Использование феромонных ловушек для борьбы со стволовыми вредителями леса
Использование
феромонных ловушек для борьбы со стволовыми вредителями леса
Введение
лес вредитель феромон защита
Защита лесов от вредителей и болезней одна из важнейших задач в деле
сохранения лесов и повышения их производительности. Лесозащитные мероприятия
взаимосвязаны со всеми лесохозяйственными и лесокультурными работами,
направленными на создание условий, необходимых для лучшего развития лесных
насаждений и предотвращения распространения в них вредителей и болезней.
Мероприятия по защите лесов от вредителей и болезней осуществляют все
лесохозяйственные организации, несущие ответственность за состояние лесов.
Общее руководство работой по защите лесов от вредителей и болезней в лесном
хозяйстве страны осуществляет Государственный комитет РФ по лесному хозяйству.
В краях и областях эту работу выполняют соответствующие органы лесного
хозяйства, а также лесохозяйственные предприятия (лесхозы) при консультации и с
помощью научно-исследовательских институтов лесного хозяйства и лесных опытных станций.
В лесах Томской области главными очагами размножения вредных насекомых
являются лесосеки, гари и реже насаждения, поврежденные первичными вредителями.
Лесосеки в некоторых районах области еще до сих пор находятся в
антисанитарном состоянии; на месте рубок остаются, порубочные остатки, вершины,
сучья, щепа и проч. На неочищенных лесосеках размножаются черные усачи хвойных
деревьев, рогохвосты, короеды, златки и сверлилы; некоторые виды короедов и
усачей, развиваясь под корой или в древесине порубочных остатков и
лесоматериалов, при своем дополнительном питании нападают на деревья около
лесосек, у которых грызут кору и сердцевину побегов и веточек, вызывая
ослабление и усыхание деревьев.
Целью данной работы является испытание феромонных ловушек для защиты
лесов Томской области от стволовых вредителей. Данная цель поставила следующие
задачи данного исследования.
. Ознакомиться с использованием половых феромонов в борьбе с вредителями
леса
. Провести испытание нового феромонного препарата «Деналол» для борьбы с
короедом-стенографом.
. Оценить эффективность этого препарата в условиях Западной Сибири.
Актуальность данной работы определяется тем, что защита лесов Томской
области от вредных лесных насекомых имеет большое хозяйственное значение. Пока
же ежегодно десятки тысяч кубометров заготовляемой деловой древесины
переводятся в низшие сорта в результате повреждения насекомыми, часть же
деловой древесины становится совершенно непригодной для использования в
строительстве.
Несмотря на большой экономический ущерб, приносимый вредными лесными
насекомыми сосновым насаждениям и лесоматериалам, эти насекомые до сих пор
остаются не изученными. Надо отметить, что изучение вредных лесных насекомых
Западной Сибири было начато Геблером в 1848 г. В своей работе он отмечает 13
видов короедов и 4 вида усачей-дровосеков южной части бывшей Томской губернии.
С.Д. Лавров (1926) для окрестностей города Омска указывает 20 видов
вредных лесных насекомых. Прозоров (1929) в своей работе «Гари в сосновых
лесах, как очаги заражения» для Барнаульского округа Чумышской лесной дачи
отмечает 23 вида короедов, 27 видов усачей (из них 17 видов вредных), 8 видов -
златок. Мамаев (1929) для Каменского округа указывает 2 вида златок, 4 вида
усачей, 2 вида слоников, 6 видов короедов, 2 вида хрущей. Месоед и Егоров
(1934) для ленточных боров Лебяжинской лесной дачи Барнаульского округа
приводят 11 видов короедов, 10 видов усачей, 5 видов златок, 2 вида слоников, 4
вида хрущей, 1 вид пилильщиков и 5-видов чешуекрылых.
В работах, непосредственно касающихся Томской области, отрывочные
сведения о вредных лесных насекомых можно найти в статьях экспедиции Зичи
(1901), С.М. Чугунова (1909), Г.Э. Иоганзена (1914 - 1915 и 1925), М.Д.
Русского (1937) и других. Кроме того следует указать исследования Е.Ф. Киселевой.
Они касаются, главным образом, одной из главнейших групп лесных вредителей, а
именно семейства короедов (1927, 1937, 1946) и также семейства усачей (1926).
Отдельно надо сказать об авторах, которые занимались изучением феромонов
насекомых. Начало этим исследованиям положили работы зарубежных исследователей
Дж. Харборна и М. Джекобсона. В своих трудах они рассмотрели особенности
феромонов различных насекомых, в частности, половые феромоны и предложили
способы их использования для борьбы с различными вредителями. В отечественной
науки к проблеме изучения феромонов обращались такие исследователи как Буда
В.Г., Скиркявичус А.В., Булеза В.В., Сафонкин А.Р., Мыттус Э.Р., Вайткявичене
Г.Б., Воронкова В.В., Щербакова Г.Д., Бочарова Н.И., Емельянов В.А., Зинкевич
Э.П., Исмаилов В.Я., Олещенко И.Н., Орлов В.Н., Алексеенко А.В. и многие другие
1. Стволовые вредители леса и борьба с ними
.1 Биологические особенности стволовых вредителей
На протяжении всего периода роста и развития лесные насаждения могут повреждаться
различными вредителями. По мере роста и формирования посадок появляются виды,
повреждающие листву деревьев и кустарников. Особое значение при этом
приобретают многоядные листогрызущие вредители (кольчатый шелкопряд,
златогузка), а также специализированные вредители листвы (ясеневая шпанка,
ильмовый ногохвост). Этих вредителей принято называть первичными. В дальнейшем
появляются и вредители скелетных частей - стволов, сучьев.
К стволовым, или вторичным, вредителям леса относятся насекомые из семейства
короедов Ipidae, усачей Cerambycidae, златок Buprestidae, долгоносиков
Curuliorudae, рогохвостов Siricidae, бабочек-древоточцев Cossidae, стеклянниц
Aegeriidae и некоторых других (Ильинский, 1962). Главная биологическая
особенность этих насекомых - скрытое обитание внутри ствола и ветвей - в лубе,
камбии и древесине, отчего их называют также скрытностволовыми вредителями. У
большинства видов ходы под корой и в древесине прокладывают личинки. У короедов
не только личинки, но и жуки обитают скрытно под корой или в древесине, где они
прокладывают ходы, размножаются и питаются (Воронцов, 1975).
Стволовые вредители причиняют деревьям значительный физиологический вред,
отчего деревья погибают. При прокладке ходов в древесине деревьям причиняется и
технический вред, приводящий к порче древесины. Поселение некоторых стволовых
вредителей не вызывает непосредственной гибели деревьев, но содействует их еще
большему ослаблению, появлению суховершинности, заражению опасными
заболеваниями.
Активность отдельных видов стволовых вредителей различна. По этому
признаку их можно разделить на три группы.
К группе физиологических, или физиолого-технических, вредителей относят
виды насекомых, поселяющихся на незначительно ослабленных деревьях, часто с
местным ослаблением отдельных ветвей, корневых лап или секторов ствола.
Поселение этих вредителей не угрожает деревьям непосредственной гибелью.
Нередко эти насекомые поселяются на одном и том же дереве в течение нескольких
поколений, что приводит к дальнейшему ослаблению дерева, заселению насекомыми
второй и третьей групп и после этого - к гибели. На срубленных деревьях и
лесоматериалах физиологические вредители не поселяются. К этой группе
вредителей относятся: алтайский усач, большой еловый лубоед, древесница
въедливая, бурый сосновый усач, черный сосновый усач, малый еловый черный усач,
большой пихтовый усач и др.
Вторая группа - это собственно вторичные вредители, поселяющиеся на
ослабленных, но еще жизнеспособных деревьях, а также на срубленных деревьях и
лесоматериалах. При массовой численности и отсутствии ослабленных деревьев
насекомые этой группы могут нападать на более здоровые деревья и после
многократных попыток их заселять и приводить к гибели. К этой группе относятся
большинство опасных видов короедов, усачей, златок, долгоносиков и рогохвостов.
К третьей группе относятся насекомые, поселяющиеся на деревьях,
потерявших свою жизнестойкость, отчего их можно назвать третичными вредителями
(Справочник по защите леса от вредителей и болезней, 1980). Эти насекомые
предпочитают заселять срубленные деревья, лесоматериалы, пни, валеж, порубочные
остатки. К ним относится ряд видов короедов, усачей, златок и др., (например,
полосатый древесинник, серый длинноусый усач, валежный короед и многие другие).
Наибольшую опасность и хозяйственную значимость имеют вредители первых двух
групп, из третьей - виды, причиняющие древесине технический вред (например,
полосатый древесинник).
Рассмотрим основные виды вредителей жуков-короедов, обитающих в лесах
Западной Сибири.
Серьезный вредитель сосны обыкновенный, кедра сибирского и корейского,
ели восточной - это шестизубчатый короед, или стенограф (Ips sexdentatus
Boern). Короед заселяет район толстой коры стволов чаще крупномерных растущих
ослабленных и срубленных деревьев, а также лесоматериалы. Жуки начинают летать
в конце второй декады мая при среднесуточной температуре 10 - 13º С, когда распускаются почки у дуба и
зацветает черемуха. Отложив яйца, жуки приступают к возобновительному питанию,
затем снова откладывают яйца, из которых развивается сестринское поколение.
Молодые жуки первого поколения вылетают в июле, несколько позднее жуки
сестринского поколения. Дополнительное и восстановительное питание жуки
проходят в местах развития или на других деревьях, где они прогрызают под корой
бессистемные каналы. Зимуют жуки в местах развития и дополнительного питания
или в подстилке вблизи усохших деревьев. Стенограф свето- и теплолюбив.
Предпочитает изреженные, прогреваемые и освещенные участки леса (Справочник по
защите леса от вредителей и болезней, 1980).
Другой вредитель - вершинный короед (Ips acuminatus Gyll) - в массе
размножается на сосне обыкновенной, вредит и другим хвойным породам. Зимовка,
периоды лета, сроки развития, число генераций, а также питание молодых и старых
жуков, как у шестизубчатого короеда. Вершинный короед заселяет растущие
ослабленные и срубленные деревья в районе тонкой коры, сучьев и ветвей, в массе
размножается на тонкокорых лесоматериалах и порубочных остатках. Вредит
соснякам всех возрастов. Предпочитает изреженные, освещенные и прогреваемые
места (Справочник по защите леса от вредителей и болезней, 1980).
Короед типограф (Ips typographus L.) - опаснейший вредитель елей обыкновенной, сибирской, аянской,
восточной. Вредит также кедру. Встречается и на других хвойных. Зимуют
различные стадии типографа: закончившие развитие молодые жуки - в подстилке в
радиусе 2 - 3 м вокруг свежеотработанного дерева, недопитавшиеся молодые жуки -
под корой в местах развития, молодые неразвитые жуки, куколки и личинки - здесь
же, но большинство из последних трех стадий вымерзает. Лет жуков начинается в
первой половине мая, когда среднесуточная температура достигает 15 - 20° С, а
температура подстилки, где зимуют жуки, 10° С. Фенологический сигнал -
распускание почек березы. Развитие потомства продолжается 60 - 70 дней. За этот
период сумма среднесуточных температур составляет около 740° С. Молодые жуки,
вылупившись из куколок, продолжают питание здесь же, под корой, и начинают
вылетать с первой декады июня, основывая второе поколение. В холодное и влажное
лето в средней полосе, а на севере ежегодно развивается только одно поколение.
Помимо основных, у типографа наблюдаются 2 - 3 сестринских поколения,
развивающихся на 2 - 3 недели позднее основных. Типограф заселяет стволы
растущих и срубленных деревьев в районе тонкой и толстой коры, неокоренные
лесоматериалы и порубочные остатки. Этот вредитель активно нападает на
жизнеспособные ели с зеленой хвоей, часто без внешних признаков ослабления
деревьев. Предпочитает условия умеренной освещенности и влажности (Воронцов,
1975).
Короед двойник (Ips duplicatus Sahlb) - обычный спутник короеда типографа, предпочитающий
селиться выше его, в районе тонкой коры и на вершине деревьев, в более
освещенных условиях. Начало лета и сроки развития несколько запаздывают по
сравнению с таковыми у короеда типографа. На зимовке жуки располагаются в
подстилке более дисперсно, в остальном не отличаются от него (Воронцов, 1978).
Короед гравер, или гравер обыкновенный (Pityogenes chalcographus L.) -
повреждает различные хвойные, предпочитая ель. Зимует в различных стадиях в
местах развития под корой деревьев. Личинки и молодые жуки обычно
перезимовывают благополучно. Лет жуков начинается одновременно с лётом
типографа и двойника, но более растянут - продолжается до июля. Молодые жуки
вылетают с конца июля, когда в жаркое и сухое лето может начаться развитие
второй генерации. Для развития потомства гравера необходима сумма
среднесуточных температур около 1000° С. Гравер поселяется на стволах в районе
тонкой коры и вершины, а также на ветвях растущих и срубленных деревьев, в
массе - на порубочных остатках и тонких лесоматериалах. На молодых деревьях
заселяет весь ствол. Светолюбив, но прямого солнечного освещения избегает
(Воронцов, 1978).
Продолговатый, или большой лиственничный, короед (Ips subelongatus Motsh)
вредит лиственнице сибирской, даурской, Сукачева. Зимуют жуки в местах
развития, в толще коры комлевой части ствола, в подстилке. Лет жуков - с
середины мая - в июне, когда завершается охвоение деревьев. Чуть позже жуки повторно
откладывают яйца, из которых развивается сестринское поколение. Молодые жуки
первого поколения появляются в первой декаде июля. Второе поколение завершает
развитие в сентябре. В северных районах ареала развивается лишь одно поколение.
Дополнительное и восстановительное питание проходит в местах развития и на
новых деревьях в минирных ходах. Заселяет весь ствол. Предпочитает условия
умеренной освещенности и влажности. Активно нападает на незначительно
ослабленные деревья, селится на срубленных деревьях и лесоматериалах
(Ильинский, 1962).
Западный непарный короед (Xyleborus (Anisandrus) dispar F.) повреждает
многие лиственные и некоторые хвойные, но чаще дуб, каштан, граб, орех грецкий.
Зимуют молодые жуки в маточных ходах. Здесь они спариваются. Вылетают в конце
апреля - в мае. Самка прогрызает в древесине маточный ход с несколькими
ответвлениями. Яйца откладывают кучками в 2 - 3 приема. Личинки питаются в
маточных ходах мицелием гриба, вносимого самкой. Окукливание в июне - июле.
Генерация одногодовая. Заселяет вершины и ветви, реже стволы ослабленных, но
еще жизнеспособных деревьев (нередко одни и те же в течение ряда лет),
содействуя их дальнейшему ослаблению и усыханию. На срубленные деревья,
лесоматериалы и пни не нападает (Ильинский, 1962).
По срокам развития различают весеннюю и летнюю фенологические подгруппы
стволовых вредителей. Насекомые, входящие в весеннюю подгруппу, заселяют
деревья в апреле-мае, в конце июня - в июле у них уже вылетает молодое
поколение. Вредители летней подгруппы заселяют деревья в июне - августе. Их
потомство остается обычно зимовать под корой или в древесине и завершает
развитие на следующий год или позже. От первопричины ослабления, биологических
особенностей древесной породы, условий местопроизрастания и образа жизни вредных
насекомых зависит процесс ослабления деревьев, их заселения стволовыми
вредителями и отмирания. Установлено 5 типов ослабления (отмирания) и заселения
деревьев: комлевой, или корневой, вершинный, одновременный, стволовый и местный
(Маслов и др., 1973).
Комлевой, или корневой тип ослабления и отмирания деревьев вызывается
причинами, нарушающими функции корневой системы или водопроводящей системы
комлевой части ствола: заражение дерева корневой губкой, повреждение устойчивым
низовым пожаром, затопление корней водой и т. п. Защитная реакция нарушается,
прежде всего, в комлевой части ствола, откуда и начинается заселение стволовыми
вредителями. Верхняя часть ствола и крона сохраняют на время защитную реакцию и
подвергаются нападению стволовых вредителей позднее, нередко, когда в комлевой
части ствола вредители уже готовы к вылету или даже вылетели. Появление в
комлевой части ствола первых поселенцев зависит от срока ослабления дерева.
Например, сосна в очаге корневой губки, ослабленная грибом, к весне заселяется
в районе толстой коры большим сосновым лубоедом, летающим в апреле-мае. Если
корневая губка ослабит сосну летом, то дерево на этом же участке ствола может
заселиться синей сосновой златкой или другими вредителями летней фенологической
подгруппы.
В результате нападения первопоселенцев на дереве образуется исходная, или
начальная, группировка стволовых вредителей. Последующее заселение дерева
стволовыми вредителями зависит от степени его ослабления, условий погоды,
обилия в лесу вредных насекомых и других причин. При резком снижении
устойчивости дерева, сухой и жаркой погоде и при большом числе вредителей
дерево может заселяться в районе тонкой коры уже через 1-2 недели после
заселения района толстой коры, в противном случае - через 2-3 месяца и более. При
летнем заселении комлевой части ствола нередки случаи, когда район тонкой коры
и кроны заселяется лишь весной следующего года. На сосне в верхней части ствола
в зависимости от срока заселения могут быть обнаружены малый сосновый лубоед,
вершинный короед и другие вредители. Группировки вредителей, образующиеся после
заселения верхней части ствола, получили наименование производных. На
заключительных этапах отмирания дерева к указанным вредителям как в нижней, так
и в верхней части ствола подселяются третичные вредители. На сосне это могут
быть полосатый древесинник, серый длинноусый усач, комлевые усачи и др.
Группировки насекомых на дереве становятся окончательными (Журавлев и др.,
1974).
Вершинный тип ослабления и отмирания деревьев обусловлен нарушением функции
кроны или водопроводящей системы верхней части ствола: повреждение хвои или
листвы насекомыми, поражение кроны сосудистыми заболеваниями или промышленными
эмиссиями и т.п. В этом случае дерево заселяется стволовыми вредителями и
отмирает в обратном порядке - вначале заселяются и сохнут крона и ствол в
районе тонкой коры, затем ствол в районе толстой коры. В обратном порядке, но в
основном аналогично формируются на дереве указанные группировки вредных
насекомых (Журавлев и др., 1974).
Одновременный тип ослабления и отмирания дерева наблюдается при
одновременном нарушении функций корневой системы и кроны, например, при
объедании хвои гусеницами в очаге корневой губки. По этому типу отмирают также
угнетенные деревья в насаждении. В зависимости от срока ослабления насекомые
заселяют деревья одновременно сверху донизу. Группировки вредителей
складываются по мере подселения одних видов насекомых к другим.
