и 5 видов сумчатых грибов; из анадары Броутона - 3 вида
анаморфных грибов (таб. 2).
Таблица 2. Количество видов мицелиальных грибов различных
групп в обследованных двустворчатых моллюсках
|
Таксоны грибов
|
Mizuhopecten
yessoensis
|
Crenomytilus
grayanus
|
Modiolus
modiolus
|
Anadara
broughtoni
|
|
Zygomycota
|
2
|
0
|
0
|
0
|
|
Ascomycota
|
0
|
3
|
5
|
0
|
|
Anamorphic
fungi
|
16
|
19
|
18
|
3
|
|
Итого
|
18
|
22
|
23
|
3
|
Количество видов мицелиальных грибов различных
таксономических групп ассоциированных с обследованными двустворчатыми
моллюсками показано на рисунке 6.
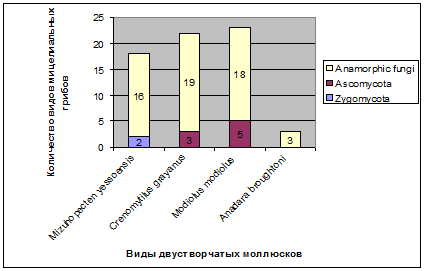
Рис. 6. Количество видов мицелиальных грибов различных групп
в двустворчатых моллюсках
Большинство видов мицелиальных грибов, выделенных из
двустворчатых моллюсков, являются анаморфными грибами. Зигомицеты были выделены
только из органов приморского гребешка. Аскомицеты были найдены в мидии Грея и
модиолусе. Таксономический состав мицелиальных грибов ассоциированных с
анадарой Броутона составляют анаморфные грибы (рис. 6). Можно предположить, что
видовой состав грибных комплексов зависит от видовой принадлежности
двустворчатых моллюсков. Возможно, такое распределение связано с
физиологическими особенностями и образом жизни моллюска, а именно,
приуроченностью к определённым грунтам и экологическими условиями обитания, так,
анадара - единственный из обследованных нами моллюсков ведёт зарывающийся образ
жизни, в связи с чем обладает гемиподобными веществами, окрашивающими её кровь
в красный цвет.
Как говорилось выше, таксономический состав мицелиальных
грибов, ассоциированных с двустворчатыми моллюсками Mizuhopecten yessoensis,
Crenomytilus grayanus, Modiolus modiolus и Anadara broughtoni составляет 42
вида (таб. 3).
Таблица 3. Таксономический состав грибов ассоциированных с
двустворчатыми моллюсками Mizuhopecten yessoensi, Modiolus modiolus
Crenomytilus grayanus и Anadara broughtoni
|
Виды грибов
|
Mizuhopecten
yessoensis
|
Crenomytilus
grayanus
|
Modiolus
modiolus
|
Anadara
broughtoni
|
|
Acremonium sp
|
|
|
+
|
|
|
Alternaria
alternata(Fr.)Keissl
|
+
|
+
|
|
|
|
Alternaria
tenuissima(Fr.)Wiltshire
|
+
|
|
|
|
|
Aspergillus
(glaucus series)
|
+
|
|
|
|
|
Aspergillus
candidus Link
|
|
+
|
|
|
|
A. nidulans
(Eidam) G. Winter
|
|
|
+
|
|
|
A. janus Raper
et Thom
|
|
+
|
|
|
|
A. niger Tiegh.
|
|
+
|
+
|
|
|
A. ochraceus K.
Wilh.
|
|
+
|
+
|
|
|
A. parasiticus
Speare
|
|
+
|
|
|
|
A. sulphureus
Thom et Church
|
|
|
+
|
|
|
A. versicolor
(Vuill.) Tirab.
|
|
+
|
+
|
|
|
Aspergillus sp1
|
+
|
|
|
|
|
Aspergillus sp.
