Соотношение онто- и филогенеза. Закон зародышевого сходства
Медицинский
институт
Кафедра
Биологии
Курсовая
работа по дисциплине
«Биология»
Тема:
«Соотношение онто и филогенеза. Закон зародышевого сходства»
Содержание
Введение
Глава
1. Соотношение онто и филогенеза
.1
Понятие онтогенеза и филогенеза
.2
Понятие филогенеза
.3
Соотношение онтогенеза и филогенеза
Глава
2. Закон зародышевого сходства
.1
Закон зародышевого сходства К. Бэра
.2
Биогенетический закон Э. Геккеля
Заключение
Список
используемых источников информации
онтогенез филогенез зародышевое
сходство
Введение
Цель выполнения курсовой работы:
Показать соотношение онтогенеза и филогенеза,
раскрыть взаимосвязь этих понятий. Также рассмотреть закон зародышевого
сходства.
Онтогенез и филогенез является основой многих
биологических наук, основой эволюционного учения. Термины «онтогенез» и
«филогенез» используются для описания развития, поэтому между этими различными
понятиями существуют и признаки различия, и признаки сходства. Поэтому в этой
работе будет рассмотрено понятие онтогенеза и филогенеза, будет дана
сравнительная характеристика этих понятий. Также особое внимание будет уделено
закону зародышевого сходства К. Бэра, будет приведена сравнительная иллюстрация
этого закона.
Также будут рассмотрены законы, сформулированные
в дальнейшем другими учеными на основе закона зародышевого сходства К. Бэра.
Глава 1. Соотношение онто и филогенеза
.1 Понятие онтогенеза
Онтогенез - это индивидуальное развитие
организма, в ходе которого происходит преобразование его морфофизиологических,
физиолого-биохимических и цитогенетических признаков. Говоря более простым
языком, онтогенез или индивидуальное развитие -это период времени, который
длится с момента зачатия и заканчивается в момент смерти.
Поэтому в онтогенезе выделяют 2 периода
развития: эмбриональный и постэмбриональный.
В ходе эмбрионального периода эмбрион человека находится в полости матки
матери, где получает все необходимые питательные вещества и кислород, а также
отдает продукты обмена. Такой обмен веществ между материнским организмом и эмбрионом
становится возможен благодаря наличию специального органа - плаценты.
У большинства многоклеточных животных,
независимо от сложности их организации, стадии эмбрионального развития, которые
проходит зародыш, едины. В эмбриональном периоде выделяют три основных этапа:
дробление, гаструляцию и первичный органогенез.
Дробление. Развитие организма начинается со
стадии одной клетки. Оплодотворенное яйцо - это клетка и одновременно уже
организм на самой ранней стадии его развития. В результате многократных делений
одноклеточный организм превращается в многоклеточный. Возникшее при
оплодотворении путем слияния сперматозоида и яйцеклетки диплоидное ядро через
несколько минут начинает делиться, вместе с ним делится и цитоплазма.
Образующиеся клетки с каждым делением уменьшаются в размерах, поэтому процесс
деления носит название дробления. В период дробления накапливается клеточный
материал для дальнейшего развития. Завершается дробление образованием
многоклеточного зародыша - бластулы. Бластула имеет полость, наполненную
жидкостью, так называемую первичную полость тела.
В тех случаях, когда в цитоплазме яйцеклетки
желтка мало (как у ланцетника) или относительно немного (как у лягушки),
дробление бывает полным, т. е. яйцеклетка делится целиком.
Иначе протекает период дробления у птиц.
Свободная от желтка цитоплазма составляет всего 1% от общего объема яйцеклетки
курицы; вся остальная цитоплазма яйцеклетки, а следовательно и зигота,
заполнена массивом желтка. Если присмотреться к куриному яйцу, на одном из его
полюсов непосредственно на желтке можно увидеть маленькое пятнышко - бластулу,
или зародышевый диск, образовавшийся в результате дробления свободного от
желтка участка цитоплазмы, содержащего ядро. В таких случаях дробление называют
неполным. Неполное дробление свойственно и некоторым рыбам и рептилиям.
Во всех случаях - и у ланцетника, и у амфибий, и
у птиц, а также у других животных - общий объем клеток на стадии бластулы не
превышает объема зиготы. Другими словами, митотическое деление зиготы не
сопровождается ростом образовавшихся дочерних клеток до объема материнской, и
размеры их в результате ряда последовательных делений прогрессивно уменьшаются.
Эта особенность митотического деления клеток в ходе дробления наблюдается при
развитии оплодотворенных яиц у всех животных.
Некоторые другие черты дробления также
свойственны различным видам животных. Например, все клетки в бластуле имеют
диплоидный набор хромосом, одинаковы по строению и отличаются друг от друга
главным образом количеством содержащегося в них желтка. Такие клетки, лишенные
признаков специализации для выполнения определенных функций, называют
неспециализированными (или недифференцированными) клетками. Другая особенность
дробления - чрезвычайно короткий митотический цикл бластомеров по сравнению с клетками
взрослого организма. Во время очень короткой интерфазы происходит только
удвоение ДНК.