Стволовый тип ослабления и отмирания отличается тем, что защитная реакция
дерева нарушается в середине ствола (например, окольцевание его смоляным
раком). Заселение дерева вредные насекомые начинают от места повреждения и
распространяются вверх и вниз (Журавлев и др., 1974).
Местный тип ослабления и отмирания происходит при частичном повреждении
дерева: механическое повреждение ствола или корневой лапы (затес, ошмыг),
поражение отдельных ветвей или вершины каким-либо заболеванием и т.п. Насекомые
заселяют только отдельные части дерева (пораженную вершину или ветви, участки
ствола выше места повреждения), что не угрожает жизни всего дерева. Избыток в
лесу видов насекомых, не характерных для данного типа ослабления, может
привести к заселению и отмиранию деревьев по ложному типу (Журавлев и др.,
1974).
Очаги стволовых вредителей преимущественно комплексные. В них
одновременно размножаются несколько видов вредных насекомых (Маслов и др.,
1973.). Роль отдельных из них в гибели деревьев неодинакова. Защитные меры
осуществляются против всего комплекса видов, но с учетом особенностей каждого
из них. В связи с этим в справочнике описание биологии главнейших стволовых
вредителей, особенностей надзора и мер борьбы дается по группам видов,
заселяющих отдельные древесные породы.
1.2 Закономерности массовых размножений стволовых вредителей
Основное условие для массового размножения стволовых вредителей -
физиологическое ослабление насаждений. Поэтому важнейшим фактором, регулирующим
динамику численности стволовых вредителей, является наличие пищи в необходимом
количестве и соответствующего качества. Все факторы среды, ослабляющие
насаждения и обеспечивающие появление пищи, относят к группе факторов,
содействующих массовому размножению стволовых вредителей. К этим факторам
относятся: климатические - главным образом, ослабление насаждений в годы засух
или избыточного увлажнения осадками; стихийные-повреждение леса бурей, снегом,
ожеледью, пожарами и др.; биотические - повреждение леса хвое- и листогрызущими
насекомыми, поражение различными заболеваниями; антропогенные - связанные с
хозяйственной деятельностью человека (ослабление леса неправильной рубкой,
подсочкой, промышленными эмиссиями и т. п.) (Справочник по защите леса от
вредителей и болезней, 1980).
Важнейшим аспектом является специфическое воздействие, оказываемое на
насаждения той или иной древесной породы конкретным ослабляющим фактором.
Второй по значению фактор - погода, оказывающая на размножение стволовых
вредителей прямое и косвенное влияние. Прямое влияние погоды выражается в
ускорении и замедлении развития вредных насекомых. У поливольтинных видов это
сказывается в увеличении или снижении числа генераций в год (Маслов и др.,
1973). Экстремальные погодные условия могут подавлять размножение вредителей,
став причиной массовой гибели насекомых. Косвенное влияние погоды проявляется
через ослабление или оздоровление насаждений.
Главная причина естественного затухания очагов стволовых вредителей в
большинстве случаев заключается в исчерпывании пищевых ресурсов вследствие
полного использования ослабленных деревьев или их оздоровления. Размножившиеся
в массе вредители рассеиваются по окружающим насаждениям, большая часть из них
гибнет в попытках заселить более здоровые деревья. По периферии очага в
примыкающих насаждениях при этом наблюдается повышенный отпад деревьев.
Важное, но не определяющее регулирующее значение имеют конкурентные
межвидовые и внутривидовые взаимоотношения. Межвидовая конкуренция за пищу
возможна между отдельными видами стволовых вредителей, входящих в экологическую
группировку на заселенном дереве, а также между насекомыми и грибными
паразитами (рост мицелия опенка под корой иногда заметно снижает выживаемость
короедов ели и сосны). Своеобразны взаимоотношения между вредными короедами и
короедами-крошками из рода Crypturgus. Короеды-крошки проникают под кору сосны
и ели, используя входные отверстия, маточные ходы и яйцевые камеры
короедов-хозяев. В короткий срок, прокладывая 2-3-этажные ходы в толще коры,
короеды-крошки ее сильно протачивают и делают непригодной для пищи потомства
вредного вида, тем самым увеличивая его смертность. Отмечены также случаи
поедания яиц и жуков короеда-хозяина жуками короеда-крошки. Еловый
короед-крошка может ограничивать численность короеда типографа до 80 % и более
(Вредители сельскохозяйственных культур и лесных насаждений, 1975).
Внутривидовая конкуренция наиболее выражена у короедов. Высокая плотность
поселения им необходима, чтобы преодолеть защитную реакцию дерева. Однако
вследствие этого обостряется конкуренция особенно на стадии личинки.
Роль энтомофагов, как ограничивающих размножение факторов, велика. У
стволовых вредителей имеется огромное количество врагов из мира теплокровных
животных, птиц, насекомых и других членистоногих, червей, микроорганизмов.
Обычно стволовыми вредителями питается целый комплекс энтомофагов, и все же не
отмечены случаи полного затухания очагов этих вредителей под влиянием их
врагов. Конечная выживаемость стволовых вредителей, особенно короедов, низкая,
не более 15 % числа отложенных яиц, а чаще намного меньше (Маслов и др., 1973).
Стволовых вредителей можно обнаружить в любом лесу, в том числе здоровом,
на отставших в росте, угнетенных, а также единичных больных, ветровальных и
буреломных деревьях. Эти деревья выполняют роль резерваций для стволовых
вредителей. Действующим очагом стволовых вредителей считают ослабленное
насаждение или его часть (лесопатологический выдел), в котором количество
заселенных или свежеотработанных насекомыми деревьев (суммарный отпад)
превышает норму естественного отпада. При этом деревья относятся
преимущественно к основному пологу древостоя и расположены группами, куртинами
или сплошь. Насаждение в возрасте молодняка относится к очагу, если в нем
заселено вредителями не менее 5-7 % деревьев, в средневозрастных насаждениях -
не менее 3-5 %, в приспевающих, спелых и перестойных - не менее 2-3 %
(Справочник по защите леса от вредителей и болезней, 1980).
Наиболее часто стволовые вредители образуют локальные очаги размножения,
действующие на небольшой площади независимо друг от друга. Пандемические очаги
охватывают огромные территории областей или даже природных зон. Они возникают и
развиваются под влиянием причин глобального характера (засухи, эпифитотии) в
насаждениях ели обыкновенной, аянской и восточной, пихты кавказской,
европейской, дуба и ильмовых. Миграционные очаги возникают при массовом
перелете (миграции) насекомых из затухающих очагов в соседние ослабленные
насаждения или вырубки.
По длительности развития очаги подразделяются на эпизодические и
хронические. Первые из них действуют от 1 до 10 лет, вторые более 10 лет,
нередко десятки лет (Маслов и др., 1973). В процессе развития эпизодические и
хронические очаги могут проходить три фазы: первую - начальную, или роста
численности, когда в ослабленном насаждении нарастает численность стволовых
вредителей, в результате чего отпад деревьев начинает превышать естественную
норму; вторую - массового размножения, или максимальной численности, - в очаге
наблюдается наибольшая активность стволовых вредителей и соответственно этому
максимальный отпад в древостое; третью - кризиса, характеризующуюся резким
снижением численности вредных насекомых и спадом интенсивности усыхания
древостоя.
.3 Надзор, учет и прогноз стволовых вредителей
Задача специального надзора заключается в своевременном выявлении
массового размножения наиболее опасных видов стволовых вредителей, определении
состояния и динамики их очагов. Надзор осуществляется по группам стволовых
вредителей основных древесных пород. При этом учитывают, прежде всего,
физиологических и собственно вторичных вредителей, из третичных - технических
вредителей. В отдельных случаях надзор проводят за отдельными видами наиболее
опасных и специфичных вредителей, но учитывают при этом и их возможных
спутников.
Надзор подразделяют на рекогносцировочный и детальный (Наставление по
надзору, учету и прогнозу массовых размножений стволовых вредителей леса,
1975). Цель рекогносцировочного надзора - своевременное обнаружение массового
размножения стволовых вредителей, их видового состава и площади очага. Он
заключается в осмотре ослабленных насаждений, выявлении свежезаселенных
деревьев и глазомерной оценке их количества в процентах от общего числа
деревьев в древостое. За вредителями весенней подгруппы надзор проводится в
конце мая - в июне, за летней - в августе. При обнаружении очага определяют
видовой состав вредителей и площадь очага. Если очаг не обнаружен, повторный
осмотр осуществляют в следующие сроки надзора до момента полного оздоровления
насаждения. В горных лесах рекогносцировочный надзор проводят по маршрутным
ходам, прокладываемым по дорогам, тропам, водоразделам с последующим осмотром
выявленных очагов усыхания в насаждениях.
При необходимости более квалифицированной оценки состояния очага и
назначения санитарно-оздоровительных мероприятий проводят детальный надзор. Его
задача - определение динамики массового размножения стволовых вредителей и
угрозы жизнеспособности насаждений. Детальный надзор проводят в те же сроки,
что и рекогносцировочный. При этом надзоре применяется метод детальных
обследований с закладкой в очагах временных пробных площадей. При надзоре за
крупными однотипными очагами используют метод рекогносцировочных обследований с
закладкой временных пробных площадей в наиболее характерных, участках. Вместо
временных проб допустим метод перечета деревьев по непровешенным ходовым
линиям. В обоих случаях при перечете учитывают не менее 100 деревьев главной
породы.
Детальный надзор за развитием хронических очагов проводят на стационарных
пробных площадях, закладываемых не менее чем в трех наиболее характерных
повреждаемых участках насаждений. На стационарных пробных площадях деревья (не
менее 150 на пробу) нумеруют масляной краской, что позволяет проследить
динамику их индивидуального состояния (Наставление по надзору, учету и прогнозу
массовых размножений стволовых вредителей леса, 1975). Стационарные пробы
закладывают на 10 лет. После этого размещение их пересматривают. В случаях
более скорого усыхания насаждений стационарные пробы перезакладывают ранее
указанного срока. В особо важных случаях метод стационарных проб применим и для
надзора за развитием эпизодических очагов. При детальном надзоре уточняют
первопричину ослабления насаждений, определяют их состояние, устанавливают тип
отмирания деревьев, учитывают показатели размножения вредных насекомых,
определяют тип и фазу очага. Все эти данные используют для прогнозирования
размножения стволовых вредителей и разработки мероприятий по борьбе с ними.
Состояние насаждений определяют перечетом деревьев на пробных площадях с
использованием 6-балльной шкалы категорий состояния. В хвойных насаждениях
руководствуются при этом следующими придержками: I) условно здоровые деревья -
без внешних признаков ослабления; II) ослабленные - деревья со слабоажурной
кроной, укороченным приростом или поврежденной до 1/3
общего количества хвоей, с усыханием отдельных ветвей, повреждением отдельных
корневых лап или небольшим местным отмиранием ствола; III) сильно ослабленные -
деревья с ажурной кроной и матовой хвоей, с сильно укороченным приростом или
без него, с повреждением и усыханием до 2/3 хвои или
ветвей, суховершинные, со значительными повреждениями ствола и корневых лап;
IV) усыхающие - с сильноажурной и бледно-зеленой, желтеющей и осыпающейся
хвоей, с повреждением более 2/3 хвои, суховершинные, а
также с признаками заселения стволовыми вредителями; V) свежий сухостой -
усохшие в текущем году, с желтой или бурой хвоей или без нее; короеды вылетают
или вылетели, другие вредители еще могут быть под корой или в древесине; VI)
старый сухостой - деревья, усохшие в прошлые годы, без хвои, кора и мелкие
веточки частично или полностью осыпались, все вредители вылетели. Градации
ослабленности деревьев лиственных пород устанавливаются аналогичные по степени
ажурности и усыхания кроны, водяным побегам, повреждениям ствола и корневых
лап, заселенности стволовыми вредителями и пр. (Справочник по защите леса от
вредителей и болезней, 1980).
Ветровал и бурелом учитывается отдельно с подразделением на не
заселенный, заселенный и отработанный вредителями. Распределение деревьев по
категориям состояния выражается в процентах от общего числа деревьев данной породы
на пробе. Затем определяют среднюю категорию состояния насаждения как
средневзвешенную величину. Усыхание не всегда сопровождается заселением
вредителями, поэтому отдельно выделяются заселенные или отработанные вредными
насекомыми деревья.
Для определения показателей численности стволовых вредителей проводят
энтомологический анализ модельных деревьев из числа заселенных или
свежеотработанных вредными насекомыми. Техника анализа сводится в основном к
следующему. Заселенные или свежеотработанные стволовыми вредителями деревья (не
менее 3 на пробную площадь) срубают и очищают от сучьев. Устанавливают
протяженность (м) районов толстой и тонкой коры. Определяют расположение и
протяженность (м) фактических районов поселения главнейших видов вредителей и
сопоставляют их с протяженностью свойственных им оптимальных районов поселения
на данном дереве, выражая это соотношение в процентах (за 100 % берется
протяженность оптимального района поселения, характерного для данного вида). В
пределах фактических районов поселения каждого из главнейших вредителей
закладывают учетные палетки, на которых определяют плотность поселения,
продукцию, реальную плодовитость (у короедов), пораженность вредителей
энтомофагами и заболеваниями (Маслов и др., 1973).
Наиболее точный, но и наиболее трудоемкий способ учета, при котором
закладывают 3 круговые палетки, располагая их в начале, середине и конце
районов поселения. Длина палетки 50 см, для мелких видов короедов - не более 30
см, для усачей - до 1 м. Палетки совмещают, если районы поселения отдельных
видов вредителей совпадают или близки. Измерив окружность ствола в пределах
палетки, определяют ее площадь (дм2). Полученные результаты учета
пересчитывают на 1 дм2. Наименее точен способ прямоугольной палетки
размером 20´50 см, т.
е. 10 дм2. Ее закладывают в середине района поселения вредителя
(Справочник по защите леса от вредителей и болезней, 1980).
Плотность поселения, или абсолютную заселенность, у моногамных видов
короедов определяют как число маточных ходов на 1 дм2 поверхности коры
заселенного дерева а. Родительское поколение в целом характеризуется выражением
2а как сумма самцов и самок. У полигамных видов короедов плотность поселения
характеризуется как плотность на 1 дм2 брачных камер b и маточных ходов а, а родительское
поколение суммой а+b. Плотность
личинок усачей, златок, долгоносиков определяют числом личинок а под корой на 1
дм2 или их уходов в древесину b. Для некоторых видов усачей определяют иногда плотность
насечек, в которые самка отложила яйца.
Продукция Р, или численность молодого поколения, выражается числом
куколок или молодых жуков под корой или в древесине, либо количеством лётных
отверстий на 1 дм2.
Используя показатели плотности поселения и продукции, рассчитывают
энергию размножения С как отношение численности молодого поколения к
родительскому. Для моногамных видов короедов энергия размножения равна
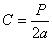
для
полигамных видов
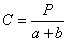
Для
усачей, златок и других вредителей определяют коэффициент выживания как
отношение плотности поселения нынешнего года (или продукции) к аналогичным
показателям прошлого года. Коэффициент выживания определяют так же, как
отношение числа особей, вылетевших из древесины, к числу особей, углубившихся в
нее. Плодовитость короедов, как было указано выше, можно определить косвенным
путем, измерив длину 10 маточных ходов и установив среднюю его протяженность
(мм) (Наставление по надзору, учету и прогнозу массовых размножений стволовых
вредителей леса, 1975).
Пораженность
паразитами и заболеваниями оценивают числом погибших особей в процентах от
общего числа потомства. Деятельность птиц можно определить таким же образом или
по проценту расклеванной коры. Об эффективности деятельности хищных насекомых
судят по их абсолютной плотности на 1 дм2.
Учет
специфических вредителей ведется иначе. Древесницу въедливую в стадии молодых
гусениц учитывают по числу заселенных побегов, выделяющихся увяданием и
наличием буровой муки у развилок ветвей, а после вылета бабочек - по экзувиям,
торчащим из лётных отверстий в нижней части ствола. Для более точного учета
гусениц раскалывают 2-метровые отрубки нижней части не менее чем трех деревьев.
Древоточца пахучего учитывают при детальном надзоре тем же способом. Для учета
заселенности деревьев большой тополевой стеклянницей и большим тополевым усачом
необходимо раскалывать 1,5-метровые отрубки комлевой части тополей и верхние
корни (не менее 3 деревьев на пробу). Поселения тополевого скрытнохоботника
подсчитывают, раскалывая 0,5-метровые отрубки стволиков заселенных деревьев
(также не менее 3) (Справочник по защите леса от вредителей и болезней, 1980).
В
крупных таежных массивах в пределах действующих или проектируемых лесосырьевых
баз очаги стволовых вредителей выявляют в основном при авиапатрулировании. При
этом обязателен специальный надзор за развитием очагов путем рекогносцировочных
обследований с закладкой временных пробных площадей и анализа модельных
деревьев. Техника учетных работ та же, но надзор проводят только раз в году - в
августе.
На
больших площадях поврежденного леса предварительно проводят аэровизуальный
облет или обследуют очаги с воздуха. В лесах вне действующих или проектируемых
лесосырьевых баз ограничиваются наземной проверкой очагов, выявленных с
воздуха. Надзор за сибирским шелкопрядом и другими массовыми хвоегрызущими
вредителями таежных лесов рекомендуется сочетать с надзором за состоянием
древостоев и их поврежденностью стволовыми вредителями.
Различают
долгосрочные и краткосрочные прогнозы. Долгосрочный прогноз составляется на 1-3
года. Задача этого прогноза заключается в определении возможности появления
очагов стволовых вредителей, а если они возникли, - сроков и степени
максимальной поврежденности древостоев.
Цель
краткосрочного прогноза - установить угрозу насаждениям в период поселения и
развития последующей генерации вредителей. Для видов с одногодовой и более
длительной генерацией он составляется на следующий год. Для видов с 2 и более
генерациями в год он составляется при надзоре за весенней подгруппой на летний
период, при надзоре за летней подгруппой - на весну следующего года. Для этого
используют полученные при надзоре показатели состояния насаждений и численности
вредных насекомых.
1.4 Меры борьбы со стволовыми вредителями
Перечислим основные меры борьбы со стволовыми вредителями.
Лесохозяйственная профилактика. Лесохозяйственные профилактические
мероприятия направлены на создание условий, исключающих ослабление деревьев и
заселение их стволовыми вредителями. Выращивание устойчивых насаждений путем
подбора древесных пород в соответствии с условиями местопроизрастания, выбора
оптимальной густоты, схемы посадки и смешения с другими древесными и
кустарниковыми породами - важное условие для предотвращения их ослабления и
усыхания. Особое значение придается также селекции, отбору и разведению
устойчивых видов, сортов и форм древесных пород.
Используемая в хозяйствах система и технология рубок главного и
промежуточного пользования не должна ослаблять и расстраивать насаждения.