2
|
|
|
|
+
|
|
Aureobasidium
pullulans(De Bary.)Arnaud
|
+
|
|
+
|
|
|
Cladosporium
cladosporioides(Fresen.) de Vries
|
+
|
|
+
|
|
|
Cladosporium
oxysporum Berk et Curt
|
+
|
+
|
|
|
|
Cladosporium
sphaerospermum Penz.
|
+
|
+
|
+
|
|
|
Cladosporium
sp.
|
|
|
|
+
|
|
Eurotium
herbariorum Link
|
|
+
|
|
|
|
Geomyces
pannorum(Link) Singler et Carmichl
|
+
|
+
|
+
|
+
|
|
Mucor
circinelloides
|
+
|
|
|
|
|
Myxotrichum
chartarum Kunze
|
|
|
+
|
|
|
Penicillium
aurantiogriseum Dierck
|
|
|
+
|
|
|
Penicillium
citrinum Thom
|
+
|
+
|
+
|
|
|
P. citreonigrum
Dierckx
|
|
+
|
|
|
|
P. commune Thom
|
|
+
|
+
|
|
|
P. decumbens
Thom
|
|
+
|
|
|
|
P. griseofulvum
Dierckx
|
|
|
+
|
|
|
P. jensenii W.
Zalessky
|
|
+
|
+
|
|
|
Penicillium
verrucosum Dierckx var cyclopium (Wesfling) Samson, Stolk et Hadlok
|
+
|
+
|
+
|
|
|
Penicillium
sp.+
|
+
|
|
+
|
|
|
.Penicillium
sp.2
|
+
|
+
|
|
|
|
Penicillium
sp.3
|
+
|
+
|
|
|
|
Ch. globosum
Kunze
|
|
|
+
|
|
|
Ch.
megalocarpum Bainier
|
+
|
|
|
|
Ch.
spiculipilium L.M. Ames
|
|
|
+
|
|
|
Ch. spinosum
Chivers
|
|
|
+
|
|
|
Chaetomium sp.
|
|
+
|
+
|
|
|
Rhizopus
nigricans Ehrenb.
|
+
|
|
|
|
|
Trichoderma
aureoviride Rifai
|
+
|
|
|
|
|
Wardomyces
inflatus (Marchae) Hennebert
|
+
|
+
|
+
|
|
|
Итого 42
|
18
|
22
|
23
|
3
|
В тканях Mizuhopecten yessoensis было обнаружено 18 видов
мицелиальных грибов, принадлежащих к 9 родам (Alternaria, Aspergillus,
Aureobasidium, Cladosporium, Geomyces, Mucor, Penicillium, Rhizopus,
Trichoderma, Wardomyces). Наиболее широко представлен род Penicillium(5 видов).
Из внутренних органов Crenomytilus grayanus было выделено и
идентифицировано 22 вида грибов, относящихся к 8 родам (Alternaria,
Aspergillus, Cladosporium, Eurotium, Geomyces, Penicillium, Chaetomium и
Wardomyces).Наиболее широко представлены рода Aspergillus (6 видов) и
Penicillium (8 видов).
В органах Modiolus modiolus обнаружено 23 вида мицелиальных
грибов, относящихся к 9 родам (Acremonium, Aspergillus, Aureobasidium
Cladosporium, Geomyces, Myxotrichum, Penicillium, Chaetomium и Wardomyces).
Наиболее широко представлен род Penicillium(7 видов).
В тканях Anadara broughtoni было обнаружено всего 3 вида
грибов, принадлежащих 3 родам (Aspergillus, Cladosporium и Geomyces).
Виды мицелиальных грибов Alternaria tenuissima, Aspergillus
(glaucus series), Aspergillus sp1, Mucor circinelloides, Rhizopus nigricans и
Trichoderma aureoviride были найдены только в приморском гребешке; Aspergillus
candidus, Aspergillus janus, Aspergillus parasiticus, Eurotium herbariorum,
Penicillium citreonigrum, Penicillium decumbens и Chaetomium megalocarpum
обнаружены лишь у мидии Грея; Acremonium sp, Aspergillus nidulans, Aspergillus
sulphureus, Myxotrichum chartarum, Penicillium aurantiogriseum, Penicillium
griseofulvum, Chaetomium globosum, Chaetomium spiculipilium и Chaetomium
spinosum найдены только в модиолусе; Aspergillus sp. 2 и Cladosporium sp.1 были
найдены только у анадары Броутона.
Лишь один вид (Geomyces pannorum) обнаружен во всех
обследованных моллюсках. Наиболее устойчивой к заселению грибами оказалась
анадара Броутона - в ней обнаружено только 3 вида грибов.
.2 Распределение мицелиальных грибов во внутренних органах
двустворчатых моллюсков
Распределение мицелиальных грибов во внутренних органах мы
изучили у Mizuhopecten yessoensis, Modiolus modiolus и Crenomytilus grayanus
(рис. 7).