Гаструляция. Бластула, как правило, состоящая из
большого числа бластомеров (например, у ланцетника из 3000 клеток), в процессе
развития переходит в новую стадию, которую называют гаструлой (от греч.гастер -
желудок). Зародыш на этой стадии состоит из отчетливо различимых пластов клеток
- так называемых зародышевых листков: наружного, или эктодермы (от греч.эктос -
находящийся снаружи), и внутреннего, или энтодермы (от греч. энтос -
находящийся внутри). Совокупность процессов, приводящих к образованию гаструлы,
называют гаструляцией.
У ланцетника гаструляция осуществляется путем
впячивания одного из полюсов бластулы внутрь, по направлению к другому, у
других животных - либо путем расслоения стенки бластулы, либо путем обрастания
массивного вегетативного полюса мелкими клетками анимального полюса.
У многоклеточных животных, кроме
кишечнополостных, параллельно с гаструляцией или, как у ланцетника, вслед за
ней возникает и третий зародышевый листок - мезодерма (от греч. мезос -
находящийся посередине), которая представляет собой совокупность клеточных
элементов, расположенных между экто- и энтодермой в первичной полости тела -
бластоцеле. С появлением мезодермы зародыш становится трехслойным.
Таким образом, сущность процесса гаструляции
заключается перемещении клеточных масс. Клетки зародыша практически делятся и
не растут. Однако на этой стадии начинается использование генетической
информации клеток зародыша, появляются первые признаки дифференцировки.
Дифференцировка, или дифференцирование, - это
процесс ее возникновения и нарастания структурных и функциональных различий
между отдельными клетками и частями зародыша. С морфологической точки зрения
дифференцирование выражается в том, что образуются несколько сотен типов клеток
специфического строения, отличающихся друг от друга. Из неспециализированных
клеток бластулы постепенно возникают клетки эпителия кожи, эпителия кишечника,
легких, появляются нервные, мышечные клетки и т.д. С биохимической точки зрения
специализация клеток заключается в способности синтезировать определенные
белки, свойственные только данному типу клеток. Лимфоциты синтезируют защитные
белки - антитела, мышечные клетки - сократительный белок миозин. Каждый тип
клеток образует «свои», свойственные только ему белки. Биохимическая
специализация клеток обеспечивается избирательной - дифференциальной
активностью генов, т. е. в клетках разных зародышевых листков - зачатков
определенных органов и систем - начинают функционировать разные группы генов.
У разных видов животных одни и те же зародышевые
листки дают начало одним и тем же органам и тканям. Это значит, что они
гомологичны. Так, из клеток наружного зародышевого листка - эктодермы - у
членистоногих, хордовых, в том числе у рыб, амфибий, рептилий, птиц и
млекопитающих, формируются кожные покровы и их производные, а также нервная
система и органы чувств. Гомология зародышевых листков подавляющего большинства
животных - одно из доказательств единства животного мира.
Органогенез. После завершения гаструляции у
зародыша образуется комплекс осевых органов: нервная трубка, хорда, кишечная
трубка. У ланцетника осевые органы формируются следующим образом: эктодерма на
спинной стороне зародыша прогибается по средней линии, превращаясь в желобок, а
эктодерма, расположенная справа и слева от него, начинает нарастать на его
края. Желобок - зачаток нервной системы - погружается под эктодерму, и края его
смыкаются. Образуется нервная трубка. Вся остальная эктодерма - зачаток кожного
эпителия.
Спинная часть энтодермы, располагающаяся
непосредственно под нервным зачатком, обособляется от остальной энтодермы и
сворачивается в плотный тяж - хорду. Из оставшейся части энтодермы развиваются
мезодерма и эпителий кишечника. Дальнейшая дифференцировка клеток зародыша
приводит к возникновению многочисленных производных зародышевых листков -
органов и тканей. В процессе специализации клеток, входящих в состав
зародышевых листков, из эктодермы образуются нервная система, органы чувств,
эпителий кожи, эмаль зубов; из энтодермы - эпителий кишки, пищеварительные
железы - печень и поджелудочная железа, эпителий жабр и легких; из мезодермы -
мышечная ткань, соединительная ткань, в том числе рыхлая соединительная ткань,
хрящевая и костная ткани, кровь и лимфа, а также кровеносная система, почки,
половые железы.
Постэмбриональный период развития
В момент рождения или выхода организма из
яйцевых оболочек заканчивается эмбриональный и начинается постэмбриональный
период развития. Постэмбриональное развитие может быть прямым или
сопровождается превращением (метаморфозом).
При прямом развитии (у пресмыкающихся, птиц,
млекопитающих) из яйцевых оболочек или из тела матери выходит организм
небольших размеров, но с уже заложенными всеми основными органами,
свойственными взрослому животному. Постэмбриональное развитие в этом случае
сводится в основном к росту и половому созреванию.
При развитии с метаморфозом из яйца выходит
личинка, обычно устроенная проще взрослого животного, со специальными личиночными
органами, во взрослом состоянии отсутствующими. Личинка питается, растет, и со
временем личиночные органы заменяются органами, свойственными взрослым особям.