Необходимо регулировать, а местами и запрещать пастьбу скота в лесу, приводящую
к механическим повреждениям корней, уплотнению почвы, уничтожению травяного
покрова, подлеска и подроста. Более эффективной должна быть защита леса от
пожаров, хвое- и листогрызущих насекомых (Надзор, учет и прогноз массовых
размножений хвое- и листогрызущих насекомых в лесах СССР, 1965), различных
заболеваний и других форм ослабления леса, в том числе обусловленных
неправильной хозяйственной деятельностью человека.
Санитарная профилактика. «Санитарные правила в лесах Российской
Федерации», утвержденные приказом Рослесхоза от 15 января 1998 г. № 10 (с
изменениями от 24 декабря 1998 г.), устанавливают цели и виды
санитарно-оздоровительных мероприятий и санитарные требования, обеспечивающие
сохранение биологической устойчивости лесных насаждений при ведении лесного
хозяйства и лесопользовании в лесах Российской Федерации, а также при
производстве работ, не связанных с ведением лесного хозяйства и
лесопользованием. «Санитарные правила в лесах Российской Федерации» (1998 г.) обязательны
к исполнению всеми юридическими лицами и физическими лицами, производящими
работу в лесу, регламентируют санитарные требования при рубке и подсочке леса,
хранении и транспортировке лесоматериалов. Цель правил не только предотвратить
непосредственный вред насаждениям и лесопродукции от стволовых вредителей, но и
снизить общий запас этих насекомых в лесу и тем самым ликвидировать угрозу их
массового размножения. Выборочные санитарные рубки проводятся с целью
оздоровления насаждений, предупреждения распространения и (или) ликвидации
очагов стволовых вредителей и опасных инфекционных заболеваний. Их проводят в
насаждениях с повышенным, по сравнению с естественным, текущим отпадом
деревьев, наличием ветровала, бурелома, снеголома, снеговала, пораженных болезнями
и заселенных стволовыми вредителями деревьев. Величина естественного текущего
отпада определяется по таблицам хода роста, принятым для лесоустроительных
работ на территории региона (Приказ Рослесхоза от 15 января 1998 г. № 10
"Об утверждении Санитарных правил в лесах Российской Федерации" (с
изменениями от 24 декабря 1998 г.)).
Отвод лесосек под санитарные рубки в очагах стволовых вредителей в зоне
действия лесовозных и лесохозяйственных дорог проводится не более чем за три
месяца, а в остальных случаях не более чем за полгода до их проведения.
Лесосеки под санитарные рубки в лиственных насаждениях отводятся только в
весенне-летний период (май - август) при наличии листвы на деревьях, кроме
участков ветровала и бурелома. Отвод лесосек под санитарные рубки, проводимые в
весенне-летний период, осуществляется не более чем за один месяц до начала
рубки.
При всех видах санитарных рубок должна быть обеспечена очистка лесосек от
порубочных остатков и своевременная вывозка древесины из леса либо ее немедленная
защита согласно нормативным документам по защите заготовленной древесины и
лесоматериалов от заселения вредителями и поражения болезнями.
Очистка мест рубок от порубочных остатков производится в соответствии с
Правилами отпуска древесины на корню в лесах Российской Федерации,
утвержденными постановлением Правительства Российской Федерации от 01.06.98 №
551 (Постановление Правительства Российской Федерации от 01.06.98 № 551 «Об
утверждении Правил отпуска древесины на корню в лесах Российской Федерации»), и
региональными правилами рубок главного пользования. В очагах стволовых
вредителей, сосудистых и некрозно-раковых болезней порубочные остатки подлежат
обязательному сжиганию с соблюдением требований Правил пожарной безопасности в
лесах Российской Федерации.
Хранение в лесу заготовленной древесины хвойных и лиственных пород
допускается при условии обязательной ее защиты от заселения стволовыми
вредителями и поражения грибами в сроки согласно зонированию территории
Российской Федерации по срокам запрета хранения (оставления) в лесу неокоренной
(незащищенной) древесины и методами, соответствующими нормативным документам по
защите заготовленной древесины и лесоматериалов от заселения вредителями и
поражения болезнями.
Особенно опасно захламление лесосек крупными порубочными остатками
(толщиной более 5 см), на которых наиболее успешно развиваются опасные виды
вредителей. Для поддержания численности валежных видов короедов, за счет
которых сохраняются многие энтомофаги стволовых вредителей, рекомендуется
весной, после осенне-зимне-весенних рубок, укладывать мелкие сучья в кучи в
сосновых насаждениях в объеме 50 % по окраинам вырубок, в еловых насаждениях
30-40 % по всей площади вырубок (в противопожарных целях кучи сучьев
опахивают). При угрозе массового размножения стволовых вредителей на порубочных
остатках, где невозможен огневой способ уничтожения, разрешается химическая
обработка (при низкой численности энтомофагов) (Приказ Рослесхоза от 15 января
1998 г. № 10 "Об утверждении Санитарных правил в лесах Российской
Федерации" (с изменениями от 24 декабря 1998 г.)).
Для создания благоприятных условий поселения и развития вредных насекомых
при всех видах рубок высота пней не должна превышать 1/3
диаметра среза. При рубке деревьев тоньше 30 см - не выше 10 см, считая высоту
от шейки корня. Окорка пней производится лишь при опасности размножения
каких-либо вредителей (например, дубовой двупятнистой златки). Окорка не
препятствует заселению пней большим сосновым долгоносиком и
короедами-корнежилами, поэтому в случае опасности их размножения рекомендуется
корчевка пней или опрыскивание их инсектицидами.
Санитарные правила запрещают оставлять в лесу и на расстоянии менее 2 км
от леса заготовленную неокоренную или не защищенную химическими или иными
способами от вредителей и невывезенную древесину. Календарные сроки этих
мероприятий дифференцированы в зональном разрезе. Древесину хвойных пород,
дуба, ясеня, ильмовых, заготовленную в весенне-летний период, необходимо
вывезти из леса, или окорить, или обработать инсектицидами в 10-дневный срок.
Для защиты древесины, оставляемой на летний период в лесу, применяют окорку,
обработку инсектицидами или организуют ее хранение сухим и влажным способами.
При сухом способе древесину после полной или частичной окорки укладывают в
рыхлые штабеля, в южных районах - в плотные. Одновременно проводят ее защиту от
грибных заболеваний и растрескивания (антисептирование, обмазку торцов и т.
д.). При влажном способе древесину, уложенную в плотные штабеля, затеняют,
орошают или погружают в воду. Наиболее целесообразно орошение штабеля
дождеванием. На севере штабель укладывают на снежно-ледяное основание,
переслаивая бревна снегом. Если хранящуюся этим способом древесину все же
заселяют вредные насекомые, ее окоряют или обрабатывают инсектицидами. Перевозить
заселенную стволовыми вредителями древесину запрещено, ее следует
предварительно окорить или подвергнуть химобработке. Не разрешается
обрабатывать химикатами древесину, подлежащую сплаву (Справочник механизатора
лесного хозяйства, 1977).
При рубках ухода за лесом следует выбирать деревья, заселенные
вредителями, пораженные болезнями, а также ослабленные, суховершинные и т.д.
Подсочка в очагах вредителей и болезней запрещена. Перед началом подсочки
следует провести выборочные санитарные рубки, а при ее ведении - не подсачивать
больные и угнетенные деревья. В случае возникновения в заподсоченном насаждении
очага вредителей и болезней подсочку приостанавливают.
Необходимость борьбы, объем и сроки ее проведения определяют по
результатам надзора, долгосрочного и краткосрочного прогнозов, а также с учетом
условий и возможностей хозяйства. Вначале принимаются меры организационного
характера: усиливают систему надзора, вносят коррективы в планы рубок,
обеспечивают необходимой техникой, средствами и др. Выбор конкретных
оздоровительных мероприятий зависит от степени и характера ослабления
древостоя, фазы очага, биологических особенностей древесной породы и вредных
насекомых, хозяйственной целесообразности.
В незначительно ослабленных древостоях принятые меры должны содействовать
их оздоровлению. В сомнительных случаях решение вопроса о борьбе откладывают до
следующего срока надзора, когда жизнестойкость насаждений проявится более
четко. Сильно ослабленные насаждения, если их нельзя спасти, назначают в
сплошную санитарную рубку до массового размножения стволовых вредителей. В
эпизодических очагах борьбу назначают в год возникновения очага, в крайнем
случае на следующий год, когда вредителей мало и борьба наиболее эффективна и
наименее трудоемка. В хронических очагах необходимо устранить причину
ослабления насаждений. Если это невозможно, борьба со стволовыми вредителями
должна быть направлена на снижение интенсивности усыхания насаждений, улучшение
санитарного состояния лесов. В таких очагах борьбу со стволовыми вредителями
следует осуществлять систематически. При этом в определенных случаях уборке
подлежат не только усыхающие, заселенные вредителями деревья, но и сильно
ослабленные (например, корневой губкой или голландской болезнью) деревья,
являющиеся пищевой базой для насекомых. Своевременная уборка ветровала,
бурелома и отдельных больных деревьев содействует снижению резерваций стволовых
вредителей. Борьба считается эффективной, если после ее проведения не будет
нового заселения деревьев. В противном случае, а также если сохраняется
ослабленность насаждений, надзор за их состоянием продолжается, а при
необходимости меры борьбы применяются повторно (Справочник по защите леса от
вредителей и болезней, 1980).
К основным оздоровительным мероприятиям относятся выборочные и сплошные
санитарные рубки, выкладка ловчих деревьев. При выборочных санитарных рубках
вырубают деревья, заселенные стволовыми вредителями, зараженные болезнями, а
также убирают ветровал, бурелом, снеголом, снеговал и мертвые деревья.
Важнейшее истребительное мероприятие - выборка свежезаселенных деревьев. Ее
проводят с учетом двух фенологических групп стволовых вредителей - весенней и
летней. Деревья, заселенные вредителями весенней подгруппы, вырубают в июне, до
вылета молодых жуков короедов. Деревья, заселенные вредителями летней
подгруппы, вырубают осенью или зимой, но отбирают деревья в рубку в августе - в
начале сентября. Отбор производят с учетом признаков заселения, характерных для
того или иного типа отмирания деревьев данной породы. С целью уничтожения
вредителей вырубленные деревья окоряют или опрыскивают инсектицидами до начала
их вылета из-под коры. При окорке кору и сучья сжигают или закапывают в землю
(Справочник по защите леса от вредителей и болезней, 1980).
В случаях, если выборочные санитарные рубки могут привести насаждения к
расстройству, назначают сплошные санитарные рубки. Обычно к ним прибегают при
угрозе усыхания более 30-10 % деревьев. При этом следует учитывать устойчивость
древесной породы к разреживанию в конкретных условиях роста, возраст и ценность
древостоев, условия эксплуатации и т. д. При этих рубках принимают меры (окорку
или химобработку) к уничтожению поселившихся вредителей и защите от них
незаселенной части древесины. Сплошные санитарные рубки целесообразнее проводить
в осенне-зимний период.
После выборочных санитарных рубок практикуют выкладку ловчих деревьев.
Такое мероприятие рекомендуется проводить в относительно устойчивых
насаждениях, чтобы растущие ослабленные деревья не отвлекали насекомых от
ловчего материала. В качестве ловчих используют ветровал, бурелом, свободные от
заселения участки ствола усыхающих деревьев, а также больные, ослабленные
деревья. В феврале - марте выкладывают ловчие деревья для вредителей весенней
подгруппы, в июне - для летней. Целесообразно выкладывать деревья группами по
2-3 шт., в количестве, обусловленном численностью молодого поколения
вредителей. Так, если численность молодого поколения высока, ловчие деревья
выкладывают в количестве, равном половине заселенных Деревьев в очаге, при
средней численности - от 1/2 до 1/4
количества заселенных деревьев, при низкой - не выше 1/4
количества заселенных деревьев. При низкой численности молодого поколения
выкладывать ловчие деревья нецелесообразно.
Для тенелюбивых видов насекомых ловчие деревья выкладывают в тени, для
светолюбивых - на свету. Следует избегать условий крайне высокой затененности и
освещенности. Ловчие деревья выкладывают с кроной, комлем на пень или
подкладку. После полного заселения дерева и прекращения откладки яиц насекомыми
ловчие деревья следует окорить, кору и ветви сжечь или закопать в землю.
Целесообразна выкладка ловчих деревьев, опрыснутых инсектицидами.
Химические меры борьбы. В связи с длительным периодом лета стволовых
вредителей химическая борьба с ними может быть успешной лишь при использовании
препаратов, сохраняющих свою токсичность в течение достаточно длительного
времени. Этим требованиям отвечают 16 %-ная минерально-масляная эмульсия
гамма-изомера ГХЦГ, технический гексахлоран и технический гамма-изомер гексахлорана.
Инсектициды применяют для истребительной и защитной обработки древесины,
опрыскивания ловчих деревьев, обработки порубочных остатков и пней, а также
мест зимовок короедов, химической защиты ослабленных деревьев. Для борьбы с
физиологическими вредителями (древесницой въедливой, тополевыми стеклянницами и
др.) рекомендуется использовать препараты системного действия (Справочник по
пестицидам (гигиена применения и токсикология), 1977).
Истребительную обработку используют для уничтожения вредителей взамен
окорки заселенной древесины, заготовленной при санитарных рубках леса, или
лесопродукции, заселенной насекомыми при ее хранении в лесу летом. Обработку
проводят перед началом вылета жуков, при появлении куколок вредителей.
Заселенную древесину не обрабатывают, если обнаружена высокая степень
пораженности вредителей паразитическими и хищными насекомыми (70 % и более).
Для истребительной обработки древесины, заселенной короедами, используют 2-3
%-ную (по препарату) минерально-масляную эмульсию 16 %-ного гамма-изомера ГХЦГ
или 1-2 %-ный раствор технического ГХЦГ в дизельном топливе, соляровом и других
легких минеральных маслах. Если древесина заселена усачами или златками,
концентрацию рабочих жидкостей повышают в 2-3 раза (максимальная растворимость
технического ГХЦГ около 4 %). Расход рабочей жидкости от 0,3 до 1 л/м2
поверхности коры, в зависимости от ее характера: сосна, ель - 0,3-0,4 л/м2;
ильмовые, дуб 0,6-0,8; лиственница до 1 л/м2. При обработке
гладкокорых материалов расход рабочей жидкости минимальный, толстокорых и
сильно трещиноватых - максимальный. При мелкокапельном опрыскивании древесины
указанные нормы расхода рабочих жидкостей снижают на 1/3
(Журавлев и др., 1974).
Химическую защитную обработку проводят, чтобы предотвратить заселение
вредными насекомыми свежезаготовленной неокоренной лесопродукции, оставляемой
на хранение в лесу на летний период, и не прибегать к дорогой и трудоемкой
окорке. Древесину осенне-зимней заготовки обрабатывают весной перед лётом
весенней подгруппы стволовых вредителей, сосновую древесину осенне-зимней
заготовки - в первой половине апреля, при поздней весне - в последнюю декаду
апреля. Еловую древесину опрыскивают на 2 недели позднее, во второй половине
апреля - начале мая. Древесина лиственницы подлежит обработке с середины мая,
ильмовых пород - не позднее конца апреля, а при наличии в лесу ильмовых
лубоедов - в первой декаде апреля. Дубовую древесину обрабатывают во
второй-третьей декадах апреля, при отсутствии красного дубового усача - в
первой половине мая. Древесину весенне-летней заготовки опрыскивают в
10-дневный срок. Для защиты неокоренной древесины используют те же препараты и
нормы расхода, что и при истребительной обработке заселенной лесопродукции.
На складах древесину обрабатывают послойно по мере укладки ее в штабеля.
В условиях таежной зоны допускается опрыскивание заштабелеванной древесины.
Расход рабочей жидкости при этом составляет 1 л/м2 поверхности
штабеля. Штабель плотно уложенной древесины обрабатывают сверху и с боковых
сторон. При обработке штабелей особенно эффективно мелкокапельное опрыскивание
при помощи аэрозольных генераторов. Расход рабочей жидкости при такой обработке
можно снизить в 2-4 раза. Чтобы предотвратить разложение инсектицида от
действия солнечных лучей, штабель прикрывают лапником или деревянными щитами.
За обработанной древесиной устанавливают контроль. При обнаружении успешных
поселений вредителей обработку повторяют (Справочник по защите леса от
вредителей и болезней, 1980).
Отравленные ловчие деревья не отпугивают стволовых вредителей, окорка их
не требуется. Применяя обработку инсектицидами, число подлежащих выкладке
ловчих деревьев можно уменьшить вдвое. Ловчие деревья обрабатывают
инсектицидами перед лётом стволовых вредителей. Для этой цели используют только
16 %-ную эмульсию гамма-ГХЦГ в тех же концентрациях, что и при истребительной
обработке древесины. Норма расхода рабочей жидкости та же. Обрабатывают ствол и
крону. Масляные растворы ожигают тонкую кору ловчих деревьев, поэтому применять
их для этой цели не рекомендуется. В южных районах через 1-2 месяца иногда
требуется повторная обработка ловчих деревьев вследствие разложения
инсектицида. Порубочные остатки, собранные в кучи или валы, опрыскивают 2-3
%-ной эмульсией гамма-ГХЦГ или 1-2 %-ным масляным раствором технического ГХЦГ.
Расход рабочей жидкости - 1 л на 1 м3 остатков, в южных районах -
1,5-2 л/м3.
Для обработки пней дуба при опасности заселения их двупятнистой и другими
узкотелыми златками, а также большим и малым дубовым усачами используют 3-5
%-ную эмульсию гамма-гексахлорана. Расход на пень 0,3-0,6 л рабочей жидкости.
Масляные растворы, ожигающие поросль, всходы и подрост, для этой цели не
используют. Этим способом можно защищать от заселения пни и других древесных
пород. Для опрыскивания зимовок большого соснового лубоеда применяют 2-3 %-ную
эмульсию или 1-2 %-ный раствор гамма-гексахлорана. Расход рабочей жидкости на
дерево 0,4-0,6 л. Осенью или ранней весной обрабатывают комлевую часть ствола и
корневые лапы, где располагаются зимующие жуки лубоеда. Для борьбы с короедами
типографом и двойником осенью или весной опрыскивают лесную подстилку в радиусе
1-2 м вокруг отработанных этими короедами деревьев. Расход рабочей жидкости
0,25-0,5 л/м2. Против зимующих жуков ильмовых лубоедов поздней
осенью обрабатывают нижнюю часть стволов растущих деревьев на высоту до 3-4 м.
Расход рабочей жидкости тех же препаратов 0,60,8 л/м2 поверхности
обрабатываемых деревьев (Справочник по защите леса от вредителей и болезней,
1980).