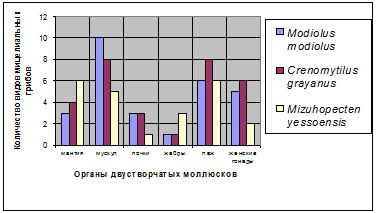
Рис. 7. Количество видов мицелиальных грибов во внутренних
органах моллюсков Mizuhopecten yessoensis Modiolus modiolus и Crenomytilus grayanus
Оно следующее: у Mizuhopecten yessoensis в мантии обнаружено
6 видов (Alternaria alternata, Alternaria tenuissima, Aspergillus sp. 1,
Cladosporium brevi-compactum, Cladosporium oxysporum и Penicillium sp. 2), в
мускуле 5 видов мицелиальных грибов (Cladosporium cladosporioides, Cladosporium
oxysporum, Penicillium sp.1, Penicillium sp. 3 и Rhizopus nigricans), в почках
1 вид (Penicillium sp.1), в жабрах 3 вида (Rhizopus nigricans, Penicillium
citrinum, Cladosporium sphaerospermum), в пищеварительной железе 6 видов
(Alternaria sp, Alternaria tenuissima, Aspergillus (glaucus feries),
Cladosporium oxysporum, Trichoderma aureoviride, Penicillium sp.) и в женских
гонадах 2 вида (Rhizopus nigricans и Penicillium sp.1)
У Modiolus modiolus в мантии было обнаружено 3 вида
(Aspergillus sp. 1, Penicillium aurantiogriseum, Penicillium sp. 1), в мускуле
10 видов (Aspergillus nidulans, A. sulphureus, A. versicolor, Aspergillus sp.,
Cladosporium. cladosporioides, Penicillium commune, P. griseofulvum, P.
jensenii, Penicillium sp.1, Chaetomium spiculipilium), в почках 3 вида
(Acremonium sp., Aspergillus niger, Penicillium sp.1), в жабрах 1 вид
(Penicillium citrinum), в пищеварительной железе 6 видов (Aspergillus
ochraceus, A. sulphureus, A. versicolor, Aureobasidium pullulans, Cladosporium
cladosporioides, Penicillium sp.1), в женских гонадах 5 видов (Cladosporium
sphaerospermum, Chaetomium globosum, Ch. spinosums, Chaetomium sp., Myxotrichum
chartarum).
У Crenomytilus grayanus в мантии было обнаружено 4 вида
(Aspergillus niger, A. versicolor, Cladosporium oxysporum, Penicillium sp. 3),
в мускуле 8 видов (Aspergillus janus, A. ochraceus, A. versicolor, Cladosporium
sphaerospermum, Eurotium herbariorum, Penicillium citrinum, P. citreonigrum, P.
jensenii), в почках 3 вида (Aspergillus candidus, A. ochraceus, Penicillium
citrinum), в жабрах 1 вид (Chaetomium megalocarpum), в пищеварительной железе 8
видов (Aspergillus parasiticus, A. versicolor, Penicillium citrinum, P.
citreonigrum, P. commune, P. decumbens, P. jensenii, Chaetomium sp.), в женских
гонадах 6 видов (Alternaria alternata, Aspergillus ochraceus, A. versicolor,
Cladosporium oxysporum, Penicillium citrinum, Penicillium sp. 2).
Наиболее заселёнными грибами органами являются: у
Mizuhopecten yessoensis - мантия и пищеварительная железа (по 6 видов), у
Modiolus modiolus - мускул (10 видов), у Crenomytilus grayanus - мускул и
пищеварительная железа (по 8 видов). Наименьшее количество видов грибов: у
приморского гребешка обнаружено в почках (1 вид), у мидии Грея и модиолуса
наименее заселёнными оказались жабры (1 вид) (рис. 7).
Мы не можем пока сказать, что вызвало такое распределение,
так как нет никаких данных о взаимоотношениях мицелиальных грибов с
двустворчатыми моллюсками.
Возможно, что характер распределения мицелиальных грибов во
внутренних органах двустворчатых моллюсков свидетельствует о степени иммунной
защиты этих гидробионтов (Зверева, Высоцкая, 2005). Согласно этому, моллюск
вида Modiolus modiolus, является наименее защищенным к заселению морскими
грибами. Одним из наименее устойчивых органов у изученных нами видов моллюсков,
является мускул. Самыми неблагоприятными органами для развития морских грибов,
являются почки и жабры моллюсков.
Мускул-замыкатель у Mizuhopecten yessoensis имеет меньшее,
среди изученных видов моллюсков, количество мицелиальных грибов (5 видов). Хотя
нет гарантий, что среди них нет видов, продуцирующих токсины, малое количество
видов грибов уменьшает вероятность токсикоинфекций.