Следовательно, при метаморфозе разрушаются личиночные органы и возникают
органы, присущие взрослым животным.
Например, метаморфоза может служить также
развитие насекомых. Гусеницы бабочек или личинки стрекоз резко отличаются по
строению, образу жизни и среде обитания от взрослых животных и напоминают своих
предков - кольчатых червей.Таким образом, метаморфоз связан с переменой образа
жизни или среды обитания. Значение метаморфоза заключается, во-первых, в том,
что свободноживущие личинки прикрепленных или паразитических животных
способствуют расселению вида. Кроме того, личиночные формы некоторых животных
живут в иных условиях и имеют другие источники питания, чем взрослые особи: это
снижает интенсивность конкуренции за пищу и в целом остроту борьбы за
существование внутри вида.
Постэмбриональный период развития имеет разную
продолжительность. Например, поденки в личиночном состоянии живут 2-3 года, а в
половозрелом - от 2-3 часов до 2-3 суток в зависимости от видовой
принадлежности. В большинстве же случаев постэмбриональный период более
продолжителен. У человека он включает стадию полового созревания, стадию
зрелости и стадию старости.
У млекопитающих и человека наблюдается известная
зависимость продолжительности жизни от длительности полового созревания и
беременности. Обычно продолжительность жизни превышает дорепродуктивный период
онтогенеза в 5-8 раз.
Постэмбриональное развитие сопровождается
ростом. Различают рост неопределенный, продолжающийся в течение всей жизни, и
определенный, ограниченный каким-то сроком. Неопределенный рост наблюдается у
древесных форм растений, некоторых моллюсков, из позвоночных - у рыб, крыс.
У многих животных рост прекращается вскоре после
достижения половой зрелости. У человека рост заканчивается к 20-25 годам [9].
Выделяют следующие видоизмененные периоды
онтогенеза, имеющие эволюционное значение.
Диапауза. В эмбриональном периоде ряда
позвоночных существует диапауза, т.е. остановка развития на более или менее
продолжительный период. Она имеет приспособительное значение. Так, у сумчатых и
грызунов развитие эмбрионов останавливается в случае совпадения по времени
развития нового потомства со вскармливанием самкой предыдущего помета. Это
лактационная пауза, продолжающаяся обычно несколько суток. В других отрядах
млекопитающих наблюдаются длительные, до 7 мес. обязательные диапаузы. К
примеру, у соболя оплодотворение происходит в июле. Оплодотворенное яйцо
дробится, процесс развития доходит до стадии бластоцисты и тормозится.
Имплантация и дальнейшее развитие начинаются только в марте. Описанная диапауза
возникла в эволюции названного вида животных путем отбора на рождение потомства
в сезон, наиболее благоприятный для вскармливания.
Деэмбрионизация. Под этим термином понимают
сильное укорочение собственно эмбрионального периода, протекающего под
оболочками яйца. Деэмбрионизация, наблюдающаяся у плацентарных млекопитающих,
сочетается с резким уменьшением желтка в их яйцеклетках и установлением связи с
материнским организмом через плаценту.
Эмбрионизация. Процесс эмбрионизации заключается
в удлинении времени защищенности зародыша от внешней среды благодаря
зародышевым оболочкам и материнскому организму. Зародыш в это время проходит
периоды, соответствующие собственно эмбриональному и личиночному. Развивающийся
организм защищен до момента формирования ювенильной стадии. После рождения не
происходит существенной перестройки организма, и развитие идет прямое, т.е. без
метаморфоза.
Эмбрионизация развития особенно проявилась в
процессе эволюции наземных позвоночных. Уже у многих земноводных, например у
рогатой лягушки Соломоновых островов, все развитие происходит под яйцевыми
оболочками, из которых вылупляется уже претерпевшее метаморфоз животное.
Полнаяэмбрионизация имеет место у пресмыкающихся и птиц в связи с переходом их
к наземному существованию. Особое изменение эмбрионального и личиночного
периодов произошло у плацентарных млекопитающих. Наряду с деэмбрионизацией, о
чем говорилось выше, у них произошла максимальнаяэмбрионизация, но не под
оболочками яйца, а внутриутробно. Период свободного личиночного развития
полностью исчез.
Проходящие под яйцевыми оболочками или
внутриутробно предплодный и плодный периоды развития вместе равнозначны
личиночному периоду. Они следуют непосредственно за эмбриональным периодом,
включают средние и более поздние фазы морфогенеза, характеризуются наличием
провизорных органов, переходом к активному питанию, началом функционирования
органов чувств (слуха, вкуса и др.).
К провизорным органам у млекопитающих относят
зародышевые оболочки, некоторые отделы кровеносной системы, временные структуры
кожного покрова. У яйцекладущих и сумчатых млекопитающих зародыш на предплодных
этапах уже переходит к питанию молоком матери, а у птиц и плацентарных
млекопитающих - заглатывает амниотическую жидкость. Начало функционирования
органа слуха описано у зародышей страуса эму, которые энергично реагируют на
крик взрослых птиц уже на последней четверти инкубационного периода.