Для защиты ослабленных деревьев при комлевом типе их отмирания используют
масляную эмульсию гамма-изомера гексахлорана и раствор технического
гексахлорана в тех же концентрациях и с тем же расходом рабочей жидкости, что и
при защитной обработке древесины. Обрабатывают нижнюю часть стволов деревьев до
высоты 3 - 4 м.
При вершинном типе ослабления у деревьев опрыскивают кроны. Для этого
применяют только эмульсию гамма-гексахлорана с концентрацией по препарату 5 %.
Расход рабочей жидкости около 10 л на дерево высотой 5 - 7 м (Справочник
механизатора лесного хозяйства, 1977).
Биологический метод борьбы. Согласно уставу Международной организации
биологической борьбы (МОББ), принятому в 1971 г., под биологическим методом
понимают использование живых существ или продуктов их жизнедеятельности для
предотвращения или уменьшения ущерба, причиняемого вредными организмами.
К числу живых существ, применяемых в борьбе с вредителями растений,
относятся их естественные враги, т.е. хищные и паразитические насекомые и
клещи, нематоды и простейшие, млекопитающие и птицы, болезнетворные организмы -
бактерии, вирусы, грибы. Из продуктов жизнедеятельности живых существ в
биологической борьбе в последние годы начинают применять половые аттрактанты,
или половые феромоны, а также гормоны и их синтетические аналоги (Справочник по
защите леса от вредителей и болезней, 1980).
Известны также успешные опыты по использованию половых аттрактантов в
качестве средства для истребления вредителей. Это достигается применением
феромона в виде инсектицидных ловущек, созданием „самцового вакуума" и дезориентацией
самцов. Так, применение в инсектицидных ловушках смеси феромона и инсектицида
позволяло снизить расход последнего до 0,5...2 г/га (Крушев, 1973).
2. Использование феромонов в борьбе с вредителями леса
.1 Общая характеристика феромонов
Для поддержания жизнеспособности популяций, для обеспечения стабильных
показателей ее воспроизводства очень существенное значение имеет внутривидовая
химическая коммуникация, которая осуществляется и регулируется с помощью
феромонов. Название феромон происходит от греческих слов jέρω - приносить, причинять, производить,
управлять и ορμάω (ορμαίω) - приводить в движение, побуждать,
стремиться, желать.
По определению Петера Карлсона и Мартина Люшера, предложивших этот
термин, «феромоны - это вещества, вырабатываемые и выделяемые в окружающую
среду живыми организмами (часто с помощью специализированных желез) и
вызывающие специфическую ответную реакцию (характерное поведение или
характерный процесс развития) у воспринимающих их особей того же биологического
вида» (Барбье, 1978).
С учетом новых данных можно дать такой вариант определения термина
«феромон».
Феромоны - это комплексы (наборы, комбинации) веществ или индивидуальные
вещества, выделяемые во внешнюю среду организмами, несущие функцию передачи
информации и часто вызывающие определенную реакцию (характерное поведение,
физиологические процессы, процессы развития) либо, напротив, ингибирующие те
или иные формы поведения или физиологические процессы у воспринимающих их
особей обычно того же биологического вида.
Высказывалось предположение (Киршенблат, 1958) использовать для
обозначения всех биологически активных веществ, которые выделяются во внешнюю
среду и оказывают воздействие на другие организмы (т. е. и тех веществ, которые
стали называть феромонами), термин «телергоны». Однако укоренился именно термин
«феромоны».
Среди феромонов выделяют две группы: феромоны-релизеры и
феромоны-праймеры (или праймер-феромоны - primer pheromones). Впервые такое деление провел Е.О. Уилсон (Харборн,
1985).
Феромоны-релизеры вызывают немедленные поведенческие эффекты (изменение
поведения) у реципиентов. Примеры феромонов-релизеров - половые феромоны,
феромоны тревоги, следа и феромоны мечения территории.
Праймер-феромоны вызывают длительные физиологические эффекты (изменения)
в воспринимающем организме. Примеры праймер-феромонов - вещества, регулирующие
принадлежность особей общественных насекомых к определенной «касте», а также
вещества, которые, по-видимому, могут воздействовать на срок наступления
половой зрелости у самок млекопитающих или синхронизировать менструальные циклы
у женщин (Введение в проблемы биохимической экологии, 1990).
Внутривидовые эколого-биохимические взаимодействия организмов имеют еще
некоторые аспекты, выходящие за рамки классических представлений о феромонах.
Так, у некоторых организмов (и позвоночных и беспозвоночных) установлены
факты ингибирования роста под влиянием экзометаболитов других особей того же
вида (Шварц и др., 1976).
Однако наиболее обширный фактический материал в области химических
внутривидовых взаимодействий касается именно феромонов.
Хотя феромоны найдены у самых различных представителей животного мира,
большая часть наших сведений об этих соединениях получена на основании опытов с
насекомыми. Отчасти это объясняется тем, что с помощью феромонов сравнительно
легко управлять активностью насекомых, тогда как млекопитающие значительно
труднее поддаются такому контролю. Отчасти же это связано с тем, что изучение
феромонов насекомых имеет практическое значение. Идентификация феромонов у насекомых
сразу же дает в руки исследователей средство управления размером популяции, а в
случае насекомых-вредителей - и средство борьбы с ними.
Выделение разнообразных веществ из организма насекомого складывается из 3
различных процессов - экскреции, секреции и инкреции, или внутренней секреции.
Экскреция - процесс выделения ненужных и вредных для организма веществ -
экскретов, образующихся вне пищеварительной системы в процессе обмена веществ.
К числу экскретов относятся растворимые продукты азотного обмена, связанные в
виде мочевой кислоты и ее солей, а также катионы калия, натрия, кальция,
магния, щавелевая и фосфорная кислоты и др. К органам выделения экскретов
относятся мальпигиевы сосуды, нижнегубные железы у первичнобескрылых и клетки
жирового тела.
Инкреция - процесс выделения секретов специальными (эндокринными)
железами, лишенными выводных протоков, непосредственно в кровь. Такие секреты
называют гормонами. Попадая в кровь, гормоны транспортируются ею во все части
тела и регулируют обмен веществ и развитие насекомых. Наиболее развиты 3 пары
желез: нейросекреторные клетки головного мозга, преднегрудные, или
проторакальные железы, и прилежащие тела.
Секреция - процесс выделения веществ - секретов, нужных организму.
Секреты выделяются различными железами, называемыми экзокринными. К ним
относятся слюнные железы и железы средней кишки, выделяющие пищеварительные
ферменты, а также восковые, лаковые, шелкоотделительные железы, выделяющие
вещества механической защиты тела (Бей-Биенко, 1971.).
Большой практический интерес представляют железы, которые выделяют
биологически активные вещества, служащие для химического воздействия на других
животных. Эти вещества Я.Д. Киршенблат (Киршенблат, 1958) назвал телергонами.
Они подразделяются на гетеротелергоны, воздействующие на другие виды животных,
и гомотелергоны, или феромоны, воздействующие на особей своего вида.
К гетеротелергонам относятся вещества химической защиты насекомого от
врагов (яд жалящих перепончатокрылых - пчел, ос), сильнопахнущие, и другие
отпугивающие вещества - репелленты (например, секрет из азотнокислых солей и
окиси азота, выделяемый при опасности жуком бомбардиром, взрывается на воздухе
с шумом и образованием облачка).
К феромонам относятся вещества, определяющие концентрацию насекомых при
использовании источников пищи или поиска мест для спаривания (феромоны
скучивания), выявленные у клопов, прямокрылых, жуков короедов; феромоны
тревоги, или обороны - у жалящих перепончатокрылых, термитов, тлей; феромоны,
контролирующие развитие половых желез - у рабочих пчел; выведение маток - у
пчел и других живущих большими сообществами насекомых (Харборн, 1985).
Выделяют в зависимости от функций несколько групп феромонов, в том числе:
1) половые феромоны; 2) феромоны тревоги; 3) феромоны аггрегации; 4) следовые феромоны;
5) феромоны метки (marking pheromones); 6) полифункциональные феромоны
(Харборн, 1985; Скиркявичюс, 1986, 1988). Указанные группы феромонов
установлены для наземных беспозвоночных, прежде всего насекомых (рис. 1).
Для изучения феромонов и разработки методов их практического
использования большое значение имел прогресс методов их идентификации, который
привел к тому, что для характеристики структуры компонентов феромона в
некоторых случаях достаточно вещества, получаемого всего от нескольких особей насекомых
(Лебедева и др., 1984). Дополнительная информация о методах изучения феромонов
дана в работах В.В. Воронковой (Воронкова, 1987).
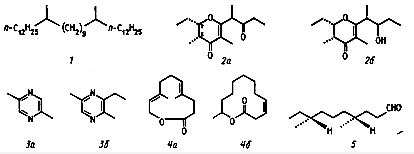
Рис. 1. Некоторые феромоны насекомых (Введение в проблемы биохимической
экологии, 1990)
Наиболее тщательно разработаны методы работы с феромонами чешуекрылых.
Все способы выделения феромонов чешуекрылых подразделяются на три группы:
1) смыв компонентов феромонов с поверхности феромонной железы и вообще кончика
брюшек при коротком контакте растворителя с биологическим материалом, 2)
экстракция при длительном контакте растворителя с биологическим материалом, 3)
сбор из паровой фазы над железой (Воронкова, 1987). В зависимости от метода
выделения феромонов получают разное процентное соотношение компонентов (что
может быть связано с их разной летучестью, т. е. парциальным давлением паров).
Минорные в экстракте компоненты могут увеличивать свои относительные количества
в паровой фазе за счет большей летучести паров. В.В. Воронкова отмечает, что
необходимо исследовать и экстракты желез, и паровую фазу (Воронкова, 1987).
Феромоны тревоги найдены у перепончатокрылых (муравьев, пчел), термитов,
тлей, клопов. Особенно разнообразны феромоны тревоги у общественных насекомых
(Харборн, 1985).
Сигналы тревоги могут вырабатываться разными железами. Например, у
муравьев Acanthomyops claviger пять феромонов тревоги вырабатывают железы
Дюфура, находящиеся в брюшке, а пять других феромонов тревоги - мандибулярные
железы.
Одно и то же вещество может служить феромоном тревоги для разных видов.
Например, гексен-2-аль является активным началом секрета, образуемого при
тревоге красноклопом Dysdercus intermedius (сем. Pyrrhocoridae) и муравьем
Crematogaster africana. Обычно же феромоны тревоги у разных организмов все-таки
отличаются, хотя специфичность этих феромонов ниже, чем специфичность половых
феромонов.
Есть примеры веществ, служащих одновременно и защитными веществами, и
феромонами тревоги. Так, двойную функцию несет ряд метилциклогептанов,
найденных в анальных железах муравьев Acteca, а также муравьиная кислота
муравьев Formica (Харборн, 1985).
Феромоны тревоги, как и другие феромоны насекомых, могут быть
представлены целым букетом веществ. Так, муравьи-портные (Oecophilla longinoda)
реагируют на нападение хищника эмиссией смеси четырех углеводородов, куда
входят: 1-гексаналь, 1-гексанол, 2-бутил-2-октеналь (biting marker),
3-ундеканон (Харборн, 1985).
Термин «феромон тревоги», по-видимому, малоудачен. В ряде случаев более
удачным могло бы быть выражение «феромон возбуждения» или «феромон агрессии».
Весьма условно к «феромонам тревоги» можно относить и «феромоны мобилизации»
(recruitment pheromones), которые выделяются некоторыми муравьями перед атакой
на колонии термитов.
Большинство феромонов тревоги насекомых образуется и выделяется затем из
подчелюстных или анальных желез и из жалящего аппарата. Их образование часто
связано с синтезом защитных веществ. Вступая в «битву», общественные насекомые
выпрыскивают через мандибулы секрет нижнечелюстных желез на врага, который
таким образом «получает ярлык» агрессора. Тревога передается другим членам
общества путем диффузии паров феромонов в воздухе.
Жалящие аппараты пчел и ос содержат несколько желез, которые продуцируют
феромоны тревоги. Сама ядовитая железа часто образует вещества тревоги, которые
выделяются вместе с ядом. Осы рода Vespa впрыскивают яд, содержащий вещества
тревоги, а медоносная пчела при ужаливании оставляет метку в виде
изоамилацетата, указывающую другим пчелам место, которое необходимо жалить.
Большинство феромонов тревоги, идентифицированных у насекомых, имеет
относительно простую структуру (рис. 2). У некоторых муравьев они представлены
простыми углеводородами, такими, как ундекан, тридекан и пентадекан. У других
видов обнаружены альдегидные и кетонные производные этих углеводородов. Запахи
тревоги у видов сем. Formicinae, Hymenoptera, Isoptera и Myrmicinae могут
давать, вероятно, компоненты эфирных масел, в том числе цитронеллол, цитраль, α-пинен, терпинолен и лимонен. У тлей
роль феромонов тревоги выполняют, очевидно, более сложные терпеноиды, такие,
как ненасыщенный моноциклический сесквитерпен гермакрен А. Гермакрен А очень
лабилен, он разрушается вскоре после удаления нападающего хищника. Это создает
для тлей то преимущество, что они могут почти немедленно вернуться к месту
кормления.
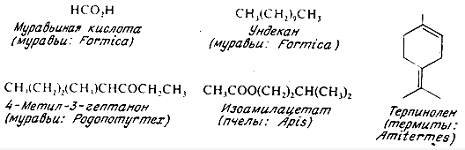
Рис. 2. Структура феромонов тревоги насекомых
Агрегационные феромоны у жесткокрылых не всегда легко отличить от половых
феромонов. Агрегационные феромоны довольно подробно изучены у жуков-короедов и
лубоедов. Чаще всего сигналом к скоплению служит смесь нескольких веществ,
являющихся друг для друга синергистами. Вот некоторые примеры агрегационных
феромонов, представленных сочетанием нескольких веществ (Лебедева и др., 1984):brevicomis
- бревикомин, фронталин, пинокарвон, Е-пинокарвеол, миртенол,
вербенонcalligraphus - ипсдиенол, ипсенол, Е-вербонол. cembrae - ипсдиенол,
ипсенол, 3-Ме-бутен-3-ол-1. amitinus - ипсенол, ипсдиенол, амитинол, (-)-Z-вербенол, (+)-Е-вербенол
Изучение агрегационных феромонов некоторых жуков весьма осложнено тем,
что нет четкой границы между агрегационными феромонами и репеллентами (Лебедева
и др., 1984).
Так, у Dendroctonus adjunctus бревикомин, испускаемый самцами, действует
на самок как агрегационный (привлекающий) феромон, а на других самцов того же
вида как репеллент.
Самцы D. pseudotsugae испускают комбинацию (+)-фронталина и
3-метил-2-циклогексен-1-она (МСН), которая действует на других самцов и самок
этого же вида как репеллент. Однако эти же два вещества испускаются самками
данного вида в сочетании с третьим веществом, (-)-3-метил-циклогексен-1-олом;
эта смесь вместе с камфеном дерева-хозяина действует на других особей D.
pseudotsugae (обоего пола) противоположным образом - как агрегационный феромон.
По-видимому, агрегацию некоторых жуков на кормовых деревьях нельзя
понять, опираясь на традиционную концепцию агрегационных феромонов, поскольку
реально действующим аттрактивным началом является (в экологии жуков некоторых
видов) не просто феромон, а комплекс феромон+летучие вещества растения
(Лебедева и др., 1984). Этот комплекс (агрегационный аттрактант) можно выделить
из воздуха над участком дерева, заселенным питающимися
короедами-первопоселенцами,
Данное обстоятельство (как и родство между феромонами и репеллентами)
представляется важным, поскольку оно еще раз подчеркивает несовершенство
используемого концептуального аппарата и терминологии (в данном случае
экологически неадекватным оказался ранее безупречно служивший термин
«феромон»), что создает дополнительную проблему в понимании экологической сути
явлений.
Следовые феромоны (trail pheromonos) входят в состав пахучих смесей,
которыми насекомые могут метить свой корм и пути, ведущие к нему. По-видимому,
сейчас идентифицированы лишь основные компоненты некоторых из этих смесей. Ниже
приведены некоторые следовые феромоны:
Муравей-листорез Atta texana - метиловый эфир 4-метилпиррол 2-карбоновой
кислоты
Термит Zootermopsis nevadensis - капроновая кислота
Пчела-мелипона Trogona subterranea - 2 изомера цитраля (нераль и
геранцаль)
О высокой эффективности феромонов следа говорит следующее: муравьи Atta
texana распознают свой следовой феромон (метил-4-метилпиррол-2-карбоксилат) в
концентрации 0,08 пг/см, или 3,48·108 молекул/см. Отсюда следует,
что для мечения пути вокруг всего земного шара достаточно 0,33 мг этого
вещества.
2.2 Половые феромоны
Наиболее изучены половые феромоны, или половые аттрактанты. Они
выделяются в ничтожно малых количествах, достаточных, однако для привлечения
другого пола. Так, секреции одной неоплодотворенной самки достаточно для
привлечения десятков тысяч особей самцов того же вида, расположенных от нее на
расстоянии десятков, иногда сотен метров. Некоторые феромоны синтезированы и
уже используются в борьбе с отдельными видами вредных насекомых или для учета
их появления и численности в природе.
Термином «половые феромоны» обозначают соединения, выделяемые самками как
для привлечения самцов, находящихся на некотором расстоянии, так и для
побуждения их к спариванию, если они находятся вблизи. Этот же термин применяют
к соединениям, выделяемым самцами для возбуждения самок. Последние соединения
иногда называют афродизиаками, хотя, строго говоря, этот термин относится к
лекарственным препаратам, усиливающим либидо у человека. Половые феромоны -
наиболее изученная группа химических средств взаимодействия насекомых, и в
настоящее время они найдены и охарактеризованы у множества различных видов.
Только у Lepidoptera обнаружено свыше 200 видов, выделяющих женские феромоны, и
не менее 60 видов, у которых аттрактанты выделяют самцы.
Простейшим по химической структуре половым аттрактантом является
валериановая кислота - женский феромон щелкуна калифорнийского. Однако
большинство феромонов относится к ненасыщенным спиртам, ацетатам или карбоксикислотам
с длинной цепью (табл. 1).
Среди них лучше всего изучена 9-кетодеценовая кислота, выделяемая
пчелиной маткой (царицей) и привлекающая трутней к совокуплению с ней. Однако
это только одно из 32 соединений подобной структуры, обнаруженных в голове матки.
Родственная ей 9-гидроксидеценовая кислота, например, также представляет собой
активное соединение, вызывающее образование и стабилизацию роя рабочих пчел.
В некоторых случаях роль феромонов играют алифатические циклические
соединения, например фронталин и экзобревикомин - феромоны жуков родов
Brevicomis и Ips. Феромоны долгоносика хлопкового (Anthonomus grandis)
составляют производные циклогексана. Существуют также ароматические феромоны.