Мы не можем точно сказать, из-за чего происходит такое
неравномерное распределение по внутренним органам, так как данная группа
мицелиальных грибов ещё недостаточно изучена.
мицелиальный гриб двустворчатый моллюск
10.3 Показатели биоразнообразия мицелиальных грибов,
ассоциированных с обследованными двустворчатыми моллюсками
Показатели биоразнообразия мицелиальных грибов,
ассоциированных с обследованными видами двустворчатых моллюсков, приведены в
Таблице 4.
Таблица 4. Показатели биоразнообразия и выравненности
комплексов грибов, ассоциированных с двустворчатыми моллюсками
|
Виды моллюсков
|
Число видов
мицелиальных грибов
|
Средне число
пропагул всех видов грибов на 1 г мягкого тела моллюска
|
Число пропагул
самого обильного вида (на 1г)
|
Индекс
разнообразия Шеннона
|
Средняя
выравненность
|
Индекс
Бергера-Паркера
|
|
Mizuhopecten
yessoensis
|
18
|
0,244±0.01
|
0,085±0
|
0,925±0.05
|
0,32±0.02
|
0,348±0.02
|
|
Crenomytilus
grayanus
|
22
|
0,381±0.02
|
0,132±0.01
|
1,348±0.07
|
0,436±0.02
|
0,347±0.02
|
|
Modiolus
modiolus
|
23
|
1,52±0.08
|
0,6±0.03
|
3,063±0.15
|
0,191±0.01
|
0,395±0.02
|
|
Anadara
broughtoni
|
3
|
0,185±0.01
|
0.123±0.01
|
0,472±0.02
|
0,43±0.02
|
0.667±0.03
|
*Данные достоверны при уровне значимости 95%, P<0,05
Среди изученных видов моллюсков самый высокий показатель
биоразнообразия наблюдается у комплекса грибов, ассоциированного с моллюском
Modiolus modiolus. Однако, этому же виду соответствует наименьшая выравненность
грибных сообществ (0,191) и наибольшая степень доминирования (0,395). Это
говорит о низкой устойчивости грибных комплексов, ассоциированных с
двустворчатым моллюском вида Modiolus modiolus. Наименьшее значение
биологического разнообразия соответствует ассоциантам моллюска Mizuhopecten
yessoensis. Показатели выравненности и доминирования невысоки, что также
свидетельствует о низкой устойчивости сообщества мицелиальных грибов.
Небольшое количество грибов, обнаруженное в моллюске вида
Anadara broughtoni (3 вида) не позволяет сделать никаких выводов о
биоразнообразии грибных комплексов. Можно только отметить низкую численность
грибов, ассоциированных с данным моллюском, о чем говорилось выше.
Возможно, различие показателей биологического разнообразия
грибных комплексов обусловлено зависимостью видового состава мицелиальных
грибов от вида двустворчатого моллюска.
Расхождения грибных комплексов, ассоциированных с
двустворчатыми моллюсками, мы высчитывали в программе STATISTICA 6, по
коэффициенту расхождения (Эвклидово расстояние) (рис. 8).
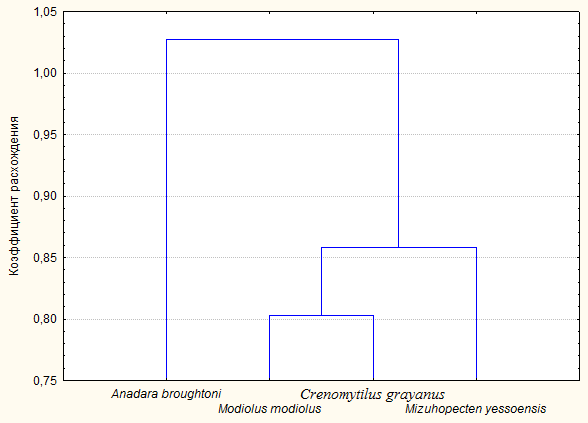
Рис. 8. Дендрограмма расхождения грибных комплексов,
ассоциированных с двустворчатыми моллюсками Mizuhopecten yessoensi, Modiolus
modiolus Crenomytilus grayanus и Anadara broughtoni
Как видно из дендрограммы, наиболее близки по видовому
составу мицелиальных грибов моллюски Modiolus modiolus и Crenomytilus grayanus.