Период метаморфоза у наземных позвоночных
соответственно упрощается, так как плод в значительной степени похож на
взрослую особь. Однако отбрасывание зародышевых оболочек, открытие
эмбриональных заращений, изменения в органах дыхания, кровообращения и в коже в
момент родов и в первую неделю после родов представляют собой перестройки,
соответствующие метаморфозу.
Ряд биологов в составе постэмбрионального
периода онтогенеза выделяют собственно постэмбриональный период (с момента
рождения до момента приобретения половой зрелости) и период старения (от
момента приобретения половой зрелости до смерти). Выделение этих периодов
целесообразно в биологии, поскольку ряд организмов погибают сразу после
размножения. В медицине же эти периоды выделять не принято, поскольку каждый
здоровый человек достигает половой зрелости, а затем проходит период старения.
Видоизмененным периодом онтогенеза является
неотения. В эволюции многих групп позвоночных наблюдается тенденция к более
раннему достижению половой зрелости. Наиболее ярко эта тенденция выражена у
хвостатых земноводных. Так, в семействе Ambistomatidae личинки (аксолотли)
приобрели возможность размножаться до наступления периода метаморфоза и
превращения во взрослую стадию, однако способности превращаться во взрослую
стадию они полностью не потеряли.
Известны популяции таких животных, в которых
сосуществуют неотенические и взрослые формы. Это частичная неотения. При полной
неотении ювенильный и взрослый периоды выпадают полностью и о его существовании
у предков судят только по близкородственным видам. Некоторые ученые полагают,
что на основе преобразования личиночных стадий мог возникнуть ряд крупных
таксонов. Считают, что неотения могла явиться основным направлением эволюции
бескилевых бегающих птиц и человека [1].
Основные характеристики онтогенеза
Исходная запрограммированность процессов.
Наличие уникальной неизменной генетической программы развития, сформированной
вследствие мейоза и оплодотворения
Необратимость онтогенеза. При реализации
генетической программы невозможен возврат к предыдущим стадиям
Углубление специализации: по мере развития
уменьшается вероятность смены траектории онтогенеза
Адаптивный характер: поливариантность онтогенеза
обеспечивает возможность приспособления к различным условиям
Неравномерность темпов: скорость процессов роста
и развития изменяется.
Целостность и преемственность отдельных этапов.
Признаки, появляющиеся на более поздних стадиях, базируются на признаках,
проявляющихся на ранних стадиях
Наличие цикличности: существует цикличность
старения и омоложения
Наличие критических периодов, связанных с
выбором пути в узловых точках (точках бифуркации) или с преодолением
энергетических порогов [12].
Основные типы онтогенеза
. Онтогенез организмов с чередованием ядерных
фаз при споровом мейозе (большинство растений и грибов).
. Онтогенез организмов с чередованием полового и
бесполого размножения без смены ядерных фаз. Метагенез - чередование поколений
у Кишечнополостных. Гетерогония - чередованиепартеногенетического и
амфимиктического поколений у червей, некоторых членистоногих и низших хордовых.
. Онтогенез с наличием личиночных и промежуточных
стадий: от первично-личиночного анаморфоза до полного метаморфоза. При
недостатке питательных веществ в яйце личиночные стадии позволяют завершить
морфогенез, а также в ряде случаев обеспечивают расселение особей.
. Онтогенез с выпадением отдельных стадий.
Утрата личиночных стадий и/или стадий бесполого размножения: пресноводные
гидры, олигохеты, большинство брюхоногих моллюсков. Утрата конечных стадий и
размножение на ранних этапах онтогенеза: неотения[8].
Также онтогенез можно классифицировать как прямой
и непрямой онтогенез. Непрямой онтогенез характерен для насекомых, которые
проходят стадию личинки. В медицине знание непрямого онтогенеза некоторых
насекомых имеет большое значение, поскольку многие насекомые являются
паразитами человека [7].
Для человека же характерен прямой онтогенез.
Прямой онтогенез в свою очередь делится на неличинковый и внутриутробный.
Неличинковый сводится к развитию взрослой особи из яйца без прохождения стадии
личинки. Внутриутробный прямой онтогенез характерен для человека.
.2 Понятие филогенеза
Филогенез- историческое развитие живых
организмов: как всего органического мира Земли, так и отдельных таксонов (от
царств до видов). Термин «филогенез» ввел Э. Геккель в 1866 г.
Раздел биологии, изучающий филогенез и его
закономерности, называется филогенетикой. Исследование филогенеза и
реконструкция его необходимы для развития общей теории эволюции и построения
естественной системы организмов; выводы филогенетики важны также для
исторической геологии и стратиграфии.
Часто термин «филогенетический» используется как
синоним термина «эволюционный»; однако следует иметь в виду, что филогенетика
изучает не механизмы эволюции, а лишь констатирует родственные связи между
таксонами.
Выражение «филогенетические преобразования»
следует понимать как преобразования в ходе исторического развития группы
организмов [2].