Фенол служит женским феромоном некоторых личинок жуков; очевидно, он образуется
из тирозина, получаемого с пищей. Феромоном моли Leucania impuris является
бензальдегид. Единственный известный феромон алкалоидной природы образуют самцы
бабочек Danaus в андрокониях крыла.
Таблица 1. Структура некоторых типичных половых феромонов насекомых
|
Структура и название
|
Пол
|
Организм
|
|
СН3(СН2)3СО2Н
Валериановая кислота
|
♀
|
Личинка щелкуна
калифорнийского (Limonius californicus)
|
|
СН3СО(СН2)5CH
= СНСО2Н транс-9-Кето-2-деценовая кислота
|
♀
|
Пчела медоносная (Apis mellifera)
|
|
СН3(СН2)3СН
= СН(СН2)6ОАс цис-7-Додеценилацетат
|
♀
|
Капустная совка (Trichoplusia ni)
|
|
СН3(СН2)2СН
= СН(СН2)7ОАс цис-8-Додеценилацетат
|
♀
|
Восточная плодожорка (Grapholitha molesta)
|
|
СН3СН2СН
= СН(СН2)10ОАс цис-11-Тетрадеценилацетат
транс-11-Тетрадеценилацетат
|
♀
|
Дубовая листовертка (Archips semiferanus)
|
|
СН3(СН2)15ОАс
Гексадеканилацетат СН3(СН2)4СН = СН(СН2)10ОАс
цис-11-октадеценилацетат
|
♂
|
Бабочка (Lycorea ceres ceres)
|
|
СН3(СН2)9СО(СН2)3СН
= СН(СН2)4СН3 цис-6-генэйкозен-11-он
|
♀
|
Волнянка (Orygia pseudotsugata)
|
Феромоны содержатся в насекомых в очень низкой концентрации, и для их
выделения и идентификации требуется огромное число особей. Каждая самка
волнянки псевдотсуговой содержит около 40 нг феромона в последних сегментах
брюшка. Чтобы выделить в достаточном количестве материал для определения,
необходимо иметь не менее 6000 насекомых. Для извлечения 1,5 мг феромона из
хлопковой моли было использовано около 1 млн. неоплодотворенных самок.
У любого насекомого химическая структура преобладающего феромона обычно
очень специфична, и небольшое изменение молекулы может привести к исчезновению
или уменьшению ее активности. Большинство углеводородных феромонов содержит
изолированную двойную связь, положение и стереохимия (цис- или транс-) которой
имеет существенное значение. Это можно проиллюстрировать на примере феромона
капустной совки (Trichoplusia ni). При тестировании ряда его синтетических
аналогов было обнаружено, что все они значительно менее активны, чем природный
феромон, а большинство вообще не обладает активностью (рис. 3) (Джекобсон,
1976).
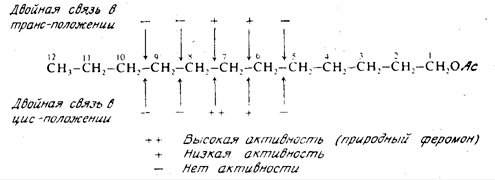
Рис. 3. Влияние положения двойной связи в ацетатах С12-спиртов
на их активность как половых аттрактантов у самцов капустной совки. Ас -
ацетат.
Природный феромон - это цис-7-алкенилацетат. Его синтетический
транс-7-изомер проявляет некоторую активность. По-видимому, существует общая
закономерность: если не меняется положение двойной связи в молекуле, в
определенной степени активны и цис- и транс-формы. У дубовой листовертки
природный феромон самки представляет собой смесь транс- и цис-изомеров в
отношении 2 : 1, что достаточно необычно. Природный феромон волнянки
псевдотсуговой - чистый цис-6-генэйкозен-11-он, однако его синтетическая
транс-форма также активно привлекает самцов этой моли (Харборн, 1985).
Хотя структурные требования для активности половых феромонов сравнительно
жестки, не исключено, что совершенно неродственные соединения оказывают
одинаковое действие. Это доказано на примере таракана американского
(Periplaneta americana), половое возбуждение которого наряду с природным
женским феромоном могут вызывать вещества, присутствующие в экстрактах из
растительных тканей (Харборн, 1985). Одно из таких активных соединений,
найденное у голосеменных, идентифицировано как D-борнилацетат (рис. 4) и
активно в концентрации 0,07 мг/см2.
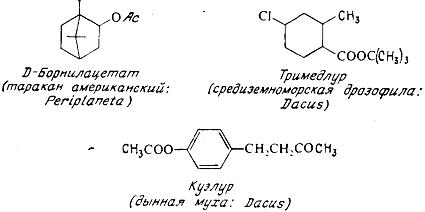
Рис. 4. Структура некоторых соединений, имитирующих половые феромоны
Следует отметить, что L-изомер в 100 раз менее активен, чем D-изомер.
Таким образом, в данном случае стереохимия имеет большое значение. Летучие
соединения, действующие как феромоны на таракана, найдены также у ряда
покрытосеменных; при обследовании 100 видов у восьми из них обнаружены активные
компоненты неизвестной природы. Некоторые синтетические органические соединения
также могут имитировать активность природных феромонов. Два из них нашли
применение. Это тримедлур, активный в отношении средиземноморской дрозофилы, и
куэлур, который привлекает самцов дынной мухи (Dacus cucurbitae). Строение этих
двух соединений изображено на рис. 4. В некоторых случаях при изучении химии
полового поведения были выявлены специфические взаимоотношения между растением
и насекомым, которое на нем живет и размножается. Так, Риддифорд и Вильяме
(Харборн, 1985) обнаружили, что павлиноглазки Полифема (Antherea polyphemus)
могут спариваться только в присутствии листьев красного дуба (Quercus rubra).
Анализ показал, что эманация листьев содержит летучий углеводород
транс-2-гексаналь, попадание которого на рецепторы антенн самок служит триггером
для выделения ими феромонов, которые в свою очередь побуждают самцов к
спариванию. Подобная ситуация, возможно, наблюдается и у насекомых других
видов, например у жуков-долгоносиков Rhabdoscelum obscurus, которые не выделяют
половых аттрактантов, если кормятся не на сахарной свекле (Харборн, 1985); в
этом случае триггерное действие предположительно оказывают неизвестные
растительные соединения, специфичные для сахарной свеклы.
Пути биосинтеза половых феромонов изучены пока еще недостаточно, что связано
с практическими трудностями введения меченых предшественников в железы
насекомых. Однако полагают, что обычно половые феромоны синтезируются в теле
насекомых. Большинство более простых феромонов (табл. 1) - производные жирных
кислот. Они могут синтезироваться непосредственно путем конденсации
соответствующего числа ацетатных единиц или непрямым путем, а из жирных кислот
обычных липидов - в результате декарбоксилирования, укорочения цепей и т. п.
Только в некоторых случаях, например как у бабочек-данаид и короедов, половые
феромоны происходят из пищи. У этих насекомых выделенное из растительного
материала вещество используется или непосредственно, или после соответствующей
химической модификации. Одно время считали, что женские феромоны дубовой
листовертки - смесь тетрадеценилацетатов (табл. 1) - также имеют пищевое
происхождение, но в более поздних работах было показано, что это не так
(Харборн, 1985).
Рассмотрим особенности половых феромонов основных жуков-короедов.
Большой сосновый лубоед - Blastophagus piniperda L. Самки первыми нападают на
дерево-прокормитель - сосну - и вбуравливаются в флоэму коры. В опытах с
ольфактометром самки, извлеченные из коры, а также самцы в брачный период не
привлекали ни самцов, ни самок. Следовательно, в этот период самцы и самки,
извлеченные из флоэмы сразу же после вбуравливания в нее, не оказывают
воздействия друг на друга. Из задней кишки самок и самцов Большого соснового
лубоеда были выделены транс-Вербенол и вербенон.
Короед типограф - Ips typographus L. Свежая древесина
ели, пораженная самцами, привлекает больше жуков обоих полов, чем еще не
заселенная самцами древесина. Феромон выделяется как при весеннем, так и втором
лёте. Феромон короеда типографа привлекает также три вида широко
распространенных хищников (Thanasimus formicarius, Medetera signaticornis и Epuraea pygmaea).
Ренвик и Питман и др. выделили транс-вербенол (33) из эфирного экстракта
задней кишки самок и вербеной из экстракта задней кишки самцов Dendroctonus
frontalis. Как показали Вите и Крозиер, именно вербеной, а не вербенол вызывает
скопление этих насекомых.
В 1968 г. Ренвик и Вите с помощью газовой хроматографии летучих веществ
выделили из пентанового экстракта задней кишки самок соединение, вызывающее при
полевых испытаниях скопления особей обоего пола. Исследователи не смогли
определить строение полученного соединения, но определили методом
масс-спектроскопии его молекулярный вес - 142. Идентификацию этого соединения
успешно осуществили Кинзер и др., которые ранее выделили это соединение из
самцов D. breuicomis (соединение 36). Несмотря на то что названные авторы
выделили его из другого вида и не доказали его идентичности с соединением,
полученным Ренвиком и Вите из D. frontalis, они назвали это соединение
фронталином. Оба соединения (фронталин и транс-вербенол) позднее выделил
методом газовой хроматографии Костер из задней кишки вышедших из древесины
девственных самок D. frontalis. У девственных самок содержание фронталина и
транс-вербенола в 4,1 и 1,9 раза соответственно было выше, чем у жуков, покинувших
древесину. Вите и Ренвик предлагают проводить сравнение результатов газовой
хроматографии с результатами полевых испытаний для выявления активных
соединений, вызывающих агрегацию жуков-короедов D. breuicomis и D- frontalis.
С помощью вакуумного насоса просасывали воздух (20 л/мин) через ящик,
содержащий кусочки бревен желтой сосны, зараженные самцами короедами Ips
confusus. После этого воздух пропускали через 2 охлажденные ловушки (в первой -
4°С и во второй - 70°С).
В опытах с полевым ольфактометром оттаявшее содержимое обеих ловушек
обладало сильным привлекающим действием. Привлекающее соединение экстрагировали
из его водной эмульсии петролейным эфиром и концентрировали, отгоняя
растворитель при 80°С. Полученное бесцветное масло при хранении в холодильнике
не теряло активности в течение нескольких недель. Однако оно теряло активность
при продолжительном нагревании выше 85°С.
Ренвик и др. пропускали через газовый хроматограф воздух из сосуда,
содержащего заднюю кишку самцов, питавшихся свежей флоэмой желтой сосны.
Соединения, вышедшие из колонки (1525 см ´ 0,313 см) с 5%-ным апиезоном на хромосорбе W, промытом
кислотой, с временем удерживания 6 мин при 110° (скорость тока азота 25 мл/мин)
сильно привлекали мигрирующих жуков.
В 1966 г. Вуд и др. описали методы получения больших количеств феромона
из буровой муки, образуемой самцами в килограммовых количествах. Было
обнаружено, что отдельные самцы выбрасывают по 9,4 мг буровой муки в день в
течение примерно 15 дней. Для выделения аттрактанта буровую муку замачивали и
растирали в теплом бензоле, затем отгоняли растворитель и перегоняли
концентрированный остаток в специальном приборе при температуре 85 - 90°С и
давлении 0,01 мм. Дистиллят хроматографировали на колонке с силикагелем и
активную фракцию (активность проявлялась в количестве 10-6 г)
элюировали смесью бензол - эфир. Экстракт разделяли на фракции с помощью
препаративной газовой хроматографии на колонке (91,5 ´ 0,9375 см) с 8%-ным SE 30 на
хромосорбе G с запрограммированной температурой
от 80 до 200°С (скорость тока 45 мл/мин). Активная фракция (0,5 г из 2,5 кг
буровой муки) выходила через 14-23 мин между 2 другими пиками, которые были
идентифицированы как нонанал- и геранилацетаты. При дальнейшей хроматографии на
полярной колонке (8%-ный карбовакс 20М) на хромосорбе G при 120°С получили 14
мг вещества, активного в концентрации 5·10-5 г. Вещество было
выделено в виде частично разделившихся минорных пиков. Три компонента,
обусловивших совместно привлекающую активность, были идентифицированы
сочетанием методом ИК-, масс- и ЯМР-спектроскопии как
(-)-метил-6-метилен-7-октен-4-ол (38), циc-вербенол (39) и
(+)-2-метил-6-метилен-2,7-октадиен-4-ол (40) (рис. 5). Методы выделения и
идентификации этих соединений изложены в обзоре Сильверстайна и др.
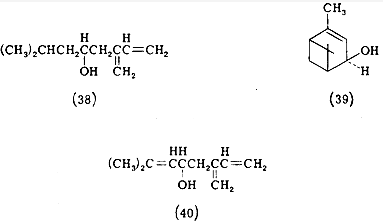
Рис. 5. Выделенные феромоны Ips confusus
В 1966 г. с помощью газовой хроматографии Питман и др. выделили из задней
кишки, мальпигиевых сосудов и экскрементов самцов, питавшихся в течение двух
дней в небольших бревнах желтой сосны, неидентифицированную фракцию, которая
вызывала ответную реакцию у ползающих жуков. Хотя авторы полагали, что
выделяемый ими материал содержал лишь единственный феромон, в действительности
это была смесь веществ (Джекобсон, 1976).
Вите считает, что по привлечению жуков к приманке нельзя судить об
активности препарата на разных стадиях его выделения, как это делали Вуд и др.
Однако последние показали, что хемоклинотаксис жуков в лабораторном
аль-фактометре вполне равнозначен реакции полета насекомых на половой феромон.
Простой полевой метод, в котором струю воздуха пропускали через
контейнеры, содержащие куски древесины, зараженной короедом Ips confusus, показал, что к аттрактанту
чувствительны особи обоего пола. Широкие алюминиевые цилиндры укрепляли вертикально
и горизонтально, и в них помещали зараженные, заведомо аттрактивные куски
древесины. Воздух продували с помощью вентилятора над зараженными поленьями и
направляли его либо на бревно или дерево, чтобы вызвать нападение на них
вредителей, либо на дощечку или стекло для сбора привлеченных насекомых.
Модификацию этого метода использовали для получения большого количества
аттрактивного материала. Насекомых ловили во вращающиеся сетчатые или
чашевидные ловушки, в которые помещали аттрактант. Такая вращающаяся сетчатая
ловушка с аттрактантом I.
confusus выловила через 7,5 ч 39 I.
confusus, 35 I. latidens+I. guildi, 10 Dendroctonus
valens, 72 других видов жука-короеда и 25 хищников жуков-короедов.
Как показали полевые испытания, привлечение насекомых в природе подчинено
определенному суточному ритму с максимумами от 8 до 10 ч утра и с 2 до 6 ч
вечера. Жуки предпочитали места, защищенные от ветра; что же касается
освещения, то оно в меньшей степени влияло на ответную реакцию, хотя прямого
солнечного света насекомые избегали. С подветренной стороны ответная реакция
жуков проявлялась по ольфакторному градиенту. При освоении нового лесного
массива колония насекомых может перемещаться под влиянием аттрактанта на
расстояния порядка 500 - 1000 м.
Три соединения, входящие в состав феромона, были испытаны в безлесных
кустарниковых зонах в 1 км от природной зараженной сосновой (Pinus ponderosa)
вырубки в Калифорнии. В этой зоне преобладали ветры, направленные из долины к
месту заражения. Использовали ловушки цилиндрической формы из жесткой ткани,
покрытой клеем, которые были установлены на палках высотой 1,5 м. Воздух (50 см3/мин)
пропускали через сосуд, содержащий аттрактант, и затем через ловушку. Как смесь
1,5 мг соединения (38) с 1 мг соединения (39) и с 0,5 мг соединения (40), так и
кусок древесины, зараженный 20 самцами, сильно привлекали жуков. Экстракт из 2
г буровой муки самцов привлекал жуков в меньшей степени. Растворы этих трех
соединений в гексане или гептане, нанесенные на ватные или бумажные тампоны,
вызывали значительно более слабую ответную реакцию. Самцы и самки 2 хищников Enoclerus lecontei и Temnochila virescens var. chlorodia, а также Tomicobia tibialis, паразитирующие на Ips, реагировали на синтетический
аттрактант. Бедард и Браун описали портативную переносную систему с испарителем
и ловушкой для оценки в природе эффективности химических аттрактантов
насекомых, особенно для I. confusus.
Райе испытал в полевых ольфактометрах и клейких ловушках (Tanglefoot) большое количество монотерпенов,
обнаруженных в растении-хозяине, пораженном I. confusus. α-Пинен и β-пинен были высокоаттрактивными для
хищников (Е. lecontei и Т. virescens var. chlorodia); 3-карен, мирцен и лимонен были менее аттрактивны. Гептан был очень
аттрактивен для Temnochila virescens, но не для Е. lecontei.
2.3 Использование половых феромонов для уничтожения
вредителей
В практике лесозащиты аттрактанты могут применяться самостоятельно для
надзора за вредителями и борьбы с ними или совместно с инсектицидами и другими
средствами. Совместное применение пищевых аттрактантов с инсектицидами повышает
эффективность химической борьбы, уменьшая расход инсектицидов и опасность
загрязнения среды. При помощи аттрактантов возможно также привлечение на
определенные места (отдельные участки леса или деревья) вредных насекомых с
последующим уничтожением их различными способами и средствами (Справочник по
защите леса от вредителей и болезней, 1980).
В защите растений применяются синтетические аттрактанты. Для многих видов
насекомых выяснена химическая структура половых аттрактантов, синтез же освоен
только для некоторых из них, а способы применения находятся в стадии
разработки. В решении этих вопросов, имеются значительные трудности: Для
выделения в чистом виде и расшифровки химической структуры аттрактантов требуется
огромное количество насекомых или продуктов их жизнедеятельности, собрать
которые не всегда возможно. Большинство половых аттрактантов - это сложные
соединения, часто являющиеся смесью стереоизомеров, биологическая активность
которых различна, или комбинацией нескольких химических веществ. Нередко
главный компонент содержит вторичные примеси и регуляторы летучести,
усиливающие действие аттрактанта. Часто при начальном синтезе аттрактант
содержит сопутствующие вещества, маскирующие или задерживающие его действие.
Эти примеси могут блокировать способность обонятельных рецепторов насекомых
воспринимать запаховые сигналы. Необходимость синтеза химически чистых веществ
значительно удорожает производство аттрактантов (Скиркявичюс, 1988).
Сложна разработка способов практического применения аттрактантов.
Недостаточно изучена биоэкология и реакции поведения взрослых насекомых и
динамика их популяций. Не ясны вопросы и технологии внесения аттрактантов в
насаждения, характер их распространения и воздействия на насекомых.
Среди половых аттрактантов вредных лесных насекомых в России наиболее
изучен половой аттрактант непарного шелкопряда и осуществлен синтез этого
вещества (Сегаль, 1983).