Они имеют самый низкий коэффициент расхождения (около 0.81), а, следовательно,
самый высокий коэффициент сходства. Это было ожидаемо, так как оба моллюска
принадлежат к одному семейству(Mytilidae). Хотя даже это значение довольно
высоко, что говорит о значительном расхождении грибных комплексов даже у относительно
близких видов.
Полученные данные по соотношению различных групп мицелиальных
грибов, их биологическому разнообразию и степени расхождения доказывают, что
систематическое положение двустворчатого моллюска и его образ жизни влияют на
видовой состав ассоциированных с ним грибных комплексов.
Выводы
1. В ходе проведённого исследования нами было выделено
около 200 штаммов, идентифицировано 42 вида мицелиальных грибов.
2. Таксономический состав мицелиальных грибов,
ассоциированных с двустворчатыми моллюсками Modiolus modiolus, Crenomytilus
grayanus, Mizuhopecten yessoensis и Anadara broughtoni, составляет 42 вида
мицелиальных грибов, относящихся к 14 родам, из них - 7 видов сумчатых грибов,
33 вида анаморфных грибов и 2 вида зигомицетов.
. Наиболее заселёнными грибами органами являются: у
Mizuhopecten yessoensis - мантия и пищеварительная железа (по 6 видов), у
Modiolus modiolus - мускул (10 видов), у Crenomytilus grayanus - мускул и
пищеварительная железа (по 8 видов). Наименьшее количество видов грибов: у
приморского гребешка обнаружено в почках (1 вид), у мидии Грея и модиолуса
наименее заселёнными оказались жабры (1 вид).
. Наиболее близкими по видовому составу мицелиальных
грибов являются моллюски видов Modiolus modiolus и Crenomytilus grayanus.
. Грибные комплексы, ассоциированные с двустворчатыми
моллюсками, имеют различные показатели биоразнообразия и низкую степень
выравненности, что говорит о зависимости этих комплексов от видовой
принадлежности двустворчатого моллюска. Возможно, это так же связано с
характером грунта (гранулометрическим составом и содержанием органического
вещества), или образом жизни моллюска.
Полученные данные по соотношению различных групп мицелиальных
грибов, их биологическому разнообразию и степени расхождения доказывают, что
систематическое положение двустворчатого моллюска и его образ жизни влияют на
видовой состав ассоциированных с ним грибных комплексов.
Список литературы
1. Артемчук Н.Я. Микофлора морей СССР. М.: Наука, 1981. 192 с.
2. Бирюлина
М.Г. Современные запасы мидии в заливе Петра Великого // Вопросы гидробиологии
некоторых районов Тихого океана. Владивосток: ДВНЦ АН СССР, 1972а.-С. 11-21.
3. Брегман
Ю.Э. Взаимосвязь роста и энергетического обмена у некоторых промысловых донных
беспозвоночных залива Посьета (Японское море): Автореф. дис. канд. биол. наук.
- Владивосток, 1973. - 33 с.
. Временная
инструкция по технологии выращивания приморского гребешка после годичного
подращивания в садках, ТИНРО 1987. 11 с.
. Галышева
Ю. А., Христофорова Н. К. Среда и макрозообентос залива Восток Японского моря в
условиях рекреационного воздействия // Изв. ТИНРО. - 2007. - Vol. 149. - P.
6. Дудка
И.А. Водные несовершенные грибы СССР. Киев: Наукова думка, 1985. 188 с.
7. Дудка
И.А., Вассер С.П. Грибы. Справочник миколога и грибника. Киев: Наукова думка,
1987. 535 с.
8. Жюбикас
И.И Некоторые данные по биологии Pecten yessoensis Jay в Курило - Сахалинском
районе. - Вести. ЛГУ. Сер. Биология, 1969, №21, вып. 4, с. 21 - 32.
9. Зверева
Л. В., Щукина Г. Ф., Высоцкая М.А. Мицелиальные грибы - эпибионты
двустворчатого моллюска Corbicula japonica. Современная микология в России.
Первый съезд микологов России. Тезисы докладов. М.: Национальная академия
микологии, 2002
. Зверева
Л.В., Высоцкая М.А. Микобиота ресурсных видов двустворчатых моллюсков
российских вод Японского моря// Морские прибрежные экосистемы. Материалы второй
Международной научно-практической конференции. М.: Изд-во ВНИРО, 2005. С.
42-44.
. Лоция
северо-западного побережья Японского моря от реки Туманная до мыса Белкина. ГУНио
НО. М., 1981. - 319 с
. Михайлов
В.В., Кузнецова Т.А., Еляков Г.Б. Морские микроорганизмы и их вторичные
биологически активные метаболиты. Владивосток: Дальнаука, 1999. 132 с.