Геккель предложил использовать для исследования
филогенеза метод тройного параллелизма - сопоставление данных палеонтологии,
сравнительной анатомии и эмбриологии. Ныне в филогенетике всё шире используются
данные генетики, биохимии, молекулярной биологии, этологии, биогеографии,
физиологии, паразитологии. Филогенез большинства групп носит характер
адаптивной радиации. Графическое изображение филогенеза - родословное (или
филогенетическое) древо. Основная движущая сила, определяющая адаптивный
характер филогенетических преобразований организмов, - естественный отбор.
Конкретные направления филогенеза ограничиваются исторически сложившимися
особенностями генетической системы, морфогенеза и фенотипа каждой конкретной
группы. Любые филогенетические преобразования происходят посредством
перестройки онтогенезов особей; при этом приспособит, ценность могут иметь
изменения любой стадии индивидуального развития. Таким образом, филогенез представляет
собой преемственный ряд онтогенезов последовательных поколений.
Филогенез различных групп организмов изучен
неравномерно, что определяется разной степенью сохранности ископаемых остатков,
древностью данной группы и т. д. Наиболее исследован филогенез позвоночных
(особенно высших групп), из беспозвоночных - филогенез моллюсков, иглокожих,
членистоногих, плеченогих. Плохо изучен филогенез прокариот и низших растений.
Дискуссионной остаётся проблема происхождения различных типов организмов и
взаимоотношений между ними [1].
.3 Соотношение онтогенеза и филогенеза
Итак, онтогенезом называется индивидуальное
развитие организма, а филогенезом - историческое развитие группы организмов.
Понятия онтогенеза и филогенеза неразрывно связаны между собой: с точки зрения
эволюционной теории, историческое развитие живой природы представляет собой
чреду онтогенезов.
Термины «онтогенез» и «филогенез» используются
для описания развития, поэтому между этими различными понятиями существуют и
признаки различия, и признаки сходства. В Таблице1 представлена сравнительная
характеристика онто и филогенеза.
Таблица1
Сравнительная характеристика онтогенеза и
филогенеза
|
Критерии
для сравнения
|
Онтогенез
|
Филогенез
|
|
Признаки
различия
|
|
Исходная
запрограммированность процессов
|
Наличие
уникальной неизменной генетической программы развития, сформированной
вследствие мейоза и оплодотворения
|
Генофонд
эволюционирующей группы непрерывно изменяется, ряд изменений генофонда связан
с адаптациогенезом
|
|
Продолжительность
и периодизация
|
Протекает
в сжатые сроки (часы, месяцы, годы), существует начало и окончание
|
Протекает
в исторически длительные сроки (многие тысячи и миллионы лет); принципиально
не ограничен
|
|
Признаки
сходства
|
|
Обратимость
или необратимость
|
Необратим:
невозможен возврат к предыдущим стадиям
|
Необратим:
исчезнувший признак не может вновь появиться в прежнем виде
|
|
Углубление
специализации
|
Специализация
углубляется: по мере развития уменьшается вероятность смены траектории
онтогенеза
|
Прогрессирующая
специализация: группа, вступившая на путь специализации, в дальнейшем
развитии будет идти по пути все более глубокой специализации
|
|
Наличие
адаптивной радиации
|
Поливариантность
онтогенеза обеспечивает возможность приспособления к различным условиям
|
Существует
правило адаптивной радиации: группа, у которой появляется безусловно
прогрессивный признак или совокупность таких признаков, дает начало множеству
новых групп, формирующих множество новых экологических ниш и даже выходящих в
иные среды обитания
|
|
Равномерность
или неравномерность процессов
|
Скорость
процессов роста и развития изменяется
|
Темпы
эволюционных преобразований различны: типа эволюции: брадителлическая
(медленные темпы), горотеллическая(средние темпы) итахителлическая (быстрые
темпы)
|
|
Целостность
и преемственность отдельных этапов
|
Признаки,
появляющиеся на более поздних стадиях, базируются на признаках, проявляющихся
на ранних стадиях
|
Существует
правило интеграции биологических систем: новые, эволюционно молодые группы
организмов вбирают в себя все эволюционные достижения предковых групп.
|
|
Наличие
цикличности
|
Существует
цикличность старения и омоложения
|
Существует
цикличность, отраженная в виде правила смены фаз: различные механизмы
эволюции закономерно сменяют друг друга
|
Таким образом, можно сделать следующие выводы:
Онтогенез - повторение филогенеза
Онтогенез - основа филогенеза [8].
Глава 2. Закон зародышевого сходства
.1 Закон зародышевого сходства К. Бэра
Исследователи начала XIX в. впервые стали
обращать внимание на сходство стадий развития эмбрионов высших животных со
ступенями усложнения организации, ведущими от низкоорганизованных форм к
прогрессивным. В 1828 г. Карл фон Бэр сформулировал закономерность, которую
называют Законом Бэра: "Чем более ранние стадии индивидуального развития
сравниваются, тем больше сходства удается обнаружить". Сопоставляя стадии
развития зародышей разных видов и классов хордовых, К. Бэр сделал следующие
выводы.