Половой аттрактант непарного шелкопряда, продуцируемый самками,
обеспечивает их встречу с самцами. Формируется он в специальных железах,
расположенных между последним и предпоследним сегментами брюшка самок. Железы
могут выдвигаться наружу и втягиваться внутрь, регулируя таким образом
выделение в атмосферу привлекающего вещества. Вещество распространяется путем
диффузии и под воздействием потоков воздуха. Самцы реагируют на появление в
воздухе ничтожно малого количества привлекающего вещества (нескольких десятков
молекул) и могут прилетать к самкам, удаленным на расстояние до 4 км.
Девственные самки непарного шелкопряда, заключенные в садочки, или
бензольные экстракты из их половых желез, нанесенные на кусочки фильтровальной
бумаги, помещенные на покрытые клеем картонные или из другого материала листы
или в специальные ловушки, применялись в ряде стран для обнаружения и надзора
за непарным шелкопрядом. Однако использование живых самок и их экстрактов в
настоящее время нецелесообразно в связи с получением синтетических аттрактантов
(Мыттус, 1983).
В результате 30-летних работ американский ученый М. Джекобсон в 1960 г.
выделил из 500 тыс. самок непарного шелкопряда 20 мг феромона, который назвал
гиптолом. Однако при более тщательном исследовании было установлено, что
вещество с таким названием не является половым феромоном. Оно присутствует в
выделениях феромонных желез самок, но само по себе не привлекает самцов.
Истинный половой феромон непарного шелкопряда был установлен спустя 10 лет. Им
оказался цис-7,8-эпокси-2-метилоктадекан, названный диспарлюром. Количество
диспарлюра, вырабатываемое одной самкой непарного шелкопряда, приблизительно
равно 0,1 мкг. Этого количества достаточно, чтобы привлечь около 1 млн самцов.
Он распространяется в воздухе путем диффузии согласно законам физики. Это
соединение при лабораторных и полевых испытаниях обнаружило исключительно
высокую биологическую активность и было определено в качестве основного
полового аттрактанта непарного шелкопряда (Джекобсон, 1976).
В Советском Союзе высокоактивные образцы диспарлюра были получены рядом
лабораторий. Разрабатывается технология промышленного производства этого
аттрактанта и его препаративных форм. Диспарлюр характеризуется высокой
устойчивостью и длительностью действия в чрезвычайно малом количестве. В
природных условиях он активен в количестве 10-9 г, или 0,001 мкг на
ловушку. Наиболее эффективные дозировки составляют обычно не более 5 мкг.
Биологическая активность диспарлюра сохраняется в течение всего периода
лета бабочек. Продолжительность действия диспарлюра, а также его дозировка
зависят: от типа ловушек, метеорологических и других условий, влияющих на
скорость испарения и рассеивания диспарлюра.
Диспарлюр помимо самцов непарного шелкопряда привлекает также самцов
монашенки. Он может использоваться в лесозащите для выявления насаждений,
заселенных непарным шелкопрядом и монашенкой, и для надзора за ними. С этой
целью применяют специальные ловушки различных типов, содержащие аттрактант и
ловчую клеевую поверхность. По количеству бабочек, привлеченных в ловушки,
оценивается численность вредителей. При помощи ловушек можно своевременно
обнаруживать начало массового размножения непарного шелкопряда и монашенки.
Высокая биологическая активность диспарлюра открывает возможности его
использования не только для надзора, но и для борьбы с непарным шелкопрядом и
монашенкой. Применение диспарлюра в качестве средства борьбы представляется
перспективным в популяциях с низкой численностью вредителей. Предусматриваются
два возможных способа его применения:
. Использование ловушек с диспарлюром для вылавливания и уничтожения
самцов. Самцы при отыскании самок будут улавливаться ловушками. Чтобы свести к
минимуму встречи самцов с самками в природе соотношение между количеством
устанавливаемых ловушек и самок должно быть достаточно высоким.
. Свободный выпуск концентрированного феромона (диспарлюра) в насаждения
с целью нарушения ориентировки самцов при поисках самок - так называемый метод
дезориентации. Предполагается, что при насыщении воздуха в насаждении запахом
синтетического диспарлюра будут нарушены нормальные реакции самцов при попытках
обнаружить истинный запах полового феромона, выделяемого самками. Самцы не
смогут отыскать самок, что приведет к отсутствию или уменьшению спаривания и
снижению численности популяции вредителя (Введение в проблемы биохимической
экологии, 1990).
Оба метода находятся в стадии разработки.
Привлечение самцов на аттрактант, выделяемый самками, присуще другим
видам бабочек (волнянкам, коконопрядам, листоверткам, пяденицам и т. д.) и
пилильщикам. Ведутся работы по изысканию и созданию синтетических аттрактантов
для этих групп лесных насекомых.
У других групп насекомых, важнейших вредителей леса, наиболее изучены и
своеобразны феромоны (аттрактанты) короедов. Различают аттрактанты первичного и
вторичного привлечения. Первичные выделяются ослабленными и срубленными
деревьями. У хвойных пород к их числу относятся монотерпеновые компоненты
живицы α- и β-пинены, лимонен, камфен и другие
соединения. У лиственных пород из привлекающих веществ известны ванилин,
сиреневый альдегид и ряд других, входящих в состав продуктов окисления лигнина.
Эти вещества являются привлекающими только в слабых концентрациях и как бы
сигнализируют насекомым о снижении устойчивости Деревьев и пригодности для
поселения. Эти же вещества при высокой концентрации, свойственной жизнеспособным
деревьям, оказывают на короедов отпугивающее действие (Исмаилов и др., 1986).
Привлечение запахом, выделяемым ослабленными деревьями, - начальный этап
при выборе короедами объектов поселения.
Более сильными привлекающими свойствами характеризуются аттрактанты
вторичного привлечения, которым принадлежит решающая роль при заселении
деревьев короедами. Эти аттрактанты продуцируются самими насекомыми -
жуками-первопоселенцами: у моногамных короедов самками, у полигамных - самцами,
но в том и другом случае привлекают особей обоего пола. Аттрактант
секретируется специальными железами задней кишки, других отделов кишечника и
мальпигиевых сосудов и выделяется наружу в составе экскрементов и буровой муки.
Аттрактант начинает продуцироваться после прохождения пищи через кишечник, так
как секреторная активность желез стимулируется под действием пищевых
метаболитов.
Ловушки с аттрактантами первичного и вторичного привлечения можно
использовать для выявления присутствия короедов в лесах, а также для массового
вылова жуков с целью предотвращения заселения ими деревьев. Перспективно
применение аттрактантов совместно с инсектицидами на ловчих деревьях. Возможны
и другие способы практического использования аттрактантов.
В результате работ, проведенных в США по изучению феромонов короедов р.
Dendroctonus, были выделены и затем изготовлены промышленными фирмами следующие
феромонные препараты: транс-вербенол, фронталин, бревикомин.
Эти препараты используют в сочетании, так как бревикомин выделяют самки
жуков, а фронталин - самцы. Короеды р. Ips. продуцируют инсдиенол и ипсенол
(Харборн, 1985).
На основании этих данных в Норвегии А. Бакке разработал смесь для борьбы
с типографом. Она была изготовлена фирмой «Целамерк» (ФРГ). Эта смесь состоит
из ипсдиенола, цис-вербенола и диметилвенилкарбенола в соотношении 1 : 1 : 10.
Использовались пластинки, пропитанные феромонной смесью. Их прикрепляли к
ловчим деревьям.
В СССР группой исследователей институтов системы Госкомлеса СССР также
был разработан феромон короеда-типографа. Он является агрегационным и
привлекает летающих жуков обоих полов, а также других видов р. Ips. Феромон
содержит много компонентов, выполняющих различные функции в жизни жуков:
ипсдиенол, ипсенол, цис-вербенол, транс-вербенол и др. К сожалению, в ловушки
попадают не только короеды, но и их хищники (Справочник по защите леса от
вредителей и болезней, 1980).
Лесным ведомством США осуществлялась большая программа по вылову
струйчатого заболонника, который распространяет голландскую болезнь ильмовых
пород. Для этого использовался феромонный препарат мултилур (Харборн, 1985).
Разработку и испытание новых аттрактантов проводят в нашей стране
ученые-химики и биологи. Они разрабатывают не только сами препараты, но и
способы их применения, изучают наилучшие формы ловушек, методы их размещения в
насаждениях, находят связь между улавливающей способностью ловушек и уровнем
численности вида насекомого в насаждениях.
Применяют клеевые и клеево-инсектицидные ловушки для бабочек,
изготовляемые из картона, пластмассы, гофрированного пластик-картона. Для жуков
используют полиэтиленовые и металлические, пластмассовые и пластиковые ловушки,
а также отрубки деревьев или специальные ловчие деревья, на которые прикрепляют
привлекающее вещество, нанесенное на пористый субстрат (диспенсер). Диспенсер
помещают в полиэтиленовый пакетик или пробирку с проницаемой пробкой, из
которых аттрактант постепенно распространяется в воздух. Прилетающие насекомые
вылавливаются и погибают, попадая на клей, водную поверхность или соприкасаясь
с инсектицидом. Большое значение имеют форма ловушек, место расположения,
защищенность от воздействия погодных факторов, удобство осмотра и очищения от
насекомых, возможность многократного использования. Ловчие деревья, снабженные
диспенсерами с феромонами короедов, заселяются ими и впоследствии окоряются или
уничтожаются и утилизируются. Использование этого метода против стволовых
вредителей перспективно.
Половые феромоны используются для борьбы с вредителями в США. При этом
применяют два подхода. Первый состоит в установке ловушек для самцов путем
размещения некоторого количества феромона самок в районе, где необходимо
провести борьбу с данным видом. Это препятствует ориентации самцов на природный
феромон, выделяемый самками, благодаря чему не происходит спаривания. Количество
феромона, необходимого для ловушки, может быть совсем небольшим. Полевые
испытания показали, что для нарушения ориентации самцов на выделения самок
капустной совки достаточно 17 мг феромона, размещенного в 100 точках
пространства объемом 27 м3. Второй подход заключается в
распространении менее специфичного летучего вещества, которое маскирует эффект
феромона, мешая восприятию сигнала (Харборн, 1985).
Хотя теоретически с помощью ловушек насекомое-вредитель может быть
полностью уничтожено, на практике множество факторов ограничивают эффективность
половой приманки. Наиболее широко, синтетические аттрактанты используют для
выявления (путем вылавливания насекомых) зоны заражения и размера популяции
данных насекомых. Эта информация может быть затем использована для определения
необходимых мер борьбы, таких, как опрыскивание зоны соответствующим
пестицидом.
По крайней мере в двух случаях полевые опыты подтвердили пригодность
феромонов непосредственно для борьбы с вредителями. Так, Гастон и др. успешно
боролись с хлопковой молью (Pectinophora gassypiella), личинки которой пожирают
хлопчатник, с помощью синтетического госсиплура (Харборн, 1985). Это вещество
нарушает нормальную связь между взрослыми бабочками через феромоны. Стоимость
процедуры, которая состоит в контролируемом распространении из специальных
контейнеров смеси феромонов в гексане, сопоставима со стоимостью применения
соответствующих инсектицидов. Другое синтетическое соединение - мультилур -
помещали в клейкие ловушки и использовали для борьбы с ильмовым короедом
Scolytus multisfriatus в восточных штатах США (Харборн, 1985). Массовый лов
жуков в ловушки, однако, должен сопровождаться такой обработкой пораженных
листьев, чтобы насекомые, которые избежали ловушек, не имели больше источника
пищи. Разрушительное действие короедов на деревья вяза определяется главным
образом тем, что они открывают дорогу для цератоцистиса вязового (Ceratocysfis
ulmae).
3. Материалы и методы исследования
.1 Описание проводимых исследований
Полевая производственная практика проходила в Зоркальцевском кедровнике
Тимерчинского лесничества Тимирязевского лесхоза, исследования проводились с 24
мая по 7 августа 2004 года.
Темой исследований является: «Использование феромонных ловушек для
уничтожения короеда стенографа».
Цель и задачи исследования: изучение эффективности различных видов
феромонов. Для этого ставились следующие задачи:
1. Проведение учета численности.
2. Сбор материала.
. Определение эффективности феромонов.
В данной полевой производственной практике объектом исследования был
короед стенограф. За время практики, было совершенно пять выездов в
Зоркальцевский кедровник. Первый выезд был 24 мая. На площади размером 375 га
было установлено 60 феромонных ловушек. Ловушки устанавливались на четырёх
участках.
Ловушки были расставлены на расстоянии 50 - 100 м. друг от друга.
Контроль над ловушками осуществлялся 1 раз в 10 дней. Во время второго выезда
(3 июня) из 31 ловушки только в одной был обнаружен один короед стенограф. В
течение всего обследования в ловушках были обнаружены такие виды насекомых, как
щелкуны, усачи, муравьи, злотки и др.
Нам удалось собрать некоторый материал, не смотря на то, что большинство
ловушек было испорчены в результате сырой дождливой погоды в июле месяце.
3.2 Описание феромонной ловушки
Феромонные ловушки предназначены для мониторинга короеда стенографа и
борьбы с ним методом массового отлова. В основе ловушки усечённая воронка
диаметром 16-30 см. с прилепленным к ней сборником для жуков. Сборник снабжён
отверстиями для отвода дождевой воды. Над воронкой установлен крестообразный
барьер высотой в 1,5 - 1,8 раза превышающий ширину воронки (см. рис. 6).
Комплектация одной ловушки:
. Верхняя часть ловушки - два согнутых по длине прямоугольника из
пластика, скрепленные вверху веревкой для подвешивания ловушки, а внизу -
скрепкой для подвешивания диспенсера - 1 шт.
. Воронка - 1 шт.
. Короткая Г-образная проволока - 1 шт.
. Стакан - сборник жуков - 1 шт.
. Скрепки для прикрепления воронки к верхней части ловушки - 4 шт.
Рис. 6. Феромонная ловушка.
Внутри ловушки крепиться диспансер, представляющий собой препаративную
форму синтетического агрегационного феромона короеда.
Диспенсер с феромоном подвешивается на скрепку, соединяющую внизу верхнюю
часть ловушки и, таким образом, он находится ниже верхнего края воронки.
Готовую ловушку (с раскрытым и прикрепленным к ней диспенсером) подвешивают с
помощью веревки в нужное место.
Инструкция по применению феромонного диспенсера для короеда-стенографа.
Диспенсер раскрывается только перед помещением в ловушку или на ловчее дерево.
Диспенсеры легко отрываются от общей ленты. В верхней части диспенсера
(там, где находится отверстие для закрепления его в ловушке), пленка запаяна не
до конца. Чтобы раскрыть диспенсер, необходимо потянуть за его недопаянные
части в разные стороны (см. рис. 7).
Рис. 7. Феромонный диспенсер для короеда стенографа.
При этом диспенсер раскрывается пополам, и в ловушку вывешивается только
та половина (рабочая), в которую запаян картонный прямоугольник (!) и на
которой написано "Вертенол". Эта рабочая половина диспенсера
прикрепляется к нижнему перекрестью ловушки с помощью скрепки.
4. Испытание феромонных ловушек для борьбы с короедом
стенографом в Томской области
.1 Феромонный препарат для борьбы с шестизубым короедом
(стенографом) Ips sexdentatus
Лабораторией исследования феромонов ВНИИ Химических Средств Защиты
Растений разработан, зарегистрирован и успешно применялся в 2001 - 2003 гг.
феромонный препарат «Вертенол» для подавления вспышки массового размножения
короеда-типографа Ips typographies, вредителя ели в Подмосковье. Также был
разработан и зарегистрирован феромонный препарат «Деналол» для сибирского
шелкопряда Dendrolimus supercms sibiricus, мониторинг которого успешно
проводится в течение 3-х лет в пяти регионах Сибири и Дальнего Востока.
Создание феромонного препарата включает в себя несколько этапов:
выделение феромона из насекомого (сбор из феромонного облачка или
экстракция кончиков брюшек);
идентификация веществ, выделенных из насекомого методами ГЖХ, ГЖХ-МС;
испытания в лаборатории методом ЭАГ имеющихся веществ;
синтез компонентов феромона;
испытания препарата в полевых условиях.
Характеристика феромонного препарата для борьбы со стенографом
Назначение. Для надзора и борьбы с короедом стенографом (Ips
Sexdentatus), вредителем хвойных пород.
Способ применения. Диспенсер помещается в барьерную ловушку или
прикрепляется на ловчее дерево.
Безопасность. Безопасен для полезных насекомых и абсолютно безвреден для
окружающей среды.
Препаративная форма. Специально разработанный диспенсер, содержащий
агрегационный феромон короеда стенографа.
Достоинства препарата:
специфичность (привлекает только короеда стенографа);
малые нормы расхода;
природное происхождение действующих веществ;
безвредность для окружающей среды.
4.2 Результаты исследования
Испытания феромонной ловушки на протяжении времени показало, что
количество насекомых, попавшихся в эти ловушки довольно большое (см. таблицу
2). При этом помимо вредителей леса присутствует некоторое количество
насекомых, попавших в эти ловушки случайно (боярышница, коровка, шмель и др.).
Возможно, часть из них была привлечена погибшими насекомыми и скопившейся в
ловушках дождевой водой.