. Мотавкин.
П.А. Приморский гребешок. - Владивосток: ИБМ ДВО АН СССР, 1986. - 244 с.
14. Мэгарран
Э. Экологическое разнообразие и его измерение: пер. с англ. М.: Мир, 1992. 184
с.
. Пивкин
М.В., Алешко С.А., Красохин В.Б., Худякова Ю.В. Комплексы грибов губок южного
побережья острова Сахалин // Биол. моря. 2006.
16. Правила
рыболовства для Дальневосточного рыбохозяйственного бассейна, ТИНРО 2007.
. Скалкин
В.А. Биология и промысел приморского гребешка. Владивосток: Дальневосточное
книжное издательство, 1966. 30 с.
. Скарлато
О.А. Двустворчатые моллюски дальневосточных морей СССР (отряд Dysodonta).
Определитель по фауне СССР. - Л.: ЗИН АН СССР, 1960, т. 71. - 150 с.
. Скарлато
О.А. Двустворчатые моллюски умеренных широт западной части Тихого океана. - Л.:
Наука, 1981. - 479 с.
. Христофорова
Н. К., Наумов Ю.A., Арзамастев И.С. Тяжелые металлы в донных осадках залива
Восток (Японское море) // Изв. ТИНРО. - 2004. - Vol. 136. - P.
. Явнов
С. В. Атлас двустворчатых моллюсков дальневосточных морей Росси.// Атласы
промысловых и перспективных для промысла гидробионтов дальневосточных морей России.
- Владивосток: «Дюма», 2000. - 168 с.
. Alderman
D.J. Fungal diseases of marine animals // Recent Advances in Aquatic Mycology /
ed. E.B.G. Jones, ed. N. Y.: Wiley, 1976. P. 223-260.
. Alderman
D.J. Fungal infe'ction of crawfish (Palinurus elephas) exoskeleton // Trans.
Brit. mycol. Soc. 1973. V. 61. P. 595-597.
. Alderman
D.J. Fusarium solani causing an exoskeletal pathology in cultured lobsters,
Homarus vulgaris// Trans. Brit. Mycol. 1981. Vol. 76. pp. 25-27.
. Barlocher
F, Kendrick B. Fungi in the diet of Gammarus pseudolimnaeus (Amphipoda) //
Oikos. 1973. V. 24. P. 295-300.
. BarlocherF.,
KendrickB. Leaf-conditioning by microorganisms//Oecologia. 1975b. V. 20. P.
359-362.
. Becker
G. Biological investigations on marine borers in Berlin-Dahlem // Marine Boring
and Fouling Organisms / ed. D.L. Ray. / Univ. of Washington Press. Seattle,
1959. P. 62-83.
. Becker
G., Kampf W.-D., Kohlmeyer J. Zur Ernahrung der Holzbohrasseln der Gattung
Limnoria II Naturwissenschaften. 1957. Bd 44. S. 473-474.
. Biirlocher
F., Kendrick B. Assimilation efficiency of Gammarus pseudolimnaeus (Amphipoda)
feeding on fungal mycelium or autumn-shed leaves // Oikos. 1975a. V. 26. P.
55-59.
. Blant
J., Сорр В., Munro M., Northcote P., Prinsep M. Marine natural products // Nat.
Prod. Rep. 2004. V. 21. P. 1-49.
. Blaylock
R.B., OverstreetR.M., Klich M.A. Mycoses in red snapper {Lutjanus campechanus)
caused by two deuteromycete fungi (Penicillium corylophilum and Cladosporium
sphaerospermum) II Hydrobiologia. 2001. V. 460. P. 221-228.
. Corte
A.M. Osservazioni sul genere Lulworthia Suth. E sui suoi rapporti con Limnoria
Menzies e segnalazioni di alter specie // Bot. Ital. 1975. V. 109. P. 227-237.
. Cuomo
V., Palomba I., Guerriero A., D'Ambrosio M., Pietra F. Antimicrobial activities
from marine fungi //J. Mar. Biotechnol. 1995. V. 2. P. 199-204.
. Geiser
DM., Taylor J. W., Ritchie K.B., Smith, G. W. Cause of sea fan death in the
West Indies // Nature. 1998. V. 394. P. 137-138.
. Gessner
R. V., Goos R.D., Sieburth J.M. The fungal microcosm of the internodes of
Spartina altermflora II Mar. Biol. 1972. V. 16. P. 269-273.