. Эмбрионы животных одного типа на ранних
стадиях развития сходны.
. Они последовательно переходят в своем развитии
от более общих признаков типа ко все более частным. В последнюю очередь
развиваются признаки, указывающие на принадлежность эмбриона к определенному
роду, виду, и, наконец, индивидуальные черты.
. Эмбрионы разных представителей одного типа
постепенно обособляются друг от друга (рис. 1).
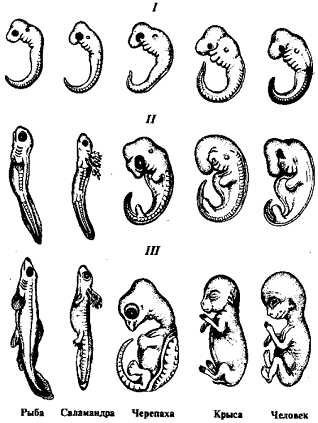
Рисунок 1. Сходство зародышей разных классов
позвоночных на разных этапах
К. Бэр, не будучи эволюционистом, не мог
связывать открытые им закономерности индивидуального развития с процессом
филогенеза. Поэтому сделанные им обобщения имели значение не более чем
эмпирических правил.
Развитие эволюционной идеи в последующем
позволило объяснить сходство ранних зародышей их историческим родством, а
приобретение ими все более частных черт с постепенным обособлением друг от
друга - действительным обособлением соответствующих классов, отрядов, семейств,
родов и видов в процессе эволюции.
Эволюционное учение, разработанное Ч. Дарвином,
ярко высветило фундаментальное значение проблемы онтогенетического развития. Зародышевое
сходство объясняется теперь действительным родством организмов, а их
постепенное расхождение (эмбриональная дивергенция) служит очевидным отражением
исторического расхождения данных форм (филогенетической дивергенции). В
зародыше потомков, писал Дарвин, мы видим «смутный портрет» предков.
Следовательно, по индивидуальному развитию можно проследить историю данного
вида [1].
.2 Биогенетический закон Э.Геккеля
Ф. Мюллер в работе «За Дарвина» (1864г.)
сформулировал мысль, что изменения онтогенетического развития, лежащие в основе
процесса эволюции, могут выражаться в изменениях ранних или поздних стадий
развития органов. В первом случае сохраняется лишь общее сходство молодых
эмбрионов. Во втором случае наблюдается продление и усложнение онтогенеза, связанное
с надставкой стадий и повторением (рекапитуляцией) в индивидуальном развитии
признаков более далеких взрослых предков.
Работы Мюллера послужили основой для формулировки Э. Геккелем (1866) основного
биогенетического закона, согласно которому онтогенез есть краткое и быстрое
повторение филогенеза. То есть органическая особь повторяет во время быстрого и
краткого течения своего индивидуального развития важнейшие из тех изменений
формы, через которые прошли ее предки во время медленного и длительного течения
их палеонтологического развития по законам наследственности и изменчивости.
Признаки взрослых предков, которые повторяются в эмбриогенезе потомков, он
назвал палингенезами. Приспособления к эмбриональным или личиночным стадиям
получили название ценогенезов.
Однако представления Геккеля сильно отличались
от взглядов Мюллера в вопросе о соотношении онтогенеза и филогенеза в процессе
эволюции. Мюллер считал, что эволюционно новые формы возникают путем изменения
хода индивидуального развития, свойственного их предкам, т.е. изменения
онтогенеза первичны по отношению к филогенетическим изменениям. По Геккелю,
наоборот, филогенетические изменения предшествуют изменениям индивидуального
развития. Эволюционно новые признаки возникают не в ходе онтогенеза, а у
взрослого организма. Эволюционирует взрослый организм, и в процессе этой
эволюции признаки сдвигаются на более ранние стадии онтогенеза.
Таким образом, возникла проблема соотношения
онтогенеза и филогенеза, которая не разрешена и до сих пор.
Геккель в отличие от Мюллера считал, что на
филогенез оказывают влияние лишь удлинение онтогенеза путем надставки стадий, а
все другие стадии остаются без изменения. Следовательно, Геккель принял только
второй путь исторических изменений онтогенеза (по Мюллеру) и оставил в стороне
изменение самих стадий онтогенеза как основы филогенетических преобразований.
Именно на этой форме взаимообусловленности онтогенеза и филогенеза делали
акцент Дарвин и Мюллер. Трактовка биогенетического закона в понимании Мюллера
позднее была развита А.Н. Северцовым (1910-1939) в теории филэмбриогенезов.
Северцов разделял взгляды Мюллера о первичности онтогенетических изменений по
отношению к изменениям взрослых организмов и рассматривал онтогенез не только
как результат филогенеза, но и как его основу. Онтогенез не только удлиняется
путем прибавления стадий: он весь перестраивается в процессе эволюции; он имеет
свою историю, закономерно связанную с историей взрослого организма и частично
ее определяющую.