Таблица 2. Количество насекомых, попавших в ловушку
|
№ учета
|
Виды насекомых
|
Общее количество
|
Среднее число на 1 ловушку
|
|
1выезд
|
боярышница (Aporia crataedi)
|
7
|
0,116
|
|
муравей (Formica polyctena Foerst.)
|
17
|
0,283
|
|
толстоголовая оса большая (Crabro cribrarius)
|
3
|
0,05
|
|
могильщик погребальный (Necrophorus verpillo)
|
8
|
0,133
|
|
рагий-сыщик (Rhagium unduisitor)
|
11
|
0,183
|
|
щелкун (Slateridae)
|
13
|
0,216
|
|
клоп итальянский (Graphosoma lineatum)
|
6
|
0,1
|
|
щитоноска (Coccus hesperidum L.)
|
1
|
0,016
|
|
щелкун красноногий (Melanotus rufipes)
|
10
|
0,166
|
|
щелкун блестящий (Selatosomis aeneus)
|
4
|
0,066
|
|
шмель лесной (Botbus silvasum)
|
6
|
0,1
|
|
грибоед красноголовый (Tripeax rissica)
|
11
|
0,183
|
|
долгоносик полосатый (Sitona eineatus)
|
2
|
0,033
|
|
коровка (Сoccineceidae)
|
1
|
0,016
|
|
синий рогохвост (Paururus juvencus)
|
11
|
0,183
|
|
черный сосновый усач (Monochamus galloprovincialis oliv)
|
2
|
0,033
|
|
сосновая тля (Lachnus pineus Mordk),
|
12
|
0,2
|
|
щитник двузубчатый (Picromesus bidens),
|
1
|
0,016
|
|
сибирский гравёр (Melolonthe hippocasteni Fabr)
|
0,033
|
|
смолевка сосновая (Pissodes pini)
|
2
|
0,033
|
|
короед стенограф (Ips sexdentatus)
|
1
|
0,016
|
|
2выезд
|
боярышница (Aporia crataedi)
|
9
|
0,115
|
|
муравей (Formica polyctena Foerst.)
|
6
|
0,1
|
|
толстоголовая оса большая (Crabro cribrarius)
|
2
|
0,033
|
|
могильщик погребальный (Necrophorus verpillo)
|
8
|
0,133
|
|
рагий-сыщик (Rhagium unduisitor)
|
6
|
0,1
|
|
щелкун (Slateridae)
|
4
|
0,066
|
|
щелкун красноногий (Melanotus rufipes)
|
8
|
0,133
|
|
щелкун блестящий (Selatosomis aeneus)
|
3
|
0,05
|
|
грибоед красноголовый (Tripeax rissica)
|
8
|
0,133
|
|
долгоносик полосатый (Sitona eineatus)
|
2
|
0,033
|
|
синий рогохвост (Paururus juvencus)
|
5
|
0,083
|
|
черный сосновый усач (Monochamus galloprovincialis oliv)
|
2
|
0,033
|
|
сосновая тля (Lachnus pineus Mordk),
|
14
|
0,23
|
|
сибирский гравёр (Melolonthe hippocasteni Fabr)
|
5
|
0,083
|
|
комлевый бурый усач (Criocephelus rusticus L)
|
3
|
0,05
|
|
большой сосновый долгоносик
(Hylobis abietis).
|
8
|
0,133
|
|
щитник черношипный (Carpocoris fuscispinus)
|
1
|
0,016
|
|
большой хвойный рогохвост (uroceras gigas)
|
1
|
0,016
|
|
3выезд
|
боярышница (Aporia crataedi)
|
1
|
0,033
|
|
могильщик погребальный (Necrophorus verpillo)
|
3
|
0,1
|
|
рагий-сыщик (Rhagium unduisitor)
|
6
|
0,203
|
|
щелкун (Slateridae)
|
4
|
0,133
|
|
щелкун красноногий (Melanotus rufipes)
|
5
|
0,166
|
|
щелкун блестящий (Selatosomis aeneus)
|
1
|
0,033
|
|
грибоед красноголовый (Tripeax rissica)
|
1
|
0,033
|
|
долгоносик полосатый (Sitona eineatus)
|
2
|
0,066
|
|
синий рогохвост (Paururus juvencus)
|
4
|
0,133
|
|
черный сосновый усач (Monochamus galloprovincialis oliv)
|
2
|
0,066
|
|
сосновая тля (Lachnus pineus Mordk),
|
3
|
0,1
|
|
бронзовка золотистая (Agrilus viridis)
|
26
|
0,866
|
|
усач-инквизитор (Rhagium inquisitor)
|
1
|
0,033
|
|
черный длинноусый усач (Acenthocinus aedilis L).
|
2
|
0,066
|
|
4выезд
|
усач-инквизитор (Rhagium inquisitor)
|
4
|
0,133
|
|
муравей (Formica polyctena Foerst.)
|
1
|
0,033
|
|
толстоголовая оса большая (Crabro cribrarius)
|
3
|
0,1
|
|
могильщик погребальный (Necrophorus verpillo)
|
5
|
0,167
|
|
рагий-сыщик (Rhagium unduisitor)
|
2
|
0,066
|
|
щелкун (Slateridae)
|
2
|
0,066
|
|
щелкун красноногий (Melanotus rufipes)
|
2
|
0,066
|
|
щелкун блестящий (Selatosomis aeneus)
|
1
|
0,033
|
|
грибоед красноголовый (Tripeax rissica)
|
4
|
0,133
|
|
долгоносик полосатый (Sitona eineatus)
|
1
|
0,033
|
|
синий рогохвост (Paururus juvencus)
|
5
|
0,167
|
|
черный сосновый усач (Monochamus galloprovincialis oliv)
|
2
|
0,066
|
|
сосновая тля (Lachnus pineus Mordk),
|
8
|
0,266
|
|
сибирский гравёр (Melolonthe hippocasteni Fabr)
|
3
|
0,1
|
|
большой сосновый долгоносик
(Hylobis abietis).
|
2
|
0,066
|
|
щитник черношипный (Carpocoris fuscispinus)
|
1
|
0,033
|
|
большой хвойный рогохвост (uroceras gigas)
|
1
|
0,033
|
|
5выезд
|
усач-инквизитор (Rhagium inquisitor)
|
2
|
0,066
|
|
боярышница (Aporia crataedi)
|
1
|
0,033
|
|
толстоголовая оса большая (Crabro cribrarius)
|
1
|
0,033
|
|
щелкун (Slateridae)
|
7
|
0,233
|
|
щелкун красноногий (Melanotus rufipes)
|
2
|
0,066
|
|
щелкун блестящий (Selatosomis aeneus)
|
3
|
0,01
|
|
грибоед красноголовый (Tripeax rissica)
|
3
|
0,01
|
|
долгоносик полосатый (Sitona eineatus)
|
2
|
0,066
|
|
синий рогохвост (Paururus juvencus)
|
6
|
0,2
|
|
черный сосновый усач (Monochamus galloprovincialis oliv)
|
2
|
0,066
|
|
сосновая тля (Lachnus pineus Mordk),
|
7
|
0,233
|
|
сибирский гравёр (Melolonthe hippocasteni Fabr)
|
1
|
0,033
|
|
комлевый бурый усач (Criocephelus rusticus L)
|
2
|
0,066
|
|
большой сосновый долгоносик
(Hylobis abietis).
|
2
|
0,066
|
|
щитник черношипный (Carpocoris fuscispinus)
|
1
|
0,033
|
|
смолевка сосновая (Pissodes pini)
|
2
|
0,066
|
|
щитник черношипный (Carpocoris fuscispinus)
|
2
|
0,066
|
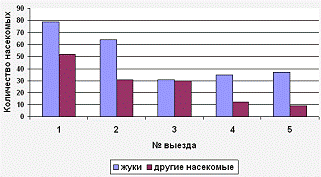
Рис. 8. Соотношение жуков и других насекомых в разные выезды
Хотя количество короедов оказалось незначительным, в целом жуки являются
преобладающей группой на протяжении всего времени учета (Рис. 8).
Из гистограммы (Рис. 8) видно, что во все дни нашего выезда количество
жуков, попавших в ловушку, всегда превышало количество других насекомых
(муравьев, шмелей, ос, бабочек). При этом только в третий учет это соотношение
почти сравнялось, но зато во второй учет количество жуков в два раза превысило
количество попавших других насекомых, в четвертый выезд - в 2,5 раза, а в пятый
выезд более чем в три раза.
Все это подтверждает, что данный феромонный препарат является эффективным
для борьбы с жуками вредителями. Несмотря на то, что в установленные ловушки
попал всего один короед-стенограф, в них попадали другие жуки-вредители. Так,
из таблицы 1 видно, что очень большое количество в ловушки попало таких
жуков-вредителей как рагий-сыщик (Rhagium unduisitor), могильщик погребальный
(Necrophorus verpillo), щелкун красноногий (Melanotus rufipes), грибоед красноголовый (Tripeax rissica).
В ловушку также попало большое количество бронзовок золотистых и
сибирской сосновой тли.
Все это говорит о том, что данный препарат может быть использован не для
борьбы с короедом стенографом, а с другими видами жуков стволовых вредителей.
Причин, почему в ловушки не попали именно короеды стенографы может быть
множество. Это может быть связанно и с климатическими отличиями между
Подмосковным и Сибирским регионом, и с особенностью погоды во время прохождения
практики (Большую часть практики стояла дождливая погода, что могло повлиять на
результаты исследования). Так климат Подмосковья является менее влажным, чем
сибирский. Другой причиной может являться нарушение технологии получения
феромона.
Заключение
Феромоны - это биологически активные вещества, выделяемые насекомыми в
окружающую среду для коммуникации между особями своего вида и вызывающие их
поведенческие или физиологические реакции. Половые и агрегационные феромоны
широко применяются в странах с развитым сельским и лесным хозяйством в
интегрированной системе защиты растений как основное средство мониторинга
насекомых-вредителей. На сегодняшний день это наиболее рентабельный путь
обнаружения и оценки численности вредителей по сравнению с другими известными
методами.
Существует два способа использования феромонов как средства борьбы с
вредителями: метод массового отлова насекомых в ловушки, снабженные
диспенсером, испускающим феромон, и метод дезориентации (распыление феромона и
как следствие - невозможность найти партнера для спаривания).
Преимущества феромонных препаратов для борьбы с вредителями заключаются:
во-первых, в их специфичности, так как феромон действует только на
определенный вид вредителя, а значит, такой препарат безвреден для нецелевой
фауны;
во-вторых, в природном происхождении действующих веществ и их малых
количествах, необходимых для применения (от десятка мкг до 1-2 мг на ловушку),
что делает этот способ борьбы безвредным для окружающей среды;
в-третьих, из-за малых количеств феромонов для применения возможен синтез
веществ на лабораторных установках, что также уменьшает нагрузку на окружающую
среду.
Нами было проведено исследование по использованию разработанного в Томске
феромонного препарата «Деналол» для борьбы с короедом-типографом. Феромонные
ловушки использовались в качестве опыта, впервые в условиях Томской области.
Выводы
Проведенные исследования показали:
. Эффективность данных феромонных ловушек очень мала.
. Это связано либо с недоработкой феромона, либо с тем, что погодные
условия Томской и Московской области различны. За все время эксперимента в наши
ловушки попал всего один короед. Зато в установленные ловушки попадали другие
насекомые. Так, в третий выезд, в 23 ловушку попало 26 бронзовок золотистых (Agrilus viridis), что трудно назвать простым совпадением. Кроме этого
в установленные ловушки в каждый наш выезд попадали различные виды щелкунов и
боярышниц. Так, особо большое количество было рагий-сыщиков (Rhagium unduisitor) и могильщиков погребальных (Necrophorus verpillo).
. В ловушки попадали не только вредные насекомые, но и полезные (муравьи,
шмели, осы и др.). Возможно, что это попадание было случайным.
. Данный препарат не эффективен как средство борьбы с короедом-стенографом,
но, при определенных доработках он может быть использован для борьбы с другими
видами лесных вредителей.
Список использованной литературы
.Барбье М.
Введение в биохимическую экологию. - М.: Мир, 1978. - 229 с.
.Бей-Биенко
Г.Я. Общая энтомология. - М.: Высшая школа, 1971. - 327 с.
.Биологическая
борьба с вредными насекомыми и сорняками: Пер. с англ. / Под ред. Б.И.
Рукавишникова. - М.: Наука, 1968. - 261 с.
.Биологические
средства защиты растений: Сб. ст. / Под ред. Е.М. Шумакова и др. - М.: Наука,
1974. - 174 с.
.Бондаренко
Н.В. Биологическая защита растении. - Л.: Колос. Ленингр. отд-ние, 1978. - 203
с.
.Бугаев В.А.
Основы лесоустройства. - Воронеж: Изд-во Воронеж. ун-та, 1982. - 232 с.
.Буда В.Г.,
Скиркявичус А.В. Основы феромонной коммуникации у насекомых // Хеморецепция
насекомых. - 1982. - № 7. - С. 24 - 36.
.Булеза В.В.,
Сафонкин А.Р., Мыттус Э.Р. и др. Половые аттрактанты листоверток-вредителей
сада // Химическая коммуникация животных. - М.: Наука, 1986. - С. 57 - 65.
.Вайткявичене
Г.Б., Скиркявичус А.В. Феромон пчелиной матки и морфологические аспекты его
восприятия рабочими пчелами и трутнями Apes mellifera L. // Хеморецепция
насекомых. - 1987. - № 9. - С. 30 - 47.
.Введение в
проблемы биохимической экологии: Биотехнология, сельское хозяйство, охрана
среды / М.М. Телитченко, С.А. Остроумов. - М.: Наука, 1990. - 228 с.
.Ведерников
Н.Е., Яковлев В.Г. Защита хвойных сеянцев от болезней. - М.: Лесная
промышленность, 1972. - 89 с.
.Воробьева
Л.И. Техническая микробиология. - М.: Изд-во МГУ, 1987. - 168 с.
.Воронкова
В.В. Методы выделения феромонов чешуекрылых // Агрохимия. - 1987. - № 4. - С.
124 - 132.
.Воронкова
В.В., Щербакова Г.Д., Бочарова Н.И. и др. О выделении и идентификации феромона
озимой совки Agrotis segetim // Химическая коммуникация животных. - М.: Наука,
1986. - С. 37 - 43.
.Воронцов
А.И. Лесная энтомология. - М.: Лесная промышленность, 1975. - 250 с.
.Воронцов
А.И. Патология леса. - М.: Лесная промышленность, 1978. - 272 с.
.Вредители
сельскохозяйственных культур и лесных насаждений: В 3 т. / Под ред. В.П.
Васильева. - Киев: Урожай, 1975.
.Горожанкина
С.М., Константинов В.Д. География тайги Западной Сибири. - Новосибирск: Наука.
Сиб. отд-ние, 1979. - 188 с.
.Гричанов
И.Я. Нарушение химической коммуникации бабочек хлопковой совки (Heliothis
armigera hubner) компонентами полового феромона самок и антиферомонами //
Химическая коммуникация животных. - М.: Наука, 1986. - С. 73 - 79.
.Гулий В.В.,
Голосова М.А. Вирусы в защите леса от вредных насекомых. - М., Лесная
промышленность, 1975. - 167 с.
.Джекобсон М.
Половые феромоны насекомых: Пер. с англ. - М.: Мир, 1976. - 391 с.
.Егоров Н.С.,
Олескин А.В., Самуилов В.Д. Биотехнология: проблемы и перспективы. - М.: Высшая
школа, 1987. - 159 с.
.Емельянов
В.А., Мыттус Э.Р. Половые феромоны насекомых. - Елгава: Латв.
сельскохозяйственная академия, 1987. - 37 с.
.Журавлев
И.И., Крангауз Р.А., Яковлев В.Г. Болезни лесных деревьев и кустарников. - М.:
Лесная промышленность, 1974. - 157 с.
.Зинкевич
Э.П. Природа феромонов // Человек и природа. - 1984. - № 7. - С. 3 - 9.
.Ильинский
А.И. Определитель вредителей леса. - М.: Сельхозиздат, 1962. - 392 с.
.Ильичев А.Л.
Роль отдельных компонентов в многокомпонентной феромонной системе чешуекрылых
// Биологические науки. - 1988. - № 10. - С. 5 - 15.
.Исмаилов
В.Я., Олещенко И.Н., Орлов В.Н., Алексеенко А.В. Новый подход к регулированию
численности насекомых с помощью синтетических половых феромонов // Химическая
коммуникация животных. - М.: Наука, 1986. - С. 109 - 111.
.Кедровые
леса Сибири / И.В. Семечкин, Н.П. Поликарпов, А.И. Ирошников и др. -
Новосибирск: Наука, 1985. - 256 с.
.Киршенблат
Я.Д. Телергоны - химические средства взаимодействия животных. - М.: Наука,
1974. - 107 с.
.Киршенблат
Я.Д. Телергоны и их биологическое значение // Успехи современной биологии. -
1958. - Т. 46. - С. 316 - 326.
.Крушев Л.Т.
Биологические методы защиты леса от вредителей. - М.: Лесная промышленность,
1973. - 192 с.
.Лебедева
К.В., Мельников Н.Н. Последние достижения в области применения и исследования
феромонов чешуекрылых // Агрохимия. - 1987. - № 3. - С. 127 - 136.
.Лебедева
К.В., Миняйло В.А., Пятнова Ю.Б. Феромоны насекомых. - М.: Наука, 1984. - 268
с.
.Маслов А.Д.,
Кутеев Ф.С., Прибылова М.В. Стволовые вредители леса. - М.: Лесная
промышленность, 1973. - 144 с.
.Микроорганизмы
в борьбе с вредными насекомыми и клещами: Пер. с англ. / Под ред. М.С.
Гилярова. М.: Сельхозиздат, 1976. - 304 с.
.Мыттус Э.Р.,
Гранат Д.А. Применение феромонов для защиты растений. - Таллинн: Эст. НИИНТИ,
1983. - 30 с.
.Надзор, учет
и прогноз массовых размножений хвое- и листогрызущих насекомых в лесах СССР /
Под ред. А.И. Ильинского и И.В. Тропина. - М.: Лесная промышленность, 1965. -
526 с.
.Наставление
по авиационно-химической борьбе с вредителями леса / И.В. Тропин, П.А. Зубов. -
М.: Гослесхоз СССР, 1972. - 84 с.
.Наставление
по использованию птиц для защиты лесов от вредителей / К.И. Благосклонов, А.В.
Карпенко. - М.: Гослесхоз СССР, 1975. - 44 с.
.Наставление
по надзору, учету и прогнозу массовых размножений стволовых вредителей леса /
А.Д. Маслов, Ф.С. Кутеев, М.В. Прибылова. - М.: Гослесхоз СССР, 1975. - 89 с.
.Негруцкий
С.Ф. Корневая губка. - М.: Лесная промышленность, 1973. - 200 с.
.Осмоловский
Г.Е., Бондаренко Н.В. Энтомология: Учебник. - 2-е изд., перераб. и доп. - Л.:
Колос. Ленингр. отд-ние, 1980. - 359 с.
.Павлов И.Ф.
Агротехнические и биологические методы защиты растений. - М.: Сельхозиздат,
1976. - 274 с.
.Постановление
Правительства Российской Федерации от 01.06.98 № 551 «Об утверждении Правил
отпуска древесины на корню в лесах Российской Федерации» // СЗ РФ. - 1998. - №
23. - Ст. 2553.
.Приказ
Рослесхоза от 15 января 1998 г. № 10 "Об утверждении Санитарных правил в
лесах Российской Федерации" (с изменениями от 24 декабря 1998 г.) // СЗ
РФ. - 1998. - № 3. - Ст. 372.
.Рябчинская
Т.А., Колесова Д.А., Мыттус Э.Р. Аттрактивность синтетических половых феромонов
для садовых листоверток (Lepidoptera, Tortricidae) // Зоологический журнал. -
1986. - Т. 65. - № 10. - С.1490 - 1499.
.Сегаль Г.М.
Феромоны и другие природные продукты // Агрохимия. - 1983. - № 6. - С. 96 -
103.
.Скиркявичюс
А.В. Феромонная коммуникация насекомых. - Вильнюс: Мокслас, 1986. - 292 с.
.Скиркявичюс
А.В. Феромоны. - Вильнюс: Ин-т зоологии и паразитологии АН ЛитССР, 1988. - 367
с.