. Hohnk
W. Ober die submersen Pilze an der rumanischen Schwarzmeerkiiste nahe Constanza
// Veroff. Inst. Meeresforsch. Bremerh. 1967. Suppl. 10. P. 149-158.
. Hohnk
W. Uber den pilzlichen Befall kalkiger Hartteile von Meerestieren // Ber.
Dtsch. Wiss. Komm. Meeresforsch. 1969. Bd. 20. S. 129-140.
38. Höller U., Wright A.D., Matthee G.F., Konig G.M., Draeger S., Aust
H.J., Schulz B. Fungi from marine sponges: diversity, biological activity and
secondary metabolites// Mycological research.2000. Vol. 104. pp. 1354-1365.
39. Hyde
K.D., Jones E.B., Leano E., Pointing S.B., Poonyth A.D., Vrijmoed L.P. Role of
fungi in marine ecosystems// Biodiversity and Conservation. 1998. Vol. 7. pp.
1147-1161.
. Johnson
P.T., Charman F.A. Infection with diatoms and other microorganisms in sea
urchin spines (Strongilocentrotus franciscanus)// J. Invertebr. Pothol. 1970.
Vol.17. pp. 94-106.
41. Johnson
T.W., Sparrow F.K. Fungi in Oceans and Estuaries. N.Y.: Weinheim, 1961. 668 p.
. Kim,
K., Kim P.D., Alker A.P., Harvell C.D. Chemical resistance of gorgonian corals
against fungal infections// Marine Biol. 2000.Vol.137. pp. 393-401.
. Kira
T. Shells of the western Pacific in color. Hokusha, Japan. 1962, 224 p., 72 pl
44. Kobayashi
M., Uehara H., Matsunami K., Aoki S., Kitagawa I. Trichoharzin, a new
polyketide produced by the imperfect fungus Tricoderma harzianum separated from
the marine sponge Mycale Cecillia //Tetrahedron Lett. 1993. V. 34. P.
7925-7928.
. Kohlmeyer
J. Beobachtungen iiber mediterrane Meerespilze sowie das Vorkommen von marinen
Moderfaule-Erregern in Aquariumszuchten holzzerstorender Meerestiere // Ber.
Dtsch. Bot. Ges. 1958. V. 71. P. 98-116.
. Kohlmeyer
J. Halophile Pilze von den Ufern Frankreichs // Nova Hedwigia. 1962a. V. 4. P.
389-20.
. Kohlmeyer
J. Marine fungi deteriorating chitin of hydrozoa and keratin-like annelid tubes
// Mar. Biol. 1972a. V. 12..P. 277-284.
. Kohlmeyer
J. New genera and species of higher fungi from the deep sea (1615-5315 m) //
Rev. Mycol. 1977. V. 41. P. 189-206.
. Kohlmeyer
J. The role of marine fungi in the penetration of calcareous substances // Am.
Zool. 1969d. V. 9. P. 741-746.
. Kohlmeyer
J. Uber Pleospora gandefroyi Patouillard // Willdenowia. 1962b. V. 3. P.
315-324.
. Kohlmeyer
J., Becker G, Kampf W. D. Versuche zur Kenntnis der Emahrung der Holzbohrassel
Limnoria tripunctata und ihre Beziehung zu holzzerstorenden Pilzen // Z. Angew.
Zool. 1959. V. 46. P. 457-89.
52. Kohlmeyer
J., Kohlmeyer E. Marine mycology. N.Y.: Academic Press, 1987, 690 p.
. Kohlmeyer
J., Kohlmeyer E. On the life history of marine Ascomycetes: Halosphaeria
mediosetigera and H. circumvestita //Nova Hedwigia. 1966. V. 12. P. 189-202.
. Kohlmeyer
J., Volkmann-Kohlmeyer B. New species of Koralionastes (Ascomycota) from the
Caribbean and Australia// Canadian Journal of Botany 1990. Vol. 68, pp.
1554-1559.
. KohlmeyerJ.,
Kohlmeyer E. Marine mycology, The Higher Fungi. N.Y., 1979. 690 p
56. KohlmeyerJ.,
Volkmann-Kohlmeyer В. Two Ascomycotina from coral reefs in the Caribbean and
Australia // Cryptogamic Botany. 1992. V. 2. P. 367-374.
. Kolliker
A. On the frequent occurence of vegetable parasites in the hard tissues of the
lower animals // Q. J. Microasc. Sci. 1860a. V. 8. P. 171-187.