Филэмбриогенезы - это эмбриональные изменения,
связанные с филогенетическим развитием взрослого организма. В процессе эволюции
перестраиваются все стадии развития. Новые изменения падают нередко на
последние стадии формообразования. Онтогенез усложняется путем прибавления, или
надставки, стадий (анаболия). Только в этом случае имеются все предпосылки для
повторения в онтогенезе исторических этапов развития данных частей у далеких
предков (рекапитуляция). Онтогенез может, однако, изменяться и на любых иных
стадиях развития, отклоняя при этом все позднейшие стадии от прежнего пути
(девиация). Наконец, возможно и изменение самих зачатков органов или частей
(архаллаксис). Тогда весь онтогенез оказывается измененным и в индивидуальном
развитии потомков не сохраняется указаний на последовательность прохождения
исторических этапов развития их предков [1].
Биологическая сущность биогенетического закона
Э. Геккеля
Биогенетический закон Геккеля и теория
филэмбриогенезов Северцова играют важную роль в развитии морфологии и самого
эволюционного учения. Изучение индивидуального развития животных дало
достаточно доказательств их исторического развития. Биогенетический закон
является важной составной частью разработанного Э. Геккелем метода тройного
параллелизма, с помощью которого производят реконструкцию филогенеза. Этот
метод основан на сопоставлении данных морфологии, эмбриологии и палеонтологии.
Морфологи при реконструкции филогенеза до сих пор пользуются геккелевским
принципом, согласно которому онтогенез потомков кратко повторяет,
рекапитулирует этапы филогенеза предков.Опираясь только на основной
биогенетический закон, невозможно объяснить процесс эволюции: бесконечное
повторение пройденного само по себе не рождает нового. Так как жизнь существует
на Земле благодаря смене поколений конкретных организмов, эволюция ее протекает
благодаря изменениям, происходящим в их онтогенезах. Эти изменения сводятся к
тому, что конкретные онтогенезы отклоняются от пути, проложенного предковыми
формами, и приобретают новые черты.
К таким отклонениям относятся, например, ценогенезы
- приспособления, возникающие у зародышей или личинок и адаптирующие их к
особенностям среды обитания. У взрослых организмов ценогенезы не сохраняются.
Примерами ценогенезов являются роговые образования во рту личинок бесхвостых
земноводных, облегчающие им питание растительной пищей. В процессе метаморфоза
у лягушонка они исчезают и пищеварительная система перестраивается для питания
насекомыми и червями. К ценогенезам у плацентарных млекопитающих и человека -
плаценту с пуповиной.
Другой тип филогенетически значимых
преобразований филогенеза - филэмбриогенезы. Они представляют собой отклонения
от онтогенеза, характерного для предков, проявляющиеся в эмбриогенезе, но
имеющие адаптивное значение у взрослых форм. Так, закладки волосяного покрова
появляются у млекопитающих на очень ранних стадиях эмбрионального развития, но
сам волосяной покров имеет значение только у взрослых организмов.
Такие изменения онтогенеза, будучи полезными, закрепляются естественным отбором
и воспроизводятся в последующих поколениях. В основе этих изменений лежат те же
механизмы, которые обусловливают врожденные пороки развития: нарушение
пролиферации клеток, их перемещения, адгезии, гибели или дифференцировки.Однако
от пороков их так же, как и ценогенезы, отличает адаптивная ценность, т.е. полезность
и закрепленность естественным отбором в филогенезе.
В зависимости от того, на каких этапах
эмбриогенеза и морфогенеза конкретных структур возникают изменения развития,
имеющие значение филэмбриогенезов, различают три их типа.
. Анаболии, или надставки, возникают после того,
как орган практически завершил свое развитие, и выражаются в добавлении
дополнительных стадий, изменяющих конечный результат. К анаболиям относят такие
явления, как приобретение специфической формы тела камбалой лишь после того, как
из икринки вылупляется малек, неотличимый от других рыб, а также появление
изгибов позвоночника, сращение швов в мозговом черепе, окончательное
перераспределение кровеносных сосудов в организме млекопитающих и человека.
. Девиации - уклонения, возникающие в процессе
морфогенеза органа. Примером может являться развитие сердца в онтогенезе
млекопитающих, у которых оно рекапитулирует стадию трубки, двухкамерное и
трехкамерное строение, но стадия формирования неполной перегородки, характерной
для пресмыкающихся, вытесняется развитием перегородки, построенной и
расположенной иначе и характерной только для млекопитающих. В развитии легких у
млекопитающих также обнаруживается рекапитуляция ранних стадий предков, позднее
морфогенез идет по-новому.
. Архаллаксисы - изменения, обнаруживающиеся на
уровне зачатков и выражающиеся в нарушении их расчленения, ранних
дифференцировок или в появлении принципиально новых закладок. Классическим
примером архаллаксиса являетсяразвитие волос у млекопитающих, закладка которых
наступает на очень ранних стадиях развития и с самого начала отличается от
закладок других придатков кожи позвоночных. По типу архаллаксиса возникают
хорда у примитивных бесчерепных, хрящевой позвоночник у хрящевых рыб ,
развиваются нефроны вторичной почки у пресмыкающихся.