.Словарь-справочник
энтомолога / Под ред. В.Н. Щеголева. - М.: Наука, 1958. - 260 с.
.Соколов Д.В.
Корневая гниль от опенка и борьба с ней. - М.: Лесная промышленность, 1964. -
183 с.
.Справочник
механизатора лесного хозяйства. - 2-е изд. - М.: Лесная промышленность, 1977. -
296 с.
.Справочник
по защите леса от вредителей и болезней / И.В. Тропин, Н.М. Ведерников, Р.А.
Крангауз и др. - М.: Лесная промышленность, 1980. - 376 с.
.Справочник
по пестицидам (гигиена применения и токсикология) / Под ред. Л.И. Медведя. -
Киев: Урожай, 1977. - 376 с.
.Стадницкий
Г.В., Юрченко Г.И., Сметанин А.Н. и др. Вредители шишек и семян хвойных пород.
- М.: Лесная промышленность, 1978. - 168 с.
.Старк В.Н. и
др. Вредители и болезни полезащитных лесных насаждений. - Л.: Колос. Ленингр.
отд-ние, 1951. - 261 с.
.Суитмен X.
Биологический метод борьбы с вредными насекомыми и сорными растениями: Пер. с
англ. - М.: Сельхозиздат, 1964. - 186 с.
.Тропин И.В.
Краткий справочник по химическим средствам защиты леса от вредителей и
болезней. - М.: Лесная промышленность, 1973. - 152 с.
.Тропин И.В.
Химическая защита леса от насекомых. - М.: Лесная промышленность, 1968. - 380
с.
.Тыщенко В.П.
Основы физиологии насекомых: В 2-х ч. - Л.: Наука, 1977.
.Харборн Дж.
Введение в экологическую биохимию: Пер. с анг. - М.: Мир, 1985. - 312 с.
.Черемисинов
Н.А., Негруцкий С.Т., Лешковцева И.И. Грибы и грибные болезни деревьев и
кустарников. - М.: Лесная промышленность, 1970. - 392 с.
.Шварц С.С.,
Пястолова О.А., Добринская Г.А., Рункова Г.Г. Эффект группы в популяциях водных
животных и химическая экология. - М.: Наука, 1976. - 152 с.
.Щеголев В.Н.
Сельскохозяйственная энтомология. - Л.: Наука, 1960. - 274 с.
Приложение
Таблица 1. Выезд 3 июня. Ясный солнечный день.
|
Ловушка
|
Обнаруженные насекомые
|
|
1 ловушка
|
1 боярышница (Aporia crataedi), 5 муравьев (Formica polyctena Foerst.), 1
толстоголовая оса большая (Сrabro cribrarius)
|
|
3 ловушка
|
1 могильщик погребальный (Necrophorus verpillo), 1 рагий-сыщик (Rhagium unduisitor), 4 щелкуна (Slateridae)
|
|
4 ловушка
|
1 клоп итальянский (Graphosoma lineatum), 1 щитоноска (Coccus
hesperidum L.), 3 щелкуна
красноногова (Melanotus rufipes), 1
щелкун блестящий (Selatosomis aeneus), 2
шмеля лесного (Botbus silvasum).
|
|
7 ловушка
|
1 могильщик погребальный (Necrophorus verpillo), 5 муравьев (Formica polyctena Foerst.), 2
грибоеда красноголовых (Tripeax rissica)
|
|
9 ловушка
|
2 боярышницы (Aporia crataedi), 1 долгоносик полосатый (Sitona
eineatus)
|
|
10 ловушка
|
1 коровка (Сoccineceidae), 3 муравья (Formica polyctena Foerst.).
|
|
12 ловушка
|
1 щелкун красноногий (Melanotus rufipes), 2 синих рогохвоста (Paururus juvencus)
|
|
15 ловушка
|
2 черных сосновых усача (Monochamus galloprovincialis oliv), 1
толстоголовая оса большая (Crabro cribrarius).
|
|
16 ловушка
|
1 грибоед красноголовый (Tripeax rissica), 3 сосновая тля (Lachnus pineus Mordk),
|
|
17 ловушка
|
4 клопа итальянских (Graphosoma lineatum),
|
|
19 ловушка
|
1 синий рогохвост (Paururus juvencus), 3 щелкуна красноногова (Melanotus rufipes), 4 муравья (Formica polyctena Foerst.), 1
могильщик погребальный (Necrophorus verpillo)
|
|
20 ловушка
|
3 рагий-сыщик (Rhagium unduisitor), 1 щелкун (Slateridae), 1
щелкун блестящий (Selatosomus aeneus)
|
|
23 ловушка
|
2 рагий-сыщик (Rhagium unduisitor), 1 бояр. (Aporia
crataedi), 1 щелкун блест. (Selatosomis aeneus), 2 шмеля лесного (Botbus
silvasum).
|
|
26 ловушка
|
2 щелкуна (Slateridae), 1 щитник двузубчатый (Picromesus bidens),
|
|
29 ловушка
|
2 грибоед красноголовый (Tripeax rissica), 1 щелкун красноногий (Melanotus rufipes), 5 сосновая тля (Lachnus pineus Mordk),
|
|
31 ловушка
|
4 сосновой тли (Lachnus pineus Mordk), 1
долгоносик полосатый (Sitona eineatus).
|
|
36 ловушка
|
1 рагий-сыщик (Rhagium unduisitor), 1 боярышница (Aporia
crataedi), 2 синих рогохвоста (Paururus juvencus)
|
|
37 ловушка
|
1 толстоголовая оса большая
(Crabro cribrarius), 2
сибирский гравёр (Melolonthe hippocasteni Fabr)
|
|
39 ловушка
|
2 могильщика погребальных (Necrophorus verpillo), 1 рагий-сыщик (Rhagium unduisitor), 1 грибоед красноголовый (Tripeax rissica), 2 шмеля лесного (Botbus
silvasum).
|
|
40 ловушка
|
1 щелкун (Slateridae)
|
|
44 ловушка
|
1 грибоед красног. (Tripeax rissica), 1 могильщик пог. (Necrophorus verpillo), 1 рагий-сыщик (Rhagium unduisitor), 1 щелкун (Slateridae)
|
|
48 ловушка
|
1 рагий-сыщик (Rhagium unduisitor). 3 синий рогохвост (Paururus juvencus), 1 короед стенограф (Ips
sexdentatus)
|
|
49 ловушка
|
1 черный сосновый усач (Monochemus gelloprovincialis oliv), 1
рагий-сыщик (Rhagium unduisitor), 1
щелкун (Slateridae), 1 щелкун блестящий (Selatosomus aeneus).
|
|
52 ловушка
|
1 синий рогохвост (Paururus juvencus), 1 могильщик погребальный (Necrophorus verpillo).
|
|
54 ловушка
|
1 щелкун (Slateridae), 2 грибоед красноголовый (Tripeax rissica), 1 боярышница (Aporia crataedi), 1
синий рогохвост (Paururus juvencus),
|
2 грибоед красноголовый (Tripeax rissica), 1 могильщик погребальный (Necrophorus verpillo), 1 щелкуна красноногова (Melanotus rufipes), 1 синий рогохвост (Paururus juvencus).
|
|
59 ловушка
|
1 щелкуна красноногий (Melanotus rufipes), 1 боярышница (Aporia
crataedi), 1 щелкун (Slateridae).
|
|
60 ловушка
|
1 щелкун (Slateridae), 2 смолевка сосновая (Pissodes pini)
|
Таблица № 2. Выезд 13 июня. Пасмурный день.
|
Ловушка
|
Обнаруженные насекомые
|
|
1 ловушка
|
1 рагий-сыщик (Rhagium unduisitor), 2 грибоед красноголовый (Tripeax rissica
|
|
6 ловушка
|
2 боярышница (Aporia crataedi), 1 могильщик погребальный (Necrophorus verpillo), 1 щелкун (Slateridae).
|
|
8 ловушка
|
1 комлевый бурый усач 5
сосновая тля (Lachnus pineus
Mordk), 1 щелкуна крас. (Melanotus rufipes), 2 большой сосновый долг. (Hylobis abietis).
|
|
14 ловушка
|
1 синий рогохвост (Paururus juvencus), 1 толстоголовая оса большая (Crabro cribrarius), 1 щелкун (Slateridae).
|
|
19 ловушка
|
2 могильщик погребальный (Necrophorus verpillo), 1 боярышница (Aporia
crataedi).
|
|
24 ловушка
|
1 рагий-сыщик (Rhagium unduisitor), 2 большой сосновый долгоносик (Hylobis abietis), 1 щитник черношипный (Carpocoris fuscispinus), 1 большой хвойный рогохвост (Uroceras gigas)
|
|
27 ловушка
|
1 грибоед красноголовый (Tripeax rissica), 2 боярышница (Aporia
crataedi), 1 синий рогохвост (Paururus juvencus)
|
|
28 ловушка
|
2 щелкун красноногий (Melanotus rufipes), 1 могильщик погребальный (Necrophorus verpillo), 1 рагий-сыщик (Rhagium unduisitor).
|
|
32 ловушка
|
1. щелкун блестящий (Selatosomus aeneus), 1 долгоносик полосатый (Sitona
eineatus).
|
|
33 ловушка
|
1 боярышница (Aporia crataedi), 2 щелкуна красноногова (Melanotus rufipes).
|
|
40 ловушка
|
2 грибоед красноголовый (Tripeax rissica)
|
|
44 ловушка
|
1 комлевый бурый усач (Criocephelus rusticus L) 1
рагий-сыщик (Rhagium unduisitor), 2
боярышница (Aporia crataedi), 1
синий рогохвост (Paururus juvencus)
|
|
45 ловушка
|
4 сосновая тля (Lachnus pineus Mordk), 1
щелкун блестящий (Selatosomus aeneus), 1
сибирский гравёр (Melolonthe hippocasteni Fabr)
|
|
46 ловушка
|
1 могильщик погребальный (Necrophorus verpillo), 4 щелкуна красноногова (Melanotus rufipes), 1 синий рогохвост (Paururus juvencus).
|
|
47 ловушка
|
1 толстоголовая оса большая
(Crabro cribrarius), 1
сибирский гравёр (Melolonthe hippocasteni Fabr).
|
|
49 ловушка
|
3 сосновая тля (Lachnus pineus Mordk), 1
грибоед красноголовый (Tripeax rissica), 2
муравья (Formica polyctena Foerst.), 1 боярышница (Aporia
crataedi), 1 черный сосновый усач (Monochamus galloprovincialis oliv).
|
|
52 ловушка
|
1 рагий-сыщик (Rhagium unduisitor).
|
|
53 ловушка
|
1 могильщик погреб. (Necrophorus verpillo) 3 муравья, 2 сосновая тля (Lachnus pineus Mordk), 1
большой сосновый долгоносик (Hylobis abietis).
|
|
55 ловушка
|
1 черный сосновый усач (Monochamus galloprovincialis oliv), 1
большой сосновый долгоносик (Hylobis abietis)
|
|
56 ловушка
|
1 щелкун красноногий (Melanotus rufipes), 1 муравей.
|
|
57 ловушка
|
1 синий рогохвост (Paururus juvencus), 2 щелкун (Slateridae), 1
большой сосновый долгоносик (Hylobis abietis).
|
|
58 ловушка
|
1 комлевый бурый усач (Criocephelus rusticus L), 2
могильщик погребальный (Necrophorus verpillo), 1
долгоносик полосатый (Sitona eineatus).
|
|
59 ловушка
|
1 щелкун блестящий (Selatosomus aeneus), 3 сибирский гравёр (Melolonthe hippocasteni Fabr).
|
|
60 ловушка
|
1 рагий-сыщик (Rhagium unduisitor), 2 грибоед красноголовый (Tripeax rissica), 1 большой сосновый долгоносик (Hylobis abietis).
|
Таблица № 3. Выезд 23 июня. Дождь.
|
Ловушка
|
Обнаруженные насекомые
|
|
3 ловушка
|
2 рагий-сыщик (Rhagium unduisitor).
|
|
4 ловушка
|
1 щелкун красноногий (Melanotus rufipes), 1 рагий-сыщик (Rhagium unduisitor).
|
|
5 ловушка
|
2 синий рогохвост (Paururus juvencus), 1 могильщик погребальный (Necrophorus verpillo), 1 боярышница (Aporia
crataedi).
|
|
6 ловушка
|
1 грибоед красноголовый (Tripeax rissica), 1 черный сосновый усач (Monochamus galloprovincialis oliv), 2
щелкун (Slateridae).
|
|
10 ловушка
|
1 синий рогохвост (Paururus juvencus), 1 черный сосновый усач (Monochamus galloprovincialis oliv).
|
|
11 ловушка
|
1 усач-инквизитор (Rhagium inquisitor), 1 рагий-сыщик (Rhagium unduisitor), 3 сосновая тля (Lachnus pineus Mordk).
|
|
12 ловушка
|
2 щелкун (Slateridae), 1 щелкун блестящий (Selatosomus aeneus).
|
|
13 ловушка
|
2 щелкуна красноногова (Melanotus rufipes), 1 могильщик погребальный (Necrophorus verpillo).
|
|
19 ловушка
|
2 рагий-сыщик (Rhagium unduisitor), 1 долг. Полос. (Sitona
eineatus), 1 щелкун красн. (Melanotus rufipes) 4 сосновая тля (Lachnus pineus Mordk).
|
|
23 ловушка
|
26 бронзовка золотистая
(Agrilus viridis)
|
|
28 ловушка
|
1 щелкун красноногий (Melanotus rufipes), 1 долгоносик полосатый (Sitona
eineatus), 2 черный длинноусый усач (Acenthocinus aedilis L).
|
|
30 ловушка
|
1 могильщик погребальный (Necrophorus verpillo), 1 синий рогохвост (Paururus juvencus)
|
Таблица № 4. Выезд 2 июля. Пасмурно, периодами дождь
|
Ловушка
|
Обнаруженные насекомые
|
|
3 ловушка
|
1 усач-инквизитор (Rhagium inquisitor), 2 щелкун красноногий (Melanotus rufipes).
|
|
4 ловушка
|
3 могильщик погребальный (Necrophorus verpillo), 2 сибирский гравёр (Melolonthe hippocasteni Fabr).
|
|
6 ловушка
|
1 сибирский гравёр (Melolonthe hippocasteni Fabr) 1
грибоед красноголовый (Tripeax rissica), 1
могильщик погребальный (Necrophorus verpillo), 1 рагий-сыщик
(Rhagium unduisitor).
|
|
7 ловушка
|
1 рагий-сыщик (Rhagium unduisitor), 1 щелкун блестящий (Selatosomus aeneus).
|
|
8 ловушка
|
1 усач-инквизитор (Rhagium inquisitor).
|
|
9 ловушка
|
2 сосновая тля (Lachnus pineus Mordk).
|
|
12 ловушка
|
1 муравей (Formica polyctena Foerst.)
|
|
13 ловушка
|
1 синий рогохвост (Paururus juvencus).
|
|
15 ловушка
|
1 грибоед красноголовый (Tripeax rissica), 1 сосновая тля (Lachnus pineus Mordk).
|
|
17 ловушка
|
1 могильщик погребальный (Necrophorus verpillo), 2 толстоголовая оса большая (Crabro cribrarius).
|
|
19 ловушка
|
1 синий рогохвост (Paururus juvencus).
|
|
22 ловушка
|
5 сосновая тля (Lachnus pineus Mordk), 1
грибоед красноголовый (Tripeax rissica).
|
|
23 ловушка
|
1 черный длинноусый усач (Acenthocinus aedilis L).
|
|
24 ловушка
|
1 усач-инквизитор (Rhаgium inquisitor), 1 грибоед красноголовый (Tripeax rissica)
|
|
26 ловушка
|
1 толстоголовая оса большая
(Сrabro cribrarius),3
синий рогохвост (Paururus juvencus)
|
|
27 ловушка
|
1 долгоносик полосатый (Sitona eineatus).
|
|
29 ловушка
|
1 усач-инквизитор (Rhagium inquisitor) 2 щелкун (Slateridae),
|
|
30 ловушка
|
2 большой сосновый
долгоносик (Hylobis abietis), 1
щитник черношипный (Carpocoris fuscispinus),
1 большой хвойный рогохвост (uroceras gigas), 1
черный сосновый усач (Monochamus galloprovincialis oliv).
|
Таблица № 5. Выезд 12 июля. Дождь.
|
Ловушка
|
Обнаруженные насекомые
|
|
2 ловушка
|
1 комлевый бурый усач (Criocephelus rusticus L), 3 сосновая
тля (Lachnus pineus
Mordk), 2 большой сосновый долгоносик (Hylobis abietis).
|
|
3 ловушка
|
2 щитник черношипный (Carpocoris fuscispinus)
|
|
6 ловушка
|
1 синий рогохвост (Paururus juvencus), 1 толст. оса большая (Crabro
cribrarius), 1 щелкун (Slateridae), 1
грибоед красн. (Tripeax rissica).
|
|
8 ловушка
|
1 комлевый бурый усач (Criocephelus rusticus L), 1
долгоносик полосатый (Sitona eineatus).
|
|
10 ловушка
|
4 сосновая тля (Lachnus pineus Mordk), 1
щелкун блестящий (Selatosomus aeneus).
|
|
13 ловушка
|
1 большой сосновый
долгоносик (Hylobis abietis), 1
грибоед красноголовый (Tripeax rissica), 1
синий рогохвост (Paururus juvencus).
|
|
14 ловушка
|
5 щелкун (Slateridae), 1 долгоносик полосатый (Sitona eineatus).
|
|
18 ловушка
|
1 боярышница (Aporia crataedi), 1 синий рогохвост (Paururus juvencus), 1 черный сосновый усач (Monochamus galloprovincialis oliv), 1
большой сосновый долгоносик (Hylobis abietis)
|
|
20 ловушка
|
1 грибоед красноголовый (Tripeax rissica), 1 сибирский гравёр (Melolonthe hippocasteni Fabr).
|
|
22 ловушка
|
1 синий рогохвост (Paururus juvencus), 1 черный сосновый усач (Monochamus galloprovincialis oliv).
|
|
26 ловушка
|
1 усач-инквизитор (Rhagium inquisitor), 1 щелкун красноногий (Melanotus rufipes),
|
|
27 ловушка
|
1 синий рогохвост (Paururus juvencus), 2 щелкуна красноногова (Melanotus rufipes), 1 щелкун блестящий (Selatosomus aeneus).
|
|
28 ловушка
|
1 грибоед красноголовый (Tripeax rissica), 1 синий рогохвост (Paururus juvencus).
|
|
29 ловушка
|
2 смолевка сосновая (Pissodes pini). 1 усач-инквизитор (Rhagium inquisitor) 1 щелкун (Slateridae).
|