. Kölliker A. On the frequentocurence of vegetable parasites in the
hard stucture of animals. - Proc. Roy. Soc. London, 1860, pp. 95-99.
. Kolliker
A. Uber das ausgebreitete Vorkommen von pflanzlichen Parasiten in den
Hartgebilden niederer Thiere // Z. Wiss. Zool. 1860b. V. 10. P. 215-232.
. Lane
C.E. The teredo // Sci. Am. 1961. V. 204. P. 132-142.
. Meyers
S.P, Feder W.A., Tsue K.M. Studies of relationships among nematodes and
filamentous fungi in the marine environment // Dev. Ind. Microbiol. 1964. V. 5.
P. 354-364.
. Meyers
S.P, Hopper B.E. Attraction of the marine nematode, Metoncholaimus sp., to fungal
substrates // Bull. Mar. Sci. 1966. V. 16. P. 142-150.
. Meyers
S.P, Hopper B.E. Studies on manne fungal nematode associations and plant
degradation // Helgol. Wiss. Meeresunfers. 1967. V. 15. P. 270-281.
. Molitoris
H.P, Schaumann K. Physiology of marine fungi: a screening programme for growth
and enzyme production // The biology of marine fungi. Cambridge: Cambridge
University Press, 1986. P. 34-47
. Mortensen
T. A monograph of the Echinoidea. Part 3. Aulodonta. Copenhagen.: C.A. Reitzel,
1940, 205 p
66. Namikoshi
M., Akano K., Kobayashi H., Koike Y., Kitazawa A., Rondonuwu A.B., Pratasik
S.B. Distribution of marine filamentous fungi associated with marine sponges in
coral reefs of Palau and Bunaken Island, Indonesia// Journal of Tokyo
University of Fisheries 2002. Vol. 88, pp. 15-20.
. Newell
S. Y, Fell J. W. Preliminary experimentation in the development of natuial food
analogues for culture of detritivorous shrimp // Univ. Miami Sea Grant. Tech.
Bull. 1975. V. 30. Pt 1-9. P. 1-115.
. Pivkin
M.V. Filamentous fungi associated with holothurians from the sea of Japan, of
the Primorye coast of Russia// Biol. Bull. 2000, Vol. 198. pp. 101-109.
. Pivkin
M. V. Filamentous fungi associated with holothurians from the Sea of Japan, off
the Primorye coast of Russia // Bioligical Bulletin. 2000. V. 198. P. 101-109.
70. Ritchie
D. A fungus flora of the sea // Science. 1954. V. 120. P. 578-579.
. Sallenave
C., Pouchus Y.F.,. Bardouil M., Lassus P., Roquebert M.F., Verbist J.F.
Bioaccumulation of mycotoxins by shellfish: Contamination of mussels by
metabolites of a Trichoderma koningii strain isolated in the marine
environment// Toxicon 1999. Vol.37. pp. 77-83.
. Sallenave-Namont
C., Pouchus Y.F., Robiou du Pont T., Lassus P., Verbist J.F. Toxigenic
saprophytic fungi in marine shellfish farming areas// Mycopatol. 2000. Vol.149.
pp.21-25.
73. Sterflinger
K., Hain M., Scholz J., Wasson K. Fungal infections of a colonial marine
invertebrate: diversity and morphological consequences// Facies №45. -
Erlangen, 2001. pp. 31-38.
. Stipanovic
R., Howell C. The structure gliovirin, a new antibiotic from Gliocladium virens
II J. Antibiot. 1982. V. 35. P. 1326-1330.
. Tubaki
K. Studies on the Japanese marine fungi. Lignicolous group. Annu. Rep. Inst.
Ferment. №4. - Osaka, 1969. - pp. 12-41.
. Turner
R.D. Identification of marine wood-boring mollusks // Marine Borers, Fungi and
Fouling Organisms of Wood/ eds E.B.G. Jones, S.K. Eitringham. Org. Econ. Coop.
Dev. P., 1971. P. 17-64.
77. Vishwakiran
Y, Thakur N.L., Raghukumar S., Yennawar P.L., Anil A. C. Spatial and Temporal
Distribution of Fungi and Wood-Borers in the Coastal Tropical Waters of Goa,
India // Botanica Marina. 2001. V. 44. P. 47-56.
. Vitellaro-Zuccarello
L. infrastructure of the byssal apparatus of Mytilus galloprovincialis.
Associated fungal hyphae // Mar. Biol. 1973. V. 22. P. 225-230.