Ясно, что при эволюции за счет анаболии в
онтогенезах потомков полностью реализуется основной биогенетический закон, т.е.
происходят рекапитуляции всех предковых стадий развития. При девиациях ранние
предковые стадии рекапитулируют, а более поздние заменяются развитием в новом
направлении. Архаллаксисы полностью не допускают рекапитуляции в развитии
данных структур, изменяя сами их зачатки.
В эволюции онтогенеза наиболее часто встречаются
анаболии как филэмбриогенезы, лишь в малой степени изменяющие целостный процесс
развития. Девиации как нарушения морфогенетического процесса в эмбриогенезе
часто отметаются естественным отбором и встречаются поэтому значительно реже.
Наиболее редко в эволюции проявляются архаллаксисы в связи с тем, что они изменяют
весь ход эмбриогенеза, и если такие изменения затрагивают зачатки жизненно
важных органов или органов, имеющих значение эмбриональных организационных
центров, то часто они оказываются несовместимыми с жизнью [1].
Кроме ценогенезов и филэмбриогенезов в эволюции
онтогенеза могут обнаруживаться еще и отклонения времени закладки органов -
гетерохронии - и места их развития - гетеротопии. Как первые, так и вторые
приводят к изменению взаимосоответствия развивающихся структур и проходят
жесткий контроль естественного отбора. Сохраняются лишь те гетерохронии и
гетеротопии, которые оказываются полезными. Примерами таких адаптивных
гетерохронии являются сдвиги во времени закладок наиболее жизненно важных
органов в группах, эволюционирующих по типу арогенеза. Так, у млекопитающих, и
в особенности у человека, дифференцировка переднего мозга существенно опережает
развитие других его отделов.
Гетеротопии приводят к формированию новых
пространственных и функциональных связей между органами, обеспечивая в
дальнейшем их совместную эволюцию. Так, сердце, располагающееся у рыб под
глоткой, обеспечивает эффективное поступление крови в жаберные артерии для
газообмена. Перемещаясь в загрудинную область у наземных позвоночных, оно
развивается и функционирует уже в едином комплексе с новыми органами дыхания -
легкими, выполняя и здесь в первую очередь функцию доставки крови к дыхательной
системе для газообмена.
Гетерохронии и гетеротопии в зависимости от
того, на каких стадиях эмбриогенеза и морфогенеза органов они проявляются,
могут быть расценены как филэмбриогенезы разных типов. Гетеротопия семенника у
человека из брюшной полости через паховый канал в мошонку, наблюдающаяся в
конце эмбриогенеза после окончательного его формирования, - типичная анаболия
[10].
Ценогенезы, филэмбриогенезы, а также гетеротопии
и гетерохронии, оказавшись полезными, закрепляются в потомстве и
воспроизводятся в последующих поколениях до тех пор, пока новые адаптивные
изменения онтогенеза не вытеснят их, заменив собой. Благодаря этому онтогенез
не только кратко повторяет эволюционный путь, пройденный предками, но и
прокладывает новые направления филогенеза в будущем [1].
Заключение
В данной работе были рассмотрено соотношение
онто и филогенеза, дана сравнительная таблица этих понятий. Поэтому можно
сделать вывод, что онтогенез-это основа филогенеза и онтонгенез - это
повторение филогенеза в более краткосрочном периоде времени.
Именно поэтому в 19 веке ученый-биолог К. Бэр на
основе эмпирических исследований сформулировал закон зародышевого сходства,
который в дальнейшем был принят всеми эволюционистами мира, в том числе и
Ч.Дарвином. Также были сформулирован ряд других законов, в частности
биогенетических закон Э. Геккеля.
В настоящий момент эти законы являются основой
многих биологических наук, таких как генетика, анатомия человека, эмбриология.
Онтогенез не только кратко повторяет
эволюционный путь, пройденный предками, но и прокладывает новые направления
филогенеза в будущем.
Список используемых источников информации
Биология
в 2 книгах. Под редакцией В.Н. Ярыгина. -М.: Высшая школа, 2005.
«Биология.
Современная иллюстрированная энциклопедия.» Гл. ред. А. П. Горкин; М.: Росмэн,
2006.)
Иорданский
Н.Н. Эволюция жизни. М.: Академия, 2001
Мамонтов
С.Г. Биология. Общие закономерности. - М.: Дрофа, 2002.
Пехов
А.П. Биология: медицинская биология, генетика и паразитология.-М.:
ГЭОТАР-Медиа, 2011.
Тейлор
Д., Грин Н., Стаут У.Биология: в 3-х томах. Перевод с англ./ Под редакцией Р.
Сопера-М.: Мир, 2005
Яшин
А.А. Живая материя. Онтогенез жизни и эволюционная биология.-М.: «ЛКИ», 2010
<http://afonin-59-bio.narod.ru/>Общая
и теоретическая биология: генетика, эволюция, цитология, экология -
учебно-методический комплекс
<http://patologii.net/>
1.